
Abstract
Nowadays, biofuels, especially bioethanol, are becoming increasingly popular as an alternative to fossil fuels. Zymomonas mobilis is a desirable species for bioethanol production due to its unique characteristics, such as low biomass production and high-rate glucose metabolism. However, several factors can interfere with the fermentation process and hinder microbial activity, including lignocellulosic hydrolysate inhibitors, high temperatures, an osmotic environment, and high ethanol concentration. Overcoming these limitations is critical for effective bioethanol production. In this review, the stress response mechanisms of Z. mobilis are discussed in comparison to other ethanol-producing microbes. The mechanism of stress response is divided into physiological (changes in growth, metabolism, intracellular components, and cell membrane structures) and molecular (up and down-regulation of specific genes and elements of the regulatory system and their role in expression of specific proteins and control of metabolic fluxes) changes. Systemic metabolic engineering approaches, such as gene manipulation, overexpression, and silencing, are successful methods for building new metabolic pathways. Therefore, this review discusses systems metabolic engineering in conjunction with systems biology and synthetic biology as an important method for developing new strains with an effective response mechanism to fermentation stresses during bioethanol production. Overall, understanding the stress response mechanisms of Z. mobilis can lead to more efficient and effective bioethanol production.
Graphical Abstract

Similar content being viewed by others

Background
Currently, there is a potential threat of air pollution and health problems due to the emission of greenhouse gases. Therefore, the search for a preferred energy source is imperative. Among the potential energy sources, bioethanol has received great attention [1]. Bioethanol is derived from three main feedstocks, sugar, starch, and lignocellulosic biomass. In addition to indigenous strains of ethanol-producing microorganisms (Zymomonas mobilis and Saccharomyces cerevisiae) [2], such microorganisms can also include halophilic (e.g., Candida sp.) [3] or thermotolerant species (e.g., Kluyveromyces marxianus) [4] as well as genetically or metabolically engineered bacterium (e.g., Escherichia coli and Scheffersomyces stipitis) [5, 6]. Nevertheless, Z. mobilis, is a highly regarded bioethanol producer and some of its characteristics, including high specific ethanol yields and productivity, and more efficient ATP formation, make it a valuable strain for industrial use and research compared to S. cerevisiae [7].
The fermentation process exposes microorganisms to a variety of cellular and process stress conditions, many of which negatively affect cell viability and ethanol yield [8]. Most cellular-related stress conditions are due to situations related to cellular metabolism, such as the accumulation of ethanol and oxidative stress. Process-related stress factors include a variety of obstacles such as high temperatures, nutrient deficiencies, and impurities [9, 10].
Stress response systems within microorganisms can interact with each other through complex global regulatory networks, allowing the cell to respond simultaneously to different types of stress [11]. According to this concept, cellular responses to these environmental fluctuations are divided into two categories: physiological responses and molecular responses. It should be noted that both types of reactions are interrelated. The physiological response is a network of cellular activities that cause physiological changes, including altering the phospholipid composition and properties of the membrane, increasing the content of specific amino acids and sugars, altering the growth rate and bioethanol production, modulating ion exchange processes, and altering the ATPase activity of the plasma membrane to conserve energy. All of these physiological responses help the cell resist and counteract stress-induced damage [12]. Molecular responses are mediated by a number of regulatory proteins and transcription factors that control gene expression, so changes in the expression of these proteins can indirectly lead to phenotypic changes and ultimately physiological changes in the cell [13]. Therefore, it is necessary to understand the cellular mechanism that contributes to overcoming these stress situations in order to improve the high level of stress-tolerant microorganisms. The general stress situations that Z. mobilis may be exposed to during bioethanol production are shown in Fig. 1.

Bioethanol fermentation stress conditions and adaptation. Schematic view of process-related stress (osmotic, high temperature, lignocellulosic inhibitors, nutrient shortage, contamination with other microorganisms) and cellular-related stress (ethanol and oxidative stress) situations during bioethanol production that leads to activation of stress response mechanism in Z. mobilis
Although Z. mobilis has inherent properties to function appropriately under a variety of stresses, there are also some engineering techniques such as adaptive laboratory evolution [14], global transcription machinery engineering (gTME) [15], and genome shuffling [16] that have been successfully used in Z. mobilis species adapted to furfural, acetic acid, and ethanol stresses [15,16,17]. With the advent of biological systems and the advances in computer models, it is now possible to target genetic and metabolic changes, resulting in desirable strains with high tolerance to stress conditions. Interestingly, systems metabolic engineering has also been able to play an important role in increasing the production of products and macromolecules [18].
This review is intended to provide an overview of the stress conditions to which Z. mobilis is exposed during bioethanol production and the mechanisms for overcoming these cellular barriers from physiological and cellular aspects. Accordingly, we have discussed in the following concepts: (1) the physiological responses under stress conditions, especially related to cell membrane lipid composition, growth rate and ethanol production; (2) regulatory network elements (regulatory RNAs and proteins) controlling gene expression and metabolism; (3) metabolic and genetic engineering approaches applied to obtain stress-resistant strains. Furthermore, to achieve a better understanding of Z. mobilis performance under industrial stress situations, we compared Z. mobilis with other potential ethanol-producing microorganisms (bacteria and yeast), that their stress response mechanisms were thoroughly determined and presented in previous reports.
Effect of stress conditions on physiology and characteristics of Z. mobilis
Ethanol stress condition
Due to the increase in ethanol concentration at the end of the fermentation process, this product is one of the most common stresses in the ethanol production industry. Moreover, reactive oxygen species like superoxide and peroxide increase during the ethanol fermentation process. These compounds are toxic, and their generation is related to Fe-S containing enzymes in the respiratory chain. High concentrations of ethanol lead to ethanol stress, which increased ROS concentration and finally encountered cells with an oxidative stress condition [9]. Inhibition of cell growth, reduction in cell volume and growth rate occur at low ethanol concentrations, while at relatively high ethanol concentrations there is a decrease in cell vitality and an increase in cell death [19]. Therefore, increasing membrane fluidity, low glucose utilization, and energy production that hinder microbial growth and metabolism are examples of changes in ethanol stress conditions.
Effects on cell membrane structure and composition
One of the physiological changes in the action of alcohols such as ethanol is the negative effect on the cell membrane and fluidity. Therefore, it seems that strengthening the membrane is a crucial way to counteract the harmful effect of a high ethanol concentration [20].
Lipidomics analysis of the membrane composition of Z. mobilis revealed a high level of vaccenic acid composition in the logarithmic phase and lower levels of myristic acid, palmitic acid, and palmitoleic acid. This composition changes under ethanol stress so that the content of glyceric, palmitic, and stearic acids increases with a significant reduction in lipid/ protein content [20]. In addition, studies indicate that vaccenic acid, an unsaturated fatty acid, accounted for more than 75% of the lipid structure under normal conditions, whereas this amount decreased significantly under ethanol stress. The content of cardiolipin and phosphatidylcholine in the cell membrane also increased, while phosphatidylethanolamine and phosphatidylglycerol, the most abundant glycolipids in the cell membrane, decreased under such stress conditions [21]. For other ethanol-producing strains such as S. cerevisiae, under normal conditions, the primary fatty acids in the cell membrane are palmitic acid, palmitoleic acid, oleic acid, and stearic acid, whereas under stress conditions, the amount of phosphatidylserine increases [22]. Studies by Dombek et al. on E. coli showed high levels of vaccenic acid, low levels of palmitic acid, and a decrease in the ratio of phospholipid/protein components in the presence of 4% ethanol [23]. it can be inferred that changes in lipid composition are an evolutionary adaptation that increases survival under ethanol stress. According to these results, it appears that the unsaturated/saturated (U/S) ratio decreases during ethanol stress as a response mechanism to control and reduce membrane fluidity. In a study by Huffer et al. the content of unsaturated and saturated fatty acids was investigated in the presence of different ethanol concentrations in E. coli K12, Z. mobilis, K. marxianus and S. cerevisiae. The results show that with increasing ethanol content, the ratio of U/S decreases. In Z. mobilis and E. coli K12, this decrease was about 60% and 40%, respectively [24].
Hopanoids in bacteria such as Z. mobilis and ergosterols in yeast species such as S. cerevisiae and K. marxianus are important sterol-like compounds that control membrane function. They appear to exert a strong influence on membrane properties by increasing van der Waals forces between lipid molecules and decreasing the penetration of small molecules. Therefore, this property is advantageous for the living cells at high ethanol concentration. For example, in S. cerevisiae, genes such as erg24, erg3, and erg2 (responsible for ergosterol biosynthesis) are up-regulated under stress conditions with ethanol concentration higher than 10% [25]. On the other hand, in K. marxianus, the expression of genes such as erg3 (C-5 sterol desaturase), erg11 (lanosterol 14-α-demethylase), and erg25 (C-4 methyl sterol oxidase) decreased in the presence of 6% ethanol [26]. In addition, hopanoids in Z. mobilis account for about 30% of the bacterial dry weight [24, 27]. The abundance of hopanoids and their polar head groups, the upregulation of genes, including hnpA and hnpB, responsible for hopanoid biosynthesis in Z. mobilis at 5% ethanol concentration, and the high content of tetrahydroxybacteriohopane (THBH) and hopanols at high ethanol concentration appear to maintain membrane stability and play a key role in determining ethanol tolerance [27]. Despite some studies suggesting an increase in hopanoid biosynthesis-related genes, the total concentration of terpenoids decreased upon ethanol stress in Z. mobilis [21]. Thus, there are still controversial hypotheses about the role of these types of terpenoids in the response to ethanol [20, 21]. Consequently, a high content of saturated and unbranched fatty acids in combination with complex structures such as glycolipids and steroids increases the ability of membranes to resist the negative effects of ethanol. Table 1 summarizes the studies reported on Z. mobilis, S. cerevisiae, E. coli, and K. marxianus during ethanol stress and compares the changes of lipid composition in these microorganisms.
Effects on growth rate and metabolism
In addition to lipid composition, ethanol stress can also affect bacterial growth rate and glucose consumption. Under ethanol stress conditions, the growth rate of Z. mobilis decreases, significantly. A study by He et al.on Z.mobilis showed that the bacterium could not grow in the presence of 5% ethanol concentration [21]. When comparing ethanol-treated and untreated cells, it was found that ethanol-treated cells reached their maximum cell density in longer periods than untreated cells [21]. Comparison of growth inhibition in four species, including E. coli K12, Z. mobilis, S. cerevisiae, and K. marxianus, showed that Z. mobilis could tolerate a higher ethanol concentration than the other species [24].
As shown in Z. mobilis, most metabolic pathways are slowed in response to stressful conditions [20]. In addition, changes in gene regulation were observed under stress conditions. Most up-regulated genes were related to energy production and stress response proteins, whereas most down-regulated genes were related to translation and ribosomal structure [20, 21]. It can be concluded that the decrease in glucose consumption and bioethanol production might be related to the lower expression of enzymes responsible for glycolytic processes. In addition, the destruction of the membrane leads to a loss of water, cofactors and intermediates necessary for the activity of the enzyme [24]. Growth inhibition was also observed in other strains. In S. cerevisiae and E. coli, ethanol stress conditions affected cell division, growth, and viability in such a way that high ethanol titers resulted in cell death in S. cerevisiae and rounded and swollen cells in E. coli [28, 29]. It seems that in this case the changes in the structures somehow correlate with changes in the structure of the lipid bilayer. According to this concept, variations in lipid membrane content significantly affect selective permeability and alter cellular efflux, leading to changes in cell turgor pressure and cell shape.
Effects on cellular enzymes and metabolites
In ethanol-producing strains, the accumulation of sugars and amino acids also appears to be a physiological response to ethanol stress. Studies on the role of sugars in the response to ethanol stress in S. cerevisiae show that genes encoding TPS1 and TPS2, which are responsible for trehalose biosynthesis, are upregulated under in such conditions [25]. Glucose-fructose oxidoreductase (GFOR) expression, which is responsible for sorbitol production, is also increased in Z. mobilis in response to ethanol stress [30]. Accumulation of sorbitol and trehalose in the cytoplasm with the participation of Hsp proteins seems to stabilize proteins and prevent their denaturation [25, 30, 31]. In addition, amino acids can protect cells from damage caused by freezing, desiccation, and oxidative stress during the fermentation process [32]. Proline and tryptophan are important amino acids that increase cell tolerance to ethanol stress. According to some reports, the role of proline (PRO1 gene) in ethanol tolerance has been observed in S. cerevisiae [33]. While the amino acid proline is necessary for ethanol tolerance in yeast, tryptophan appears to play an important role in both S. cerevisiae and Z. mobilis [19, 29]. In Z. mobilis, metabolomics analysis revealed that the expression of the tryptophan operon, which consists of two genes encoding this amino acid, is upregulated 2-fold [20]. The increased expression of tryptophan biosynthesis was also observed in S. cerevisiae as a result of high expression of TAT2, TRP2, and TRP5 genes [33]. Therefore, it should be noted that the accumulation of this compound in the cytoplasm can strengthen membrane stability and prevent protein aggregation in the cytoplasm during refolding, which is also toxic to the cell [29, 34].
(Table 1)
Lignocellulosic hydrolysate inhibitors stress condition
Lignocellulosic biomass is one of the possible feedstocks for bioethanol production. Inhibitors formed during the pretreatment of lignocellulose are classified into five groups: Furanaldehydes, organic acids, aromatic compounds, alcohols and inorganic compounds [35]. The accumulation of these compounds in the growth culture produces a stress state for the microorganisms that impedes their activity by reducing growth rate, decreasing bioethanol production, inhibiting metabolic enzymes, and altering cell components and transcription levels [36, 37]. In addition, inhibitors, such as furfural, can induce oxidative stress by causing the accumulation of reactive oxygen species inside the cell. Consequently, this accumulation could damage various cellular components, including DNA, lipids, and proteins [38].
Cellular responses of Z mobilis ZM4 to representative biomass-derived inhibitors such as formic acid, acetic acid, furfural, 5-hydroxymethylfurfural, and phenol were quantified using proteomics and metabolomics methods. Changes in protein expression were observed in DNA replication, DNA recombination, DNA repair, DNA transcription, RNA translation, amino acid biosynthesis, central carbon metabolism, cell wall/membrane biogenesis, and energy production and metabolism [39].
Effects on DNA
The negative effects of furfural on DNA are consistent with the accumulation of ROS, including hydrogen peroxide, superoxide anions, and hydroxyl radicals, which lead to DNA damage and the destruction of proteins and lipids [38]. In Z. mobilis, DNA destruction is restored by upregulation of genes related to the DNA repair system and recombination, such as dnaA, uvrA, uvrB, recJ, recF [36].
Effects on lipid membrane composition
Another effect of lignocellulosic compounds on strains is the inhibition of genes related to lipid metabolism. In Z. mobilis, in the presence of high furfural concentration, the expression of some genes related to membrane composition decreased, including oprM (lipoprotein biosynthesis), kpsC (polysaccharide capsule biosynthesis), and flagellar proteins [36]. Other membrane compounds such as hopanoids were significantly reduced in the presence of furfural. These results suggest that furfural has a negative effect on membrane stability and that membrane integrity fluctuates under such stress [36]. One reaction mechanism to improve membrane stabilization is to increase the content of long-chain fatty acids in the membrane. This phenomenon was observed in S. stipitis in the presence of three lignocellulosic inhibitors (HMF, acetic acid, and vanillin). In this strain, up-regulation of genes such as INO1 (inositol synthetase), FAA22 (long-chain-fatty-acid CoA ligase), FAA24 (long-chain-fatty-acid CoA ligase 2), and YBO9 (very-long-chain 3-oxoacyl-CoA reductase), responsible for long-chain fatty acid production, enhances the ability of the microorganism to compete with these stresses [40].
Effects of membrane transporters
In addition to membrane proteins, there were some transporters whose expression changed markedly under stress with phenolic aldehydes in Z. mobilis [41]. For example, the adenine triphosphate binding cassette (ABC) and resistance-nodulation-cell division (RND) were up-regulated for about 3 and 7-fold in Z. mobilis, which seems to have a positive effect in exporting inhibitors out of the cell [41]. The induction of ABC transporter genes such as snq2 and pdr5 in S. cerevisiae was also observed under vanillin stress, which seems to have a positive effect on the excretion of vanillin from the cell [42].
Effects on proteins
Under stress conditions with lignocellulosichydrolysates (LCH), inhibition of protein synthesis was observed in most ethanologenic strains. Inhibition of protein synthesis in Z. mobilis and S. cerevisiae was studied under furfural and vanillin stress conditions [36, 42]. In S. cerevisiae, inhibition of protein synthesis led to the formation of stress granules and p-bodies in the cytoplasm; however, such phenomenon was not observed in Z. mobilis [43]. On the other hand, most of the down-regulated genes in these two microorganisms were associated with ribosomal synthesis, such as rbfA, rbsR, rplI, and rpsF in Z. mobilis [36] and Rpa 12, Rpa 190, and Rpc11 in S. cerevisiae [42, 44]. In addition, genes related to amino acid metabolism and tRNA synthesis were downregulated in the presence of lignocellulose inhibitors. These results suggest that inhibition of protein synthesis provides these microbial species with the potential energy they need to survive under such stressful conditions [44].
Although protein synthesis was inhibited under lignocellulosic inhibitor stress, the expression of some other protein increased in such situations. Most of these genes are related to carbon metabolic pathways, including the ED pathway and the tricarboxylic acid (TCA) cycle. Proteins, including Zwf (glucose-6-phosphate dehydrogenase), Adh (alcohol dehydrogenase), Ald (aldehyde dehydrogenase), Cit1 (citrate synthase), Ihd2 (isocitrate dehydrogenase), Sdh2 (succinate dehydrogenase) that play a vital role in the metabolism of carbohydrates were up-regulated [36, 43, 45]. NADH and NADPH will be produced during these processes, which seems to have a significant role in stress tolerance by controlling the redox balance.
On the other hand, NADPH produced through these mechanisms is necessary for protecting cells against ROS and oxidative stress induced by hydrolysate inhibitors. NADPH is utilized by many stress protection enzymes, including thioredoxin peroxidase and glutathione oxidoreductase [38]. Glutathione and glutathione oxidoreductase enzymes are essential for oxidative stress responses. Consequently, sulfate assimilation and cysteine biosynthesis will increase in response to hydrolysate inhibitors’ stress conditions [46].
In a study by Skerker et al., they determined that the increased demand for sulfite and cysteine is due to the heightened needs of glutathione, formed from glutamate and cysteine, during Z. mobilis growth in stressful conditions. They proposed that the cysC and cysHIJ genes are crucial for Z. mobilis’s tolerance to hydrolysates [47]. Interestingly, genes including ZMO0426 (FeS assembly protein SufBD), ZMO0427 (SufS subfamily cysteine desulfurase), and ZMO0005 (encoding sulfate adenylyltransferase subunit, CysD) were up-regulated which seems to contribute to protecting Z. mobilis 8b cells from oxidative stress caused by furfural [48].
In addition, sulfate assimilation as a response to furfural stress has been reported in E. coli and S. cerevisiae as well. Studies on E. coli revealed that cys genes were up-regulated in response to furfural detoxification, and cysteine supplementation could alleviate furfural toxicity [49]. Moreover, cysteine supplementation resulted in furfural, acetate, and ethanol tolerance in Z. mobilis. This indicates that the cysteine pool is necessary in microorganisms, and GSH (a cysteine-containing tripeptide) is the required antioxidant in response to oxidative stresses induced by hydrolysate inhibitors [50].
Along with metabolic pathways leading to NADH and NADPH production, some reductases, including oxidoreductase and NADH: flavin oxidase, play a crucial role in stress factors. For example, in Z. mobilis, under phenolic aldehydes such as vanillin, 4-hydroxybenzaldehyde, and syringaldehyde reductase enzymes were up-regulated, resulting in a reduction of these compounds into 4-hydroxy benzyl alcohol, vanillyl alcohol, and syringyl alcohol, respectively [51].
Accordingly, it can be concluded that detoxification of vanillin, furfural, and HMF is a kind of NADH-NADPH-dependent process that can alleviate inhibitors’ toxicity in the cell [43, 45]. It seems that up-regulation in enzymes and proteins that role in metabolic pathways is among efficient physiological responses, which is also common in other ethanologenic species such as S. cerevisiae and S. stipitis (Table 1).
Therefore, during ethanol fermentation, microorganisms activate DNA repair systems, produce cofactors such as NADH and NADPH, and activate genes that produce reductase enzymes to convert lignocellulosic inhibitors into less toxic compounds and detoxify cells.
High temperature stress condition
Ethanol fermentation is known to be an exothermic process, so the microorganisms have to cope with this high temperature [52]. Moreover, it is worth they state that high temperature fermentation allows simultaneous saccharification and fermentation (SSF). Interestingly, this process reduces the risk of contamination with other microorganisms [53]. Most ethanol-producing microorganisms, specifically Z. mobilis and S. cerevisiae, are among mesophilic species with the ability to grow in temperatures between 30–35℃ [52, 54]. Therefore, the use of microorganisms capable of reacting and growing under such stress conditions is of great importance.
Heat shock stress can adversely affect ethanologenic strains by destroying cellular macromolecules and impairing cellular metabolism [55]. In Z. mobilis, the increased temperature may hinder cell growth and impact cell viability, so several responses are required to adapt to this condition [52].
Effects on cell growth and ethanol fermentation
The first effect of increased temperature is the growth rate and ethanol production, observed in Z. mobilis, S. cerevisiae, and P. kudriavzevii, which is also summarized in Table 1. The results of a study by Samappito et al. showed that the growth rate of the thermally adapted strain at 37℃ and 40℃ was about 1.1 and 62.5 times higher than that of the wild type. In addition, ethanol production at 39℃ and 41℃ in thermoadapted strains was 1.8 and 38.6 times higher than wild type, respectively [44]. The thermotolerant strains of S. cerevisiae were able to grow at temperatures above 40℃ (normally they grow at 25–30℃) and achieved a growth rate of 43 and an ethanol yield of 9.2% [56]. From these results, it is clear that strains with the ability to respond and tolerate such high-temperature stress conditions are of great importance.
Effects on cell morphology and membrane composition
The physiological stress responses of Z. mobilis to heat shock conditions are complicated and involve it includes the expression of several genes related to membrane stabilization, transport, and cell division. In addition, high temperatures can affect cell morphology. In Z. mobilis, for example, elongated cells were observed at high temperatures in the wild types. This appears to be a consequence of high temperature negatively affecting bacterial DNA and inhibiting cell division. In this case, the expression of genes related to cell division, including MinD and MinC, is inactive and activates the correct placement of division sites [52]. In high temperature, Z. mobilis increase the expression of genes related to membrane structures and proteins as a response. For instance, overexpression of TolB and TolQ proteins seems to impact the membrane’s stability, positively [52]. In gram-negative bacteria, a tol-pal gene constitutes two operons of tolQRA and tolBpal. Proteins encoded by this operon are related to LPS and porins of the outer membrane, which affect membrane stability by regulating the translocation and assembly of membrane components [52, 57].
In addition, because ethanol fermentation is an exothermic process, high temperatures may expose microorganisms to oxidative stress due to the accumulation of endogenous reactive oxygen species. The accumulation of ROS in the cell can have detrimental effects, including the disruption of membrane lipids, proteins, and DNA [58]. In Z. mobilis, ZmcytC (cytochrome c peroxidase), ZMO1573 (iron-dependent peroxidase), Zmcat (catalase), Zmsod (superoxide dismutase), and ZmahpC (alkyl hydroperoxide reductase) are among the important genes involved in the response to oxidative stress in Z. mobilis during high-temperature stress conditions. Studies have shown that ZmCytC catalyzes peroxidase reactions with reduced ubiquinol-1 or NADH as an electron donor in membrane fractions, accepting electrons from the bc1 complex via cytochrome c552 to convert H2O2 to water in the periplasm [59]. Therefore, ZmCytC is critical for Z. mobilis survival at high temperatures.
Effects on nucleic acid structure and DNA repair system
High temperatures can affect intracellular structures, including DNA, tRNA, and rRNA, because an increase in temperature can alter their three-dimensional structure by disrupting hydrogen bonds. In this case, the expression of genes that play a crucial role in DNA-RNA restoration, including RadA, RadC, XseA, and SpoU is critical for coping with such stress conditions [60]. The activity of these proteins reduces DNA damage and activates repair systems. In addition, as discussed earlier in this section, elongation of Z. mobilis cells occurs under stress conditions at high temperatures [52], in this case, the expression of genes related to cell partitioning and division, including MinD and MinC inactive potential sites and activate correct placement of division sites [44]. On the other hand, the accumulation of sugars and carbohydrates confers thermotolerance to cells in some cases. For example, the accumulation of sorbitol in Z. mobilis or trehalose in S. cerevisiae and P. kudriavzevii leads to the protection of proteins from denaturation, destruction, and aggregation [30, 44, 61]. The main physiological changes under high-temperature stress conditions in Z. mobilis, S. cerevisiae, and P. kudriavzevii are listed and compared in Table 1.
Effects on metabolic pathways and intracellular metabolites
High temperature fermentation also has a negative effect on enzyme activity. This negative effect is due to inhibition of genes encoding important enzymes, including PDC and ADH. A comparison between thermotolerant and wild-type strains of Z. mobilis showed that these genes were strongly expressed in the thermoadapted strains [62]. In such conditions, it seems that accumulation of sorbitol [30] and high expression of proteins controlling translation such as HrpB (an ATP-dependent helicase) [52] positively influence the expression of such enzymes in wild-type strain. Inducing gene expression in proteins involved in ethanol production at high temperatures was also evaluated in other microorganisms. High expression of genes encoding hexokinase, glyceraldehyde-3-phosphate dehydrogenase, alcohol dehydrogenase and, isocitrate dehydrogenase in S. cerevisiae, and high expression of adh and tdh2 genes in P. kudriavzevii are some of the examples [56, 60]. Accordingly, it appears that high-temperature stress conditions may negatively affect glycolysis, TCA, and PE pathways, which play important roles in metabolism, growth, and bioethanol production. Therefore, as a response mechanism, expression of genes related to these metabolic processes enhances and controls bioethanol synthesis under heat shock and also helps the cell to produce sufficient energy needed to restore destroyed proteins and macromolecules [62]. On the other hand, the accumulation of sugars and carbohydrates confer thermotolerance to cells in some cases. For example, accumulation of sorbitol in Z. mobilis or trehalose in S. cerevisiae and P. kudriavzevii results in the protection of proteins from denaturation, destruction, and aggregation [30, 56, 60]. The main physiological changes under high-temperature stress conditions in Z. mobilis, S. cerevisiae, and P. kudriavzevii are listed and compared in Table 1.
In general, Z. mobilis can maintain cell activity and overcome the adverse effects of high temperature by altering physiological properties, biosynthesizing membrane proteins and transporters to improve stability, producing metabolites and proteins to restore and protect intracellular structures.
High glucose and osmotic stress condition
In most batch fermentation processes, the optimal sugar concentration is about 190 g/l, and sugar concentration greater than 500 g/l can lead to a stress condition that results in substrate inhibition and reduces bioethanol production [63]. Cellular metabolism is reduced under such stress conditions, and the enzymes responsible for glucose breakdown are inhibited [64]. When cells are exposed to hyperosmotic extracellular stress, they generally employ two strategies to overcome the negative effects. First, the accumulation of osmolytes such as polyols, sugars, betaines, and ectoins, also known as compatible solutes, and second, osmoadaptation to the saline cytoplasm, in which an efflux of ions such as K+, Na+, and Cl− is transported into the cell and accumulates in the cytoplasm [64]. Studies on Z. mobilis have shown that both strategies are used to overcome such stress conditions. Two main effects of osmotic stress on Z. mobilis are discussed below.
Effects on cell membrane composition and transporters
According to studies on membrane transporters, 30 genes encoding membrane proteins are differentially expressed under osmotic pressure, including the clay transporter system, ABC proteins, the type I secretion system, sugar transporters, and heavy metal transporters [65]. TolC type I protein is a significant protein that is highly expressed under osmotic stress conditions. In Z. mobilis, this protein was highly expressed under osmotic stress conditions, although its function in this species has not been elucidated comprehensively [65]. TonB-dependent transporters (TBDT) is another membrane protein inclusively expressed under osmotic stress conditions in Z. mobilis [65]. This outer membrane protein is responsible for siderophore transportation, vitamin B12, nickel compounds, and carbohydrates [66]. Under normal conditions, this protein is not expressed by Z. mobilis, whereas its expression is increased in Z. mobilis at high glucose concentration. Although, although the function of this protein in Z. mobilis is still unknown, its high expression may contribute to osmotic stress tolerance [65]. In addition, transmembrane proteins such as heavy metal transporters, Mg2+ transporter proteins, and chloride channel proteins are upregulated [65] is consistent with this phenomenon that efflux of ions inside the cytoplasm will be increased in response to high glucose concentration. Therefore, changes in cell membrane protein structure are a mechanism that is used to cope with the negative effect of hyper-osmotic conditions. This phenomenon is also observed in other ethanologenic yeasts such as S. cerevisiae and K. marxianus [67, 68].
Effects on growth rate and metabolism
Z. mobilis is able to grow at a glucose concentration of 10 to 15% [30]. In a study performed by Sturch et al., the result revealed that by enhancing initial glucose concentration in the media from 20 g/l to 200 g/l, specific bacterial growth rate and cell yields will be decreased, whereas lag-time of growth will be increased significantly [65]. Wild type of Z. mobilis can produce ethanol from initial sources of glucose, fructose, and sucrose through the ED pathway by the activity of two enzymes of pyruvate decarboxylase (PDC) and alcohol dehydrogenase (ADH). Under osmotic stress conditions, the expression of these two enzymes will be decreased significantly, which also clearly explains the reduction of ethanol production by Z. mobilis [69]. One of the key enzymes involved in osmoadaptation in Z. mobilis is the enzyme glucose-fructose oxidoreductase (GFOR), which is encoded by the gfo gene. This periplasmic enzyme accounts for approximately 1% of the soluble protein in the cell [30]. In addition, this enzyme can convert glucose and fructose to gluconolactone and sorbitol, respectively [70].
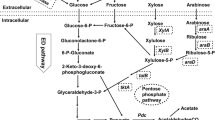
Sorbitol is one of the compatible solutes whose accumulation in the cell increases the specific growth rate and volumetric ethanol production due to the positive effect of this compound on the expression level of the pdc and adh genes [30, 34]. Although more studies still needed to be done to understand the molecular effect of sorbitol, studies indicated that the presence of sorbitol inside the cell would stabilize the cell membrane and recover the cell fluidity under osmotic stress conditions [69]. In addition, under conditions of osmotic stress, reactive oxygen species can be generated that can damage DNA, cell membrane lipids, and especially structural and functional proteins [65]. Comparison of protein synthesis under osmotic stress conditions in the presence and absence of sorbitol revealed that this sugar plays a significant role in protein synthesis and protection [69]. The accumulation of compatible solutes is also observed in other ethanologenic species such as S. cerevisiae and K. marxianus. In these two microorganisms, the accumulation of glycerol leads to osmoadaptation [67]. Interestingly, comparison of the production of compatible solutes in Z. mobilis, K. marxianus, and S. cerevisiae revealed that the production of sorbitol and glycerol is an enzymatic process. As shown in Fig. 2, the production of sorbitol in Z. mobilis occurs through the activity of the GFOR enzyme, and in S. cerevisiae the production of glycerol is controlled by the gpd1 gene, which encodes glycerol-3-phosphate dehydrogenase [68, 71]. It can be inferred that in Z. mobilis, exerting several mechanisms from membrane remodeling to substrate accumulation give rise to its osmoadaptation ability.

Comparing compatible solute production mechanism inZ. mobilis and S. cerevisiae during high glucose and osmotic stress conditions. In Z. mobilis (a), expression of gfo gene encodes glucose-fructose oxidoreductase, and in S. cerevisiae (b), gpd1 gene encodes glycerol-3-phosphate dehydrogenase contributes to tolerance to high glucose concentrations
Effect of stress conditions on expression of regulatory proteins in Z. mobilis
Regulatory network in Z. mobilis
Under stress conditions, different strains employ a variety of regulatory networks to cope with external stress conditions. These regulatory networks consist of three main components: Transcription factors, sigma factors, and regulatory RNAs [72]. Table 1 summarizes studies conducted on key regulatory networks activated during the four major fermentation stress conditions (ethanol, lignocellulose, temperature, and osmotic stress) in Z. mobilis and compares them with other ethanol-producing species.
Transcription factors
Several transcription factors control gene expression in microorganisms, and can respond to different stress conditions by regulating gene expression about 54 repressors and activators have been found in the genome of Z. mobilis [73]. Lrp/AsnC family, Xre family, LysR family, LytR family, TetR family, MarR family, RpiR family, Psp family, GntR family, and LacI are some of the major transcriptional regulators expressed in Z. mobilis under a variety of stress conditions (Table 2) [21, 36, 65].
Sigma factors
Sigma factors are another group of proteins that play an important role in the regulatory system [74,75,76]. Since the sigma factors are almost completely characterized in E. coli, comparison was made with Z. mobilis. Seven sigma factor proteins have been discovered in E. coli, including sigma 70, sigma 54, sigma 38, sigma 32, sigma 28, sigma 24, and sigma 19 [42, 76]. According to analyses the analyses compiled in Table 3, the sigma factor proteins in Z. mobilis, including sigma-D, sigma-N, sigma-H, sigma-F, and sigma-E, have structural similarity of 62%, 35%, 40%, 36%, and 30%, respectively, with the to sigma factors in E. coli. Compared to sigma factors in Z. mobilis, only five homologous proteins have been observed in this species. Sigma 38 and sigma 19 have not been detected in Z. mobilis species [44]. In E. coli, and sigma factor H, sigma factor E, sigma factor N are activated under heat shock stress conditions [76]. These regulatory proteins are activated in E. coli for the same purpose as in Z. mobilis with the same function. For example, sigma-H and sigma-E are activated during heat shock to control protein folding in the cytoplasm and periplasmic region, respectively [76]. Furthermore, activation of Sigma-N at high temperatures leads to induction of PspF and stabilization of the cell membrane in E. coli [70]. However, sigma-N has the same function (PspF activation) in Z. mobilis and E. coli, this protein role in ethanol and osmotic stress response in Z. mobilis. Apart from Sigma-N’s significant role in both Z. mobilis and E. coli, the two species also exhibit significant similarity (> 60%) in their Psp transcription factors, as described in Table 3. Therefore, PspA and PspF appear to stabilize the membrane structure under stress conditions and induce the psp operon in both species. In Table 3, the general characteristic of sigma factors in Z. mobilis and their similarity to the same proteins in E. coli are summarized.
Regulatory RNAs
Regulatory small RNAs also play an important role in modulating gene expression under various stress conditions such as ethanol, temperature, pH, and oxidative stress [77]. These regulatory compounds maintain mRNA stability and translation [78]. In addition, they fulfil their regulatory function and regulate gene expression by base-pairing with the 5´ UTR region of mRNA [78]. In Z. mobilis, fifteen sRNAs known as Zms have been discovered in which Zms2, Zms4, Zms6, and Zms18 seem to play a vital role in response to different stress conditions [34, 81]. In Z. mobilis, the 5´UTR region was found to control gene expression of the RNA chaperone Hfq under ethanol stress.
Therefore, the UTR region controls cell performance under ethanol stress conditions by up and down- regulating genes expressing Hfq proteins, thioredoxin reductase, etc.; however, no significant UTR regulatory activity was detected in Z. mobilis under xylose and acetate stress [79]. In the following section, the role of each protein during stress response in Z. mobilis will be discussed.
Regulatory networks during bioethanol fermentation stress condition
Ethanol stress condition
As we have already discussed, ethanol stress has some effects on the physiological structure of the cell, including changes in membrane proteins, ethanol production, growth rate, DNA replication, and membrane permeability [20, 21]. These changes are controlled by the regulatory system that controls gene expression under such environmental conditions. Transcriptome analysis in Z. mobilis revealed that the expression of 33 transcription factors decreased under ethanol stress conditions, including MarR, XRE, and HxIR, whose expression changed 1.9, 1.5, and 1.8 fold respectively under high ethanol concentrations [21]. TetR family transcriptional regulators are another group of the regulatory system that controls gene expression. These regulatory factors have a variety of roles, such as controlling metabolism, antibiotic resistance, and other physiological changes in the cell [80]. LysR and RpiR are also transcriptional factors regulating genes involved in quorum sensing, motility, and carbon metabolism [81]. Transcriptome analysis of Z. mobilis showed that these three factors (TetR, LysR, and RpiR) are abundant under ethanol stress situations [21]. Considering that these factors are upregulated under ethanol stress and the role of transcription factors, it is reasonable to assume that high expression of these proteins induces the expression of ABC transporters and increases sugar degradation, regulates cell metabolism, and stimulates fatty acid synthesis to compensate for the deleterious effects of ethanol on the physiological aspect of Z mobilis.
In addition, there were two genes, ZMO1107 and ZMO0347, whose expression significantly decreased in Z. mobilis under ethanol stress [21]. Analysis and homologous study on these two genes revealed that they have approximately 50% and 70% identity to Lrp/AsnC family and Hfq chaperone proteins in E. coli, respectively. In most bacterial strains, Lrp proteins are responsible for the metabolism of amino acids; for instance, in E. coli, its’ role is in the catabolism of alanine amino acid [82]. Compared to ethanologenic yeasts such as K. marxianus, metabolism of amino acids under ethanol stress is a critical activity that occurs under the control of the transcription factor GCN4 [26]. GCN4 is a type of transcription factor that is widely expressed in yeast cells [83]. This transcription factor is the main central domain for protein and amino acid biosynthesis, especially under stress conditions [26].
An important factor regulating gene expression under stress conditions, particularly heat shock and ethanol stress, is heat shock protein (Hsp). Hsps are functionally divided into two groups. Some Hsps, including GroEL, DnaK, GrpE, and DnaJ, are chaperones that attach to misfolded proteins and change their folding properly when the cell is exposed to temperature stress conditions. Other Hsps function as proteases such as Clp, Lon, and FtsH, which are responsible for removing and denaturing misfolded proteins before cell dysfunction occurs [84]. Genomic analysis of Z. mobilis revealed several complete ORFs for these Hsps from different chaperones to various ATP-dependent proteases. These proteins will be up-regulated in stress conditions like ethanol and heat shocks in Z. mobilis to combat misfold protein aggregation in the cell [73]. In an investigation by Thanonkeo et al., the expression of two Hsp proteins under ethanol concentration from 3.5 to 14% was studied in Z. mobilis. The comparison of groEL and groES expression between stressed and non-stressed cells suggested that the expression of these two proteins increased about three to six-fold under ethanol stress conditions [85]. However, according to studies by He et al., no significant change was observed in the expression of molecular chaperones including DnaJ, DnaK, Hsp70, GroES, GroEL, and Hsp33 in Z. mobilis [21].
It appears that these proteins are necessary for the enhancement of ethanol tolerance in other ethanologenic species such as K. marxianus and S. cerevisiae, either [25, 26, 86]. In K. marxianus, transcriptome analysis showed three Hsp proteins, including Hsp60, Hsp26, and Hsp78, significantly upregulate under ethanol stress conditions [26]. Different Hsp proteins were also observed in S. cerevisiae under ethanol stress, including Hsp104, Hsp82, Hsp70, Hsp12, Hsp90, etc [19, 25, 87]. . . Ethanol stress may have some negative effects on cellular components and denature proteins; consequently, upregulation of these proteins may refold proteins and prevent aggregation of misfolded proteins within the cell [25, 26, 86].
Genomic analysis of Z. mobilis revealed that this microorganism is able to express another transcription protein called Psp family [73]. Studies by He et al. revealed that three types of Psp proteins, including PspA (1.64-fold change), PspB, and PspC showed differential expression under ethanol stress [21], so it is suggested that under ethanol stress conditions, these Psp proteins activate some other proteins that help the cell to recover cell membrane and alleviate the negative effect of ethanol on cell physiology.
Sigma factors were also upregulated under ethanol stress conditions. According to studies, sigma E (1.3-fold), rpoD (1.7-fold), rpoN (1.2-fold), fliA (1.4-fold) were differentially expressed, suggesting that their expression is critical for maintaining cell survival under stress conditions [21]. Upregulation of rpoD is consistent with the expression of Ton-B transporters. According to studies, this transporter regulates under the control of ECF sigma factors, a protein in the group of sigma-70 that regulates the expression of periplasmic and outer membrane proteins [66]. According to recent studies on the role of sigma factors in Z. mobilis, sigma32 and sigma24 do not appear to be responsible for the response to high ethanol concentrations [61]. Homology and sequence analysis of sigma factor proteins in Z. mobilis is compared with E. coli in Table 3.
Besides proteins that can act as regulatory elements, there are also RNAs that can be involved in regulatory processes, which are also more efficient than transcription proteins because they do not need to be translated and directly bind to mRNAs [78]. Studies on such small RNAs in Z. mobilis revealed that four regulatory RNAs have significant functions in gene expression. Three sRNAs (Zms2, Zms6, and Zms18) showed remarkable differential expression under different ethanol concentrations from 0 to 5% [34]. Moreover, other experiments on the role of sRNA in response to ethanol stress showed that, in addition to Zms6, Zms4 is also among vital regulatory elements in Z. mobilis [88]. These regulatory RNAs participate in different cell functions such as expression of ABC transporters, Ton B receptors, carbohydrate utilization, and cell motility [34, 88]. Besides small RNA that was previously studied, some regulatory regions in 5´ untranslated regions control the expression of genes in stress conditions. In a study by Cho et al., they used transcriptomic data to analyze and identify the 5´UTR regions in Z. mobilis. The results suggest that there are 101 UTR regions in the genome of these species that act as a regulatory modules. Of all these untranslated regions, 36 UTRs function as regulators in stress responses. To test this concept, they constructed 36 5´UTR- GFP libraries and analyzed the gene expression of GFP proteins under stress and standard conditions (a strain without 5´UTR region used as control). The results showed that two strains containing the 5′UTR of RNA binding protein Hfq and the 5′UTR of thioredoxin reductase had significant fluorescent changes under 5% ethanol concentration compared to control. Furthermore, this experiment was also performed under xylose and acetate stresses in which the result showed no significant changes in regulatory regions of strain that contain the 5′UTR of thioredoxin reductase [79]. In summary, in Z. mobilis, ethanol stress leads to some physiological changes in which the expression of transcription factors such as TetR, Psp, and RpiR helps the cell to regulate cell metabolism, membrane viability, fatty acid biosynthesis, and various transporters. Along with transcription factors, sRNAs including Zms2, Zms4, Zms6, and Zms18, by controlling the regulation of ABC transporters, carbon utilization, and cell motility, are another group of regulators that their expression is critical for cell performance under such stress environment. The overall physiological and molecular responses in Z. mobilis during ethanol stress are summarized in Fig. 3(a).

Physiological and molecular stress responses inZ. mobilis during ethanol and lignocellulosic inhibitor stress conditions. a) Ethanol stress: Ethanol stress induces changes in the lipid membrane composition of Z. mobilis strains, resulting in a decrease in the level of unsaturated fatty acids to saturated fatty acids. During normal conditions, PspF binds to PspA protein. However, during ethanol stress, this protein is separated, leading to increased expression of the psp operon and high levels of PspA production. PspA proteins attach to the cell membrane structure, maintaining membrane structure and stability. Furthermore, during ethanol stress, the GFOR enzyme converts glucose to gluconolactate and sorbitol. The accumulation of sorbitol, as well as Hsp proteins and GroEL/GroES chaperones, controls protein conformation and folding. Interestingly, increasing the expression of tryptophan operon increases the level of this amino acid in the cytoplasm, preventing protein accumulation inside the cell. Additionally, the activity of regulatory RNAs (Zms), transcription factors such as TetR and sigma-70 enhance the expression of membrane transporters (ABC transporters and Ton-B), which significantly control membrane permeability under ethanol stress
b) Lignocellulosic inhibitor stress: Under stress conditions, reactive oxygen species (ROS) such as OH−, O2−, and H2O2 may be produced, which can denature DNA structure. In response, DNA repair systems and proteins such as RecJ, DnaA, RecF, and UvrB are activated to maintain DNA conformation and stability. Regulatory networks such as LacI and sigma-70 increase the expression of genes that regulate TCA enzymes, leading to increased NADH and NADPH levels. Together with oxidoreductase enzymes, these cofactors convert lignocellulosic inhibitors such as furfural, HMF, and vanillin into low-toxic structures. The activity of Lon and DnaJ proteins helps retain protein structure and fold. Transcription factors such as TetR increase the expression of transmembrane proteins such as ABC transporters and RND, which inhibit the negative effects of lignocellulosic inhibitors and direct them outside the cell
Lignocellulosic hydrolysate inhibitor stress condition
Lignocellulose inhibitors resulting from hydrolysis activity also affect the regulation of various genes and proteins, leading to bacterial tolerance to such stress conditions. Transcriptome profiling of Z.mobilis under furfural stress response showed that some proteins are upregulated, and some are downregulated as a stress response mechanism. Under furfural stress response, chaperone proteins like DnaJ and proteases like Lon gene remarkably upregulated, while groEL- groES did not show significant changes in their regulation [36]. Moreover, upregulation of transcriptional regulators such as LysR, LytR, GntR, TetR, LacI was also observed under furfural stress conditions in Z. mobilis [36]. LytR is a regulatory protein in the LytTR two-component response regulators that control cell wall structure and function and bacterial growth under stress conditions. This protein in Z. mobilis has approximately 54% similarity to Skn7 protein in S. cerevisiae, according to blastp analysis. This protein is also part of a two-component system called SLN1-YPD1-SKN7 in eukaryotic cells that serves as a transcription factor in signaling pathways under osmotic and oxidative stress conditions. Interestingly, this protein plays a central role in controlling cell wall structure and integrity during external environmental stress in S. cerevisiae [89].
Another factor that significantly affects the regulation of genes under stress conditions is Hfq, a global regulatory system that acts as an RNA-binding protein. Hfq is an RNA-binding protein that regulates gene expression during post-transcriptional levels. Hfq plays various functions in the RNA metabolism of bacteria, including stabilizing small non-coding RNAs (sRNAs) and aiding in their interactions with messenger RNAs (mRNAs), which consequently impact the stability and translation of the specific targeted mRNAs [90]. This protein is homologous to Lms proteins found in eukaryotic cells in both functions and structures. Studies showed that this protein has a significant role in resistance to lignocellulosic inhibitors such as furfural, HMF, acids, and ethanol [91]. In an experiment by Yang et al., they use Z. mobilis and its mutant to determine the effect of byproducts such as furfural, vanillin, and HMF produced under the fermentation process on the growth rate and production of ethanol by the microorganism. The results showed that the mutant, which has functional Hfq, has a shorter lag phase and high cell density than the wild type. These results assumed that Hfq is an essential protein for optimal growth of Z. mobilis under such stress conditions [91].
The genome sequence of Z. mobilis revealed that it has rpoH (sigma-32 factor), rpoE (sigma-24 factor), rpoN (sigma-54 factor), rpoD, rpoA, and fliA (sigma F), but microarray analysis revealed that only rpoD and fliA are differentially expressed under furfural stress conditions [36]. Sigma 70 is another regulatory factor that helps RNA polymerase recognize the transcription site and promotors encoded by the rpoD gene. The effect of rpoD on the growth and performance of bacteria under furfural stress was investigated by comparing control strains and mutant strains (ZM4-rpoD) [92]. The results determined that increased furfural concentration under growth profile will also increase in mutant strains. The importance of this protein on the cell under stress conditions relies on its function, which is controlling and regulating gene expression. Thus, this protein promotes the expression of genes involved in metabolic activity, enabling the cell to cope with stressful situations [92].
In addition, the effect of two regulatory factors, SigE and Hfq, was studied under furfural and acetic stress conditions. The comparison of two recombinant strains that highly expressed Sigma E and Hfq proteins with the wild type revealed that these two factors significantly affect the tolerance of Z. mobilis to stress conditions [93]. However, the increase in furfural and acetic acid concentration in the medium reduced growth rate and ethanol production in all strains, but the recombinant strain was less affected. Moreover, comparing two recombinant strains revealed that ZM4- sigE has more ethanol production and growth rate than ZM4-hfq recombinant strain [93].
In general, under LCH inhibitors stress conditions in Z. mobilis, sigma factors such as fliA and sigma D along with Hfq proteins are among important proteins regulating gene expression in bacteria. Furthermore, upregulation of transcription factors such as TetR, LacI, GntR, LysR, and LytR by regulating the expression of ABC transporters, Na+/H+ transporters, cell division, bacterial growth, and cell membrane structure helps the cell to compensate for the negative effect of LCH inhibitors stresses during bioethanol production on bacterial growth and function. The physiological and molecular stress response during LCH inhibitors stress conditions and the regulatory networks activate under such stress conditions is illustrated in Fig. 3(b).
High temperature stress condition
The molecular response of Z. mobilis to different temperatures is complex and involves the expression of genes control protein quality, membrane stability, translation, transferring proteins, and DNA repair system [44]. The heat shock proteins GroEL, GroES, and mHsp70 are among the most abundant proteins in the cell under heat stress [44]. Investigations on the expression of these proteins under heat shock in Z. mobilis revealed that the expression of groEL and groES had been increased about three to six-fold under heat shock stress compared with non-stressed cells [44].
In conclusion, Hsp proteins are among critical factors that their expression can protect peptidoglycan and prevent protein denaturation under high temperatures. Besides heat shock proteins, other proteins such as sigma factors play a crucial role in adaptation to heat shock stresses. Among several sigma factors sequenced in Z. mobilis genome, sigma 24 has a significant role in responses to heat shock and high temperatures [61]. In the study performed by Benoliel et al., results determined that sigma factor 32 and 24 regulates gene expression in both heat shock and osmotic stress in Z. mobilis [61].
According to transcriptome analysis in Z. mobilis, no significant changes in the expression of transcription factors were observed under high temperature, except WrbA protein, a regulatory protein that binds to tryptophan inhibitors (TrpR) and controls the expression of this amino acid [52]. Moreover, proteins that control protein quality, such as M16 peptidase (Zn- dependent protein) and DegP protein, a periplasmic serine peptidase, were also differentially expressed [52]. These proteins remove misfolded proteins in the periplasmic region to control the negative impact of their accumulation on the cell membrane under heat shocks [94].
Compared to S. cerevisiae, according to a study by Kim et al., profiling transcription regulators that are expressed at temperatures above 40℃ revealed that factors such as Hsf1, Msn2, Msn4, and Yap1 were significantly increased in thermotolerant strain, while the wild strain did not have any changes in these kinds of proteins [56]. These regulatory factors are summarized and compared in Table 1 in Z. mobilis, S. cerevisiae, and P. kudriavzevii. In general, it appears that under high-temperature stress in Z. mobilis, proteins quality control system and Hsp proteins are the essential elements for controlling cell tolerance to such stress conditions (Fig. 4 (a)).

Physiological and molecular stress responses inZ. mobilis during high temperature and high glucose stress conditions. a) High-temperature stress: In Z. mobilis, high temperatures can negatively affect its cell performanceThe elevated temperature can disrupt DNA structure by breaking hydrogen bonds between nucleobases, resulting in denaturation. To counteract this, the DNA repair system is activated, aided by proteins such as RadA, RadC, and Xse, to stabilize and refold the DNA. Conversely, FtsH is inhibited during this stress condition. This protein typically binds to Sigma-32 to inhibit its activity and expression under normal conditions. However, in high-temperature stress conditions, FtsH inhibition activates Sigma-32, which, in conjunction with other chaperones and proteins, refolds denatured proteins and stabilizes their structure. Notably, WrbA protein binds to TrpR (the trp operon inhibitor) to increase tryptophan expression, preventing protein aggregation within the cytoplasm. In the periplasmic region, DegP and M16 proteins degrade misfolded proteins to prevent aggregation within the cell. Finally, increased expression of transmembrane proteins and MinC and MinD proteins helps regulate membrane structure and cell division under high-temperature stress, respectively
b) High glucose stress: Elevating the concentration of glucose in the extracellular environment induces a series of intracellular signals that ultimately upregulate the expression of genes, such as the gfo gene. The increased activity of the GFOR enzyme facilitates the conversion of extracellular glucose into sorbitol. The accumulation of sorbitol inside the cell, in conjunction with chaperones like DnaJ, DnaK, and Hsp20, helps regulate and refold denatured proteins. Moreover, sorbitol enhances the expression of PDC and ADH enzymes, which control bioethanol production. Additionally, the transcription factors Sigma-54 and PspF increase the expression of the psp operon, promoting membrane stabilization during glucose stress. Transmembrane proteins, including TolC, play a crucial role in hemostasis by facilitating the flow of proteins and sugars outside the cell
High glucose and osmotic stress condition
During osmotic stress, transcriptome analysis in Z. mobilis revealed some changes and upregulations in the expression of factors and proteins, including RNA polymerase sigma factor 32, sigma 54-specific transcriptional activator PspF, and catalase [65]. According to studies on Z. mobilis, heat shock proteins such as Hsp20, heat shock protein DnaJ domain-containing protein, and chaperone DnaJ domain-containing protein were upregulated in hyperosmotic stress conditions [65]. Sigma 54, which regulates psp operon, was also upregulated, resulting in the upregulation of two major Psp proteins, PspA and PspB [65]. High expression of the Psp family transcription factor was also observed during ethanol stress. This transcription factor may play a critical role during the bioethanol fermentation process and act as a regulatory protein in response to various stresses. In contrast, other sigma factors showed a significant decrease in expression during osmotic stress conditions. Sigma 70 is a vital sigma factor that plays an essential role in cell growth under standard conditions. The species that expressed sigma 70 under high glucose concentration have lower cell densities because the increased expression of sigma 70 may result in a decreased expression of transporter genes and osmoprotectants production such as sorbitol which may result in a slow adaptation to osmotic shocks [61]. Comparing Hsp production with other species, different Hsp proteins, including Hsp26, Hsp42, Hsp31, Hsp82, and Hsp12, were significantly changed in hyperosmotic stress conditions in ethanologenic yeasts such as K. marxianus and S. cerevisiae (Table 1) [68, 95].
In summary, upregulation of sigma factors in Z. mobilis under hyperosmotic stress conditions regulates the expression of heat shock proteins to degrade misfolded and unfolded proteins. Expression of sigma 54 leads to the regulation of Psp proteins, a group of regulatory proteins that control cell envelope structure. Finally, regulation of transporters and efflux proteins, carbohydrate metabolism, and bacterial cell growth are affected by the expression and activation of transcriptional regulators such as LysR, AsnC, and Xre under hyperosmotic conditions Fig. 4(b) presents all regulatory factors and proteins that contribute to the response mechanism to osmotic fermentation condition in Z. mobilis.
During bioethanol production, cells may simultaneously encounter multiple types of stress. These stressors can have antagonistic or synergistic effects on microbial cells. A study on the cellular responses of S. cerevisiae to high temperature and osmotic stress revealed that heat, acetic acid, ethanol, and oxidative stressors resulted in synergistic adverse effects, which consequently reduced the growth, ethanol production rate, and long-term thermal stability of S. cerevisiae and ethanol productivity at high temperatures [96]. Another investigation of the synergistic effect of temperature and ethanol on S. cerevisiae revealed that fermentation temperature and high ethanol concentration influenced the dynamic behavior of S. cerevisiae, leading to variations in biomass, ethanol, and glycerol production [97]. The combined effects of cold, acid, and ethanol stress on the membrane stability of Oenococcus oeni were investigated. The results indicated that ethanol and acid stress had severe effects on cell viability and increased membrane fluidity, whereas cold shock reduced the deleterious effects of these two stress conditions on the cell and maintained cell viability [98]. Moreover, changes in cell viability and membrane structure were observed in E. coli during a combination of temperature and osmotic stress [99].
In general, physiological responses in Z. mobilis during ethanol fermentation stress conditions include several changes, including changes in cell membrane structure, such as (1) changes in membrane proteins and transporters, as well as phospholipid structure to maintain membrane stability; (2) activation of the DNA repair system that alleviates the negative effects of ROS accumulation inside the cell and their influence on DNA structure; (3) activation of proteins such as heat shock proteins that degrade and refold misfolded proteins; and (4) accumulation of sugars (sorbitol) and amino acids (tryptophan) inside the cell to stabilize the cell membrane and prevent protein aggregation in the cytoplasm. In addition, the molecular mechanism of response to ethanol, lignocellulosic hydrolysate inhibitors, high temperature, and osmosis stresses includes changes in the expression of regulatory proteins, such as transcription factors and sigma factors, as well as regulatory RNAs.
Some of these responses are commonly seen under several stress conditions and can be attributed to the responses that occur under multi-stress conditions, including the accumulation of Hsp proteins such as DnaK and DnaJ, which are widespread during ethanol, lignocellulosic hydrolysate inhibitors, high temperature, and osmotic stress; the accumulation of sorbitol and tryptophan during ethanol and osmotic stress situations; and the activation of the DNA repair system during high temperature and lignocellulosic hydrolysate inhibitor stress conditions. Regulatory proteins such as sigma-32 function under both high temperature and hyperosmotic stress, sigma-54 is highly expressed under both ethanol and osmotic stress, and transcription factors such as LysR, TetR, and Psp are highly expressed during furfural, ethanol, and hyperosmotic stress. These factors are attractive elements that may react with a combination of these inhibitors during ethanol fermentation.
Impact of systems metabolic engineering on stress response and resistance in Z. mobilis
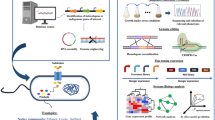
As mentioned earlier, biomass refining produces several toxic and inhibitory compounds. A combination of systems biology and synthetic biology leads to the application of targeted genetic manipulation methods such as overexpression and silencing of a specific gene, DNA shuffling, gTME, and evolutionary engineering, which are seen as a promising way to improve strains and achieve significant yields and bioethanol titers [100]. In other words, current systems metabolic engineering combined with classical metabolic engineering tools, systems biology, and synthetic biology, as depicted in Fig. 5, can acquire efficient new strains with desired characteristics [18]. In addition to toxic inhibitory compounds that can impede the production of bioethanol, other notable characteristics of Z. mobilis, such as a narrow range of substrates, competing pathways, and the detrimental impact of carbon catabolite repression (CCR) on sugar utilization, can also influence the fermentation process [101]. It is feasible to use systems metabolic engineering techniques to create more resilient Z. mobilis strains for effective biofuel production. Various genetic techniques, including forward methods (NTG, transposon mutagenesis, genome shuffling, error-prone PCR) and reverse genetics (techniques guided by omics), have been employed to enhance the intrinsic inhibitor tolerance capacity of Z. mobilis strains [37].

Systems metabolic engineering. Schematic view of systems metabolic engineering and its combination with classical metabolic engineering methods, systems biology and computational modeling, synthetic biology, and Adaptive laboratory evolution
We reviewed some of these methods and articles on this concept in the following sections.
Synthetic and systems biology
Systems biology combines biological components into a systems view and defines synthetic biology as a field in which new and artificial biological elements and pathways are constructed and designed to develop a new biological system [102]. Despite these definitions, it is worth mentioning that synthetic and systems biology are interrelated in a way that by applying knowledge of systems biology to the design of synthetic biology tools, we can in turn gain insight into systems biology [100].
One of the engineering methods that is widely used to produce recombinant strain is global transcriptional machinery engineering (gTME) that targets sigma factors and global transcription factors for strain improvement [15]. According to Lui et al., this technique designs a synthetic regulatory network with transcriptomics data which is categorized in the systems biology domain [100]. In Z. mobilis, gene manipulating using gTME methods elicits mutant strains with high ethanol and furfural stress tolerance. These mutations occurred in RpoD, Sigma70, a housekeeping gene controlling bacterial growth and promotor recognition, transcribed by RNA polymerase [15]. In ethanol stress, a random mutation in RpoD results in the production of strains that can tolerate 9% ethanol concentration compared to the wild types that can grow in the presence of 5% ethanol stress [15, 21].
Furthermore, glucose consumption in the mutant (0.64% of glucose remained) was more than control strain (5.43% of glucose remains) in the presence of 9% glucose in Z. mobilis [15]. After 24 h of incubation with 3 g/l furfural, only 22% of glucose remained in the mutant strain media, while in the control strain (non-mutant RpoD), 58% of glucose remained contact in the presence of high furfural concentration [92]. In the case of ethanol production, for ethanol and furfural mutant strains, ethanol production was about 13–14 g/l and 9.8 g/l, respectively, which was also more than the control strains [15, 92]. Mutation in sigma 70 alters ADH and PDC enzymes’ expression and activity. In the presence of 3 g/l furfural, enzyme activity changed about 3.1 to 4.6-fold compared to wild type [92].
Similarly, in the presence of 9% ethanol stress, PDC enzyme activity increased for 1.6 to 2.6 than the wild type [15]. So, it seems that the high expression of these enzymes is a kind of stress response exerted by Z. mobilis strains. Therefore, it can be concluded that tailoring the housekeeping sigma factor can alter gene expression and promotor recognition which results in obtaining strains with high tolerance to stress conditions.
It appears that the reprogramming of cellular metabolism by the gTME method is one of the best examples of the use of synthetic biology tools to alter microbes at the system level and generate highly tolerant strain. Another widely used technique for strain improvement is DNA shuffling. In a study by Wang et al., Z. mobilis strain with high tolerance to furfural and acetic acid was obtained by genome shuffling technique named Z. mobilis 532 (ZM532) [16]. Better growth rate, ethanol production, and glucose utilization in the presence of 7 g/l acetic acid and 3 g/l furfural were obtained, while these concentrations of inhibitors have negative impacts on wild type strains. In this study, genome shuffling revealed that several genes such as radA gene, which encodes DNA repair protein that controls DNA structure during stresses [103], CPSase large subunit, and arginine-tRNA ligase that controls arginine biosynthesis, increased tolerance to furfural and acetic acid stress condition [16, 103]. In addition, molecular analysis of the Z. mobilis 532 mutant revealed that genes and proteins involved in central carbon metabolism (Pgk, gpmA, glucokinase, and alcohol dehydrogenase), DNA repair system (RecF and the DNA mismatch repair enzyme) proteins, and a variety of molecular chaperones (peptidylprolyl isomerase and the Hsp20 family protein) were up-regulated, leading to tolerance in mutant strains, especially ZM532 [104]. Besides reports on overexpression of transcriptional regulators, there are also other reports on the overexpression of exogenous transcriptional factors in Z. mobilis. IrrE is a protein expressed by Deinicoccus strain in response to radiation. However, recent studies indicated that this protein can act as a regulator for recognizing the promotor region of genes such as recA (Recombinase A) and pprA (pleiotropic protein promoting DNA repair). Producing a recombinant strain that overexpresses IrrE protein can lead to tolerance to ethanol and acid and osmotic stress condition in Z. mobilis [105].
Another example of using synthetic biology tools to manipulate Z. mobilis was gene integration and transposition. Integration of Pfu-sHSP (under high-temperature stress conditions ) from Pyrococcus furious and yfdZ, metB, xylA, xylB, tktA and talB (amino acids biosynthesis and xylose utilization) from E. coli into Z. mobilis genome [106]. Accordingly, such a recombinant Z. mobilis strain can tolerate high temperatures and malnutrition. Moreover, efflux pumps seem to be another promising way to improve stress-resistant microbes [54]. In Z. mobilis, knocking out and downregulating of efflux pumps operon consists of ZMO0282 (efflux transporter, RND family, MFP subunit), ZMO0283 (efflux pump membrane transporter), and ZMO0285 (RND efflux system, outer membrane lipoprotein, NodT family) genes improves its tolerance toward furfural stress condition [54]. Furthermore, overexpression of nhaA gene that encodes hydroxylamine reductase (about 16-fold change in expression) in Acr mutant strain of Z. mobilis leads to overexpression of Na+/H+ antiporters and finally, improvement of strains that can tolerate and grow in the presence of 20 g/l acetic acids (an acidic-pH stress condition). In contrast, the wild type could tolerate only 12 g/l acetic acid in its growth environment [107, 108]. It can be concluded that the NhaA antiporter protein controls ion concentration and the balance of hemostasis inside and outside the cell membrane by ejecting H+ from the cell. Interestingly, a homolog of this protein was found in S. cerevisiae, and studies indicate the important effect of this protein in tolerance to acid stress [108]. Overexpression of genes can result in recombinant strains that tolerate a variety of stress levels. For instance, a recombinant Z. mobilis named Z mobilis R301, which overexpresses the groESL genes, can withstand multiple stress conditions, including high temperatures (40 °C), high sugar concentrations (30 g/L), and high ethanol concentrations (102.57 g/L) [109, 110]. Additionally, the overexpression of ZMO1721, a dioxygenase-encoding gene in Z. mobilis ZM4, along with NADH-dependent reductase enzymes involved in the reduction of phenolic aldehydes to phenolic alcohols, enhances the synthesis of phenolic aldehydes (4-hydroxybenzaldehyde, syringaldehyde, and vanillin), glucose consumption, and ethanol production. This system can serve as a practical synthetic biology tool to address phenolic stress conditions during bioethanol production [111]. Overexpression of some transcription factors also resulted in recombinant strains that can be active under stress conditions. In a study by Nouri et al., overexpression of transcription factors such as sigE and hfq was performed to obtain strains that can tolerate acetic acid (5 g/l) and furfural (3 g/l) under stress conditions. Comparison of the two recombinant strains revealed that recombinant strain ZM4-sigE performed better than strain ZM4-hfq in glucose utilization, ethanol production, and growth rate under furfural and acetic acid stress conditions; therefore, it appears that overexpression of regulatory factor sigE in Z. mobilis may be an effective way to produce a desirable strain [93]. Moreover, overexpression of hfq resulted in tolerance of Z. mobilis to ethanol stress, which was attributed to the downregulation of genes related to flagellar biosynthesis and heat stress response proteins. Additionally, there was an upregulation of genes related to sulfate assimilation and cysteine biosynthesis, leading to a reduction in reactive oxygen species induced during ethanol stress [90].
Recently, atmospheric and room temperature plasma (ARTP) has become a potent mutagenesis method that is quick, safe, and effective [112]. Using multiplex atmospheric and room temperature plasma (mARTP), mutants with enhanced resistance to acetic acid and low pH were generated. In the presence of high acetic acid concentrations, mutants AQ8-1 and AC8-9 demonstrated improved growth and ethanol production, adapting to acetic acid and low pH stressors by adjusting their NADH/NAD+ ratio [113].
Inhibiting competitive pathways for biofuel production, redirecting metabolic flux, and enhancing substrate utilization capability by employing CRISPR/Cas-based genome editing tools have been utilized in microorganisms to improve biofuel production [114]. Genome editing using the clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated (Cas) system has been utilized to generate recombinant Z. mobilis strains exhibiting the desired characteristics [114, 115]. The Type I-F CRISPR/Cas system, featuring Cas3 nuclease/helicase and CRISPR–Cas12a, has been successfully detected and utilized for genome editing in Z. mobilis ZM4 [116, 117]. The native Type I-F CRISPR-Cas system was employed to produce the plasmid-free mutant strain of Z. mobilis, known as ZMNP. In comparison to the wild-type strain ZM4, Z. mobilis ZMNP showed improved tolerance to inhibitors, including furfural and ethanol. Additionally, several genes and proteins crucial for ROS detoxification and the oxidative stress response were found to be increased in ZMNP. These genes and proteins include glutathione reductase, MauG, thioredoxin, glutaredoxin, and alkyl hydroperoxide reductase subunit C (AhpC). It was discovered that ZMNP’s hydrolysate tolerance was influenced by increased cysteine production (including ZMO1685 (serA1), ZMO1684 (serC), and ZMO0748 (cysK)) and the activation of stress response genes [118].
In silico genome-scale metabolic modeling and mathematical modeling are two of the most potent systems biology methods for creating and modifying microbes. Z. mobilis has been the subject of medium- and genome-scale stoichiometric reconstructions. Create microbes in such a way that the metabolic fluxes are redirected to produce more of the target products than is necessary to severely impair the behavior of the cells as a whole. Determination of intracellular fluxes based on the intricate stoichiometric connection of metabolites that make up the metabolic network is made possible by genome-scale modeling and constraint-based flux analysis. The advantage of genome-scale modeling is that it can be used in conjunction with other high-throughput methods, such as gene expression data, and provides a comprehensive prediction of the effects of genetic and environmental perturbations on cellular metabolism [119]. For instance, the genome-scale reconstructed metabolic model of Z. mobilis ZM4, ZmoMBEL601, comprises 579 metabolites and 601 metabolic reactions with physiological characteristics, including the ED pathway, incomplete pentose phosphate pathway, oxidative phosphorylation mechanisms, and high ethanol-producing ability [120]. In another experiment, genome-scale metabolic modeling (iZM363) of Z. mobilis ATCC31821 (ZM4) was performed to accurately predict the phenotypic behaviors and metabolic states in Z. mobilis. In addition, the functional role of the adh and pdc genes in the ethanologenic activity of Z. mobilis was confirmed by subsequent comparative analysis, as well as gene essentiality and flux coupling analyses [121].
A high-quality genome-scale metabolic model was used for Z. mobilis. The final model, called iZM516, contained data on three cell compartments, 1389 processes, 516 genes, and 1437 metabolites. Of all the reported GEMs of Z. mobilis, iZM516 had the highest MEMOTE score (91%), demonstrating excellent quality and accuracy was used to develop metabolic engineering techniques for biochemical production and to suggest a route for the synthesis of various biotechnological applications [122].
Adaptive laboratory evolution
Adaptive laboratory evolution (ALE) is another metabolic engineering technique that lends itself to strain evolution and optimization through the accumulation of beneficial mutations [123]. The ALE technique was performed for phenotypic properties optimization and environmental adaptation. Applying this method in Z. mobilis results in introducing two mutant strains with high capability to tolerate ethanol concentrations of more than 70–90 g/l. These two mutants were designated as ER79ag and ER79ap that both have a mutation in clpP, clpB, and spot/relA gene; however, the position of mutations in these two strains were different [124]. ClpB is an ATP-dependent chaperone that belongs to the Hsp100-family and is a member of the AAA (ATPase associated with diverse cellular activities) chaperone. These proteins, in cooperation with other chaperones, including DnaK, DnaJ, and GrpE, are responsible for protein refolding and segregation [125]. ClpP is also a protease enzyme complex with other AAA + chaperones and denaturate misfolded or unfolded proteins [126]. It seems that mutation in these two proteins can have a crucial role in improving Z. mobilis tolerance to high ethanol concentration and controlling protein quality under such stress conditions. According to an investigation on mutant strains of Z. mobilis, it seems that Z. mobilis strains with mutations in spoT/relA perform better than other strains in a medium with 70 g/l ethanol [124]. In most bacteria, including E. coli, in response to a starvation condition, a stringent response system will be activated that produces and releases a compound called (p)ppGpp. A high concentration of (p)ppGpp under stress conditions results in down-regulation of genes responsible for RNA coding, flagellar synthesis, chemotaxis response, metabolite synthesis, and transporters, while up-regulated genes such as rpoS, rpoH, and rpoE [127]. In conclusion, the spoT/ relA mutation may increase the production of (p)ppGpp and accelerate the response to high ethanol concentration [124]. In another study by Yang et al., they used the ALE technique to obtain mutants of Z. mobilis adapted to acidic and low pH conditions. In this study, two significant mutants designated as 3.6 M and 3.5 M were derived, which grew at pH 3.8 [128].
Another mutation that leads to Z. mobilis tolerance to acidic pH occurred in the ppk gene, which encodes a phosphate kinase enzyme. The activity of this enzyme release poly P structures, which according to previous studies, control protein degradation, hemostasis, expression of sigma factor S, and DNA repair system under environmental stress condition in prokaryotes [129]. Mutation and differential expression in membrane transporters were also observable in 3.6 M mutant strain which performed better under 3.8 pH conditions. These transporters, specifically RND efflux system, ABC transporters, F1/F0 ATP synthase, are responsible for the efflux of ions, different organic compounds, and protons to reduce cytoplasm acidity and control hemostasis of the cell under acidic stress conditions [128].
Another critical target for metabolite engineering to produce Z. mobilis strain tolerates stress conditions performed by manipulating the nhaA gene [107, 108, 130]. In an experiment, a combination of two NTG and ALE techniques led to the derivation of two Z. mobilis mutants (ZMA-167 and ZMA-142) that were able to grow in acetic acid stress conditions (195mM acid) [130]. Genetic analysis revealed that ZMA-167 is more tolerable to acetic acid stress conditions, and this property is related to a mutation in the terminator of the nhaA gene. This mutation leads to overexpression of Na+/H+ antiporters and hydroxylamine reductase and finally improved resistance to acetic acid stress conditions [130]. According to other reports, the methods of ALE in Z. mobilis have led to the development of two mutant strains, ZMA7-2 and ZMF3-3, which can tolerate high concentrations of acetic acid (7 g/l) and furfural (3 g/l) due to overexpression of pdc and adh under such stress conditions [17]. In conclusion, it is worth saying that the ALE method can be considered a powerful tool for improving strains with desired phenotypic characteristics.
Conclusion
Zymomonas mobilis is regarded as a suitable microorganism for bioethanol production through the PE pathway and activity of PDC and ADH enzymes. Although various inhibitors are produced during these metabolic pathways and fermentation processes, the physiological and molecular stress responses exerted by this species can result in significant adaptation. This series of actions can alter the cell interactome to overcome stress conditions. Changes in the molecular network of Z. moblis occurs at four levels of the genome, transcriptome, proteome, and metabolome during bioethanol stress. However, all four levels can directly or indirectly affect one another.
In general, under ethanol fermentation stress conditions, inhibitors such as ethanol, lignocellulosic hydrolysates inhibitors, high temperature, and osmotic stress resulted in a series of changes in gene expression and transcriptome in Z. mobilis and alterations in proteomes and metabolomes. Up-regulation and down-reglation of proteins including transcription factors like Lrp/AsnC family, Xre family, LysR family, LytR family, TetR family, MarR family, RpiR family, Psp family, GntR family, and LacI, as well as sigma factors such as sigma-D, sigma-N, sigma-H, sigma-F, and sigma-E are among these changes.
In addition, the expression of a variety of proteins, such as the enzymes responsible for bioethanol production, especially PDC and ADH; proteins that are responsible for controlling folded and misfolded proteins and preventing their aggregation, such as GroEL/GroES, Hsp, Lon, DnaJ, and DnaK; and proteins that activate the DNA repair system during stress conditions, such as RecA, Xse, RadA, and RadC, are influenced by these stressful environmental conditions.
Finally, alterations in the expression and repression of genes result in changes in the metabolome, specifically the contents of sugars (sorbitol), amino acids (tryptophan), and lipid membrane composition (mainly changes in lipid membrane content and structure during fermentation stress conditions that result in membrane stabilization).
Advancements in omics approaches, including genomic, transcriptomic, proteomic, metabolomics, and fluxomics, as well as computer modeling, help identify crucial components of the cell and efficient regulatory networks that facilitate stress responses. Using these data, in conjunction with the application of genetic engineering methods and the design of metabolic pathways, strains that are capable of producing ethanol and are resistant to stresses encountered during the fermentation process can be identified. Employing ALE and synthetic biology methods for Z. mobilis has successfully developed resistant strains. However, further investigations are required in this area. For example, future investigations could explore the application of systems metabolic engineering methods in small RNA (sRNA), the application of new genome editing tools such as CRISPR-Cas9, and the implementation of genome-scale metabolic modeling (GEM), which simulates metabolic fluxes in silico using algorithms like flux balance analysis (FBA), could be performed for developing recombinant Z. mobilis strains. Ultimately, comprehending the processes underlying stress response and the roles played by genes and proteins in this process aids in identifying possible targets for future metabolic engineering and systems biology research, as well as offering perspectives on the application of this bacterium in biotechnology.
Data availability
All data are included in the manuscript and additional information, and further queries about sharing data can be directed to the corresponding author.
Abbreviations
- GHG:
-
Green House Gases
- EMP:
-
Embden-Meyerhof-Parnas
- ED:
-
Entner–Doudoroff pathway
- GFOR:
-
Glucose-Fructose-Oxidoreductase
- HMF:
-
Hydroxy methyl furfural
- ABC:
-
Adenine triphosphate binding cassette
- RND:
-
Resistance-Nodulation Cell-Division
- MFS:
-
Major Facilitator Superfamily
- PPP:
-
Pentose-Phosphate-Pathway
- TCA:
-
Tri-Carboxylic Acid cycle
- ALD:
-
Aldehyde dehydrogenase
- ADH:
-
Alcohol dehydrogenase
- gTME:
-
global Transcription Machinery Engineering
- ALE:
-
Adaptive Laboratory Evolution
- PE:
-
Pyruvate to Ethanol pathway
- PDC:
-
pyruvate decarboxylase
- TBDT:
-
TonB- dependent transporter
- GPD:
-
Glycerol-3-phophate dehydrogenase
- TFBS:
-
Transcription Factor Binding Site
- Lrp:
-
Leucine-responsive regulatory protein
- Dam:
-
Deoxy adenosine methylase
- TFTR:
-
TetR-Family Transcription Regulator
- AAA:
-
ATPase association with diverse cellular activities
- CCR:
-
Carbon Catabolite Repression
- TAF:
-
Transcription Associated Factor
References
Lin Y, Tanaka S. Ethanol fermentation from biomass resources: current state and prospects. Appl Microbiol Biotechnol. 2006;69:627–42.
Ajit A, Sulaiman AZ, Chisti Y. Production of bioethanol by Zymomonas mobilis in high-gravity extractive fermentations. Food Bioprod Process. 2017;102:123–35.
Khambhaty Y, Upadhyay D, Kriplani Y, Joshi N, Mody K, Gandhi MR. Bioethanol from Macroalgal Biomass: utilization of Marine yeast for production of the same. Bioenergy Res. 2013;6:188–95.
Hu N, Yuan B, Sun J, Wang S-A, Li F-L. Thermotolerant Kluyveromyces marxianus and Saccharomyces cerevisiae strains representing potentials for bioethanol production from Jerusalem artichoke by consolidated bioprocessing. Appl Microbiol Biotechnol. 2012;95:1359–68.
Agbogbo F, Coward-Kelly G. Cellulosic ethanol production using the naturally occurring xylose-fermenting yeast, Pichia stipitis. Biotechnol Lett. 2008;30:1515–24.
Zhou S, Yomano LP, Shanmugam KT, Ingram LO. Fermentation of 10% (w/v) sugar to D(-)-lactate by engineered Escherichia coli B. Biotechnol Lett. 2005;27:1891–6.
Yang S, Fei Q, Zhang Y, Contreras LM, Utturkar SM, Brown SD, et al. Zymomonas mobilis as a model system for production of biofuels and biochemicals. Microb Biotechnol. 2016;9:699–717.
Almeida JRM, Bertilsson M, Gorwa-Grauslund MF, Gorsich S, Lidén G. Metabolic effects of furaldehydes and impacts on biotechnological processes. Appl Microbiol Biotechnol. 2009;82:625–38.
Pérez-Gallardo RV, Briones LS, Díaz-Pérez AL, Gutiérrez S, Rodríguez-Zavala JS, Campos-García J. Reactive oxygen species production induced by ethanol in Saccharomyces cerevisiae increases because of a dysfunctional mitochondrial iron – sulfur cluster assembly system. FEMS Yeast Res. 2013;13:804–19.
Bai FW, Anderson WA, Moo-Young M. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnol Adv. 2008;26:89–105.
Guan N, Li J, Shin H, dong, Du G, Chen J, Liu L. Microbial response to environmental stresses: from fundamental mechanisms to practical applications. Appl Microbiol Biotechnol Appl Microbiol Biotechnol. 2017;101:3991–4008.
Denich TJ, Beaudette LA, Lee H, Trevors JT. Effect of selected environmental and physico-chemical factors on bacterial cytoplasmic membranes. J Microbiol Methods. 2003;52:149–82.
Hu XH, Wang MH, Tan T, Li JR, Yang H, Leach L, et al. Genetic dissection of ethanol tolerance in the budding yeast Saccharomyces cerevisiae. Genetics. 2007;175:1479–87.
Sarkar P, Mukherjee M, Goswami G, Das D. Adaptive laboratory evolution induced novel mutations in Zymomonas mobilis ATCC ZW658: a potential platform for co-utilization of glucose and xylose. J Ind Microbiol Biotechnol. 2020;47:329–41.
Tan F, Wu B, Dai L, Qin H, Shui Z, Wang J, et al. Using global transcription machinery engineering (gTME) to improve ethanol tolerance of Zymomonas mobilis. Microb Cell Fact. 2016;15:1–9.
Wang W, Wu B, Qin H, Liu P, Qin Y, Duan G, et al. Genome shuffling enhances stress tolerance of Zymomonas mobilis to two inhibitors. Biotechnol Biofuels. 2019;12:1–12.
Shui ZX, Qin H, Wu B, Ruan Z, yong, Wang Lshang, Tan FR, et al. Adaptive laboratory evolution of ethanologenic Zymomonas mobilis strain tolerant to furfural and acetic acid inhibitors. Appl Microbiol Biotechnol. 2015;99:5739–48.
Ko YS, Kim JW, Lee JA, Han T, Kim GB, Park JE, et al. Tools and strategies of systems metabolic engineering for the development of microbial cell factories for chemical production. Chem Soc Rev Royal Soc Chem. 2020;49:4615–36.
Ma M, Liu ZL. Mechanisms of ethanol tolerance in Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2010;87:829–45.
Yang S, Pan C, Tschaplinski TJ, Hurst GB, Engle NL, Zhou W, et al. Systems Biology Analysis of Zymomonas mobilis ZM4 ethanol stress responses. PLoS ONE. 2013;8:e68886.
He, Wu B, Shui ZX, Hu QC, Wang WG, Tan FR, et al. Transcriptome profiling of Zymomonas mobilis under ethanol stress. Biotechnol Biofuels. 2012;5:1–10.
Henderson CM, Block DE. Examining the role of membrane lipid composition in determining the ethanol tolerance of Saccharomyces cerevisiae. Appl Environ Microbiol. 2014;80:2966–72.
Dombek KM, Ingram LO. Effects of ethanol on the Escherichia coli plasma membrane. J Bacteriol. 1984;157:233–9.
Huffer S, Clark ME, Ning JC, Blanch HW, Clark DS. Role of alcohols in growth, lipid composition, and membrane fluidity of yeasts, bacteria, and archaea. Appl Environ Microbiol. 2011;77:6400–8.
Li R, Xiong G, Yuan S, Wu Z, Miao Y, Weng P. Investigating the underlying mechanism of Saccharomyces cerevisiae in response to ethanol stress employing RNA-seq analysis. World J Microbiol Biotechnol. 2017;33:1–13.
Diniz RHS, Villada JC, Alvim MCT, Vidigal PMP, Vieira NM, Lamas-Maceiras M, et al. Transcriptome analysis of the thermotolerant yeast Kluyveromyces marxianus CCT 7735 under ethanol stress. Appl Microbiol Biotechnol. 2017;101:6969–80.
Brenac L, Baidoo EEK, Keasling JD, Budin I. Distinct functional roles for hopanoid composition in the chemical tolerance of Zymomonas mobilis. Mol Microbiol. 2019;112:1564–75.
Ma M, Han P, Zhang R, Li H. Ultrastructural changes of Saccharomyces cerevisiae in response to ethanol stress. Can J Microbiol. 2013;59:589–97.
Ingram LO, Vreeland NS. Differential effects of ethanol and hexanol on the Escherichia coli cell envelope. J Bacteriol. 1980;144:481–8.
Loos H, Kramer R, Sahm H, Sprenger GA. Sorbitol promotes growth of Zymomonas mobilis in environments with high concentrations of sugar: evidence for a physiological function of glucose- fructose oxidoreductase in osmoprotection. J Bacteriol. 1994;176:7688–93.
Gomes FCO, Pataro C, Guerra JB, Neves MJ, Corrêa SR, Moreira ESA, et al. Physiological diversity and trehalose accumulation in Schizosaccharomyces Pombe strains isolated from spontaneous fermentations during the production of the artisanal Brazilian cachaça. Can J Microbiol. 2002;48:399–406.
Takagi H. Proline as a stress protectant in yeast: physiological functions, metabolic regulations, and biotechnological applications. Appl Microbiol Biotechnol. 2008;81:211–23.
Hirasawa T, Yoshikawa K, Nakakura Y, Nagahisa K, Furusawa C, Katakura Y, et al. Identification of target genes conferring ethanol stress tolerance to Saccharomyces cerevisiae based on DNA microarray data analysis. J Biotechnol. 2007;131:34–44.
Cho SH, Lei R, Henninger TD, Contreras LM. Discovery of ethanol-responsive small RNAs in Zymomonas mobilis. Appl Environ Microbiol. 2014;80:4189–98.
Bhatia SK, Jagtap SS, Bedekar AA, Bhatia RK, Patel AK, Pant D, et al. Recent developments in pretreatment technologies on lignocellulosic biomass: Effect of key parameters, technological improvements, and challenges. Bioresour Technol. 2020;300:122–724.
He M, Wu B, Shui Z, Hu Q, Wang W, Tan F, et al. Transcriptome profiling of Zymomonas mobilis under furfural stress. Appl Microbiol Biotechnol. 2012;95:189–99.
Yang Y, Hu M, Tang Y, Geng B, Qiu M, He Q, et al. Progress and perspective on lignocellulosic hydrolysate inhibitor tolerance improvement in Zymomonas mobilis. Bioresour Bioprocess. 2018;5:1–12.
Allen SA, Clark W, McCaffery JM, Cai Z, Lanctot A, Slininger PJ, et al. Furfural induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisiae. Biotechnol Biofuels. 2010;3:1–10.
Chang D, Yu Z, Islam ZU, French WT, Zhang Y, Zhang H. Proteomic and metabolomic analysis of the cellular biomarkers related to inhibitors tolerance in Zymomonas mobilis ZM4. Biotechnol Biofuels. 2018;11:1–23.
Zhu Y, Wu L, Zhu J, Xu Y, Yu S. Transcriptome and metabolome analysis of Pichia stipitis to three representative lignocellulosic inhibitors. Arch Microbiol. 2019;201:581–9.
Yi X, Gu H, Gao Q, Liu ZL, Bao J. Transcriptome analysis of Zymomonas mobilis ZM4 reveals mechanisms of tolerance and detoxification of phenolic aldehyde inhibitors from lignocellulose pretreatment. Biotechnol Biofuels BioMed Cent. 2015;8:1–15.
Cao W, Zhao W, Yang B, Wang X, Shen Y, Wei T, et al. Proteomic analysis revealed the roles of YRR1 deletion in enhancing the vanillin resistance of Saccharomyces cerevisiae. Microb Cell Fact. 2021;20:1–17.
Ishida Y, Nguyen TTM, Izawa S. The yeast ADH7 promoter enables gene expression under pronounced translation repression caused by the combined stress of vanillin, furfural, and 5-hydroxymethylfurfural. J Biotechnol. 2017;252:65–72.
Samappito J, Yamada M, Klanrit P, Thanonkeo P. Characterization of a thermo-adapted strain of Zymomonas mobilis for ethanol production at high temperature. 3 Biotech. 2018;8:1–9.
Zhu Y, Wu L, Zhu J, Xu Y, Yu S. Quantitative proteomic analysis of xylose fermentation strain Pichia stipitis CBS 5776 to lignocellulosic inhibitors acetic acid, vanillin and 5-hydroxymethylfurfural. FEMS Microbiol Lett. 2018;365:1–8.
Yang S, Pan C, Hurst GB, Dice L, Davison BH, Brown SD. Elucidation of Zymomonas mobilis physiology and stress responses by quantitative proteomics and transcriptomics. Front Microbiol. 2014;5:246.
Skerker JM, Leon D, Price MN, Mar JS, Tarjan DR, Wetmore KM, et al. Dissecting a complex chemical stress: chemogenomic profiling of plant hydrolysates. Mol Syst Biol. 2013;9:674.
Yang S, Franden MA, Wang X, Chou YC, Hu Y, Brown SD, et al. Transcriptomic profiles of Zymomonas mobilis 8b to Furfural Acute and Long-Term stress in both glucose and xylose conditions. Front Microbiol. 2020;11:13.
Miller EN, Jarboe LR, Turner PC, Pharkya P, Yomano LP, York SW, et al. Furfural inhibits growth by limiting sulfur assimilation in ethanologenic Escherichia coli strain LY180. Appl Environ Microbiol. 2009;75:6132–41.
Yan X, Wang X, Yang Y, Wang Z, Zhang H, Li Y, et al. Cysteine supplementation enhanced inhibitor tolerance of Zymomonas mobilis for economic lignocellulosic bioethanol production. Bioresour Technol. 2022;349:126878.
Franden MA, Pienkos PT, Zhang M. Development of a high-throughput method to evaluate the impact of inhibitory compounds from lignocellulosic hydrolysates on the growth of Zymomonas mobilis. J Biotechnol. 2009;144:259–67.
Charoensuk K, Sakurada T, Tokiyama A, Murata M, Kosaka T, Thanonkeo P, et al. Thermotolerant genes essential for survival at a critical high temperature in thermotolerant ethanologenic Zymomonas mobilis TISTR 548. Biotechnol Biofuels BioMed Cent. 2017;10:1–11.
Hoshida H, Akada R. High-temperature Bioethanol Fermentation by Conventional and Nonconventional yeasts. In: Sibirny A, editor. Biotechnology of yeasts and Filamentous Fungi. Springer; 2017. pp. 39–61.
Birch RM, Walker GM. Influence of magnesium ions on heat shock and ethanol stress responses of Saccharomyces cerevisiae. Enzyme Microb Technol. 2000;26:678–87.
Caspeta L, Castillo T, Nielsen J. Modifying yeast tolerance to inhibitory conditions of ethanol production processes. Front Bioeng Biotechnol. 2015;3:184.
Kim IS, Kim YS, Kim H, Jin I, Yoon HS. Saccharomyces cerevisiae KNU5377 stress response during high-temperature ethanol fermentation. Mol Cells. 2013;35:210–8.
Lazzaroni JC, Germon P, Ray MC, Vianney A. The tol proteins of Escherichia coli and their involvement in the uptake of biomolecules and outer membrane stability. FEMS Microbiol Lett. 1999;177:191–7.
Noor R, Murata M, Yamada M. Oxidative stress as a trigger for growth phase-specific σ-dependent cell lysis in Escherichia coli. J Mol Microbiol Biotechnol. 2009;17:177–87.
Charoensuk K, Irie A, Lertwattanasakul N, Sootsuwan K, Thanonkeo P, Yamada M. Physiological importance of cytochrome c peroxidase in ethanologenic thermotolerant Zymomonas mobilis. J Mol Microbiol Biotechnol. 2011;20:70–82.
Chamnipa N, Thanonkeo S, Klanrit P, Thanonkeo P. The potential of the newly isolated thermotolerant yeast Pichia kudriavzevii RZ8-1 for high-temperature ethanol production. Brazilian J Microbiol. 2018;49:378–91.
Benoliel T, Rubini MR, de Souza Baptistello C, Janner CR, Vieira VR, Torres FA, et al. Physiological effects of overexpressed sigma factors on fermentative stress response of Zymomonas mobilis. Brazilian J Microbiol. 2020;51:65–75.
Matsushita K, Azuma Y, Kosaka T, Yakushi T, Hoshida H, Akada R, et al. Genomic analyses of thermotolerant microorganisms used for high-temperature fermentations. Biosci Biotechnol Biochem. 2016;80:655–68.
Chang YH, Chang KS, Chen CY, Hsu CL, Chang TC, Jang H, Der. Enhancement of the efficiency of bioethanol production by Saccharomyces cerevisiae via gradually batch-wise and fed-batch increasing the glucose concentration. Fermentation. 2018;4:45.
Bremer E, Krämer R. Responses of microorganisms to osmotic stress. Annu Rev Microbiol. 2019;73:313–34.
Zhang K, Shao H, Cao Q, He M, Wu B, Feng H. Transcriptional analysis of adaptation to high glucose concentrations in Zymomonas mobilis. Appl Microbiol Biotechnol. 2015;99:2009–22.
Noinaj N, Guillier M, Barnard TJ, Buchanan SK. TonB-dependent transporters: regulation, structure, and function. Annu Rev Microbiol. 2010;64:43–60.
Kitichantaropas Y, Boonchird C, Sugiyama M, Kaneko Y, Harashima S, Auesukaree C. Cellular mechanisms contributing to multiple stress tolerance in Saccharomyces cerevisiae strains with potential use in high-temperature ethanol fermentation. AMB Express. 2016;6:1–14.
Gao J, Yuan W, Li Y, Xiang R, Hou S, Zhong S, et al. Transcriptional analysis of Kluyveromyces marxianus for ethanol production from inulin using consolidated bioprocessing technology. Biotechnol Biofuels. 2015;8:1–17.
Sootsuwan K, Thanonkeo P, Keeratirakha N, Thanonkeo S, Jaisil P, Yamada M. Sorbitol required for cell growth and ethanol production by Zymomonas mobilis under heat, ethanol, and osmotic stresses. Biotechnol Biofuels. 2013;6:1–13.
De Barros M, Celligoi MAPC. Synthesis of sorbitol by Zymomonas mobilis under high osmotic pressure. Brazilian J Microbiol. 2006;37:324–8.
Albertyn J, Hohmann S, Thevelein JM, Prior BA. GPD1, which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol Cell Biol. 1994;14:4135–44.
Fu Y, Chen L, Zhang W. Regulatory mechanisms related to biofuel tolerance in producing microbes. J Appl Microbiol. 2016;121:320–32.
Seo JS, Chong H, Park HS, Yoon KO, Jung C, Kim JJ, et al. The genome sequence of the ethanologenic bacterium Zymomonas mobilis ZM4. Nat Biotechnol. 2005;23:63–8.
Guo Z, Olsson L. Physiological response of Saccharomyces cerevisiae to weak acids present in lignocellulosic hydrolysate. FEMS Yeast Res. 2014;14:1234–48.
Feklístov A, Sharon BD, Darst SA, Gross CA. Bacterial sigma factors: a historical, structural, and genomic perspective. Annu Rev Microbiol. 2014;68:357–76.
Chung HJ, Bang W, Drake MA. Stress response of Escherichia coli. Compr Rev inFood Sci Food Saf. 2006;5:52–64.
Gottesman S, McCullen CA, Guillier M, Vanderpool CK, Majdalani N, Benhammou J, et al. Small RNA regulators and the bacterial response to stress. Cold Spring Harb Symp Quant Biol. 2006;71:1–11.
Storz G, Vogel J, Wassarman KM. Regulation by small RNAs in Bacteria: expanding frontiers. Mol Cell. 2011;43:880–91.
Cho SH, Haning K, Shen W, Blome C, Li R, Yang S, et al. Identification and characterization of 5’ untranslated regions (5’UTRs) in Zymomonas mobilis as regulatory biological parts. Front Microbiol. 2017;8:2432.
Deng W, Li C, Xie J. The underling mechanism of bacterial TetR/AcrR family transcriptional repressors. Cell Signal. 2013;25:1608–13.
Maddocks SE, Oyston PCF. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology. 2008;154:3609–23.
Cho BK, Barrett CL, Knight EM, Park YS, Palsson B. Genome-scale reconstruction of the Lrp regulatory network in Escherichia coli. Proc Natl Acad Sci. 2008;105:19462–7.
Arndt K, Fink GR. GCN4 protein, a positive transcription factor in yeast, binds general control promoters at all 5’ TGACTC 3’ sequences. Proc Natl Acad Sci. 1986;83:8516–20.
Roncarati D, Scarlato V. Regulation of heat-shock genes in bacteria: from signal sensing to gene expression output. FEMS Microbiol Rev. 2017;41:549–74.
Thanonkeo P, Sootsuwan K, Leelavacharamas V, Yamada M. Cloning and transcriptional analysis of groES and groEL in ethanol-producing bacterium Zymomonas mobilis TISTR 548. Pakistan J Biol Sci. 2007;10:13–22.
Li P, Fu X, Chen M, Zhang L, Li S. Proteomic profiling and integrated analysis with transcriptomic data bring new insights in the stress responses of Kluyveromyces marxianus after an arrest during high-temperature ethanol fermentation. Biotechnol Biofuels. 2019;12:1–13.
Stanley D, Bandara A, Fraser S, Chambers PJ, Stanley GA. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J Appl Microbiol. 2010;109:13–24.
Han R, Haning K, Gonzalez-rivera JC, Yang Y, Li R, Cho SH, et al. Multiple small RNAs interact to co-regulate ethanol tolerance in Zymomonas mobilis. Front Bioeng Biotechnol. 2020;8:155.
Levin DE. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: the cell wall integrity signaling pathway. Genetics. 2011;189:1145–75.
Tang Y, Wang Y, Yang Q, Zhang Y, Wu Y, Yang Y, et al. Molecular mechanism of enhanced ethanol tolerance associated with hfq overexpression in Zymomonas mobilis. Front Bioeng Biotechnol. 2022;10:1098021.
Yang S, Pelletier DA, Lu TS, Brown SD. The Zymomonas mobilis regulator hfq contributes to tolerance against multiple lignocellulosic pretreatment inhibitors. BMC Microbiol. 2010;10:1–11.
Tan FR, Dai LC, Wu B, Qin H, Shui ZX, Wang JL, et al. Improving furfural tolerance of Zymomonas mobilis by rewiring a sigma factor RpoD protein. Appl Microbiol Biotechnol. 2015;99:5363–71.
Nouri H, Moghimi H, Marashi SA, Elahi E. Impact of hfq and sigE on the tolerance of Zymomonas mobilis ZM4 to furfural and acetic acid stresses. PLoS ONE. 2020;15:e0240330.
Chang Z. The function of the DegP (HtrA) protein: protease versus chaperone. IUBMB Life. 2016;68:904–7.
Saini P, Beniwal A, Vij S. Physiological response of Kluyveromyces marxianus during oxidative and osmotic stress. Process Biochem. 2017;56:21–9.
Woo J, Yang K, Kim S, Blank LM, Park J. High temperature stimulates acetic acid accumulation and enhances the growth inhibition and ethanol production by Saccharomyces cerevisiae under fermenting conditions. Appl Microbiol Biotechnol. 2014;98:6085–94.
Aldiguier AS, Alfenore S, Cameleyre X, Goma G, Uribelarrea JL, Guillouet SE, et al. Synergistic temperature and ethanol effect on Saccharomyces cerevisiae dynamic behaviour in ethanol bio-fuel production. Bioprocess Biosyst Eng. 2004;26:217–22.
Chu-ky S, Tourdot-Marechal R, Marechal P-A, Guzzo J. Combined cold, acid, ethanol shocks in Oenococcus oeni: effects on membrane fluidity and cell viability. Biochim Biophys Acta - Biomembr. 2005;1717:118–24.
Mille Y, Beney L, Gervais P. Viability of Escherichia coli after combined osmotic and thermal treatment: a plasma membrane implication. Biochim Biophys Acta - Biomembr. 2002;1567:41–8.
Liu D, Hoynes-O’Connor A, Zhang F. Bridging the gap between systems biology and synthetic biology. Front Microbiol. 2013;4:55718.
Zhang K, Lu X, Li Y, Jiang X, Liu L, Wang H. New technologies provide more metabolic engineering strategies for bioethanol production in Zymomonas mobilis. Appl Microbiol Biotechnol. 2019;103:2087–99.
Xu X, Liu Y, Du G, Liu L. Systems biology, synthetic biology, and metabolic engineering. In: Long L, Guocheng D, Liu Y, editors. Systems and synthetic metabolic engineering. Elsevier, Amnsterdam; 2020. pp. 1–31.
Charlier D, Le Nguyen P, Roovers M. Regulation of carbamoylphosphate synthesis in Escherichia coli: an amazing metabolite at the crossroad of arginine and pyrimidine biosynthesis. Amino Acids. 2018;50:1647–61.
Shabbir S, Wang W, Nawaz M, Boruah P, Kulyar MFe, Chen A. Molecular mechanism of engineered Zymomonas mobilis to furfural and acetic acid stress. Microb Cell Fact. 2023;22:88.
Zhang Y, Ma R, Zhao Z, Zhou Z, Lu W, Zhang W, et al. irrE, an exogenous gene from deinococcus radiodurans, improves the growth of and ethanol production by a Zymomonas mobilis strain under ethanol and acid stresses. J Microbiol Biotechnol. 2010;20:1156–62.
Zhang X, Wang T, Zhou W, Jia X, Wang H. Use of a Tn5-based transposon system to create a cost-effective Zymomonas mobilis for ethanol production from lignocelluloses. Microb Cell Fact. 2013;12:1–12.
You KM, Rosenfield CL, Knipple DC. Ethanol tolerance in the yeast Saccharomyces cerevisiae is Dependent on Cellular Oleic Acid Content. Appl Environ Microbiol. 2003;69:1499–503.
Yang S, Land ML, Klingeman DM, Pelletier DA, Lu TYS, Martin SL, et al. Paradigm for industrial strain improvement identifies sodium acetate tolerance loci in Zymomonas mobilis and Saccharomyces cerevisiae. Proc Natl Acad Sci. 2010;107:10395–400.
Charoenpunthuwong K, Klanrit P, Chamnipa N, Thanonkeo S, Yamada M, Thanonkeo P. Optimization Condition for Ethanol Production from Sweet Sorghum Juice by recombinant Zymomonas mobilis overexpressing groESL genes. Energies. 2023;16:5284.
Kaewchana A, Techaparin A, Boonchot N, Thanonkeo P, Klanrit P. Improved high–temperature ethanol production from sweet sorghum juice using Zymomonas mobilis overexpressing groESL genes. Appl Microbiol Biotechnol. 2021;105:9419–31.
Yi X, Mei J, Lin L, Wang W. Overexpression of Dioxygenase Encoding Gene accelerates the Phenolic Aldehyde Conversion and ethanol fermentability of Zymomonas mobilis. Appl Biochem Biotechnol. 2021;193:3017–27.
Zhang X, Zhang C, Zhou QQ, Zhang XF, Wang LY, Chang HB, et al. Quantitative evaluation of DNA damage and mutation rate by atmospheric and room-temperature plasma (ARTP) and conventional mutagenesis. Appl Microbiol Biotechnol. 2015;99:5639–46.
Wu B, Qin H, Yang Y, Duan G, Yang S, Xin F, et al. Engineered Zymomonas mobilis tolerant to acetic acid and low pH via multiplex atmospheric and room temperature plasma mutagenesis. Biotechnol Biofuels. 2019;12:1–13.
Shanmugam S, Ngo H, Wu Y, Advanced CRISPR. Cas-based Genome Editing Tools for Microbial Biofuels production: a review. Renew Energy. 2020;149:1107–19.
Cao QH, Shao HH, Qiu H, Li T, Zhang YZ, Tan XM. Using the CRISPR/Cas9 system to eliminate native plasmids of Zymomonas mobilis ZM4. Biosci Biotechnol Biochem. 2017;81:453–9.
Zheng Y, Han J, Wang B, Hu X, Li R, Shen W, et al. Characterization and repurposing of the endogenous type I-F CRISPR – cas system of Zymomonas mobilis for genome engineering. Nucleic Acids Res. 2019;47:11461–75.
Shen W, Zhang J, Geng B, Qiu M, Hu M, Yang Q, et al. Establishment and application of a CRISPR–Cas12a assisted genome–editing system in Zymomonas mobilis. Microb Cell Fact. 2019;18:1–11.
Geng B, Liu S, Chen Y, Wu Y, Wang Y, Zhou X, et al. A plasmid-free Zymomonas mobilis mutant strain reducing reactive oxygen species for efficient bioethanol production using industrial effluent of xylose mother liquor. Front Bioeng Biotechnol. 2022;10:1110513.
Kalnenieks U, Pentjuss A, Rutkis R, Stalidzans E, Fell DA. Modeling of Zymomonas mobilis central metabolism for novel metabolic engineering strategies. Front Microbiol. 2014;5:75668.
Lee KY, Park JM, Kim TY, Yun H, Lee SY. The genome-scale metabolic network analysis of Zymomonas mobilis ZM4 explains physiological features and suggests ethanol and succinic acid production strategies. Microb Cell Fact. 2010;9:1–12.
Widiastuti H, Kim JY, Selvarasu S, Karimi IA, Kim H, Seo JS, et al. Genome-scale modeling and in silico analysis of ethanologenic bacteria Zymomonas Mobilis. Biotechnol Bioeng. 2011;108:655–65.
Wu Y, Yuan Q, Yang Y, Liu D, Yang S, Ma H. Construction and application of high-quality genome-scale metabolic model of Zymomonas mobilis to guide rational design of microbial cell factories. Synth Syst Biotechnol. 2023;8:498–508.
Portnoy VA, Bezdan D, Zengler K. Adaptive laboratory evolution — harnessing the power of biology for metabolic engineering. Curr Opin Biotechnol. 2011;22:590–4.
Carreón-Rodríguez OE, Gutiérrez-Ríos RM, Acosta JL, Martinez A, Cevallos MA. Phenotypic and genomic analysis of Zymomonas mobilis ZM4 mutants with enhanced ethanol tolerance. Biotechnol Rep. 2019;23:e00328.
Sangpuii L, Dixit SK, Kumawat M, Apoorva S, Kumar M, Kappala D, et al. Comparative roles of clpA and clpB in the survival of S. Typhimurium under stress and virulence in poultry. Sci Rep. 2018;8:4481.
Moreno-Cinos C, Goossens K, Salado IG, Van Der Veken P, De Winter H, Augustyns K. ClpP protease, a promising antimicrobial target. Int J Mol Sci. 2019;20:2232.
Gaca AO, Colomer-Winter C, Lemos JA. Many means to a common end: the intricacies of (p)ppGpp metabolism and its control of bacterial homeostasis. J Bacteriol. 2015;197:1146–56.
Yang Q, Yang Y, Tang Y, Wang X, Chen Y, Shen W, et al. Development and characterization of acidic-pH-tolerant mutants of Zymomonas mobilis through adaptation and next-generation sequencing-based genome resequencing and RNA-Seq. Biotechnol Biofuels. 2020;13:1–17.
Seufferheld MJ, Alvarez HM, Farias ME. Role of polyphosphates in microbial adaptation to extreme environments. Appl Environ Microbiol. 2008;74:5867–74.
Liu YF, Hsieh CW, Chang YS, Wung BS. Effect of acetic acid on ethanol production by Zymomonas mobilis mutant strains through continuous adaptation. BMC Biotechnol. 2017;17:1–10.
Si M, Chen C, Zhong J, Li X, Liu Y, Su T, et al. MsrR is a thiol-based oxidation-sensing regulator of the XRE family that modulates C. Glutamicum oxidative stress resistance. Microb Cell Fact. 2020;19:1–14.
Deochand DK, Grove A. MarR family transcription factors: dynamic variations on a common scaffold. Crit Rev Biochem Mol Biol. 2017;52:595–613.
Perera IC, Grove A. Molecular mechanisms of ligand-mediated attenuation of DNA binding by MarR family transcriptional regulators. J Mol Cell Biol. 2010;2:243–54.
Zhu Y, Nandakumar R, Sadykov MR, Madayiputhiya N, Luong TT, Gaupp R, et al. RpiR homologues may link Staphylococcus aureus RNAIII synthesis and pentose phosphate pathway regulation. J Bacteriol. 2011;193:6187–96.
Suvorova IA, Korostelev YD, Gelfand MS. GntR family of bacterial transcription factors and their DNA binding motifs: structure, positioning and co-evolution. PLoS ONE. 2015;10:e0132618.
Jovanovic G, Engl C, Mayhew AJ, Burrows PC, Buck M. Properties of the phage-shock-protein (psp) regulatory complex that govern signal transduction and induction of the psp response in Escherichia coli. Microbiology. 2010;156:2920–32.
Ravcheev DA, Khoroshkin MS, Laikova ON, Tsoy OV, Sernova NV, Petrova SA, et al. Comparative genomics and evolution of regulons of the LacI-family transcription factors. Front Microbiol. 2014;5:294.
Sharma-Kuinkel BK, Mann EE, Ahn JS, Kuechenmeister LJ, Dunman PM, Bayles KW. The Staphylococcus aureus LytSR two-component regulatory system affects biofilm formation. J Bacteriol. 2009;191:4767–75.
Acknowledgements
Not applicable.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
S.A and H.N wrote the manuscript. S.A, H.N and G.P edit the manuscript. H.M and H.N supervised the project and revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Ethics approval is not applicable as this article does not describe any studies involving human participants or animals.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Asefi, S., Nouri, H., Pourmohammadi, G. et al. Comprehensive network of stress-induced responses in Zymomonas mobilis during bioethanol production: from physiological and molecular responses to the effects of system metabolic engineering. Microb Cell Fact 23, 180 (2024). https://doi.org/10.1186/s12934-024-02459-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-024-02459-1