
Abstract
Demand of flowers is increasing with time worldwide. Floriculture has become one of the most important commercial trades in agriculture. Although traditional breeding methods like hybridization and mutation breeding have contributed significantly to the development of important flower varieties, flower production and quality of flowers can be significantly improved by employing modern breeding approaches. Novel traits of significance have interest to consumers and producers, such as fragrance, new floral color, change in floral architecture and morphology, vase life, aroma, and resistance to biotic and abiotic stresses, have been introduced by genetic manipulation. The clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein (Cas) system has recently emerged as a powerful genome-editing tool for accurately changing DNA sequences at specific locations. It provides excellent means of genetically improving floricultural crops. CRISPR/Cas system has been utilized in gene editing in horticultural cops. There are few reports on the utilization of the CRISPR/Cas9 system in flowers. The current review summarizes the research work done by employing the CRISPR/Cas9 system in floricultural crops including improvement in flowering traits such as color modification, prolonging the shelf life of flowers, flower initiation, and development, changes in color of ornamental foliage by genome editing. CRISPR/Cas9 gene editing could be useful in developing novel cultivars with higher fragrance and enhanced essential oil and many other useful traits. The present review also highlights the basic mechanism and key components involved in the CRISPR/Cas9 system.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In ornamentals, various breeding strategies have been used in the past to produce new cultivars. Selection, hybridization, and mutation breeding are all traditional breeding procedures that have been employed since time to produce a variety of colors and forms, crop architecture in the case of foliage plants, and resistance to biotic and abiotic stresses. However, in ornamental plants, traditional methods have several limitations, including the fact that they are time-consuming and that majority of the ornamentals are heterozygous, resulting in polyploidy, and complex genetic components transmission [1, 2]. As a result, new approaches for improving decorative plant production are the need of time. Genome sequencing technology has had a substantial impact on the improvement of flowering crops in recent decades. More than 69 ornamental plants (whole-genome/ draft genome) have been sequenced yet [3]. The nuclear genome of cultivated hybrid rose [4], wild rose [5], scarlet sage [6], petunia [7], sunflower [8], orchids [9], and other ornamentals has been sequenced. Transcriptome data are available for chrysanthemums [10, 11], osmanthus [12], petunia [13], hydrangea [14], lavender [15], and carnation [16]. The chloroplast genome sequence is also available for a few ornamental plants [17]. Genome sequence information on important decorative plant species would be immensely beneficial in ornamental plant breeding and research following the debut of next-generation sequencing (NGS) technologies in 2005. Genetic transformation is an essential requirement for the development and improvement of flowering crops when distinguished with mutation breeding, which delivers a “subtractive” one-point improvement in flowers [18]. Ever since the very first transformation in ornamentals described in 1987, genetic transformation is acknowledged as an important method to develop desirable cultivars [19, 20].
CRISPR/Cas9 is an approach for genetic modification with immense capability and simpler implementation that has accelerated crop improvement research. CRISPR/Cas9 genome-editing technology has been employed in agriculture as well as horticultural crops and found to be effective in crop improvement [20,21,22,23]. CRISPR/Cas-based system has progressed to allow for a wide range of applications. These include CRISPR interference (CRISPRi) and CRISPR activator (CRISPRa) gene regulators, as well as development as a base editor, prime editor, and epigenetic editor. It can also be utilized for imaging and chromatin and RNA targeting. Its application has revolutionized a wide range of biological domains, including biomedical and agricultural research. CRISPR has also been created as a diagnostic tool to aid in the detection and screening of both human and plant diseases, and it was even used during the coronavirus (COVID-19) pandemic [24].
CRISPR/Cas9 Genome Editing
Significant improvement has been made in the development of effective approaches for targeted editing of the plant genomes, over the last three decades [25, 26]. Before the introduction of the CRISPR/Cas9 technology in 2013, plant genome engineering relied on sequence-specific nucleases such as zinc finger nucleases (ZENs) and transcription activator-like effectors nucleases (TALENs) [27]. There are two types of genome-editing tools that used double-strand break (DSB) nucleases. MNs (meganucleases), ZFNs, and TALENs come under the first group, that use protein–DNA interactions to enable sequence-specific DNA binding [28, 29], and the second group is further divided into two sub-groups: (a) RNA-guided system (CRISPR/Cas9 and targetrons) [30, 31] and (b) DNA-based-guided systems [structure-guided endonuclease (SGNs), peptide nucleic acids (PNAs), and triplex-forming oligonucleotides (TFOs)] [32,33,34].
The presence of pre-configured CRISPR/Cas9 ribonucleoproteins (RNPs) enables DNA-free genome editing in plants, without the need for codon optimization or highly specialized regulators for expression in host cells. An efficient protoplast transformation by manipulating the molar ratio of Cas9 and sgRNAs was optimized for grape and apple. The targeted mutagenesis insertion and deletion rate was analyzed with the help of targeted deep sequencing. Direct delivery of CRISPR/Cas9 RNPs to the protoplast system enables targeted gene editing and paves the way to the generation of DNA-free genome edited in plants [35]. Moreover, Cas9 RNPs break the target DNAs shortly after transfection and are thus quickly destroyed in cells, significantly lowering the number of undesired alterations at off-target sites [36, 37]. Preassembled CRISPR/Cas9 ribonucleoproteins can also help circumvent GMO regulations over their release as cultivars since there is no transgene integration [38, 39].
Off-target effects, which are usually caused by unanticipated cleavage at genomic locations that are similar to the target sequences, are one of the key concerns about gRNA design [40]. There are few online off-target prediction tools such as Burrows-Wheeler Alignment Tool (BWA), Bowtie [41], CCTop [42], and Cas-OFFinder [43] that can be used to predict probable off-target sites [44].
To date, the major utilization of the CRISPR/Cas9 technology in plants has been to create gene knockouts. It has also been successfully employed to create gene knockouts in ornamental plants to induce genetic alterations in Petunia inflate [45], Petunia hybrid [27, 37, 46, 47], Chrysanthemum morifolium [48], Dendrobium officinale [49], Torenia [39], Ipomoea nil [50,51,52], Lilium longiflorum and Lilium pumilum [53], and Phalaenopsis equestris [54]. These reports suggest that CRISPR/Cas9-induced mutagenesis is effective in ornamental plants. The alteration produced is precise [55, 56] and could be passed on to succeeding generations by employing the classic Mendelian segregation [57]. Further in-depth information about CRISPR technology, reports by many research groups [34, 44, 58,59,60] and may be referred for future research on the subject.
Among several transformed and genetically modified (GM) ornamental plants, few regulatory approvals have been made [61]. Certainly, the regulatory approval process has a direct effect on the economics of product development. Only three ornamental species are recorded in the International Service for the Acquisition of Agri-biotech Applications (ISAAA) database as permitted biotech/GM crops: carnation, petunia, and rose. To date, only carnation and rose are allowed in the market of a few nations, conditional to their GMO regulations [61, 62]. Only two GM carnation cultivars are now allowed to be sold as cut flowers in Europe [63]. Many GM crops, both edible and in-edible, are still under research or in limited field trials and are ready for commercialization, despite considerable opposition from some farmers and NGOs being concerned about their safety and effects on biodiversity.
The Fundamental Mechanism of the CRISPR/Cas9 System
Prokaryotes show adaptation and heritable immunity is one of the most exciting findings in microbiology in recent decades [64]. In archaea and bacteria, the CRISPR/Cas9 system has been identified as a component of the immune system that guards the host against invading phages including plasmid DNA [65]. Tandem repetitions of viral genomic segments were discovered in bacterial cells to be consistently interspaced with non-repetitive sequences, a phenomenon known as CRISPR [66]. The sequence of 29-nucleotides, separated by several 32-nucleotide spacer sequences, was discovered first in the E. coli genome [67]. Similar sequences were discovered in various bacteria and archaea [21, 68].
By acquiring phage-derived spacer sequences that protect against re-infection, the CRISPR array acts as a repository for previous infections [65]. The Cas9 proteins are encoded by a cluster of genes near the CRISPR array, which controls the three steps of the process: adaptation, CRISPR RNA (crRNA) synthesis, and interference. To offer a memory of infection, bacteria may transfer portions of external DNA into their own genomes during adaptation. When the CRISPR array is translated into two short RNAs, crRNA and trans-activating CRISPR RNA (“tracrRNA”), memory is retained [21, 64] (Fig. 1).

The CRISPR array serves as memory storage for previously acquired infections. The Cas proteins are encoded by a cluster of genes situated near to the CRISPR array, which controls the three phases of immunity. Non-repetitive sequences were discovered to be frequently interspersed between tandem repeats
Following infection, the interference machinery recognizes a particular region in the target nucleic acids by complementary base pairing, causing Cas9 enzyme-catalyzed target cleavage [69]. CRISPR/Cas9 system is designed to protect prokaryotes from viruses by undermining the selfish, often hostile instructions encoded by invading mobile genomic elements (MGEs). The type II system, which has originated from Streptococcus pyogenes, controls the cleavage of DNA at specific site which makes it a good option for genetic modification [70,71,72]. Several reports have detailed the biology of the CRISPR/Cas9 system, [64, 69, 73, 74].
To create a single molecule, tracrRNA and crRNA can be linked artificially, forming a single-guide RNA (sgRNA) that is complementary to a particular segment of a target sequence (about 20 nucleotides) at a location where a protospacer adjacent motif (PAM) is present [68]. The PAM is a short sequence motif found close to the protospacer on the invading DNA. Cas9 is led by sgRNA and utilizes the PAM sequence to identify the target region and distinguishes between self and non-self nucleotide sequences. The CRISPR locus is protected against self-cleavage by the absence of the PAM sequence in the host genome. In addition, sgRNA includes a "scaffold" sequence required for Cas9 engagement. The sgRNA guides Cas9 and uses the PAM sequence to identify the target location [21, 75].
The CRISPR/Cas9 defense pathway is divided into three steps (Fig. 2). At the time of adaptation cycle, the Cas1–Cas2 complex, which is made up of dual Cas1 dimers and a single Cas2 dimer, a protospacer from intruding viral DNA which is inserted as a new spacer into the host’s CRISPR array.

The three stages of the CRISPR-Cass9 defense system are as follows: Cas complex gets the protospacer from invading viral DNA during the adaptation stage and incorporates it as a new spacer into the host’s CRISPR array. The CRISPR array is subsequently translated into a lengthy pre-crRNA, Cas9 proteins then convert into mature crRNAs. With the aid of Cas9 proteins, these crRNAs generate RNP complexes. During the interference stage, Cas nucleases are directed by mature crRNAs, to the relevant foreign DNA using complementary base pairing to detect the invading DNA. The invading genome is eliminated when the target sequence is cleaved
The CRISPR array is subsequently transcribed into a long pre-crRNA, which is then processed into mature crRNAs by Cas9 proteins or, in certain cases, cellular RNases, each of which contains a transcribed spacer and a portion of the repeat sequence, thereafter the crRNAs form ribonucleoprotein complexes. During the interference phase, mature crRNAs drive Cas9 nucleases to the relevant foreign DNA by using complementary base pairing to identify the invading DNA. The invading genome is destroyed when the target sequence is cleaved. Multi-Cas9 protein complex acts as an interference mechanism in class 1 systems, class 2 systems, on the other hand uses one Cas9 protein to cleave the target [64]. Endonuclease cleaves DNA in a specific region of the cell, causing homology-directed repair (HDR), non-homologous end joining (NHEJ) or microhomology-mediated end joining (MEEJ) [57, 76]. HDR is a genetic modification that enables the results precision editing of repair templates [77]. MMEJ is an error-prone repair approach that combines insertions and deletions with micro homologous sequence placement inside damaged ends prior to joining [78]. Because NHEJ lacks a DNA repair template, it is also prone to errors, which frequently results in inactivation mutations [79, 80]. NHEJ is the most prevalent kind of DSB repair, which is defined by the fusion of the broken ends, might be resulting in one or more random nucleotides insertions or deletions (indels), leading to partial repair and mutations [81]. NHEJ is primarily responsible for DSB repair in somatic cells [27, 82]. When DSBs are repaired, NHEJ typically results in gene deletion or protein function impairment. Chemically stabilized double-stranded oligo-deoxy-nucleotides (dsODNs) donors with 5-phosphorylated ends can likewise be used to drive targeted insertion via NHEJ [83, 84]. The target has a low mutation frequency owing to the difficulty in adjusting the artificial DNA repair template [21, 85, 86].
Approaches to Applying CRISPR/Cas9 System in Plants
Major Web-Based Resources
One of the most fundamental elements is to guide precise changes in the genomic sequences in the desired way, which is highly dependent on sgRNA-directed identification of targeted DNA sequences. CRISPR-PLANT, CRISPR-P, and CRISPR-GE are online sgRNA design resources for plants. These tools usually provide empirical CRISPR/Cas vectors and methods that are highly valuable for wet lab investigations. By gathering sgRNA data from large-scale CRISPR/Cas research, several web-accessible databases have been developed. These platforms not only provide useful resources for sgRNA selection, but also highlight the major parameters that influence sgRNA efficacy and specificity, allowing for further sgRNA design optimization [44].
Transformation Method
The majority of CRISPR/Cas9 studies have used Agrobacterium-mediated transformation to introduce exogenous DNA into ornamental plants. A key advantage of Agrobacterium-mediated transformation is its higher incidence of single transgene insertion. Moreover, several of the most often employed A. tumefaciens strains in plant transformation (e.g., AGL1, C58C1, GV3101, LBA4404, and EHA105) show enhanced resistant tissue tolerance or greater adaptability to the preferred plant species, which can boost transformation efficiency. The hypervirulent strains EHA105, AGL1, and LBA4404 are likely to be the outcome of increased vir gene induction. These strains are employed to transform recalcitrant or monocot plants, whereas milder strains are utilized to transform non-recalcitrant dicotyledonous plants [87]. In plants that are sensitive to Agrobacterium-mediated leaf-disk transformation, sequential transformation is an alternative for Cas9-mediated genome alteration. Because the significantly larger size of Cas9 expression cassette (usually greater than 5 kb), manipulation in destination vectors is difficult. To overcome this problem, sequential transformation method of introducing Cas9 and sgRNA expression cassettes sequentially into plants makes CRISPR/Cas9-mediated genetic changes smoother. The efficiency of CRISPR/Cas9-mediated targeted genome mutagenesis is unaffected by genome size [27]. Although recent CRISPR/Cas9 toolkits, based on Golden Gate ligation or Gibson Assembly, have made it easier to assemble numerous sgRNA expression cassettes [88, 89], sequential transformation retains some advantages, including improved transformation capabilities.
The transmission of CRISPR/Cas9 nucleases through Agrobacterium tumefaciens allows for the continuous expression of CRISPR complexes, leading to a variety of chimera genotypes in tissues or organisms. In petunia, Yu et al. [37] used a Cas9 RNP-mediated transformation technique and discovered no chimeric genotypes in every regenerated plant based on next-generation sequencing data, as reported previously by Woo et al. [38]. Cas9 RNPs cleave the target DNAs as they are transected and then decay quickly in protoplasts, resulting in genotypes that are homozygous or heterozygous instead of being chimeric. When using the CRISPR/Cas9 system to target a particular gene, orthologous or paralogous genes that are found frequently in plant speciation must be examined. A full change in flower color was achieved in petunia by simultaneously targeting both loci of F3H (A and B) with CRISPR/Cas9 RNPs [37].
Due to the high percentage of chimeras and low efficiency of transformation, only a few Lilium transformations have been reported. For the first time, Yan et al. [53] used L. pumilum embryogenic callus as a target material for Agrobacterium-mediated transformation. Each embryogenic cell of the callus can develop into a somatic embryo; it could diminish or eliminate chimeras if used as explants for Agrobacterium-mediated transformation. Although the transformation cycle is quite long, cell proliferation and transformation efficiency are high when embryogenic callus is employed as a recipient, and chimeras are rare. On the other hand, despite the disadvantages of low transformation efficiency and a high number of chimeras, using vegetative tissue scales as a transformation recipient is a reasonably easy operation that takes far less time [53].
Plant protoplasts offer a flexible system for DNA-free genome editing. Protoplasts are plant cells without cell walls that provide a useful transient system for evaluating the efficacy of gene-editing reagents before moving forward with a more labor-intensive transformation process [90, 91]. PEG-mediated transfection and electro-transfection are two popular ways of delivering plasmid DNA, in vitro transcripts, or RNPs into protoplasts for CRISPR cassette transient expression. Once successful protoplast isolation, transfection, and regeneration have been established in a plant species, it could be used as a high-throughput platform for gene editing [92].The usage of the protoplast technology can be advantageous because a high level of transgenic expression could be achieved [76, 93]. The protoplast transient expression system has been demonstrated to be an effective tool for CRISPR/Cas9 genetic modification in plants. A robust mechanism aids Cas9 transfection in Petunia hybrida. Cas9 and gRNA complex-mediated genetic manipulation through plasmid vector elicited target alteration frequencies of 21 and 13% in rice and maize protoplast systems, respectively, after 72 and 48 h post-transfection [76, 94]. Within 24 h of transfection, almost comparable mutation frequencies (20%) were generated in the petunia protoplast system, demonstrating that the direct delivery of RNPs is more successful than the vector-based approach. In animal cell lines, direct administration of recombinant Cas9 proteins resulted in genetic variation ranging from 16 to 72% [36]. Differences in mutation rates between plant and animal systems could be attributed to experimental differences. Direct administration of recombinant Cas9 protein caused specific mutations in Arabidopsis, rice, and tobacco protoplasts with 17 to 23% efficiency, which was comparable with previous findings of Subburaj et al. [46] in petunia protoplasts [38]. One of the significant advantages of employing direct Cas9 protein administrations is the ability to induce mutations quickly and precisely [36, 38].
The introduction of CRISPR/Cas9 components via floral dip method is both cost-effective and simple. This is the most widely used approach for genome editing in Arabidopsis. Due to limited flower and seed production, floral-dip-mediated administration of CRISPR/Cas9 components is confined to a few plants such as Arabidopsis, flax, and tomato [95]. The key benefit of this delivery method was that it did not necessitate the use of a plant tissue culture facility.
Co-delivery of developmental regulators (DRs) with CRISPR reagents via Agrobacterium can expedite and improve gene-editing efficiency in plants. Developmental regulators are genes involved in dictating meristem identity in plants. Ectopic expression of DRs in plants has resulted in somatic embryogenesis [96]. This phenomenon was leveraged to induce de novo meristems in somatic tissues by injecting Agrobacterium cultures co-delivering DRs and gene-editing cassettes directly into soil-grown plants [92].
Heritable changes have traditionally been achieved through persistent expression of CRISPR cassettes and regeneration of transgenic progenies. Tobacco rattle virus (TRV) [97], Barley stripe mosaic virus (BMSV) [24], and Sonchus yellow net rhabdovirus (SYNV) [98] are autonomously replicating viral vectors delivered into plants via Agrobacterium that provide an alternative for heritable gene-editing in plants.
Nanotechnology has made significant advances during the last decade especially in relation to plant genetic engineering. Recent reports have highlighted the importance of nanomaterials such as carbon dots, carbon nanotubes (CNTs), and mesoporous silicon nanoparticles (MSNs) to deliver biomolecules such as DNA, RNA, RNPs, and proteins [99]. In plants, nanoparticle-mediated administration of DNA and proteins into nuclear and chloroplast genomes has been achieved [100, 101].
Biolistics, also known as particle bombardment, is a popular method for plant transformation especially in plants that are not amenable to Agrobacterium infection. However, random incorporation of DNA at multiple genomic loci and labor-intensive production of explants such as calli or immature embryos are two major limitations of biolistic delivery. In cereal crops, however, bombarding RNPs instead of plasmid DNA was effectively demonstrated to produce marker-free gene-edited plants [102,103,104].
Pollen transformations by Agrobacterium and biolistic method bypasses tissue culture regeneration step but frequently results in pollen with poor viability [105, 106]. Furthermore, pollen-tube conversions can lead to chimerism [107]. While pollen magnetoreception has made progress in addressing these issues [106], it is still limited to dicots [108]. Nanoparticles as delivery engines for plant genome editing are also promising [109], further progress is needed to make plant gene editing easier [92].
CRISPR/Cas9 Specificity
Watanabe et al. [50] observed that roughly 75% (24/32) of transgenic plants were biallelic mutants (mutations that occur on both alleles of a single gene) at the locus of interest in one generation, thus the CRISPR/Cas9 system has been found to be an extremely effective and useful tool for next-generation breeding. Moreover, the efficacy of editing is dependent on sgRNA [56, 57], and researchers are working to improve sgRNA design to boost its activity [110]. Using an optimization tool, the editing efficacy in Ipomoea nil can be boosted further [50, 111, 112]. One-third of stable transgenic plants suggest that CRISPR/Cas9 system mutation frequency in Ipomoea nil is significantly higher compared to higher plants [113]. Hoshino et al. [114] presented a high-quality genome sequencing of Ipomoea nil that enables the creation of sgRNA intended to prevent targeted mutagenesis. The application of CRISPR/Cas9 technology on Ipomea nil could provide a useful model for learning some of the key features including flower color, shape, and floral lifespan.
Kui et al. [49] demonstrated that the CRISPR/Cas9 system is capable of precisely inserting mutations into targets. However, compared with the efficacy of different targets in each candidate gene, they discovered differed mutation rates of these target sites and some targets to be completely immutable. One reason could be a difference in the greater chromatin structure of certain target regions. Some target locations on the chromosomes may be tightly folded, whereas others may be more loosely folded, resulting in a variable Cas9 protein and sgRNA complex accessibility. An appropriate expression system to drive Cas9 and sgRNA is necessary for the initial utilization of genome editing in plants. A higher mutation frequency may result from the use of more effective target locations. The effectiveness of genome editing appeared to vary among callus clones; it is likely that the higher amount of Cas9 expression tends to boost mutation efficiency, as seen in rice calli [48, 115].
The ability to perform targeted deletions between two Cas9 cut sites is a key feature of CRISPR/Cas9 technology. The capability of the system was originally tested using transient systems [116, 117]. The efficiency of fragment deletion was also shown to be adversely linked with the distance between two paired gRNA/Cas9 cut sites employing transient systems [118]. Other parameters that include cell type and cell division phase, length of deletion, the efficiency of various sgRNAs, orientation and base structure of two paired sgRNAs, and target locations and context may contribute to effective deletion [57]. After identifying genome-edited transgenic plants, T-DNA containing Cas9 can be separated through selfing or crossing, and the progeny plants that are Cas9-free can be analyzed [45].
Mutation Detection
PCR/restriction enzyme assay, TaqMan qPCR test, T7 Endonuclease I, and high-resolution melting (HRM) analysis are available to evaluate the effect of genetic mutation [119]. As compared to the above approaches, sequencing analysis is preferred for evidence-based genome editing. The novel genetic engineering methods are much more accurate than the basic tools of genetic engineering developed earlier.
Applications of CRISPR/Cas9 System in Floriculture
Flower color is among the most essential characteristics of commercial flower production and it is mostly produced from betalains, carotenoids, and flavonoids [63, 120]. Flower pigmentation had first been suppressed by inserting antisense chalcone synthase (CHS) or dihydroflavonol-4-reductase (DFR) genes into transgenic lines, which produced flower alterations in 0–89% of transgenic lines; however, the extent of color lightening differed between lines [121]. Floral color suppression was next achieved using RNAi-targeted CHS or ANS (anthocyanidin synthase) genes, revealing that RNAi was a more effective (more than 50%) strategy for producing a stable white flower color phenotype in torenia [122, 123]. RNAi has also been used to block F3H gens in torenia plants, resulting in white blooms [39, 124].
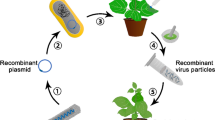
Nowadays, desired flower colors have been generated through the application of genetic modification techniques, Fig. 3. In this context, Watanabe et al. [51] used CRISPR Cas9-mediated mutagenesis to modify color in higher plants by modifying the carotenoid cleavage dioxygenase (CCD) gene in Ipomea nil and obtained pale yellow petals mutant (55.5%) plants. In a previous study, Watanabe et al. [50] targeted dihydroflavonol-4-reductase-B (DFR-B) gene, which encodes an anthocyanin biosynthetic enzyme, and observed changes in stem color throughout the initial phase of plant tissue culture. Because of biallelic changes in the Cas9 cleavage site in DFR-B, 75% of transgenic plants developed anthocyanin-free white flowers, with single base insertion or deletion of more than two bases. Nishihara et al. [39] used the CRISPR/Cas9 system to detect color changes in the Torenia fournieri from blue to white (approximately 80%) due to genetic variation of flavone 3-hydrolase (F3H), which encodes the critical enzyme for flavonoid biosynthesis. Su et al. [125] also explored T. fournieri, and observed that the abnormal expression of TfCYC2 (CYCLOIDEA) or TfRAD1 (RADIALIS) disturbed the asymmetric corolla pigmentation pattern, resulting in severely dorsal flowers, which ensures that the CYC-RAD module controls petal shape and corolla pigmentation. When TfCYC2 was down regulated or TfRAD1 was knocked out, dorsal petal identity was lost. The CRISPR/Cas9 technique was utilized to modify the phytoene desaturase (PDS) gene, which is a key enzyme in the production of carotenoids and necessary for chlorophyll biosynthesis. Zhang et al. [27] altered petunias using a CRISPR/Cas9 construct that targeted PDS and obtained a 55–87% albino phenotype. Also in Lilium pumilum (DC Fisch) and Lilium lonfiflorum (White heaven), the LpPDS gene was knocked out, and the mutants were observed to be completely albino, albino green, and pale yellow. The overall mutation rates for Fisch and White heaven cultivars were 69.57% and 63.64%, respectively [53]. Tasaki et al. [126] targeted genes in the Japanese gentian including anthocyanin 5/3'-aromatic acyltransferase (Gt5/3’AT), anthocyanin 5-O-glycosyltransferase (Gt5GT), and anthocyanin 3'-O-glycosyltransferase (Gt3GT) and concluded that for the production of blue flowers, glycosylation, following acylation of the 3'-hydroxy B-ring group in delphinidin aglycone is required. Tasaki et al. [127] also further described a type of glutathione S-transferases (GST) indicating that GST1 is a gene that is involved in anthocyanin movement in gentian flowers and is required for gentiodelphin accumulation.

CRISPR/Cas9 genome editing application for ornamental plants improvement is depicted schematically
Yu et al. [37] used Cas9-ribonucleoprotein delivery to create a petunia mutant line with mutations across both F3H genes, resulting in a pale purple-pink flower color. Only the f3ha-f3hb mutant plants had visible changes, pale purplish-pink flower color, while the rest, including plants with a single-copy gene knockout, had purple-violet flowers that resembled wild-type petunia. Chib et al. [128] established a CRISPR/Cas9-based system for future research advancements in saffron (Crocus sativus L.). Zhang et al. [129] presented evidence for redefining DPL’s role (DPL: THE R2R3-MYB transcription factor, which is connected to anthocyanin, has been linked to the regulation of corolla tube venation) in Petunia plants. The disappearance of the vein-associated anthocyanin pattern above the abaxial surface of the flower bud was produced by the CRISPR/Cas9-mediated mutation at DPL, but not corolla tube venation, suggesting that DPL had no influence over corolla tube venation development. Nitarska et al. [130] utilized the CRISPR/Cas9 system to knock out flavonoid 3’-hydroxylase (F3'H) from red blooming poinsettias (Euphorbia pulcherrima) cultivar ‘Christmas Eve,’ expecting plants with orange bracts and strong pelargonidin accumulation. The enzyme F3'H is required for the synthesis of cyanidin-type anthocyanins, which give poinsettia bracts their red color. Despite the fact that F3'H was not totally inactivated, transgenic plants' bract color changed from vivid red to brilliant reddish-orange, and cyanidin levels reduced considerably as compared to wild type. The F3’H encodes a significant enzyme in the flavonoid/anthocyanin synthesis pathway and has been mentioned as a potential target in floral color engineering.
Flower longevity is among the most important characteristics of attractive flowering plants. The majority of the flowering plants show a decrease in floral lifetime due to increased ethylene production [131]. Previously, Liu et al. [132] used virus-induced silencing to silence suspected ethylene signaling components EIL1 and EIL2 in petunia. They also observed that the EIL genes are functionally redundant and that down regulating the two genes affects floral bloom lifespan. However, EIL gene silencing resulted in no phenotypic changes in transgenic plants other than increased flower longevity, demonstrating that EIL genes have a specific function [132, 133]. Flower lifetime can be increased by reducing ethylene production, which can be reduced by targeting a critical enzyme in the ethylene metabolism [1-aminocyclopropane-1-carboxylate oxidase (ACO)] [134, 135]. Xu et al. [47] targeted 1-aminocyclopropane-1-1carboxylase (PhACO) genes (PhACO1, PhACO3, and PhACO4) and observed that PhACO1-edited mutant lines produced significantly less ethylene and had significantly longer flower lifespan. In another study, Xu et al. [136] modified the PhACO3 and PhACO4 genes in petunia cv. Mirage Rose through CRISPR/Cas9 editing showed a reduction in ethylene and extended flower longevity. EPHEMERAL1 (EPH1) is a NAC transcription factor and is thought to be a key regulator of Japanese Morning Glory (Ipomoea nil) petal senescence. Petal senescence was delayed after targeted mutagenesis of the EPH1 gene [52]. Ethylene is also thought to be an essential seed germination regulator. Naing et al. [137] observed that three distinct petunia mutants (phaco1, phaco3, and phaco4) revealed considerably lower germination percentage, along with delayed germination time and seedling growth when compared to wild-type plants. To reduce the negative impacts of ethylene suppression on seed germination and plant growth, it is critical to reevaluate when modifying the ethylene production route to maintain postharvest flower quality.
The initiation and growth of flowers are important stages in the life cycle of an ornamental plant. CRISPR/Cas9 system-based genome editing can improve various floral traits for profit [61]. Orchids are the members of the Orchidaceae family, the second largest family of flowering plants, and have significant commercial importance on a global scale [138]. However, very little genome sequencing data on orchid species are available such as Dendrobium catenatum [9], Dendrobium officinale [139], Gastrodiaelata [140], Phalaenopsis aphrodite [141], and Phalaenopsis equestris [142]. Kui et al. [49] targeted five genes in the orchid family's lignocellulose biosynthesis pathway (9C3H, 4CL, C4H, CCR, and IRX) and revealed that the CRISPR/Cas9 system may cause mutation at a rate of 10–100% for each target. Tong et al. [54] successfully produced several MADS-box mutants (97.9%) in the orchid Phalaenopsis equestris; however Semiarti et al. [143] observed low transformation efficiency (0.96%) in Phalaenopsis anabilis targeting, but genome editing using the CRISPR/Cas9 system is complex and challenging due to the huge size of the genome and higher genomic ploidy levels, as well as a higher amount of repetitive sequences. There is only one study in which the CRISPR/Cas9 technology was applied to Chrysanthemum [48]. CRISPR/Cas9 system targeted Chiridius poppei yellowish-green fluorescent protein gene (CpYGFP) and induced mutation efficiency of 0–28.9%. In the Petunia hybrida protoplast system, Subburaj et al. [46] demonstrated site-directed mutagenesis via direct administration of pure Cas9 protein preassembled with guide RNA to efficiently create mutations in nitrate reductase (NR) genes. The use of CRISPR/Cas9 to create frameshift indel mutations in PiSSK1 (S = locus F-box-interacting SKP-like 1) gene in Petunia inflata demonstrates the gene's involvement in self-incompatibility and supports the notion that SLF-containing SCF complexes are essential for compatible pollination [45].
The CRISPR/Cas9 system has evolved into a versatile and cutting-edge tool that allows scientists to manufacture any desirable alterations in plants voluntarily, allowing plant mutants to be created. Unlike traditional genetic approaches that require extensive breeding cycles, CRISPR alters a desired feature in a site-specific manner in a few generations [37]. To the best of our knowledge, this report includes all of the CRISPR/Cas9 research on ornamental plants that have been reported to date, Table 1.
Conclusion
CRISPR/Cas9 technology has been accepted to be effective in genome editing to achieve desirable characteristics in flowering crops. Genome editing in a targeted manner of genes that regulate desirable traits like flowering promotion, both by raising the number of flowers and altering flowering time and longevity, color spectrum, aromas, and creation of innovation in flower structure can help to develop desirable genotypes that can fulfill the void of modern demands and also, be fruitful for the investors or producers. It can be concluded that editing specific genes is an excellent way to enhance the characteristics of existing floriculture plants and allows us to make the most of the usage of ornamental plants for global competitiveness.
However, there are a few key drawbacks to this cutting-edge technology that must be addressed. Plant tissue culture-based gene transformation is critical for CRISPR/Cas9 based genome editing in plants. Agrobacterium-mediated gene transformation is currently the most efficient technique to obtain genome editing events, although it is confined to a small number of plant species. Hence, establishing a new transformation process that does not require plant regeneration is critical. Although plant tissue culture-based approaches can produce genome-edited plants, various difficulties, such as induced mutations or somaclonal changes during plant tissue culture, as well as the lengthy process, have hampered CRISPR/Cas9 technology's applicability. Thus, finding a delivery approach that is not dependent on tissue culture could progress the CRISPR/Cas9 genome editing.
The most common method of administering gene-editing reagents is through Agrobacterium, which has a narrow host range, and some plant species are resistant to transformation by Agrobacterium. The use of A. rhizogenes, which can substantially minimize the time between reagent administration and mutation evaluation, as well as expand the spectrum of species changed are two further ways to circumvent the regeneration process [92, 144, 145].
Off-target effects are a major concern with CRISPR/Cas9 genome editing as it may have an impact on precise breeding by changing other essential agricultural features. When Cas/sgRNA genes are incorporated into the genome, they become permanently produced in the cells, potentially causing off-target effects. Off-target effects may be reduced if the Cas9/sgRNA genes are not introduced into the plant genome and only reside in the target cells for a short duration [24]. Off-target effects can also be reduced by designing high-fidelity sgRNA and using the correct Cas9 enzymes and genome editing tools. According to Zhang and Zhang [146], combining the proofreading enzyme with the Cas9 enzyme will repair any error induced by an off-target event.
Future Perspective
Many ornamental plants are highly heterozygous, have huge genome size, polyploidy, higher chromosome number, long lifespan, self-sterility, or incapacity to produce seeds which indicate a major obstacle to using conventional breeding methods. Nowadays, the floriculture industry needs additional and more cultivars with superior characteristics such as flowering promotion, both by increasing the number of flowers and changing flowering time, floral longevity, color spectrum, aromas, and creation of innovation in flower structure.
CRISPR/Cas9 technology, which first appeared in early 2013, has been more popular in the field of genome editing due to its simplicity of use, economic, and wide applicability. We would like to mention here that there is no report available on increasing essential oil traits with the fragrance that is used in the cosmetic industry. Rose contains a fragrant and essential oil, and the nuclear genome of the hybrid rose plant [4] and wild rose [5] has been sequenced. This information can be exploited further. Advances in functional studies on key genes involved in the scent of flowers and essential oils may depend on the development of mutation technology with CRISPR/Cas9 that can lead to more efficient results, so CRISPR/Cas9-based genome editing study must be carried out to fill the gap of scent and essential oil-based flower crops. Deep knowledge about the tools of the CRISPR/Cas9 system, particularly strategies that enable the development of non-transgenic ornamentals using advanced delivery systems, will contribute to the emergence of better qualitative traits, and bring about creative ideas for competitive and sustainable flower production around the world. Therefore, non-transgenic gene-editing technologies could provide us with new varieties with augmented traits and could fulfill the void of modern demands, and also, be fruitful for the investors or producers.
Abbreviations
- CpYGF:
-
Chiridius Poppei Yellowish-green fluorescent protein
- PDS:
-
Phytoene desaturase
- NR:
-
Nitrate reductase
- C3H:
-
Coumarate 3-hydroxylase
- CH4:
-
Cinnamate 4-hydroxylase
- 4CL:
-
4-Coumarate: coenzyme A ligase
- CCR:
-
Cinnamoyl coenzyme A reductase
- IRX:
-
Irregular xylem 5
- TfRAD:
-
Torenia Fournieri RADIALSIS-like
- CAPS:
-
Cleaved amplified polymorphic sequence
- DBP:
-
DNA-binding proteins
- DFR:
-
Dihydroflavonol-4-reductase
- EPH1:
-
EPHEMERAL1
- T7E1:
-
T7 endonuclease 1
- SSK1:
-
S-locus F-box-interacting SKP1- like 1
- CCD4:
-
Carotenoid cleavage dioxygenase 4
- F3H:
-
Flavanone 3’ –hydroxylase
- ACO:
-
1-Aminocyclopropane-1-carboxylate oxidase
- DPL:
-
Deep purple
- NGS:
-
Next generation sequencing
- PCR:
-
Polymerase chain reaction
- Gt5GT:
-
Anthocyanin 5-O-glycosyltransferase
- Gt3’GT:
-
Anthocyanin 3’-O-glycosyltransferase
- Gt5/3’AT:
-
Anthocyanin 5/3’-aromatic acyltransferase
- GST:
-
Glutathione S-transferases
References
Azadi, P., Bagheri, H., Nalousi, A. M., Nazari, F., & Chandler, S. F. (2016). Current status and biotechnological advances in genetic engineering of ornamental plants. Biotechnology Advances, 34, 1073–1090.
Sharma, R., & Messar, Y. (2017). Transgenics in ornamental crops: creating novelties in economically important cut flowers. Current Science, 113, 43–52.
Zheng, T., Li, P., Li, L., & Zhang, Q. (2021). Research advances in and prospects of ornamental plant genomics. Horticulture Research. https://doi.org/10.1038/s41438-021-00499-x
Qi, W., Chen, X., Fang, P., Shi, S., Li, J., Liu, X., & Zhang, Z. (2018). Genomic and transcriptomic sequencing of Rosa hybrida provides microsatellite markers for breeding, flower trait improvement and taxonomy studies. BMC Plant Biology, 18, 1–11.
Nakamura, N., Hirakawa, H., Sato, S., Otagaki, S., Matsumoto, S., Tabata, S., & Tanaka, Y. (2018). Genome structure of Rosa multiflora, a wild ancestor of cultivated roses. DNA Research, 25, 113–121.
Dong, A. X., Xin, H. B., Li, Z. J., Liu, H., Sun, Y. Q., Nie, S., & Mao, J. F. (2018). High-quality assembly of the reference genome for scarlet sage, Salvia splendens, an economically important ornamental plant. GigaScience, 7, giy068.
Bombarely, A., Moser, M., Amrad, A., Bao, M., Bapaume, L., Barry, C. S., & Kuhlemeier, C. (2016). Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nature Plants, 2, 1–9.
Badouin, H., Gouzy, J., Grassa, C. J., Murat, F., Staton, S. E., Cottret, L., & Langlade, N. B. (2017). The sunflower genome provides insights into oil metabolism, flowering and Asterid evolution. Nature, 546, 148–152.
Zhang, G. Q., Xu, Q., Bian, C., Tsai, W. C., Yeh, C. M., Liu, K. W., & Liu, Z. J. (2016). The Dendrobium catenatumLindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Scientific Reports, 6, 1–10.
Wang, J., Wang, H., Ding, L., Song, A., Shen, F., Jiang, J., & Chen, F. (2017). Transcriptomic and hormone analyses reveal mechanisms underlying petal elongation in Chrysanthemum morifolium ‘Jinba.’ Plant Molecular Biology, 93, 593–606.
Sasaki, K., Mitsuda, N., Nashima, K., Kishimoto, K., Katayose, Y., Kanamori, H., & Ohmiya, A. (2017). Generation of expressed sequence tags for discovery of genes responsible for floral traits of Chrysanthemum morifolium by next-generation sequencing technology. BMC Genomics, 18, 1–14.
Wang, X., Cai, F., Zhang, C., Zhang, M., Li, Y., & Duan, Y. (2019). Characterization of the complete chloroplast genome of the ornamental plant Osmanthus cooperi. Mitochondrial DNA Part B, 4, 2314–2315.
Wong, J., Mudd, E. A., Hayes, A., & Day, A. (2019). The chloroplast genome sequence of the ornamental plant Petunia hybrida. Mitochondrial DNA Part B, 4, 249–250.
Lee, J., Lee, S. C., Joh, H. J., Lee, H., Sung, S. H., Kang, J. H., & Yang, T. J. (2016). The complete chloroplast genome sequence of a Korean indigenous ornamental plant Hydrangea serrata for. fertilisNakai (Hydrangeaceae). Mitochondrial DNA Part B, 1, 27–28.
Li, H., Li, J., Bai, H., Shi, L., & Wang, H. (2019). The complete chloroplast genome sequence of Lavandula dentata (Lamiaceae) and its phylogenetic analysis. Mitochondrial DNA Part B, 4, 2135–2136.
Raman, G., & Park, S. (2015). Analysis of the complete chloroplast genome of a medicinal plant, Dianthus superbus var. longicalyncinus, from a comparative genomics perspective. PLoS ONE, 10, e0141329.
Sadhukhan, A., & Huo, H. (2020). Improvement of floriculture crops using genetic modification and genome editing techniques. In A. Bhattacharya & V. Parkhi (Eds.), CRISPR/Cas genome editing (pp. 69–90). Springer.
Singh, G., Srivastava, M., & Misr, P. (2015). Genetic transformation for quality improvement in ornamental climbers. In A. Shahzad, S. Sharma, & S. A. Siddiqui (Eds.), biotechnological strategies for the conservation of medicinal and ornamental climbers (pp. 351–365). Springer.
Meyer, P., Heidmann, I., Forkmann, G., & Saedler, H. (1987). A new petunia flower colour generated by transformation of a mutant with a maize gene. Nature, 330, 677–678.
Ahn, C. H., Ramya, M., An, H. R., Park, P. M., Kim, Y. J., Lee, S. Y., & Jang, S. (2020). Progress and challenges in the improvement of ornamental plants by genome editing. Plants, 9, 687.
Corte, L. E. D., Mahmoud, L. M., Moraes, T. S., Mou, Z., Grosser, J. W., & Dutt, M. (2019). Development of improved fruit, vegetable, and ornamental crops using the CRISPR/Cas9 genome editing technique. Plants, 8, 601.
Zhu, H., Li, C., & Gao, C. (2020). Applications of CRISPR–Cas in agriculture and plant biotechnology. Nature Reviews Molecular Cell Biology, 21, 661–677.
Shipman, E. N., Yu, J., Zhou, J., Albornoz, K., & Beckles, D. M. (2021). Can gene editing reduce postharvest waste and loss of fruit, vegetables, and ornamentals. Horticulture Research, 8, 1–21.
Li, C., Brant, E., Budak, H., & Zhang, B. (2021). CRISPR/Cas: A Nobel Prize award-winning precise genome editing technology for gene therapy and crop improvement. Journal of Zhejiang University-Science B, 22, 253–284.
Bortesi, L., & Fischer, R. (2015). The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnology Advances, 33, 41–52.
Puchta, H., & Fauser, F. (2014). Synthetic nucleases for genome engineering in plants: Prospects for a bright future. The Plant Journal, 78, 727–741.
Zhang, B., Yang, X., Yang, C., Li, M., & Guo, Y. (2016). Exploiting the CRISPR/Cas9 system for targeted genome mutagenesis in petunia. Scientific Reports, 6, 1–8.
Normile, D. (2017). China sprints ahead in CRISPR therapy race. Science, 358, 20–21.
Wolfs, J. M., Hamilton, T. A., Lant, J. T., Laforet, M., Zhang, J., Salemi, L. M., & Edgell, D. R. (2016). Biasing genome-editing events toward precise length deletions with an RNA-guided TevCas9 dual nuclease. Proceedings of the National Academy of Sciences USA, 113, 14988–14993.
Jiang, J., Zhang, L., Zhou, X., Chen, X., Huang, G., Li, F., & Ying, Q. L. (2016). Induction of site-specific chromosomal translocations in embryonic stem cells by CRISPR/Cas9. Scientific Reports, 6, 1–9.
Blasco, R. B., Karaca, E., Ambrogio, C., Cheong, T. C., Karayol, E., Minero, V. G., & Chiarle, R. (2014). Simple and rapid in vivo generation of chromosomal rearrangements using CRISPR/Cas9 technology. Cell Reports, 9, 1219–1227.
Delacôte, F., Perez, C., Guyot, V., Duhamel, M., Rochon, C., Ollivier, N., & Duchateau, P. (2013). High frequency targeted mutagenesis using engineered endonucleases and DNA-end processing enzymes. PLoS ONE, 8, e53217.
Qiu, P., Shandilya, H., D’Alessio, J. M., O’Connor, K., Durocher, J., & Gerard, G. F. (2004). Mutation detection using Surveyor™ nuclease. BioTechniques, 36, 702–707.
Guha, T. K., & Edgell, D. R. (2017). Applications of alternative nucleases in the age of CRISPR/Cas9. International Journal of Molecular Sciences, 18, 2565.
Malnoy, M., Viola, R., Jung, M. H., Koo, O. J., Kim, S., Kim, J. S., et al. (2016). DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Frontiers in Plant Science, 7, 1904.
Kim, S., Kim, D., Cho, S. W., Kim, J., & Kim, J. S. (2014). Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Research, 24, 1012–1019.
Yu, J., Tu, L., Subburaj, S., Bae, S., & Lee, G. J. (2021). Simultaneous targeting of duplicated genes in Petunia protoplasts for flower color modification via CRISPR-Cas9 ribonucleoproteins. Plant Cell Reports, 40, 1037–1045.
Woo, J. W., Kim, J., Kwon, S. I., Corvalán, C., Cho, S. W., Kim, H., & Kim, J. S. (2015). DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nature Biotechnology, 33, 1162–1164.
Nishihara, M., Higuchi, A., Watanabe, A., & Tasaki, K. (2018). Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant Biology, 18, 1–9.
Zhang, X. H., Tee, L. Y., Wang, X. G., Huang, Q. S., & Yang, S. H. (2015). Off-target effects in CRISPR/Cas9-mediated genome engineering. Molecular Therapy-Nucleic Acids, 4, e264.
Langmead, B., Trapnell, C., Pop, M., & Salzberg, S. L. (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biology, 10, 1–10.
Stemmer, M., Thumberger, T., del Sol Keyer, M., Wittbrodt, J., & Mateo, J. L. (2015). CCTop: An intuitive, flexible and reliable CRISPR/Cas9 target prediction tool. PLoS ONE, 10(4), e0124633.
Bae, S., Park, J., & Kim, J. S. (2014). Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics, 30, 1473–1475.
Li, C., Chu, W., Gill, R. A., Sang, S., Shi, Y., Hu, X., & Zhang, B. (2022). Computational tools and resources for CRISPR/Cas genome editing. Genomics, Proteomics & Bioinformatics. https://doi.org/10.1016/j.gpb.2022.02.006
Sun, L., & Kao, T. H. (2018). CRISPR/Cas9-mediated knockout of PiSSK1 reveals essential role of S-locus F-box protein-containing SCF complexes in recognition of non-self S-RNases during cross-compatible pollination in self-incompatible Petunia inflata. Plant Reproduction, 31, 129–143.
Subburaj, S., Chung, S. J., Lee, C., Ryu, S. M., Kim, D. H., Kim, J. S., & Lee, G. J. (2016). Site-directed mutagenesis in Petunia×hybrida protoplast system using direct delivery of purified recombinant Cas9 ribonucleoproteins. Plant Cell Reports, 35, 1535–1544.
Xu, J., Kang, B. C., Naing, A. H., Bae, S. J., Kim, J. S., Kim, H., & Kim, C. K. (2020). CRISPR/Cas9-mediated editing of 1-aminocyclopropane-1-carboxylate oxidase1 enhances Petunia flower longevity. Plant Biotechnology Journal, 18, 287–297.
Kishi-Kaboshi, M., Aida, R., & Sasaki, K. (2017). Generation of gene-edited Chrysanthemum morifolium using multicopy transgenes as targets and markers. Plant and Cell Physiology, 58, 216–226.
Kui, L., Chen, H., Zhang, W., He, S., Xiong, Z., Zhang, Y., & Cai, J. (2017). Building a genetic manipulation tool box for orchid biology: Identification of constitutive promoters and application of CRISPR/Cas9 in the orchid, Dendrobium officinale. Frontiers in Plant Science, 7, 2036.
Watanabe, K., Kobayashi, A., Endo, M., Sage-Ono, K., Toki, S., & Ono, M. (2017). CRISPR/Cas9-mediated mutagenesis of the dihydroflavonol-4-reductase-B (DFR-B) locus in the Japanese morning glory Ipomoea (Pharbitis) nil. Scientific Reports, 7, 1–9.
Watanabe, K., Oda-Yamamizo, C., Sage-Ono, K., Ohmiya, A., & Ono, M. (2018). Alteration of flower colour in Ipomoea nil through CRISPR/Cas9-mediated mutagenesis of carotenoid cleavage dioxygenase 4. Transgenic Research, 27, 25–38.
Shibuya, K., Watanabe, K., & Ono, M. (2018). CRISPR/Cas9-mediated mutagenesis of the EPHEMERAL1 locus that regulates petal senescence in Japanese morning glory. Plant Physiology and Biochemistry, 131, 53–57.
Yan, R., Wang, Z., Ren, Y., Li, H., Liu, N., & Sun, H. (2019). Establishment of efficient genetic transformation systems and application of CRISPR/Cas9 genome editing technology in Lilium pumilum DC. Fisch. and Lilium longiflorum White Heaven. International Journal of Molecular Sciences, 20, 2920.
Tong, C. G., Wu, F. H., Yuan, Y. H., Chen, Y. R., & Lin, C. S. (2020). High-efficiency CRISPR/Cas-based editing of Phalaenopsis orchid MADS genes. Plant Biotechnology Journal, 18, 889.
Feng, Z., Mao, Y., Xu, N., Zhang, B., Wei, P., Yang, D. L., & Zhu, J. K. (2014). Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proceedings of the National Academy of Sciences USA, 111, 4632–4637.
Zhang, H., Zhang, J., Wei, P., Zhang, B., Gou, F., Feng, Z., & Zhu, J. K. (2014). The CRISPR/C as9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnology Journal, 12, 797–807.
Zhang, D., Li, Z., & Li, J. F. (2016). Targeted gene manipulation in plants using the CRISPR/Cas technology. Journal of Genetics and Genomics, 43, 251–262.
Li, J., Li, H., Chen, J., Yan, L., & Xia, L. (2020). Toward precision genome editing in crop plants. Molecular Plant, 13, 811–813.
Gaj, T., Sirk, S. J., Shui, S. L., & Liu, J. (2016). Genome-editing technologies: Principles and applications. Cold Spring Harbor Perspectives in Biology, 8, a023754.
Chen, L., Tang, L., Xiang, H., Jin, L., Li, Q., Dong, Y., & Zhang, G. (2014). Advances in genome editing technology and its promising application in evolutionary and ecological studies. Gigascience, 3, 2047–2217.
Chandler, S. F., & Sanchez, C. (2012). Genetic modification; the development of transgenic ornamental plant varieties. Plant Biotechnology Journal, 10, 891–903.
Darqui, F. S., Radonic, L. M., Hopp, H. E., & Lopez Bilbao, M. G. (2017). Biotechnological improvement of ornamental plants. Ornamental Horticulture, 23, 279.
Boutigny, A. L., Dohin, N., Pornin, D., & Rolland, M. (2020). Overview and detectability of the genetic modifications in ornamental plants. Horticulture Research. https://doi.org/10.1038/s41438-019-0232-5
Hille, F., Richter, H., Wong, S. P., Bratovič, M., Ressel, S., & Charpentier, E. (2018). The biology of CRISPR-Cas: Backward and forward. Cell, 172, 1239–1259.
Barrangou, R., Fremaux, C., Deveau, H., Richards, M., Boyaval, P., Moineau, S., & Horvath, P. (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science, 315, 1709–1712.
Jansen, R., Embden, J. D. V., Gaastra, W., & Schouls, L. M. (2002). Identification of genes that are associated with DNA repeats in prokaryotes. Molecular Microbiology, 43, 1565–1575.
Ishino, Y., Shinagawa, H., Makino, K., Amemura, M., & Nakata, A. (1987). Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. Journal of Bacteriology, 169, 5429–5433.
Mojica, F. J., & Rodriguez-Valera, F. (2016). The discovery of CRISPR in archaea and bacteria. The FEBS Journal, 283, 3162–3169.
Shivram, H., Cress, B. F., Knott, G. J., & Doudna, J. A. (2021). Controlling and enhancing CRISPR systems. Nature Chemical Biology, 17, 10–19.
Makarova, K. S., Aravind, L., Wolf, Y. I., & Koonin, E. V. (2011). Unification of Cas protein families and a simple scenario for the origin and evolution of CRISPR-Cas systems. Biology Direct, 6, 1–27.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., & Charpentier, E. (2012). A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science, 337, 816–821.
Pan, C., Ye, L., Qin, L., Liu, X., He, Y., Wang, J., & Lu, G. (2016). CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Scientific Reports, 6, 1–9.
Liao, C., & Beisel, C. L. (2021). The tracrRNA in CRISPR biology and technologies. Annual Review of Genetics, 55, 161–181.
Pramanik, D., Shelake, R. M., Kim, M. J., & Kim, J. Y. (2021). CRISPR-mediated engineering across the central dogma in plant biology for basic research and crop improvement. Molecular Plant, 14(1), 127–150.
Mojica, F. J., Díez-Villaseñor, C., García-Martínez, J., & Almendros, C. (2009). Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology, 155, 733–740.
Shan, Q., Wang, Y., Li, J., Zhang, Y., Chen, K., Liang, Z., & Gao, C. (2013). Targeted genome modification of crop plants using a CRISPR-Cas system. Nature Biotechnology, 31, 686–688.
Bassett, A. R., Tibbit, C., Ponting, C. P., & Liu, J. L. (2013). Highly efficient targeted mutagenesis of Drosophila with the CRISPR/Cas9 system. Cell Reports, 4, 220–228.
Yanik, M., Ponnam, S. P. G., Wimmer, T., Trimborn, L., Müller, C., Gambert, I., & Stieger, K. (2018). Development of a reporter system to explore MMEJ in the context of replacing large genomic fragments. Molecular Therapy-Nucleic Acids, 11, 407–415.
Chen, K., Wang, Y., Zhang, R., Zhang, H., & Gao, C. (2019). CRISPR/Cas genome editing and precision plant breeding in agriculture. Annual Review of Plant Biology, 70, 667–697.
Afzal, S., Sirohi, P., & Singh, N. K. (2020). A review of CRISPR associated genome engineering: Application, advances and future prospects of genome targeting tool for crop improvement. Biotechnology Letters, 42, 1611–1632.
Jia, H., & Wang, N. (2014). Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE, 9, e93806.
Puchta, H. (2005). The repair of double-strand breaks in plants: Mechanisms and consequences for genome evolution. Journal of Experimental Botany, 56, 1–14.
Lu, Y., Tian, Y., Shen, R., Yao, Q., Wang, M., Chen, M., & Zhu, J. K. (2020). Targeted, efficient sequence insertion and replacement in rice. Nature Biotechnology, 38, 1402–1407.
Gao, C. (2021). Genome engineering for crop improvement and future agriculture. Cell, 184, 1621–1635.
Branzei, D., & Foiani, M. (2008). Regulation of DNA repair throughout the cell cycle. Nature Reviews Molecular Cell Biology, 9, 297–308.
Puchta, H., Dujon, B., & Hohn, B. (1996). Two different but related mechanisms are used in plants for the repair of genomic double-strand breaks by homologous recombination. Proceedings of the National Academy of Sciences USA, 93, 5055–5060.
Basso, M. F., Arraes, F. B. M., Grossi-de-Sa, M., Moreira, V. J. V., Alves-Ferreira, M., & Grossi-de-Sa, M. F. (2020). Insights into genetic and molecular elements for transgenic crop development. Frontiers in Plant Science, 11, 509.
Xing, H. L., Dong, L., Wang, Z. P., Zhang, H. Y., Han, C. Y., Liu, B., & Chen, Q. J. (2014). A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biology, 14, 1–12.
Ma, X., Zhang, Q., Zhu, Q., Liu, W., Chen, Y., Qiu, R., & Liu, Y. G. (2015). A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Molecular Plant, 8, 1274–1284.
Nadakuduti, S. S., Starker, C. G., Ko, D. K., Jayakody, T. B., Buell, C. R., & Voytas, D. F. (2019). Evaluation of methods to assess in vivo activity of engineered genome-editing nucleases in protoplasts. Frontier in Plant Sciences, 10, 110.
Lin, Q., Zong, Y., Xue, C., Wang, S., Jin, S., & Zhu, Z. (2020). Prime genome editing in rice and wheat. Nature Biotechnology, 38, 582–585.
Laforest, L. C., & Nadakuduti, S. S. (2022). Advances in delivery mechanisms of CRISPR gene-editing reagents in plants. Frontiers in Genome Editing. https://doi.org/10.3389/fgeed.2022.830178
Xie, K., & Yang, Y. (2013). RNA-guided genome editing in plants using a CRISPR–Cas system. Molecular Plant, 6, 1975–1983.
Liang, Z., Zhang, K., Chen, K., & Gao, C. (2014). Targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. Journal of Genetics and Genomics, 41, 63–68.
Sandhya, D., Jogam, P., Allini, V. R., Abbagani, S., & Alok, A. (2020). The present and potential future methods for delivering CRISPR/Cas9 components in plants. Journal of Genetic Engineering and Biotechnology, 18(1), 1–11.
Lowe, K., Wu, E., Wang, N., Hoerster, G., Hastings, C., & Cho, M.-J. (2016). Morphogenic regulators baby boom and wuschel improve monocot transformation. The Plant Cell, 28, 1998–2015.
Cody, W. B., Scholthof, H. B., & Mirkov, T. E. (2017). Multiplexed gene editing and protein overexpression using a tobacco mosaic virus viral vector. Plant Physiology, 175, 23–35.
Ma, X., Zhang, X., Liu, H., & Li, Z. (2020). Highly efficient DNA-free plant genome editing using virally delivered CRISPR-Cas9. Nature Plants, 6, 773–779.
Mujtaba, M., Wang, D., Carvalho, L. B., Oliveira, J. L., Santo-Pereira, A. D. E., & Sharif, R. (2021). Nanocarrier-mediated delivery of miRNA, RNAi, and CRISPR-Cas for plant protection: current trends and future directions. ACS Agricultural Science and Technology, 1, 417–435.
Demirer, G. S., Zhang, H., Matos, J. L., Goh, N. S., Cunningham, F. J., & Sung, Y. (2019). High aspect ratio nanomaterials enable delivery of functional genetic material without DNA integration in mature plants. Nature Nanotechnology, 14, 456–464.
Kwak, S. Y., Lew, T. T. S., Sweeney, C. J., Koman, V. B., Wong, M. H., & Bohmert- Tatarev, K. (2019). Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers. Nature Nanotechnology, 14, 447–455.
Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., & Mark Cigan, A. (2016). Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nature Communications, 7, 1–7.
Liang, Z., Chen, K., Li, T., Zhang, Y., Wang, Y., & Zhao, Q. (2017). Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nature Communications, 8, 1–5.
Banakar, R., Eggenberger, A. L., Lee, K., Wright, D. A., Murugan, K., & Zarecor, S. (2019). High-frequency Random DNA Insertions upon co-delivery of CRISPR-Cas9 ribonucleoprotein and selectable marker plasmid in rice. Scientific Reports, 9, 1–13.
Wang, J., Li, Y., & Liang, C. (2008). Recovery of transgenic plants by pollen-mediated transformation in Brassica Juncea. Transgenic Research, 17, 417–424.
Zhao, X., Meng, Z., Wang, Y., Chen, W., Sun, C., & Cui, B. (2017). Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nature Plants, 3, 956–964.
Ali, A., Bang, S. W., Chung, S.-M., & Staub, J. E. (2015). Plant transformation via pollen tube-mediated gene transfer. Plant Molecular Biology Reporter, 33, 742–747.
Vejlupkova, Z., Warman, C., Sharma, R., Scheller, H. V., Mortimer, J. C., & Fowler, J. E. (2020). No evidence for transient transformation via pollen magnetofection in several monocot species. Nature Plants, 6, 1323–1324.
Demirer, G. S., Silva, T. N., Jackson, C. T., Thomas, J. B., Ehrhardt, D. W., & Mortimer, J. C. (2021). Nanotechnology to advance CRISPR-Cas genetic engineering of plants. Nature Nanotechnology, 16, 243–250.
Doench, J. G., Fusi, N., Sullender, M., Hegde, M., Vaimberg, E. W., Donovan, K. F., & Root, D. E. (2016). Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nature Biotechnology, 34, 184–191.
Lei, Y., Lu, L., Liu, H. Y., Li, S., Xing, F., & Chen, L. L. (2014). CRISPR-P: A web tool for synthetic single-guide RNA design of CRISPR-system in plants. Molecular Plant, 7, 1494–1496.
Naito, Y., Hino, K., Bono, H., & Ui-Tei, K. (2015). CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics, 31, 1120–1123.
Rani, R., Yadav, P., Barbadikar, K. M., Baliyan, N., Malhotra, E. V., Singh, B. K., et al. (2016). CRISPR/Cas9: A promising way to exploit genetic variation in plants. Biotechnology Letters, 38, 1991–2006.
Hoshino, A., Jayakumar, V., Nitasaka, E., Toyoda, A., Noguchi, H., Itoh, T., et al. (2016). Genome sequence and analysis of the Japanese morning glory Ipomoea nil. Nature Communications, 7, 1–10.
Mikami, M., Toki, S., & Endo, M. (2015). Parameters affecting frequency of CRISPR/Cas9 mediated targeted mutagenesis in rice. Plant Cell Reports, 34, 1807–1815.
Li, J. F., Norville, J. E., Aach, J., McCormack, M., Zhang, D., Bush, J., & Sheen, J. (2013). Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nature Biotechnology, 31, 688–691.
Lowder, L. G., Zhang, D., Baltes, N. J., Paul, J. W., III., Tang, X., Zheng, X., & Qi, Y. (2015). A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiology, 169, 971–985.
Xie, K., Minkenberg, B., & Yang, Y. (2015). Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proceedings of the National Academy of Sciences USA, 112, 3570–3575.
Peng, C., Wang, H., Xu, X., Wang, X., Chen, X., Wei, W., & Xu, J. (2018). High-throughput detection and screening of plants modified by gene editing using quantitative real-time polymerase chain reaction. The Plant Journal, 95, 557–567.
Tanaka, Y., Brugliera, F., Kalc, G., Senior, M., Dyson, B., Nakamura, N., et al. (2010). Flower color modification by engineering of the flavonoid biosynthetic pathway: Practical perspectives. Bioscience, Biotechnology, and Biochemistry, 74, 1760.
Aida, R., Yoshida, K., Kondo, T., Kishimoto, S., & Shibata, M. (2000). Copigmentation gives bluer flowers on transgenic torenia plants with the antisense dihydroflavonol-4-reductase gene. Plant Science, 160, 49–56.
Fukusaki, E. I., Kawasaki, K., Kajiyama, S. I., An, C. I., Suzuki, K., Tanaka, Y., & Kobayashi, A. (2004). Flower color modulations of Torenia hybrida by downregulation of chalcone synthase genes with RNA interference. Journal of Biotechnology, 111, 229–240.
Nakamura, N., Fukuchi-Mizutani, M., Miyazaki, K., Suzuki, K., & Tanaka, Y. (2006). RNAi suppression of the anthocyanidin synthase gene in Torenia hybrida yields white flowers with higher frequency and better stability than antisense and sense suppression. Plant Biotechnology, 23, 13–17.
Ono, E., Fukuchi-Mizutani, M., Nakamura, N., Fukui, Y., Yonekura-Sakakibara, K., Yamaguchi, M., & Tanaka, Y. (2006). Yellow flowers generated by expression of the aurone biosynthetic pathway. Proceedings of the National Academy of Sciences USA, 103, 11075–11080.
Su, S., Xiao, W., Guo, W., Yao, X., Xiao, J., Ye, Z., & Luo, D. (2017). The CYCLOIDEA–RADIALIS module regulates petal shape and pigmentation, leading to bilateral corolla symmetry in Torenia fournieri (Linderniaceae). New Phytologist, 215, 1582–1593.
Tasaki, K., Higuchi, A., Watanabe, A., Sasaki, N., & Nishihara, M. (2019). Effects of knocking out three anthocyanin modification genes on the blue pigmentation of gentian flowers. Scientific Reports, 9, 1–10.
Tasaki, K., Yoshida, M., Nakajima, M., Higuchi, A., Watanabe, A., & Nishihara, M. (2020). Molecular characterization of an anthocyanin-related glutathione S-transferase gene in Japanese gentian with the CRISPR/Cas9 system. BMC Plant Biology, 20, 1–14.
Chib, S., Thangaraj, A., Kaul, S., Dhar, M. K., & Kaul, T. (2020). Development of a system for efficient callus production, somatic embryogenesis and gene editing using CRISPR/Cas9 in Saffron (Crocus sativus L.). Plant Methods, 16, 1–10.
Zhang, B., Xu, X., Huang, R., Yang, S., Li, M., & Guo, Y. (2021). CRISPR/Cas9-mediated targeted mutation reveals a role for AN4 rather than DPL in regulating venation formation in the corolla tube of Petunia hybrida. Horticulture Research. https://doi.org/10.1038/s41438-021-00555-6
Nitarska, D., Boehm, R., Debener, T., Lucaciu, R. C., & Halbwirth, H. (2021). First genome edited poinsettias: Targeted mutagenesis of flavonoid 3′-hydroxylase using CRISPR/Cas9 results in a colour shift. Plant Cell Tissue and Organ Culture (PCTOC), 147, 49–60.
Pandey, S., Ranade, S. A., Nagar, P. K., & Kumar, N. (2000). Role of polyamines and ethylene as modulators of plant senescence. Journal of Biosciences, 25, 291–299.
Liu, F., Hu, L., Cai, Y., Lin, H., Liu, J., & Yu, Y. (2016). Molecular characterization and functional analysis of two petunia PhEILs. Frontiers in Plant Science, 7, 1606.
Tieman, D. M., Ciardi, J. A., Taylor, M. G., & Klee, H. J. (2001). Members of the tomato LeEIL (EIN3-like) gene family are functionally redundant and regulate ethylene responses throughout plant development. The Plant Journal, 26, 47–58.
John, P. (1997). Ethylene biosynthesis: The role of 1-aminocyclopropane-1-carboxylate (ACC) oxidase, and its possible evolutionary origin. Physiologia Plantarum, 100, 583–592.
Boatright, J. L. (2000). Transcriptional regulation of PhACO1 in Petunia hybrida (Doctoral dissertation, Purdue University).
Xu, J., Naing, A. H., Bunch, H., Jeong, J., Kim, H., & Kim, C. K. (2021). Enhancement of the flower longevity of petunia by CRISPR/Cas9-mediated targeted editing of ethylene biosynthesis genes. Postharvest Biology and Technology, 174, 111460.
Naing, A. H., Xu, J., & Kim, C. K. (2021). Editing of 1-aminocyclopropane-1-carboxylate oxidase genes negatively affects petunia seed germination. Plant Cell Reports, 41, 209.
Cardoso, J. C., Zanello, C. A., & Chen, J. T. (2020). An overview of orchid protocorm-like bodies: Mass propagation, biotechnology, molecular aspects, and breeding. International Journal of Molecular Sciences, 21, 985.
Yan, L., Wang, X., Liu, H., Tian, Y., Lian, J., Yang, R., & Sheng, J. (2015). The genome of Dendrobium officinale illuminates the biology of the important traditional Chinese orchid herb. Molecular Plant, 8, 922–934.
Yuan, Y., Jin, X., Liu, J., Zhao, X., Zhou, J., Wang, X., & Huang, L. (2018). The Gastrodiaelata genome provides insights into plant adaptation to heterotrophy. Nature Communications, 9, 1–11.
Chao, Y. T., Chen, W. C., Chen, C. Y., Ho, H. Y., Yeh, C. H., Kuo, Y. T., & Shih, M. C. (2018). Chromosome-level assembly, genetic and physical mapping of Phalaenopsis aphrodite genome provides new insights into species adaptation and resources for orchid breeding. Plant Biotechnology Journal, 16, 2027–2041.
Cai, J., Liu, X., Vanneste, K., Proost, S., Tsai, W. C., Liu, K. W., & Liu, Z. J. (2015). The genome sequence of the orchid Phalaenopsis equestris. Nature Genetics, 47, 65–72.
Semiarti, E., Nopitasari, S., Setiawati, Y., Lawrie, M. D., Purwantoro, A., Widada, J., & Asano, Y. (2020). Application of CRISPR/Cas9 genome editing system for molecular breeding of orchids. Indonesian Journal of Biotechnology, 25, 61–68.
Yoshida, K., Ma, D., & Constabel, C. P. (2015). The MYB182 protein down-regulates proanthocyanidin and anthocyanin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant Physiology, 167, 693–710.
Triozzi, P. M., Schmidt, H. W., Dervinis, C., Kirst, M., & Conde, D. (2021). Simple, efficient and open-source CRISPR/Cas9 strategy for multi-site genome editing in Populus tremula× alba. Tree Physiology, 41, 2216–2227.
Zhang, D., & Zhang, B. (2020). SpRY: Engineered CRISPR/Cas9 harnesses new genome-editing power. Trends in Genetics, 36, 546–548.
Fan, D., Liu, T., Li, C., Jiao, B., Li, S., Hou, Y., & Luo, K. (2015). Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Scientific reports, 5, 1–7.
Funding
No funding is involved for the present manuscript preparation or no financial help taking from anywhere to complete this manuscript.
Author information
Authors and Affiliations
Contributions
All the authors equally contributed in the manuscript preparation.
Corresponding authors
Ethics declarations
Conflict of interest
There is no conflict of interest with any one either within the institute or outside the institute.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sirohi, U., Kumar, M., Sharma, V.R. et al. CRISPR/Cas9 System: A Potential Tool for Genetic Improvement in Floricultural Crops. Mol Biotechnol 64, 1303–1318 (2022). https://doi.org/10.1007/s12033-022-00523-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12033-022-00523-y