
Abstract
In this review, a comprehensive discussion exclusively on bacterial xylanases; their gene organization; different factors and conditions affecting enzyme yield and activity; and their commercial application have been deliberated in the light of recent research findings and extensive information mining. Improved understanding of biological properties and genetics of bacterial xylanase will enable exploitation of these enzymes for many more ingenious biotechnological and industrial applications.
Similar content being viewed by others

Introduction
Xylan is the second most abundant polysaccharide in nature present in both hard woods and annual plants. This homopolysaccharide is as abundant as cellulose accounting for approximately one-third of the renewable organic carbon sources on the earth (Kamble and Jadhav 2012). Structure of xylan varies among different plant species and its homopolymer backbone chain can be substituted with different side chain groups at various positions (Wang et al. 2014). Owing to its heterogeneity and complexity, complete hydrolysis of xylan requires variety of cooperatively acting enzymes collectively known as xylanases. Among various groups of microorganisms, bacteria and fungi are endowed with powerful xylanolytic machinaries. Xylanolytic microorganisms have been reported from various extreme environments, such as thermal springs (Bouacem et al. 2014), marines (Annamalai et al. 2009), Antarctic environments (Bradner et al. 1999), and soda lakes (Huang et al. 2015). Xylanases have wide range of industrial and biotechnological applications. Their commercial exploitation in the area of food (Harris and Ramalingam 2010), feed, and paper and pulp industries (Polizeli et al. 2005) are well documented. Recently, xylanases are also being used to increase the sugar recovery from agricultural residues for biofuel production (Gonçalves et al. 2015).
Due to huge industrial applications, a significant research effort has been devoted towards mining and characterization of xylanases. Initial focus had been on the fungal xylanases due to their high activity, but constraints faced during mass production and industrial applications, placed bacterial xylanases as a tough competitor in the industrial arena. Distribution, properties, genetics, and application of the bacterial xylanases have been discussed in this review to highlight their real potential as a promising source.
Xylan and xylanases
Structurally xylan is a homopolymer of D-xylopyranose residues in β (1 → 4) linkages with a degree of polymerization ranging from 150 to 200. This backbone is substituted by some of the sugars and organic acids, such as arabinose, glucuronic acid, ferulic acid, etc. Xylans are broadly categorized into four major groups based on its substituents, viz., homoxylan, arabinoxylan, glucuronoxylan, and glucuronoarabinoxylan. Homoxylans contain xylose residues only, and can be either linear or branched (Sun et al. 2011). Arabinoxylans consist of a (1 → 4)-β-xylan main chain, but is substituted with α-arabinosyl residues. The β-(1 → 4)-linked d-xylopyranosyl residues are substituted with one α-(1 → 2)-linked 4-O-methyl-d-glucuronic acid in the case of glucuronoxylan; while in glucuronoarabinoxylans, the same backbone is linked to arabinofuranose and uronic acid (Gröndahl et al. 2003; Bergmans et al. 1996). The side chains determine the solubility, physical conformation, and reactivity of xylan molecule with other hemicellulosic components, and, hence, greatly influence the mode and extent of enzymatic cleavage.
Due to complexity in its structure, xylan needs different enzymes collectively termed as xylanases for its complete hydrolysis. Xylanases basically belong to hydrolase group of enzymes, precisely to glycoside hydrolases. Sequence-based glycoside hydrolase classification has placed xylanase in two families GH10 and GH11, but xylanases are also found in other glycoside hydrolase families, such as GH5, 7, 8, and 43. Plant, fungal, and bacterial enzymes comprise GH10 family, whereas GH11 family which is structurally unrelated includes only fungal and bacterial enzymes (Lafond et al. 2014). At least ten subfamilies of xylanases, some of which are restricted to fungi (xylanases Ia, Ib, Ic, II, IIIa, IIIb, and IV), and others to bacteria (A, B, C). GH10 family is composed of both endo-1,4-β-xylanases and endo-1,3-β-xylanases, but the majority being endo-1,4-β-xylanases with few endo-1,3-β-xylanases. They have greater catalytic versatility and can catalyze hydrolysis of even cellulose and aryl β-d-cellobiosides. In mixed linkage xylan, GH10 xylanases can attack on β-1,4-linkages that precede a β-1,3-linkage, but not the ones that immediately follow β-1,3-linkages. GH10 xylanases can attack β-1,3-linkages flanked on both sides by β-1,4-linkages. This family of xylanases can also tolerate replacement of one or two consecutive xylose residues by glucose residues in the substrate. Members of this family are capable of hydrolyzing the aryl β-glycosides of xylobiose and xylotriose at the aglyconic bond. Furthermore, these enzymes are highly active on short xylooligosaccharides, thereby indicating small substrate-binding sites (Pollet et al. 2010). Analyses of crystal structure, kinetic activity on xylooligosaccharides, and end products have indicated that family 10 xylanases typically have four-to-five substrate-binding sites. Most of the GH 10 xylanases typically have high molecular mass, low pI, and (α/β)8-barrel fold conformation (Teplitsky et al. 2000). GH11 members are monospecific, as they consist exclusively of true endo-β-1,4-xylanases that cleave internal β-1,4-xylosidic bonds. Their catalytic versatility is lower than GH10 members, and the products of their action can be further hydrolyzed by the family 10 enzymes. These are considered as “true xylanases”, because of their exclusive action on d-xylose containing substrates. Members of this family have low molecular mass, high pI, and a wide range of pH optima varying from 2 to 11. Like GH10 xylanases, these enzymes can hydrolyze the aryl β-glycosides of xylobiose and xylotriose at the aglyconic bond, but, unlike GH10, they cannot cleave cellulose or aryl β-d-cellobiosides. GH11 preferably cleaves un-substituted regions of the backbone, since they cannot attack the xylosidic linkage towards the non-reducing-end next to a branched xylose. Structure of GH11 enzymes principally consist of 3 β-pleaded sheets and one α-helix (Biely et al. 1997; Collins et al. 2005; Pollet et al. 2010). Some genera of fungi and bacteria produce more than one subfamily of xylanases. This means a possible duplication of family 11 xylanases might have occurred before the divergence between eubacteria and other domains (Sapag et al. 2002). GH5 family of xylanases is not as prevalent as of their GH10 and GH11 families but play an important role in complementing the action of GH10 and GH11 enzymes during depolymerization of glucuronoxylans in lignocellulosic fibres (Gallardo et al. 2010). GH8 xylanases have both high pI and molecular weight. Six enzymes from this family have been shown to have xylan degrading activity (Collins et al. 2012; Hong et al. 2014). GH8 xylanases have substrate specificity and action pattern similar to GH11 enzymes (Collins et al. 2006).
Xylanolytic bacteria and their xylanases
Xylanases are produced by fungi, bacteria, yeast, marine algae, protozoa, snails, crustaceans, insect, seeds, etc. (Polizeli et al. 2005) Bacterial genera, such as Bacillus, Cellulomonas, Micrococcus, Staphylococcus, Paenibacillus, Arthrobacter, Microbacterium, Pseudoxanthomonas, and Rhodothermus have been reported to produce xylanases (Subramanian and Prema 2000; Beg et al. 2001; Gupta et al. 2001; Chapla et al. 2012). Among the actinomycetes group, Streptomyces, Actinomadura, Nonomuraea, etc. are commonly reported for xylanase production (Prakash et al. 2013).
Prevalence of the xylanolytic bacteria have been reported from most of the bacterial groups. Various kinds of xylanases with thermostability, cold adaptivity, or alkalo-stability have been mined and purified from diverse microorganisms, including extremophilic bacteria (Table 1). Among bacteria, Bacillus was found to be a potential source of xylanases, and a number of bacilli, such as B. circulans, B. stearothermophilus, B. amyloliquefaciens, B. subtilis, B. pumilus, and B. halodurans (Subramanian and Prema 2002; Thomas et al. 2014; Banka et al. 2014; Gupta et al. 2015) have been reported to have considerable xylanolytic activity. Thermostable xylanases active at temperature as high as 60–70 °C have been reported from Bacillus spp., Stenotrophomonas maltophila, Rhodothermus marinus, Thermotoga sp., Clostridium thermocellum, and Streptomyces sp. (Kumar and Satyanarayana 2013; Raj et al. 2013; Thomas et al. 2014). Although cold-adaptive xylanases are not very common, but bacteria, such as Flavobacterium frigidarium and Clostridium sp. PXLY1 (Akila and Chandra 2003; Humphry et al. 2001) have been reported to produce such xylanases. Bacteria have an advantage over fungi for xylanase production as pH optimum for bacterial xylanases are in neutral or alkaline range, whereas for fungal xylanases, it is in the acidic range. The low pH requirement for growth of fungi and production of fungal xylanases necessitates additional steps in the subsequent stages which make fungal xylanases less attractive. Firmicutes, such as B. pumilus, B. halodurans, and Geobacillus thermoleovorans (Verma and Satyanarayana 2012), and actinomycetes, such as Streptomyces sp. abd Actinomadura sp. (Luo et al. 2015; Taibi et al. 2012) have been reported as potential source of alkali stable xylanases. Xylanases with such unique features have found diverse application in various industries.
Comparison of bacterial and fungal xylanases
Majority of the bacterial xylanases belong to GH10 family, whereas fungal xylanases majorly belong to GH11 family (Liu et al. 2011). Endoxylanases from bacteria and fungi display exclusively single subunit protein structures with molecular weight (MW) ranging from 8.5 to 85 kDa and their isoelectric point (pI) values between 4.0 and 10.3 and most of them show glycosylation (Polizeli et al. 2005). In-silico structural comparison of bacterial and fungal xylanases shows difference in their secondary structure especially at the loop areas. Bacillus circulans and B. subtilis mainly have beta sheet structure, while fungal xylanase, especially Aspergillus niger, have alpha helices and also have difference in various loop regions though both belonged to class GH11. A common thing found in both fungal and bacterial xylanases is that the nucleophile and proton donor are always the Glutamic acid, though their position may change (Mathur et al. 2015). Fungal xylanases are produced simultaneously with cellulase that increases length of downstream processing; while in bacteria, xylanase are produced alone mostly, thereby reducing the downstream process time. Low pH requirement for growth of fungi and production of cellulase necessitates additional steps in downstream processing which make fungal xylanase less attractive (Subramanian and Prema 2002).
Genetics of bacterial xylanases
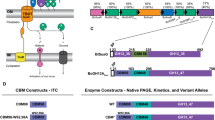
Bacillus subtilis and Clostridium acetobutylicum are two well-studied xylanolytic firmicutes. Whole genome sequencing of B. subtilis str 168 has provided in sight of the organization of the genes-coding xylanolytic enzymes and transporters for the utilization of xyloses. The genes-coding xylanolytic enzymes of B. subtilis str 168 are located in the chromosomal DNA. Four major genes, viz., xynA (1241 bp), xynB (1601 bp), xynC (641 bp), and xynD (1241 bp) are reported (Fig. 1a). xynA encodes for endo-1,4 beta-xylanase (GH11 family) and depolymerizes xylan to produce methyl glucurono xylotetraose and xylooligosaccharides. xynB codes for β-xylosidase, while xynC encodes GH30 family endoxylanase (glucuronoxylanases). Generation of a series of aldouronates with an increasing number of xylose residues and a single methylated glucuronoxylan (MeG) linked-1, 2 to penultimate xylose residue from reducing-end is a characteristic of XynC. xynD encodes arabinoxylan arabinofuranohydrolase that catalyzes the release of free arabinose from methyl galacturono xylans. XynD has higher activity on arabinoxylans compared to oligo-arabinoxylosides. β-xylanase is synthesized constitutively and in contrast to many other extracellular enzymes, it is synthezised mostly during exponential phase. On the other hand, β-xylosidase synthesis has been found to be strongly induced by xylose and xylan (Lindner et al. 1994). Xylose induction for the genes of the xynCB and xylAB operons had been described previously by Gartner et al. (1988) and Hastrup (1988). xylA (1337 bp) encodes for xylose isomerase and xylB codes for xylulose kinase. Upstream to the xynB, another gene xynP (1392 bp) encodes for sugar (glycoside-pentoside-hexuronide) transporter protein. The genes of the xynCB and xylAB (involved in xylose utilization) operons are controlled by the transcriptional regulator encoded by xylR (1053 bp). Expression of xylAB is negatively regulated at the transcriptional level by the regulator XylR.

a Organization of xylanase genes on chromosome of B. subtilis str 168. b Organization of xylanase gene on plasmid of C. acetobutylicum ATCC824. c Diversity of xylanolytic bacteria based on composition and copy number of xylanase genes (clustering was performed based on weighted pair average and Manhattan distances using Multi-Experiment Viewer)
In case of Clostridium acetobutylicum ATCC 824, the xylanase encoding genes are located both on plasmid and chromosome. The organization of xylanase encoding genes on plasmid is presented in Fig. 1b. Like B. subtilis, C. acetobutylicum also have xynB (957 bp) and xynD (1376 bp) genes. C. acetobutylicum contains at least two homologous copies of xynC (CAP_0118, 1766 bp and CAP_0119, 1758 bp). xynD encodes GH43 endo-1,4 beta-xylanase, while xynA encodes a GH35 endoxylanase. In addition, C. acetobutylicum encodes (CAP_0114, 1625 bp) another GH35 endoxylanase which also includes merged domains for arabinoxylan arabinofurano hydrolase, β-xylosidase, and α-l-arabinofuranosidase. The plasmid harbours a 1283 bp gene (CAP_0117) for β-xylosidase. xynD (CA_C 3452) encoding another GH43 hydrolase is also located on chromosome. Upstream to this xynD gene, xynT (1377 bp) encodes a xyloside transporter protein. In C. acetobutylicum, the xylose utilization pathway also includes a xylulokinase (XylB, CAC2612), a xylose proton-symporter (XylT, CAC1345), and a transcriptional regulator (XylR, CAC3673). xynD and xynT along with xyl genes are under transcriptional control of XylR. Bacteria are endowed with varying xylanolytic activity with distinct properties due to diversity in their xylanase genes. Table 2 presents list of genes and their coded products which have been thoroughly characterized at proteomic level, and protein models are available at public databases. However, the differential ability of the bacteria to degrade xylan with varying efficiency and distinct mode of action also depend on the composition and organization of xylanase genes. Global distribution of gene copy number among the xylan degrading gene cassette represented by four genes encoding endo-1,4-beta-xylanase (xynA), xylan 1,4-beta-xylosidase (xynB), glucuronoarabinoxylan endo-1,4-beta-xylanase (xynC), and oligosaccharide reducing-end xylanases (rexA). Whole genome sequence data of 74 xylanolytic bacteria selected from diverse ecological niche, such as soil, rumen, human gut etc., that are available at JGI (Joint Genome Institute) was used to study the xylan degrading gene cassette. The abundance of xylan degrading genes in terms of gene copy number among the selected xylanolyitc bacteria was used to create dual dendrogram. Among the four genes, xynA was most widely distributed, whereas rexA was very rare among the isolates. Presence and high abundance of particular xylan degrading genes help in natural selection of particular bacteria in a specialized ecological niche. For instance, cattle rumen-associated cellulo-xylanolytic bacterium Fibrobacter succinogenes S85 (Fig. 1c) has the highest copy number of rexA gene that endowed the bacterium to synthesise xylo-oligosachharide which supports the growth of other bacteria in rumen. Paenibacillus mucilaginosus KNP414 and Ruminococcus albus 7 were having the highest copy number of all xylan degrading gene and could be very useful for complete structural degradation of hemicellulosic biomass. Knowledge of relative gene copy number and gene composition through whole genome survey could play a vital role in selection of ecological niches for isolation experiments and further selection of bacterial group for mining novel xylanolytic bacteria.
Cloning and expression of xylanase genes
For commercial level production of xylanase, it is necessary to select bacteria which can overproduce xylanase. Recombinant DNA technology is one of the important techniques which provide ways to enhance enzyme production. Xylanase genes from different bacteria have been cloned into suitable hosts for overexpression (Kulkarni et al. 1999). Two xylanase genes (xynA and xynB) from a cellulolytic bacterium Ruminococcus flavefaciens17 were cloned and expressed in a heterologous host E. coli (Flint et al. 1991). Xylanase gene has also been cloned from an extreme thermophilic bacterium Dictyoglomusthermophilum Rt46B.1, employing a genomic DNA library screening approach. Sequencing the subclone is shown to contain a single complete open reading frame coding for a single-domain xylanase, XynA, with a putative length of 352 amino acids (Gibbs et al. 1995). A xylanase gene of 1383 bp coding for 460 amino acids with a molecular mass of 51.3 kDa was cloned from a marine bacterium Vibrio sp. strain XY-214 and expressed in E. coli DH5α. The transformant enzyme hydrolyzed β-1,3-xylan to produce several xylooligosaccharides (Araki et al. 2000). Two genes XynA- and XynB-encoding xylanases from Paenibacillus sp. KCTC 8848P were cloned and expressed in Escherichia coli. The structural gene of XynA 636 bp, encoded a protein of 212 amino acids, while the XynB gene consisted of 951 bp open reading frame for a protein of 317 amino acids. The xylanases of E. coli transformants were released into the extracellular culture fluid in the absence of xylan (Lee et al. 2000). A xylanase gene of 642 bp length was cloned from Bacillus subtilis B10 isolated for degumming of ramie blast fibres. Xylanase gene was expressed and xylanase activity was measured. The xylanase distribution in extracellular, intracellular, and periplasmic fractions was about 22.4, 28.0, and 49.6 %, respectively (Huang et al. 2006). An alkalistable and thermostable xylanase gene Myxl was cloned from a metagenomic library constructed from DNA extracted from the compost-soil. Sequence analysis of the clone revealed a xylanase gene of 1077 bp. The deduced protein sequence (358 amino acids) displayed homology with glycosyl hydrolase (GH) family 11 xylanases. The gene was subcloned into pET28a vector and expressed in E. coli BL21 (DE3). The recombinant xylanase (rMxyl) exhibited activity over a broad range of pH and temperature with optima at pH 9.0 and 80 °C. The recombinant xylanase is highly thermostable having T1/2 of 2 h at 80 °C and 15 min at 90 °C (Verma et al. 2013a, b). A xylanase gene was isolated from Bacillus brevis and expressed in E.coli BL21. The recombinant xylanase was predominantly secreted to culture medium and showed mesophilic nature (optimum activity at 55 °C and pH 7.0). The cell free culture medium exhibited 30 IU/ml xylanse activity. The enzyme did not show any cellulose activity and was active under wide range of temperature and pH (Goswami et al. 2014).
Factors affecting xylanase yield and activity
Although an efficient xylanase producing microorganism is a prerequisite for industrial fermentation of xylanases, the yield during fermentation process is a function of factors, such as nutritional requirements and environmental conditions. Among them, nutritional factors, the carbon sources present in the media, are the most important factors for xylanase production. A number of substrates, such as wheat bran, rice bran, soy meal, sugarcane bagasse, etc., have been found to be suitable substrates for xylanase production (Kumar et al. 2009; Raj et al. 2013). Murugan et al. (2011) reported xylanase production (117 U/ml) using saw dust as substrate for Arthrobacter sp. MTCC6915 under solid-state fermentation (SSF), while Sepahy et al. (2011) reported xylanase activity (249.308 IU/ml) in Bacillus mojavensis AG137 using oat bran under submerged fermentation. Bacillus subtilis ASH have been reported to produce xylanase with wheat bran, wheat straw, rice husk, saw dust, gram bran, groundnut, and maize bran under SSF (Sanghi et al. 2008). Boonchuay et al. (2016) reported xylanase production from Streptomyces thermovulgaris TISTR1948 using corn cob. Wheat bran has been reported to be the best substrate for the maximum production of xylanase enzyme (Sanghi et al. 2008; Kapoor et al. 2008; Nagar et al. 2010; Kumar et al. 2013). The reason might be due to the fact that it contains 54 % carbohydrates (pentoses and hexoses), 14 % protein, minerals, amino acids, and vitamins (El-Sharnouby et al. 2012), which supports the growth of the bacterium and hence xylanase production. In addition, substrate accessibility, rate, and quantity of release of the xylooligosaacharides, their chemical nature, and quantity of xylose released also influence the production of xylanases during fermentation process. Guha et al. (2013) reported sugarcane bagasse as best inducer for xylanase production from Bacillus sp. Yeast extract, beef extract, peptone, etc. can serve as good nitrogen source for yielding xylanase. Enhanced levels of xylanase from Streptomyces cyaneus SN32, Bacillus sp., Streptomyces sp., Streptomyces sp. CA24 have been reported using yeast extract, peptone, and beef extract as organic nitrogen source (Porsuk et al. 2013). Apart from the substrates, the method of fermentation influences the number of isoforms produced by bacterium. For example, wheat bran induced six isoforms under SSF, but only one under submerged fermentation in Simplicillium obclavatum MTCC 9604. Surprisingly, three among six isoforms were of similar molecular weight, but with different pI which helps them to achieve effective hydrolysis of xylan in wider pH range (Roy et al. 2013). In the case of Providencia sp. strain X1, it produced seven isoforms with wheat bran, but only two with birchwood xylan (Raj et al. 2013). The need for multiple forms of xylanase could be due to the complexity in lignocellulosic biomass to achieve maximum sugar yield, and they could arise from post-translational modification of a gene product, such as differential glycosylation or proteolysis (Sharma and Chadha 2011). Besides nutritional components, other bioprocess parameters, such as pH of the medium, incubation temperature, agitation, initial load of inoculum, etc., also influence the xylanase production during fermentation process. Among these factors, pH and temperature determine the activity and stability of enzyme. Xylanases obtained from bacterial sources are known to be active and stable in wide range of pH and temperature. Xylanase enzyme exhibit its optimal activity wide ranges of incubation conditions, such as temperature from 30 to 60 °C, pH from 5.0 to 9.0. In many cases, the optimum conditions for production are differ from its activity. For example, the temperature for xylanse production was 30 and 37 °C, but it showed its optimal activity at 60 and 55 °C, respectively (Murugan et al. 2011; Sepahy et al. 2011). Inoculum size of 1 % and agitation rate of 120 or 200 proved to be the suitable parameters for maximum production of xylanase under submerged fermentation (Mittal et al. 2012; Raj et al. 2013; Kaur et al. 2016). Presence of carboxymethyl cellulose, dextrose, peptone, and beef extract supported the maximum xylanase production in the range from 161.4 to 176.4 U/ml (Murugan et al. 2011). Table 3 elaborates factors, such as substrate, pH, and temperature, affecting the activity of some selected bacterial xylanases.
Application of xylanases
Most of the bacterial xylanases are alkali- and/or thermostable and could be free of cellulose activity. These features make them ideal candidate for industrial applications. A number of bacterial xylanases, including their production technology, have been patented for various applications, and have been marketed for numerous commercial applications worldwide (Table 4). Some of the major applications of bacterial xylanases are discussed below.
Paper and pulp industry
Xylanase enzymes to be used for biobleaching of pulps must be active at high temperatures and alkaline pH, and must not contain cellulolytic enzymes to preserve the cellulose fibres (Polizeli et al. 2005). Any cellulase activity will have serious economic implications in terms of cellulose loss, degraded pulp quality, and increased effluent treatment cost. Xylanases from various microbes have been screened for their efficiency in enzyme aided bleaching of wood pulp. Many bacterial xylanases binds to both cellulose and insoluble xylan, but the enzyme has activity against only xylan. Thus, fungal-based xylanases have a lesser degree of hemicellulose removal for a given charge than those of bacterial-based enzymes. In comparison to fungal xylanases, bacterial enzymes tend to produce a significant kappa reduction. The fungal enzymes are thought not to exhibit the same level of tenacity toward carbohydrate binding domains; hence, they are less aggressive than bacterial-based enzymes (Wong et al. 1988). Use of xylanase can significantly reduce the amount of chlorine used for bleaching of pulp (Viikari et al. 1986; Paice et al. 1988; Clarke et al. 1991; Jeffries et al. 1992) Biobleaching of eucalyptus kraft pulp using xylanase from B. pumilus ASH5 resulted in reduction of chlorine and chlorine dioxide consumption by 20 and 10 %, respectively, besides improved brightness and whiteness of pulp (Battan et al. 2007). Chapla et al. (2012) reported similar result using cellulose free thermostable xylanase from Paenibacillus sp. ASCD2 in prebleaching of eucalyptus kraft pulp.
Deinking of waste paper
One of the important steps in recycling of waste paper is the deinking process which involves dislodgement of ink particles from paper. Conventionally, large amount of chlorine, chlorine-based derivatives, sodium hydroxide, sodium carbonate, sodium silicate, hydrogen peroxide, hypochlorites, and chelating agents have been used which resulted in hazardous effluent disposal problem (Maity et al. 2012). Recently, xylanase and laccase enzymes have been reported to remove ink from effluent generated from paper and pulp industries (Dhiman et al. 2014; Chandra and Singh 2012). An attempt has been made to deink old newsprint using the combined applications of xylanase and laccase. Combined action resulted in significant increase in brightness (11.8 %), whiteness (39 %) and physical properties like breaking length (34.8 %), burst factor (2.77 %), and tear factor (2.4 %). Synergism between xylanase and laccase during co-treatment would have positively influenced physical properties, whereas chemical treatment for deinking would has deteriorated freeness and strength of the recycled paper (Gupta et al. 2015). Kumar and Satyanarayana (2014) tried different combinations of commercial cellulase and xylanase from Bacillus haloduransTSEV1. They found that xylanase dose of 1.2 U/mg along with commercial cellulase at 1.2 U/mg was efficient in ink removal.
Animal feeds
Xylanases are used to improve digestibility of animal feed. Low viscosity in some cereals, such as maize and sorghum, is attributed to their arabinoxylan content which renders them unsuitable as feed. Xylanase addition to feeds improves digestion in the initial part of digestive tract resulting in better energy use (Harris and Ramalingam 2010). Clarkson et al. (1999) reported better digestibility of cereal feeds for poultry resulting in reduction in feed conversion ratio (FCR) using endoxylanases from Acidothermus cellulolyticus. Babalola et al. (2006) observed improved apparent nitrogen and fiber absorption as well as feed transit time. In general, a combination of different enzymes is supplemented with animal feed to achieve favorable results. A wheat based diet supplemented with combination of phytase, xylanase, and β-glucanase improved the performance of broiler chickens. Improved performance is attributed to reduced digesta viscosity, increased apparent metabolisable energy, reduced relative weight, and length of small intestine (Wu et al. 2004). A number of bacterial xylanases have been commercially available for uses as feed additive.
Bakery industry
Presence of hemicelluloses, such as arabinoxylans, in wheat, the key raw material of baking industry, pose serious constraints in making of good quality dough. The ability of xylanases to solubilize water unextractable arabinoxylan (WU-AX) and its low activity on water extractable arabinoxylan (WE-AX) results in better performance during bread making as a result of its solubilized arabinoxylan (Courtin and Delcour 2002). Bacterial xylanases like GH11 endoxylanases from B. subtilis solubilize WU-AX, in particular, increases the viscosity of dough and has a negative effect on gluten agglomeration (Butt et al. 2008). Xylanase transforms water insoluble hemicellulose into soluble form, which binds water in the dough, therefore, decreasing the dough firmness, increasing volume, and creating finer and more uniform crumbs (Butt et al. 2008).
Biofuel production
Conversion of abundant lignocellulosic plant biomass into biofuels presents a viable option for improving energy security and reducing greenhouse emissions (Kumar et al. 2009). One of the major limitations is the recalcitrant nature of the plant cell wall, which is composed mostly of lignocellulosic materials, such as lignin, cellulose, and hemicelluloses. Xylanase, together with other hydrolytic enzymes, such as cellulases and laccases can be used for the generation of biofuels, such as ethanol, from lignocellulosic biomass. Bacteria are rich source of thermostable xylanases and are generally preferred for lignocellulose hydrolysis. Robust thermostability of bacterial xylanases make them well suited for the harsh processing conditions required during deconstruction of lignocellulose to fermentable products. Bhalla et al. (2015) reported higher hydrolytic conversion of Birchwood xylan (68.9 %) with Geobacillus sp. strain WSCUF1 as compared to commercial enzymes, such as Celic HTec2 (49.4 %) and Accelerase XY (28.92 %), at 70 °C. The supplementation of xylanases during enzymatic saccharification increased the availability of reducing sugars which ultimately results in high bioethanol production.
Pharmaceutical industry
Hydrolytic products of xylan known as xylooligosaccharides (XOs) are mixtures of oligosaccharides formed by xylose residues linked through β-1,4 linkages. On the basis of number of xylose residues, they are known as xylobiose, xylotriose, xylotetrose, and so on. Xylobiose is ideal XO for food application as prebiotic. XOS are produced from xylan containing lignocellulosic biomass (LCM) by chemical, enzymatic (Katapodis et al. 2002), and combination of both the methods (Kokubo and Ikemizu 2004). For the production of XOs, the enzyme complex should have low exoxylanase or β-xylosidase activity, to prevent the production of high amounts of xylose, which has inhibitory effects on XOs production (Liu and Liu 2008; Boonchuay and Chaiyaso 2012; Verma et al. 2013a, b; Jain et al. 2015). XOs have a huge commercial market due to their health promoting properties. Besides chemical processes, enzymatic hydrolysis of xylans using endoxylanases from bacterial genera, such as Bacillus, Streptomyces, Rhodothermus, Thermobifida, etc., are being used. Detailed genomic information has provided opportunities for engineering bacterial strains to produce desired XOs. e.g. Rhee et al. (2014) engineered B. subtilis str 168 to produce acidic XOs.
Future perspectives
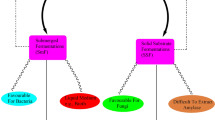
In many aspects of biotechnological application, bacterial xylanases are advantageous as compared to fungal xylanases. Stability and activity in alkaline range, high thermosatbility, low cellulase activity, greater aggression towards xylans due to high tenacity of carbohydrate domains, and ability to selectively solubilize arabinoxylan make bacterial xylanases amenable to specific industrial application. However, it is very difficult to mine super-xylanases with all these features. Most of the available bacterial xylanases have only few of these properties which make industrial processes to depend much on expensive and hazardous chemical processes. Random mutation through UV or chemical mutagenesis has been a classical way to improve xylanolytic strains for high xylanase activities or inhibiting undesirable activities. However, availability of information on whole genome sequence and metabolic pathways have paved the way for engineering xylanase producing strains suitable for specific industrial application, such as biobleaching of paper pulps, production of biofuel, or XOs. Unlike silencing some properties, adding useful features to xylanase enzymes seems to be more challenging. With the boom of the bioinformatics and public access to whole genome sequences, now, it is possible to design xylanases with industrially desirable features. The techniques used in drug designing like in-silico mutagenesis or simulation of molecular dynamics can be very helpful for designing super xylanases as well as engineering superbugs with multiple catalytic activity. Many of the industrial applications, such as biofuel production, need an array of enzymatic activity for biomass deconstruction. Engineering fusion xylanases with cellulolytic and lignolytic activity may be the choice of the future for biofuel production. However, before going into the advanced techniques, the basic research on isolation of xylanolytic bacteria with novel/high yielding xylanases remains the foremost challenge. Figure 2 presents a comprehensive summary of the probable strategies for designing or engineering super-xylanases. Selection of ecological niche and group of target bacteria is very crucial and can be guided by the genomic information on the abundance of genes responsible for xylanase activities. In the present era of molecular biology and biotechnology, it is not very far to have multi-speciality superxylanases from bacteria which will be better than the best fungal xylanases.

Strategies for designing super-xylanases with novel properties
Conclusion
Increased awareness about environmental pollution by chemical operations in many industries paved a way for usage of more energy efficient, economically viable, and environmental friendly means, such as enzymes. Xylanases are known to have many applications in various industrial processes. Till now, most of the known xylanases are obtained from fungi. Bacterial xylanases offer more competitive and milder operating conditions that put them as a quick substitution for fungal xylanases. Thermo and alkali stable bacterial xylanases have been studied from many bacteria. This gives us a fair idea about genomics and their biochemistry. Modern techniques have further simplified the screening and identification procedures for xylanases. Combination of various methods would help further speed up the research and understanding of bacterial xylanases, so as to have an efficient enzyme to satisfy the want for environmental protection. An in-depth understanding of biology and biochemistry of bacterial xylanases will offer a better scope to search or tailor highly efficient xylanases for effective utilization for industrial purposes.
References
Akila G, Chandra T (2003) A novel cold-tolerant Clostridium strain PXYL1 isolated from a psychrophilic cattle manure digester that secretes thermolabile xylanase and cellulase. FEMS Microbiol Lett 219:63–67
Annamalai N, Thavasi R, Jayalakshmi S, Balasubramanian T (2009) Thermostable and alkaline tolerant xylanase production by Bacillus subtilis isolated from marine environment. Indian J Biotechnol 8:291–297
Araki T, Inoue N, Morishita T (1998) Purification and characterization of β-1,3-xylanase from a marine bacterium, Alcaligenes sp. XY-234. J Gen Appl Microbiol 44:269–274
Araki T, Tani S, Maeda K, Hashikawa S, Nakagawa H, Morishita T (1999) Purification and characterization of β-1,3-xylanase from marine bacterium Vibrio sp. Xy-214. Biosci Biotechnol Biochem 63:2017–2019
Araki T, Hashikawa S, Morishita T (2000) Cloning, sequencing, and expression in Escherichia coli of the new gene encoding β-1,3-xylanase from a Marine Bacterium, Vibrio sp. strain XY-214. Appl Environ Microbiol 66:1741–1743
Babalola T, Apata DF, Atteh JO (2006) Effect of β-xylanase supplementation of boiled castor seed meal-based diets on the performance, nutrient absorbability and some blood constituents of pullet chicks. Trop Sci 46:216–223
Bai W, Xue Y, Zhou C, Ma Y (2012) Cloning, expression and characterization of a novel salt tolerant xylanase from Bacillus sp. SN5. Biotechnol Lett 34:2093–2099
Bajaj BK, Singh N (2010) Production of xylanase from an alkali-tolerant Streptomyces sp. 7b under solid-state fermentation, its purification and characterization. Appl Biochem Biotechnol 162:1804–1818
Banka AL, Guralp SA, Gulari E (2014) Secretory expression and characterization of two hemicellulases, xylanase, and β-xylosidase, isolated from Bacillus Subtilis M015. Appl Biochem Biotechnol 174:2702–2710
Barabote RD, Parales JV, Guo Y-Y, Labavitch JM, Parales RE, Berry AM (2010) Xyn 10A, a thermostable endoxylanase from Acidothermus cellulolyticum 11B. Appl Environ Microb 76:7363–7366
Battan B, Sharma J, Dhiman SS, Kuhad RC (2007) Enhanced production of cellulase-free thermostable xylanase by Bacillus pumilus ASH and its potential application in paper industry. Enzyme Microb Technol 41:733–739
Beg QK, Kapoor M, Mahajan L, Hoondal GS (2001) Microbial xylanases and their industrial applications: a review. Appl Microbiol Biotechnol 56:326–338
Bergmans MEF, Beldman G, Gruppen H, Voragen AGJ (1996) Optimisation of the selective extraction of (glucurono) arabinoxylans from wheat bran: use of barium and calcium hydroxide solution at elevated temperatures. J Cereal Sci 23:235–245
Bhalla A, Bischoff KM, Sani RK (2015) Highly thermostable xylanase production from a thermophilic Geobacillus sp. strain WsUcF1 utilizing lignocellulosic biomass. Front Bioeng Biotechnol 3:84
Biely P, Vrsanská M, Tenkanen M, Kluepfel D (1997) Endo-β-1,4-xylanase families: differences in catalytic properties. J Biotechnol 57:151–166
Boonchuay P, Chaiyaso T (2012) Enzymatic production of xylooligosaccharides from corncob using endoxylanase from Streptomyces thermovulgaris TISTR1948. In: 1st Mae Fah Luang University international conference
Boonchuay P, Takenaka S, Kuntiya A, Techapun C, Leksawasdi N, Seesuriyachan P, Chaiyaso T (2016) Purification, characterization, and molecular cloning of the xylanase from Streptomyces thermovulgaris TISTR1948 and its application to xylooligosaccharide production. J Mol catal B: Enzym 129:61–68
Bouacem K et al (2014) Partial characterization of xylanase produced by Caldicoprobacter algeriensis, a new thermophilic anaerobic bacterium isolated from an Algerian hot spring. Appl Biochem Biotechnol 174:1969–1981
Boucherba N, Gagaoua M, Copinet E, Bettache A, Duchiron F, Benallaoua S (2014) Purification and characterization of the xylanase produced by Jonesia denitrificans BN-13. Appl Biochem Biotechnol 172:2694–2705
Bradner J, Gillings M, Nevalainen K (1999) Qualitative assessment of hydrolytic activities in Antarctic microfungi grown at different temperatures on solid media. World J Microbiol Biotechnol 15:131–132
Butt MS, Tahir-Nadeem M, Ahmad Z, Sultan MT (2008) Xylanases and their applications in baking industry. Food Technol Biotechnol 46:22–31
Chandra R, Singh R (2012) Decolourisation and detoxification of rayon grade pulp paper mill effluent by mixed bacterial culture isolated from pulp paper mill effluent polluted site. Biochem Eng J 61:49–58
Chapla D, Patel H, Madamwar D, Shah A (2012) Assessment of a thermostable xylanase from Paenibacillus sp. ASCD2 for application in prebleaching of eucalyptus kraft pulp. Waste Biomass Valor 3:269–274
Clarke JH, Laurie JI, Gilbert HJ, Hazlewood GP (1991) Multiple xylanases of Cellulomonas fimi are encoded by distinct genes. FEMS Microbiol Lett 83:305–309
Clarkson KA, Morgan AJ, Wang ZC (1999) Xylanase from Acidothermus cellulolyticus. US Patent 5902581 A
Collins T, Gerday C, Feller G (2005) Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol Rev 29:3–23
Collins T, Hoyoux A, Dutron A, Georis J, Genot B, Dauvrin T, Arnaut F, Gerday C, Feller G (2006) Use of glycoside hydrolase family 8 xylanases in baking. J Cereal Sci 43:79–84
Collins T, Feller G, Gerday C, Meuwis MA (2012) Family 8 enzymes with xylanolytic activity. US Patent 8309336
Courtin C, Delcour JA (2002) Arabinoxylans and endoxylanases in wheat flour bread-making. J Cereal Sci 35:225–243
Dheeran P, Nandhagopal N, Kumar S, Jaiswal YK, Adhikari DK (2012) A novel thermostable xylanase of Paenibacillus macerans IIPSP3 isolated from the termite gut. J Ind Microbiol Biotechnol 39:851–860
Dhiman SS, Garg G, Sharma J, Kalia VC, Kang YC, Lee JK (2014) Reduction in acute ecotoxicity of paper mill effluent by sequential application of xylanase and laccase. PLoS One 9:1–13
Ellis JT, Magnuson TS (2012) Thermostable and alkalistable xylanases produced by the thermophilic bacterium Anoxybacillus flavithermus TWXYL3. ISRN Microbiol 2012:517524
El-Sharnouby GA, Aleid SM, Al-Otaibi MM (2012) Nutritional quality of biscuit supplemented with wheat bran and date palm fruits (Phoenix dactylifora L.). Food Nutr Sci 3:322–328
Flint HJ, McPerson CA, Martin J (1991) Expression of two xylanase genes from the rumen cellulolytic bacterium Ruminococcusflavefaciens 17 cloned in pUC13. J Gen Microbiol 137:123–129
Gallardo Ó et al (2010) Characterization of a family GH5 xylanase with activity on neutral oligosaccharides and evaluation as a pulp bleaching aid. Appl Environ Microbiol 76:6290–6294
Gartner D, Geissenddrfer M, Hillen W (1988) Expression of the Bacillus subtilis xyl operon is repressed at the level of transcription and is induced by xylose. J Bacteriol 170:3102–3109
Gibbs MD, Reeves RA, Berqquist PL (1995) Cloning, sequencing, and expression of a xylanase gene from the extreme thermophile Dictyoglomus thermophilum Rt46B.1 and activity of the enzyme on fiber-bound substrate. Appl Environ Microbiol 61:4403–4408
Gonçalves GA et al (2015) Synergistic effect and application of xylanases as accessory enzymes to enhance the hydrolysis of pretreated bagasse. Enzyme Microb Technol 72:16–24
Goswami GK, Krishnamohan M, Nain V, Aggarwal C, Ramesh B (2014) Cloning and heterologous expression of cellulose free thermostable xylanase from Bacillus brevis. SpringerPlus 1:1–6
Gröndahl M, Teleman A, Gatenholm P (2003) Effect of acetylation on the material properties of glucuronoxylan from aspen wood. Carbohydr Polym 52:359–366
Guha S, Bhutty S, Khurana SP, Kohli UK (2013) Optimization of cultural conditions for production of thermo-alkali tolerant xylanase from Bacillus sp. Int J Res Pure Appl Microbiol 3:116–120
Guo B, Chen XL, Sun CL, Zhou BC, Zhang YZ (2009) Gene cloning, expression and characterization of a new cold-active and salt-tolerant endo-β-1,4-xylanase from marine Glaciecola mesophila KMM 241. Appl Microbiol Biotechnol 84:1107–1115
Gupta S, Kuhad RC, Bhushan B, Hoondal GS (2001) Improved xylanase production from a haloalkalophilic Staphylococcus sp. SG-13 using inexpensive agricultural residues. World J Microbiol Biotechnol 17:5–8
Gupta V, Garg S, Capalash N, Gupta N, Sharma P (2015) Production of thermo-alkali-stable laccase and xylanases by co-culturing of Bacillus sp. and B. halodurans for biobleaching of kraft pulp and deinking of waste paper. Bioprocess Biosyst Eng 38:947–956
Harris AD, Ramalingam C (2010) Xylanases and its application in food industry: a review. J Exp Sci 1:1–11
Hastrup S (1988) Analysis of the B. subtilis xylose regulon. In: Cranesan AT, Hock JA (eds) Genetics and biotechnology of bacilli, vol 2. Academic Press Inc, NewYork, pp 79–84
Hong P-Y, Iakiviak M, Dodd D, Zhang M, Mackie RI, Cann I (2014) Two new xylanases with different substrate specificities from the human gut bacterium Bacteroides intestinalis DSM 17393. Appl Environ Microbiol 80:2084–2093
Huang J, Wang G, Xiao L (2006) Cloning, sequencing and expression of the xylanase gene from a Bacillus subtilis strain B10 in Escherichia coli. Biores Technol 97:802–808
Huang X, Lin J, Ye X, Wang G (2015) Molecular characterization of a thermophilic and salt-and alkaline-tolerant xylanase from Planococcus sp. SL4, a strain isolated from the sediment of a soda lake. J Microbiol Biotechnol 25:662–671
Humphry DR, George A, Black GW, Cummings SP (2001) Flavobacterium frigidarium sp. nov., an aerobic, psychrophilic, xylanolytic and laminarinolytic bacterium from Antarctica. Int J Syst Evol Microbiol 51:1235–1243
Irfan M, Nadeem M, Syed Q, Baig S (2012) Effect of medium composition on xylanase production by Bacillus subtilis using various agricultural wastes. American-Eurasian J Agric Environ Sci 12:561–565
Jain I, Kumar V, Satyanarayana T (2015) Xylooligosaccharides: an economical prebiotic from agroresidues and their health benefits. Ind J Exp Biol 53:131–142
Jeffries TW, Patel RN, Sykes MS, Klungness J (1992) Enzymatic solutions to enhance bonding, bleaching and contaminant removal. In: MRS Proceedings, vol 266. Cambridge University Press, p 277
Kamble RD, Jadhav AR (2012) Isolation, purification, and characterization of xylanase produced by a new species of Bacillus in solid state fermentation. Int J Microbiol 2012:683193
Kapoor M, Nair LM, Kuhad RC (2008) Cost-effective xylanase production from free and immobilized Bacillus pumilus strain MK001 and its application in saccharification of Prosopis juliflora. Biochem Engg J 38:88–97
Katapodis P, Kavarnou A, Kintzios S, Pistola E, Kekos D, Macris BJ, Christakopoulos P (2002) Production of acidic xylooligosaccharides by a family 10 endoxylanase from Thermoascus aurantiacus and use as plant growth regulators. Biotechnol Lett 24:1413
Kaur P, Bhardwaj NK, Sharma J (2016) Process optimization for hyper production of xylanase via statistical methodology from isolated Bacillus pumilus 3GAH using lignocellulosic waste. Biocatal Agric Biotechnol 6:159–167
Khandeparkar RDS, Bhosle NB (2006) Isolation, purification and characterization of the xylanase produced by Arthrobacter sp. MTCC 5214 when grown in solid-state fermentation. Enzyme Microb Technol 39:732–742
Kiddinamoorthy J, Anceno AJ, Haki GD, Rakshit SK (2008) Production, purification and characterization of Bacillus sp. GRE7 xylanase and its application in eucalyptus kraft pulp biobleaching. World J Microb Biotechnol 24:605–612
Kokubo I, Ikemizu S (2004) Histamine-release inhibitors containing xylooligosaccharides. Japan patent JP 2004059481:2004
Kulkarni N, Shendye A, Rao M (1999) Molecular and biotechnological aspects of xylanases. FEMS Microbiol Rev 23:411–456
Kumar V, Satyanarayana T (2013) Biochemical and thermodynamic characteristics of thermo-alkali-stable xylanase from a novel polyextremophilic Bacillus halodurans TSEV1. Extremophiles 17:797–808
Kumar V, Satyanarayana T (2014) Production of endoxylanase with enhanced thermostability by a novel polyextremophilic Bacillus halodurans TSEV1 and its applicability in waste paper deinking. Proc Biochem 49:386–394
Kumar S, Singh SP, Mishra IM, Adhikari DK (2009) Recent advances in production of bioethanol from lignocellulosic biomass. Chem Eng Technol 32:517–526
Kumar L, Kumar D, Nagar S, Gupta R, Garg N, Kuhad RC, Gupta VK (2013) Modulation of xylanase production from alkaliphilic Bacillus pumilus VLK-1 through process optimization and temperature shift operation 3. Biotech 4:345–356
Lafond M, Guais O, Maestracci M, Bonnin E, Giardina T (2014) Four GH11 xylanases from the xylanolytic fungus Talaromyces versatilis act differently on (arabino) xylans. Appl Microbiol Biotechnol 98:6339–6352
Lee HY, Shin DJ, Cho NC et al (2000) Cloning, expression and nucleotide sequences of two xylanase genes from Paenibacillus sp. Biotechnol Lett 22:387–392
Lindner C, Stülke J, Hecker M (1994) Regulation of xylanolytic enzymes in Bacillus subtilis. Microbiology 140:753–757
Liu MQ, Liu GF (2008) Expression of recombinant Bacillus licheniformis xylanase A in Pichia pastoris and xylooligosaccharides released from xylans by it. Protein Express Purif 57:101
Liu L, Cheng J, Chen H, Li X, Wang S, Song A, Wang M, Wang B, Shen J (2011) Directed evolution of a mesophilic fungal xylanase by fusion of a thermophilic bacterial carbohydrate-binding module. Process Biochem 46:395–398
Luo L, Cai J, Wang C, Lin J, Du X, Zhou A, Xiang M (2015) Purification and characterization of an alkaliphilic endo-xylanase from Streptomyces althioticus LMZM and utilization in the pulp paper industry. J Chem Technol Biot 91:1093–1098
Maity C, Ghosh K, Halder SK, Jana A, Adak A, Mohapatra PKD, Pati BR, Mondal KC (2012) Xylanase Isozymes from the Newly Isolated Bacillus sp.CKBx1D and optimization of its deinking potentiality. Appl Biochem Biotechnol 167:1208–1219
Mathur N, Goswami GK, Pathak AN (2015) In silico study of Bacillus brevis xylanase—structure prediction and comparative analysis with other bacterial and fungal xylanase. Biomed Data Min 4:112. doi:10.4172/2090-4924.1000112
Mittal A, Nagar S, Gupta VK (2012) Production and purification of high levels of cellulase-free bacterial xylanase by Bacillus sp. SV-34S using agro-residue. Ann Microbiol 63:1157–1167
Mohana A, Shah A, Divecha J, Madamwar D (2008) Xylanase production by Burkholderia sp. DMAX strain under solid state fermentation using distillery spent wash. Biores Technol 99:7553–7564
Murugan S, Arnold D, Pongiya U, Narayanan PM (2011) Production of xylanase from Arthrobacter sp. MTCC 6915 using saw dust as substrate under solid state fermentation. Enzyme Res. doi:10.4061/2011/696942
Nagar S, Gupta VK, Kumar D, Kumar L, Kuhad RC (2010) Production and optimization of cellulase-free, alkali-stable xylanase by Bacillus pumilus SV-85S in submerged fermentation. J Ind Microbiol Biotechnol 37:71–83
Nagar S, Mittal A, Kumar D, Gupta VK (2012) Production of alkali tolerant cellulase free xylanase in high levels by Bacillus pumilus SV-205. Int J Biol Macromol 50:414–420
Paice MG, Bernier R, Jurasek L (1988) Viscosity-enhancing bleaching of hardwood kraft pulp with xylanase from a cloned gene. Biotechnol Bioeng 32:235–239
Park D-J, Lee Y-S, Chang J, Fang S-J, Choi Y-L (2013) An β-1,4-xylanase with exo-enzyme activity produced by Paenibacillus xylanilyticus KJ-03 and its cloning and characterization. J Microbiol Biotechnol 23:397–404
Pathania S, Sharma N, Verma SK (2012) Optimization of cellulase-free xylanase produced by a potential thermoalkalophilic Paenibacillus sp. N1 isolated from hot springs of Northern Himalayas in India. J Microbiol Biotechnol Food Sci 2:1–24
Polizeli M, Rizzatti A, Monti R, Terenzi H, Jorge JA, Amorim D (2005) Xylanases from fungi: properties and industrial applications. Appl Microbiol Biotechnol 67:577–591
Pollet A, Delcour JA, Courtin CM (2010) Structural determinants of the substrate specificities of xylanases from different glycoside hydrolase families. Crit Rev Biotechnol 30:176–191
Porsuk I, Özakin S, Bali B, Yilmaz EI (2013) A cellulase-free, thermoactive, and alkali xylanase production by terrestrial Streptomyces sp. CA24. Turk J Biol 37:370–375
Poosarla VG, Chandra TS (2014) Purification and characterization of novel halo-acid-alkali-thermo-stable xylanase from Gracilibacillus sp. TSCPVG. Appl Biochem Biotechnol 173:1375–1390
Prakash D, Nawani N, Prakash M, Bodas M, Mandal A, Khetmalas M, Kapadnis B (2013) Actinomycetes: a repertory of green catalysts with a potential revenue resource. BioMed Res Int 2013:264020
Raj A, Kumar S, Singh SK (2013) A highly thermostable xylanase from Stenotrophomonas maltophilia: purification and partial characterization. Enzyme Res 2013:429305. doi:10.1155/2013/429305
Rhee MS, Wei L, Sawhney N, Rice JD, John FJS, Hurlbert JC, Preston JF (2014) Engineering the xylan utilization system in Bacillus subtilis for production of acidic xylooligosaccharides. Appl Environ Microbiol 80:917–927
Roy S, Dutta T, Sarkar TS, Ghosh S (2013) Novel xylanases from Simplicillium oclavatum MTCC 9604: comparative analysis of production, purification and characterization of enzyme from submerged and solid state fermentation. SpringerPlus 2:382
Sanghi A, Garg N, Sharma J, Kuhar K, Kuhad RC, Gupta VK (2008) Optimization of xylanase production using inexpensive agro-residues by alkalophilic Bacillus subtilis ASH in solid-state fermentation. World J Microb Biotechnol 24:633–640
Sapag A, Wouters J, Lambert C, De Ioannes P, Eyzaguirre J, Depiereux E (2002) The endoxylanases from family 11: computer analysis of protein sequences reveals important structural and phylogenetic relationships. J Biotechnol 95:109–131
Sepahy AA, Ghazi S, Sepahy MA (2011) Cost-effective production and optimization of alkaline xylanase by indigenous Bacillus mojavansis AG137 fermented on agricultural waste. Enzyme Res 2011:593624
Shang SM, Qian L, Zhang X, Li KZ, Chagan I (2013) Themoanaerobacterium calidifontis sp. nov., a novel anaerobic, thermophilic, ethanol-producing bacterium from hot springs in China. Arch Microbiol 195:439–445
Sharma M, Chadha BS (2011) Production of hemicellulolytic enzymes for hydrolysis of lignocellulosic biomass. In: Pandey A, Larroche C, Ricke SC, Dussap C, Gnanasonou E (eds) Biofuels, alternative feed stocks and conversion processes, 1st edn. Academic, USA, pp 214–217
Shi H, Zhang Y, Zhang H, Huang Y, Li X, Wang F (2014) Cloning, over-expression and characterization of a thermo- tolerant xylanase from Thermotoga thermarum. Biotechnol Lett 36:587–593
Subramaniyan S (2012) Isolation, purification and characterisation of low molecular weight xylanase from Bacillus pumilus SSP-34. Appl Biochem Biotechnol 166:1831–1842
Subramanian S, Prema P (2000) Cellulase-free xylanases from Bacillus and other microorganisms. FEMS Microbiol Lett 183:1–7
Subramanian S, Prema P (2002) Biotechnology of microbial xylanases: enzymology, molecular biology and application. Crit Rev Biotechnol 22:33–46
Sun YC, Wen JL, Xu F, Sun RC (2011) Structural and thermal characterization of hemicelluloses isolated by organic solvents and alkaline solutions from Tamarix astromongolica. Biores Technol 102:5947–5951
Taibi Z, Saoudi B, Boudelaa M, Trigui H, Belghith H, Gargouri A, Ladjama A (2012) Purification and biochemical characterization of a highly thermostable xylanase from Actinomadura sp. strain Cpt20 isolated from poultry compost. Appl Biochem Biotechnol 166:663–679
Teplitsky A, Shulami S, Moryles S, Shoham Y, Shoham G (2000) Crystallization and preliminary X-ray analysis of an intracellular xylanase from Bacillus stearothermophilus T-6. Act Cryst Sect D: Biol Crystallogr 56:181–184
Thomas L, Ushasree MV, Pandey A (2014) An alkali-thermostable xylanase from Bacillus pumilus functionally expressed in Kluyveromyces lactis and evaluation of its deinking efficiency. Biores Technol 165:309–313
Verma D, Satyanarayana T (2012) Cloning, expression and applicability of thermo-alkali-stable xylanase of Geobacillus thermoleovorans in generating xylooligosaccharides from agro-residues. Biores Technol 107:333–338
Verma D, Anand A, Satyanarayana T (2013a) Thermostable and alkalistable endoxylanase of the extremely thermophilic bacterium Geobacillus thermodenitrificans TSAA1: cloning, expression, characteristics and its applicability in generating xylooligosaccharides and fermentable sugars. Appl Biochem Biotechnol 170:119–130
Verma D, Kawarabayasi K, Miyazaki K, Satyanarayana T (2013b) Cloning, expression and characteristics of a novel alkali-stable and thermostable xylanase encoding gene (Mxyl) retrieved from compost-soil metagenome. PLoS One 8:1–8
Viikari L, Ranua M, Kantelinen A, Sundquist J, Linko M (1986) Bleaching with enzymes. In: Proceedings of 3rd international conference on biotechnology in the pulp and paper industry. STFI, Stockholm, pp 67–69
Wang SY, Hu W, Lin XY, Wu ZH, Li YZ (2012) A novel cold-active xylanase from the cellulolytic myxobacterium Sorangium cellulosum So9733-1: gene cloning, expression, and enzymatic characterization. Appl Microbiol Biotechnol 93:1503–1512
Wang G, Huang X, Ng TB, Lin J, Ye XY (2014) High phylogenetic diversity of glycosyl hydrolase Family 10 and 11 Xylanases in the sediment of lake Dabusu in China. PLoS One 9:e112798
Wong K, Tan L, Saddler JN (1988) Multiplicity of beta-1,4-xylanase in microorganisms: functions and applications. Microbiol Rev 52:305
Wu YB, Ravindran V, Thomas DG, Birtles MJ, Hendriks WH (2004) Influence of phytase and xylanase, individually or in combination, on performance, apparent metabolisable energy, digestive tract measurements and gut morphology in broilers fed wheat-based diets containing adequate level of phosphorus. Br Poul Sci 45:76–84
Xin F, He J (2013) Characterization of a thermostable xylanase from a newly isolated Kluyvera species and its application for biobutanol production. Biores Technol 135:309–315
Zheng H, Ming-zhe S, Ling-cai M, Hai-sheng P, Xiu-qing Z, Zheng Y, Wen-hui Zeng, Zhang J, Hu J, Lu F, Sun J (2014) Purification and characterization of a thermostable xylanase from Paenibacillus sp. NF1 and its application in xylooligosaccharides production. J Microbiol Biotechnol 24:489–496
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Chakdar, H., Kumar, M., Pandiyan, K. et al. Bacterial xylanases: biology to biotechnology. 3 Biotech 6, 150 (2016). https://doi.org/10.1007/s13205-016-0457-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-016-0457-z