
Abstract
The SCUBE [Signal peptide-Complement C1r/C1s, Uegf, Bmp1 (CUB)-Epithelial growth factor domain-containing protein] family consists of three proteins in vertebrates, SCUBE1, 2 and 3, which are highly conserved in zebrafish, mice and humans. Each SCUBE gene encodes a polypeptide of approximately 1000 amino acids that is organized into five modular domains: (1) an N-terminal signal peptide sequence, (2) nine tandem epidermal growth factor (EGF)-like repeats, (3) a large spacer region, (4) three cysteine-rich (CR) motifs, and (5) a CUB domain at the C-terminus. Murine Scube genes are expressed individually or in combination during the development of various tissues, including those in the central nervous system and the axial skeleton. The cDNAs of human SCUBE orthologs were originally cloned from vascular endothelial cells, but SCUBE expression has also been found in platelets, mammary ductal epithelium and osteoblasts. Both soluble and membrane-associated SCUBEs have been shown to play important roles in physiology and pathology. For instance, upregulation of SCUBEs has been reported in acute myeloid leukemia, breast cancer and lung cancer. In addition, soluble SCUBE1 is released from activated platelets and can be used as a clinical biomarker for acute coronary syndrome and ischemic stroke. Soluble SCUBE2 enhances distal signaling by facilitating the secretion of dual-lipidated hedgehog from nearby ligand-producing cells in a paracrine manner. Interestingly, the spacer regions and CR motifs can increase or enable SCUBE binding to cell surfaces via electrostatic or glycan-lectin interactions. As such, membrane-associated SCUBEs can function as coreceptors that enhance the signaling activity of various serine/threonine kinase or tyrosine kinase receptors. For example, membrane-associated SCUBE3 functions as a coreceptor that promotes signaling in bone morphogenesis. In humans, SCUBE3 mutations are linked to abnormalities in growth and differentiation of both bones and teeth. In addition to studies on human SCUBE function, experimental results from genetically modified mouse models have yielded important insights in the field of systems biology. In this review, we highlight novel molecular discoveries and critical directions for future research on SCUBE proteins in the context of cancer, skeletal disease and cardiovascular disease.
Similar content being viewed by others


Introduction
Three SCUBE [Signal peptide-complement C1r/C1s, Uegf, and Bmp1 (CUB)-Epithelial growth factor domain-containing proteins] proteins in vertebrates comprise a small family with critical functions in a wide variety of physiological and pathological processes. The human SCUBE genes were originally identified from cultured endothelial cells (ECs), but rodent homologs were identified on the basis of their distinctive embryonic expression patterns. Over the past two decades, significant progress has been made in understanding the functions of SCUBEs in normal physiology and disease. In this review, we provide a detailed survey of the crucial known and emerging roles of SCUBE proteins. We begin by recounting the identification, phylogenetic relationships, and protein domain organization and functions of the proteins. Then for each SCUBE family member, we highlight the gene expression patterns, posttranslational control, developmental and physiological functions, and associated diseases. We also discuss several potential translational and therapeutic applications as well as future research directions related to the SCUBE family.
Discovery and phylogenetic analysis
The first identified SCUBE family member (SCUBE1) was isolated from developing mouse urogenital ridge [1] and human vascular endothelial [2] cDNA libraries. Subsequent genomic and homology searches led to the discovery of two additional family members, SCUBE2 [3, 4] and SCUBE3 [5] (Fig. 1A). Based on a comprehensive search using the Genome Browser website (http://genome.ucsc.edu), the three SCUBE genes were found to be highly conserved in zebrafish, mice and humans. Thus, the three proteins comprise a small, evolutionarily conserved family (Fig. 1B) (Table 1) [1,2,3,4,5,6,7,8,9,10]. Despite their conservation in vertebrates, homologous genes have not been identified in the genomes of single-cell yeast (Saccharomyces cerevisiae) or invertebrate animals, such as fruit flies (Drosophila melanogaster) or nematodes (Caenorhabditis elegans). This species distribution is in line with the idea that SCUBE genes might be involved in the development or function of anatomical features specific to vertebrates or animals with vasculature, such as the neural tube or blood vessels. Moreover, comparative genetic analyses revealed that the organization of SCUBE genes in zebrafish, mouse and humans consistently follows a modular arrangement. The N-terminal signal peptide, every epidermal growth factor (EGF)-like domain, and single cysteine-rich (CR) motifs are each encoded by single exon. Meanwhile, the spacer region is encoded by five exons, and the CUB domain is encoded by two exons (Fig. 1A, C).

Protein domains, phylogenetic analysis, genomic organization, sequence alignment, and sequence identity of SCUBE protein family. A Graphic illustration of the domain structure of human SCUBE1, 2 and 3 (see Table 1). SP, signal peptide sequence; E, EGF-like domain (grey shade indicated the calcium-binding EGF module); Cys-rich, cysteine-rich repeats; CUB, the CUB domain. “Y” marks the potential N-linked glycosylated sites of each SCUBE protein. Protein domain sequence identity shared among human SCUBE members is calculated at the bottom; the highest homology was found in the CUB domain (83%), followed by the EGF-like (73%) and Cys-rich (66%) domains. The spacer region appears to have the lowest homology (34%) and may be associated with the unique functions of each SCUBE member. B Phylogenetic tree of the SCUBE family. Similarity of human (h), mouse (m), zebrafish (z), eagle (e), lizard (l), or frog (f) SCUBE protein sequences was analyzed by using the Lasegene MEGALIGN program (ClustalW algorithm). The length of each pair of branches represents the phylogenic distance between sequence pairs. Below the tree is a scale indicating the number of “Nucleotide Substitutions” for both DNA and protein sequences. C Genomic structure of human SCUBE1. SCUBE1 gene consists of 22 exons spanning about 140 kb on chromosome 22. The exon–intron boundaries are well preserved among the SCUBE gene members, which suggests possible gene duplication during evolution. Of note, SCUBE genomic organizations derived from zebrafish, mouse and human genomes (http://genome.ucsc.edu) follow a modular arrangement, with the signal peptide sequence, each epidermal growth factor (EGF)-like domain, and Cys-rich motif, each encoded by a single exon. In addition, the spacer region is encoded by five exons and the CUB is encoded by two exons. D Amino acid sequence identity of SCUBE family. Data in the upper right (blue shading) represent the sequence identity in EGF-like domain, and data on the bottom left (orange shading) represent the sequence identity in spacer region between SCUBE proteins (upper panel). Data on the upper right (gray shading) present the sequence identity in Cys-rich domain, and data on the bottom left (yellow shading) present the sequence identity in CUB domain between SCUBE proteins (lower panel). E Sequence alignment, sequence identity, and structural comparison of EGF-like repeats of human SCUBE proteins. Protein sequences of SCUBE1, SCUBE2 and SCUBE3 used for alignment were derived from NCBI Reference Sequence Database, as listed in Table 1. Symbols use the one-letter code for amino acids. A dash indicates a gap. The conserved cysteines are numbered 1 to 6, and the predicted disulfide bonds form as follows: C1-C3, C2-C4, and C5-C6. Amino acids indicated below highlight the canonical calcium binding (cb) consensus sequence (D/N)-X-(D/N)-E/Q-Xm (D/N*) Xn (Y/F), where m and n are variable and the asterisk indicates potential β-hydroxylation [17]. Of note, two triplets of cbEGF1-3 or cbEGF7-9 modules (amino acid residue boundaries marked in red fonts) are well conserved in all SCUBE members. F Structural comparison of SCUBE3 second and seventh cbEGF-like domains with human NOTCH1 fifth cbEGF-like domain. 3D models of the second and seventh EGF-like domains of human SCUBE3 obtained from AlphaFold2 (AF-Q8IX30-F1-model_v3) [187] were superimposed on the fifth cbEGF-like domain of human NOTCH1 (PDB 5FM9) [221]. These EGF-like domains contain a short calcium binding motif sequence signature: (D/N)-X-(D/N)-E/Q-Xm-(D/N*)-Xn-(Y/F), where m and n are variable and the asterisk indicates potential β-hydroxylation [17]. This canonical calcium coordination module is found near at the N-terminus. The residues coordinating calcium ion (in green) are shown as sticks, and the coordination as pink dotted lines. The five calcium-binding motif residues indicated in E are presented as cyan sticks; Asn and Phe/Try between C3 and C4 of these EGF-like domains are consistent with the substrate sequence pattern of Asp/Asn-β-hydroxylase [222, 223]. The three paired cysteines (C1:C3, C2:C4, and C5:C6) are presented as yellow sticks. Atoms are colored, with oxygen in red, nitrogen in blue and sulfur in gold. This figure was produced using PyMOL (Schrödinger, LLC. The PyMOL molecular graphics system, version 1.8, https://pymol.org). Nt, NH2 terminus; Ct, COOH terminus
The SCUBE genes give rise to polypeptides of about 1000 amino acids with five major protein domains: (1) an N-terminal leader sequence, (2) nine tandem EGF-like repeats, (3) a large spacer region that undergoes N-glycosylation, (4) three stretches of six cysteine residues (designated CR motifs) with conserved amino acid spacing between the cysteines, and (5) a CUB domain at the C-terminus (Fig. 1A). The amino acid sequence of SCUBE3 is 65.6% identical with SCUBE1 and 58.5% identical with SCUBE2, while SCUBE1 and SCUBE2 share 65.6% identity. The CUB domain is the most highly conserved region, sharing 82.2–90.1% (~ 85.2%) identity among all three members. The EGF-like repeats range from 66.0 to 76.7% (~ 72.7%) identity and CR motif region is 62.2–67.7% (~ 65.8%) identical among all three members (Fig. 1D). The spacer region is the most diverse (34% identity), which suggests that this region might be centrally involved in molecular functions that are uniquely associated with individual SCUBE members. In the next section, we provide further details about the overall structures and functions of each SCUBE protein domain.
Domain organization and function
Signal peptide and EGF-like domain
SCUBE proteins have an N-terminal signal peptide domain that is important for protein secretion and localization to the membrane. The adjacent EGF-like domains are composed of stretches of about 30–40 residues and are located near the N-terminus of the SCUBE protein. The EGF-like domain is an evolutionarily conserved protein domain that was first identified in EGF but also exists in many other secreted or membrane-localized proteins. These EGF-like domain-containing proteins are mostly of animal origin and include growth factors, adhesive selectin molecules, extracellular matrix protein, and clotting factors [11]. The domain is frequently found as tandem copies with various degrees of conservation. These tandem repeats tend to fold together to become a single linear solenoid structure that works as a functional unit. In general, each EGF-like domain contains six cysteine residues that form three intramolecular disulfide bonds in a conserved pattern (i.e., C1–C3, C2–C4 and C5–C6) [11] (Fig. 1E). Whether the EGF-like domains are present in the extracellular portion of a membrane protein or in a secreted protein, a typical function of the domain is to facilitate homophilic [12, 13] or heterophilic protein–protein interactions, such as thrombin–thrombomodulin [14], Notch1-Jagged-1 [15], and low-density lipoprotein receptor–proprotein convertase subtilisin/kexin type 9 [16] interactions. In addition, some specialized EGF-like domains are capable of binding calcium and are thus termed calcium-binding EGF (cbEGF) domains. These cbEGF domains carry the characteristic six cysteine residues but also have an additional calcium-binding motif. The motif sequence is: (D/N)–X–(D/N)–E/Q–Xm–(D/N*)–Xn-(Y/F), where m and n are variable and the asterisk indicates a potential β-hydroxylation site [17] (Fig. 1F). Extracellular cbEGFs are known to participate in protein–protein and protein–carbohydrate interactions [18]. A previous biochemical study showed that triple cbEGF repeats could form a rigid, protease-resistant conformation upon calcium binding [19]. Therefore, when a cbEGF domain-containing protein is secreted or exposed to the calcium-rich extracellular environment, tandem cbEGF repeats may acquire a stiff, protease-resistant structure that can facilitate protein–protein interactions.
Intriguingly, we found that six of the nine EGF-like repeats in SCUBE proteins contain calcium-binding consensus sequences and are organized in triplet modules (i.e., cbEGF1-3 or cbEGF7-9) (Fig. 1A, E). When overexpressed, these SCUBE proteins are secreted glycoproteins that can form homomeric or heteromeric complexes with other SCUBE proteins via EGF-like domain-mediated interactions [2, 5, 20, 21] (Table 2) and are stably associated with the cell surface [2, 4, 5]. Further deletion mapping and cell aggregation assays revealed that the EGF modules are involved in reciprocal (trans) or lateral (cis) interactions between SCUBE molecules [20, 22]. Moreover, the triplet cbEGF7–9 comprises a minimal unit (i.e., sufficient but not necessary) for the calcium-dependent trans-homophilic association of SCUBE1 with adjacent cells [20]. In addition, the EGF-like repeats in SCUBE proteins can interact with and modulate signaling activities of growth factors. For instance, SCUBE2 interacts with vascular endothelial growth factor (VEGF) [23, 24], and SCUBE3 interacts with fibroblast growth factor 8 (FGF8) (Table 2) [9].
Spacer region
The spacer region is a relatively large domain situated in the middle of the SCUBE protein, separating the EGF-like domains near the N-terminus from the CR repeats and CUB domain. The spacer is encoded by five exons and respectively contains 231, 227 and 241 amino acid residues in human SCUBE1, 2 and 3 proteins. A potential N-linked glycosylation motif (N–X–S/T consensus sequence, where X is any amino acid except proline) [25] is conserved within the spacer domains of all three SCUBE members (Figs. 1A and 2). To date, no detailed structural information has been published regarding the spacer region. However, our cellular and molecular analyses revealed that this domain functions to tether SCUBE1 protein to the plasma membrane, and it is also necessary for SCUBE1 secretion [20, 26]. Furthermore, we identified a stretch of positively charged residues within the spacer region (amino acids 501–550 of SCUBE1) that interacts electrostatically with anionic heparan sulfate proteoglycans on the plasma membrane [26]. Although it has not been confirmed experimentally, SCUBE2 or SCUBE3 might both utilize similar mechanisms for membrane anchorage, as stretches of positively charged residues were also found in the spacer regions of these molecules.

Sequence alignment of the spacer regions, Cys-rich domains, and CUB domains in the human SCUBE family. The margins of the spacer region and the CUB domain are derived from PFAM analysis and are marked by arrows. Amino acids residues that are identical in at least 2 protein sequences are highlighted in black. Potential N-linked glycosylation sites shared among the SCUBE proteins are denoted with a filled red circle. The minimal furin cleavage sites (RXXR), which are present in SCUBE1 and SCUBE3 but not SCUBE2, are boxed
Cysteine-rich domain
Between the spacer domain and the CUB domain lies the CR domain, which consists of three repeated motifs with six cysteine residues in each repeat. Of note, this 51-amino-acid motif with six cysteines bears little resemblance to the EGF-like domains (~ 40 amino acid residues). An important feature of the three CR motifs is that they can undergo N-linked glycosylation, as each motif in a SCUBE family member carries up to five potential well-conserved N-glycosites (Fig. 1A and Fig. 2). While the precise functions and structures of these motifs are currently unknown, our biochemical and molecular studies revealed that the CR domains are required for extracellular secretion and cell-surface expression of SCUBE1 in vitro [2, 20]. Most importantly, N-glycosylation at four different Asn residues (positions 679, 750, 779 and 789) within the CR domain appears to be essential for SCUBE1 cell-surface binding activity [26]. Further mutagenesis analysis verified that three of the four N-glycosites (i.e., Asn679, Asn750, and Asn789) are indeed utilized in vivo, and any given N-glycan is sufficient to allow the CR motif to bind the cell surface, possibly via an as yet unidentified glycan-lectin interaction [26].
CUB domain
Following the CR domain, a SCUBE protein contains a single CUB domain and a highly conserved C-terminal tail (Figs. 1A, 2). The CUB domains are named after the proteins in which they were originally discovered: human complement proteases C1r and C1s, embryonic sea urchin protein Uegf, and human bone morphogenetic protein (BMP) 1 [27]. A 110-amino-acid module known as the CUB domain is nearly always present in extracellular and plasma membrane proteins. CUB domain-containing proteins participate in a wide variety of biological processes, including developmental patterning [28], axon guidance and angiogenesis [29], cell signaling [30], receptor-mediated endocytosis [31], hemostasis [32], fertilization [33], neurotransmission [34], tissue repair [35], tumor suppression [36], complement activation [37], and inflammation [38]. Although the functional roles of many CUB domains are largely unknown, their structures typically exhibit a β-sandwich fold [39] and some can mediate protein–protein interactions [40], homodimerization [37, 41], heterodimerization [33], and participate in the recognition of substrates and binding partners [42]. In a subset of CUB domains endowed with calcium-binding activity (i.e., cbCUBs), coordination of the calcium ion involves three acidic residues (one glutamic acid and two aspartic acid residues). Structural studies provided further evidence that calcium binding induces and stabilizes the conformation of the cbCUB domain, potentially facilitating its interaction with protein ligands [42, 43].
Along the same lines, the CUB domain of SCUBE proteins can bind numerous growth factors and their cognate receptors to modulate the signaling strength of the growth factors. In particular, the SCUBE1 CUB domain can bind BMP2 [20], fms-like receptor tyrosine kinase 3 (FLT3) and FLT3 ligand [44]. Meanwhile, the CUB domains of SCUBE2 or SCUBE3 can interact with BMP2/4 [21], Sonic hedgehog (SHH) [45,46,47], Indian hedgehog (IHH) [45], and VEGF receptor 2 (VEGFR2) [23, 24] or FGF8 [9], FGF receptor 4 [9] and transforming growth factor β (TGF-β) type II receptor [48]. These interactions may serve to fine-tune the signaling intensities of many key cell signaling events (Table 2). Notably, the direction of effect by these SCUBE proteins (enhancing or suppressing the signaling strength) largely depends on whether the SCUBE protein is expressed in a signal-producing or signal-receiving cell. For instance, when BMP2 is co-expressed with a SCUBE1 C-terminal fragment containing the CR and CUB domains, secretion of BMP2 is markedly reduced, attenuating its long-range signaling activity [20, 22]. On the other hand, when overexpressed SCUBE1 is present on the surface of signal-receiving cells, it binds to BMP2/4/7 ligands and their cognate receptors, acting as a BMP coreceptor to increase BMP signaling activity (Fig. 3A) [8, 49, 50].

Pathological involvement of SCUBE1. A SCUBE1 modulates BMP signaling activity in a context-dependent manner. When expressed in “signal-producing” cells, the C-terminal CR and CUB domains are released by an as yet undetermined proteinase, allowing them to bind and inhibit the secretion of mature BMP into the culture medium. SCUBE1 therefore acts as an antagonist for long-range BMP signaling activity during brain development [20]. In contrast, when SCUBE1 is expressed in “signal-responding” cells, it forms a complex with BMP ligand and its receptors. SCUBE1 thereby acts as a BMP coreceptor to augment BMP signaling activity critical for protecting against kidney ischemia–reperfusion (I/R) injury or for primitive hematopoiesis [8, 49]. B Proposed model for adhesive function of SCUBE1 in platelet aggregation and thrombus formation. Under normal conditions, plasma SCUBE1 is expressed at low levels primarily by endothelial cells and platelets. In addition, SCUBE1 is stored in the α-granules of resting platelets. Upon pathological stimulation, activated platelets secrete large amounts of SCUBE1 into the circulation and high levels of SCUBE1 are also found on the platelet surface. The surface SCUBE1 on nearby activated platelets is trans-homophilically crosslinked by soluble SCUBE1, acting through its sticky EGF-like repeats to promote platelet agglutination and stabilize platelet plugs. Targeting the adhesive modules of SCUBE1 with a specific monoclonal antibody might be a potentially useful anti-thrombotic strategy [72, 86]. C SCUBE1-promoted BMP signaling protects against kidney ischemia/reperfusion (I/R) injury. I/R-induced SCUBE1 protein in proximal tubular epithelial cells serves as a BMP coreceptor to enable renoprotective BMP7 signaling, which stimulates epithelial repair and regeneration through proliferative, anti-apoptotic, and anti-inflammatory effects [49]. D Potential immunotherapy strategy and the pathological function of surface SCUBE1 in MLL-r leukemias. Left: SCUBE1 is an immediate downstream target of the HOXA9/MEIS1 transcriptional regulatory complex, which is activated by MLL-fusion proteins like MLL-AF9 and is crucial for sustaining leukemic transition. Surface SCUBE1 functions as a FLT3 coreceptor in MLL-r leukemias, facilitating the interaction between FLT3 ligand and FLT3. This action increases downstream LYN-AKT activation (tyrosine phosphorylation) to enhance leukemic cell proliferation and survival, thereby promoting leukemogenesis. Right: An anti-SCUBE1 ADC conjugated to an antimitotic agent (MMAE) leads to significant cell killing specifically in MLL-AF9 leukemias. Thus, surface expression of SCUBE1 on MLL-r leukemia cells may be useful as a target for immunotherapy. ADC antibody–drug conjugate; MMAE monomethyl auristatin E. MLL-r, mixed-lineage leukemia gene-rearranged; BMP bone morphogenetic protein 1
Summary of SCUBE protein domain functions
In summary, data from cell culture experiments has shown that SCUBE molecules may be secreted or membrane-tethered N-glycoproteins, which are capable of forming homomeric or heteromeric complexes with other members [2, 5, 20, 21]. The cis or trans-oligomeric interactions of SCUBE proteins are mediated by EGF-like repeats and may be dependent on calcium binding [20, 22, 51]. In addition, a minimum of one triplet cbEGF7-9 is sufficient to drive self-adhesive interaction of SCUBE1 to an adjacent cell surface [20]. Although the spacer region and the CR motif alone are each sufficient for cell-surface tethering, cooperative actions of the two domains within full-length SCUBE1 are required for its secretion [26]. A molecular map revealed an essential polycationic area in the spacer, which most likely interacts electrostatically with anionic heparan sulfate proteoglycans on the cell surface to facilitate membrane attachment [26]. Furthermore, N-glycans of CR repeats greatly influence membrane localization, possibly due to lectin-mediated membrane presentation [26]. It is well known that the CUB domain, which is present in many developmentally regulated proteins, forms a β-barrel. In agreement with its previously reported functions in protein–protein or protein–carbohydrate interactions, we and others have clearly demonstrated that BMP proteins, hedgehog (HH) proteins, and many other growth factors or their receptors can bind to SCUBE proteins. These observations suggest that SCUBE proteins may serve as crucial regulators of growth factor signaling in diverse physiological and pathological processes (Table 2).
In the following sections, we elaborate further on the structural basis of SCUBE protein function and discuss the basic and clinical implications of each SCUBE family member. We also provide suggestions as to how current knowledge of SCUBE proteins may be translated to develop diagnostic, prognostic and therapeutic strategies for various human disorders.
SCUBE1
Regulation of gene expression and posttranslational control
SCUBE1 was identified as the first member of the SCUBE family. It was discovered in samples from human vascular ECs and the developing mouse urogenital ridge in the early 2000’s [1, 2]. In human samples, northern blot analysis revealed that SCUBE1 mRNA is strongly expressed in highly vascularized tissues, such as liver, kidney and lung, with somewhat lower expression in brain, colon and spleen. Consistent with these findings, endothelial expression of SCUBE1 was demonstrated in monkey tissues by in situ hybridization, which showed that the gene is expressed in arterial, venous and microvascular ECs [2]. In addition, whole-mount or embryonic section in situ hybridization showed that mouse Scube1 transcripts are expressed in the developing limb buds, nervous system, gonad, surface ectoderm, somites, and mesenchymal component of numerous tissues in the early head [1, 52]. For instance, Scube1 was found to have strong, restricted expression in facial structures, such as the medial and lateral nasal processes, the maxilla and mandible, the caudal pharyngeal arches, and the dental papilla of both incisor and molar teeth, indicating a potential role during early craniofacial development (Table 3) [52]. Indeed, we reported that the first targeted Scube1 knockout (KO) mice lacking functional CR and CUB domains exhibit an absent cranial vault and exencephaly. Moreover, we concluded that these phenotypes probably arise due to disruption of SCUBE1 antagonization of BMP family members via the CR and CUB domains (see below for further discussion) [20]. Additionally, Scube1 mRNA is expressed throughout branching morphogenesis and in the urogenital system, suggesting that it plays a crucial role in the differentiation of metanephric mesenchymal cells during renal development [1, 53].
Genome mapping showed that SCUBE1 is localized to human chromosome 22q13.2 and Scube1 is found on mouse 15qE1 [1, 2]. There are no definitive reports of disease-associated SCUBE1 mutations in humans, nor are there reports of N-ethyl-N-nitrosourea (ENU)-induced loss-of-function alleles causing Scube1-associated phenotypes in mice. One single nucleotide polymorphism (SNP) within SCUBE1, rs5759224 (intron genetic variant), was significantly associated with venous thromboembolism [54], and two de novo variants, NM_173050: exon 19: c.G2428A: p.E810K (damaging missense) and exon 9: c967 + 1G > A (likely gene disruption), are associated with obsessive–compulsive disorder [55]. Although our previous clinical investigation showed that acute myeloid leukemia (AML) patients with high levels of SCUBE1 expression had worse overall and disease-free survival rates [44], we were unable to detect genetic gains or amplifications of SCUBE1 in either AML or myelodysplastic syndrome cohorts (http://cbioportal.org). Instead, homozygous deletion of SCUBE1 was identified in juvenile AML at a very low incidence (0.34%, 1 in 295 cases) [56]. Meanwhile, a natural variant, G398R, a SNP (dbSNP: rs129415), was detected in myelodysplastic syndrome at a low frequency of 0.39% (4 in 1026 cases). Further research will be necessary to determine whether chromosomal deletion of this SCUBE1 SNP has any clinical impact.
SCUBE1 expression in AML and primary AML samples has been examined, revealing that SCUBE1 expression is upregulated in mixed-lineage leukemia gene-rearranged (MLL-r) AML cells, as compared to healthy hematopoietic stem and progenitor cells, peripheral blood cells, or leukemic cells devoid of the MLL gene rearrangement [44]. Despite the absence of a unique transcriptional regulator for SCUBE1, recent research has shown that in MLL-r leukemia, homeobox A9 (HOXA9) and Meis homeobox 1 (MEIS1) act together to upregulate SCUBE1 [44]. HOXA9 is a homeobox protein with an important role in hematopoiesis and leukemogenesis. Its expression is primarily in less-differentiated cells, such as hematopoietic stem cells and progenitor cells, and is downregulated after lineage differentiation [57, 58]. The similarity of expression patterns for HOXA9 and SCUBE1 suggests that homeobox proteins, including HOXA9, might regulate SCUBE1 during primitive hematopoiesis in mammals. In addition, our studies of renal ischemia–reperfusion (I/R) injury showed that SCUBE1 is a stress-responsive gene in ECs (positive for cluster of differentiation 31/CD31 and platelet endothelial cell adhesion molecule-1/PECAM-1) of the glomerulus and the peritubular capillary network [59] as well as proximal tubular epithelial cells [49]. The regulatory mechanisms underpinning this I/R-mediated induction of SCUBE1 remain unclear. Nevertheless, the 1.5 kb proximal promoter region of SCUBE1 has more than five hypoxia-inducible factor (HIF)-binding sites (A/GCTGA), which are likely to be involved in the ischemia-induced SCUBE1 expression in renal ECs and tubular cells. Further investigations will be required to test whether these binding sites are indeed functional.
By substituting glutamine for asparagine at all N-linked glycosites, we created an N-glycan-deficient SCUBE1-ΔN construct. This construct was expressed in HEK-293T cells to explore the biological significance of N-glycosylation of SCUBE1 [26]. Both SCUBE1-wild type and SCUBE1-ΔN mutant cells produced comparable quantities of secreted protein, which implies that the ΔN mutant is structurally intact. However, due to the absence of N-glycans, the SCUBE1-ΔN mutant protein had a lower molecular mass, and it also exhibited poor cell-surface expression and regulatory activity as a BMP coreceptor [26]. Furthermore, mRNA injection of scube1-wild-type but not scube1-ΔN into scube1-knockdown zebrafish embryos could restore deficiencies in stem cell leukemia (scl) and GATA-binding factor 1 (gata1), two hematological and erythroid biomarkers. Thus, SCUBE1 function during hematopoiesis appears to depend on its N-glycan-mediated cell-surface expression and its ability to modulate BMP signaling in vivo [26].
Developmental and physiological function
Brain development
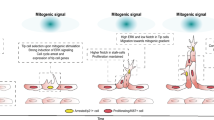
Zebrafish genetic studies showed that the ty97 allele (nonsense mutation in scube2 encoding a truncated null protein lacking the CR and CUB domains) impairs HH and BMP signaling. These findings suggest that the CR and/or CUB domains are indispensable for Scube2 function [6, 7, 10]. To evaluate the biological function of the corresponding C-terminal portion of SCUBE1 in vivo, we generated a targeted disruption of mouse Scube1, which encodes a truncated protein lacking the CR and CUB domains [20]. This mutant allele was named Scube1Δcub, based on the lack of CUB immunoreactivity in the animal tissues [20]. Homozygous Scube1Δcub/Δcub mice showed phenotypes of acrania, lack of brain tissue and death shortly after birth. Histological analysis of E12.5 embryos suggested that a brain defect between the diencephalon and myelencephalon led to exencephaly that was restricted to the midbrain, probably due to excessive neural tissue proliferation and everted cranial neural folds. In particular, hyperplasia was seen in the trigeminal and vestibulocochlear ganglia, with thickening of the forebrain neuroepithelium wall; the hindbrain neural tube was less impacted [20]. Overall, these brain malformation phenotypes are reminiscent of those in animals with BMP antagonist deficiencies, such as mice lacking Noggin and/or Chordin [60, 61]. These observations were in line with our molecular and biochemical findings that the CR and CUB domains are essential for SCUBE1 function in reducing long-range BMP signaling activity when co-expressed in the same cells [20]. Taken together, these studies strongly suggest that SCUBE1 plays a crucial function in the early phases of central nervous system development, presumably by regulating the activity of BMPs through its C-terminal CR and CUB domains (Fig. 3A).
Platelet function
Our previous immunohistochemistry results showed weak anti-SCUBE1 staining in microvascular ECs within a wide variety of vascular tissues, including the kidney, lung, gut and heart [62]. Yet, the immunoreactive signal was strongly restricted to the fibrin and platelet-rich regions of organized thrombi [62]. These findings are consistent with the notion that endothelial SCUBE1 might be released into the bloodstream, where it accumulates in thrombi formed by fibrin clots. SCUBE1 may also be expressed in platelets, similar to other endothelial membrane proteins like von Willebrand factor, P-selectin, and thrombomodulin [63,64,65]. Indeed, the expression of SCUBE1 was confirmed both at mRNA and protein levels in human platelets [62]. Further immunogold electron microscopy revealed that SCUBE1 is present in the membranes of α-granules of platelets, suggesting the organelles could be a potential location for SCUBE1 storage. SCUBE1 labeling was also seen in thin channels or open canaliculi, implying that SCUBE1 may be localized or linked to the membranes [62]. Furthermore, flow cytometry assays demonstrated that P-selectin and SCUBE1 were both expressed on the surface of thrombin-induced activated platelets [62].
The apparent molecular mass of SCUBE1 in resting platelets is about 130 kDa [20], which corresponds to the molecular mass of the full-length product [2]. In contrast, thrombus lysates reproducibly show a proteolytic fragment at ~ 95 kDa. It is known that the CUB domain of platelet-derived growth factor C or D must be proteolytically removed to reveal the physiologically active growth factor core domain [66,67,68,69]. Therefore, locally available fibrinolytic and coagulation proteases at a developing thrombus may perform a similar cleavage to release an active fragment from surface-bound or soluble SCUBE1. Although the precise nature of SCUBE1 processing is unclear, a proposed cleavage site (R535RXR538) for the furin-like protease [70] has been identified in the spacer region and may account for the serum-dependent proteolytic cleavage of SCUBE1 [71]. Consistently, a 95 kDa proteolytic SCUBE1 fragment is greatly increased in plasma from acute coronary syndrome patients who are experiencing plaque rupture/erosion and subsequent platelet activation; the same fragment is undetectable in healthy people [71]. A detailed analysis of the function of soluble SCUBE1 shows that it has a critical role in platelet agglutination and thrombus formation [62, 72]. Although soluble SCUBE1 alone cannot stimulate platelet activation, the SCUBE1 EGF-like repeat-containing protein fragment was shown to enhance ristocetin-induced platelet agglutination [62], which is analogous to platelet-subendothelial adhesion [73]. Since typical platelet aggregation stimulates the binding of fibrinogen to glycoprotein IIb–IIIa, the insensitivity of adenosine diphosphate or collagen-induced aggregation to additional SCUBE1 fragments further suggests a role for soluble SCUBE1 in platelet adhesion as opposed to platelet aggregation. Moreover, the EGF-like repeats of SCUBE1 on the surface of platelets likely mediate platelet self-adhesion [20]. For soluble SCUBE1, both its EGF-like repeats and its CUB domains have potential to interact with platelet endothelial aggregation receptor-1, a transmembrane receptor that contains EGF-like repeats and is expressed on the platelet surface, where it functions in platelet contact-induced activation [74] to mediate platelet agglutination [72].
Hematopoiesis
All vertebrates, including zebrafish, undergo primitive and definitive waves of hematopoiesis. In addition to its function in platelets, SCUBE1 has been shown to play a critical role in primitive hematopoiesis of zebrafish [8]. Knockdown of scube1 in zebrafish embryos with antisense morpholino-oligonucleotides leads to the downregulation of marker genes for early primitive hematopoietic precursors [stem cell leukemia (scl)], erythroid lineage [GATA-binding factor 1 (gata1) and hemoglobin beta embryonic-1 (hbbe1)] as well as early and late myelomonocytic lineages [early: transcription factor PU. 1 (pu.1), and late: myeloperoxidase (mpo) and plastin-2 (l-plastin)] [8]. However, knockdown of scube1 does not affect the expression of an early endothelial marker [fli-1 proto-oncogene (fli1a)], nor does it affect vascular morphogenesis. Importantly, genetic overexpression of bmp restores the expression of scl in the posterior lateral mesoderm during early primitive hematopoiesis in scube1 morphants. This result suggests that scube1 may function as a Bmp coreceptor to promote Bmp signaling, as seen in biochemical and molecular studies. Combined, these findings in zebrafish suggest that scube1 controls Bmp activity during zebrafish embryogenesis, which is essential for primitive hematopoiesis and places SCUBE1 at an early position in the regulatory hierarchy (Fig. 3A) [8].
Pathological function
Acute coronary syndrome (ACS) and acute ischemic stroke (AIS)
Activated platelets are critical players in the initiation and progression of atherosclerosis [75], and SCUBE1 is a platelet granule protein that is exposed on the platelet surface upon activation [62]. Therefore, we performed immunostaining to examine the levels of SCUBE1 in the subendothelial matrices of atherosclerotic lesions, and found strong SCUBE1 staining in these areas [62]. The widespread, diffuse SCUBE1 staining pattern is suggestive of an extracellular localization of the protein in atherosclerotic plaques [62]. Moreover, SCUBE1 expression is not observed in macrophages or smooth muscle cells associated with atheromatous plaques, indicating that the SCUBE1 associated with atheroma may have an exogenous origin. Platelets and their granular contents, including von Willebrand factor, P-selectin and cluster of differentiation 40 (CD40) ligand, are closely associated with atherosclerotic plaques and play an important role in the development of disease [76,77,78]. Since SCUBE1 is generated by platelets and ECs, it might play a role in the pathophysiology of arteriosclerosis when it is deposited inside the subendothelial matrix of plaques. In addition, an adhesion assay showed that matrix-bound SCUBE1 can support platelet adhesion, and its soluble form can enhance ristocetin-induced platelet agglutination [62]. Together, these observations suggest that SCUBE1 might function as a key platelet-endothelial adhesion molecule in the pathophysiological development of cardiovascular disease.
According to a report from the World Health Organization, nearly 17.9 million people died from cardiovascular disease in 2019, accounting for 32% of all global deaths [79]. The most common three cardiovascular diseases are myocardial infarction, stroke and venous thromboembolism. In broad terms, thrombosis is defined as the formation of clots in arteries or veins. In the cases of myocardial infarction and ischemic stroke, the clots that form in an artery consist of platelets, while the clots in venous thromboembolism consist of red blood cells [80]. Platelet activation and aggregation are widely recognized as crucial reactions in arterial thrombosis and are responsible for ischemic complications in ACS or AIS. Since activated platelets can release soluble SCUBE1 into circulation [62], we hypothesized that the plasma level of SCUBE1 might serve as a reliable biomarker of platelet activation in acute atherothrombotic diseases. Indeed, our findings in clinical samples revealed that the plasma SCUBE1 concentration was high in ACS (n = 40) and AIS (n = 40) patients (median 205 and 95.1 ng/ml, p < 0.01), but it was virtually undetectable in healthy controls (n = 40) and people with chronic coronary artery disease (n = 83) [71]. At least four separate studies have included ELISA-based measurements of plasma SCUBE1 in healthy people (n = 40, 50, 50, 24), with average concentrations ranging from 30 to 50 ng/ml [71, 81,82,83]. The upregulation of plasma SCUBE1 is detectable early symptom onset (6 h) and remains so for at least three days. As such, the concentration of SCUBE1 in plasma can be considered an independent indicator of stroke severity based on the National Institute of Health Stroke Scale (β = 3.18, p < 0.001) [71]. In addition, small fragments of SCUBE1 have been identified in samples from ACS patients, which suggests that some proteolytic regulation/activation of plasma SCUBE1 may occur under pathological conditions [71]. Overall, these observations show that plasma SCUBE1 levels are markedly elevated in AIS and ACS patients but not coronary artery disease patients. As such, plasma SCUBE1 can be considered as a potential biomarker of platelet activation in acute thrombotic disease [71].
While the most effective reperfusion method for acute myocardial infarction is percutaneous coronary intervention with stenting, the intervention may lead to stent thrombosis involving activated platelets. Depending on the time delay after percutaneous coronary intervention, stent thrombosis can be classified as early (within 30 days of stenting) or late (within 1 year) stent thrombosis-elevation myocardial infarction (STEMI) [84]. Although stent thrombosis is an uncommon event, it is becoming a significant global health issue due to the increasing number of percutaneous coronary interventions undertaken around the world. Strikingly, the fatality in STEMI can be up to 18% [85], so accurate predictors for STEMI are urgently needed. Bolayir et al. found that the plasma level of SCUBE1 may be a promising biomarker for both early and late STEMI [81, 82]. In particular, the authors showed that the plasma level of SCUBE1 in patients with STEMI can reach ~ 100 ng/ml, and it can be considered an independent prognostic factor. Furthermore, the plasma SCUBE1 level was shown to be positively correlated with soluble CD40 ligand (sCD40L) level in hemodialysis patients, signaling myocardial infarction.
Platelet activation and thrombosis
To further define the precise role of plasma SCUBE1 in vivo, we generated a second version of mutant (Δ) mice that lack soluble but retain membrane-bound SCUBE1. To generate these mice, we removed the genetic sequences encoding a portion of the spacer region and the entire region encoding the three CR repeats. However, the C-terminal CUB domain was still transcribed in-frame in the mutant Δ/Δ mouse platelets [72]. In the plasma SCUBE1-depleted Δ/Δ mice, major platelet receptor expression is unaffected, yet Δ/Δ platelet-rich plasma exhibits abnormal platelet aggregation in response to adenosine diphosphate and collagen. Notably, recombinant SCUBE1 protein treatment improves platelet aggregation in wild-type platelet-rich plasma, and it also restores platelet aggregation in Δ/Δ platelet-rich plasma. The deficit in active plasma SCUBE1 attenuates arterial thrombosis in mice and provides protection from lethal thromboembolism after collagen-epinephrine treatment. Collectively, these observations suggest that plasma SCUBE1 contributes to platelet aggregation by joining adjacent activated platelets during thrombosis. Hence, it is possible that inhibition of soluble SCUBE1 could exert antithrombic actions (Fig. 3B) (see below for further discussion).
Our studies to this point had clearly shown that soluble SCUBE1 is not only a potential biomarker of platelet activation [71], but it is also an active participant in thrombosis [20, 62, 72]. However, we still did not know whether the adhesive EGF-like repeat module is crucial for its activity, and we also did not know the specific contributions of SCUBE1 produced in ECs and platelets during thrombosis. To address these questions, we generated a third version of mutant (Δ2) mice, which lack the entire EGF-like repeat module, and we used these mice to further evaluate the functional significance of this module during in vivo thrombogenesis [86]. The platelet-rich plasma from Δ2 mice exhibits a profound deficiency in platelet aggregation, as induced by a wide range of agonists [adenosine diphosphate (ADP), collagen, thrombin agonist protease-activated receptor 4 (PAR-4) peptide, and thromboxane A2 analogue U46619]. Moreover, genetic deletion of EGF-like repeats also prevents deadly thromboembolism in Δ2 mice and reduces arterial thrombosis [86]. A flow chamber assay was also used to demonstrate that blocking antibodies against the EGF-like repeats in whole blood from wild-type mice can significantly reduce platelet deposition and thrombus formation on collagen-coated surfaces under arterial shear rates. By performing reciprocal bone-marrow transplantation in wild-type and Δ2 mice, we generated animals that only produce SCUBE1 in either ECs or platelets. The animals that only produce SCUBE1 in ECs exhibit a prolonged time to carotid arterial thrombosis after ferric chloride administration, which suggests that platelet-derived SCUBE1 is a crucial factor in arterial thrombosis. This effect is likely mediated by the adhesive EGF-like repeats in platelet-derived SCUBE1, so targeting the adhesive motifs of SCUBE1 may be a potentially useful anti-thrombotic strategy (Fig. 3B).
sCD40L is a tumor necrosis factor expressed in several immune-specific cells, including platelets [87]. Like SCUBE1, sCD40L is stored in platelet α-granules, and it is released upon activation. Furthermore, sCD40L is a biomarker for platelet activation [88], ACS [89] and sepsis [90]. Plasma sCD40L and SCUBE1 levels are correlated in patients with cardiovascular disease [83, 91,92,93] and in in patients with hypertension [83, 92]. According to the ambulatory blood-pressure monitoring data classifications, a ≥ 10% decrease in night-time blood pressure is called “dipper” hypertension, and a < 10% decrease is “non-dipper” hypertension [83, 94]. Guzel et al. analyzed SCUBE1 plasma levels in dipper and non-dipper hypertensive patients [83]. Although the plasma SCUBE1 level is elevated in both classes of hypertensive patients, it is significantly higher in non-dipper patients than dipper patients. However, the level of sCD40L is not significantly different when comparing between the two classes. Since non-dipper hypertension is considered to be more severe than dipper hypertension, SCUBE1 can be used to differentiate between the classes and is considered a biomarker for severe hypertension.
Pulmonary hypertension
Pulmonary hypertension is high blood pressure in pulmonary arteries, which supply the lungs. The condition is characterized by a mean pulmonary artery pressure of ≥ 25 mmHg with a normal pulmonary artery wedge pressure of ≤ 15 mmHg, in addition to increased pulmonary vascular resistance > 240 dyn·s·m−5 [95]. Pulmonary hypertension affects 1% of all adults globally and has a 5-year mortality rate of approximately 40% [95, 96]. This chronic and progressive disease can lead to heart failure and death when left untreated. Pathogenic mutations in the BMP receptor 2 (BMPR2) can cause one of the most severe types of pulmonary hypertension. In line with our studies implicating SCUBE1 in BMP signaling [8, 20, 49], it was shown that SCUBE1 is downregulated in pulmonary arterial hypertension (PAH) with BMPR2 mutation [97]. The study further showed that the SCUBE1 expression level is decreased in ECs derived from human induced pluripotent stem cell ECs with BMPR2 mutation as compared with wild-type controls. Induction of hypoxia or treatment with the inflammatory cytokine interleukin-1β were used to trigger PAH. In BMPR2-mutated cells, these treatments reduce SCUBE1 expression at the transcript and protein levels and also reduce the secreted protein level, whereas most examined EC-specific genes (e.g., VEGF and endothelial nitric oxide synthase) are upregulated [97]. Moreover, SCUBE1 was shown to be downregulated via HIF-1α in this context. The study showed that SCUBE1 acts by regulating Smad1/5/8 signaling, which is relevant to BMPR2 but not TGF-β–related SMAD2/3 signaling. Consistent with the cell-based results, the authors also found reduced SCUBE1 levels in plasma and lung tissue both in the animal model of PAH and in patients with PAH. Therefore, SCUBE1 downregulation in the context of pulmonary hypertension could be a specific biomarker of disease.
Kidney injury
As mentioned in previous sections, zebrafish Scube1 forms a complex with Bmp ligands and its receptors to act a BMP coreceptor and augment BMP signal activity [8]. Nevertheless, it remained unknown if mammalian SCUBE1 can bind to BMP7 and enhance activation of its downstream signaling pathway. This activity would be relevant to kidney injury, as BMP7 is a preventive factor for renal I/R damage [98, 99], which may also facilitate renal tubular cells proliferation and healing after I/R injury. Recently, we demonstrated that proximal tubular cells exhibit I/R-induced upregulation of SCUBE1 expression together with renoprotective BMP7 expression. Further molecular and pharmacological studies showed that SCUBE1 promotes BMP7 signaling by acting as a BMP coreceptor, directly binding to BMP7 and its receptors [49]. For this study, we used Scube1-deletion mutant (Δ2) mice to clarify the pathophysiological role of SCUBE1 following kidney I/R damage. The Δ2 mice show more severe histopathologic features than wild-type littermates, in terms of tubular necrosis and dilation and loss of brush border. The Δ2 mice also have increased inflammatory response, including neutrophil infiltration, and induction of tumor necrosis factor-α, monocyte chemoattractant protein-1 and interleukin-6, during the resolution phase of I/R damage [49]. Furthermore, the Δ2 mice show reduced BMP signaling strength (in terms of phosphorylated Smad1/5/8) together with increased apoptosis and decreased proliferation of kidney tubular cells compared to controls [49]. Together, these results reveal a novel cell-autonomous function of I/R-induced SCUBE1 protein in proximal tubules. We can therefore conclude that surface SCUBE1 functions as a coreceptor to promote BMP7-mediated proliferative, anti-inflammatory, and anti-apoptotic effects to protect from injury and promote epithelial repair and regeneration (Fig. 3C) [49]. However, another paracrine mode of action has also been proposed, whereby SCUBE1 is secreted from peritubular capillary ECs after I/R injury and mediates renal remodeling and cellular replacement by increasing tubular cell proliferation [59]. Nevertheless, the SCUBE1-induced signaling pathway that mediates its reparative paracrine effects on adjacent kidney tubules is still largely unknown [59]. Most importantly, it remains an open question whether stress-induced expression of endothelial or epithelial SCUBE1 may play some role in pathophysiological conditions other than the kidney.
Acute myeloid leukemia (AML)
Although the SCUBE1 plasma level is high in the context of some cancers, such as renal and breast cancer, the exact functions of SCUBE1 in cancer cells remain largely unknown and uninvestigated. In AML, SCUBE1 is known to be abundantly expressed on the MLL-r AML cell membrane and predicts a poor prognosis in de novo AML, and we recently showed that the SCUBE1 gene is a direct target of HOXA9/MEIS1 [44, 100]. These findings raise the possibility that SCUBE1 may have oncogenic properties. For instance, knockdown of SCUBE1 in human MLL-r AML cells reduces leukemic cell engraftment in an in vivo model, and it lowers cell viability and enhances apoptosis in vitro. Along the same lines, ablation of Scube1 in murine hematopoietic progenitor cells greatly reduce the potential for oncogenic MLL-AF9-mediated leukemic initiation, even though SCUBE1 overexpression was insufficient to promote neoplastic transformation. We also assessed whether Scube1 plays a role in sustaining MLL-r AML development in vivo using the Scube1 conditional knockout mouse model [44]. The survival of mice receiving MLL-AF9 leukemia cells is dramatically improved by tamoxifen-induced deletion of Scube1. Collectively, our observations strongly imply that SCUBE1 plays important roles in the development and persistence of MLL-r leukemia [44].
In an effort to further clarify the molecular pathways affected by membrane SCUBE1 during leukemogenesis, we performed unbiased proteomic proximity labeling and mass spectrometry analysis to identify membrane proteins in close proximity to surface SCUBE1 [44]. FLT3 and its direct signaling component lck/yes-related novel protein tyrosine kinase (LYN, a non-receptor protein tyrosine kinase) are both associated with cell-surface SCUBE1 in MLL-r AML cells. Further gain- and loss-of-function, biochemical, and molecular studies demonstrated that the membrane-tethered SCUBE1 binds to extracellular ligand-binding domains of FLT3, which promotes signaling along the FLT3-LYN axis in order to enhance leukemic growth and survival signals [44]. Thus, our results showed that aberrantly upregulated SCUBE1 binds to the HOXA9/MEIS1 target FLT3, stabilizing FLT3 protein and promoting its activation. This mechanism appears to represent a feed-forward mechanism that serves to enhance the FLT3–LYN signaling and also reinforce the signaling network downstream of HOXA9 and MEIS1 during leukemogenesis. Overall, this study shed light on critical pathological roles of SCUBE1, a newly identified transcriptional target of HOXA9/MEIS1, in MLL-r AML initiation and maintenance (Fig. 3D).
Potential translational applications and therapeutic strategies
As stated above, SCUBE1 concentrations are low in plasma derived from PAH animal models as well as patients with PAH [97]. In addition, plasma SCUBE1 concentrations are closely associated with hemodynamic markers of disease severity. Thus, SCUBE1 is implicated as a contributor to BMPR2-related PAH pathogenesis, which suggests that it may be a relevant target for therapeutics, diagnosis, and/or prognosis of the disease [97].
Given the susceptibility of Δ2 mutants after I/R injury and the novel BMP7-enhancing effects of upregulated SCUBE1 in response to I/R stress [49, 59], targeting SCUBE1 might be a feasible treatment strategy for acute kidney injury. This could be accomplished by infusion of recombinant SCUBE1 protein, gene therapy to induce targeted renal SCUBE1 expression, or injection of compounds that can effectively and specifically upregulate renal SCUBE1 protein. Moreover, plasma SCUBE1 level is greatly increased due to secretion from platelets and/or ECs in pathologic conditions, making it a possible biomarker of platelet activation in the context of ACS, AIS, hemodialysis or hypertension. Similarly, injured or regenerating tubular cells may release membrane-tethered SCUBE1 into the urine, which may have diagnostic or prognostic utility for acute kidney injury. Clinical studies will be required to test the utility of urinary SCUBE1 as a biomarker and to assess whether supplementation of exogenous SCUBE1 protein could provide therapeutic benefit for kidney disease.
Because our cumulative in vitro/ex vivo platelet binding/aggregation assays [20, 62] and in vivo genetic ablation studies [72, 86] suggested that the EGF-like repeats of SCUBE1 are essential for thrombogenesis, we tested whether blocking the EGF-like repeats of SCUBE1 might prevent thrombosis. To investigate the effects of particular antibodies targeting EGF-like repeats of SCUBE1 on platelet adhesion and thrombus formation, we performed proof-of-concept experiments using a flow chamber. Two different monoclonal antibodies, mAb #2 and mAb #7, were respectively raised against EGF1-3 or EGF4-6 of SCUBE1. To assess the formation of thrombi on a collagen-coated microfluidic device, whole blood from wild-type mice pre-incubated with isotype control antibody, anti-SCUBE1 mAb #2, mAb #7 or anti-CUB was perfused through the flow chamber at a high shear rate of 1000 s−1 [86]. We found that treatment with mAb #2 or mAb #7 only slightly increases initial platelet adhesion, but it significantly decreases subsequent platelet recruitment and thrombus formation. In contrast, control IgG and anti-CUB mAb seem to have no impact on platelet adhesion or thrombus formation, indicating that the specific targeting of EGF-like repeats by mAb #2 and mAb #7 is crucial for the effect [86]. Furthermore, mAb #7 blocks ristocetin-induced platelet agglutination [72] and protects wild-type mice against fatal thromboembolism triggered by collagen and epinephrine (60% survival of treated mice, compared with 0% survival in saline and isotype control groups). mAb #7 also attenuated arterial thrombosis formation due to ferric chloride injury [72]. Importantly, mice injected with anti-thrombotic SCUBE1 mAb #2 or #7 exhibit no evidence of prolonged bleeding times [72, 86]. Collectively, these findings demonstrate the critical functions of EGF-like domains in the recruitment and deposition of platelets during aggregation. Antibodies targeting SCUBE1 EGF-like repeats may therefore be promising tools for the prevention of thrombosis (Fig. 3B).
Once an antibody binds to a cell-surface target, its internalization and transport to lysosomes allows an antibody–drug conjugate (ADC) to execute its killing effect, so long as the cytotoxic payload can be released from the lysosomes [101]. We therefore generated an anti-SCUBE1 mAb #1 (a different clone from the above-mentioned antithrombotic mAb #2 and mAb #7) that could be rapidly bound, efficiently endocytosed to lysosomes and degraded within 24 h. This timeline suggested that the mAb could be efficiently internalized and directed to lysosomes [44]. We then produced a SCUBE1-targeting ADC by linking mAb #1 to an enzymatically cleavable valine-citrulline linker and an anti-microtubule cytotoxic agent monomethyl auristatin E (MMAE). Importantly, the ADC (anti-SCUBE1-valine-citrulline-monomethyl auristatin E) retains binding affinity similar to the parental antibody [44]. In addition, the ADC can selectively kill SCUBE1-expressing MLL-r leukemia cells (THP-1 or NOMO-1) but not SCUBE1-negative KG-1a or K562 leukemic cells in vitro, and it can suppress MLL-r leukemia growth in a xenograft model [44]. The ADC-receiving mice also show no signs of antigen-independent toxicity, as indicated by weight loss. Collectively, these findings demonstrate the selectivity of the ADC and support the idea that surface SCUBE1 has potential as a therapeutic target and MLL-r-specific biomarker (Fig. 3D).
SCUBE2
Regulation of gene expression and posttranslational control
Research on SCUBE2 has been largely driven by findings from zebrafish genetic studies [6, 7, 10] and human breast cancer genomic studies [102,103,104,105,106]. Notably, a series of reports using a positional cloning approach showed that the zebrafish homologue of scube2 is required for HH signaling during the development of the ventral spinal cord, dorsal aorta, and slow muscle [6, 7, 10]. Meanwhile, human SCUBE2 has been implicated in breast cancer biology. As such, SCUBE2 was the only overlapping breast cancer-associated gene identified in a microarray gene expression profiling study [102] involving a cross-platform comparison of gene lists that define a molecular signature for breast cancer (70 genes) [103, 104] and recurrence score models [105, 106] for predicting the prognosis of breast cancers.
The zebrafish genetic analyses suggest that Scube2 acts upstream of the HH response mechanism [6, 7, 10] and that it can attenuate Bmp activity [7]. Of note, a nonsense mutation in the ty97 allele encodes a truncated Scube2 product lacking the CR and CUB domains, which suggests that these domains are indispensable for Scube2 function [6, 7, 10]. In line with these genetic studies, molecular and biochemical experiments showed that SCUBE2 can positively regulate SHH signaling via separate modes of action on signal-producing and signal-receiving cells. In the first mechanism, the soluble form of SCUBE2 facilitates the release and solubilization of dual-lipidated SHH ligand by acting as a chaperone for dual-lipidated HH or as an enhancer of HH shedding. In the dual-lipidated HH ligand, a palmityl moiety is attached at the N-terminus of HH by palmityl transferase [107] and a cholesteryl residue is linked at the C-terminus by autocatalysis [108]; this form of HH is mainly involved in long-range signaling [46, 47, 109,110,111]. In the second mechanism, membrane-bound SCUBE2 within lipid rafts (cholesterol-concentrated membrane microdomains enriched with cholesterol-modified HH ligand) can bind, concentrate and promote the presentation of SHH to the patched homolog 1 receptor in signal-responding cells [4]. Therefore, SCUBE2 positively modulates HH signaling activity, either by promoting the release of HH ligand from signal-producing cells or by enhancing cellular signaling in the signal-receiving cells. In addition, a zebrafish genetic study showed that Scube2 can attenuate Bmp activity [7]. Similarly, our clinical, biochemical and molecular studies showed that in the context of breast cancer, high SCUBE2 expression suppresses breast tumor cell proliferation and is associated with a favorable prognosis for invasive breast cancer, likely because the CR and CUB domains directly bind BMP proteins and antagonize long-range BMP signaling [112].
A commercially available cDNA clone for human SCUBE2 represents a splice variant lacking the ninth EGF-like repeat and a portion of the spacer region [2]. Therefore, we swapped in a cDNA fragment to correct the defective region in this clone [4]. The resulting human SCUBE2 full-length cDNA encodes a polypeptide containing a fully organized protein domain structure in line with zebrafish scube2 (Table 1) [4]. Interestingly, a major difference between SCUBE2 and SCUBE1 is that SCUBE2 carries an extra 30 amino acid residues (encoded by a single exon) between the fifth and sixth EGF-like repeats (Fig. 1A) [4]. This added stretch contains a potential N-linked glycosylation site and also a minimal furin cleavage site (R276XXR279), which can allow for release of an N-terminal cleaved fragment containing the EGF1–5 repeats from the rest of the protein [4]. Of note, this furin consensus site was ablated to improve the yield of full-length SCUBE2 protein in a biochemical study on SHH signaling [113]. Currently, the biological relevance of this extra stretch of 30 amino acids in SCUBE2 remains unknown, but it can be postulated that proteolytic cleavage might represent a regulatory mechanism that could be critical for SCUBE2 function. Analogously, upregulated SCUBE2 in breast cancer primary tumor cells can be proteolytically cleaved by matrix metalloproteinase 2 (MMP-2) to separate the C-terminal CR and CUB domains from the N-terminal EGF-like repeats and the spacer region [112]. It was shown that only the C-terminal CR and CUB domains but not the full-length protein or the ty97 mutant lacking the CR and CUB domains can potently suppress the secretion of mature BMP2 into conditioned medium [112]. Consistent with these data, overexpressed full-length SCUBE2 and ty97-encoded protein are secreted into culture medium and properly targeted to the cell surface, while the fragment containing only C-terminal CR and CUB domains is defective in membrane association and secretion [112]. Intriguingly, the C-terminal fragment is subject to regulatory processing via MMP-2. Since the fragment is incapable of membrane binding or secretion, it confines BMP ligands within the signal-producing cells, thus preventing BMP secretion and long-range signaling activity [112]. The findings from our breast cancer studies might therefore partly explain why the zebrafish Scube2 can attenuate Bmp in a permissive manner [7]. The anti-Bmp activity of zebrafish Scube2 may depend on the concurrent presence of some as yet unidentified cofactor, although MMP-2 is a likely candidate. Further investigations will be needed to test this possibility.
Human SCUBE2 and mouse Scube2 are respectively located at chromosome 11p15.4 and 7qE3. Like SCUBE1, no germline SCUBE2 mutations are directly associated with human diseases, and no ENU-induced phenotypes are associated with mutations in mouse Scube2. Nevertheless, a genome-wide association study on aberrant plasma β2-glycoprotein I levels identified an intronic SNP (rs963167) from SCUBE2 that is significantly associated with the β2-glycoprotein I phenotype [114]. β2-Glycoprotein I levels are associated with autoimmune diseases, hemostasis, atherogenesis and angiogenesis. However, the authors were unable to detect a significant association between the identified SCUBE2 variant and coronary artery disease in an in silico follow-up study [114]. Although the genome-wide association study provided evidence that SCUBE2 is a genetic determinant of plasma β2-glycoprotein I level, further investigations will be required to elucidate the putative role of SCUBE2 in β2-glycoprotein I-associated cardiovascular and autoimmune diseases. In addition, a recent study using targeted sequencing of previously identified linkage regions revealed a missense variant (rs61751342) within the chromosome 11 quantitative trait locus that co-segregates with carotid plaque [115]. The rs61751342 missense variant is located in the DENN domain-containing 2B gene (DENND2B) and is an expression quantitative trait locus for SCUBE2 in the atrial appendage (p = 3.5 \(\times\) 10–5, GTEx Consortium) [115].
Developmental and physiological function
Regulation of the HH signal pathway
Zebrafish is the first in vivo model in which Scube2 activity was found to be involved in the control of the HH signal pathway. Furthermore, this regulatory mechanism was observed during development of slow muscle, the ventral spinal cord, and the dorsal aorta [6, 7, 10, 116]. In addition, a zebrafish genetic and biochemical study showed that Scube2 is also involved in maintaining the structure and function of the notochord, a transiently existing tissue that exerts both mechanical and signaling cues during vertebrate embryonic development [117]. In particular, Scube2 interacts with Emilin3 in the extracellular matrix of the notochord sheath, which inhibits the regulatory activity of Scube2 on release of HH ligands by notochord cells [117]. Thus, Emilin3 ensures proper notochord patterning by supporting the integrity and function of the notochord sheath [117].
The HH signaling pathway was first identified in Drosophila and has been found to be highly conserved in vertebrates. Although there is only one HH in the fruit fly, the mammalian HH family includes three ligand proteins, SHH, IHH and desert hedgehog (DHH), all of which are involved in morphogenesis and organogenesis [118, 119]. During prenatal skeletal development, IHH is mostly expressed in prehypertrophic chondrocytes, and it serves to regulate chondrocyte differentiation and proliferation, guiding the differentiation of perichondrial mesenchymal cells into osteoblasts [120,121,122,123,124]. In the postnatal period, chondrocyte-derived IHH plays a crucial role in preserving the growth plate, trabecular bone development, and skeletal growth [125]. Using whole-mount and section in situ hybridization, the mouse Scube2 transcript was found to be expressed in the embryonic neuroectoderm, the developing face, heart, vasculature, and the perichondrium of endochondral skeletal structures, such as developing ribs, lumbar vertebrae and long bones of limbs (Table 3) [3, 126]. The primitive mesenchymal cells that constitute the perichondrium surrounding the growth-plate cartilage develop into osteoblasts that later form the cortical and trabecular bones [127]. Nonetheless, it remains unknown whether mammalian SCUBE2 participates in endochondral bone formation or other developmental processes, as has been observed for zebrafish scube2.
Endochondral bone formation
Our study demonstrated that soluble SCUBE2 is more effective than SCUBE1 or SCUBE3 at modulating IHH signaling in vitro [45]. Further experiments in mice showed that Scube2 plays a role in osteogenesis by facilitating Ihh-stimulated osteoblast formation from mesenchymal progenitor cells. In line with the these findings in vitro and the role of Ihh in orchestrating skeletogenesis, we further found that genetic knockout of Scube2 (deletion of all coding exons) results in poor Ihh-mediated chondrocyte differentiation and proliferation, along with defects in osteoblast differentiation and deficient endochondral bone formation in bone-marrow mesenchymal stromal cell cultures [45]. Together, these findings suggest that Scube2 is an essential determinant of Ihh-dependent endochondral bone formation. Particularly, SCUBE2 protein produced in perichondral cells may influence prehypertrophic chondrocyte-derived IHH signaling by facilitating the release and mobilization of IHH to promote differentiation of osteoblasts during endochondral bone formation (Fig. 4).

SCUBE2 is essential for controlling IHH signaling during endochondral ossification. SCUBE2 protein is expressed in peri-chondrial cells and may be secreted (dashed line with arrow). This secreted SCUBE2 could modulate IHH signals derived from pre-hypertrophic chondrocytes (solid line with arrow) by promoting the release and mobilization of IHH, which drives differentiation into osteoblasts during endochondral bone formation. Green arrow indicates the differentiation direction toward hypertrophic chondrocytes within endochondral bone. IHH Indian hedgehog
Neovascularization
VEGF is an essential angiogenic factor that drives proangiogenic signaling by binding to VEGFR2, which is also subject to regulation by VEGFR2 coreceptors such as neurophilins [128]. Since vascular EC-derived SCUBE2 is known to be a peripheral membrane protein that binds and functions as a coreceptor for other signaling receptors [4, 45, 51], we conducted biochemical and molecular studies that revealed a role for SCUBE2 in the VEGF/VEGFR2 signaling pathway [23]. In our study, we found that SCUBE2 overexpression increases VEGF-induced EC proliferation and capillary-like network formation on Matrigel, while its knockdown has the opposite effect. We also showed that similar to VEGF, the transcriptional and translational levels of endothelial SCUBE2 are also upregulated by HIF-1α. Consistently, recovery of blood flow after hind-limb ischemia and VEGF-induced neovascularization in implanted Matrigel plugs are impaired in EC-specific Scube2-knockout mice. Co-immunoprecipitation and binding assays further showed that VEGF and VEGFR2 both bind to SCUBE2, suggesting that SCUBE2 acts as a coreceptor to facilitate VEGF binding and augment VEGFR2 signal activity (Fig. 5). SCUBE2 deficiency (knockout or knockdown) suppresses VEGF-mediated activation of proliferation and proangiogenic signals, while SCUBE2 overexpression enhances the signaling readouts, including phosphorylation of VEGFR2 and activation of mitogen-activated protein kinase (MAPK)/AKT.

Physiological and pathological roles of endothelial SCUBE2 in modulating VEGF signaling during angiogenesis and potential immunotherapy approach. VEGF-stimulated VEGFR2 signaling is essential for angiogenesis and vasculogenesis under physiological conditions, such as during embryonic development (left panel). However, under pathological circumstances (e.g., intratumor hypoxia), hypoxia-inducible HIF-1 upregulates SCUBE2. The SCUBE2 protein localizes on the tumor EC cell surface, where it functions as a coreceptor with VEGFR2 to facilitate VEGF binding and enhance its downstream signaling. Thus, SCUBE2 promotes VEGF-induced tumor angiogenesis [23] (middle panel). SCUBE2 is internalized into the endosome-lysosomal protein degradation pathway when SP.B1 mAb attaches to EC-surface SCUBE2. This internalization decreases the VEGF-VEGFR2 association and prevents tumor angiogenesis (right panel). EC, endothelial cell. VEGF vascular endothelial growth factor; VEGFR2 VEGF receptor 2
Despite the production of SCUBEs by endothelial cells [2, 5], ablation of any single Scube gene does not cause any overt vascular phenotype during development [20, 21, 45, 129]. Thus, SCUBEs may exhibit functional redundancy, or other Scube genes may compensate for a single gene knockout during embryogenesis. Owing to the fact that SCUBE proteins may interact as homodimeric or heterodimeric complexes [2, 5], it should be possible to infer whether SCUBEs cooperate genetically or functionally during vasculogenesis. We therefore initiated a study to dissect the mode of cooperation of scube genes in zebrafish [130]. Knockout alleles were generated by genome editing for each scube gene. As expected, no major phenotypes were observed in the vasculature of any single scube KO mutant, confirming the redundancy or compensatory effects of the scube genes in zebrafish development [130]. In contrast, the scube1/scube2 double knockout shows severe defects in EC proliferation, filopodia extension and migration, which disrupt vascular lumen formation in vasculogenesis and angiogenesis, further impairing the development of distinct vasculatures in different organs [130]. These findings are in line with our recent studies showing that EC SCUBE2 acts as a coreceptor for VEGFR2 and regulates VEGF-induced tube formation and proliferation of ECs [23, 24]. In particular, SCUBE2 regulates VEGFR2-mediated signaling in the context of postnatal ischemia-induced angiogenesis [23] and also in hypoxic pathological tumor angiogenesis [24]. Further genetic, pharmacological and molecular studies in zebrafish suggest that Scube1 and Scube2 may collaborate at the level of cell-surface receptors to promote Vegfa signaling during embryonic vascularization (Fig. 6A) [130]. Additionally, our observations imply that it may be possible to treat VEGF-mediated proliferative vascular disorders by modifying VEGF signaling with therapeutic agents that target SCUBE1 and/or SCUBE2.

Functional studies of SCUBE family using the zebrafish model. A Scube1 and Scube2 from zebrafish endothelial cells work together to support Vegfa signaling during embryonic vascularization. Endothelial Scube1 and Scube2 form a complex under physiological conditions, such as during the development of embryonic vessels. This complex may promote Vegf-induced Vegfr2 phosphorylation and its downstream signaling for proper vasculogenesis and angiogenesis [130]. B Zebrafish Scube3 acts as an Fgf coreceptor during fast muscle fiber development. We utilized a combination of molecular, biochemical, and genetic methods to reveal a new biological function for SCUBE3 as a key regulator of fast muscle precursors. SCUBE3 potentially acts as a coreceptor at the plasma membrane to promote Fgf8 signaling for the differentiation of fast muscle precursors in zebrafish [9]
Pathological functions
Breast cancer
The first clinical implication of SCUBE2 in breast cancer biology was from a gene expression profiling study in which the authors made cross-platform comparisons of lists of genes [103, 104] or recurrence score models [105, 106]. The authors concluded that the breast cancer gene expression profiles overlap by only one gene, SCUBE2 [102]. We discovered that endogenous SCUBE2 protein is expressed on the ductal epithelial or vascular EC surface of healthy breast tissue using immunohistochemistry with an anti-SCUBE2 polyclonal antibody. Additionally, in terms of disease-free survival, patients with positive SCUBE2-expressing tumors (55%; 86/156) have a superior prognosis to those with negative SCUBE2-expressing tumors. Our clinical association and functional studies also suggest that SCUBE2 may be a valuable biomarker indicating good clinical prognosis. Breast cancer cell proliferation and tumor growth were found to be significantly influenced by changes in SCUBE2 protein levels [112]. Molecular and biochemical analyses revealed that SCUBE2 protein is cleaved into N-terminal and C-terminal fragments by MMP-2 protease. The N-terminal EGF-like repeats mediate calcium-dependent cell–cell homophilic adhesion, interact with E-cadherin (a master tumor suppressor), and inhibit the β-catenin signaling pathway, whereas the C-terminal CUB domain directly bind to and antagonize BMP activity in an autocrine manner [22, 112]. During TGF-β-induced epithelial–mesenchymal transition (EMT), SCUBE2 mRNA is downregulated in response to hypermethylation of CpG islands by DNA methyltransferase 1. By causing feed-forward induction of a positive regulator of E-cadherin, forkhead box A1, ectopic SCUBE2 overexpression can stabilize and facilitate E-cadherin-containing adherens junction formation. In turn, this action leads to epithelial transition and inhibits the migration and invasion of aggressive MDA-MB-231 breast cancer cells (Fig. 7; Table 4) [51].

SCUBE2 functions as a tumor suppressor, preventing the migration and invasion of breast cancer cells by reversing epithelial-mesenchymal transition (EMT). By promoting the expression of forkhead box A1, an E-cadherin positive regulator, and subsequent transactivation of E-cadherin, SCUBE2 increases the formation of E-cadherin-containing adherens junctions. This process causes breast cancer cells to undergo epithelial transition by reversing EMT. During EMT, SCUBE2 is epigenetically silenced by binding of DNA methyltransferase 1 at its CpG islands
In several clinical studies, SCUBE2 was found to be more highly expressed in non-metastatic tumor tissues than in normal tissues [131]. The expression of SCUBE2 predicts a favorable outcome in epidermal growth factor receptor (EGFR)-negative and estrogen receptor (ER)α-positive breast cancers [132,133,134,135]. Furthermore, low expression of SCUBE2 is associated with reduced time to disease-related mortality [135,136,137,138,139] and contributes to tumorigenesis in triple-negative breast cancer [140, 141]. The expression of SCUBE2 is also correlated with levels of microRNAs, such as hsa-miR-9-5p and miR-155-5p [137, 142]. In addition, SCUBE2 expression is upregulated in breast cancer stem cells, promotes EMT, and enhances metastasis of triple-negative breast cancer by promoting NOTCH signaling. [143]. These findings imply that SCUBE2 may be a potential druggable molecular target for treating triple-negative breast cancer that has spread and become aggressive (Table 4). Moreover, SCUBE2 expression and function in different types of breast cancer appear to be dynamically regulated at various stages of disease progression. In the earlier stages, SCUBE2 acts as a tumor suppressor, but is downregulated later during TGF-β-induced EMT. Most importantly, SCUBE2 is expressed in breast cancer stem cells and plays a role in the metastasis of triple-negative breast cancer. Since the roles of SCUBE2 in breast cancer appear to be complex and context-dependent, further investigations are needed on this topic.
Other cancer types
In several types of cancer, SCUBE2 is thought to act as a tumor suppressor. For instance, SCUBE2 is commonly downregulated in non-small-cell lung cancer (NSCLC) [144], glioma tissues [145], colorectal cancer tissues [146], parathyroid adenoma tissues [147], gastric carcinoma tissues [148], and invasive regions of bladder cancer [149]. Moreover, loss of SCUBE2 expression in different cancers is associated with pathological progression, in terms of clinical stage, tumor size, vascular invasion, lymph node status, depth of invasion and histological grade, and it is an independent predictor of poor prognosis [148,149,150,151]. Meanwhile, overexpression of SCUBE2 inhibits the proliferation of NSCLC, glioma, and colorectal cancer cells, and it also suppresses cell migration and invasion [144,145,146]. Kaposi sarcoma is a rare type of cancer that develops in the lining of blood and lymph vessels. A whole-genome sequencing and genome-wide linkage analysis identified two homozygous variants in a familial clustering of classical Kaposi sarcoma in an Iranian family. One identified variant is in SCUBE2 (c.1489G > A [p.Pro497Ser]), and the other is in CDHR5 [c.2269C > T (p.Gly757Ser)], which encodes a Mucin protein (Table 4) [152].
Tumor angiogenesis
Angiogenesis can be a double-edged sword in health and disease, as it is centrally involved in both normal tissue homeostasis and tumorigenesis, supporting the growth and dissemination of cancer cells. SCUBE2 acts as coreceptor for VEGFR2 expressed on the surface of ECs, which enhances VEGF-mediated signaling in adult angiogenesis [23]. However, it remains unclear whether SCUBE2 plays an important role in pathological angiogenesis. To address this issue, we performed a study investigating the pathological role of SCUBE2 in the vasculature of tumors. Our immunohistochemistry results showed that SCUBE2 is more strongly expressed in the ECs of several types of carcinomas compared to those in normal tissues. We also established tumor models using Lewis lung carcinoma cells (LLC), melanoma cells (B16F10), and mouse testis Leydig tumor cells (MLTC-1) that express very low levels of or no SCUBE2. These models showed that genetic knockout of SCUBE2 in ECs significantly decreases tumor vascular density and reduces xenograft tumor growth [143]. The abnormalities in angiogenesis seen in Scube2-knockout mice suggest that tumor cells should become starved of nutrients and oxygen, leading to apoptosis and necrosis. Correspondingly, Scube2-knockout mice display increased apoptosis (TUNEL assay) and markedly lower levels of tumor cell proliferation (Ki67 immunostaining) compared to wild types [143]. Together, these findings suggest that endothelial SCUBE2 is essential for angiogenesis that ensures tumor cell survival (Fig. 5).
Cardiovascular diseases
SCUBE2 is significantly downregulated in ECs after in vitro tumor necrosis factor-α and interleukin-1β treatments and after in vivo lipopolysaccharide injection [2]. Therefore, SCUBE2 may play important roles in inflammation-related diseases. The functions of SCUBE2 in inflammation-related diseases have been explored in several studies. In one study, a co-culture model of the blood–brain barrier was exposed to Mycobacterium tuberculosis, which damages the barrier and decreases the expression levels of tight-junction protein, type IV collagen, and SCUBE2. In addition, the SCUBE2-related HH pathway is downregulated by M. tuberculosis stimulation. The molecular mechanism of this phenomenon was also determined. Intriguingly, downregulation of SCUBE2 after exposure to M. tuberculosis limits release of SHH from astrocytes and its delivery to the protein patched homolog 1 (PTCH1) at the surface of ECs [153]. Another study showed that traumatic brain injury leads to the induction of MMP-9, which inhibits SCUBE2-mediated SHH release. These events cause damage to tight junctions and disrupt the blood–brain barrier [154]. In other work, researchers showed that in patients with dyslipidemia and type 2 diabetes, SCUBE2 is highly expressed and positively correlated with endothelin-1, a potent vasoconstrictor peptide produced by vascular ECs [155]. Moreover, SCUBE2 is upregulated in the injured carotid artery and aortic sinus after carotid artery ligation in wild-type mice and low-density lipoprotein receptor-knockout mice with high-fat-diet-induced atherosclerosis [156]. In a recent clinical study, the authors identified a missense variant (rs61751342) that co-segregates with carotid plaques. This variant is located in DENND2B and is an expression quantitative trait locus for SCUBE2 in the atrial appendage [115]. Despite the lack of detailed studies exploring the molecular mechanism of SCUBE2 in arteriosclerosis, it has been suggested that SCUBE2 may play an important role in the development of atherosclerotic plaques [115, 156].
Our previous studies showed that SCUBE2 is expressed in vascular ECs, where it genetically interacts with SCUBE1 and functions as a coreceptor for VEGFR2 to regulate VEGF-induced angiogenesis [23, 24, 130]. Furthermore, we showed that soluble SCUBE1 actively participates in thrombogenesis [20, 62, 72, 86]. Since thrombi frequently form in the atrial appendage of patients with atrial fibrillation or mitral valve disease [157], SCUBE2 may be considered as a potential therapeutic target for cardiac disease [115]. Nevertheless, functional studies are still needed to validate and further elucidate the putative roles of SCUBE2 in the pathological development of these heart conditions. Another genetic study revealed that Tibetans who live at high altitudes often carry an 662-bp intronic insertion in SCUBE2 that is linked to improved lung performance [158]. This genomic structural variant was therefore postulated to contribute to high-altitude adaptation. Our studies showed that SCUBE2 is upregulated by HIF-1α at both the mRNA and protein levels in lung ECs and that endothelial SCUBE2 acts as a novel coreceptor for VEGFR2 and potentiates VEGF-mediated signaling in adult angiogenesis (Fig. 5) [23]. Thus, our findings provide a likely mechanistic explanation for the observed association between the SCUBE2 structural variant and better lung function in Tibetans [158].
Autoimmune disease
Multiple studies have linked SCUBE2 to different autoimmune diseases, including systemic sclerosis and rheumatoid arthritis. One recent study investigated the association of SCUBE levels with ultrasonographic skin thickness and other clinical characteristics in systemic sclerosis patients. The serum level of SCUBE2 was negatively correlated with the forced vital capacity value, and it was positively correlated with the Duruöz Hand Index and serum C4 level [159]. In another study, synovial tissue of rheumatoid arthritis rats showed a high level of SCUBE2 and low level of miR-543, which can target SCUBE2 and interact with a long non-coding RNA called plasmacytoma variant translocation 1. Furthermore, downregulation of plasmacytoma variant translocation 1 induces apoptosis of rheumatoid arthritis fibroblast-like synoviocytes via the miR-543–SCUBE2 axis [160].
Translational application and therapeutic strategy
We recently demonstrated that EC-surface SCUBE2 is an essential VEGFR2 coreceptor in the context of pathological tumor angiogenesis [23]. Therefore, we further examined the possibility of pharmacologically treating solid tumors to inhibit SCUBE2 surface expression with a function-blocking mAb against SCUBE2 (SP.B1) developed by conventional hybridoma technology. The SP.B1 antibody recognizes only murine and human SCUBE2 in ECs and specifically recognizes the spacer region, which essential for SCUBE2 anchoring on the cell surface. Importantly, the mAb does not cross-react with SCUBE1 or SCUBE3. Treatment with SP.B1 in mice significantly decreased the growth and the vascular density of xenografted melanoma, as well as lung, pancreatic, testis, and colorectal tumors [143]. Further mechanistic investigations showed that SP.B1 binding to SCUBE2 causes the protein to be internalized for lysosomal degradation. This internalization reduces the SCUBE2 cell-surface level, attenuating VEGF binding and downstream signaling, as indicated by phosphorylation of VEGFR2 and stimulation of AKT/MAPK (Fig. 5). Another crucial finding in this study was that combination treatments of the anti-SCUBE2 mAb with chemotherapy (docetaxel and gemcitabine) or anti-VEGF antibody are more effective than single-agent therapies. Therefore, an anti-SCUBE2 mAb that induces protein internalization may provide a powerful adjuvant treatment to block tumor angiogenesis [143].
Global inactivation of SCUBE2 in mice does not appear to have any negative effects on the vascular system under normal physiological conditions. Since SCUBE2 is specifically upregulated by hypoxia and highly expressed in tumor vasculature, the anti-SCUBE2 mAb may provide a relatively nontoxic means of targeting tumor angiogenesis. As previously mentioned, we have shown that the mRNA and protein levels of SCUBE2, but not SCUBE1 or SCUBE3, are significantly upregulated after 12 h exposure of human umbilical vein endothelial cells (HUVECs) to hypoxia, and this upregulation occurs concomitantly with HIF-1α accumulation [23]. Additionally, our chromatin immunoprecipitation assay and promoter mutation analysis verified that endogenous HIF-1α can indeed bind to a DNA region containing a consensus HIF-binding motif (A/GCTGA) within the promoter region of SCUBE2, transactivating expression of SCUBE2 in HUVECs exposed to hypoxia [23]. Still, systematic toxicity studies are needed to determine the safety of combination treatments consisting of anti-SCUBE2 mAb and cytotoxic chemotherapeutic agents (e.g., gemcitabine or docetaxel) or anti-VEGF antibody (e.g., bevacizumab). Nevertheless, our studies to this point have shown that SCUBE2 acts a crucial VEGFR2 coreceptor during tumor angiogenesis, and targeting SCUBE2 on the cell surface may be an effective anti-angiogenic treatment strategy that may be especially useful in combination with cytotoxic chemotherapies.
Apart from its endothelial upregulation by pathological hypoxic conditions, our quantitative DNA methylation, methylation-specific PCR and chromatin immunoprecipitation analyses of breast tumor epithelial cells demonstrated that SCUBE2 expression is inactivated due to DNA hypermethylation at CpG islands during TGF-β–induced EMT, and this hypermethylation is due to the recruitment and binding of DNA methyltransferase 1 [51]. This mechanism was also highlighted in a recent study on the regulation of SCUBE2 methylation by (-)-epigallocatechin-3-gallate (EGCG), a highly abundant catechin in green tea. The authors found that treatment with EGCG reverses DNA methylation of SCUBE2 in breast cancer [161]. In addition, the study showed that EGCG inhibits breast cancer cell viability and suppresses the migration and invasion capacities of the cells, while knockdown of endogenous SCUBE2 blocks these inhibitory effects of EGCG in breast cancer cells. EGCG treatment was also shown to enhance SCUBE2 gene expression, elevate E-cadherin expression and decrease vimentin expression. Furthermore, it was revealed that EGCG significantly decreases the DNA methylation status of SCUBE2 by attenuating the expression and activity of DNA methyltransferase. These results suggest that pharmacological upregulation of the tumor suppressor SCUBE2 might be a potential therapeutic approach for breast cancer.
SCUBE3
Regulation of gene expression and posttranslational control
SCUBE3 is the third and last member of the SCUBE protein family to be identified. By conducting a homology search, we identified and characterized SCUBE3 in the human genome, as it shares an overall 60% protein identity and a similar protein domain organization with other SCUBE members (Fig. 1A, D). Quantitative RT-PCR and northern blot analyses revealed that the gene is prominently expressed in human primary cultured osteoblasts and mouse long bones, such as the humerus and femur; lower expression levels are present in human HUVECs and the heart [5]. In agreement with human SCUBE3 expression in osteoblasts and long bones, mouse Scube3 transcripts are expressed in the peripheral skeleton, including the ribs, limbs, and vertebra during cartilaginous condensation [129, 162, 163]. Similar to other SCUBE members, overexpressed human SCUBE3 protein exists as a secreted glycoprotein tethered to the cell surface that can form homodimers or heterodimers with other SCUBE members [5]. Secreted human SCUBE3 protein can be further proteolytically processed by an as yet unknown serum-associated protease to separate the N-terminal EGF-like repeats from the C-terminal CUB domain [5]. As seen with SCUBE1, SCUBE3 contains a minimal consensus sequence for furin-like protease (R537XXR540) within the spacer region, which might be responsible for its proteolytic cleavage in the presence of fetal bovine serum [5]. Based on its high expression in osteoblasts and long bones, we originally postulated that SCUBE3 might act locally and/or distally through a proteolytic mechanism to modulate bone cell homeostasis during development, repair and/or regeneration. In support of this hypothesis, recent human genetic studies have shown that bi-allelic inactivating variants in SCUBE3 on chromosome 6p21.3 contribute to a developmental disorder involving, dental abnormalities, certain skeletal features, growth retardation, and a distinctive craniofacial appearance [21, 164]. Likewise, screening of mouse ENU-induced mutagenesis phenotypes led to the identification of recessive mutations in Scube3 on chromosome 17qA3.3 that confer severe spine deformity, skeleton abnormalities, and altered bone metabolism (see below for details) [164, 165].
Developmental and physiological function
Hair growth
According to whole-mount and radiolabeled in situ hybridization, mouse Scube3 transcripts are primarily localized to the neuroectoderm at embryonic day (E)8.5. At E9.5 to E10.5, Scube3 expression is observed in the branchial arches and frontonasal region, as well as the ventral dermomyotome of somites and limb buds [162, 163]. Of note, expression of Scube3 in the central nervous system is restricted to ventral regions within the neural tube and cranial ganglia, including the trigeminal. In addition, mouse Scube3 mRNA is expressed in a number of tissues during embryonic development, including ectodermal, endodermal and mesodermal derivatives (Table 3). Scube3 transcripts are highly expressed in organs that develop via interactions between epithelium and mesenchyme, such as tooth germs and hair follicles [162, 163]. Indeed, a recent report showed that Scube3 is expressed only in the growing but not resting hair follicles in dermal papillae [166]. In the study, new hair was induced in response to the injection of SCUBE3 protein, and pharmacological inhibition of TGF-β could rescue a mutant hair hyper-activating phenotype [166]. The authors therefore concluded that HH regulates mesenchymal niche function in the hair follicle via SCUBE3–TGF-β signaling [166]. Thus, SCUBE3 was shown to be a mesenchymal niche factor that activates hair growth.
Of note, the first targeted mutation allele of Scube3 (Scube3tm1Dge/H) replaces the coding sequence of exons 2 and 3 with a neomycin-lacZ selectable marker cassette. Thus, only the EGF1-2 repeats are removed, and the allele still produces a partial Scube3 protein containing the C-terminal spacer region, CR and CUB domains. Since characterization of mice with this allele did not reveal any developmental abnormalities [163], it is possible that the partial Scube3 protein might retain functionality, at least during embryonic development [163]. Although this mutant line did not provide functional insights into the role of Scube3 during mouse development, it is a useful reporter that has been used to verify the strong expression of Scube3 in the epithelial compartments of developing teeth and hair follicles.
Chondrogenesis and osteogenesis
To further investigate the developmental functions of SCUBE3, a second targeted Scube3 allele was generated by replacing all of the coding exons with a ZEN-Ub1 promoter-driven expression-selection cassette [lacZ-(pA)-hUbiPro-neo-(pA)] [129]. The resulting tm1(KOPM)Vlcg allele contains a lacZ reporter for verification of its expression, and it was used to create heterozygous animals (Scube3+/−) on a C57/Bl6 background. Reporter activity domains in the Scube3+/− animals were generally consistent with the embryonic expression domains reported previously for wild-type Scube3 [162] or another Scube3tm1Dge/H reporter [163]. Interestingly, mice that are homozygous for the tm1(KOPM)Vlcg allele (Scube3−/−) are born at expected Mendelian ratios, show no overt embryonic phenotypes, and are both viable and fertile [129]. Furthermore, mouse Scube3 does not appear to affect the HH pathway, unlike zebrafish scube3, which cooperates with Scube2 to modulate HH signaling [116]. The Scube3-knockout embryos retain normal HH signaling activity, as indicated by expression of Ptch1, a known downstream transcriptional target of the HH pathway. Therefore, mouse Scube3, at least on a C57/Bl6 genetic background, seems to be dispensable for gross embryonic development and survival. These results suggest functional redundancy of Scube3 with other family members during development. However, the authors did not rule out a potentially exclusive role for the protein in postnatal tissue homeostasis or regeneration. In line with this suggestion, our analysis of this knockout mouse revealed that it has impaired BMP-mediated chondrogenesis and osteogenesis, which result in craniofacial and dental abnormalities, decreased body size, and faulty endochondral bone development (Fig. 8) [21]. Our molecular studies further demonstrated that membrane-associated SCUBE3 may function as a BMP2/4 coreceptor, attract BMP receptor complexes to raft microdomains, and enhance BMP signaling, perhaps by facilitating the interaction of BMP ligands with BMP type I receptors (Fig. 8) [21]. Consistent with these findings, mice homozygous for an ENU-induced mutant allele (Scube3N294K/N294K) exhibit skeletal abnormalities, including small body size, short kinked tails, abnormal digit positioning, malformations of the thoracic and lumbar vertebrae, and short femora, as well as other aberrations in parameters related to bone metabolism [165]. In this mutant allele, an asparagine-to-lysine exchange at protein position 294 (N294K) lies within the cbEGF7 repeat containing a calcium-binding consensus sequence [(D/N)-X-(D/N)-E/Q-Xm- (D/N*)-Xn-(Y/F) [17], where m or n are variable and the asterisk marks potential β-hydroxylation]. Our homology modeling suggested that the side-chain of β-hydroxylated N294 forms a hydrogen bond with D277 and serves as one metal-acceptor among oxygen atoms of other residues. In contrast, the N294K mutant forms a stronger hydrogen network with D277, I278 and E280, with the nitrogen of K294 likely occupying the position of calcium (Fig. 9A, B). Hence, the N294K exchange might lead to reduced calcium-binding capacity and/or conformational changes that affect the well-documented function of the cbEGF domains in mediating protein–protein interactions. Indeed, our molecular analysis showed that the SCUBE3-N294K mutant protein is expressed on the cell surface, but it does not readily form a complex with the BMP type IA receptor. This lack of interaction serves to abrogate osteogenic BMP2/4-induced signaling activity (Fig. 9C–F). Together, these results might provide a potential molecular explanation for the morphological abnormalities of the skeleton observed in this ENU-induced Scube3N294K/N294K mutant [165]. Similarly, another ENU-induced mutant allele (Scube3C301Y) was reported to cause recessive spine deformities, such as severe kyphosis and kinked tail [164]. As with the N294K mutation, this C301Y mutation resides within the cbEGF7 domain, and it is expected to disrupt a key disulfide bond between conserved C2-C4 cysteines that is likely essential for proper domain folding and function. Consequently, this disease-causing SCUBE3 missense mutation impairs the interaction between SCUBE3 and SCUBE1 and reduces Smad signaling strength [164].

Schematic diagram depicts the role of SCUBE3 in enhancing the BMP signaling critical for endochondral bone formation. A When SCUBE3 (+/+) is present, it facilitates BMP2/4 ligand binding to type II receptors (BMPR-II) and recruits BMP type I receptors (BMPRIA or BMPRIB) to generate a SCUBE3-enhanced signaling complex. This complex relocates to the lipid raft (green-colored area) in response to BMP stimulation, which increases activation of the signaling cascade required for concerted endochondral bone formation. B In the absence of SCUBE3, lipid rafts show reduced BMP2 and BMP receptor levels. Thus, BMP downstream signaling and function are attenuated. Scube3 deficiency in mice results in short stature, short-limbed skeletal dysplasia, misalignment of the front teeth, and kinked tail. In addition, bone mineral density is markedly lower in Scube3-deficient (−/−) mice compared to controls

SCUBE3-N294K mutant has a defective interaction with the BMP type IA receptor, diminishing osteogenic BMP signaling. A–B Close-up of SCUBE3 cbEGF7 domains in N294 and N294K mutant. 3-D structure of mouse SCUBE3 seventh EGF-like domain (D277 to C316) was extracted from the AlphaFold2 model (AF-Q66PY1-F1-model_v3) [187]. Calcium ion (green sphere) was taken from the fifth EGF-like domain of human NOTCH1 (PDB 5FM9) [221] and coordinated by the oxygen atoms of D277, E280 and N294 side-chains and carboxyl atoms of I278, T295 and S298 backbones (shown as pink dotted lines with carbon atoms colored in yellow, oxygen atoms in red, and nitrogen atoms in blue). Additionally, hydrogen bonds (red dotted lines) form between the nitrogen atom of the N294 side-chain and the carboxyl atom of the D277 side chain and between the hydroxyl atom of D277 and the backbone nitrogen of G297 to stabilize the overall conformation of the cbEGF7 domain (A). For the N294K mutant, the lysine residue is shown in ‘stick-mode’ with carbon atoms colored in cyan. The mutant K294 forms a hydrogen-bond network (red dotted lines) with oxygen atoms from the side-chains of D277 and E280 and backbone of I278. Of note, the nitrogen of K294 possibly excludes and occupies the position of calcium. This figure was produced using PyMOL (Schrödinger, LLC. The PyMOL molecular graphics system, version 1.8, https://pymol.org) (B). C Protein domain structure depicting the amino acid exchange of N294K lying within the calcium-binding seventh EGF-like domain (cbEGF7) of SCUBE3. D Cell-surface distribution of SCUBE3-wild-type (WT) and -N294K protein. The expression plasmids encoding FLAG-tagged SCUBE3-WT or -N294K were transfected in HepG2 cells for 2 days. Immunofluorescence staining of cells with anti-FLAG (red) antibody. Nuclei were visualized by DAPI staining (blue). Scale bar = 10 μm. E SCUBE3-N294K mutation blunts BMP2/4-stimulated signaling activity. The BMP-responsive luciferase reporter (BRE-luc) and pRL-TK alone or in combination with the designated expression vectors were transfected into HepG2 cells. Luciferase activity was assessed 24 h after transgenic cells were treated with or without BMP2 or BMP4 (50 ng/ml). Firefly luciferase values were normalized to Renilla activity to show relative luciferase activity. The experiments were performed 3 times in triplicate. Data are mean ± SD. **p < 0.01. F Effect of the SCUBE3-N294K mutant on interactions with BMP signaling cascade receptors. In HEK-293T cells, HA-tagged BMPR-IA, HA-tagged BMPR-IB, or Myc-tagged BMPR-II constructs were co-transfected with plasmids expressing FLAG-tagged SCUBE3-WT or -N294K. In order to identify protein–protein interactions, transfected cell extracts were immunoprecipitated after 48 h and used for western blot analysis with the designated antibodies
Together, the above mouse studies firmly implicate Scube3 as a critical regulator in bone morphology and metabolism, as well as body size and weight. Likewise, a genome-wide association study in humans identified SNPs of SCUBE3 among adult and pediatric height-associated loci [167, 168]. In pigs, SCUBE3 is also associated with body height, body length, and rump circumference [169]. Of note, apart from bone-related phenotypes, Scube3N429K mutant mice also showed disturbed renal function, hearing loss, altered inner and middle ear anatomy, and behavioral abnormalities [165].
Fast muscle development
In addition to performing studies on mice, we identified and characterized scube3 in the zebrafish model system [9]. In that study, we found that myod1 expression was selectively inhibited in lateral fast-muscle precursors upon scube3 knockdown by antisense morpholino oligonucleotides, but it was largely unaltered in adaxial slow-muscle precursors. Immunofluorescence labeling of fast- but not slow-muscle myosin was also significantly reduced in scube3 morphants. Further experiments revealed fgf8 as a crucial modulator of zebrafish scube3-mediated rapid muscle development. Additional biochemical and molecular research then showed that SCUBE3 enhances FGF8 signaling by acting as a FGF coreceptor [9]. Thus, zebrafish Scube3 may modify Fgf8 signaling during development in order to serve as an important upstream regulator of fast fiber myogenesis (Fig. 6B).
Pathological function
Cardiac hypertrophy
In addition to our loss-of-function studies in mice, we generated the first transgenic mice overexpressing SCUBE3 to further explore the function of SCUBE3 in vivo [170]. Expression was controlled by a type I collagen promoter, which induces strong expression of green fluorescent protein in isolated tail tendons and bone, but has relatively lower expression in other type I collagen-producing tissues [171, 172]. Male transgenic mice that overexpress SCUBE3 appear to grow normally from the embryonic period to adulthood. However, when the transgenic mice reach an age of ~ 8 months, the animals begin to display cardiac hypertrophy, as revealed by echocardiography and histopathological examination. Furthermore, pressure overload augments left-ventricle hypertrophy (i.e., it occurs more rapidly and severely) in transgenic animals. Our research indicates that the accelerated onset and progression of cardiac hypertrophy in transgenic mice may be due to pathological induction of SCUBE3 expression and elevated TGF-β1 level after pressure overload. Additionally, biochemical and molecular experiments showed that the C-terminal CR and CUB domains of SCUBE3 can interact with TGF-β1 and facilitate TGF-β1-mediated transcriptional activation [170]. Overall, this study suggested that upregulation of SCUBE3 modulates pathological TGF-β signaling to negatively impact cardiac growth and/or remodeling [170].
Lung cancer
In line with the pathological role of the SCUBE3-TGF-β signaling axis, our collaborative studies showed that soluble SCUBE3 can act via TGF-β to stimulate negative outcomes in lung cancer cells. In this context, SCUBE3 is cleaved by MMP-2 or MMP-9 to release its C-terminal CR and CUB domains. This fragment binds TGF-β receptors and elicits TGF-β-Smad2/3 signaling pathways, which drive EMT, migration and invasion of lung cancer cells as well as extracellular matrix deposition and angiogenesis [48]. Consistent with these findings, knockdown of SCUBE3 in a cellular model of NSCLC markedly reduces intratumor vessel density at the core region [173]. Moreover, microarray analysis of SCUBE3-knockdown lung tumors showed downregulation of angiogenic and EMT-related genes, including MMP-2, MMP-9, MMP-14, fibronectin, early growth response protein 1, hairy/enhancer-of-split related with YRPW motif protein 1, and interleukin 8 [173]. In agreement with these laboratory studies, a clinical association study correlated high SCUBE3 expression with low expression of the epithelial marker E-cadherin and high expression of the mesenchymal marker vimentin [174]. Importantly, the clinical data also showed that patients with high expression of SCUBE3 have a relatively poor survival rate, and SCUBE3 expression can serve as an independent prognostic factor for NSCLC [174] (Table 4).
Other cancer types
Consistent with the role of SCUBE3 in promoting angiogenesis in vitro and in vivo, a recent molecular study showed that DEP domain-containing 1B enhances melanoma angiogenesis and metastasis by stabilizing SCUBE3. DEP domain-containing 1B binds competitively with ubiquitin ligase cell division cycle 16, thereby preventing degradation of SCUBE3 via the ubiquitin–proteasome pathway [175]. Likewise, SCUBE3 promotes breast cancer progression via the activation of TGF-β1/TWIST1 signaling [176], and its expression can also independently serve as an indicator of poor prognosis in breast cancer [177]. In addition, SCUBE3 promotes hepatocellular carcinoma tumorigenesis by regulating cyclin E1 via the TGF-β/PI3K/AKT/GSK3β pathway [178]. SCUBE3 was also found to regulate glioma cell proliferation [179] and promote proliferation of osteosarcoma cells, and its expression is correlated prognosis of patients with osteosarcoma [180,181,182]. A public database search (www.proteinatlas.org) showed that high SCUBE3 protein expression can serve as a marker of unfavorable prognosis for human thyroid cancer (Table 4).
Besides these tumor-promoting functions of SCUBE3, genome-wide methylation analysis revealed that SCUBE3 is a tumor suppressor that may be epigenetically inactivated in renal cell carcinoma [183]. In this study, the authors used a genome-wide analysis to identify genes frequently methylated and silenced in renal cell carcinoma. Using methylated DNA immunoprecipitation and whole-genome arrays combined with high-density expression arrays, the study showed that the promoter region of SCUBE3 is frequently methylated in primary renal cell carcinoma tumor samples (19% of examined cases) [183]. Moreover, knockdown of SCUBE3 increased the occurrence of anchorage-independent colonies (> 200 μm) in HEK-293 cells by 71%. Also, methylation of SCUBE3 is associated with significantly increased overall risk of death (P = 0.009) and cancer death or relapse (p = 0.0046) in patients with renal cell carcinoma [183] (Table 4).
Skeletal disorders
In addition to the associations of aberrant SCUBE3 expression with pathogenesis and clinical outcomes of various types of cancers, mutations in SCUBE3 (GenBank: NM_152753, see Table 1) are associated with human skeletal disease. Eight bi-allelic inactivating variants in SCUBE3 were first discovered and characterized by our research in 18 affected people from nine independent families who shared the same phenotype of unusual craniofacial features, short stature, skeletal deformities and dental abnormalities [21]. Subsequently, another team identified different recessive SCUBE3 mutations in a patient with undiagnosed skeletal disease [164] (Fig. 10A, B). The discovered variations, which comprised missense, frameshift, nonsense, and canonical splice site modifications as well as a complicated intragenic rearrangement, were dispersed throughout the entire coding sequence of SCUBE3. Two variants are expected to alter transcript processing [c.2239 + 1G > A (p.Val747Aspfs*46); c.2599 + 2T > C], and the other three are expected to cause truncations [c1717C > T (p.Arg573*); c.2785C > T (p.Arg929*); c.1521_1522insGC (p.Ile508Alafs*74)]. Total RNA isolated from the affected patient carrying c.2239 + 1G > A confirmed that a cryptic donor site of exon 17 was activated, four intronic flanking nucleotides were retention, and the coding sequence (p.Val747Aspfs*46) was prematurely terminated. However, a biological sample from the proband carrying the c.2599 + 2T > C variant was unavailable for functional validation. In order to confirm the aberrant splicing of the genomic region of SCUBE3 containing exons 18 to 21, we employed a minigene splicing assay for a number of mRNA variants expected to yield prematurely truncated proteins (p.Asn801Thrfs*127 was the most common out-of-frame product) [21] (Fig. 10A, B).

Genomic organization, protein domains, and predicted protein domain structure of SCUBE3. A–B Graphic display of the genomic organization of SCUBE3 and the protein domain structure of its encoded product. Impacts of the disease-causing variants on protein coding are indicated. Our team first reported and characterized eight bi-allelic inactivating variants in SCUBE3 from 18 affected individuals of nine unrelated families who showed a similar phenotype of distinctive craniofacial appearance, dental anomalies, skeletal features, and reduced growth (closed red circles) [21]. Another team then identified recessive mutations of SCUBE3 from a patient with undiagnosed skeletal disease (open red circles) [164]. Two ENU-induced mutant alleles in Scube3 associated with spine deformity including severe kyphosis and kinked tail [164], as well as skeletal abnormalities and altered bone metabolism [165] are also shown (closed black triangles above the diagram) (A). Genomic location of the disease-causing variants of SCUBE3. C.2599 + 2T > C is predicted to result in multiple transcripts with abnormal processing, with p.Asn801Thrfs*127 representing the most common out-of-frame product [21]. Each exon is shown with size to scale, but introns are not. Also, the colors were used to coordinate specific protein domains and their coding exons. For instance, the signal peptide sequence and corresponding exon 1 are both shown in grey. In addition, exons 19 and 20, and the encoded the CUB domain are shown in purple (B). C–D Residues C97 and G204 of human SCUBE3 from AlphaFold2 model. Three disulfide bonds between six cysteine side chains are shown in ‘stick-mode’ in the second EGF-like domain of human SCUBE3; for instance, C97 (in cyan) connects with C110. These six cysteine residues are crucial for disulfide connectivity in structure folding and stability (C). G204 (in green) located on the fifth EGF-like domain is in contact with F101 of the second EGF-like domain and residues M130, M131, G132 and S133 of the third EGF-like domain (atoms colored with carbon in yellow, oxygen in red and nitrogen in blue) in this model. The substitution of residue G204 into D204 may perturb the interactions between the three EGF-like modules in human SCUBE3 (D). E The position of residue I815 in the CUB domain of human SCUBE3. Ribbon model represents human SCUBE3 CUB domain (C804 to Y916) from AlphaFold2 prediction (AF-Q8IX30-F1-model_v3) [187] with the β-strands numbered (in white) based on the bovine spermadhesin CUB domain [224]. I815 is shown in green, and residues within a distance of 5 Å are in yellow. Side chains of these residues buried in the hydrophobic core are shown in ‘stick-mode’ with oxygen atoms in red and nitrogen atoms in blue. Of note, I815 and surrounding residues, including I896, W832, L808, I834, Y913, I844, I911 and F909, form a cluster of buried hydrophobic amino acids in the core of the domain. Replacement of I815 with a polar threonine residue is predicted to destabilize the structure of the CUB domain
Four different missense changes were identified, all of which affect residues that are well-conserved among SCUBE members from different species [21]. A substitution c.291C > G (p.Cys97Trp) within the second EGF-like repeat affects one of the six conserved cysteines that form the disulfide bonds essential for stabilizing the EGF module structure. Thus, the protein should lose one of the three conserved disulfide bridges, causing a structural rearrangement of the EGF motif (Fig. 10C). Pathogenic variations in EGF-like modules at equivalent positions of fibrillin-1, fibrillin-2, and latent TGF-β binding protein 3 were previously reported [184,185,186], further supporting the putative functional importance of this variant. The second missense mutation [c.611G > A (p.Gly204Asp)] results in the exchange of a conserved glycine residue within the EGF5 module. The non-conservative substitution with aspartic acid is predicted to cause local structural conflicts with adjacent EGF-like modules (Fig. 10D). Based on the AlphaFold model of human SCUBE3 (CUB domains share 80% sequence identity) [187], a non-polar Ile815 reside is conserved and located on the β3 strand buried in the core of the CUB domain (Fig. 10E). The third missense variant [c.2444T > C (p.Ile815Thr)] involves a replacement of a non-polar isoleucine with a polar threonine. The structure of the hydrophobic core is expected to become unstable as a result of this replacement, which includes residues Leu808, Trp832, Ile834, Ile844, Ile896, Phe909, Ile911, and Tyr913 (Fig. 10E), thus perturbing the overall stability and folding of the CUB domain and its interaction with binding partners. The fourth missense mutation [c.2785C > G (pArg929Gly)] similarly changes a conserved positively charged arginine to a small glycine that might disrupt folding of the region and impair the proper function and binding ability of the CUB domain.
Only the cbEGF7 module is removed as a consequence of the complete deletion of exon 8 [c829_952del (p.Arg282_Cys322del)] by a complicated intragenic rearrangement in SCUBE3 (combined deletion and inverted duplication). Of note, two ENU-induced mutant alleles, Scube3N294K or Scube3C301Y, with skeletal phenotypes might also have detrimental effects on the folding and function of the cbEGF7 module [164, 165]. Moreover, comparable intragenic structural rearrangements that lead to deletion of a single EGF-like module in fibrillin 1 (FBN1) or fibrillin 2 (FBN2) are known to significantly affect protein functions [188, 189].
We [21] and others [164] used transient transfection to further investigate the effects of a representative panel of variants affecting different domains of SCUBE3 (pCys97Trp, p.Arg573*, pIle815Thr, or pCys301Tyr) in terms of protein stability, secretion, membrane anchoring and function. These studies revealed four major findings. (1) The p.Cys97Trp variant protein shows less-efficient dimerization with itself and with wild-type SCUBE3 [21]. (2) The pArg573* truncated variant protein is neither expressed on the cell surface nor is it secreted in the medium, which suggests the protein might be mistargeted due to a trafficking defect and endoplasmic reticulum retention [21]. (3) The pIle815Thr mutant is defective as a BMP coreceptor and cannot promote BMP signaling in vitro [21]. (4) The pCys301Tyr variant has reduced interaction with SCUBE1 (i.e., deficient formation of the SCUBE1-SCUBE3 heterodimer) and fails to activate Smad signaling [164]. Together, these findings suggest that variants affecting the EGF-like or CUB domain can act through diverse mechanisms to cause functional deficiencies. Since mouse ENU mutagenesis screening and human genetic studies reproducibly and repeatedly show that disruption of the cbEGF7 module [due to destabilized domain folding by two disease-associated missense mutations (N294K or C301Y) or complete domain deletion by a genomic structural variant] causes functional deficits, one may conclude that the cbEGF7 module plays a critical, potentially calcium-dependent role in regulating SCUBE3 signaling and activity. Nevertheless, this hypothesis warrants testing in further in-depth structural and functional investigations.
Consistent with the common clinical features of individuals carrying bi-allelic loss-of-function SCUBE3 variants, Scube3 knockout animals show reduced growth associated with small forelimbs and hindlimbs. Furthermore, Scube3 knockout mice typically exhibit misalignment of both upper and lower incisors, and the animals suffer from several craniofacial anomalies, such as a small forehead, short and narrow face, and short frontonasal and mandibular regions (Fig. 8) [21]. On occasion, it is also possible to find a knockout mouse with a hunchback spine or protruding ribs. Overall, these phenotypes recapitulate major features of SCUBE3-associated human skeletal disorders. Therefore, these clinical studies confirm SCUBE3 as the first SCUBE member involved in Mendelian disorders. Our studies and those from another team positively identified these mutant SCUBE3 alleles, which are primarily inactivating variants, as causing identifiable developmental abnormalities. Furthermore, our molecular characterization studies attribute this novel condition to the disruption of endochondral bone formation caused by defects in BMP signaling [21, 164].
Translational application and therapeutic strategy
To date, aberrant upregulation of SCUBE3 expression has been associated with unfavorable outcomes in various tumors, including NSCLC [181], breast cancer [181], metastatic melanoma [175], and osteosarcoma [181]. In contrast to these apparent oncogenic functions, SCUBE3 might also serve as a tumor suppressor in the context of renal cell carcinoma. In patients with this disease, promoter methylation (epigenetically silenced) of SCUBE3 is associated with increased risk of mortality or relapse [181]. Given that SCUBE3 is a secreted protein, it would be of great interest to determine whether elevated circulating soluble SCUBE3 protein might be associated with disease severity or progression. If so, the circulating protein may serve as a potential novel biomarker and/or a therapeutic target.
A recent study highlighted an interesting means of targeting SCUBE3 as a therapeutic strategy. In this study, SCUBE3 level was reduced by a new marine-isolated asterosaponin, which led to G1/S cell cycle arrest of glioma cells [179]. The authors showed that the newly identified asterosaponin, CN-3 (isolated from the starfish Crocodylus novaeguineae), could decrease the proliferation of two different glioma cell lines. Cellular and molecular studies revealed that the reduction of SCUBE3 expression by CN-3 is a key mediator of its anti-proliferative effects and cell cycle arrest via the AKT/p53/p21/p27/E2F1 signaling pathway [179]. Nevertheless, the function of SCUBE3 should be further explored in glioma, both in vitro and in vivo. In addition to chemical compounds that can modulate the expression of SCUBE3 under different pathological conditions, anti-SCUBE3 antibodies might be developed to specifically downregulate its cell-surface expression. The ultimate goal of any such method of targeting SCUBE3 would most likely be to suppress the pathological SCUBE3/TGF-β/Smad signaling pathway.
Conclusions
After two decades of research on the SCUBE family, many basic and translational discoveries have been made and reported (Fig. 11), especially regarding the roles of SCUBE proteins in various cancers (Table 4). However, many more biological functions of SCUBE proteins remain to be explored and will require further investigation. For example, the publicly available Allen Brain Atlas (https://portal.brain-map.org/) shows that SCUBE genes are expressed in various regions of the brain as well as spinal cord in mice and humans. Yet, little is known about the potential physiological or pathological functions of SCUBE proteins in neural tissues. This topic certainly warrants further in-depth neurobiological studies.

Summary of SCUBE-regulated growth factor signaling pathways and their involvements in physiological and pathological processes derived mainly from our studies. To date, different SCUBE members have been shown to modulate a diverse set of signal pathways activated by growth factors including BMP, FGF, TGF-β, VEGF, FLT3 ligand, and HH ligands. By these actions, SCUBE proteins participate in a wide variety of homeostatic and pathological processes
A Ramachandran plot analysis was performed to validate the accuracy of the SCUBE protein structures predicted by AlphaFold [190]; however, we still lack experimentally determined high-resolution structures of full proteins or key domains of SCUBE proteins. For instance, the impact of calcium binding on the structure and function of the cbEGF module or the cbCUB domain in SCUBE proteins is still undefined and uninvestigated. Furthermore, because SCUBE proteins interact with molecules in many different signaling pathways, we must unravel the structural basis of these interactions by conventional crystallization or by integrative approaches with cryogenic electron microscopy [191]. The structural insights derived from studies on SCUBE proteins in complex with signaling ligand binding partners (e.g., TGF-β, SHH, VEGF or FLT3 ligand) and their receptors might assist in the development of small-molecule inhibitors or antibody-based biologics. Such tools can potentially be used to antagonize disease-promoting signaling under pathological conditions, such as solid tumors or AML.
SCUBE1 is a multifunctional protein that has different functions based on its expression in different cell types. In several ischemic contexts, SCUBE1 is released into plasma and may be useful as biomarker for cardiovascular diseases [71, 81,82,83]. Its expression is also found in renal cancer, breast cancer and leukemia. Therefore, it could also be used as a biomarker in these cancers. However, detailed clinical analyses are needed to further validate the utility of circulating SCUBE1 levels as a biomarker for different conditions. In addition, although numerous studies have suggested that elevated plasma SCUBE1 levels might serve as biomarkers for different pathological conditions, these findings must be validated in prospective clinical trials on large patient cohorts.
SCUBE1 is also considered to be a promising therapeutic target in platelet activation and AML. With regard to platelet activation, an anti-SCUBE1 antibody was able to inhibit thrombus formation [72]. However, further studies are needed to uncover other promising methods to inhibit SCUBE1, such as small-molecule inhibitors to prevent thrombus formation. Likewise, in AML, anti-SCUBE1 ADC shows promising pre-clinical results in inhibiting disease progression [44]. Moreover, SCUBE1 could be an important target in AML and other types of leukemia due to its regulation by HOXA9, which is overexpressed in a broad range of leukemias [192]. Further analysis is needed to identify all the types of leukemia in which SCUBE1 is aberrantly upregulated. Furthermore, because SCUBE1 is expressed on the cell surface of leukemic blasts, methods such as chimeric antigen receptor T-cell therapy could be developed in addition to the ADC strategy that is currently being evaluated.
Another relatively unexplored field is the pathological implications and therapeutic targeting of the well-documented SCUBE2-mediated enhancement of HH signaling [113, 193]. Given that HH signaling plays a critical role in maintaining cancer stemness and progression of various diseases [194,195,196], further investigations are needed to explore how SCUBE2–HH signaling is involved in the progression of human pathologies, such as SHH-subgroup medulloblastoma [197]. Studies are also needed to develop treatments that can target SCUBE2-enhanced HH signaling.
Because enhancement of TGF-β signaling by SCUBE3 has been linked to a variety of cancers, understanding the molecular and structural basis of enhancement action could facilitate the development of more effective cancer therapy. In addition, a thorough molecular understanding of SCUBE3-mediated effects on BMP signaling could contribute to future treatments for craniofacial or bone defect in patients with SCUBE3 loss-of-function mutations. A number of critical DNA and protein reagents and many genetically modified animal models have already been established for investigating the SCUBE family. Further investigations of the molecular functions and structural characteristics of SCUBE proteins should include collaborations among basic and clinical researchers. Such collaborative projects may uncover the clinical potential and translational utility of secreted and membrane-bound SCUBE proteins as diagnostic, prognostic and/or therapeutic targets for human diseases.
Availability of data and materials
Not applicable.
Abbreviations
- ACS:
-
Acute coronary syndrome
- ADC:
-
Antibody–drug conjugate
- AIS:
-
Acute ischemic stroke
- AML:
-
Acute myeloid leukemia
- BMP:
-
Bone morphogenetic protein
- BMPR:
-
BMP receptor
- cbEGF:
-
Calcium binding EGF
- CR:
-
Cysteine-rich
- EC:
-
Endothelial cell
- EGCG:
-
(–)-Epigallocatechin-3-gallate
- EGF:
-
Epidermal growth factor
- EMT:
-
Epithelial-mesenchymal transition
- ENU:
-
Ethyl-N-nitrosourea
- FGF8:
-
Fibroblast growth factor 8
- FLT3:
-
Fms-like receptor tyrosine kinase 3
- HH:
-
Hedgehog
- HIF:
-
Hypoxia-inducible factor
- HOXA9:
-
Homeobox A9
- HUVECs:
-
Human umbilical vein endothelial cells
- IHH:
-
Indian hedgehog
- I/R:
-
Ischemia–reperfusion
- KO:
-
Knockout
- MAPK:
-
Mitogen-activated protein kinase
- MEIS1:
-
Meis homeobox 1
- MLL-r:
-
Leukemia gene-rearranged
- MMP:
-
Matrix metalloproteinase
- NSCLC:
-
Non-small cell lung cancer
- PAH:
-
Pulmonary arterial hypertension
- SCUBE:
-
Signal peptide-complement C1r/C1s, Uegf, Bmp1 (CUB)-epithelial growth factor domain-containing protein
- sCD40L:
-
Soluble CD40 ligand
- SHH:
-
Sonic hedgehog
- SNP:
-
Single nucleotide polymorphism
- STEMI:
-
Stent thrombosis-elevation myocardial infarction
- TGF-β:
-
Transforming growth factor beta
- VEGF:
-
Vascular growth factor
- VEGFR2:
-
Vascular growth factor receptor 2
References
Grimmond S, Larder R, Van Hateren N, Siggers P, Hulsebos TJ, Arkell R, et al. Cloning, mapping, and expression analysis of a gene encoding a novel mammalian EGF-related protein (SCUBE1). Genomics. 2000;70(1):74–81.
Yang RB, Ng CK, Wasserman SM, Colman SD, Shenoy S, Mehraban F, et al. Identification of a novel family of cell-surface proteins expressed in human vascular endothelium. J Biol Chem. 2002;277(48):46364–73.
Grimmond S, Larder R, Van Hateren N, Siggers P, Morse S, Hacker T, et al. Expression of a novel mammalian epidermal growth factor-related gene during mouse neural development. Mech Dev. 2001;102(1–2):209–11.
Tsai MT, Cheng CJ, Lin YC, Chen CC, Wu AR, Wu MT, et al. Isolation and characterization of a secreted, cell-surface glycoprotein SCUBE2 from humans. Biochem J. 2009;422(1):119–28.
Wu BT, Su YH, Tsai MT, Wasserman SM, Topper JN, Yang RB. A novel secreted, cell-surface glycoprotein containing multiple epidermal growth factor-like repeats and one CUB domain is highly expressed in primary osteoblasts and bones. J Biol Chem. 2004;279(36):37485–90.
Hollway GE, Maule J, Gautier P, Evans TM, Keenan DG, Lohs C, et al. Scube2 mediates Hedgehog signalling in the zebrafish embryo. Dev Biol. 2006;294(1):104–18.
Kawakami A, Nojima Y, Toyoda A, Takahoko M, Satoh M, Tanaka H, et al. The zebrafish-secreted matrix protein you/scube2 is implicated in long-range regulation of hedgehog signaling. Curr Biol. 2005;15(5):480–8.
Tsao KC, Tu CF, Lee SJ, Yang RB. Zebrafish scube1 (signal peptide-CUB (complement protein C1r/C1s, Uegf, and Bmp1)-EGF (epidermal growth factor) domain-containing protein 1) is involved in primitive hematopoiesis. J Biol Chem. 2013;288(7):5017–26.
Tu CF, Tsao KC, Lee SJ, Yang RB. SCUBE3 (signal peptide-CUB-EGF domain-containing protein 3) modulates fibroblast growth factor signaling during fast muscle development. J Biol Chem. 2014;289(27):18928–42.
Woods IG, Talbot WS. The you gene encodes an EGF-CUB protein essential for Hedgehog signaling in zebrafish. PLoS Biol. 2005;3(3): e66.
Davis CG. The many faces of epidermal growth factor repeats. New Biol. 1990;2(5):410–9.
Balzar M, Briaire-de Bruijn IH, Rees-Bakker HA, Prins FA, Helfrich W, de Leij L, et al. Epidermal growth factor-like repeats mediate lateral and reciprocal interactions of Ep-CAM molecules in homophilic adhesions. Mol Cell Biol. 2001;21(7):2570–80.
Ishii J, Adachi H, Aoki J, Koizumi H, Tomita S, Suzuki T, et al. SREC-II, a new member of the scavenger receptor type F family, trans-interacts with SREC-I through its extracellular domain. J Biol Chem. 2002;277(42):39696–702.
Prieto JH, Sampoli Benitez BA, Melacini G, Johnson DA, Wood MJ, Komives EA. Dynamics of the fragment of thrombomodulin containing the fourth and fifth epidermal growth factor-like domains correlate with function. Biochemistry. 2005;44(4):1225–33.
Luca VC, Kim BC, Ge C, Kakuda S, Wu D, Roein-Peikar M, et al. Notch-Jagged complex structure implicates a catch bond in tuning ligand sensitivity. Science. 2017;355(6331):1320–4.
Lo Surdo P, Bottomley MJ, Calzetta A, Settembre EC, Cirillo A, Pandit S, et al. Mechanistic implications for LDL receptor degradation from the PCSK9/LDLR structure at neutral pH. EMBO Rep. 2011;12(12):1300–5.
Campbell ID, Bork P. Epidermal growth factor-like modules. Curr Opin Struct Biol. 1993;3(3):385–92.
Stacey M, Chang GW, Davies JQ, Kwakkenbos MJ, Sanderson RD, Hamann J, et al. The epidermal growth factor-like domains of the human EMR2 receptor mediate cell attachment through chondroitin sulfate glycosaminoglycans. Blood. 2003;102(8):2916–24.
Periz J, Gill AC, Knott V, Handford PA, Tomley FM. Calcium binding activity of the epidermal growth factor-like domains of the apicomplexan microneme protein EtMIC4. Mol Biochem Parasitol. 2005;143(2):192–9.
Tu CF, Yan YT, Wu SY, Djoko B, Tsai MT, Cheng CJ, et al. Domain and functional analysis of a novel platelet-endothelial cell surface protein, SCUBE1. J Biol Chem. 2008;283(18):12478–88.
Lin YC, Niceta M, Muto V, Vona B, Pagnamenta AT, Maroofian R, et al. SCUBE3 loss-of-function causes a recognizable recessive developmental disorder due to defective bone morphogenetic protein signaling. Am J Hum Genet. 2021;108(1):115–33.
Lin YC, Chen CC, Cheng CJ, Yang RB. Domain and functional analysis of a novel breast tumor suppressor protein, SCUBE2. J Biol Chem. 2011;286(30):27039–47.
Lin YC, Chao TY, Yeh CT, Roffler SR, Kannagi R, Yang RB. Endothelial SCUBE2 interacts with VEGFR2 and regulates VEGF-induced angiogenesis. Arterioscler Thromb Vasc Biol. 2017;37(1):144–55.
Lin YC, Liu CY, Kannagi R, Yang RB. Inhibition of endothelial SCUBE2 (signal peptide-CUB-EGF domain-containing protein 2), a novel VEGFR2 (vascular endothelial growth factor receptor 2) coreceptor, suppresses tumor angiogenesis. Arterioscler Thromb Vasc Biol. 2018;38(5):1202–15.
Mellquist JL, Kasturi L, Spitalnik SL, Shakin-Eshleman SH. The amino acid following an asn-X-Ser/Thr sequon is an important determinant of N-linked core glycosylation efficiency. Biochemistry. 1998;37(19):6833–7.
Liao WJ, Tsao KC, Yang RB. Electrostatics and N-glycan-mediated membrane tethering of SCUBE1 is critical for promoting bone morphogenetic protein signalling. Biochem J. 2016;473(5):661–72.
Bork P, Beckmann G. The CUB domain. A widespread module in developmentally regulated proteins. J Mol Biol. 1993;231(2):539–45.
Lee HX, Mendes FA, Plouhinec JL, De Robertis EM. Enzymatic regulation of pattern: BMP4 binds CUB domains of Tolloids and inhibits proteinase activity. Genes Dev. 2009;23(21):2551–62.
Klagsbrun M, Eichmann A. A role for axon guidance receptors and ligands in blood vessel development and tumor angiogenesis. Cytokine Growth Factor Rev. 2005;16(4–5):535–48.
Li L, Wu D, Qin X, Mi LZ. PDGF-D prodomain differentially inhibits the biological activities of PDGF-D and PDGF-B. J Mol Biol. 2022;434(16): 167709.
Christensen EI, Birn H. Megalin and cubilin: multifunctional endocytic receptors. Nat Rev Mol Cell Biol. 2002;3(4):256–66.
Kangro K, Roose E, Joly BS, Sinkovits G, Falter T, von Auer C, et al. Anti-ADAMTS13 autoantibody profiling in patients with immune-mediated thrombotic thrombocytopenic purpura. Blood Adv. 2021;5(17):3427–35.
Romero A, Romão MJ, Varela PF, Kölln I, Dias JM, Carvalho AL, et al. The crystal structures of two spermadhesins reveal the CUB domain fold. Nat Struct Biol. 1997;4(10):783–8.
Kiragasi B, Goel P, Perry S, Han Y, Li X, Dickman D. The auxiliary glutamate receptor subunit dSol-1 promotes presynaptic neurotransmitter release and homeostatic potentiation. Proc Natl Acad Sci USA. 2020;117(41):25830–9.
McGovern JA, Heinemann JR, Burke LJ, Dawson R, Parker TJ, Upton Z, et al. Stratum basale keratinocyte expression of the cell-surface glycoprotein CDCP1 during epidermogenesis and its role in keratinocyte migration. Br J Dermatol. 2013;168(3):496–503.
Kang W, Reid KB. DMBT1, a regulator of mucosal homeostasis through the linking of mucosal defense and regeneration? FEBS Lett. 2003;540(1–3):21–5.
Gregory LA, Thielens NM, Arlaud GJ, Fontecilla-Camps JC, Gaboriaud C. X-ray structure of the Ca2+-binding interaction domain of C1s. Insights into the assembly of the C1 complex of complement. J Biol Chem. 2003;278(34):32157–64.
Lun Y, Borjini N, Miura NN, Ohno N, Singer NG, Lin F. CDCP1 on dendritic cells contributes to the development of a model of Kawasaki disease. J Immunol. 2021;206(12):2819–27.
Gaboriaud C, Gregory-Pauron L, Teillet F, Thielens NM, Bally I, Arlaud GJ. Structure and properties of the Ca(2+)-binding CUB domain, a widespread ligand-recognition unit involved in major biological functions. Biochem J. 2011;439(2):185–93.
Gaboriaud C, Teillet F, Gregory LA, Thielens NM, Arlaud GJ. Assembly of C1 and the MBL- and ficolin-MASP complexes: structural insights. Immunobiology. 2007;212(4–5):279–88.
Gregory LA, Thielens NM, Matsushita M, Sorensen R, Arlaud GJ, Fontecilla-Camps JC, et al. The X-ray structure of human mannan-binding lectin-associated protein 19 (MAp19) and its interaction site with mannan-binding lectin and L-ficolin. J Biol Chem. 2004;279(28):29391–7.
Andersen CB, Madsen M, Storm T, Moestrup SK, Andersen GR. Structural basis for receptor recognition of vitamin-B(12)-intrinsic factor complexes. Nature. 2010;464(7287):445–8.
Bally I, Rossi V, Lunardi T, Thielens NM, Gaboriaud C, Arlaud GJ. Identification of the C1q-binding sites of human C1r and C1s: a refined three-dimensional model of the C1 complex of complement. J Biol Chem. 2009;284(29):19340–8.
Sahoo BK, Lin YC, Tu CF, Lin CC, Liao WJ, Li FA, et al. Signal peptide-CUB-EGF-like repeat-containing protein 1-promoted FLT3 signaling is critical for the initiation and maintenance of MLL-rearranged acute leukemia. Haematologica. 2023;108(5):1284–99.
Lin YC, Roffler SR, Yan YT, Yang RB. Disruption of Scube2 impairs endochondral bone formation. J Bone Miner Res. 2015;30(7):1255–67.
Creanga A, Glenn TD, Mann RK, Saunders AM, Talbot WS, Beachy PA. Scube/You activity mediates release of dually lipid-modified Hedgehog signal in soluble form. Genes Dev. 2012;26(12):1312–25.
Tukachinsky H, Kuzmickas RP, Jao CY, Liu J, Salic A. Dispatched and scube mediate the efficient secretion of the cholesterol-modified hedgehog ligand. Cell Rep. 2012;2(2):308–20.
Wu YY, Peck K, Chang YL, Pan SH, Cheng YF, Lin JC, et al. SCUBE3 is an endogenous TGF-beta receptor ligand and regulates the epithelial–mesenchymal transition in lung cancer. Oncogene. 2011;30(34):3682–93.
Liao WJ, Lin H, Cheng CF, Ka SM, Chen A, Yang RB. SCUBE1-enhanced bone morphogenetic protein signaling protects against renal ischemia-reperfusion injury. Biochim Biophys Acta Mol Basis Dis. 2019;1865(2):329–38.
Lincoln TM. Pertussis toxin-sensitive and insensitive guanine nucleotide binding proteins (G-proteins) are not phosphorylated by cyclic GMP-dependent protein kinase. Second Messengers & Phosphoproteins. 1991;13:99–109.
Lin YC, Lee YC, Li LH, Cheng CJ, Yang RB. Tumor suppressor SCUBE2 inhibits breast-cancer cell migration and invasion through the reversal of epithelial-mesenchymal transition. J Cell Sci. 2014;127(Pt 1):85–100.
Xavier GM, Sharpe PT, Cobourne MT. Scube1 is expressed during facial development in the mouse. J Exp Zool B Mol Dev Evol. 2009;312B(5):518–24.
Vanpoucke G, Orr B, Grace OC, Chan R, Ashley GR, Williams K, et al. Transcriptional profiling of inductive mesenchyme to identify molecules involved in prostate development and disease. Genome Biol. 2007;8(10):R213.
Heit JA, Cunningham JM, Petterson TM, Armasu SM, Rider DN. Genetic variation within the anticoagulant, procoagulant, fibrinolytic and innate immunity pathways as risk factors for venous thromboembolism. J Thromb Haemost. 2011;9(6):1133–42.
Cappi C, Oliphant ME, Péter Z, Zai G, Conceição do Rosário M, Sullivan CAW, et al. De novo damaging DNA coding mutations are associated with obsessive-compulsive disorder and overlap with tourette’s disorder and autism. Biol Psychiatry. 2020;87(12):1035–44.
Ma X, Liu Y, Liu Y, Alexandrov LB, Edmonson MN, Gawad C, et al. Pan-cancer genome and transcriptome analyses of 1,699 paediatric leukaemias and solid tumours. Nature. 2018;555(7696):371–6.
Argiropoulos B, Humphries RK. Hox genes in hematopoiesis and leukemogenesis. Oncogene. 2007;26(47):6766–76.
Ramos-Mejia V, Navarro-Montero O, Ayllon V, Bueno C, Romero T, Real PJ, et al. HOXA9 promotes hematopoietic commitment of human embryonic stem cells. Blood. 2014;124(20):3065–75.
Zhuang J, Deane JA, Yang RB, Li J, Ricardo SD. SCUBE1, a novel developmental gene involved in renal regeneration and repair. Nephrol Dial Transpl. 2010;25(5):1421–8.
Anderson RM, Lawrence AR, Stottmann RW, Bachiller D, Klingensmith J. Chordin and noggin promote organizing centers of forebrain development in the mouse. Development. 2002;129(21):4975–87.
Stottmann RW, Berrong M, Matta K, Choi M, Klingensmith J. The BMP antagonist Noggin promotes cranial and spinal neurulation by distinct mechanisms. Dev Biol. 2006;295(2):647–63.
Tu CF, Su YH, Huang YN, Tsai MH, Li LT, Chen YL, et al. Localization and characterization of a novel protein SCUBE1 in human platelets. Cardiovasc Res. 2006;71(3):486–95.
Blann AD, Nadar SK, Lip GY. The adhesion molecule P-selectin and cardiovascular disease. Eur Heart J. 2003;24(24):2166–79.
Lenting PJ, Christophe OD, Denis CV. von Willebrand factor biosynthesis, secretion, and clearance: connecting the far ends. Blood. 2015;125(13):2019–28.
Suzuki K, Nishioka J, Hayashi T, Kosaka Y. Functionally active thrombomodulin is present in human platelets. J Biochem (Tokyo). 1988;104(4):628–32.
Bergsten E, Uutela M, Li X, Pietras K, Ostman A, Heldin CH, et al. PDGF-D is a specific, protease-activated ligand for the PDGF beta-receptor. Nat Cell Biol. 2001;3(5):512–6.
Gilbertson DG, Duff ME, West JW, Kelly JD, Sheppard PO, Hofstrand PD, et al. Platelet-derived growth factor C (PDGF-C), a novel growth factor that binds to PDGF alpha and beta receptor. J Biol Chem. 2001;276(29):27406–14.
LaRochelle WJ, Jeffers M, McDonald WF, Chillakuru RA, Giese NA, Lokker NA, et al. PDGF-D, a new protease-activated growth factor. Nat Cell Biol. 2001;3(5):517–21.
Li X, Ponten A, Aase K, Karlsson L, Abramsson A, Uutela M, et al. PDGF-C is a new protease-activated ligand for the PDGF alpha-receptor. Nat Cell Biol. 2000;2(5):302–9.
Duckert P, Brunak S, Blom N. Prediction of proprotein convertase cleavage sites. Protein Eng Des Sel. 2004;17(1):107–12.
Dai DF, Thajeb P, Tu CF, Chiang FT, Chen CH, Yang RB, et al. Plasma concentration of SCUBE1, a novel platelet protein, is elevated in patients with acute coronary syndrome and ischemic stroke. J Am Coll Cardiol. 2008;51(22):2173–80.
Wu MY, Lin YC, Liao WJ, Tu CF, Chen MH, Roffler SR, et al. Inhibition of the plasma SCUBE1, a novel platelet adhesive protein, protects mice against thrombosis. Arterioscler Thromb Vasc Biol. 2014;34(7):1390–8.
Kasirer-Friede A, Frojmovic MM. Ristocetin- and thrombin-induced platelet aggregation at physiological shear rates: differential roles for GPIb and GPIIb-IIIa receptor. Thromb Haemost. 1998;80(3):428–36.
Kauskot A, Di Michele M, Loyen S, Freson K, Verhamme P, Hoylaerts MF. A novel mechanism of sustained platelet alphaIIbbeta3 activation via PEAR1. Blood. 2012;119(17):4056–65.
Huo Y, Schober A, Forlow SB, Smith DF, Hyman MC, Jung S, et al. Circulating activated platelets exacerbate atherosclerosis in mice deficient in apolipoprotein E. Nat Med. 2003;9(1):61–7.
Dong ZM, Brown AA, Wagner DD. Prominent role of P-selectin in the development of advanced atherosclerosis in ApoE-deficient mice. Circulation. 2000;101(19):2290–5.
Mach F, Schonbeck U, Sukhova GK, Atkinson E, Libby P. Reduction of atherosclerosis in mice by inhibition of CD40 signalling. Nature. 1998;394(6689):200–3.
Methia N, Andre P, Denis CV, Economopoulos M, Wagner DD. Localized reduction of atherosclerosis in von Willebrand factor-deficient mice. Blood. 2001;98(5):1424–8.
Kim HC. Epidemiology of cardiovascular disease and its risk factors in Korea. Glob Health Med. 2021;3(3):134–41.
Mackman N. Triggers, targets and treatments for thrombosis. Nature. 2008;451(7181):914–8.
Bolayir HA, Gunes H, Kivrak T, Sahin O, Akaslan D, Kurt R, et al. The role of SCUBE1 in the pathogenesis of no-reflow phenomenon presenting with ST segment elevation myocardial infarction. Anatol J Cardiol. 2017;18(2):122–7.
Bolayir HA, Kivrak T, Gunes H, Akaslan D, Sahin O, Bolayir A. The role of SCUBE1 in the development of late stent thrombosis presenting with ST-elevation myocardial infarction. Rev Port Cardiol (Engl Ed). 2018;37(5):375–81.
Guzel M, Dogru MT, Simsek V, Demir V, Alp C, Kandemir H, et al. Influence of circadian blood pressure alterations on serum SCUBE-1 and soluble CD40 ligand levels in patients with essential hypertension. Am J Cardiovasc Dis. 2019;9(4):42–8.
Gori T, Polimeni A, Indolfi C, Raber L, Adriaenssens T, Munzel T. Predictors of stent thrombosis and their implications for clinical practice. Nat Rev Cardiol. 2019;16(4):243–56.
D’Ascenzo F, Bollati M, Clementi F, Castagno D, Lagerqvist B, de la Torre Hernandez JM, et al. Incidence and predictors of coronary stent thrombosis: evidence from an international collaborative meta-analysis including 30 studies, 221,066 patients, and 4276 thromboses. Int J Cardiol. 2013;167(2):575–84.
Liao WJ, Wu MY, Peng CC, Tung YC, Yang RB. Epidermal growth factor-like repeats of SCUBE1 derived from platelets are critical for thrombus formation. Cardiovasc Res. 2020;116(1):193–201.
Chakrabarti S, Varghese S, Vitseva O, Tanriverdi K, Freedman JE. CD40 ligand influences platelet release of reactive oxygen intermediates. Arterioscler Thromb Vasc Biol. 2005;25(11):2428–34.
Prasad KS, Andre P, He M, Bao M, Manganello J, Phillips DR. Soluble CD40 ligand induces beta3 integrin tyrosine phosphorylation and triggers platelet activation by outside-in signaling. Proc Natl Acad Sci USA. 2003;100(21):12367–71.
Heeschen C, Dimmeler S, Hamm CW, van den Brand MJ, Boersma E, Zeiher AM, et al. Soluble CD40 ligand in acute coronary syndromes. N Engl J Med. 2003;348(12):1104–11.
Chew M, Rahman M, Ihrman L, Erson A, Zhang S, Thorlacius H. Soluble CD40L (CD154) is increased in patients with shock. Inflamm Res. 2010;59(11):979–82.
Ulusoy S, Ozkan G, Mentese A, Yavuz A, Karahan SC, Sumer AU. Signal peptide-CUB-EGF domain-containing protein 1 (SCUBE1) level in hemodialysis patients and parameters affecting that level. Clin Biochem. 2012;45(16–17):1444–9.
Ozkan G, Ulusoy S, Mentese A, Karahan SC, Cansiz M. New marker of platelet activation, SCUBE1, is elevated in hypertensive patients. Am J Hypertens. 2013;26(6):748–53.
Bilir B, Soysal-Atile N, Ekiz Bilir B, Yilmaz I, Bali I, Altintas N, et al. Evaluation of SCUBE-1 and sCD40L biomarkers in patients with hypothyroidism due to Hashimoto’s thyroiditis: a single-blind, controlled clinical study. Eur Rev Med Pharmacol Sci. 2016;20(3):407–13.
Mancia G, Fagard R, Narkiewicz K, Redon J, Zanchetti A, Bohm M, et al. 2013 ESH/ESC guidelines for the management of arterial hypertension: the Task Force for the Management of Arterial Hypertension of the European Society of Hypertension (ESH) and of the European Society of Cardiology (ESC). Eur Heart J. 2013;34(28):2159–219.
Hoeper MM, Ghofrani HA, Grunig E, Klose H, Olschewski H, Rosenkranz S. Pulmonary hypertension. Dtsch Arztebl Int. 2017;114(5):73–84.
Farber HW, Miller DP, Poms AD, Badesch DB, Frost AE, Muros-Le Rouzic E, et al. Five-year outcomes of patients enrolled in the REVEAL registry. Chest. 2015;148(4):1043–54.
Sun W, Tang Y, Tai YY, Handen A, Zhao J, Speyer G, et al. SCUBE1 controls BMPR2-relevant pulmonary endothelial function: implications for diagnostic marker development in pulmonary arterial hypertension. JACC Basic Transl Sci. 2020;5(11):1073–92.
Almanzar MM, Frazier KS, Dube PH, Piqueras AI, Jones WK, Charette MF, et al. Osteogenic protein-1 mRNA expression is selectively modulated after acute ischemic renal injury. J Am Soc Nephrol. 1998;9(8):1456–63.
Vukicevic S, Basic V, Rogic D, Basic N, Shih MS, Shepard A, et al. Osteogenic protein-1 (bone morphogenetic protein-7) reduces severity of injury after ischemic acute renal failure in rat. J Clin Invest. 1998;102(1):202–14.
Barabe F, Gil L, Celton M, Bergeron A, Lamontagne V, Roques E, et al. Modeling human MLL-AF9 translocated acute myeloid leukemia from single donors reveals RET as a potential therapeutic target. Leukemia. 2017;31(5):1166–76.
Hartley-Brown M, Richardson P. Antibody-drug conjugate therapies in multiple myeloma-what’s next on the horizon? Explor Target Antitumor Ther. 2022;3(1):1–10.
Fan C, Oh DS, Wessels L, Weigelt B, Nuyten DS, Nobel AB, et al. Concordance among gene-expression-based predictors for breast cancer. N Engl J Med. 2006;355(6):560–9.
van’t Veer LJ, Dai H, van de Vijver MJ, He YD, Hart AA, Mao M, et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature. 2002;415(6871):530–6.
van de Vijver MJ, He YD, van't Veer LJ, Dai H, Hart AA, Voskuil DW, et al. A gene-expression signature as a predictor of survival in breast cancer. N Engl J Med. 2002;347(25):1999–2009.
Paik S, Shak S, Tang G, Kim C, Baker J, Cronin M, et al. A multigene assay to predict recurrence of tamoxifen-treated, node-negative breast cancer. N Engl J Med. 2004;351(27):2817–26.
Paik S, Tang G, Shak S, Kim C, Baker J, Kim W, et al. Gene expression and benefit of chemotherapy in women with node-negative, estrogen receptor-positive breast cancer. J Clin Oncol. 2006;24(23):3726–34.
Chen MH, Li YJ, Kawakami T, Xu SM, Chuang PT. Palmitoylation is required for the production of a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. Genes Dev. 2004;18(6):641–59.
Lewis PM, Dunn MP, McMahon JA, Logan M, Martin JF, St-Jacques B, et al. Cholesterol modification of sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell. 2001;105(5):599–612.
Jakobs P, Exner S, Schurmann S, Pickhinke U, Bandari S, Ortmann C, et al. Scube2 enhances proteolytic Shh processing from the surface of Shh-producing cells. J Cell Sci. 2014;127(Pt 8):1726–37.
Jakobs P, Schulz P, Ortmann C, Schurmann S, Exner S, Rebollido-Rios R, et al. Bridging the gap: heparan sulfate and Scube2 assemble Sonic hedgehog release complexes at the surface of producing cells. Sci Rep. 2016;6:26435.
Jakobs P, Schulz P, Schurmann S, Niland S, Exner S, Rebollido-Rios R, et al. Ca2+ coordination controls sonic hedgehog structure and its Scube2-regulated release. J Cell Sci. 2017;130(19):3261–71.
Cheng CJ, Lin YC, Tsai MT, Chen CS, Hsieh MC, Chen CL, et al. SCUBE2 suppresses breast tumor cell proliferation and confers a favorable prognosis in invasive breast cancer. Cancer Res. 2009;69(8):3634–41.
Wierbowski BM, Petrov K, Aravena L, Gu G, Xu Y, Salic A. Hedgehog pathway activation requires coreceptor-catalyzed, lipid-dependent relay of the sonic hedgehog ligand. Dev Cell. 2020;55(4):450-67.e8.
Athanasiadis G, Sabater-Lleal M, Buil A, Souto JC, Borrell M, Lathrop M, et al. Genetic determinants of plasma β2-glycoprotein I levels: a genome-wide association study in extended pedigrees from Spain. J Thromb Haemost. 2013;11(3):521–8.
Dueker ND, Beecham A, Wang L, Dong C, Sacco RL, Blanton SH, et al. Rare variants in previously identified linkage regions associated with carotid plaque in Dominican Republic families. PLoS ONE. 2022;17(1): e0250799.
Johnson JL, Hall TE, Dyson JM, Sonntag C, Ayers K, Berger S, et al. Scube activity is necessary for Hedgehog signal transduction in vivo. Dev Biol. 2012;368(2):193–202.
Corallo D, Schiavinato A, Trapani V, Moro E, Argenton F, Bonaldo P. Emilin3 is required for notochord sheath integrity and interacts with Scube2 to regulate notochord-derived Hedgehog signals. Development. 2013;140(22):4594–601.
Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol. 2005;6(4):306–17.
McMahon AP, Ingham PW, Tabin CJ. Developmental roles and clinical significance of hedgehog signaling. Curr Top Dev Biol. 2003;53:1–114.
Chung UI, Schipani E, McMahon AP, Kronenberg HM. Indian hedgehog couples chondrogenesis to osteogenesis in endochondral bone development. J Clin Invest. 2001;107(3):295–304.
Long F, Chung UI, Ohba S, McMahon J, Kronenberg HM, McMahon AP. Ihh signaling is directly required for the osteoblast lineage in the endochondral skeleton. Development. 2004;131(6):1309–18.
Long F, Zhang XM, Karp S, Yang Y, McMahon AP. Genetic manipulation of hedgehog signaling in the endochondral skeleton reveals a direct role in the regulation of chondrocyte proliferation. Development. 2001;128(24):5099–108.
Mak KK, Kronenberg HM, Chuang PT, Mackem S, Yang Y. Indian hedgehog signals independently of PTHrP to promote chondrocyte hypertrophy. Development. 2008;135(11):1947–56.
St-Jacques B, Hammerschmidt M, McMahon AP. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999;13(16):2072–86.
Maeda Y, Nakamura E, Nguyen MT, Suva LJ, Swain FL, Razzaque MS, et al. Indian Hedgehog produced by postnatal chondrocytes is essential for maintaining a growth plate and trabecular bone. Proc Natl Acad Sci USA. 2007;104(15):6382–7.
Xavier GM, Cobourne MT. Scube2 expression extends beyond the central nervous system during mouse development. J Mol Histol. 2011;42(5):383–91.
Colnot C, Lu C, Hu D, Helms JA. Distinguishing the contributions of the perichondrium, cartilage, and vascular endothelium to skeletal development. Dev Biol. 2004;269(1):55–69.
Gelfand MV, Hagan N, Tata A, Oh WJ, Lacoste B, Kang KT, et al. Neuropilin-1 functions as a VEGFR2 co-receptor to guide developmental angiogenesis independent of ligand binding. Elife. 2014;3: e03720.
Xavier GM, Panousopoulos L, Cobourne MT. Scube3 is expressed in multiple tissues during development but is dispensable for embryonic survival in the mouse. PLoS ONE. 2013;8(1): e55274.
Tsao KC, Lin YC, Chen YT, Lai SL, Yang RB. Zebrafish Scube1 and Scube2 cooperate in promoting Vegfa signalling during embryonic vascularization. Cardiovasc Res. 2022;118(4):1074–87.
Poorhosseini SM, Hashemi M, Alipour Olyaei N, Izadi A, Moslemi E, Ravesh Z, et al. New gene profiling in determination of breast cancer recurrence and prognosis in Iranian women. Asian Pac J Cancer Prev. 2016;17(S3):155–60.
Esmaeili R, Mohammadi S, Jafarbeik-Iravani N, Yadegari F, Olfatbakhsh A, Mazaheri M, et al. Expression of SCUBE2 and BCL2 predicts favorable response in ERalpha positive breast cancer. Arch Iran Med. 2021;24(3):209–17.
Shuai C, Yuan F, Liu Y, Wang C, Wang J, He H. Estrogen receptor-positive breast cancer survival prediction and analysis of resistance-related genes introduction. PeerJ. 2021;9: e12202.
Andres SA, Wittliff JL. Co-expression of genes with estrogen receptor-alpha and progesterone receptor in human breast carcinoma tissue. Horm Mol Biol Clin Investig. 2012;12(1):377–90.
Ni H, Kumbrink J, Mayr D, Seiler A, Hagemann F, Degenhardt T, et al. Molecular prognostic factors for distant metastases in premenopausal patients with HR+/HER2− early breast cancer. J Pers Med. 2021;11(9):835.
Andres SA, Brock GN, Wittliff JL. Interrogating differences in expression of targeted gene sets to predict breast cancer outcome. BMC Cancer. 2013;13:326.
Liu M, Zhou S, Wang J, Zhang Q, Yang S, Feng J, et al. Identification of genes associated with survival of breast cancer patients. Breast Cancer. 2019;26(3):317–25.
Ahmed W, Malik MFA, Saeed M, Haq F. Copy number profiling of Oncotype DX genes reveals association with survival of breast cancer patients. Mol Biol Rep. 2018;45(6):2185–92.
Fatima A, Tariq F, Malik MFA, Qasim M, Haq F. Copy number profiling of MammaPrint genes reveals association with the prognosis of breast cancer patients. J Breast Cancer. 2017;20(3):246–53.
Jinesh GG, Flores ER, Brohl AS. Chromosome 19 miRNA cluster and CEBPB expression specifically mark and potentially drive triple negative breast cancers. PLoS ONE. 2018;13(10): e0206008.
Fei H, Chen S, Xu C. RNA-sequencing and microarray data mining revealing: the aberrantly expressed mRNAs were related with a poor outcome in the triple negative breast cancer patients. Ann Transl Med. 2020;8(6):363.
Shen Y, Zhang M, Da L, Huang W, Zhang C. Circular RNA circ_SETD2 represses breast cancer progression via modulating the miR-155-5p/SCUBE2 axis. Open Med (Wars). 2020;15(1):940–53.
Chen JH, Kuo KT, Bamodu OA, Lin YC, Yang RB, Yeh CT, et al. Upregulated SCUBE2 expression in breast cancer stem cells enhances triple negative breast cancer aggression through modulation of notch signaling and epithelial-to-mesenchymal transition. Exp Cell Res. 2018.
Yang B, Miao S, Li Y. SCUBE2 inhibits the proliferation, migration and invasion of human non-small cell lung cancer cells through regulation of the sonic hedgehog signaling pathway. Gene. 2018;672:143–9.
Guo E, Liu H, Liu X. Overexpression of SCUBE2 inhibits proliferation, migration, and invasion in glioma cells. Oncol Res. 2017;25(3):437–44.
Song Q, Li C, Feng X, Yu A, Tang H, Peng Z, et al. Decreased expression of SCUBE2 is associated with progression and prognosis in colorectal cancer. Oncol Rep. 2015;33(4):1956–64.
Imam N, Alam A, Siddiqui MF, Ahmed MM, Malik MZ, Ikbal Khan MJ, et al. Identification of key regulators in parathyroid adenoma using an integrative gene network analysis. Bioinformation. 2020;16(11):910–22.
Wang X, Zhong RY, Xiang XJ. Reduced expression of SCUBE2 predicts poor prognosis in gastric cancer patients. Int J Clin Exp Pathol. 2018;11(2):972–80.
Ottley EC, Pell R, Brazier B, Hollidge J, Kartsonaki C, Browning L, et al. Greater utility of molecular subtype rather than epithelial-to-mesenchymal transition (EMT) markers for prognosis in high-risk non-muscle-invasive (HGT1) bladder cancer. J Pathol Clin Res. 2020;6(4):238–51.
Skrzypczak M, Lattrich C, Haring J, Schuler S, Ortmann O, Treeck O. Expression of SCUBE2 gene declines in high grade endometrial cancer and associates with expression of steroid hormone receptors and tumor suppressor PTEN. Gynecol Endocrinol. 2013;29(12):1031–5.
Chen Y, Zhou C, Li H, Li H, Li Y. Identifying key genes for nasopharyngeal carcinoma by prioritized consensus differentially expressed genes caused by aberrant methylation. J Cancer. 2021;12(3):874–84.
Rinne SJ, Sipila LJ, Sulo P, Jouanguy E, Beziat V, Abel L, et al. Candidate predisposition variants in Kaposi sarcoma as detected by whole-genome sequencing. Open Forum Infect Dis. 2019;6(10):ofz337.
Brilha S, Ong CWM, Weksler B, Romero N, Couraud PO, Friedland JS. Matrix metalloproteinase-9 activity and a downregulated Hedgehog pathway impair blood-brain barrier function in an in vitro model of CNS tuberculosis. Sci Rep. 2017;7(1):16031.
Wu MY, Gao F, Yang XM, Qin X, Chen GZ, Li D, et al. Matrix metalloproteinase-9 regulates the blood brain barrier via the hedgehog pathway in a rat model of traumatic brain injury. Brain Res. 2020;1727: 146553.
Ali H, Rustam R, Aprilia D, Arizal C, Gusadri IB, Utami PR. Upregulation of SCUBE2 expression in dyslipidemic type 2 diabetes mellitus is associated with endothelin-1. Diabetes Metab Syndr. 2019;13(5):2869–72.
Ali H, Emoto N, Yagi K, Vignon-Zellweger N, Nakayama K, Hatakeyama K, et al. Localization and characterization of a novel secreted protein, SCUBE2, in the development and progression of atherosclerosis. Kobe J Med Sci. 2013;59(4):E122–31.
Al-Saady NM, Obel OA, Camm AJ. Left atrial appendage: structure, function, and role in thromboembolism. Heart. 1999;82(5):547–54.
Ouzhuluobu, He Y, Lou H, Cui C, Deng L, Gao Y, et al. De novo assembly of a Tibetan genome and identification of novel structural variants associated with high-altitude adaptation. Natl Sci Rev. 2020;7(2):391–402.
Gunduz I, Batmaz I, Bozan T, Ekinci A, Cevik R. The relationship of serum SCUBE-1, -2 and -3 levels with clinical findings and ultrasonographic skin thickness in systemic sclerosis patients. Int J Rheum Dis. 2020;23(4):526–31.
Wang J, Kong X, Hu H, Shi S. Knockdown of long non-coding RNA PVT1 induces apoptosis of fibroblast-like synoviocytes through modulating miR-543-dependent SCUBE2 in rheumatoid arthritis. J Orthop Surg Res. 2020;15(1):142.
Sheng J, Shi W, Guo H, Long W, Wang Y, Qi J, et al. The Inhibitory Effect of (-)-Epigallocatechin-3-Gallate on Breast Cancer Progression via Reducing SCUBE2 Methylation and DNMT Activity. Molecules. 2019;24(16).
Haworth K, Smith F, Zoupa M, Seppala M, Sharpe PT, Cobourne MT. Expression of the Scube3 epidermal growth factor-related gene during early embryonic development in the mouse. Gene Expr Patterns. 2007;7(5):630–4.
Xavier GM, Economou A, Senna Guimaraes AL, Sharpe PT, Cobourne MT. Characterization of a mouse Scube3 reporter line. Genesis. 2010;48(12):684–92.
Rios JJ, Denton K, Yu H, Manickam K, Garner S, Russell J, et al. Saturation mutagenesis defines novel mouse models of severe spine deformity. Dis Model Mech. 2021;14(6).
Fuchs H, Sabrautzki S, Przemeck GK, Leuchtenberger S, Lorenz-Depiereux B, Becker L, et al. The first Scube3 mutant mouse line with pleiotropic phenotypic alterations. G3 (Bethesda). 2016;6(12):4035–46.
Liu Y, Guerrero-Juarez CF, Xiao F, Shettigar NU, Ramos R, Kuan CH, et al. Hedgehog signaling reprograms hair follicle niche fibroblasts to a hyper-activated state. Dev Cell. 2022;57(14):1758-75.e7.
Gudbjartsson DF, Walters GB, Thorleifsson G, Stefansson H, Halldorsson BV, Zusmanovich P, et al. Many sequence variants affecting diversity of adult human height. Nat Genet. 2008;40(5):609–15.
Zhao J, Li M, Bradfield JP, Zhang H, Mentch FD, Wang K, et al. The role of height-associated loci identified in genome wide association studies in the determination of pediatric stature. BMC Med Genet. 2010;11:96.
Wang L, Zhang L, Yan H, Liu X, Li N, Liang J, et al. Genome-wide association studies identify the loci for 5 exterior traits in a Large White × Minzhu pig population. PLoS ONE. 2014;9(8): e103766.
Yang HY, Cheng CF, Djoko B, Lian WS, Tu CF, Tsai MT, et al. Transgenic overexpression of the secreted, extracellular EGF-CUB domain-containing protein SCUBE3 induces cardiac hypertrophy in mice. Cardiovasc Res. 2007;75:139–47.
Dacic S, Kalajzic I, Visnjic D, Lichtler AC, Rowe DW. Col1a1-driven transgenic markers of osteoblast lineage progression. J Bone Miner Res. 2001;16(7):1228–36.
Kalajzic Z, Liu P, Kalajzic I, Du Z, Braut A, Mina M, et al. Directing the expression of a green fluorescent protein transgene in differentiated osteoblasts: comparison between rat type I collagen and rat osteocalcin promoters. Bone. 2002;31(6):654–60.
Chou CH, Cheng YF, Siow TY, Kumar A, Peck K and Chang C. SCUBE3 regulation of early lung cancer angiogenesis and metastatic progression. Clin Exp Metastasis. 2013.
Zhao C, qin Q, Wang Q, Zhang J, Xu Y, Li W, et al. SCUBE3 overexpression predicts poor prognosis in non-small cell lung cancer. Biosci Trends. 2013;7(6):264–9.
Hu F, Fong KO, Cheung MPL, Liu JA, Liang R, Li TW, et al. DEPDC1B promotes melanoma angiogenesis and metastasis through sequestration of ubiquitin ligase CDC16 to stabilize secreted SCUBE3. Adv Sci (Weinh). 2022;9(10): e2105226.
Yang X, Hu J, Shi C, Dai J. Activation of TGF-β1 pathway by SCUBE3 regulates TWIST1 expression and promotes breast cancer progression. Cancer Biother Radiopharm. 2020;35(2):120–8.
Huo Q, He X, Li Z, Yang F, He S, Shao L, et al. SCUBE3 serves as an independent poor prognostic factor in breast cancer. Cancer Cell Int. 2021;21(1):268.
Xu P, Luo A, Xiong C, Ren H, Yan L, Luo Q. SCUBE3 downregulation modulates hepatocellular carcinoma by inhibiting CCNE1 via TGFβ/PI3K/AKT/GSK3β pathway. Cancer Cell Int. 2022;22(1):1.
Qiu PC, Lu YY, Zhang S, Li H, Bao H, Ji YQ, et al. Reduction of SCUBE3 by a new marine-derived asterosaponin leads to arrest of glioma cells in G1/S. Oncogenesis. 2020;9(8):71.
Heng L, Jia Z, Sun J, Zhao Y, Zhang K, Zhu Y, et al. Integrated analysis of competing endogenous RNAs network reveals potential signatures in osteosarcoma development. Technol Cancer Res Treat. 2020;19:1533033820957025.
Liang W, Yang C, Peng J, Qian Y, Wang Z. The expression of HSPD1, SCUBE3, CXCL14 and its relations with the prognosis in osteosarcoma. Cell Biochem Biophys. 2015;73(3):763–8.
Song G, Wang J, Lu J, Xu H, Zhao Z, Tang Q, et al. Role of SCUBE3 in promoting osteosarcoma cell growth and its association with prognosis. Nan Fang Yi Ke Da Xue Xue Bao. 2014;34(5):617–21.
Morris MR, Ricketts CJ, Gentle D, McRonald F, Carli N, Khalili H, et al. Genome-wide methylation analysis identifies epigenetically inactivated candidate tumour suppressor genes in renal cell carcinoma. Oncogene. 2011;30(12):1390–401.
Collod-Béroud G, Béroud C, Ades L, Black C, Boxer M, Brock DJ, et al. Marfan Database (third edition): new mutations and new routines for the software. Nucleic Acids Res. 1998;26(1):229–33.
Belleh S, Zhou G, Wang M, Der Kaloustian VM, Pagon RA, Godfrey M. Two novel fibrillin-2 mutations in congenital contractural arachnodactyly. Am J Med Genet. 2000;92(1):7–12.
Intarak N, Theerapanon T, Thaweesapphithak S, Suphapeetiporn K, Porntaveetus T, Shotelersuk V. Genotype-phenotype correlation and expansion of orodental anomalies in LTBP3-related disorders. Mol Genet Genomics. 2019;294(3):773–87.
Varadi M, Anyango S, Deshpande M, Nair S, Natassia C, Yordanova G, et al. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022;50(D1):D439–44.
Liu W, Qian C, Comeau K, Brenn T, Furthmayr H, Francke U. Mutant fibrillin-1 monomers lacking EGF-like domains disrupt microfibril assembly and cause severe marfan syndrome. Hum Mol Genet. 1996;5(10):1581–7.
Xu P, Li R, Huang S, Sun M, Liu J, Niu Y, et al. A novel splicing mutation in the FBN2 gene in a family with congenital contractural arachnodactyly. Front Genet. 2020;11:143.
Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596(7873):583–9.
Banerjee A, Bhakta S, Sengupta J. Integrative approaches in cryogenic electron microscopy: Recent advances in structural biology and future perspectives. iScience. 2021;24(2):102044.
Collins CT, Hess JL. Role of HOXA9 in leukemia: dysregulation, cofactors and essential targets. Oncogene. 2016;35(9):1090–8.
Huang P, Wierbowski BM, Lian T, Chan C, García-Linares S, Jiang J, et al. Structural basis for catalyzed assembly of the Sonic hedgehog-Patched1 signaling complex. Dev Cell. 2022;57(5):670-85.e8.
Akbar Samadani A, Keymoradzdeh A, Shams S, Soleymanpour A, Elham Norollahi S, Vahidi S, et al. Mechanisms of cancer stem cell therapy. Clin Chim Acta. 2020;510:581–92.
Jeng KS, Chang CF and Lin SS. Sonic Hedgehog Signaling in Organogenesis, Tumors, and Tumor Microenvironments. Int J Mol Sci. 2020;21(3).
Lézot F, Corre I, Morice S, Rédini F and Verrecchia F. SHH Signaling Pathway Drives Pediatric Bone Sarcoma Progression. Cells. 2020;9(3).
Wynn DT, Rodriguez-Blanco J, Long J, Yang F, Shen C, Fei D, et al. Protein arginine methyltransferase 5 regulates SHH-subgroup medulloblastoma progression. Neurooncol Adv. 2022;4(1):vdac144.
Topcu TO, Kavgaci H, Ozdemir F, Aksoy A, Erdem D, Mentese A, et al. Elevated serum levels of SCUBE1, a marker for coagulation, in patients with breast cancer. Tohoku J Exp Med. 2015;237(2):127–32.
Mentese A, Fidan E, Sumer AU, Karahan SC, Sonmez M, Altay DU, et al. Is SCUBE 1 a new biomarker for gastric cancer? Cancer Biomark. 2012;11(5):191–5.
Orr B, Grace OC, Brown P, Riddick AC, Stewart GD, Franco OE, et al. Reduction of pro-tumorigenic activity of human prostate cancer-associated fibroblasts using Dlk1 or SCUBE1. Dis Model Mech. 2013;6(2):530–6.
Levina E, Chen M, Carkner R, Shtutman M, Buttyan R. Paracrine Hedgehog increases the steroidogenic potential of prostate stromal cells in a Gli-dependent manner. Prostate. 2012;72(8):817–24.
Jiang W, Han X, Wang J, Wang L, Xu Z, Wei Q, et al. miR-22 enhances the radiosensitivity of small-cell lung cancer by targeting the WRNIP1. J Cell Biochem. 2019;120(10):17650–61.
Karaguzel E, Mentese A, Kazaz IO, Demir S, Orem A, Okatan AE, et al. SCUBE1: a promising biomarker in renal cell cancer. Int Braz J Urol. 2017;43(4):638–43.
Iwamoto T, Katagiri T, Niikura N, Miyoshi Y, Kochi M, Nogami T, et al. Immunohistochemical Ki67 after short-term hormone therapy identifies low-risk breast cancers as reliably as genomic markers. Oncotarget. 2017;8(16):26122.
Parris TZ, Danielsson A, Nemes S, Kovacs A, Delle U, Fallenius G, et al. Clinical implications of gene dosage and gene expression patterns in diploid breast carcinoma. Clin Cancer Res. 2010;16(15):3860–74.
Behring M, Ye Y, Elkholy A, Bajpai P, Agarwal S, Kim HG, et al. Immunophenotype-associated gene signature in ductal breast tumors varies by receptor subtype, but the expression of individual signature genes remains consistent. Cancer Med. 2021;10(16):5712–20.
Andres SA, Brock GN, Wittliff JL. Interrogating differences in expression of targeted gene sets to predict breast cancer outcome. BMC Cancer. 2013;13(1):1–18.
Koleck TA, Conley YP. Identification and prioritization of candidate genes for symptom variability in breast cancer survivors based on disease characteristics at the cellular level. Breast Cancer (Dove Med Press). 2016;8:29–37.
Parris TZ, Kovacs A, Aziz L, Hajizadeh S, Nemes S, Semaan M, et al. Additive effect of the AZGP1, PIP, S100A8 and UBE2C molecular biomarkers improves outcome prediction in breast carcinoma. Int J Cancer. 2014;134(7):1617–29.
Kallarackal J, Burger F, Bianco S, Romualdi A, Schad M. A 3-gene biomarker signature to predict response to taxane-based neoadjuvant chemotherapy in breast cancer. PLoS ONE. 2020;15(3): e0230313.
Koleck TA, Bender CM, Clark BZ, Ryan CM, Ghotkar P, Brufsky A, et al. An exploratory study of host polymorphisms in genes that clinically characterize breast cancer tumors and pretreatment cognitive performance in breast cancer survivors. Breast Cancer (Dove Med Press). 2017;9:95–110.
Chen JH, Kuo KT, Bamodu OA, Lin YC, Yang RB, Yeh CT, et al. Upregulated SCUBE2 expression in breast cancer stem cells enhances triple negative breast cancer aggression through modulation of notch signaling and epithelial-to-mesenchymal transition. Exp Cell Res. 2018;370(2):444–53.
Parris TZ, Aziz L, Kovács A, Hajizadeh S, Nemes S, Semaan M, et al. Clinical relevance of breast cancer-related genes as potential biomarkers for oral squamous cell carcinoma. BMC Cancer. 2014;14(1):1–11.
Yang X, Hu J, Shi C, Dai J. Activation of TGF-beta1 pathway by SCUBE3 regulates TWIST1 expression and promotes breast cancer progression. Cancer Biother Radiopharm. 2020;35(2):120–8.
Gao C, Zhuang J, Li H, Liu C, Zhou C, Liu L, et al. Development of a risk scoring system for evaluating the prognosis of patients with Her2-positive breast cancer. Cancer Cell Int. 2020;20:121.
Joosten SC, Odeh SNO, Koch A, Buekers N, Aarts MJB, Baldewijns M, et al. Development of a prognostic risk model for clear cell renal cell carcinoma by systematic evaluation of DNA methylation markers. Clin Epigenetics. 2021;13(1):103.
Xu P, Luo A, Xiong C, Ren H, Yan L, Luo Q. SCUBE3 downregulation modulates hepatocellular carcinoma by inhibiting CCNE1 via TGFbeta/PI3K/AKT/GSK3beta pathway. Cancer Cell Int. 2022;22(1):1.
Chou CH, Cheng YF, Siow TY, Kumar A, Peck K, Chang C. SCUBE3 regulation of early lung cancer angiogenesis and metastatic progression. Clin Exp Metastasis. 2013;30(6):741–52.
Song G, Wang J, Lu J, Xu H, Zhao Z, Tang Q, et al. Role of SCUBE3 in promoting osteosarcoma cell growth and its association with prognosis. Nan Fang yi ke da xue xue bao= Journal of Southern Medical University. 2014;34(5):617–21.
Han N, Lu H, Zhang Z, Ruan M, Yang W, Zhang C. Comprehensive and in-depth analysis of microRNA and mRNA expression profile in salivary adenoid cystic carcinoma. Gene. 2018;678:349–60.
Weisshuhn PC, Sheppard D, Taylor P, Whiteman P, Lea SM, Handford PA, et al. Non-linear and flexible regions of the human Notch1 extracellular domain revealed by high-resolution structural studies. Structure. 2016;24(4):555–66.
Stenflo J, Stenberg Y, Muranyi A. Calcium-binding EGF-like modules in coagulation proteinases: function of the calcium ion in module interactions. Biochim Biophys Acta. 2000;1477(1–2):51–63.
Stenflo J, Ohlin AK, Owen WG, Schneider WJ. beta-hydroxyaspartic acid or beta-hydroxyasparagine in bovine low density lipoprotein receptor and in bovine thrombomodulin. J Biol Chem. 1988;263(1):21–4.
Romão MJ, Kölln I, Dias JM, Carvalho AL, Romero A, Varela PF, et al. Crystal structure of acidic seminal fluid protein (aSFP) at 1.9 A resolution: a bovine polypeptide of the spermadhesin family. J Mol Biol. 1997;274(4):650–60.
Acknowledgements
The authors thank Dr. Ming-Jing Hwang and Ching-Shu Suen for their helpful discussion and assistance on homology modeling and structural insights into different protein domains of SCUBE family.
Funding
This work was supported by grants from Academia Sinica (AS- GC- 111- L04 to R.-B.Y.), the Biomedical Translation Research Center of Academia Sinica (AS- KPQ- 111- KNT to R.-B.Y.) and the National Science and Technology Council of Taiwan (109- 3111- Y- 001- 001, 109- 2320- B- 001- 012- MY3 and 110- 2320- B- 001- 019- MY3 to R.-B. Y. and 107- 2321- B- 001- 036- MY3, 110- 2326- B-001- 004- MY3 and 110- 2326- B- 038- 003- MY3 to Y.-C.L.).
Author information
Authors and Affiliations
Contributions
Y-CL, BKS, S-SG, and R-BY conceived the review idea, prepared figures, and wrote this review manuscript together. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lin, YC., Sahoo, B.K., Gau, SS. et al. The biology of SCUBE. J Biomed Sci 30, 33 (2023). https://doi.org/10.1186/s12929-023-00925-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12929-023-00925-3