
Abstract
The blood–brain barrier (BBB) is an essential component of the neurovascular unit that controls the exchanges of various biological substances between the blood and the brain. BBB damage is a common feature of different central nervous systems (CNS) disorders and plays a vital role in the pathogenesis of the diseases. Non-coding RNAs (ncRNAs), such as microRNAs (miRNAs), long non-coding RNA (lncRNAs), and circular RNAs (circRNAs), are important regulatory RNA molecules that are involved in almost all cellular processes in normal development and various diseases, including CNS diseases. Cumulative evidences have demonstrated ncRNA regulation of BBB functions in different CNS diseases. In this review, we have summarized the miRNAs, lncRNAs, and circRNAs that can be served as diagnostic and prognostic biomarkers for BBB injuries, and demonstrated the involvement and underlying mechanisms of ncRNAs in modulating BBB structure and function in various CNS diseases, including ischemic stroke, hemorrhagic stroke, traumatic brain injury (TBI), spinal cord injury (SCI), multiple sclerosis (MS), Alzheimer's disease (AD), vascular cognitive impairment and dementia (VCID), brain tumors, brain infections, diabetes, sepsis-associated encephalopathy (SAE), and others. We have also discussed the pharmaceutical drugs that can regulate BBB functions via ncRNAs-related signaling cascades in CNS disorders, along with the challenges, perspective, and therapeutic potential of ncRNA regulation of BBB functions in CNS diseases.
Similar content being viewed by others
Overview of non-coding RNAs
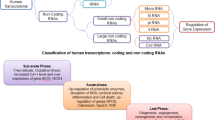
Non-coding RNAs (ncRNAs) refer to the RNA molecules that do not have the protein-coding potential, which account for the majority of RNAs and make up about 98–99% of all mammalian genomes generated RNAs [1, 2]. Although non-coding RNAs do not possess the ability to translate to proteins, increasing evidence demonstrated that they actively interact with nucleic acids or other molecules to function as vital regulatory molecules on almost all cellular processes in normal development and various diseases, including the central nervous system (CNS) disorders [1, 3,4,5]. In addition, one ncRNA is able to interact with one or more target molecules in different cellular signaling pathways, making the ncRNA-mediated regulation network even more complicated. For endogenous ncRNAs, besides structural ncRNAs such as ribosomal RNA (rRNAs) and transfer RNAs (tRNAs), ncRNAs can be broadly divided into small ncRNAs (<200 nucleotides) and long ncRNAs (>200 nucleotides). Small ncRNAs can be even subdivided into microRNAs [6], small interfering siRNA (siRNAs) [7], piwi-interacting RNAs (piRNAs) [8], small nuclear RNAs (snRNAs) [9], small nucleolar RNAs (snoRNAs), [10] and others [11]. Long ncRNAs include long intronic ncRNAs, antisense RNAs (asRNAs), promoter-associated long RNAs (PALRs), promoter upstream transcripts (PROMPTs), stable excised intron RNAs, and long stress-induced non-coding transcripts (LSINCTs) [12]. Numerous ncRNAs exhibit regulatory functions on the blood–brain barrier (BBB) during CNS diseases, which mainly include microRNAs, long non-coding RNAs, and circular RNAs.
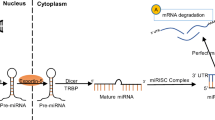
MicroRNAs (miRNAs, miRs) are a class of single-stranded non-coding RNAs with an average length of 22 nucleotides [13]. They can inhibit protein expression mainly by complementary binding to the messenger RNAs (mRNA) of target genes at the 3’ untranslated region (3’ UTR) to induce mRNA degradation or translational repression [6]. miRNAs are very stable, can be secreted to various biofluids by different types of cells, and be transported by Argonaute protein or exosomes, which can protect their degradation from RNases, suggesting them as potential biomarkers during disease biogenesis [5,6,7]. Also, miRNAs have been shown to be able to target hundreds of target genes by each of them and can modulate more than one-third of all human genes [5, 14, 15], suggesting their vast involvement and complex regulatory functions.
Long non-coding RNA (lncRNAs) are defined as a heterogeneous class of mRNA-like transcripts that are longer than 200 nucleotides but without protein-coding ability [3, 16]. Similar to mRNAs, their transcription is also involved in RNA Polymerase II (RNAPII), and most of the lncRNAs are polyadenylated [12]. In general, lncRNAs lack the open reading frame, show tissue-specific expression profile, and exhibit low levels compared to the protein-coding counterparts [3, 12]. The structure features of the lncRNAs play a key role in modulating the affinity of DNA, RNA, and proteins, in the assembly and regulation of multi-molecular complexes [3]. It has been shown that lncRNAs can regulate gene expression through different mechanisms, including modulation of gene promotor or transcription factor activity and splicing machinery, recruitment of transcriptional activator, increasing mRNAs stability or inducing their decay, acting as architectural components in the assembly of protein complexes, or served as competing endogenous RNAs (ceRNAs) to regulate the functions of miRNAs [3, 16, 17].
Unlike linear ncRNAs (e.g., microRNAs or lncRNAs), circular RNA (circRNA) is a class of covalently closed single-stranded long ncRNA that the 5’ and 3’ termini are connected by back-splicing circularization of exons of pre-mRNAs and forms a continuous loop [18, 19]. Due to the unique circular structure, circRNAs are more resistant to be degraded by RNases, thereby more stable than linear ncRNAs, and can be served as potential biomarkers in some diseases. Also, it has been suggested that circRNAs exhibit tissue-specific and cell-specific expression properties [19]. The growing interest in this relatively new type of ncRNAs gradually reveals the biological functions of circRNAs, including miRNA sponges, protein sponges or decoys, enhancer of protein functions, protein scaffolding and recruitment, and serving as templates for translation under certain circumstances [19, 20].
Overview of the blood–brain barrier and neurovascular unit in the central nervous system
The blood–brain barrier (BBB) plays a vital role in controlling the influx and efflux of various biological substances between the brain and the blood to maintain the metabolic activity and function of the brain [21]. Serval excellent articles have systematically reviewed the structure, cellular components, and biological functions of the BBB [21,22,23]. At the cellular level, BBB is formed by brain microvascular endothelial cells (BMECs), astrocytes end-feet and pericytes, and cell-cell adhesion molecules (mainly tight junctions, TJs) tightly seal a monolayer of brain endothelial cells in the capillary microvasculature to form a highly selective diffusion barrier. The diffusion barrier can selectively prevent the passive exchange of solutes and neurotoxic molecules, regulate the trafficking of macromolecules, ions, amino acids, peptides, and signaling molecules, and the entry of leukocytes [24,25,26,27] between the blood and the brain [23, 28].
The junction complex of BBB consists of tight junctions (TJs) and adherens junctions (AJs) [21]. The TJs mainly comprised of three types of integral membrane proteins, including claudins (e.g., claudin-1, claudin-2, claudin-3, claudin-5, claudin-11, etc.), occludin, junction adhesion molecules (JAMs) (e.g., JAM-A, JAM-B, JAM-C, JAM-4, JAM-5, etc.), and cytoplasmic accessory proteins (e.g., zonula occludens-1 (ZO-1), ZO-2, ZO-3, cingulin, etc.), which form the cytoplasmic bridge connecting the TJs to the cell cytoskeleton [22, 23]. At the BBB, it is broadly consented that claudin-5 is the most abundant and dominant claudin in endothelial cells, which plays a key role in maintaining BBB integrity, and its dysfunction has been associated with various neurological disorders [29,30,31,32]. The AJs mainly consist of cadherins, which join the actin cytoskeleton to form adhesive contacts between cells through intermediary submembrane proteins, catenins (e.g., β-catenin, γ-catenin) [21,22,23]. Beta-catenin and γ-catenin connect the cytoplasmic domains of cadherins to the cell cytoskeleton via α-catenin [21,22,23].
Pericytes are also crucial constituents of the BBB and brain capillary, and they are abundantly expressed in the CNS. They share a basement membrane with endothelial cells and contact with endothelium through N-cadherin and connexins [21, 33]. Pericytes play a crucial role in maintaining BBB integrity, facilitating angiogenesis, and stabilizing the structure of microvasculature [21, 23, 33, 34]. Astrocytes are also essential components of BBB and may play a decisive role in the induction of BBB characteristics and maintaining the barrier function of brain endothelial cells. Astrocytic end-feet has also been suggested as crucial checkpoints of brain metabolism [21, 23, 35]. The neurovascular unit consists of all the major cellular components of the brain, including brain endothelial cells, vascular smooth muscle cells (VSMC), pericytes, astrocytes, neurons, microglia, and perivascular cells [36]. The communication between the cells of the neurovascular unit is vital and responsible for regulating blood flow and controlling the exchange of substances across the BBB [37].
Non-coding RNAs as diagnostic and prognostic biomarkers for BBB damage in the CNS disorders
Emerging evidence has suggested the diagnostic and therapeutic values of circulating non-coding RNAs in human diseases, and some excellent articles have reviewed microRNAs, long non-coding RNAs, or circular RNAs as promising non-invasive biomarkers for numerous CNS disorders. For example, microRNAs can be served as diagnostic and prognostic biomarkers of ischemic stroke [5, 38], hemorrhagic stroke [39], traumatic brain injury (TBI) [40], spinal cord injury (SCI) [41], glioma [42], multiple sclerosis (MS) [43], Alzheimer’s disease (AD) [44], vascular cognitive impairment and dementia (VCID) [45], and others. LncRNAs have also been suggested as diagnostic and therapeutic biomarkers for stroke [46], TBI [47], SCI [48], glioma [49], MS [50], AD [51], and other diseases. In addition, several circular RNAs (e.g., circ_101222) have been suggested as potential biomarkers for neurological diseases [52]. Although these articles summarized the general potential of non-coding RNAs as diagnostic and prognostic biomarkers for CNS diseases, their regulation on BBB was not specified. Thus, our current paper reviews the non-coding RNAs able to regulate BBB functions and possess the potential to be served as promising biomarkers of CNS diseases.
microRNAs
Several microRNAs have been suggested as potential biomarkers for BBB disruption in ischemic stroke, hemorrhagic stroke, TBI, SCI, MS, breast cancer brain metastasis, AD and sepsis-associated encephalopathy (SAE). Wang et al. described that elevated miR-29b levels in white blood cells related to impaired BBB functions, which also negatively associated with NIHSS (National Institute of Health stroke scale) scores and brain infarct volume in ischemic stroke, and could potentially predict stroke outcomes as a novel circulating biomarker [53]. Zhang et al. showed that the significantly increased plasma miR-503 positively correlated with plasma von Willebrand Factor (vWF, an indicator of EC dysfunction) in ischemic stroke patients [54]. miR-503 was also an independent risk factor for ischemic stroke by logistic analysis [54]. Additionally, miR-503 was able to regulate BBB damage, brain edema, and cerebral circulation by modulating endothelial monolayer permeability, oxidative stress, and cell apoptosis, suggesting miR-503 as a promising biomarker for ischemic stroke with BBB regulation functions [54]. miR-130a was significantly elevated in the serum of intracerebral hemorrhage (ICH) patients and rat ICH models, associated with clinical outcomes in patients with deep hematoma, and was an independent indicator positively associated with perihematomal edema (PHE) volume within the first three days after the onset of ICH in patients [55]. Besides, brain edema, BBB permeability, and neurological deficit scores can be significantly alleviated by miR-130a inhibitors, making miR-130a a useful early biomarker for monitoring post-ICH PHE and predicting disease prognosis [55]. miR-21 has been proposed by several groups to be a promising biomarker for both TBI patients and preclinical animal models, and its upregulation upon TBI inhibits endothelial apoptosis, enhances junctional proteins expression, and targets angiogenic factors critical to BBB maintenance [40]. miR-155 has been suggested as a potential biomarker in spinal cord ischemic injury, as miR-155 exhibited upregulated expression in neurons and endothelial cells of spinal cord ischemic injury mice, but showed lower miR-155 expression in mice that escaped paralysis [56]. miR-155 deletion also reduced central cord edema and improved blood-spinal cord barrier (BSCB) integrity with reduced incidence of spinal cord paralysis [56]. miR-155 has also been suggested to serve as potential prognostic biomarkers for relapsing-remitting multiple sclerosis (RRMS) [57]. Different from SCI, miR-155 exhibited reduced expression in blood samples of RRMS patients. More interestingly, downregulated miR-155 significantly correlated with patients’ expanded disability status scale (EDSS) and upregulated surface receptors and cytotoxic proteins that were crucial for migration through the BBB, such as intracellular adhesion molecule 1 (ICAM-1) and integrin subunit beta 2 (ITGB-2) in CD8+ T cells [57]. In breast cancer brain metastasis, miR-105 is specifically expressed and secreted by the breast cancer cell. Cancer-secreted miR-105 is also significantly associated with metastatic progression in breast cancer patients by downregulating tight junction protein zonula occludens 1 (ZO-1), destroying the endothelial monolayer barrier function, and inducing vascular permeability. These results suggested miR-105 as an early diagnostic blood-borne marker as well as a therapeutic target for breast cancer metastasis [58]. In AD, miR-181a might be a biomarker for the progression of the disease, as miR-181a declined synchronously with the accumulation of beta-amyloid (Aβ) in APP/PS1 mouse, suggesting a reverse correlation between miR-181a levels and AD development [59]. The decline of miR-181a is also correlated with Aβ accumulation-induced pericyte apoptosis and BBB breakdown in APP/PS1 mice [59]. Furthermore, plasma miR-370-3p has been suggested as an attractive biomarker candidate in SAE [60]. Sepsis-induced BBB breakdown is considered a significant cause of SAE. Increased miR-370-3p expression was specially observed in the brain but not in other organs in SAE mouse models [60]. Plasma miR-370-3p was also specifically increased and highly sensitive for early detection in SAE in patients or animal models with BBB permeability defect, neuroinflammation, and brain apoptosis, suggesting plasma miR-370-3p a unique biomarker for SAE [60].
Long non-coding RNAs
In ischemic stroke, lncRNA XIST has been proposed to be a potential biomarker for predicting the prognosis of acute cerebral ischemia and a therapeutic target for stroke patients. LncRNA XIST exhibited increased expression during the late stages (seven days later) after the onset of ischemic stroke, and the serum levels of lncRNA XIST were significantly negatively correlated with the severity of neurological impairments in the ischemic stroke patients [61]. Silencing of lncRNA XIST significantly impaired angiogenesis by decreasing the endothelial migration and tube formation, exacerbated cerebral vascular injury by markedly reducing the expressions of KLF4 and tight junction proteins claudin-5 and ZO-1, and evidently increased the vascular permeability by upregulating the expression of E-selectin, vascular cell adhesion protein 1 (VCAM-1), ICAM-1, and p-NF-kB [61]. Another lncRNA, named lncRNA associated with breast cancer brain metastasis (BCBM) (Lnc-BM), has also been suggested as a prognostic biomarker of the progression of brain metastasis in breast cancer patients [62]. Lnc-BM is overexpressed in breast cancer tissue and is upregulated specifically in brain metastatic cells, but not in lung or bone metastatic cells [62]. High Lnc-BM expression was negatively correlated with recurrence-free survival in breast cancer patients, and high Lnc-BM expression in the primary tumor predicts an increased risk for brain metastasis [62]. In addition, elevated Lnc-BM expression promotes BCBM, while depletion of Lnc-BM effectively suppresses BCBM. Moreover, Lnc-BM enhances the STAT3 (signal transducer and activator of transcription 3)-dependent expression of ICAM1 and CCL2 (C-C chemokine ligand 2, monocyte chemoattractant protein-1, MCP-1), which mediated vascular invasion and recruitment of macrophages in the brain, respectively, to increase the cancer cell migration through BBB and to exacerbate BCBM [62].
Circular RNAs
Circular RNAs are abundantly expressed in eukaryotes, spatiotemporal specific, highly conserved, and less susceptible to degradation by RNA exonucleases; thus, they have recently been proposed to be novel biomarkers for CNS disorders, including ischemic stroke. However, there are relatively fewer publications of this type of non-coding RNAs that possess the ability to regulate BBB functions in different CNS disorders and serve as biomarkers. One circular RNA, named circRNA DLGAP4 (circDLGAP4), has recently been suggested as a promising biomarker for diagnosing and evaluating the degree of cerebral damage caused by acute ischemic insults [63]. CircDLGAP4 levels were significantly decreased in the plasma of acute ischemic stroke patients and a mouse model of ischemic stroke, and upregulation of circDLGAP4 expression significantly inhibited miR-143 activity and resulting in improved neurological deficits, decreased infarct areas, and attenuated blood–brain barrier extravasation along with ameliorated the downregulation of tight junction proteins, including claudin-5, occludin, and ZO-1 in the preclinical ischemic stroke models [63]. These findings suggested cirDLGAP4 in the plasma as a novel biomarker and therapeutic target in acute cerebrovascular protection after ischemic stroke [63].
Non-coding RNAs regulate BBB/BSCB functions in CNS disorders
Accumulating evidence has demonstrated the extensive involvement (Fig. 1) and regulatory mechanisms of different non-coding RNAs (microRNAs (Table 1), lncRNAs (Table 2), and circular RNAs (Table 3) in BBB/BSCB injuries and repairs in various CNS disorders. Herein, we summarized functional significance and molecular mechanisms of different non-coding RNAs in CNS disorders, including ischemic stroke and hemorrhagic stroke, TBI, SCI, brain tumors (glioma, glioblastoma, and brain metastasis), MS, dementia, brain infections, diabetes, SAE and others.

Altered expression profiles of BBB regulatory miRNAs, lncRNAs, and circRNAs in CNS disorders. During CNS disorders, such as ischemic stroke, hemorrhagic stroke, traumatic brain injury (TBI), spinal cord injury (SCI), glioma and brain metastasis, multiple sclerosis (MS), Alzheimer's disease (AD), vascular cognitive impairment and dementia (VCID), brain infections, diabetes, sepsis-associated encephalopathy (SAE), and others, the structure and the function of BBB has been compromised along with dysregulated non-coding RNA expressions. Modulating the expressional levels of miRNAs, lncRNAs, and circRNAs is able to effectively regulate the BBB damage and recovery during these CNS disorders. “↑”, upregulated non-coding RNA levels, and “↓” downregulated non-coding RNA levels in CNS disorders. RCVS reversible cerebral vasoconstriction syndrome; TSC tuberous sclerosis complex
Ischemic stroke
Our group is among the first to investigate the regulatory role of microRNA in cerebrovascular endothelial injury and BBB dysfunction after ischemic stroke. We demonstrated that downregulation of ischemia-induced miR-15a expression in the brain can alleviate apoptotic cell death in cerebral microvessels, and reduce BBB disruption and cerebral infarction in mice transient focal cerebral ischemia [64]. Recently, by using an endothelial cell (EC)-selective miR-15a/16-1 conditional knockout mouse model, we show that endothelial-targeted deletion of the miR-15a/16-1 cluster ameliorated BBB leakage and infiltration of peripheral immune cells after experimental ischemic stroke [65], endothelial miR-15a/16-1 cluster is also demonstrated to be a negative regulator for cerebral angiogenesis and long-term neurological recovery following ischemic stroke [66]. Other miRNAs (Fig. 1) that are involved and exhibit regulatory functions on BBB permeability or integrity after ischemic stroke including miR-29b [53], miR-34a [67,68,69], miR-150 [70], miR-122 [71], miR-210 [72], miR-21 [73], miR-539 [74], miR-155 [75, 76], miR-149-5p [77], miR-1 [78], miR-132 [79], let-7g*[80, 81], miR-98 [80], miR-126-3p/5p [82], miR-98 [83], miR-668 [84], miR-182 [85], miR-503 [54], let-7f [86], miR-30a [87], miR-376b-5p [88], miR-149-5p [89], miR-92b [90], and miR-141-3p [91].
Among these miRNAs, miR-34a, miR-210, miR-155, miR-212/312, miR-15a/16-1, miR-1, miR-182, miR-503, miR-30a, and miR-141-3p exhibit upregulated expression profile under cerebral ischemic or hypoxic conditions, and generally associate with deteriorated neurological neurobehaviors, large infarct volume, and brain edema, and BBB disruption. Downregulation of these enhanced miRNAs through antagomir or miRNA inhibitors significantly alleviated BBB permeability and infiltration of peripheral immune cells, reduced brain infarction and edema, and improved neurological functions after ischemic stroke. For example, miR-34a expression was significantly elevated in the extracted brain microvascular endothelial cells from ischemic mouse brains at the time point of BBB opening following 1-hour MCAO and reperfusion [69], and overexpression of miR-34a significantly enhanced the BBB permeability by disruption of tight junction protein ZO-1 in vitro [67, 68]. Genetic deletion of miR-34a effectively reduced the BBB leakage, alleviated the disruption of tight junction proteins ZO-1, claudin-5, and occludin, and improved neurological recovery following ischemic stroke [69]. Wang et al. demonstrated that miR-30a was significantly increased under ischemic conditions, and inhibition of miR-30a levels with inhibitor decreased the BBB permeability by preventing the degradation of tight junction proteins occludin and claudin-5, and reduction of zinc transporter ZnT4 in both brain endothelial cells and isolated cerebral microvessels of ischemic mice, which yielded ameliorated infarct volume and improved neurological deficits after cerebral ischemia in mice [87].
On the other hand, miR-29b, miR-122, miR-539, miR-149-5p, miR-132, let-7g/7g*, miR-98, miR-126-3p/5p, let-7f, miR-376b-5p, miR-92b exhibited downregulated expression profile under cerebral ischemic conditions. Restoring their expression level through miR mimics or agomirs can significantly downregulate the BBB permeability by enhancing the tight junction protein expression and pericytes coverage, decreasing pro-inflammatory cytokines, matrix metalloproteinases (MMPs), and apoptotic cell death, etc., to reduce the cerebral infarction, brain edema, and improve the overall neurological outcomes following ischemic stroke. For instance, Pan et al. demonstrated decreased expression of miR-126-3p and miR-126-5p in ischemic mouse brains [82]. Lentiviral mediated overexpression in ischemic brains significantly attenuated the decrease of tight junction proteins ZO-1 and occludin, and reduced IgG leakage to the brain tissue three days after stroke [82]. Overexpression of miR-126-3p and -5p also downregulated the expression of pro-inflammatory cytokines interleukin (IL)-1β and tumor necrosis factor (TNF)-α, accompanied by reduced protein level of cell adhesion molecules VCAM-1 and E-selectin three days after stroke [82]. These effects conferred reduced brain infarction, edema volume, and improved behavioral outcomes following ischemic stroke [82].
In addition, one miRNA was found downregulated and another one was upregulated in ischemic stroke, but the protective effects were achieved by strengthening this dysregulation. miR-150 expression was significantly suppressed under hypoxia or ischemic stroke [92], upregulation of miR-150 exhibited worse BBB permeability and decreased claudin-5 expression in both in vitro and in vivo ischemic stroke models [70]. On the contrary, downregulation of miR-150 expression contributed to BBB protection, infarct volume reduction, and neurological deficit amelioration by regulating the angiopoietin receptor Tie-2 [70]. miR-21 was found significantly elevated in the serum of ischemic stroke patients [93, 94], and upregulation of miR-21 expression by mimics significantly reduced ischemic stroke-induced infarct volume, edema, BBB disruption by decreasing the levels of p38, mitogen-activated protein kinase kinase 3 (MAP2k3), inducible nitric oxide synthase (iNOS), and MMP-9 [73].
Recent studies also demonstrated the functional roles of lncRNAs in the regulation of the BBB permeability and neurological recovery in ischemic stroke. Among these differentially expressed lncRNAs in ischemic stroke, lncRNA metastasis-associated lung adenocarcinoma transcript 1 (Malat1) [95,96,97], LOC102640519 [98], and maternally expressed gene 3 (MEG3) [99] exhibit increased expression, and lncRNA small nucleolar RNA host gene 8 (Snhg8) [100] and X-inactive-specific transcript (XIST) [61] exhibit decreased expression in brain microvascular endothelial cell (BMEC) cultures, astrocytes, or ischemic brain tissues under hypoxia or cerebral ischemia in mice (Fig. 1). Cerebral microvascular endothelial injuries mediate the initial process of BBB disruption in ischemic stroke. We and others show that lncRNA Malat1 is one of the most highly upregulated lncRNA and plays an essential role in the protection against cerebral microvascular endothelial pathophysiology during ischemic stroke [95, 96, 101]. Li et al. showed that upregulation of Malat1 promoted BMEC autophagy as a protective mechanism to enable endothelial survival under ischemic insults. Mechanistic studies revealed that Malat1 serves as a competing endogenous RNA by sponging miR-26b and upregulating the expression of uncoordinated 51-like kinase 2 (ULK2) [95]. Our group demonstrated that Malat1 exerts anti-apoptotic and anti-inflammatory functions in brain microvasculature to ameliorate ischemic cerebral vascular and parenchymal damages through binding to proapoptotic factor Bim and pro-inflammatory molecule E-selectin both in vitro and in vivo. Silencing Malat1 severely aggravated the injury of primary BMEC cultures, and worsened neurological scores, sensorimotor functions, and brain infarct size in the mouse model of ischemic stroke [96]. Astrocytes provide structural and nutritional supports for neurons and are important components of the BBB. Wang et al. showed that Malat1 expression was highly upregulated in astrocytes and animal models of ischemic stroke, however, they observed knockdown of Malat1, instead of overexpression, could protect against ischemia-induced injuries by reducing the cell apoptosis and increasing the cell viability [97]. Further investigation suggested that lncRNA Malat1 could positively regulate the expression of aquaporin-4 (AQP4) by competitively binding miR-145 to mediate the damage of astrocytes during ischemic stroke [97]. LncRNA XIST was found to exhibit decreased serum levels during the early stages of ischemic stroke patients, and silencing lncRNA XIST significantly decreased the endothelial migration and tube formation, and exacerbated cerebral vascular injury by notably reducing the expressions of krüppel-like factor 4 (KLF4) and tight junction proteins claudin-5 and ZO-1, leading to larger infarction and worse neurological functions in transient ischemic stroke mice [61].
Moreover, several circular RNAs have been demonstrated to exert protective function against ischemic stroke-induced BBB disruption in cellular and experimental animal models. Circular RNA DLGAP4 (circDLGAP4) has been found to significantly decrease in the plasma of ischemic stroke patients and in a mouse stroke model, and upregulation of circDLGAP4 expression significantly attenuated neurological deficits and decreased infarct areas and BBB damage, including reduced Evans blue extravasation, ameliorated the downregulation of tight junction proteins claudin-5, occludin, and ZO-1 [63]. CircDLGAP4 also acts as an endogenous miR-143 sponge via targeting miR-143 to downregulate endothelial to mesenchymal transition (EndoMT), an inflamed pathological condition of endothelial cells, to regulate BBB integrity under cerebral ischemic conditions [63]. The expression of circular RNA CCDC9 (circCCDC9) also decreased in ischemic stroke mice, overexpression of circCCDC9 exhibited BBB protection with decreased Evens blue leakage and brain water content, restored NO production and endothelial nitric oxide synthase (eNOS) expression in the ischemic brains. Also, overexpression of circCCDC9 inhibited apoptosis and the Notch pathway by repressing the modulator levels of Notch1, NICD, and Hes1 after cerebral ischemia/reperfusion injury in mice [102].
Hemorrhagic stroke
Hemorrhagic stroke can lead to severe BBB permeability, neuroinflammation, and cerebral edema. Accumulating investigations have revealed the participation of microRNAs in the regulation of BBB integrity during experimental hemorrhagic stroke models. Among these up or downregulated miRNAs in hemorrhagic stroke, miR-130a [55], miR-21-5p [103], miR-103-3p [104], and miR-24 [105], exhibited upregulated expression profiles under cerebral hemorrhagic conditions (Fig. 1), and inhibition of their expression exert reduced edema, alleviated BBB permeability, improved long-term neurobehavioral functions. For instance, miR-21-5p was found to exhibit elevated serum levels in elderly intracerebral hemorrhage (ICH) or patients, and intracerebroventricular (ICV) administration of miR-21-5p antagomir in aged ICH rats robustly alleviated BBB permeability, inhibited neuronal apoptosis and neuroinflammation, accelerated hematoma absorption, and enhanced neurological and cognitive functions [103]. Dual-specificity phosphatase 8 (DUSP8) was identified as a direct target of miR-21-5p, inhibition of miR-21-5p upregulated the expression of DUSP8, which induced the activation of the p-ERK /heme oxygenase-1 (HO-1) pathways to alleviate hemorrhagic injuries [103]. miR-103-3p also exhibits increased levels in a rat subarachnoid hemorrhage (SAH) model. Repression of miR-103-3p in rats significantly relieved caveolin-1 (Cav-1) loss, reduced BBB permeability, preserved microvascular integrity, and improved long-term neurobehavioral function after SAH [104]. Mechanistic studies demonstrated that miR-103-3p could directly target Cav-1 to decrease the protein expression of ZO-1 and occludin in rats following SAH insults [104].
On the other hand, miR-126-3p [106,107,108], miR-27a-3p [109], miR-193b-3p [110], and miR-26a-5p [111] exhibited downregulated expression (Fig. 1), restoring or upregulating their levels also yield beneficial effects on BBB integrity and cell survival, neuroinflammatory environment, brain infarction, edema, and neurological outcomes. Several groups investigated the beneficial effects of miR-126-3p against BBB disruption and associated neuronal injuries following ICH. Xi et al. showed that silencing of miR-126-3p led to impaired BMEC barrier integrity, reversed vascular endothelial growth factor (VEGF)- and angiopoietin-1 (Ang-1)-induced Akt activation and apoptosis inhibition [106]. On the other hand, overexpression of miR-126-3p suppressed the upregulation of phosphoinositide-3-kinase regulatory subunit 2 (PIK3R2) and maintained the activation of Akt in the perihematomal area, accompanied by inhibited neutrophil infiltration, microglial activation, and neuronal apoptosis [106]. Fu et al. demonstrated that miR-126-3p showed downregulated expression in serum and hemorrhagic area in ICH rats and downregulated serum expression in patients with ICH [107]. Inhibition of miR-126-3p impaired endothelial barrier permeability by upregulating VCAM-1 expression levels in rat BMECs, while overexpression of miR-126-3p downregulated VCAM-1 expression in the hemorrhagic area in rats following ICH [107]. Wang et al. investigated the efficiency of miR-126-3p-overexpressed bone marrow mesenchymal stem cells (MSCs) in the repair of BBB damage after ICH. They observed that miR-126 facilitated the differentiation of MSCs into vascular endothelial cells in ICH models both in vitro and in vivo [108]. They also confirmed that miR-126-overexpressed MSC alleviated the cell apoptosis, robustly reduced the expression of protease-activated receptor-1 (PAR-1) and MMP-9, while enhancing the expression of ZO-1 and claudin-5 to improve the neurological outcomes, alleviated brain water content and BBB leakage after ICH [108]. Lai et al. explored the protective effects of miR-193b-3p on early brain injury after SAH. Authors demonstrated that systematic exosomal miR-193b-3p administration effectively suppressed the expression and activity of histone deacetylase 3 (HDAC3) and upregulated the acetylation of NF-κB p65, which reduced the inflammatory cytokine expression in the hemorrhagic mouse brains, and mitigated BBB permeability, brain edema, neurodegeneration, and neurobehavioral impairments after SAH [110].
Two lncRNAs, including lncRNA small nucleolar RNA host gene 3 (Snhg3), and brown fat-enriched lncRNA 1 (Blnc1), were also found to exert BBB regulatory functions during the pathogenesis of ICH. Both LncRNA Snhg3 and Blnc1 exhibited enhanced expression under experimental ICH models (Fig. 1). Downregulation of Snhg3 improved endothelial cell proliferation and migration abilities and attenuated the apoptosis and monolayer permeability in BMVECs under oxygen and glucose deprivation (OGD) with hemin, an in vitro model of ICH [112]. Similarly, downregulation of Snhg3 in vivo improved the integrity of BBB and neurobehavioral scores, while mitigating brain water content and cell apoptosis [112]. Mechanistically, lncRNA Snhg3 can enhance the expression of TWEAK (tumor necrosis factor-like weak inducer of apoptosis) protein and its receptor Fn14 (fibroblast growth factor-inducible 14) to activate the downstream neuroinflammatory pathway STAT3, thus enhancing the secretion of MMP-2 and MMP-9, which contribute to cerebral microvascular dysfunction in ICH rats [112]. Xie et al. observed that inhibition of Blnc1 promoted endothelial cell viability, migration, and endothelial monolayer integrity in vitro. Also, suppression of Blnc1 ameliorated BBB permeability and pro-inflammatory cytokines levels, and reduced brain edema in ICH-challenged mice. Blnc1 was further confirmed to be able to positively regulate peroxisome proliferator-activated receptor gamma (PPAR-γ) levels, while Blnc1 knockdown suppressed PPAR-γ/Sirtuin 6 (SIRT6)-mediated forkhead box O-3 (FoxO3) signaling pathway in ICH mice [113].
Circular RNA circARF3 was also revealed to regulate BBB permeability in subarachnoid hemorrhage. Cai et al. observed significant downregulation of circARF3 in plasma and cerebrospinal fluid (CSF) in SAF patients with higher Fisher stages [114]. Overexpressing circARF3 robustly improved BBB integrity and neurological score, and decreased neuronal apoptosis and microglial activation in the ipsilateral basal cortex of SAF rat brains. Mechanistically, the authors demonstrated that circARF3 acted as an endogenous competitive RNA to sponge miR-31-5p, thus deactivating myeloid differentiation factor 88 (MyD88)-NF-κB pathway, to exert protection against SAH-induced BBB disruption [114].
Traumatic brain injury
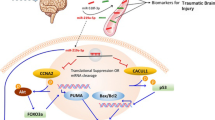
Traumatic brain injury (TBI) causes primary and secondary disruptions of microvessels’ structural and physiological integrity and results in compromised BBB integrity, which facilitates the blood-borne factors entering the brain and causes microglial activation, proliferation, and the production of pro-inflammatory factors [115]. Several miRNAs, including miR-21 [40, 116,117,118] and miR-9-5p [119], exhibited upregulated expression levels in TBI and were associated with regulating BBB integrity after TBI (Fig. 1). Ge et al. demonstrated that upregulation of miR-21 levels in rat brains conferred improved neurological recovery, alleviated BBB permeability, and reduced brain edema and lesion volume following TBI [116,117,118]. miR-21 can exert BBB protection against TBI injuries via activating the expression of VEGF and Ang-1/Tie-2 to promote the expression of tight junction proteins, such as occludin and claudin-5 to amplify BBB stabilization [116,117,118]. miR-21 was also able to inhibit the expression of PTEN (phosphatase and tensin homolog deleted on chromosome 10) and activate Akt signaling pathway to inhibit cellular apoptosis [116, 118]. In addition, miR-21 exert anti-inflammatory functions via suppressing the expression of pro-inflammatory cytokines and the NF-κB signaling pathway after TBI [118]. Wu et al. explored the contribution of miR-9-5p on the recovery of neurological function after TBI. Data revealed that upregulation of miR-9-5p significantly alleviated apoptosis, neuroinflammation, and BBB permeability in rats after TBI [119]. Authors also confirmed that miR-9-5p exerted these protective functions through targeting protein patched homolog 1 (Ptch-1) and activating the Hedgehog/AKT/Glycogen synthase kinase-3β (GSK3β) axis to inhibit the NF-κB and MMP-9 signaling pathways [119].
LncRNA KCNQ1 overlapping transcript 1 (KCNQ1OT1) was also involved in the pathogenesis of TBI and played a role in the regulation of BBB permeability after TBI. Liu et al. showed that KCNQ1OT1 was markedly overexpressed in the cerebral tissue of TBI mice, and knockdown of KCNQ1OT1 in the mouse brain exhibited alleviated neurological deficits, neuronal loss, microglial activation, pro-inflammatory cytokines expression (e.g., IL-1β, TNF-α, IL-6, etc.), and augmented anti-inflammatory cytokines (e.g., IL-10, transforming growth factor β (TGFβ), brain-derived neurotrophic factor (BDNF)) accompanied by improved BBB integrity and functions [120]. Authors also confirmed that miR-873-5p was a direct target of KCNQ1OT1, which functioned as a competitive endogenous RNA to sponge miR-873-5p, thus knocking down the levels of KCNQ1OT1 in the brain effectively decreased the levels of tumor necrosis factor receptor-related factor 6 (TRAF6) [120].
Circular RNA Lphn3 (circLphn3) was also demonstrated BBB protection in a cellular and mouse model of TBI. Cheng et al. observed that the expression of circLphn3 was substantially decreased after TBI both in vivo and in vitro [121]. Overexpression of circLphn3 enhanced the expression of tight junction proteins ZO-1, ZO-2, and occludin, to attenuate the hemin-induced high endothelial permeability in the in vitro model of BBB [121]. Authors confirmed that circLphn3 acted as a molecular sponge of miR-185-5p to upregulate the tight junction protein ZO-1 and repair the permeability of BBB after TBI [121].
Spinal cord injury
The blood-spinal cord barrier (BSCB) functions as the conceptually equivalent to BBB in the spinal cords and provides a similar functional microenvironment as BBB for the cellular constituents of spinal cords; thus, BSCB has been considered as the morphological extension of the BBB [122]. Spinal cord injury (SCI) also results in direct vascular damage and induces prominent disruption of the BSCB [123]. In this context, a number of miRNAs, including miR-27a [124], miR-129-5p [125], miR-199a-5p [126], miR-128-3p [127], miR-9 [128], have been demonstrated to exert protective effects against BSCB damage, but exhibited downregulated levels after the onset of SCI (Fig. 1). For instance, intrathecal injection of miR-129-5p mimics successfully preserved the motor function and prevented the BSCB leakage with decreased Evan blue extravasation and spinal water content [125]. Authors demonstrated that overexpression of miR-129-5p significantly reduced its molecular target high-mobility group box-1 (HMGB1), thus inhibiting Toll-like receptor (TLR)-3, IL-1β, and TNF-α levels in the injured mouse spinal cords [125]. Wang et al. showed that miR-125a-5p overexpression induced the expression of ZO-1, occludin, and VE-cadherin, which lessened the endothelial permeability and cell death in an in vitro model of SCI, suggesting miR-125a-5p as anti-apoptotic mediator in spinal cord microvascular endothelial cells after SCI [129].
Several miRNAs exert protective effects on the BSCB structure and function when they are downregulated during SCI. These miRNAs include miR-155 [56, 130], miR-429 [131], and miR-181c-5p [132] (Fig. 1). Awad et al. observed that aortic cross-clamping (ACC)-paralyzed mice exhibit higher miR-155 expression in neurons and endothelial cells than ACC mice that escaped paralysis [56]. Genetic deletion of miR-155 improved BSCB integrity, reduced central cord edema, and reduced total paralysis incidence [56]. Authors elucidated that depletion of miR-155 enhanced the protein levels of major facilitator superfamily domain-containing 2a (Mfsd2) in ECs and motoneurons to attenuate EC permeability and grey matter damage [56]. In addition, Ge et al. also confirmed that miR-155 might aggravate BSCB disruption following experimental SCI [130]. Authors showed that exosomal miR-155 from M1-polarized macrophages promotes EndoMT, a pathological condition of ECs, and impairs mitochondrial function via activating NF-κB signaling pathway after traumatic SCI by directly targeting downstream suppressor of cytokine signaling 6 (SOCS6) and inhibiting the expression of tight junction proteins ZO-1, occludin, claudin-1, -2 and -5 in vascular endothelial cells [130]. Sun et al. observed that inhibition of miR-429 expression by antagomir resulted in significantly increased levels of tight junction proteins ZO-1, occludin, and claudin-5 and reduced BSCB permeability in an extracorporeal BSCB model [129]. Authors then elucidated that miR-429 can negatively regulate KLF6 to mediated tight junction protein expression and BSCB integrity [129].
Regarding long non-coding RNA in the regulation of BSCB permeability following experimental SCI, the functional role of lncRNA taurine upregulated gene 1 (TUG1) was investigated. Jia et al. discovered that knockdown of TUG1 alleviated blood-spinal cord barrier leakage and improved hind-limb motor function by upregulating miR-29b-1-5p and suppressing metadherin (MTDH)/NF-κB pathway-mediated inflammatory cytokines expression after spinal cord ischemia-reperfusion [133].
Circular RNA has also been studied in the involvement of the pathophysiology of traumatic SCI. By using the RNA-seq technique, Wang et al. systematically studied the expression profile of circRNA in the lesion epicenter of spinal tissues after traumatic SCI, with a focus on circAbca1 [134]. Authors demonstrated that miR-135b-5p was the most significantly downregulated microRNA, and circAbca1 exhibited significant upregulation after traumatic SCI; authors then confirmed that circAbca1 plays a neuroinhibitory role by the miR-135b-5p/KLF4 axis [134], which may relate to the severe damage to the blood-spinal cord barrier observed after traumatic SCI.
Glioma, glioblastoma, and CNS metastasis
CNS tumors have grown substantially during the past two decades. Glioma constitutes the most diagnosed brain tumor type among children ages 0 to 14, glioblastoma constitutes the most common type of primary malignant brain tumor and other CNS tumor diagnosed in adults 40 years or older in the United States [135]. Glioblastoma is the most aggressive type of tumor of glioma. CNS metastasis is another type of CNS tumor, which refers to the spread of cancer cells from the primary site of the body to the CNS, and the incidence of CNS metastasis is rapidly increasing in the past decade [136]. The integrity of normal BBB was compromised during the progress of CNS tumors and resulting in a vasculature known as the blood-tumor barrier (BTB) [137]. Although BTB has been characterized as a disrupted BBB, it preserves the critical characteristics of the BBB, which still restrain the easy delivery of therapeutic agents to the tumor tissue [137], thus increasing the permeability of BTB has become a therapeutic strategy for CNS tumors. Emerging evidence has shown the extensively regulatory abilities of non-coding RNAs (miRNAs, lncRNAs, and circular RNAs, etc.) on the BBB/BTB integrity and permeability in experimental models of glioma, glioblastoma, and CNS metastasis (Fig. 1). Small non-coding RNAs that involved in BBB/BTB regulation including miR-181a [138], miR-34c [139], miR-18a [140, 141], miR-34a [142], miR-200b [143], miR-330-3p [144], piR-DQ590027/miR-17HG [145], miR-429 [146], piRNA-DQ593109/miR-330-5p [147], miR-132-3p [148], etc. for glioma and glioblastoma; and miR-105 [58], miR-181c [149], miR-509 [150], miR-101-3p [151], and miR-211 [152], etc. for brain metastasis. For example, it was described that miR-429 exhibited lower expression in glioma endothelial cells (GECs), an in vitro BTB model, and overexpression of miR-429 in GECs significantly decreased the expression of tight junction proteins ZO-1, occludin, and claudin-5, as ZO-1 and occludin were direct targets of miR-429 [146]. In addition, miR-429 can also downregulate the tight junction-associated proteins by targeting p70S6K to increase the BTB permeability in glioma models [146]. It was also reported that the expression of miR-132-3p was greatly upregulated in GECs, and miR-132-3p contributed to the increased permeability of BTB and caveolae-mediated transcellular transport by targeting its downstream molecule PTEN, and positive regulation of phosphorylated protein kinase B (p-PKB), p-Src, and p-Cav-1 (Tyr14 phosphorylation of caveolin-1) [148]. Moreover, in breast cancer brain metastasis, Pan et al. demonstrated that high levels of miR-211 drove early and specific brain colonization of tumor cells to enhance their stemness properties and trans-BBB adherence and migration to promote brain metastasis by downregulating the SOX11 (SRY-Box Transcription Factor 11)/NGN2 (neurogenin 2)-dependent axis in vivo [152].
Besides small non-coding RNAs, numerous lncRNAs exhibited upregulated expression in brain tumors, and their inhibition generally yields decreased BTB integrity and increased permeability through different pathways. These lncRNAs includes lncRNA TUG1 [153, 154], Malat1 [155, 156], Lnc-BM [62], XIST [157], NEAT1 (nuclear paraspeckle assembly transcript 1) [158], HOTAIR (homeobox transcript antisense intergenic RNA) [159], linc00174 (long intergenic non-protein coding RNA 174) [160], Lnc00462717 [161], CCRR (cardiac conduction regulatory RNA) [162]. These lncRNAs normally act as competing endogenous RNAs to bind their downstream microRNAs, then regulate the BTB permeability. For instance, Zhang et al. observed that Lnc00462717 was upregulated in GECs, and that knockdown of Lnc00462717 reduced its interaction with PTBP1 (Polypyrimidine Tract Binding Protein 1), thus significantly increasing levels of miR-186-5p to subsequently downregulate the protein expression of occludin to increase the BTB permeability [161]. LncRNA GS1-600G8.5 [163] and MIAT [164] were also highly expressed in brain metastatic cells and GECs, however, instead of suppression, overexpression can facilitate endothelial barrier permeability and promote the invasion of cancer cells across the BBB/BTB. For example, MIAT was shown to function as a miR-140-3p sponge to upregulate the expression of ZAK (ZO-1-associated kinase) and the phosphorylation of NF-κB-p65, to inhibit the expression of tight junction proteins ZO-1, occludin, and claudin-5 to increase permeability in an in vitro model of BTB [164].
Furthermore, it was observed that circular RNA USP1 (circ‐USP1) [165], circRNA_001160 [166], and circular RNA DENND4C (cDENND4C) [167] were also deeply involved and markedly upregulated in GECs. Knockdown of the cellular expression of circ‐USP1, circRNA_001160, cDENND4C disrupted the barrier integrity, increased barrier permeability accompanied by reduced tight junction-related proteins claudin-5, occludin, and ZO-1 by targeting miR-194-5p/FLI1 (Friend leukemia virus integration 1), miR-195-5p/ETV1 (Ets variant gene 1), and miR-577, respectively [165,166,167].
Multiple sclerosis
Multiple sclerosis (MS) is a chronic, autoimmune disease that affects the normal function of the CNS. A number of studies have investigated the involvement and functional significance of various non-coding RNAs in the regulation of barrier function in MS. Several miRNAs exhibited downregulated expression from MS patients’ brain/spinal cord tissue or blood samples, as well as in vitro BBB models treated with pro-inflammatory molecules (Fig. 1). These downregulated miRNAs include miR-125-5p [168], miR-320a [169], and miR-126 & miR-126* [170]. Upregulating their expression levels can generally improve brain endothelial cell barrier function by decreasing the expression of cell adhesion molecules, such as ICAM-1, VCAM-1, and E-selectin, or targeting intracellular MMPs expressions. For example, Cerutti et al. observed that reduction of endothelial miR-126 and miR-126* resulted in enhanced expression of E-selectin and VCAM-1, respectively, to enhance the firm adhesion of leukocytes and primary MS patient-derived peripheral blood mononuclear cell (PBMC) to brain endothelial cells [170]. In contrast, overexpression of miR-126 reduced the expression of VCAM1 and MCP1 (monocyte chemoattractant protein 1) expression in brain microvascular endothelial cells [170].
miR-155 exhibited upregulated cerebral expression in MS patients or experimental models [171,172,173]. Lopez-Ramirez et al. showed that loss of miR-155 reduced BBB extravasation of both experimental autoimmune encephalomyelitis (EAE) and in an acute systemic inflammation model induced by lipopolysaccharide [172]. The mechanistic investigation demonstrated that miR-155 modulated brain endothelial barrier function by targeting both cell-cell complex molecules, such as annexin-2 and claudin-1, and cell-to-extracellular matrix interactions, such as dedicator of cytokinesis 1 (DOCK-1) and syntenin-1 (SDCBP), to increase the barrier permeability [172]. It was also reported that miR-155 overexpression boosted the levels of VCAM-1 and ICAM-1, which facilitated the firm adhesion of monocytic and T cells to both unstimulated and pro-inflammatory cytokines-stimulated human brain endothelium, thus enhancing the leukocytes extravasation of the inflamed BBB [173].
As astrocytes actively participate in the formation and integrity of the BBB, the function of astrocytic lncRNA Gm13568 in the regulation of MS pathophysiology has been investigated in experimental EAE mice and primary astrocyte culture. Liu et al. observed that inhibiting Gm13568 levels in astrocytes significantly ameliorated inflammation and demyelination in EAE mice, which delayed the progress of experimental EAE [174]. Knockdown of the endogenous Gm13568 in IL-9 treated primary astrocyte culture remarkably suppressed astrocytosis and the phosphorylation of signal transducer and activator of transcription 3 (p-STAT3) as well as the production of inflammatory cytokines and chemokines (IL-6, TNF-α, interferon-inducible protein-10) through inhibiting the Notch1 pathway [174].
Moreover, circular RNA HECW2 (circ_HECW2) [175, 176] was also involved in the pathogenesis of MS. Elevated expression of circ_HECW2 leads to EndoMT, which plays a critical role in the dysfunction of BBB and contributes to BBB leakage, in both in vitro and in vivo MS experimental models [175, 176]. Yang et al. showed that circ_HECW2 functioned as a miR-30D sponge to increase the expression of ATG5 (autophagy-related 5) and activate the NOTCH pathway, then positively regulate LPS-induced EndoMT [175]. Dong et al. further revealed that circ_HECW2 also interacted with miR-30e-5p to regulate the levels of neuronal growth regulator 1, which repressed endothelial cell proliferation and exacerbated apoptosis and LPS-induced EndoMT [176].
Dementia
BBB breakdown and pericyte degeneration have been found in vascular cognitive impairment and dementia (VCID) and Alzheimer's disease (AD). In this context, non-coding RNAs have emerged as critical BBB regulators during the pathological process of these CNS disorders. In experimental AD models, it has been reported that miR-107 [177], miR-181a [59], miR-124 [178] showed downregulated expression, while miR-96 [179] and miR-424-5p [180] showed up-regulated expression in AD environments (Fig. 1). In experimental VCID models, it has been shown that miR-126 was downregulated in multiple microinfarction (MMI) model-induced vascular dementia [181], and miR-501-3p was upregulated in bilateral common carotid artery stenosis (BCAS)-induced VCID [182] (Fig. 1). These dysregulated microRNAs are typically associated with decreased endothelial cell viability, impaired BBB integrity, increased BBB permeability, and/or declined microvascular density and angiogenesis. On the other hand, overexpression of miR-107 [177], miR-181a [59], miR-124 [178], or inhibition of miR-96 [179] and miR-424-5p [180] in AD significantly abrogated beta-amyloid-induced cerebrovascular injury and BBB disruption through upregulating junctional protein expression or ameliorating pericyte apoptosis. For example, Zhang et al. reported that miR-96 could target erythroblast transformation-specific (ETS) transcription factor ERG (ETS-related gene) to inhibit ERG protein expression, which can bind to ZO-1 promoter region to downregulate the ZO-1 transcription in BMECs; thus, inhibition of miR-96 prevented ZO-1 downregulation induced by GM-CSF (granulocyte-macrophage colony-stimulating factor) in AD environments [179]. Li et al. observed that C1ql3, one of the C1q subunits, was a potential target of miR-124, and overexpression of miR-124 dramatically elevated the expression of ZO-1 and robustly rescued breakdown of the BBB, promoted angiogenesis and reduced Aβ deposition, and finally alleviated learning and memory deficit in APP/PS1 mice [178]. In an MMI-induced vascular dementia model, Yu et al. discovered that miR-126 was downregulated and negatively regulated MMP-9 and TLR4 inflammatory factor expression in endothelial cells, which are related to BBB disruption and neuroinflammation [181]. EC-targeted deletion of miR-126 exhibited significant water channel and glymphatic impairment [181], suggesting upregulation of miR-126 levels may provide beneficial therapeutic effects against vascular cognitive impairments.
Long non-coding RNA LINC00662 was also able to regulate the BBB permeability in Alzheimer's microenvironment. It was reported that LINC00662 was upregulated in beta-amyloid-incubated microvascular endothelial cells, and knockdown of LINC00662 decreased BBB permeability in AD microenvironment [183]. LINC00662 downregulated the expression of ETS-domain protein 4 (ELK4), which can bind to the promotors of ZO-1, occludin, and claudin-5 to promote their protein expression [183]. Thus, inhibition of LINC006622 resulted in enhanced expression of ELK4, increased the levels of tight junction proteins, and improved BBB integrity in the AD microenvironment [183].
Bacterial and viral infections
Cerebral infections can be caused by various bacteria, viruses, fungi, parasites, or other inflammatory factors, leading to severe brain inflammation and injuries. The integrity and normal functions of BBB are largely compromised by the infectious microorganisms, resulting in increased BBB permeability and exacerbated brain injuries [184]. In this context, emerging evidence has demonstrated the regulatory function of non-coding RNAs on BBB integrity and permeability during cerebral infections. For instance, Mishra et al. showed that HIV Tat C protein significantly impaired the BBB permeability and decreased the expression of VE-cadherin and tight junction proteins (TJPs) (claudin-5, ZO-1, occludin) [185]. Further investigation observed that HIV Tat C protein increased the expression of miR-101, which led to suppression of VE-cadherin in human BMECs [185]. Rom et al. observed the downregulation of miR-98 and let-7g* in experimental models of aseptic meningitis [186]. Overexpression of let-7 and miR-98 suppressed the secretion of CCL2 and CCL5, reducing the leukocyte adhesion and migration across the endothelium, diminishing the pro-inflammatory cytokines, and improving the BBB integrity [186]. In an experimental mouse model of cerebral malaria, plasmodium berghei ANKA (PbA) infection increased circulating exosomal miR-155 levels [187]. Genetic deletion of miR-155 ameliorated endothelial activation, preserved BBB integrity, and improved survival rate in response to infection in experimental cerebral malaria (ECM) models [187]. miR-155 antagomir administration also reduced ex vivo vascular leakage in human cerebral microvessels exposed to sera collected from children with cerebral malaria in Ugandan [187]. In an in vitro BBB model with coxsackievirus A16 (CA16) infection and an in vivo CA16 infant rhesus monkey infection model, Song and colleagues suggested that CA16 infection downregulated miR-1303 levels and upregulated MMP9 expression, which promoted the degradation of junctional proteins, including claudin4, claudin5, VE-cadherin, and ZO-1, and ultimately causing neuroinflammation and injury to the CNS [188].
Two long non-coding RNAs are involved in the regulation of bacterial meningitis-induced BBB damages. Wang et al. showed that NEAT1 levels were upregulated in glioma-exposed endothelial cells and miR-135a was a direct target of NEAT1 [189]. Downregulation of NEAT1 effectively maintained BBB integrity and decreased BBB permeability in bacterial meningitis experimental models through upregulating miR-135a and downregulating HIF1α to increase the expression of ZO-1, occludin, and claudin-5 [189]. On the other hand, overexpression of NEAT1 increased BBB permeability in both in vitro and in vivo bacterial meningitis models [189]. In an in vitro bacterial meningitis model, Xu et al. demonstrated that long non-coding RNA LncRSPH9-4 was significantly elevated and cytoplasmically distributed in meningitic E. coli-infected hBMECs. LncRSPH9-4 was able to regulate the BBB permeability by competitively sponging miR-17-5p, thereby increasing MMP3 expression, which can target the tight junction proteins ZO-1, occludin, and claudin-5 in meningitic E. coli-infected hBMECs [190].
Yang and colleagues also observed circular RNA circ_2858 mediated the BBB disruption in bacterial meningitis. Data revealed that circ_2858 was significantly upregulated by meningitic E. coli infection in human BMECs, and circ_2858 competitively bound miR-93-5p to elevate VEGFA levels [191]. The enhanced VEGFA expression led to downregulation and altered distribution of tight junction proteins such as ZO-1, occludin, and claudin-5, which eventually increased BBB permeability in bacterial meningitis [191].
Diabetes
Hyperglycemia and diabetes have been known to induce cerebrovascular stress and trigger BBB impairment and permeability, leading to severe cerebrovascular disorders, such as stroke and dementia [192,193,194]. It has been reported that several microRNAs participated in BBB regulation under diabetic conditions. Song et al. described that high glucose condition downregulated miR-Let7A expression in brain endothelial cells in vitro. Overexpression of miR-Let7A markedly attenuated endothelial cell death and the loss of tight junction proteins (claudin-5 and ZO-1), diminished the levels of pro-inflammatory factors (TNF-α and iNOS), and nitrite production in the brain endothelial cells under high glucose condition [195]. Zhao et al. demonstrated HDAC3 inhibition reduced diabetes-induced BBB permeability and rescued junction protein expression in db/db diabetic mice, and HDAC3 inhibition-mediated protective effects against BBB permeability was at least partly mediated by miR-200a [196]. Data revealed that HDAC3 inhibition significantly increased the miR-200a levels, which targeted and downregulated Keap1 (Kelch-like ECH-associated protein 1), the negative regulator of Nrf2 (nuclear factor-erythroid factor 2-related factor 2), thereby contributing to Nrf2 activation and ultimately the protection against endothelial monolayer permeability under diabetic conditions [196].
Sepsis-associated encephalopathy
Blood–brain barrier disruption induced by sepsis plays a critical role in the pathophysiology of sepsis-associated encephalopathy (SAE), which increases the influx and efflux of various circulating immune cells, detrimental pathogens, and harmful molecules between the circulation and the brain [197]. Recently, emerging studies have described the functional significance of microRNAs in regulating BBB permeability in SAE. For example, in a septic rat model, Chen et al. demonstrated that miR-181b was elevated and negatively targeted sphingosine-1-phosphate receptor 1 (S1PR1) and neurocalcin delta (NCALD) [198]. Inhibition of miR-181b levels reduced damage and permeability to the BBB via increasing the expression of S1PR1 and NCALD in septic rats [198]. Visitchanakun et al. showed increased expression of miR-370-3p in plasma and brain tissue was associated with SAE outcomes [60]. Plasma miR-370-3p also specifically increased and highly sensitive for early detection (6h) of cecal ligation and puncture (CLP)-induced SAE with BBB permeability, elevated TNF-a, and brain apoptosis [60]. Additionally, Nong et al. revealed downregulated expression of miR-126 in septic rat brain tissues, and overexpression of miR-126 significantly reduced the brain tissue water content and BBB permeability in SAE rats, and significantly increased the expression of claudin-5 and occludin. Overexpression of miR-126 also decreased the serum levels of pro-inflammatory factors TNF-α, IL-6, and IL-1β and increased the expression of anti-inflammatory IL-10 [199].
Other CNS disorders
Recent investigations also suggested that microRNAs are involved in regulating blood–brain barrier integrity in other pathophysiological events, including shear stress, development, aging, tuberous sclerosis complex, reversible cerebral vasoconstriction syndrome (RCVS), and others. For instance, shear stress-upregulated miR-27b in endothelial cells increased pericyte adhesion and pericyte recruitment of endothelial tubes to preserve the BBB integrity [200]. miR-285 and miR-132 were demonstrated to be essential to BBB development and to maintain brain vascular integrity [201]. miR-285 directly targets the Yki cofactor Mask (Multiple Ankyrin repeats Single KH domain) to suppress Yki activity and downregulate the expression of cyclin E to regulate cell cycle and keep proper cell size to maintain a functional BBB in Drosophila [201]. miR-132 regulated the brain vascular integrity by affecting adherens junction protein VE-cadherin rather than transcytosis or pericytes in larval zebrafish [202]. In aging, cerebral miR-195 levels decreased with age, and led to increased expression of thrombospondin-1 (TSP1), which can activate selective autophagy of tight junction proteins by increasing the formation of claudin-5-p62 and ZO-1-p62 complexes, resulting in TJ protein degradation and BBB permeability [203]. In tuberous sclerosis complex (TSC), high protein expression of MMPs (MMP2, 3, 9, and 14) and TIMPs (endogenous tissue inhibitors 1, 2, 3, and 4) in TSC tubers was associated with BBB dysfunction, while these dysregulated proteins can be partly rescued by miR-146a and miR-147b in tuber-derived TSC cultures [204]. In reversible cerebral vasoconstriction syndrome (RCVS), high expression of circulating miR-130a-3p was associated with BBB disruption in patients, and overexpression of miR-130a-3p also led to increased BBB permeability in vitro [205]. Neuropilin-1 (Npn-1) has been suggested to play a critical role in regulating endothelial barrier dysfunction in response to VEGF [206] or interferon-γ [207], and Mone et al. demonstrated that Npn-1 is a direct target of miR-24, which could negatively regulate Npn-1 mediated endothelial permeability [208].
Non-coding RNAs as therapeutic targets of pharmacotherapy in regulating BBB/BSCB functions in CNS disorders
Some pharmaceutical drugs preserve the ability to regulate BBB or BSCB functions by modulating non-coding RNAs, thus making these ncRNAs therapeutic targets in regulating BBB or BSCB integrity and permeability in CNS disorders. As shown in Table 4, we summarized the reported pharmacologic agents that exert BBB regulatory functions in different experimental models of CNS disorders, including ischemic stroke, intracerebral hemorrhage, TBI, SCI, AD, and others. Polydatin (PD), a natural product, has been described to enhance lncRNA MALAT1 gene expression, reduce cell toxicity, apoptosis, and inflammatory factor expression in rat brain microvascular endothelial cells [209]. Also, PD administration in rats elevated MALAT1 expressions, reduced cerebral infarct volume and brain inflammation, protected cerebrovascular endothelial cells and BBB integrity after cerebral ischemia [209]. Similarly, Alisol A 24-acetate (AA), a natural compound, has been observed to downregulate the expression of miR-92a-3p, increase ZO-1, claudin-5, and occludin expression in the OGD-insulted BMECs, which provided evidence for AA application in aging-associated BBB protection [210]. Methamphetamine abuse led to upregulation of miR-143 in isolated brain microvessels and tissues, which was also accompanied by BBB leakage [211]. Silencing miR-143 ameliorated methamphetamine-triggered permeability of endothelial cells monolayer and the BBB both in vitro and in vivo through targeting PUMA (p53 upregulated modulator of apoptosis), NF-κB, and p53 transcription factor pathway [211]. Salvianolic acid A can improve the recovery of neurological function after SCI, which could be correlated with the repair of BSCB integrity by the miR-101/Cul3 (Cullin 3)/Nrf2/HO-1/ZO-1 and occludin signaling pathway [212]. Hydrogen gas (H2) can robustly improve neurological outcomes after TBI by mitigating neurological dysfunction, alleviating brain edema, and decreasing lesion volume and BBB permeability by significantly increasing the expression of miR-21 [213]. Memantine (MEM), an N-methyl-d-aspartate (NMDA) receptor antagonist, has been demonstrated to alleviate the drastic increase of circular RNA LINC00094 in beta-amyloid-incubated microvascular endothelial cells in an in vitro BBB model. Suppression of LINC00094 levels significantly mitigated BBB permeability and upregulated the expression of ZO-1, occludin, and claudin-5 through the miR-224-5p (miR-497-5p)/Endophilin-1 axis [214].
Challenges, perspectives, and future goals
BBB dysfunction is a common pathological feature upon the onset of various CNS diseases, and the disrupted BBB further exacerbates the initial CNS damage. BBB damage has become a vital factor in determining the progression and outcomes of CNS disorders. Currently, there is no available clinical pharmacotherapy in the treatment of the BBB dysfunctions directly [216]. The involvement and regulatory functions of non-coding RNAs on BBB dysfunction in CNS disorders have been rapidly and vastly investigated during the past decade. Considerable evidence has demonstrated the effectiveness and ability of miRNAs, lncRNAs, and circRNAs in the protection of the BBB/BSCB in stroke, TBI, SCI, MS, dementia, brain infections, diabetes, SAE, and others, and in the enhancement of BTB permeability to facilitate the anti-cancer drug delivery in glioma and brain metastasis. As shown in Fig. 2, miRNAs, lncRNAs, and circRNAs modulate the integrity of BBB through a number of mechanisms, including directly or indirectly modulate TJ proteins (mostly claudin-5, occludin, and ZO-1), AJ proteins, MMPs, water channel-related proteins, angiogenesis-related proteins, inflammation-related factors, apoptosis-related molecules, oxidative stress-related factors, autophagy-related proteins, and others, in endothelial cells, pericytes, or astrocytes of the CNS. LncRNAs and circRNAs primarily act as microRNA “sponges” or competing endogenous RNAs to participate in BBB regulation in CNS diseases. There are also increasingly discovered agents in regulating BBB/BSCB functions in CNS disorders through the functions of ncRNAs. Thus, these studies have shed light on the discovery of novel pharmaceutical drugs for the treatment of BBB impairments in numerous CNS diseases. However, nowadays, the most commonly used drugs to treat BBB dysfunctions are glucocorticoids that are able to improve the tightness and contribute to BBB stabilization [216,217,218], and numerous difficulties and challenges exist on the translation of non-coding RNA-based therapeutics from bench to bedside.

Summarized mechanisms of BBB regulatory miRNAs, lncRNAs, and circRNAs in CNS disorders. MiRNAs, lncRNAs, and circRNAs modulate the integrity of BBB through different mechanisms, including direct or indirect modulation of tight junction proteins, adherens junction proteins, MMPs, water channel-related proteins, angiogenesis-related proteins, inflammation-related factors, apoptosis-related molecules, oxidative stress-related factors, autophagy-related proteins, and others, in endothelial cells, pericytes, or astrocytes of the CNS. LncRNAs and circRNAs primarily act as microRNA “sponges” or competing endogenous RNAs to regulate BBB damage and recovery in CNS diseases
One challenge might be the side-effects of the usage of one non-coding RNA in the disease. As one ncRNA can target numerous different nucleotides or molecules, such as microRNA [219], some ncRNAs play different or even opposite roles on BBB integrity among different CNS diseases, and the most used methods to experimentally modulate the expression of ncRNAs are mimics or inhibitors with systematic administration or cell cultures, strategies to increase the specificity of ncRNAs to their targets, cell types, location are critical for the efficiency and efficacy of ncRNA-based therapeutics. Also, the relationship between the expression levels of dysregulated ncRNAs and the severity of BBB/BSCB/BTB leakage in CNS disease needs to be better investigated.
Another challenge is searching for the origin of the dysregulated ncRNAs after CNS diseases. Most investigations utilize the whole blood, whole or part of CNS tissues to analyze the levels of dysregulated ncRNA, but lack of search for the actual origin (e.g., cell type, tissue type), and how did the dysregulated ncRNA transport to the affected region of the CNS. Interestingly, some studies have focused on the functional roles of cell-specific ncRNAs on BBB or cerebrovascular systems after CNS injuries [65, 66, 181]. Other studies have investigated the transportation of ncRNA from one type of neuronal cells to BBB components and the regulatory mechanisms for the BBB integrity in CNS diseases [130, 149, 163, 202].
As to investigating the roles of ncRNAs in the regulation of BBB, most studies focused on TJ proteins, especially claudin-5, occludin, and ZO-1. Other TJ proteins, AJ proteins, JAMs, and cytoplasmic accessory proteins are critical elements as well to build an intact and functional BBB, thus warranting more research efforts in the regulation of BBB by ncRNAs in CNS diseases. Moreover, pericytes and astrocytes are crucial for maintaining BBB integrity and function, but only a few studies have been involved regarding ncRNA-regulated roles in CNS diseases. For example, Wan et al. show the functions of miR-149-5p in pericytes on BBB integrity after ischemic stroke [77], Wu et al. demonstrate the protective role of miR-181a in pericytes on BBB breakdown in AD [59], and Wang et al. validate the functional role of lncRNA Malat1 in downregulating astrocyte apoptosis and the water channel protein AQP-4 in ischemic stroke [97]. Surprisingly, almost no study of ncRNA has been discovered in the regulation of the basement membrane of the BBB in CNS diseases. The components of the basement membrane might be becoming interesting targets and deserve more research attention.
In stroke studies, sex difference has been increasingly recognized as a vital factor in determining the severity of neurological outcomes between male and female animals [220, 221]. Further efforts also need to be considered in the experimental design to investigate the roles of ncRNA in the regulation of BBB functions after cerebral ischemia.
MiRNAs have been intensively studied in the regulation of BBB in different CNS diseases, but the functional significance and molecular mechanisms of lncRNAs and circRNAs in CNS diseases are relatively less studied, especially for circRNAs, which might be due to their unique circular structure. With the development of next-generation sequencing, the advance of complete genome sequence and bioinformatics technology, more miRNAs, lncRNAs, and circRNAs will be identified, and their regulatory mechanisms on BBB permeability will also be better elucidated with the advance of methodology in neuroscience.
Despite these challenges and unsolved problems, the emerging investigations and in-depth mechanism elucidation will not only advance our current knowledge of different non-coding RNAs in the regulation of structure and function of the BBB in all CNS diseases, but also pave a fundamental basis for the development of ncRNA-based therapeutics from pre-clinical animal models to human clinical applications.
Availability of data and materials
Not applicable.
Abbreviations
- ncRNAs:
-
Non-coding RNAs
- CNS:
-
Central nervous system
- BBB:
-
Blood–brain barrier
- miR:
-
MicroRNA
- lncRNA:
-
Long non-coding RNA
- circRNA:
-
Circular RNA
- TBI:
-
Traumatic brain injury
- SCI:
-
Spinal cord injury
- MS:
-
Multiple sclerosis
- AD:
-
Alzheimer's disease
- VCID:
-
Vascular cognitive impairment and dementia
- SAE:
-
Sepsis-associated encephalopathy
- rRNA:
-
Ribosomal RNA (rRNAs)
- tRNA:
-
Transfer RNAs
- piRNAs:
-
Piwi-interacting RNAs
- snRNAs:
-
Small nuclear RNAs
- snoRNA:
-
Small nucleolar RNAs
- asRNAs:
-
Antisense RNAs
- PALRs:
-
Promoter-associated long RNAs
- PROMPTs:
-
Promoter upstream transcripts
- LSINCTs:
-
Long stress-induced non-coding transcripts
- 3’ UTR:
-
3’ Untranslated region
- RNAPII:
-
RNA polymerase II
- VSMC:
-
Vascular smooth muscle cells
- AJ:
-
Adherens junction
- NIHSS:
-
National Institute of Health stroke scale
- vWF:
-
von Willebrand Factor
- ICH:
-
Intracerebral hemorrhage
- PHE:
-
Perihematomal edema
- RRMS:
-
Relapsing–remitting multiple sclerosis
- EDSS:
-
Expanded disability status scale
- NF-κB:
-
Nuclear factor-κB
- ICAM-1:
-
Intracellular adhesion molecule 1
- ITGB-2:
-
Integrin subunit beta 2
- TJ:
-
Tight junction
- ZO-1:
-
Zonula occludens 1
- Aβ:
-
Beta-amyloid
- VCAM-1:
-
Vascular cell adhesion protein 1
- BCBM:
-
Breast cancer brain metastasis
- Lnc-BM:
-
LncRNA associated with breast cancer brain metastasis
- STAT3:
-
Signal transducer and activator of transcription 3
- CCL2:
-
C–C chemokine ligand 2
- MCP-1:
-
Monocyte chemoattractant protein-1
- circDLGAP4:
-
CircRNA DLGAP4
- MMP:
-
Matrix metalloproteinase
- IL:
-
Interleukin
- TNF-α:
-
Tumor necrosis factor
- MAP2k3:
-
Mitogen-activated protein kinase kinase 3
- iNOS:
-
Inducible nitric oxide synthase
- Malat1:
-
Metastasis-associated lung adenocarcinoma transcript 1
- MEG3:
-
Maternally expressed gene 3
- Snhg8:
-
Small nucleolar RNA host gene 8
- XIST:
-
X-inactive-specific transcript
- BMEC/BMVEC:
-
Brain microvascular endothelial cell
- ULK2:
-
Uncoordinated 51-like kinase 2
- AQP4:
-
Aquaporin-4
- KLF4:
-
Krüppel-like factor 4
- circDLGAP4:
-
Circular RNA DLGAP4
- EndoMT:
-
Endothelial to mesenchymal transition
- circCCDC9:
-
Circular RNA CCDC9
- eNOS:
-
Endothelial nitric oxide synthase
- ICH:
-
Intracerebral hemorrhage
- ICV:
-
Intracerebroventricular
- DUSP8:
-
Dual-specificity phosphatase 8
- HO-1:
-
Heme oxygenase-1
- SAH:
-
Subarachnoid hemorrhage
- Cav-1:
-
Caveolin-1
- VEGF:
-
Vascular endothelial growth factor
- Ang-1:
-
Angiopoietin-1
- PIK3R2:
-
Phosphoinositide-3-kinase regulatory subunit 2
- MSC:
-
Mesenchymal stem cell
- PAR-1:
-
Protease-activated receptor-1
- HDAC3:
-
Histone deacetylase 3
- Snhg3:
-
Small nucleolar RNA host gene 3
- Blnc1:
-
Brown fat-enriched lncRNA 1
- OGD:
-
Oxygen and glucose deprivation
- TWEAK:
-
Tumor necrosis factor-like weak inducer of apoptosis
- Fn14:
-
Fibroblast growth factor-inducible 14
- PPAR-γ:
-
Peroxisome proliferator-activated receptor gamma
- SIRT6:
-
Sirtuin 6
- FOXO3:
-
Forkhead box O3
- CSF:
-
Cerebrospinal fluid
- MyD88:
-
Myeloid differentiation factor 88
- PTEN:
-
Phosphatase and tensin homolog deleted on chromosome 10
- Ptch-1:
-
Patched homolog 1
- GSK3β:
-
Glycogen synthase kinase-3β
- KCNQ1OT1:
-
KCNQ1 overlapping transcript 1
- TGFβ:
-
Transforming growth factor β
- BDNF:
-
Brain-derived neurotrophic factor
- TRAF6:
-
Tumor necrosis factor receptor-related factor 6
- circLphn3:
-
Circular RNA Lphn3
- BSCB:
-
Blood-spinal cord barrier
- HMGB1:
-
High-mobility group box-1
- TLR:
-
Toll-like receptor
- ACC:
-
Aortic cross-clamping
- Mfsd2:
-
Major facilitator superfamily domain-containing 2a
- SOCS6:
-
Suppressor of cytokine signaling 6
- TUG1:
-
Taurine upregulated gene 1
- MTDH:
-
Metadherin
- BTB:
-
Blood-tumor barrier
- GECs:
-
Glioma endothelial cells
- p-PKB:
-
Phosphorylated protein kinase B
- p-Cav-1:
-
Tyr14 phosphorylation of caveolin-1
- SOX11:
-
SRY-Box Transcription Factor 11
- NGN2:
-
Neurogenin 2
- NEAT1:
-
Nuclear paraspeckle assembly transcript 1
- HOTAIR:
-
Homeobox transcript antisense intergenic RNA
- linc00174:
-
Long intergenic non-protein coding RNA 174
- CCRR:
-
Cardiac conduction regulatory RNA
- PTBP1:
-
Polypyrimidine Tract Binding Protein 1
- ZAK:
-
ZO-1-associated kinase
- cDENND4C:
-
Circular RNA DENND4C
- FLI1:
-
Friend leukemia virus integration 1
- ETV1:
-
Ets variant gene 1
- PBMC:
-
Peripheral blood mononuclear cell
- EAE:
-
Experimental autoimmune encephalomyelitis
- DOCK-1:
-
Dedicator of cytokinesis 1
- SDCBP:
-
Syntenin-1
- MMI:
-
Multiple microinfarction
- BCAS:
-
Bilateral common carotid artery stenosis
- ETS:
-
Erythroblast transformation-specific
- ERG:
-
ETS-related gene
- GM-CSF:
-
Granulocyte–macrophage colony-stimulating factor
- ELK4:
-
ETS-domain protein 4
- PbA:
-
Plasmodium berghei ANKA
- ECM:
-
Experimental cerebral malaria
- CA16:
-
Coxsackievirus A16
- Keap1:
-
Kelch-like ECH-associated protein 1
- Nrf2:
-
Nuclear factor-erythroid factor 2-related factor 2
- S1PR1:
-
Sphingosine-1-phosphate receptor 1
- NCALD:
-
Neurocalcin delta
- CLP:
-
Cecal ligation and puncture
- RCVS:
-
Reversible cerebral vasoconstriction syndrome
- Mask:
-
Multiple Ankyrin repeats Single KH domain
- TSP1:
-
Thrombospondin-1
- TSC:
-
Tuberous sclerosis complex
- TIMP:
-
Tissue inhibitors of matrix metalloproteinases
- RCVS:
-
Reversible cerebral vasoconstriction syndrome
- Npn-1:
-
Neuropilin-1
- ATG5:
-
Autophagy Related 5
- PD:
-
Polydatin
- AA:
-
Alisol A 24-acetate
- PUMA:
-
P53 upregulated modulator of apoptosis
- Cul3:
-
Cullin 3
- H2 :
-
Hydrogen gas
- MEM:
-
Memantine
- NMDA:
-
N-Methyl-d-aspartate
- MAPK:
-
Mitogen-activated protein kinase
- MF:
-
Monomethyl fumarate
- CREB:
-
CAMP-response element binding protein
- PGC-1α:
-
Peroxisome proliferator-activated receptor-gamma coactivator-1alpha
- CYC C:
-
Cytochrome c
- S1PR2:
-
Sphingosine-1-phosphate receptor 2
- PLA2G2A:
-
Phospholipase A2 group IIA
- ALOX5:
-
Arachidonate 5-Lipoxygenase
- ITGA2B:
-
Integrin subunit alpha 2b
- JAM3:
-
Junctional adhesion molecule 3
- TJAP1:
-
Tight junction associated protein 1
- mTOR:
-
Mammalian target of rapamycin
- PI3K:
-
Phosphoinositide 3-kinases
- HMGA2:
-
High mobility group AT-hook 2
- AChE:
-
Acetylcholinesterase
- AQP11:
-
Aquaporin-11
- DUSP8:
-
Dual specificity phosphatase 8
- RUNX:
-
Runt-related transcription factor
- SOD:
-
Superoxide dismutase
- MDA:
-
Malondialdehyde
- MPO:
-
Myeloperoxidase
- SHED:
-
Stem cells from human exfoliated deciduous teeth
- CTGF:
-
Connective tissue growth factor
- MFSD2A:
-
Major facilitator superfamily domain containing 2A
- ECE1:
-
Endothelin converting enzyme 1
- SP1:
-
Specificity protein 1
- NOTCH2:
-
Notch receptor 2
- SOX:
-
Sex determining region Y-box protein
- MEF2D:
-
Myocyte enhancer factor 2D
- EMAP-II:
-
Endothelial monocyte-activating polypeptide-II
- RhoA:
-
Ras homolog family member A
- ROCKII:
-
Rho-associated protein kinase II
- PKC:
-
Protein kinase C
- MIR17HG:
-
MiR-17-92a-1 cluster host gene
- FOXR2:
-
Forkhead box R2
- PIWIL1:
-
Piwi like RNA-mediated gene silencing 1
- ITGB2:
-
Integrin subunit beta 2
- CBF:
-
Cerebral blood flow
- mBVPs:
-
Mouse brain vascular pericytes
- BMECs:
-
Brain microvascular endothelial cells
- FOXO1:
-
Forkhead box O1
- SEMA6A:
-
Semaphorin 6A
- SEMA6D:
-
Semaphorin 6D
- eef2k:
-
Eukaryotic elongation factor 2 kinase
- ceRNA:
-
Competing endogenous RNA
- Itga5:
-
Integrin a5
References
Beermann J, Piccoli MT, Viereck J, Thum T. Non-coding RNAS in development and disease: background, mechanisms and therapeutic approaches. Physiol Rev. 2016;96(4):1297–325. https://doi.org/10.1152/physrev.00041.2015.
Arraiano CM. Regulatory noncoding RNAs: functions and applications in health and disease. FEBS J. 2021;288(22):6308–9. https://doi.org/10.1111/febs.16027.
Salvatori B, Biscarini S, Morlando M. Non-coding RNAs in nervous system development and disease. Front Cell Dev Biol. 2020;8. https://doi.org/10.3389/fcell.2020.00273.
Oe S, Kimura T, Yamada H. Regulatory non-coding RNAs in nervous system development and disease. Front Biosci-Landmrk. 2019;24:1203–40. https://doi.org/10.2741/4776.
Sun P, Liu DZ, Jickling GC, Sharp FR, Yin KJ. MicroRNA-based therapeutics in central nervous system injuries. J Cerebr Blood F Met. 2018;38(7):1125–48. https://doi.org/10.1177/0271678x18773871.
O’Brien J, Hayder H, Zayed Y, Peng C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne). 2018;9:402. https://doi.org/10.3389/fendo.2018.00402.
Wei JW, Huang K, Yang C, Kang CS. Non-coding RNAs as regulators in epigenetics. Oncol Rep. 2017;37(1):3–9. https://doi.org/10.3892/or.2016.5236.
Ozata DM, Gainetdinov I, Zoch A, O’Carroll D, Zamore PD. PIWI-interacting RNAs: small RNAs with big functions. Nat Rev Genet. 2019;20(2):89–108. https://doi.org/10.1038/s41576-018-0073-3.
Valadkhan S, Gunawardane LS. Role of small nuclear RNAs in eukaryotic gene expression. Essays Biochem. 2013;54:79–90. https://doi.org/10.1042/bse0540079.
Kiss T. Small nucleolar RNAs: an abundant group of noncoding RNAs with diverse cellular functions. Cell. 2002;109(2):145–8. https://doi.org/10.1016/s0092-8674(02)00718-3.
Dahariya S, Paddibhatla I, Kumar S, Raghuwanshi S, Pallepati A, Gutti RK. Long non-coding RNA: classification, biogenesis and functions in blood cells. Mol Immunol. 2019;112:82–92. https://doi.org/10.1016/j.molimm.2019.04.011.
Gibb EA, Brown CJ, Lam WL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10:38. https://doi.org/10.1186/1476-4598-10-38.
Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–22. https://doi.org/10.1038/nrd.2016.246.
Guarnieri DJ, DiLeone RJ. MicroRNAs: a new class of gene regulators. Ann Med. 2008;40(3):197–208. https://doi.org/10.1080/07853890701771823.
Esquela-Kerscher A, Slack FJ. Oncomirs—microRNAs with a role in cancer. Nat Rev Cancer. 2006;6(4):259–69. https://doi.org/10.1038/nrc1840.
Yang RC, Xu BJ, Yang B, Fu JY, Chen HC, Wang XR. Non-coding RNAs: the extensive and interactive regulators of the blood–brain barrier permeability. RNA Biol. 2021. https://doi.org/10.1080/15476286.2021.1950465.
Yao RW, Wang Y, Chen LL. Cellular functions of long noncoding RNAs. Nat Cell Biol. 2019;21(5):542–51. https://doi.org/10.1038/s41556-019-0311-8.
Yu CY, Kuo HC. The emerging roles and functions of circular RNAs and their generation. J Biomed Sci. 2019;26. https://doi.org/10.1186/s12929-019-0523-z.
Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–91. https://doi.org/10.1038/s41576-019-0158-7.
Kristensen LS, Hansen TB, Veno MT, Kjems J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene. 2018;37(5):555–65. https://doi.org/10.1038/onc.2017.361.
Kadry H, Noorani B, Cucullo L. A blood–brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS. 2020;17(1):69. https://doi.org/10.1186/s12987-020-00230-3.
Liu WY, Wang ZB, Zhang LC, Wei X, Li L. Tight junction in blood–brain barrier: an overview of structure, regulation, and regulator substances. CNS Neurosci Ther. 2012;18(8):609–15. https://doi.org/10.1111/j.1755-5949.2012.00340.x.
Ballabh P, Braun A, Nedergaard M. The blood–brain barrier: an overview: structure, regulation, and clinical implications. Neurobiol Dis. 2004;16(1):1–13. https://doi.org/10.1016/j.nbd.2003.12.016.
Zlokovic BV. The blood–brain barrier in health and chronic neurodegenerative disorders. Neuron. 2008;57(2):178–201. https://doi.org/10.1016/j.neuron.2008.01.003.
Zlokovic BV. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat Rev Neurosci. 2011;12(12):723–38. https://doi.org/10.1038/nrn3114.
Zenaro E, Piacentino G, Constantin G. The blood–brain barrier in Alzheimer’s disease. Neurobiol Dis. 2017;107:41–56. https://doi.org/10.1016/j.nbd.2016.07.007.
Unzeta M, Hernandez-Guillamon M, Sun P, Sole M. SSAO/VAP-1 in cerebrovascular disorders: a potential therapeutic target for stroke and Alzheimer’s disease. Int J Mol Sci. 2021;22(7):3365. https://doi.org/10.3390/ijms22073365.
Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood–brain barrier. Neurobiol Dis. 2010;37(1):13–25. https://doi.org/10.1016/j.nbd.2009.07.030.
Greene C, Hanley N, Campbell M. Claudin-5: gatekeeper of neurological function. Fluids Barriers CNS. 2019;16(1):3. https://doi.org/10.1186/s12987-019-0123-z.
Jia W, Lu RC, Martin TA, Jiang WG. The role of claudin-5 in blood–brain barrier (BBB) and brain metastases (review). Mol Med Rep. 2014;9(3):779–85. https://doi.org/10.3892/mmr.2013.1875.
Lv JJ, Hu W, Yang Z, Li T, Jiang S, Ma ZQ, et al. Focusing on claudin-5: a promising candidate in the regulation of BBB to treat ischemic stroke. Prog Neurobiol. 2018;161:79–96. https://doi.org/10.1016/j.pneurobio.2017.12.001.
Greene C, Hanley N, Campbell M. Blood–brain barrier associated tight junction disruption is a hallmark feature of major psychiatric disorders. Transl Psychiat. 2020;10(1). https://doi.org/10.1038/s41398-020-01054-3.
Armulik A, Genove G, Betsholtz C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell. 2011;21(2):193–215. https://doi.org/10.1016/j.devcel.2011.07.001.
Winkler EA, Bell RD, Zlokovic BV. Central nervous system pericytes in health and disease. Nat Neurosci. 2011;14(11):1398–405. https://doi.org/10.1038/nn.2946.
Wolburg H, Noell S, Wolburg-Buchholz K, Mack A, Fallier-Becker P. Agrin, aquaporin-4, and astrocyte polarity as an important feature of the blood–brain barrier. Neuroscientist. 2009;15(2):180–93.
Bell RD, Zlokovic BV. Neurovascular mechanisms and blood–brain barrier disorder in Alzheimer’s disease. Acta Neuropathol. 2009;118(1):103–13. https://doi.org/10.1007/s00401-009-0522-3.
Iadecola C. The overlap between neurodegenerative and vascular factors in the pathogenesis of dementia. Acta Neuropathol. 2010;120(3):287–96.
Eyileten C, Wicik Z, De Rosa S, Mirowska-Guzel D, Soplinska A, Indolfi C, et al. MicroRNAs as diagnostic and prognostic biomarkers in ischemic stroke—a comprehensive review and bioinformatic analysis. Cells. 2018;7(12):249.
Martinez B, Peplow PV. Blood microRNAs as potential diagnostic markers for hemorrhagic stroke. Neural Regen Res. 2017;12(1):13–8. https://doi.org/10.4103/1673-5374.198965.
Atif H, Hicks SD. A review of MicroRNA biomarkers in traumatic brain injury. J Exp Neurosci. 2019;13:1179069519832286. https://doi.org/10.1177/1179069519832286.
Tigchelaar S, Gupta R, Shannon CP, Streijger F, Sinha S, Flibotte S, et al. MicroRNA biomarkers in cerebrospinal fluid and serum reflect injury severity in human acute traumatic spinal cord injury. J Neurotraum. 2019;36(15):2358–71. https://doi.org/10.1089/neu.2018.6256.
Huang SW, Ali ND, Zhong L, Shi J. MicroRNAs as biomarkers for human glioblastoma: progress and potential. Acta Pharmacol Sin. 2018;39(9):1405–13. https://doi.org/10.1038/aps.2017.173.
Martinez B, Peplow PV. MicroRNAs in blood and cerebrospinal fluid as diagnostic biomarkers of multiple sclerosis and to monitor disease progression. Neural Regen Res. 2020;15(4):606–19. https://doi.org/10.4103/1673-5374.266905.
Galimberti D, Villa C, Fenoglio C, Serpente M, Ghezzi L, Cioffi SMG, et al. Circulating miRNAs as potential biomarkers in Alzheimer’s disease. J Alzheimers Dis. 2014;42(4):1261–7. https://doi.org/10.3233/Jad-140756.
Zhang J, Sun P, Zhou C, Zhang XJ, Ma FF, Xu Y, et al. Regulatory microRNAs and vascular cognitive impairment and dementia. CNS Neurosci Ther. 2020;26(12):1207–18. https://doi.org/10.1111/cns.13472.
Fan JF, Saft M, Sadanandan N, Gonzales-Portillo B, Park YJ, Sanberg PR, et al. LncRNAs stand as potent biomarkers and therapeutic targets for stroke. Front Aging Neurosci. 2020;12. https://doi.org/10.3389/fnagi.2020.594571.
Li ZX, Han KW, Zhang DF, Chen JG, Xu Z, Hou LJ. The role of long noncoding RNA in traumatic brain injury. Neuropsych Dis Treat. 2019;15:1671–7. https://doi.org/10.2147/Ndt.S206624.
Wang F, Liu JZ, Wang XN, Chen JG, Kong QJ, Ye BG, et al. The emerging role of lncRNAs in spinal cord injury. Biomed Res Int. 2019;2019:1–90. https://doi.org/10.1155/2019/3467121.
Sun YZ, Wang Z, Zhou D. Long non-coding RNAs as potential biomarkers and therapeutic targets for gliomas. Med Hypotheses. 2013;81(2):319–21. https://doi.org/10.1016/j.mehy.2013.04.010.
Yang X, Wu YZ, Zhang B, Ni B. Noncoding RNAs in multiple sclerosis. Clin Epigenetics. 2018;10. https://doi.org/10.1186/s13148-018-0586-9.
Doxtater K, Tripathi MK, Khan MM. Recent advances on the role of long non-coding RNAs in Alzheimer’s disease. Neural Regen Res. 2020;15(12):2253–4. https://doi.org/10.4103/1673-5374.284990.
Zhang ZR, Yang TT, Xiao JJ. Circular RNAs: promising biomarkers for human diseases. EBioMedicine. 2018;34:267–74. https://doi.org/10.1016/j.ebiom.2018.07.036.
Wang Y, Huang J, Ma Y, Tang G, Liu Y, Chen X, et al. MicroRNA-29b is a therapeutic target in cerebral ischemia associated with aquaporin 4. J Cereb Blood Flow Metab. 2015;35(12):1977–84.
Zhang H, Pan Q, Xie Z, Chen Y, Wang J, Bihl J, et al. Implication of MicroRNA503 in brain endothelial cell function and ischemic stroke. Transl Stroke Res. 2020;11(5):1148–64. https://doi.org/10.1007/s12975-020-00794-0.
Wang MD, Wang Y, Xia YP, Dai JW, Gao L, Wang SQ, et al. High serum MiR-130a levels are associated with severe perihematomal edema and predict adverse outcome in acute ICH. Mol Neurobiol. 2016;53(2):1310–21. https://doi.org/10.1007/s12035-015-9099-0.
Awad H, Bratasz A, Nuovo G, Burry R, Meng X, Kelani H, et al. MiR-155 deletion reduces ischemia-induced paralysis in an aortic aneurysm repair mouse model: utility of immunohistochemistry and histopathology in understanding etiology of spinal cord paralysis. Ann Diagn Pathol. 2018;36:12–20. https://doi.org/10.1016/j.anndiagpath.2018.06.002.
Elkhodiry AA, Zamzam DA, El Tayebi HM. miR-155 and functional proteins of CD8+T cells as potential prognostic biomarkers for relapsing-remitting multiple sclerosis. Mult Scler Relat Dis. 2021;53:103078. https://doi.org/10.1016/j.msard.2021.103078.
Zhou W, Fong MY, Min Y, Somlo G, Liu L, Palomares MR, et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014;25(4):501–15. https://doi.org/10.1016/j.ccr.2014.03.007.
Wu Q, Yuan X, Bai J, Han R, Li Z, Zhang H, et al. MicroRNA-181a protects against pericyte apoptosis via directly targeting FOXO1: implication for ameliorated cognitive deficits in APP/PS1 mice. Aging (Albany NY). 2019;11(16):6120–33. https://doi.org/10.18632/aging.102171.
Visitchanakun P, Tangtanatakul P, Trithiphen O, Soonthornchai W, Wongphoom J, Tachaboon S, et al. Plasma miR-370-3p as a biomarker of sepsis-associated encephalopathy, the transcriptomic profiling analysis of microRNA-arrays from mouse brains. Shock. 2020;54(3):347–57.
Wang C, Dong J, Sun J, Huang S, Wu F, Zhang X, et al. Silencing of lncRNA XIST impairs angiogenesis and exacerbates cerebral vascular injury after ischemic stroke. Mol Ther Nucleic Acids. 2021;26:148–60. https://doi.org/10.1016/j.omtn.2021.06.025.
Wang S, Liang K, Hu Q, Li P, Song J, Yang Y, et al. JAK2-binding long noncoding RNA promotes breast cancer brain metastasis. J Clin Invest. 2017;127(12):4498–515. https://doi.org/10.1172/JCI91553.
Bai Y, Zhang Y, Han B, Yang L, Chen X, Huang R, et al. Circular RNA DLGAP4 ameliorates ischemic stroke outcomes by targeting miR-143 to regulate endothelial–mesenchymal transition associated with blood–brain barrier integrity. J Neurosci. 2018;38(1):32–50. https://doi.org/10.1523/JNEUROSCI.1348-17.2017.
Yin KJ, Deng Z, Hamblin M, Xiang Y, Huang H, Zhang J, et al. Peroxisome proliferator-activated receptor delta regulation of miR-15a in ischemia-induced cerebral vascular endothelial injury. J Neurosci. 2010;30(18):6398–408. https://doi.org/10.1523/JNEUROSCI.0780-10.2010.
Ma F, Sun P, Zhang X, Hamblin MH, Yin KJ. Endothelium-targeted deletion of the miR-15a/16-1 cluster ameliorates blood–brain barrier dysfunction in ischemic stroke. Sci Signal. 2020;13(626). https://doi.org/10.1126/scisignal.aay5686.
Sun P, Zhang K, Hassan SH, Zhang X, Tang X, Pu H, et al. Endothelium-targeted deletion of microRNA-15a/16-1 promotes poststroke angiogenesis and improves long-term neurological recovery. Circ Res. 2020;126(8):1040–57. https://doi.org/10.1161/CIRCRESAHA.119.315886.
Bukeirat M, Sarkar SN, Hu H, Quintana DD, Simpkins JW, Ren X. MiR-34a regulates blood–brain barrier permeability and mitochondrial function by targeting cytochrome c. J Cereb Blood Flow Metab. 2016;36(2):387–92. https://doi.org/10.1177/0271678X15606147.
Ren X, Engler-Chiurazzi EB, Russell AE, Sarkar SN, Rellick SL, Lewis S, et al. MiR-34a and stroke: assessment of non-modifiable biological risk factors in cerebral ischemia. Neurochem Int. 2019;127:73–9. https://doi.org/10.1016/j.neuint.2018.10.019.
Hu H, Hone EA, Provencher EAP, Sprowls SA, Farooqi I, Corbin DR, et al. MiR-34a interacts with cytochrome c and shapes stroke outcomes. Sci Rep. 2020;10(1):3233. https://doi.org/10.1038/s41598-020-59997-y.
Fang Z, He QW, Li Q, Chen XL, Baral S, Jin HJ, et al. MicroRNA-150 regulates blood–brain barrier permeability via Tie-2 after permanent middle cerebral artery occlusion in rats. FASEB J. 2016;30(6):2097–107. https://doi.org/10.1096/fj.201500126.
da Liu Z, Jickling GC, Ander BP, Hull H, Zhan X, Cox C, et al. Elevating microRNA-122 in blood improves outcomes after temporary middle cerebral artery occlusion in rats. J Cereb Blood Flow Metab. 2016;36(8):1374–83. https://doi.org/10.1177/0271678X15610786.
Ma Q, Dasgupta C, Li Y, Huang L, Zhang L. MicroRNA-210 suppresses junction proteins and disrupts blood–brain barrier integrity in neonatal rat hypoxic-ischemic brain injury. Int J Mol Sci. 2017;18(7):1356. https://doi.org/10.3390/ijms18071356.
Yao X, Wang Y, Zhang D. microRNA-21 confers neuroprotection against cerebral ischemia-reperfusion injury and alleviates blood–brain barrier disruption in rats via the MAPK signaling pathway. J Mol Neurosci. 2018;65(1):43–53. https://doi.org/10.1007/s12031-018-1067-5.
Fan F, Yang J, Xu Y, Guan S. MiR-539 targets MMP-9 to regulate the permeability of blood–brain barrier in ischemia/reperfusion injury of brain. Neurochem Res. 2018;43(12):2260–7. https://doi.org/10.1007/s11064-018-2646-0.
Pena-Philippides JC, Gardiner AS, Caballero-Garrido E, Pan R, Zhu Y, Roitbak T. Inhibition of MicroRNA-155 supports endothelial tight junction integrity following oxygen-glucose deprivation. J Am Heart Assoc. 2018;7(13). https://doi.org/10.1161/JAHA.118.009244.
Suofu Y, Wang X, He Y, Li F, Zhang Y, Carlisle DL, et al. Mir-155 knockout protects against ischemia/reperfusion-induced brain injury and hemorrhagic transformation. NeuroReport. 2020;31(3):235–9. https://doi.org/10.1097/WNR.0000000000001382.
Wan Y, Jin HJ, Zhu YY, Fang Z, Mao L, He Q, et al. MicroRNA-149-5p regulates blood–brain barrier permeability after transient middle cerebral artery occlusion in rats by targeting S1PR2 of pericytes. FASEB J. 2018;32(6):3133–48. https://doi.org/10.1096/fj.201701121R.
Talebi A, Rahnema M, Bigdeli MR. Effect of intravenous injection of antagomiR-1 on brain ischemia. Mol Biol Rep. 2019;46(1):1149–55. https://doi.org/10.1007/s11033-018-04580-y.
Zuo X, Lu J, Manaenko A, Qi X, Tang J, Mei Q, et al. MicroRNA-132 attenuates cerebral injury by protecting blood–brain-barrier in MCAO mice. Exp Neurol. 2019;316:12–9. https://doi.org/10.1016/j.expneurol.2019.03.017.
Bernstein DL, Rom S. Let-7g* and miR-98 Reduce stroke-induced production of proinflammatory cytokines in mouse brain. Front Cell Dev Biol. 2020;8:632. https://doi.org/10.3389/fcell.2020.00632.
Bernstein DL, Gajghate S, Reichenbach NL, Winfield M, Persidsky Y, Heldt NA, et al. let-7g counteracts endothelial dysfunction and ameliorating neurological functions in mouse ischemia/reperfusion stroke model. Brain Behav Immun. 2020;87:543–55. https://doi.org/10.1016/j.bbi.2020.01.026.
Pan J, Qu M, Li Y, Wang L, Zhang L, Wang Y, et al. MicroRNA-126-3p/-5p overexpression attenuates blood–brain barrier disruption in a mouse model of middle cerebral artery occlusion. Stroke. 2020;51(2):619–27. https://doi.org/10.1161/STROKEAHA.119.027531.
Bernstein DL, Zuluaga-Ramirez V, Gajghate S, Reichenbach NL, Polyak B, Persidsky Y, et al. miR-98 reduces endothelial dysfunction by protecting blood–brain barrier (BBB) and improves neurological outcomes in mouse ischemia/reperfusion stroke model. J Cereb Blood Flow Metab. 2020;40(10):1953–65. https://doi.org/10.1177/0271678X19882264.
He J, Zhang X. miR-668 inhibitor attenuates mitochondrial membrane potential and protects against neuronal apoptosis in cerebral ischemic stroke. Folia Neuropathol. 2020;58(1):22–9. https://doi.org/10.5114/fn.2020.94003.
Zhang T, Tian C, Wu J, Zhang Y, Wang J, Kong Q, et al. MicroRNA-182 exacerbates blood–brain barrier (BBB) disruption by downregulating the mTOR/FOXO1 pathway in cerebral ischemia. FASEB J. 2020;34(10):13762–75. https://doi.org/10.1096/fj.201903092R.
Zhao Z, Wu C, He X, Zhao E, Hu S, Han Y, et al. MicroRNA let-7f alleviates vascular endothelial cell dysfunction via targeting HMGA2 under oxygen-glucose deprivation and reoxygenation. Brain Res. 2021;1772: 147662. https://doi.org/10.1016/j.brainres.2021.147662.
Wang P, Pan R, Weaver J, Jia M, Yang X, Yang T, et al. MicroRNA-30a regulates acute cerebral ischemia-induced blood–brain barrier damage through ZnT4/zinc pathway. J Cereb Blood Flow Metab. 2021;41(3):641–55. https://doi.org/10.1177/0271678X20926787.
Zhao B, Wang P, Yu J, Zhang Y. MicroRNA-376b-5p targets SOX7 to alleviate ischemic brain injury in a mouse model through activating Wnt/beta-catenin signaling pathway. Life Sci. 2021;270: 119072. https://doi.org/10.1016/j.lfs.2021.119072.
Ghasemloo E, Oryan S, Bigdeli MR, Mostafavi H, Eskandari M. The neuroprotective effect of MicroRNA-149-5p and coenzymeQ10 by reducing levels of inflammatory cytokines and metalloproteinases following focal brain ischemia in rats. Brain Res Bull. 2021;169:205–13. https://doi.org/10.1016/j.brainresbull.2021.01.013.
Shen J, Li G, Zhu Y, Xu Q, Zhou H, Xu K, et al. Foxo1-induced miR-92b down-regulation promotes blood–brain barrier damage after ischaemic stroke by targeting NOX4. J Cell Mol Med. 2021;25(11):5269–82. https://doi.org/10.1111/jcmm.16537.
Dhuri K, Vyas RN, Blumenfeld L, Verma R, Bahal R. Nanoparticle delivered Anti-miR-141–3p for stroke therapy. Cells. 2021;10(5):1011. https://doi.org/10.3390/cells10051011.
Jeyaseelan K, Lim KY, Armugam A. MicroRNA expression in the blood and brain of rats subjected to transient focal ischemia by middle cerebral artery occlusion. Stroke. 2008;39(3):959–66. https://doi.org/10.1161/STROKEAHA.107.500736.
Tsai PC, Liao YC, Wang YS, Lin HF, Lin RT, Juo SH. Serum microRNA-21 and microRNA-221 as potential biomarkers for cerebrovascular disease. J Vasc Res. 2013;50(4):346–54. https://doi.org/10.1159/000351767.
Panagal M, Biruntha M, Vidhyavathi RM, Sivagurunathan P, Senthilkumar SR, Sekar D. Dissecting the role of miR-21 in different types of stroke. Gene. 2019;681:69–72. https://doi.org/10.1016/j.gene.2018.09.048.
Li Z, Li J, Tang N. Long noncoding RNA Malat1 is a potent autophagy inducer protecting brain microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury by sponging miR-26b and upregulating ULK2 expression. Neuroscience. 2017;354:1–10. https://doi.org/10.1016/j.neuroscience.2017.04.017.
Zhang X, Tang X, Liu K, Hamblin MH, Yin KJ. Long Noncoding RNA Malat1 regulates cerebrovascular pathologies in ischemic stroke. J Neurosci. 2017;37(7):1797–806. https://doi.org/10.1523/JNEUROSCI.3389-16.2017.
Wang H, Zheng X, Jin J, Zheng L, Guan T, Huo Y, et al. LncRNA MALAT1 silencing protects against cerebral ischemia-reperfusion injury through miR-145 to regulate AQP4. J Biomed Sci. 2020;27(1):40. https://doi.org/10.1186/s12929-020-00635-0.
Wu L, Ye Z, Pan Y, Li X, Fu X, Zhang B, et al. Vascular endothelial growth factor aggravates cerebral ischemia and reperfusion-induced blood–brain-barrier disruption through regulating LOC102640519/HOXC13/ZO-1 signaling. Exp Cell Res. 2018;369(2):275–83. https://doi.org/10.1016/j.yexcr.2018.05.029.
You D, You H. Repression of long non-coding RNA MEG3 restores nerve growth and alleviates neurological impairment after cerebral ischemia-reperfusion injury in a rat model. Biomed Pharmacother. 2019;111:1447–57. https://doi.org/10.1016/j.biopha.2018.12.067.
Tian J, Liu Y, Wang Z, Zhang S, Yang Y, Zhu Y, et al. LncRNA Snhg8 attenuates microglial inflammation response and blood–brain barrier damage in ischemic stroke through regulating miR-425-5p mediated SIRT1/NF-kappaB signaling. J Biochem Mol Toxicol. 2021;35(5): e22724. https://doi.org/10.1002/jbt.22724.
Zhang J, Yuan L, Zhang X, Hamblin MH, Zhu T, Meng F, et al. Altered long non-coding RNA transcriptomic profiles in brain microvascular endothelium after cerebral ischemia. Exp Neurol. 2016;277:162–70. https://doi.org/10.1016/j.expneurol.2015.12.014.
Wu L, Xu H, Zhang W, Chen Z, Li W, Ke W. Circular RNA circCCDC9 alleviates ischaemic stroke ischaemia/reperfusion injury via the Notch pathway. J Cell Mol Med. 2020;24(24):14152–9. https://doi.org/10.1111/jcmm.16025.
Ouyang Y, Li D, Wang H, Wan Z, Luo Q, Zhong Y, et al. MiR-21-5p/dual-specificity phosphatase 8 signalling mediates the anti-inflammatory effect of haem oxygenase-1 in aged intracerebral haemorrhage rats. Aging Cell. 2019;18(6): e13022. https://doi.org/10.1111/acel.13022.
Wang L, Zhao Y, Gang S, Geng T, Li M, Xu L, et al. Inhibition of miR-103-3p preserves neurovascular integrity through caveolin-1 in experimental subarachnoid hemorrhage. Neuroscience. 2021;461:91–101. https://doi.org/10.1016/j.neuroscience.2021.03.007.
Deng X, Liang C, Qian L, Zhang Q. miR-24 targets HMOX1 to regulate inflammation and neurofunction in rats with cerebral vasospasm after subarachnoid hemorrhage. Am J Transl Res. 2021;13(3):1064–74.
Xi T, Jin F, Zhu Y, Wang J, Tang L, Wang Y, et al. MicroRNA-126-3p attenuates blood–brain barrier disruption, cerebral edema and neuronal injury following intracerebral hemorrhage by regulating PIK3R2 and Akt. Biochem Biophys Res Commun. 2017;494(1–2):144–51. https://doi.org/10.1016/j.bbrc.2017.10.064.
Fu X, Niu T, Li X. MicroRNA-126-3p attenuates intracerebral hemorrhage-induced blood–brain barrier disruption by regulating VCAM-1 expression. Front Neurosci. 2019;13:866. https://doi.org/10.3389/fnins.2019.00866.
Wang C, Cao J, Duan S, Xu R, Yu H, Huo X, et al. Effect of MicroRNA-126a-3p on bone marrow mesenchymal stem cells repairing blood–brain barrier and nerve injury after intracerebral hemorrhage. J Stroke Cerebrovasc Dis. 2020;29(5): 104748. https://doi.org/10.1016/j.jstrokecerebrovasdis.2020.104748.
Xi T, Jin F, Zhu Y, Wang J, Tang L, Wang Y, et al. miR-27a-3p protects against blood–brain barrier disruption and brain injury after intracerebral hemorrhage by targeting endothelial aquaporin-11. J Biol Chem. 2018;293(52):20041–50. https://doi.org/10.1074/jbc.RA118.001858.
Lai N, Wu D, Liang T, Pan P, Yuan G, Li X, et al. Systemic exosomal miR-193b-3p delivery attenuates neuroinflammation in early brain injury after subarachnoid hemorrhage in mice. J Neuroinflammation. 2020;17(1):74. https://doi.org/10.1186/s12974-020-01745-0.
Zhao B, Qian M, Zhang Y, Yin F. Stem cells from human exfoliated deciduous teeth transmit microRNA-26a to protect rats with experimental intracerebral hemorrhage from cerebral injury via suppressing CTGF. Brain Res Bull. 2021;168:146–55. https://doi.org/10.1016/j.brainresbull.2020.12.006.
Zhang J, Dong B, Hao J, Yi S, Cai W, Luo Z. LncRNA Snhg3 contributes to dysfunction of cerebral microvascular cells in intracerebral hemorrhage rats by activating the TWEAK/Fn14/STAT3 pathway. Life Sci. 2019;237: 116929. https://doi.org/10.1016/j.lfs.2019.116929.
Xie L, Wang Y, Chen Z. LncRNA Blnc1 mediates the permeability and inflammatory response of cerebral hemorrhage by regulating the PPAR-gamma/SIRT6/FoxO3 pathway. Life Sci. 2021;267: 118942. https://doi.org/10.1016/j.lfs.2020.118942.
Cai L, Ge B, Xu S, Chen X, Yang H. Up-regulation of circARF3 reduces blood–brain barrier damage in rat subarachnoid hemorrhage model via miR-31-5p/MyD88/NF-kappaB axis. Aging (Albany NY). 2021;13(17):21345–63. https://doi.org/10.18632/aging.203468.
Price L, Wilson C, Grant G. Blood–brain barrier pathophysiology following traumatic brain injury. In: Laskowitz D, Grant G, editors. Translational research in traumatic brain injury. Front Neurosci. Boca Raton (FL) 2016.
Ge XT, Lei P, Wang HC, Zhang AL, Han ZL, Chen X, et al. miR-21 improves the neurological outcome after traumatic brain injury in rats. Sci Rep. 2014;4:6718. https://doi.org/10.1038/srep06718.
Ge X, Han Z, Chen F, Wang H, Zhang B, Jiang R, et al. MiR-21 alleviates secondary blood–brain barrier damage after traumatic brain injury in rats. Brain Res. 2015;1603:150–7. https://doi.org/10.1016/j.brainres.2015.01.009.
Ge X, Huang S, Gao H, Han Z, Chen F, Zhang S, et al. miR-21-5p alleviates leakage of injured brain microvascular endothelial barrier in vitro through suppressing inflammation and apoptosis. Brain Res. 2016;1650:31–40. https://doi.org/10.1016/j.brainres.2016.07.015.
Wu J, He J, Tian X, Luo Y, Zhong J, Zhang H, et al. microRNA-9-5p alleviates blood–brain barrier damage and neuroinflammation after traumatic brain injury. J Neurochem. 2020;153(6):710–26. https://doi.org/10.1111/jnc.14963.
Liu N, Sun H, Li X, Cao W, Peng A, Dong S, et al. Downregulation of lncRNA KCNQ1OT1 relieves traumatic brain injury induced neurological deficits via promoting “M2” microglia polarization. Brain Res Bull. 2021;171:91–102. https://doi.org/10.1016/j.brainresbull.2021.03.004.
Cheng YQ, Wu CR, Du MR, Zhou Q, Wu BY, Fu JY, et al. CircLphn3 protects the blood–brain barrier in traumatic brain injury. Neural Regen Res. 2022;17(4):812–8. https://doi.org/10.4103/1673-5374.322467.
Bartanusz V, Jezova D, Alajajian B, Digicaylioglu M. The blood-spinal cord barrier: morphology and clinical implications. Ann Neurol. 2011;70(2):194–206. https://doi.org/10.1002/ana.22421.
Whetstone WD, Hsu JY, Eisenberg M, Werb Z, Noble-Haeusslein LJ. Blood-spinal cord barrier after spinal cord injury: relation to revascularization and wound healing. J Neurosci Res. 2003;74(2):227–39. https://doi.org/10.1002/jnr.10759.
Li XQ, Lv HW, Wang ZL, Tan WF, Fang B, Ma H. MiR-27a ameliorates inflammatory damage to the blood-spinal cord barrier after spinal cord ischemia: reperfusion injury in rats by downregulating TICAM-2 of the TLR4 signaling pathway. J Neuroinflammation. 2015;12:25. https://doi.org/10.1186/s12974-015-0246-3.
Li XQ, Chen FS, Tan WF, Fang B, Zhang ZL, Ma H. Elevated microRNA-129-5p level ameliorates neuroinflammation and blood-spinal cord barrier damage after ischemia-reperfusion by inhibiting HMGB1 and the TLR3-cytokine pathway. J Neuroinflammation. 2017;14(1):205. https://doi.org/10.1186/s12974-017-0977-4.
Bao N, Fang B, Lv H, Jiang Y, Chen F, Wang Z, et al. Upregulation of miR-199a-5p protects spinal cord against ischemia/reperfusion-induced injury via downregulation of ECE1 in Rat. Cell Mol Neurobiol. 2018;38(6):1293–303. https://doi.org/10.1007/s10571-018-0597-2.
Wang D, Chen F, Fang B, Zhang Z, Dong Y, Tong X, et al. MiR-128-3p alleviates spinal cord ischemia/reperfusion injury associated neuroinflammation and cellular apoptosis via SP1 suppression in rat. Front Neurosci. 2020;14: 609613. https://doi.org/10.3389/fnins.2020.609613.
Chen F, Han J, Li X, Zhang Z, Wang D. Identification of the biological function of miR-9 in spinal cord ischemia-reperfusion injury in rats. PeerJ. 2021;9: e11440. https://doi.org/10.7717/peerj.11440.
Wang J, Nie Z, Zhao H, Gao K, Cao Y. MiRNA-125a-5p attenuates blood-spinal cord barrier permeability under hypoxia in vitro. Biotechnol Lett. 2020;42(1):25–34. https://doi.org/10.1007/s10529-019-02753-8.
Ge X, Tang P, Rong Y, Jiang D, Lu X, Ji C, et al. Exosomal miR-155 from M1-polarized macrophages promotes EndoMT and impairs mitochondrial function via activating NF-kappaB signaling pathway in vascular endothelial cells after traumatic spinal cord injury. Redox Biol. 2021;41: 101932. https://doi.org/10.1016/j.redox.2021.101932.
Sun R, Ge L, Cao Y, Wu W, Wu Y, Zhu H, et al. MiR-429 regulates blood-spinal cord barrier permeability by targeting Kruppel-like factor 6. Biochem Biophys Res Commun. 2020;525(3):740–6. https://doi.org/10.1016/j.bbrc.2020.02.138.
Yu D, Sun R, Shen D, Ge L, Xue T, Cao Y. Nuclear heme oxygenase-1 improved the hypoxia-mediated dysfunction of blood-spinal cord barrier via the miR-181c-5p/SOX5 signaling pathway. NeuroReport. 2021;32(2):112–20. https://doi.org/10.1097/WNR.0000000000001568.
Jia H, Li Z, Chang Y, Fang B, Zhou Y, Ma H. Downregulation of long noncoding RNA TUG1 attenuates MTDH-mediated inflammatory damage via targeting miR-29b-1-5p after spinal cord ischemia reperfusion. J Neuropathol Exp Neurol. 2021;80(3):254–64. https://doi.org/10.1093/jnen/nlaa138.
Wang WZ, Li J, Liu L, Zhang ZD, Li MX, Li Q, et al. Role of circular RNA expression in the pathological progression after spinal cord injury. Neural Regen Res. 2021;16(10):2048–55. https://doi.org/10.4103/1673-5374.308100.
Francis SS, Ostrom QT, Cote DJ, Smith TR, Claus E, Barnholtz-Sloan JS. The epidemiology of central nervous system tumors. Hematol Oncol Clin North Am. 2022;36(1):23–42. https://doi.org/10.1016/j.hoc.2021.08.012.
Nieblas-Bedolla E, Zuccato J, Kluger H, Zadeh G, Brastianos PK. Central nervous system metastases. Hematol Oncol Clin North Am. 2022;36(1):161–88. https://doi.org/10.1016/j.hoc.2021.08.004.
Arvanitis CD, Ferraro GB, Jain RK. The blood–brain barrier and blood-tumour barrier in brain tumours and metastases. Nat Rev Cancer. 2020;20(1):26–41. https://doi.org/10.1038/s41568-019-0205-x.
Ma J, Yao Y, Wang P, Liu Y, Zhao L, Li Z, et al. MiR-181a regulates blood-tumor barrier permeability by targeting Kruppel-like factor 6. J Cereb Blood Flow Metab. 2014;34(11):1826–36. https://doi.org/10.1038/jcbfm.2014.152.
Zhao L, Wang P, Liu Y, Ma J, Xue Y. miR-34c regulates the permeability of blood-tumor barrier via MAZ-mediated expression changes of ZO-1, occludin, and claudin-5. J Cell Physiol. 2015;230(3):716–31. https://doi.org/10.1002/jcp.24799.
Zhao YY, Zhao LN, Wang P, Miao YS, Liu YH, Wang ZH, et al. Overexpression of miR-18a negatively regulates myocyte enhancer factor 2D to increase the permeability of the blood-tumor barrier via Kruppel-like factor 4-mediated downregulation of zonula occluden-1, claudin-5, and occludin. J Neurosci Res. 2015;93(12):1891–902. https://doi.org/10.1002/jnr.23628.
Miao YS, Zhao YY, Zhao LN, Wang P, Liu YH, Ma J, et al. MiR-18a increased the permeability of BTB via RUNX1 mediated down-regulation of ZO-1, occludin and claudin-5. Cell Signal. 2015;27(1):156–67. https://doi.org/10.1016/j.cellsig.2014.10.008.
Zhao W, Wang P, Ma J, Liu YH, Li Z, Li ZQ, et al. MiR-34a regulates blood-tumor barrier function by targeting protein kinase Cepsilon. Mol Biol Cell. 2015;26(10):1786–96. https://doi.org/10.1091/mbc.E14-10-1474.
Ma T, Xue YX. MiRNA-200b regulates RMP7-induced increases in blood-tumor barrier permeability by targeting RhoA and ROCKII. Front Mol Neurosci. 2016;9:9. https://doi.org/10.3389/fnmol.2016.00009.
Liu J, Liu L, Chao S, Liu Y, Liu X, Zheng J, et al. The Role of miR-330-3p/PKC-alpha signaling pathway in low-dose endothelial-monocyte activating polypeptide-II increasing the permeability of blood-tumor barrier. Front Cell Neurosci. 2017;11:358. https://doi.org/10.3389/fncel.2017.00358.
Leng X, Ma J, Liu Y, Shen S, Yu H, Zheng J, et al. Mechanism of piR-DQ590027/MIR17HG regulating the permeability of glioma conditioned normal BBB. J Exp Clin Cancer Res. 2018;37(1):246. https://doi.org/10.1186/s13046-018-0886-0.
Chen L, Xue Y, Zheng J, Liu X, Liu J, Chen J, et al. MiR-429 regulated by endothelial monocyte activating polypeptide-II (EMAP-II) influences blood-tumor barrier permeability by inhibiting the expressions of ZO-1, Occludin and Claudin-5. Front Mol Neurosci. 2018;11:35. https://doi.org/10.3389/fnmol.2018.00035.
Shen S, Yu H, Liu X, Liu Y, Zheng J, Wang P, et al. PIWIL1/piRNA-DQ593109 regulates the permeability of the blood-tumor barrier via the MEG3/miR-330-5p/RUNX3 axis. Mol Ther Nucleic Acids. 2018;10:412–25. https://doi.org/10.1016/j.omtn.2017.12.020.
Gu Y, Cai R, Zhang C, Xue Y, Pan Y, Wang J, et al. miR-132-3p boosts caveolae-mediated transcellular transport in glioma endothelial cells by targeting PTEN/PI3K/PKB/Src/Cav-1 signaling pathway. FASEB J. 2019;33(1):441–54. https://doi.org/10.1096/fj.201800095RR.
Tominaga N, Kosaka N, Ono M, Katsuda T, Yoshioka Y, Tamura K, et al. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood–brain barrier. Nat Commun. 2015;6:6716. https://doi.org/10.1038/ncomms7716.
Xing F, Sharma S, Liu Y, Mo YY, Wu K, Zhang YY, et al. miR-509 suppresses brain metastasis of breast cancer cells by modulating RhoC and TNF-alpha. Oncogene. 2015;34(37):4890–900. https://doi.org/10.1038/onc.2014.412.
Harati R, Mohammad MG, Tlili A, El-Awady RA, Hamoudi R. Loss of miR-101-3p promotes transmigration of metastatic breast cancer cells through the brain endothelium by inducing COX-2/MMP1 signaling. Pharmaceuticals (Basel). 2020;13(7):144. https://doi.org/10.3390/ph13070144.
Pan JK, Lin CH, Kuo YL, Ger LP, Cheng HC, Yao YC, et al. MiR-211 determines brain metastasis specificity through SOX11/NGN2 axis in triple-negative breast cancer. Oncogene. 2021;40(9):1737–51. https://doi.org/10.1038/s41388-021-01654-3.
Cai H, Xue Y, Wang P, Wang Z, Li Z, Hu Y, et al. The long noncoding RNA TUG1 regulates blood-tumor barrier permeability by targeting miR-144. Oncotarget. 2015;6(23):19759–79. https://doi.org/10.18632/oncotarget.4331.
Cai H, Liu X, Zheng J, Xue Y, Ma J, Li Z, et al. Long non-coding RNA taurine upregulated 1 enhances tumor-induced angiogenesis through inhibiting microRNA-299 in human glioblastoma. Oncogene. 2017;36(3):318–31. https://doi.org/10.1038/onc.2016.212.
Ma J, Wang P, Yao Y, Liu Y, Li Z, Liu X, et al. Knockdown of long non-coding RNA MALAT1 increases the blood-tumor barrier permeability by up-regulating miR-140. Biochim Biophys Acta. 2016;1859(2):324–38. https://doi.org/10.1016/j.bbagrm.2015.11.008.
Kim SS, Harford JB, Moghe M, Rait A, Pirollo KF, Chang EH. Targeted nanocomplex carrying siRNA against MALAT1 sensitizes glioblastoma to temozolomide. Nucleic Acids Res. 2018;46(3):1424–40. https://doi.org/10.1093/nar/gkx1221.
Yu H, Xue Y, Wang P, Liu X, Ma J, Zheng J, et al. Knockdown of long non-coding RNA XIST increases blood-tumor barrier permeability and inhibits glioma angiogenesis by targeting miR-137. Oncogenesis. 2017;6(3): e303. https://doi.org/10.1038/oncsis.2017.7.
Guo J, Cai H, Zheng J, Liu X, Liu Y, Ma J, et al. Long non-coding RNA NEAT1 regulates permeability of the blood-tumor barrier via miR-181d-5p-mediated expression changes in ZO-1, occludin, and claudin-5. Biochim Biophys Acta Mol Basis Dis. 2017;1863(9):2240–54. https://doi.org/10.1016/j.bbadis.2017.02.005.
Sa L, Li Y, Zhao L, Liu Y, Wang P, Liu L, et al. The Role of HOTAIR/miR-148b-3p/USF1 on regulating the permeability of BTB. Front Mol Neurosci. 2017;10:194. https://doi.org/10.3389/fnmol.2017.00194.
Guo J, Shen S, Liu X, Ruan X, Zheng J, Liu Y, et al. Role of linc00174/miR-138-5p (miR-150-5p)/FOSL2 feedback loop on regulating the blood-tumor barrier permeability. Mol Ther Nucleic Acids. 2019;18:1072–90. https://doi.org/10.1016/j.omtn.2019.10.031.
Zhang C, Zhang X, Wang J, Di F, Xue Y, Lin X, et al. Lnc00462717 regulates the permeability of the blood–brain tumor barrier through interaction with PTBP1 to inhibit the miR-186-5p/Occludin signaling pathway. FASEB J. 2020;34(8):9941–58. https://doi.org/10.1096/fj.202000045R.
Li D, Li L, Chen X, Zhou C, Hao B, Cao Y. Dysregulation of lncRNA-CCRR contributes to brain metastasis of breast cancer by intercellular coupling via regulating connexin 43 expression. J Cell Mol Med. 2021;25(10):4826–34. https://doi.org/10.1111/jcmm.16455.
Lu Y, Chen L, Li L, Cao Y. Exosomes derived from brain metastatic breast cancer cells destroy the blood–brain barrier by carrying lncRNA GS1-600G8.5. Biomed Res Int. 2020;2020:7461727. https://doi.org/10.1155/2020/7461727.
He J, Xue Y, Wang Q, Zhou X, Liu L, Zhang T, et al. Long non-coding RNA MIAT regulates blood tumor barrier permeability by functioning as a competing endogenous RNA. Cell Death Dis. 2020;11(10):936. https://doi.org/10.1038/s41419-020-03134-0.
Gao Y, Wu P, Ma Y, Xue Y, Liu Y, Zheng J, et al. Circular RNA USP1 regulates the permeability of blood-tumour barrier via miR-194-5p/FLI1 axis. J Cell Mol Med. 2020;24(1):342–55. https://doi.org/10.1111/jcmm.14735.
Li H, Shen S, Ruan X, Liu X, Zheng J, Liu Y, et al. Biosynthetic CircRNA_001160 induced by PTBP1 regulates the permeability of BTB via the CircRNA_001160/miR-195-5p/ETV1 axis. Cell Death Dis. 2019;10(12):960. https://doi.org/10.1038/s41419-019-2191-z.
Wu P, Gao Y, Shen S, Xue Y, Liu X, Ruan X, et al. KHDRBS3 regulates the permeability of blood-tumor barrier via cDENND4C/miR-577 axis. Cell Death Dis. 2019;10(7):536. https://doi.org/10.1038/s41419-019-1771-2.
Reijerkerk A, Lopez-Ramirez MA, van Het Hof B, Drexhage JA, Kamphuis WW, Kooij G, et al. MicroRNAs regulate human brain endothelial cell-barrier function in inflammation: implications for multiple sclerosis. J Neurosci. 2013;33(16):6857–63. https://doi.org/10.1523/JNEUROSCI.3965-12.2013.
Aung LL, Mouradian MM, Dhib-Jalbut S, Balashov KE. MMP-9 expression is increased in B lymphocytes during multiple sclerosis exacerbation and is regulated by microRNA-320a. J Neuroimmunol. 2015;278:185–9.
Cerutti C, Edwards LJ, de Vries HE, Sharrack B, Male DK, Romero IA. MiR-126 and miR-126* regulate shear-resistant firm leukocyte adhesion to human brain endothelium. Sci Rep. 2017;7:45284. https://doi.org/10.1038/srep45284.
Maciak K, Dziedzic A, Miller E, Saluk-Bijak J. MiR-155 as an important regulator of multiple sclerosis pathogenesis. A review. Int J Mol Sci. 2021;22(9):4332.
Lopez-Ramirez MA, Wu D, Pryce G, Simpson JE, Reijerkerk A, King-Robson J, et al. MicroRNA-155 negatively affects blood–brain barrier function during neuroinflammation. FASEB J. 2014;28(6):2551–65. https://doi.org/10.1096/fj.13-248880.
Cerutti C, Soblechero-Martin P, Wu D, Lopez-Ramirez MA, de Vries H, Sharrack B, et al. MicroRNA-155 contributes to shear-resistant leukocyte adhesion to human brain endothelium in vitro. Fluids Barriers CNS. 2016;13(1):8. https://doi.org/10.1186/s12987-016-0032-3.
Liu X, Zhou F, Wang W, Chen G, Zhang Q, Lv R, et al. IL-9-triggered lncRNA Gm13568 regulates Notch1 in astrocytes through interaction with CBP/P300: contribute to the pathogenesis of experimental autoimmune encephalomyelitis. J Neuroinflammation. 2021;18(1):1–15.
Yang L, Han B, Zhang Y, Bai Y, Chao J, Hu G, et al. Engagement of circular RNA HECW2 in the nonautophagic role of ATG5 implicated in the endothelial-mesenchymal transition. Autophagy. 2018;14(3):404–18. https://doi.org/10.1080/15548627.2017.1414755.
Dong Y, Fan X, Wang Z, Zhang L, Guo S. Circ_HECW2 functions as a miR-30e-5p sponge to regulate LPS-induced endothelial-mesenchymal transition by mediating NEGR1 expression. Brain Res. 2020;1748:147114.
Liu W, Cai H, Lin M, Zhu L, Gao L, Zhong R, et al. MicroRNA-107 prevents amyloid-beta induced blood–brain barrier disruption and endothelial cell dysfunction by targeting Endophilin-1. Exp Cell Res. 2016;343(2):248–57. https://doi.org/10.1016/j.yexcr.2016.03.026.
Li AD, Tong L, Xu N, Ye Y, Nie PY, Wang ZY, et al. miR-124 regulates cerebromicrovascular function in APP/PS1 transgenic mice via C1ql3. Brain Res Bull. 2019;153:214–22. https://doi.org/10.1016/j.brainresbull.2019.09.002.
Zhang H, Zhang S, Zhang J, Liu D, Wei J, Fang W, et al. ZO-1 expression is suppressed by GM-CSF via miR-96/ERG in brain microvascular endothelial cells. J Cereb Blood Flow Metab. 2018;38(5):809–22.
Lin M, Zhu L, Wang J, Xue Y, Shang X. miR-424-5p maybe regulate blood–brain barrier permeability in a model in vitro with Abeta incubated endothelial cells. Biochem Biophys Res Commun. 2019;517(3):525–31. https://doi.org/10.1016/j.bbrc.2019.07.075.
Yu P, Venkat P, Chopp M, Zacharek A, Shen Y, Ning R, et al. Role of microRNA-126 in vascular cognitive impairment in mice. J Cereb Blood Flow Metab. 2019;39(12):2497–511. https://doi.org/10.1177/0271678X18800593.
Toyama K, Spin JM, Deng AC, Huang TT, Wei K, Wagenhauser MU, et al. MicroRNA-mediated therapy modulating blood–brain barrier disruption improves vascular cognitive impairment. Arterioscler Thromb Vasc Biol. 2018;38(6):1392–406. https://doi.org/10.1161/ATVBAHA.118.310822.
Liu Q, Zhu L, Liu X, Zheng J, Liu Y, Ruan X, et al. TRA2A-induced upregulation of LINC00662 regulates blood–brain barrier permeability by affecting ELK4 mRNA stability in Alzheimer’s microenvironment. RNA Biol. 2020;17(9):1293–308. https://doi.org/10.1080/15476286.2020.1756055.
Chaudhuri JD. Blood brain barrier and infection. Med Sci Monit. 2000;6(6):1213–22.
Mishra R, Singh SK. HIV-1 Tat C modulates expression of miRNA-101 to suppress VE-cadherin in human brain microvascular endothelial cells. J Neurosci. 2013;33(14):5992–6000. https://doi.org/10.1523/JNEUROSCI.4796-12.2013.
Rom S, Dykstra H, Zuluaga-Ramirez V, Reichenbach NL, Persidsky Y. miR-98 and let-7g* protect the blood–brain barrier under neuroinflammatory conditions. J Cereb Blood Flow Metab. 2015;35(12):1957–65. https://doi.org/10.1038/jcbfm.2015.154.
Barker KR, Lu Z, Kim H, Zheng Y, Chen J, Conroy AL, et al. miR-155 modifies inflammation, endothelial activation and blood–brain barrier dysfunction in cerebral malaria. Mol Med. 2017;23:24–33. https://doi.org/10.2119/molmed.2016.00139.
Song J, Hu Y, Li H, Huang X, Zheng H, Hu Y, et al. miR-1303 regulates BBB permeability and promotes CNS lesions following CA16 infections by directly targeting MMP9. Emerg Microbes Infect. 2018;7(1):155. https://doi.org/10.1038/s41426-018-0157-3.
Wang C, Yang Y, Cong L, Jiang Y, Du N, Zhang H. Implication of long non-coding RNA NEAT1 in the pathogenesis of bacterial meningitis-induced blood–brain barrier damage. Microvasc Res. 2021;138: 104225. https://doi.org/10.1016/j.mvr.2021.104225.
Xu B, Yang R, Fu J, Yang B, Chen J, Tan C, et al. LncRSPH9–4 facilitates meningitic Escherichia coli-caused blood–brain barrier disruption via miR-17-5p/MMP3 axis. Int J Mol Sci. 2021;22(12):6363. https://doi.org/10.3390/ijms22126343.
Yang R, Chen J, Xu B, Yang B, Fu J, Xiao S, et al. circ_2858 helps blood–brain barrier disruption by increasing VEGFA via sponging miR-93-5p during Escherichia coli meningitis. Mol Ther Nucleic Acids. 2020;22:708–21. https://doi.org/10.1016/j.omtn.2020.09.034.
Shao B, Bayraktutan U. Hyperglycaemia promotes human brain microvascular endothelial cell apoptosis via induction of protein kinase C-ssI and prooxidant enzyme NADPH oxidase. Redox Biol. 2014;2:694–701. https://doi.org/10.1016/j.redox.2014.05.005.
Shao B, Bayraktutan U. Hyperglycaemia promotes cerebral barrier dysfunction through activation of protein kinase C-beta. Diabetes Obes Metab. 2013;15(11):993–9. https://doi.org/10.1111/dom.12120.
Zhao F, Deng J, Xu X, Cao F, Lu K, Li D, et al. Aquaporin-4 deletion ameliorates hypoglycemia-induced BBB permeability by inhibiting inflammatory responses. J Neuroinflammation. 2018;15(1):157. https://doi.org/10.1186/s12974-018-1203-8.
Song J, Yoon SR, Kim OY. miR-Let7A controls the cell death and tight junction density of brain endothelial cells under high glucose condition. Oxid Med Cell Longev. 2017;2017:6051874. https://doi.org/10.1155/2017/6051874.
Zhao Q, Zhang F, Yu Z, Guo S, Liu N, Jiang Y, et al. HDAC3 inhibition prevents blood–brain barrier permeability through Nrf2 activation in type 2 diabetes male mice. J Neuroinflammation. 2019;16(1):103. https://doi.org/10.1186/s12974-019-1495-3.
Kuperberg SJ, Wadgaonkar R. Sepsis-associated encephalopathy: the blood–brain barrier and the sphingolipid rheostat. Front Immunol. 2017;8:597. https://doi.org/10.3389/fimmu.2017.00597.
Chen SL, Cai GX, Ding HG, Liu XQ, Wang ZH, Jing YW, et al. JAK/STAT signaling pathway-mediated microRNA-181b promoted blood–brain barrier impairment by targeting sphingosine-1-phosphate receptor 1 in septic rats. Ann Transl Med. 2020;8(21):1458. https://doi.org/10.21037/atm-20-7024.
Nong AN, Li QF, Huang ZJ, Xu YA, He KB, Jia YY, et al. MicroRNA miR-126 attenuates brain injury in septic rats via NF-kappa B signaling pathway. Bioengineered. 2021;12(1):2639–48. https://doi.org/10.1080/21655979.2021.1937905.
Demolli S, Doddaballapur A, Devraj K, Stark K, Manavski Y, Eckart A, et al. Shear stress-regulated miR-27b controls pericyte recruitment by repressing SEMA6A and SEMA6D. Cardiovasc Res. 2017;113(6):681–91. https://doi.org/10.1093/cvr/cvx032.
Li D, Liu YL, Pei CL, Zhang P, Pan LQ, Xiao J, et al. miR-285-Yki/Mask double-negative feedback loop mediates blood–brain barrier integrity in Drosophila. P Natl Acad Sci USA. 2017;114(12):E2365–74. https://doi.org/10.1073/pnas.1613233114.
Xu B, Zhang Y, Du XF, Li J, Zi HX, Bu JW, et al. Neurons secrete miR-132-containing exosomes to regulate brain vascular integrity. Cell Res. 2017;27(7):882–97. https://doi.org/10.1038/cr.2017.62.
Chen CY, Chao YM, Lin HF, Chen CJ, Chen CS, Yang JL, et al. miR-195 reduces age-related blood–brain barrier leakage caused by thrombospondin-1-mediated selective autophagy. Aging Cell. 2020;19(11). https://doi.org/10.1111/acel.13236.
Broekaart DWM, van Scheppingen J, Anink JJ, Wierts L, van HetHof B, Jansen FE, et al. Increased matrix metalloproteinases expression in tuberous sclerosis complex: modulation by microRNA 146a and 147b in vitro. Neuropath Appl Neuro. 2020;46(2):142–59. https://doi.org/10.1111/nan.12572.
Chen SP, Chang YA, Chou CH, Juan CC, Lee HC, Chen LK, et al. Circulating microRNAs associated with reversible cerebral vasoconstriction syndrome. Ann Neurol. 2021;89(3):459–73. https://doi.org/10.1002/ana.25965.
Becker PM, Waltenberger J, Yachechko R, Mirzapoiazova T, Sham JSK, Lee CG, et al. Neuropilin-1 regulates vascular endothelial growth factor-mediated endothelial permeability. Circ Res. 2005;96(12):1257–65. https://doi.org/10.1161/01.Res.0000171756.13554.49.
Wang Y, Cao Y, Mangalam AK, Guo Y, LaFrance-Corey RG, Gamez JD, et al. Neuropilin-1 modulates interferon-gamma-stimulated signaling in brain microvascular endothelial cells. J Cell Sci. 2016;129(20):3911–21. https://doi.org/10.1242/jcs.190702.
Mone P, Gambardella J, Wang X, Jankauskas SS, Matarese A, Santulli G. miR-24 targets the transmembrane glycoprotein neuropilin-1 in human brain microvascular endothelial cells. Noncoding RNA. 2021;7(1):9. https://doi.org/10.3390/ncrna7010009.
Ruan W, Li J, Xu Y, Wang Y, Zhao F, Yang X, et al. MALAT1 up-regulator polydatin protects brain microvascular integrity and ameliorates stroke through C/EBPbeta/MALAT1/CREB/PGC-1alpha/PPARgamma Pathway. Cell Mol Neurobiol. 2019;39(2):265–86. https://doi.org/10.1007/s10571-018-00646-4.
Lu L, Lu T, Shen J, Lv X, Wei W, Wang H, et al. Alisol A 24-acetate protects against brain microvascular endothelial cells injury through inhibiting miR-92a-3p/tight junctions axis. Aging (Albany NY). 2021;13(11):15353–65. https://doi.org/10.18632/aging.203094.
Bai Y, Zhang Y, Hua J, Yang X, Zhang X, Duan M, et al. Silencing microRNA-143 protects the integrity of the blood–brain barrier: implications for methamphetamine abuse. Sci Rep. 2016;6:35642. https://doi.org/10.1038/srep35642.
Yu DS, Wang YS, Bi YL, Guo ZP, Yuan YJ, Tong SM, et al. Salvianolic acid A ameliorates the integrity of blood-spinal cord barrier via miR-101/Cul3/Nrf2/HO-1 signaling pathway. Brain Res. 2017;1657:279–87. https://doi.org/10.1016/j.brainres.2016.12.007.
Wang L, Zhao C, Wu S, Xiao G, Zhuge X, Lei P, et al. Hydrogen gas treatment improves the neurological outcome after traumatic brain injury via increasing miR-21 expression. Shock. 2018;50(3):308–15. https://doi.org/10.1097/SHK.0000000000001018.
Zhu L, Lin M, Ma J, Liu W, Gao L, Wei S, et al. The role of LINC00094/miR-224-5p (miR-497-5p)/Endophilin-1 axis in Memantine mediated protective effects on blood–brain barrier in AD microenvironment. J Cell Mol Med. 2019;23(5):3280–92. https://doi.org/10.1111/jcmm.14214.
Shi YY, Cui HF, Qin BJ. Monomethyl fumarate protects cerebral hemorrhage injury in rats via activating microRNA-139/Nrf2 axis. Eur Rev Med Pharmacol Sci. 2019;23(11):5012–9. https://doi.org/10.26355/eurrev_201906_18093.
Ihezie SA, Mathew IE, McBride DW, Dienel A, Blackburn SL, Pandit PKT. Epigenetics in blood–brain barrier disruption. Fluids Barriers CNS. 2021;18(1):1–16.
Salvador E, Shityakov S, Forster C. Glucocorticoids and endothelial cell barrier function. Cell Tissue Res. 2014;355(3):597–605. https://doi.org/10.1007/s00441-013-1762-z.
Keil JM, Liu X, Antonetti DA. Glucocorticoid induction of occludin expression and endothelial barrier requires transcription factor p54 NONO. Invest Ophthalmol Vis Sci. 2013;54(6):4007–15. https://doi.org/10.1167/iovs.13-11980.
Hartmann C, Corre-Menguy F, Boualem A, Jovanovic M, Lelandais-Briere C. MicroRNAs: a new class of gene expression regulators. Med Sci (Paris). 2004;20(10):894–8. https://doi.org/10.1051/medsci/20042010894.
Sohrabji F, Park MJ, Mahnke AH. Sex differences in stroke therapies. J Neurosci Res. 2017;95(1–2):681–91. https://doi.org/10.1002/jnr.23855.
Girijala RL, Sohrabji F, Bush RL. Sex differences in stroke: review of current knowledge and evidence. Vasc Med. 2017;22(2):135–45. https://doi.org/10.1177/1358863X16668263.
Burek M, Konig A, Lang M, Fiedler J, Oerter S, Roewer N, et al. Hypoxia-induced MicroRNA-212/132 alter blood–brain barrier integrity through inhibition of tight junction-associated proteins in human and mouse brain microvascular endothelial cells. Transl Stroke Res. 2019;10(6):672–83. https://doi.org/10.1007/s12975-018-0683-2.
Zhang Y, Han B, He Y, Li D, Ma X, Liu Q, et al. MicroRNA-132 attenuates neurobehavioral and neuropathological changes associated with intracerebral hemorrhage in mice. Neurochem Int. 2017;107:182–90. https://doi.org/10.1016/j.neuint.2016.11.011.
Ren S, Wu G, Huang Y, Wang L, Li Y, Zhang Y. MiR-18a aggravates intracranial hemorrhage by regulating RUNX1-Occludin/ZO-1 axis to increase BBB permeability. J Stroke Cerebrovasc Dis. 2021;30(8): 105878. https://doi.org/10.1016/j.jstrokecerebrovasdis.2021.105878.
Acknowledgements
This work was supported by the National Institutes of Health, American Heart Association, and the U.S. Department of Veterans Affairs.
Funding
This work was supported by the National Institutes of Health Grant NS112181 (K.J. Yin); and American Heart Association Grant #20POST35210900 (P. Sun); K.J. Yin is also supported by the Merit Review Grant I01BX004837 from the Department of Veterans Affairs.
Author information
Authors and Affiliations
Contributions
PS and KJY conceived the concept and content of this article; PS wrote, and MHH and KJY revised the article, all authors approved the final version of the article.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sun, P., Hamblin, M.H. & Yin, KJ. Non-coding RNAs in the regulation of blood–brain barrier functions in central nervous system disorders. Fluids Barriers CNS 19, 27 (2022). https://doi.org/10.1186/s12987-022-00317-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12987-022-00317-z