
Abstract
Background
Liquid suspension culture efficiently proliferates plant cells and can be applied to ferns because it rapidly increases the fresh weight of gametophytes. This study assessed gametophyte proliferation and sporophyte production of Pteridium aquilinum var. latiusculum using a suspension culture method.
Results
The growth curve linear phase of gametophyte cells was confirmed between 9 and 18 days of culture, and the subculture cycle was determined to be 2 weeks. A double-strength MS medium (fresh weight, 18.0 g) containing 2% sucrose and NH4+:NO3− (120 mM, 40:80) was found to be the optimal liquid medium. Gametophytes obtained after suspension culture for 18 days did not normally form sporophytes in an ex vitro soil environment. However, this issue was resolved after changing the culture type or extending the culture period to 6 weeks. A short suspension culture period increased the fresh weight of fragmented and homogenized gametophytes but yielded numerous relatively immature gametophytes (globular forms of branching gametophytes, BG). Furthermore, differences in gametophyte morphogenesis and development were indicated by changes in endogenous phytohormone content. BG with immature development exhibited high accumulation of zeatin, jasmonic acid, and salicylic acid, and relatively low levels of abscisic acid and indole-3-acetic acid. The immature development of gametophytes directly affected sporophyte formation.
Conclusions
This study maximized the advantages of liquid suspension culture using eastern bracken gametophytes and provides data to resolve any associated issues, thus facilitating efficient bracken production.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Background
Ferns are very popular horticultural plants worldwide [1]. They have a very high value as indoor/outdoor ornamental and landscaping plant materials. Ferns are rich sources of protein, essential amino acids, fatty acids, fiber, vitamins, and minerals [2], and they also occupy an important position as an edible plant [3, 4]. Among the ferns, eastern bracken (Pteridium aquilinum var. latiusculum (Desv.) Underw. ex A. Heller) is a popular wild vegetable in South Korea and its allies. Production of eastern bracken in South Korea, in particular, has reached 14,031 tons (72 million USD) [5], and it is cultivated as a high value commercial crop.
A liquid suspension culture of plant cells consists of cells and cell clumps dispersed and grown in an agitated medium [6]. The number of cells in the medium increases rapidly and approaches the maximum cell density [7]. Simultaneously, as mineral salts and carbohydrates in the medium are rapidly consumed as an energy source, the selection of medium components is important. In addition, liquid culture can resolve numerous issues associated with solid cultures, such as gaseous exchange and the gradients of nutrients in the medium. Suspension culture facilitates large-scale production of various plant species and is suitable for strategically producing high amounts of useful components (e.g., functional substances, natural products, and secondary metabolites) [8]. Furthermore, suspension culture can effectively preserve and propagate species that are difficult to cultivate and propagate, such as endangered wild plants. In suspension culture, callus and green globular bodies (ferns) derived from cells of numerous plant species have been used [8]. For example, Pteris vittata [9], Nephrolepis exaltata [10], Drynaria quercifolia [11], and Platycerium bifurcatum [12] were produced in suspension culture. However, ferns are easy to produce using gametophyte instead of callus [13, 14]. The gametophyte has a high regenerative ability, and this method can be maximized in liquid suspension cultures. However, the characteristics and development of gametophytes obtained by culture may potentially differ from those of gametophytes grown naturally depending on the culture type, such as solid, liquid, and airlift fermenter [15, 16]. Furthermore, exogenous gibberellic acid4+7 affects the regulation of gametophyte morphogenesis [17], and 6-Benzylaminopurine induces male development of homogenized gametophytes [18]. As such, gametophytes are sensitive to the culture type and phytohormone, and morphogenesis and endogenous phytohormone content potentially differ in accordance with gametophyte development and maturity [19,20,21].
This study proposed the mass production of eastern bracken gametophytes and sporophytes of high commercial value using liquid suspension culture. Furthermore, gametophyte morphogenesis and endogenous phytohormone content were investigated in accordance with culture type and culture period.
Results
Gametophyte growth in suspension culture
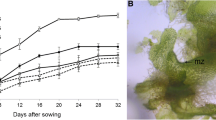
Based on the growth curve for 30 days (Fig. 1A), the gametophytes increased rapidly from the 6th day (Fig. 1B). The fresh weight of gametophytes increased rapidly between 9 and 18 days and were 4.4, 7.0, 9.0, and 14.7 g. From the 18th day, browning and senescence were observed in some gametophyte cells (Fig. 1C).

Suspension culture growth curve and fresh weight of eastern bracken gametophyte measured over 30 days. A Growth curve; B, C gametophytes in accordance with the liquid culture period (left to right: B 3, 6, 9, 12, 15, C 18, 21, 24, 27, and 30 days after inoculation). Vertical bars represent the mean ± standard error values (n = 4). zDifferent letters indicate a significant difference using Duncan’s multiple range test at P < 0.05
Conditions of gametophyte proliferation in suspension culture
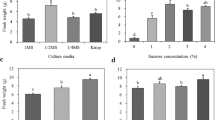
The fresh weight of gametophytes (15.9 and 18.0 g) increased the most in 1 and 2MS medium (Fig. 2A). Fresh weight was most investigated in 2MS medium containing 2–3% sucrose (Fig. 2B). Regardless of the total concentration and ratio, the fresh weight of the gametophytes peaked in the medium with the nitrogen source adjusted to 120 mM NH4+:NO3− (40:80) (Fig. 2C, D). Activated charcoal was not effective in increasing the fresh weight of gametophytes (Fig. 2E).

Effect of medium components on the fresh weight of eastern bracken gametophytes cultured for 18 days. A Culture media; B sucrose concentration; C total nitrogen concentration; D NH4+:NO3− nitrogen ratio; E charcoal concentration. Vertical bars represent the mean ± standard error values (n = 4). zDifferent letters indicate a significant difference using Duncan’s multiple range test at P < 0.05
Ex vitro sporophyte propagation in accordance with culture types
GS produced 156.7 sporophytes per pot, whereas GL, GL1st, and GL2nd hardly formed sporophytes per pot) (Table 1). However, GL2nd-GS produced 77.8 sporophytes per pot. Two weeks after sowing, all fragmented and homogenized gametophytes were successfully covered in ex vitro soil. Sporophyte formation occurred 4 weeks after sowing (Fig. 3). Nevertheless, unlike solid culture gametophytes, GL did not develop into sporophytes.

Development of eastern bracken gametophyte and sporophyte sown in accordance with the culture types in ex vitro conditions. GS, gametophyte fragmentation cultured for 6 weeks in solid medium; GL, gametophyte fragmentation cultured for 2 weeks in liquid medium; GL2nd-GS, liquid cultured gametophytes were subcultured twice at intervals of 2 weeks and then solid cultured for 6 weeks. Scale bars are 3 mm
Gametophyte morphogenesis in accordance with culture types
The morphogenesis of gametophytes obtained with different culture types was investigated (Table 2). GS mostly exhibited spatula gametophyte (SG) (89.7%) and spatula-heart gametophyte (S-HG) (10.3%). GL had a high proportion of branching gametophyte (BG) with immature morphogenesis (Fig. 4). For GL, GL1st, and GL2nd, the proportion of BG was high, from 54.5 to 82.4%. GL2nd-GS displayed a similar tendency to GS, with recovery of morphogenesis of SG (77.2%) and S-HG (22.8%). The GL series inhibited sporophyte formation regardless of morphogenesis (Additional file 1: Table S1).

Regeneration and morphology of eastern bracken gametophytes cultured in liquid medium. A Globular forms of branching gametophyte; B and C branching gametophyte; D spatula shaped; E and F spatula-heart shaped. Scale bars are 1 mm
Gametophyte endogenous phytohormone content according to culture types
Low levels of both ABA and IAA were detected in GL (1.0 and 16.9 ng· g−1) (Table 3). In contrast, high levels of zeatin, JA, and SA were detected in GL (109.4, 16.4, and 241.7 ng· g−1). However, GA3 content was not detected.
Gametophyte morphogenesis and ex vitro sporophyte propagation according to the culture period and density
As the culture period increased, the BG decreased, and the SG and S-HG percentages increased (Table 4). Furthermore, as the density of gametophytes per flask decreased, BG decreased, and S-HG increased. The most drastic change was investigated at a density of 0.1 g: 2 weeks (BG, 82.3%; SG, 10.3%; S-HG, 7.4%), 4 weeks (BG, 46.4%; SG, 33.4%; S-HG, 20.2%), and 6 weeks (BG, 7.1%; SG, 0.7%; S-HG, 92.2%).
As the suspension culture period was extended to 6 weeks, sporophyte formation was recovered (Table 5). Sporophytes cultured for 6 weeks formed 56.0–115.0 sporophytes per pot, yielding different results depending on the density. Sporophytes were formed the most in gametophytes cultured for 6 weeks at a density of 0.1 g, and the growth of the individuals was relatively good.
Discussion
A suitable subculture period is essential for long-term preservation or proliferation of plant cells [1, 22]. Prompt depletion of nutrients in the suspension medium occurs with rapid growth of the cells [23]. Cell growth in liquid suspension culture is characterized by a lag phase, exponential phase, linear phase, deceleration phase, and a stationary phase [7]. Furthermore, eastern bracken gametophyte cells undergo the same growth process. The growth curve of gametophyte cells revealed that the gametophytes entered the exponential phase from the 6th day of culture. The culture period between 9 and 18 days revealed linear growth, and the deceleration phase was estimated to occur after 18 days. The fresh weight of the gametophytes stagnated from the 21st day and entered the stationary phase. Therefore, it was determined that the subculture period occurred before the deceleration phase (2 weeks post-culture).
A medium is the most important factor for plant growth in in vitro culture and contains various components for explant growth. It contains nutrients such as mineral salts, organic matter, and carbohydrates, and often contains activated charcoal, agar, and plant growth regulators. Among these, mineral salts and carbohydrates are indispensable for explant growth and proliferation [22, 23]. In this study, the difference in fresh weight of eastern bracken gametophytes, in accordance with the medium and components, was clearly revealed. Quarter- and half-strength MS liquid medium resulted in severe nutrient depletion, and quadruple-strength MS decreased the fresh weight owing to an excessive nutrient supply. The fresh weight of gametophytes depended on sucrose and nitrogen sources. However, activated charcoal did not contribute to this process. Through successive experiments, the liquid medium suitable for eastern bracken gametophyte proliferation was found to be a double-strength MS medium containing 2% sucrose and NH4+:NO3−; 120 mM (40:80). The eastern bracken gametophytes required very rich nutrients and components. Moreover, this has been reported in studies on Equisetum arvense [24], Dicksonia sellowiana [25], Pteris tripartita [26], and Lemmaphyllum microphyllum [27].
Gametophytes produced through liquid culture for 18 days displayed a very high proliferation rate; nonetheless, unfortunately, sporophytes were not normally formed in ex vitro soil environments. This issue resulted from differences in culture type and culture period, and gametophyte morphogenesis and developmental issues were also considered. In general, gametophytes form filamentous-, spatula-, spatula-heart-, heart-, and ribbon-shaped gametophytes during development [28]. The mature gametophyte is close to heart-shaped, and early gametophyte cells, which are immature and actively dividing, were filamentous-shaped. This morphogenesis can be classified in accordance with the division and development of gametophyte cells. Sheffield et al. [16] reported that the characteristics of cultured gametophytes may differ from those of gametophytes grown under natural conditions depending on the culture method. Gametophytes obtained from the suspension culture (GL series) had more than 55% of globular forms of BG, which is similar to that of filamentous gametophytes. The percentage of BG rapidly increased with repeated subculturing, and sporophytes were hardly formed. Meanwhile, in GS used as an initial explant, no BG was obtained after culturing, and only S-HG and SG were identified. GL2nd-GS displayed similar morphology to GS. In both conditions, sporophytes were normally formed, suggesting that gametophyte morphogenesis and development are closely associated with sporophyte formation.
Gametophyte morphogenesis is closely associated with cell division and development and is characterized by differential endogenous phytohormone content. The development of gametophytes and sporophytes is regulated by various hormone systems [29]. The ABA content of GL was less than half of that of the control and GS, in which gametophytes were relatively immature (GL). As GL contains a large amount of BG, cell division is quite active. ABA accumulated more in the mature stage than in the early stage, where cell division was active. Exogenous ABA inhibited cell division and elongation of Lygodium japonicum gametophyte [30] and that of Mohria caffrorum protonema [31]. Notably, the endogenous IAA content displayed a similar trend to that of ABA. Kosakivska et al. [20] reported a similar balance between the endogenous ABA and IAA contents analyzed from Polystichum aculeatum gametophytes of the same morphogenesis stage. Although auxin is involved in the growth of gametophytes, it accumulates primarily in young sporophytes and promotes development [32]. The IAA content was the lowest in the GL with BG; however, the control and GS with S-HG and SG displayed relatively high IAA levels, suggesting that similar to ABA, auxins accumulate more in mature gametophytes than in early-stage gametophytes. Gibberellin or gibberellin-like hormones are involved in the formation of reproductive organs and sex determination in ferns [33, 34] and are produced during gametophyte development; nonetheless, only GA3 was analyzed among various gibberellins, and all types of gibberellins were not synthesized herein. In the Asplenium nidus gametophyte, only GA9 was detected in the analysis of GA1, GA3, GA4, GA7, GA9, and GA20 [19]. Endogenous gibberellin is expected to play an important role in gametophyte development and morphogenesis. Therefore, it is necessary to analyze gibberellins in the biosynthetic pathway in the future. Cytokinins (CKs) are involved in cell division and organogenesis, among which zeatin promotes gametophyte and rhizoid development [35]. CKs can be detected at all stages because they promote cell division and organogenesis for development into mature plants. Zeatin, dihydrozeatin, zeatin-riboside, and isopentenyladenine were detected at similar levels in both gametophytes and sporophytes [19]. Kosakivska et al. [20] reported zeatin glucoside and isopentenyladenin in both the SG and sporophyte stages. In this study, zeatin was detected in the control, GS, and GL. Furthermore, high levels of both JA and SA were detected in GL. Surprisingly, endogenous JA and SA contents opposite trends to ABA and IAA contents. Although JA and SA are not often-used hormones in ferns, they have been used in some studies. Camloh et al. [36] reported that JA promotes early gametophyte development. Grzyb et al. [37] reported that exogenous SA reduced the endogenous IAA content of Cyathea delgadii explants. These results suggest that endogenous JA and SA hormones may contribute to early cell division in gametophytes, and these hormones potentially interact with other phytohormones. Moreover, the concentration of endogenous phytohormones clearly plays an important role in hormone regulation. Phytohormone concentration and function exert different effects depending on the cell and hormone receptor site. Additionally, qualitative and quantitative differences in phytohormones occur between adjacent tissues of the same species. For example, differences in gibberellin content and metabolism have been detected when comparing Pisum sativum L. embryos and seed coats [38]. The differing response to phytohormones is referred to as tissue 'sensitivity' (or responsiveness) [39, 40]. It is difficult to accurately define or measure sensitivity. For example, an increase in the number of phytohormone receptors, decrease in phytohormone breakdown, reduction in metabolism leading to fewer conjugated or inactive forms, and a decrease in transport away from the active site could all be interpreted as an increase in tissue sensitivity to an exogenous phytohormone [40]. Therefore, further studies and samples are required in this regard. Unfortunately, the sensitivity of gametophytic cells was not considered in this study. This should be considered when analyzing endogenous phytohormone concentration in the future.
As described above, morphogenesis (cell division and development) and the endogenous phytohormone content of gametophytes differed according to the culture type. Consequently, immature gametophytes influenced the inhibition of sporophyte formation. Short-term liquid culture effectively increased the fresh weight; however, it did not promote the regeneration and development of fragmented gametophytes. As the liquid culture period was extended to 6 weeks, sporophyte formation resumed. Furthermore, morphogenesis was confirmed to be closely associated with gametophyte density. The same volume of medium had a different nutrient-use efficiency in accordance with the gametophyte density [6, 7], potentially leading to developmental or morphogenetic changes. In fact, at 0.1 g density, the ratio of immature BG decreased significantly, and sporophyte formation increased. Furthermore, gametophyte morphogenesis and the percentage of sporophyte formation displayed the same tendency and recovered upon 6 weeks of culture. The culture period and gametophyte density were associated with gametophyte morphogenesis and the number of sporophytes formed and displayed a highly significant correlation (P < 0.001) (Tables 4 and 5).
Conclusions
In summary, this study provides data on how to maximize the advantages of liquid suspension culture and resolve the associated issues, thus facilitating the efficient production of eastern bracken seedlings. Liquid suspension culture has been shown to rapidly increase the fresh weight of gametophytes. However, gametophytes obtained through suspension culture produced relatively immature gametophytes, which adversely affected the formation of sporophytes. In our study, the immature development of gametophytes directly affected sporophyte formation. This issue was resolved as the gametophytes matured after changing the culture type or extending the culture period to 6 weeks. Furthermore, BG with immature development exhibited high accumulation of zeatin, JA, and SA, and relatively low levels of ABA and IAA. It has been found that the content of endogenous hormones varies according to the maturity of gametophytes. This suggests that plant hormones are deeply involved in gametophyte development.
Methods
Plant materials
Sporophylls of Pteridium aquilinum var. latiusculum (Desv.) Underw. ex A. Heller (eastern bracken) were collected in a greenhouse at Chungbuk National University, Cheongju, Korea in September 2017. The sporophylls were dried in a paper box at 25 °C for 1 week, and spores and impurities were filtered out using a 100 μm sieve (Chunggye Sieve, Gunpo, Korea).
Spores were surface-sterilized and germinated in accordance with the methods of Jang et al. [41]. The spore solution (30 mg· 15 mL−1) was centrifuged (3 min, 1811×g), and the supernatant was discarded. The spores were sterilized with 1.4% (v/v) sodium hypochlorite (4% NaClO; Yuhanclorox Co., Ltd., Hwaseong, Korea) for 13 min and washed thrice with sterilized water. Finally, the spore solution was diluted to 1 mg· mL−1 of sterilized water and then inoculated in Knop medium [42]; the spores germinated at 25 °C under a 16/8 h photoperiod, with a light intensity of 30 μmol· m−2 ·s−1 photosynthetic photon flux density (PPFD). Gametophytes obtained from the spores were subcultured using double-strength MS [43] medium at eight-week intervals and then used for further experiments. The gametophyte material was transferred to the same medium without homogenization during subculture.
Liquid suspension culture and gametophyte growth curve
The growth curves of eastern bracken gametophytes were assessed under suspension culture conditions. The culture was supplemented with 100 mL double-strength MS liquid medium [3% (w/v) sucrose, pH 5.8] in a 250 mL Erlenmeyer flask and agitated (VS-203D, Vision Scientific Co., Ltd, Daejeon, Korea) at 125 rpm. Gametophytes (1 g) were chopped using a scalpel and then suspension-cultured (1 g· 100 mL−1) [41]. The gametophytes were incubated for 30 days, and the fresh weight was determined at 3-day intervals. Thereafter, all cultures were carried out using this method. All suspension cultures were maintained at 25 °C, under a light intensity of 30 μmol· m−2 ·s−1, and a 16/8 h photoperiod, unless stated otherwise.
To determine the optimal liquid media for gametophyte proliferation, gametophyte fragments were cultured on quarter-, half-, full-, double-, and quadruple-strength MS liquid medium. Thereafter, the selected optimal medium (Fig. 2A, double-strength MS liquid medium) was controlled with various sucrose concentrations (0–5%, w/v), total nitrogen sources (60, 120, and 240 mM), nitrogen ratios (0:120, 40:80, 60:60, 80:40, and 120:0 mM), and activated charcoal (0%–0.8%, w/v) to assess gametophyte proliferation. NH4Cl (CAS 12125–02-9, Samchun Chemicals, Pyeongtaek, Korea) and KNO3 (CAS 7757–79-1, Wako Pure Chemical Industries, Ltd., Osaka, Japan) were used as nitrogen sources. The fresh weight of gametophytes was measured after 18 days. The solid medium was prepared with double-strength MS containing 2% sucrose, NH4+:NO3−, 120 mM (40:80), and 0.8% (w/v) agar (pH 5.8). The solid culture was sowed so that the ratio of gametophyte to medium was 0.3 g· 30 mL−1 and then cultured for 6 weeks.
Responses of ex vitro sporophyte propagation in accordance with culture types
For ex vitro sporophyte propagation, the solid or liquid culture-derived gametophytes were fragmented and homogenized with a hand blender with 25 mL distilled water [27]. Gametophyte (1 g) was fragmented to various culture types and sowed under ex vitro soil conditions. The culture types were as follows (Additional file 1: Figure S1) fragmented gametophyte cultured for 6 weeks in solid medium (GS) and fragmented gametophyte cultured for 2 weeks in liquid medium (GL). Liquid cultured gametophytes were subcultured once or twice at 2-week intervals and then liquid cultured for 2 weeks (GL1st or GL2nd). Liquid cultured gametophytes were subcultured twice at 2-week intervals and then solid cultured for 6 weeks (GL2nd-GS). Soil was mixed using sterilized horticultural substrate (Hanareum no. 2; Shinsung Mineral Co., Ltd., Goesan, Korea) and perlite (Newpershine no. 2; GFC. Co., Ltd., Hongseong, Korea) and filled in pots (75 × 75 × 75 mm). The soil volume was mixed at a 2:1 (v/v) ratio of horticultural substrate and perlite. Pots were placed in a plastic box, covered with a glass plate, and grown for 8 weeks. Water was sprayed on the gametophyte surface every 2-day 2 weeks after sowing. After 8 weeks, the growth of gametophytes and sporophytes was assessed. Thereafter, all ex vitro sporophyte propagations were carried out using the aforementioned method. All ex vitro conditions were maintained at 25 °C, under a light intensity of 43 μmol· m−2 ·s−1, 16/8 h photoperiod, and 85 ± 5% humidity, unless stated otherwise.
Responses of gametophyte morphogenesis in accordance with culture types
Morphogenesis and fresh weight of gametophytes regenerated after culturing were assessed in accordance with the culture type. Regenerated gametophytes in each Erlenmeyer flask were washed with distilled water and enumerated, except for browning gametophytes. The total number of gametophytes was determined [A], and gametophyte morphogenesis (Fig. 4) was classified into three types [B]: globular forms of BG, SG, and S-HG. Morphogenesis (%) was determined as follows: [B – A] × 100%. Gametophytes, whose morphogenesis was classified in accordance with the culture type, were sowed ex vitro. However, morphogenesis types that were not regenerated were excluded from sowing.
Change in gametophyte endogenous phytohormone content in accordance with the culture types
The endogenous phytohormone content in the gametophytes regenerated in accordance with the culture type was analyzed. As a control group, spore-derived gametophyte was analyzed. Gametophyte extraction from each treatment was conducted as indicated below. After freeze-drying, the homogenized 50 mg sample was placed in a 2 mL tube, and 500 μL extraction solvent (2-propanol/H2O/concentrated HCl (2:1:0.002, v/v/v, %) was added. The sample was homogenized at 100 rpm and 4 °C for 30 min. After agitation, 1 mL dichloromethane was added to each sample, agitated for 30 min (100 rpm, 4 °C), and each sample was centrifuged for 5 min (13,000 × g, 4 °C) (Smart R17 Plus; Hanil Scientific Inc., Gimpo, Korea). When the layers separated, 900 μL of the supernatant was transferred into a new 2 mL tube. The supernatant was evaporated using vacuum concentrators, and the recovered sample was finally dissolved in 100 μL methanol for subsequent use. The reference materials used were 2-cis,4-trans-ABA, GA3, IAA, JA, SA, and zeatin. They were dissolved in 1 mL methanol and diluted to 1 mg· mL−1. HPLC was performed using an Agilent 1260 series system (Agilent Technologies, Palo Alto, CA, USA). Chromatographic separation was achieved using an Agilent Eclipse Plus C18 column (4.6 × 50 mm2, 3.5 μm). The HPLC mobile phase was 0.1% formic acid in water (A) and 0.1% formic acid in methanol (B). The gradient was initiated at 5% (B), increased to 95% (B) for 1 min, and maintained at 95% (B) for 4 min. It was rapidly changed to 5% (B) for 0.1 min and retained for 0.9 min. The flow rate was set at 500 μL· min−1. The column oven temperature was set to 30 °C. The injection volume was 10 μL. The mass spectrometer was an API-4000 (AB Sciex, Concord, ON, Canada) equipped with an electrospray ionization source. Analysis was conducted in the negative mode and MRM mode. BioAnalyst version 1.6.1 and analyst software version 1.6.1 programs were used for equipment operation and data analysis, respectively. During ionization, high-purity nitrogen gas was used as the spray and drying gas. The gas pressure was set to 60 psi. Ion spray voltage (−4.5 kV) and an ionization source temperature of 600 °C were used. Q1 and Q3 were analyzed using LC–MS/MS in the multiple reaction monitoring mode with unit resolution. Three replicates were analyzed for each standard solution and sample.
Responses of gametophyte morphogenesis and ex vitro sporophyte propagation in accordance with the culture period and density
Gametophyte morphogenesis and sporophyte formation were assessed in accordance with the liquid culture duration and gametophyte density. The culture duration (2, 4, and 6 weeks) and gametophyte density (fresh weight: 0.1, 0.3, 0.5, and 1.0 g per flask) were varied and evaluated after ex vitro seeding.
Data collection and statistical analysis
Gametophyte morphogenesis was observed, and gametophytes were enumerated using a microscope (SZ61; Olympus, Tokyo, Japan). Gametophyte images were captured using a CMOS camera (eXcope F630; Dixi Sci., Daejeon, Korea) and eXcope 3.7.12277 software. In vitro gametophyte proliferation was evaluated using the fresh weight of cultured gametophytes (n = 4). To investigate the effect of sporophyte production, growth parameters of sporophytes were measured, including the number of sporophytes per pot, number of leaves, leaf length, number of roots, root length, and fresh weight. Triplicate sets were used to determine the levels of ex vitro sporophyte production, and sporophyte growth was investigated using 5 plants per replicate (n = 15). However, when <15 sporophytes were formed, all sporophytes were assessed. SAS version 9.4 (SAS Institute Inc., Cary, NC, USA) was used to determine the mean ± standard error values for each treatment, and factorial analysis was performed using Duncan’s multiple range test, with a significance level of P < 0.05. The culture period and gametophyte density data were analyzed using two-way ANOVA with SAS version 9.4.
Availability of data and materials
Not applicable.
References
Fernández H, Revilla MA. In vitro culture of ornamental ferns. Plant Cell Tissue Organ Cult. 2003;73:1–13.
Greeshma AA, Sridhar KR. Nutraceutical and bioactive significance of ferns with emphasis on the medicinal fern Diplazium. In: Egamberdieva D, Tiezzi A, editors. Medically important plant biomes: source of secondary metabolites, vol. 15. Singapore: Springer; 2019. p. 115–31.
Liu Y, Wujisguleng W, Long C. Food uses of ferns in China: a review. Acta Soc Bot Pol. 2012;81:263–70.
Langhasova L, Pumprova K, Haisel D, Ekrt L, Pavicic A, Zajíčková M, Vanek T, Dvorakova M. European ferns as rich sources of antioxidants in the human diet. Food Chem. 2021;356:129637.
Korea Forest Service. Production of Forest Products. 2018. http://kosis.kr/statHtml/statHtml.do?orgId=136&tblId=DT_13648_A015&conn_path=I2. Accessed 10 May 2021.
Bligny R, Leguay J-J. Techniques of cell suspension culture. Methods Enzymol. 1978;148:3–16.
George EF. Plant tissue culture procedure-background. In: George EF, Hall MA, De Klerk G-J, editors. Plant propagation by tissue culture. Dordt: Springer; 2008. p. 1–28.
Rybczyński JJ, Tomiczak K, Grzyb M, Mikuła A. Morphogenic events in ferns: single and multicellular explants in vitro. In: Fernández H, editor. Current advances in fern research. Cham: Springer; 2018. p. 99–120.
Kato Y. Physiological and morphogenetic studies of fern gametophyte by aseptic culture III. Growth and differentiation of single cells isolated from callus tissues of Pteris vittata. Cytologia (Tokyo). 1964;29:79–85.
Byrne TE, Caponetti JD. Morphogenesis in three cultivars of Boston fern. II. Callus production from stolon tips and plantlet differentiation from callus. Am Fern J. 1992;82:1–11.
Hegde S, Menon VK, Noronha R, D’Souza L. Callus culture and an unconventional pattern of sporophyte regeneration in Drynaria quercifolia—a medical fern. In Vitro Cell Dev Biol Plant. 2006;42:508–13.
Teng WL, Teng MC. In vitro regeneration patterns of Platycerium bifurcatum leaf cell suspension culture. Plant Cell Rep. 1997;16:820–4.
Camloh M, Gogala N. In vitro culture of Platycerium bifurcatum gametophytes. Sci Hortic. 1992;51:343–6.
Douglas GE, Sheffield E. The investigation of existing and novel artificial growth systems for the production of fern gametophytes. In: Ide JM, Jermy AC, Paul AM, editors. Fern horticulture: past, present and future perspectives. Andover: Intercept; 1992. p. 183–7.
Douglas GE, Sheffield E. A new technique for the culture of fern gametophytes. Plant Cell Rep. 1990;8:632–4.
Sheffield E, Douglas GE, Cove DJ. Growth and development of fern gametophytes in an airlift fermenter. Plant Cell Rep. 1997;16:561–4.
Menéndez V, Revilla MA, Bernard P, Gotor V, Fernández H. Gibberellins and antheridiogen on sex in Blechnum spicant L. Plant Cell Rep. 2006;25:1104–10.
Menéndez V, Revilla MA, Fernández H. Growth and gender in the gametophyte of Blechnum spicant L. Plant Cell Tissue Organ Cult. 2006;86:47–53.
Menéndez V, Abul Y, Bohanec B, Lafont F, Fernández H. The effect of exogenous and endogenous phytohormones on the in vitro development of gametophyte and sporophyte in Asplenium nidus L. Acta Physiol Plant. 2011;33:2493–500.
Kosakivska IV, Vasyuk VA, Voytenko LV, Shcherbatiuk MM, Romanenko KO, Babenko LM. Endogenous phytohormones of fern Polystichum aculeatum (L.) Roth gametophytes at different stages of morphogenesis in vitro culture. Cytol Genet. 2020;54:23–30.
Romanenko KO, Babenko LM, Vasheka OV, Romanenko PO, Kosakivska IV. In vitro phytohormonal regulation of fern gametophytes growth and development. Russ J Dev Biol. 2020;51:71–83.
Murphy KP, Nicholas JR, Leifert C, Lumsden JP. Growth and mineral nutrition in micropropagated delphinium during a subculture period. In: Lumsden PJ, Nicholas JR, Davies WJ, editors. Physiology, growth and development of plants in culture. Dordt: Springer; 1994. p. 72–6.
Gamborg OL, Murashige T, Thorpe TA, Vasil IK. Plant tissue culture media. In Vitro. 1976;12:473–8.
Kuriyama A, Hojoh T, Sugawara Y, Matsushima H, Takeuchi M. A method for the rapid growth in culture of gametophytes of Equisetum arvense with antheridia. Plant Cell Physiol. 1989;30:1189–92.
Renner GDR, Randi ÁM. Effects of sucrose and irradiance on germination and early gametophyte growth of the endangered tree fern Dicksonia sellowiana Hook (Dicksoniaceae). Acta Bot Bras. 2004;18:375–80.
Ravi BX, Robert J, Gabriel M. In vitro spore germination and gametophytic growth development of a critically endangered fern Pteris tripartita Sw. Afr J Biotehnol. 2014;13:2350–8.
Jang BK, Cho JS, Park K, Lee CH. Practical methodology for gametophyte proliferation and sporophyte production in green penny fern (Lemmaphyllum microphyllum C. Presl) using mechanical fragmentation. In Vitro Cell Dev Biol Plant. 2020;56:318–24.
Praptosuwiryo TN, Isnaini Y. Morphological variations and sex expression in gametophytes of Cibotium barometz under in vitro conditions. Biodiversitas. 2017;18:312–20.
Rivera A, Conde P, Cañal MJ, Fernández H. Biotechnology and apogamy in Dryopteris affinis spp. affinis: the influence of tissue homogenization, auxins, cytokinins, gibberellic acid, and polyamines. In: Fernández H, editor. Current advances in fern research. Cham: Springer; 2018. p. 139–52.
Swami P, Raghavan V. Control of morphogenesis in the gametophyte of a fern by light and growth hormones. Can J Bot. 1980;58:1464–73.
Chia SGE, Raghavan V. Abscisic acid effects on spore germination and protonemal growth in the fern Mohria caffrorum. New Phytol. 1982;92:31–7.
Albaum HG. Inhibitions due to growth hormones in fern prothallia and sporophytes. Am J Bot. 1938;25:124–33.
Banks JA. Sex determination in the fern Ceratopteris. Trends Plant Sci. 1997;2:175–80.
Tanaka J, Yano K, Aya K, Hirano K, Takehara S, Koketsu E, Ordonio RL, Park SH, Nakajima M, Ueguchi-Tanaka M, Matsuoka M. Antheridiogen determines sex in ferns via a spatiotemporally split gibberellin synthesis pathway. Science. 2014;346:469–73.
Romanenko KO, Kosakivska IV, Babenko LM, Vasheka OV, Romanenko PO, Negretsky VA, Minarchenko VM. Effects of exogenous cytokinins on spore germination and gametophyte morphogenesis of Dryopteris filix-mas (L.) Schott in vitro culture. Cytol Genet. 2019;53:192–201.
Camloh M, Ravnikar M, Zel J. Jasmonic acid promotes division of fern protoplasts, elongation of rhizoids and early development of gametophytes. Physiol Plant. 1996;97:659–64.
Grzyb M, Kalandyk A, Mikuła A. Effect of TIBA, fluridone and salicylic acid on somatic embryogenesis and endogenous hormone and sugar contents in the tree fern Cyathea delgadii Sternb. Acta Physiol Plant. 2018;40:1.
Zhu YX, Davies PJ, Halinska A. Metabolism of gibberellin A12 and A12-aldehyde in developing seeds of Pisum sativum L. Plant Physiol. 1991;97:26–33.
Gaspar T, Kevers C, Penel C, Greppin H, Reid DM, Thorpe TA. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell Dev Biol Plant. 1996;32:272–89.
Davies PJ. Regulatory factors in hormone action: level, location and signal transduction. In: Davies PJ, editor. Plant hormones. Dordt: Springer; 2010. p. 16–35.
Jang BK, Cho JS, Kwon HJ, Lee CH. Optimal conditions for spore germination and gametophyte and sporophyte production in the autumn fern Dryopteris erythrosora. Hortic Environ Biotechnol. 2019;60:115–23.
Knop W. Quantitative untersuchungen über den ernährungsprozess der pflanzen. Landwirtsch Vers Stn. 1865;7:93–107.
Murashige T, Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant. 1962;15:473–97.
Acknowledgements
Not applicable.
Funding
This research was supported by Chungbuk National University Korea National University Development Project (2020).
Author information
Authors and Affiliations
Contributions
BKJ and CHL: Conceptualization; BKJ, JSC and SHK: Methodology; BKJ: Writing—Original Draft Preparation; BKJ and CHL: Writing—Review and Editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13007_2021_786_MOESM1_ESM.docx
Additional file 1: Table S1. Effect of gametophyte morphogenesis in accordance with culture type on sporophyte formation and growth of eastern bracken in ex vitro conditions. Figure S1. Experimental design diagram of ex vitro sporophyte propagation according to culture types and periods. zLiquid culture is more efficient for gametophyte proliferation than solid culture (shorter culture period and easier handling).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Jang, BK., Cho, JS., Kang, SH. et al. Culture types and period impact gametophyte morphogenesis and sporophyte formation of eastern bracken. Plant Methods 17, 87 (2021). https://doi.org/10.1186/s13007-021-00786-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13007-021-00786-7