
Abstract
The global rise in urbanization and industrial activity has led to the production and incorporation of foreign contaminant molecules into ecosystems, distorting them and impacting human and animal health. Physical, chemical, and biological strategies have been adopted to eliminate these contaminants from water bodies under anthropogenic stress. Biotechnological processes involving microorganisms and enzymes have been used for this purpose; specifically, laccases, which are broad spectrum biocatalysts, have been used to degrade several compounds, such as those that can be found in the effluents from industries and hospitals. Laccases have shown high potential in the biotransformation of diverse pollutants using crude enzyme extracts or free enzymes. However, their application in bioremediation and water treatment at a large scale is limited by the complex composition and high salt concentration and pH values of contaminated media that affect protein stability, recovery and recycling. These issues are also associated with operational problems and the necessity of large-scale production of laccase. Hence, more knowledge on the molecular characteristics of water bodies is required to identify and develop new laccases that can be used under complex conditions and to develop novel strategies and processes to achieve their efficient application in treating contaminated water. Recently, stability, efficiency, separation and reuse issues have been overcome by the immobilization of enzymes and development of novel biocatalytic materials. This review provides recent information on laccases from different sources, their structures and biochemical properties, mechanisms of action, and application in the bioremediation and biotransformation of contaminant molecules in water. Moreover, we discuss a series of improvements that have been attempted for better organic solvent tolerance, thermo-tolerance, and operational stability of laccases, as per process requirements.
Similar content being viewed by others

Introduction
Urbanization and industrialization have resulted in a serious contamination of water bodies, causing harmful effects to ecosystems. Biotechnologists around the world are researching and developing innovative tools and non-polluting processes to correct the effect of global pollution. However, this is challenging, owing to the quantity and diversity of pollutant molecules discharged into water bodies [1], such as plastics, herbicides, fertilizers, synthetic dyes, polycyclic aromatic hydrocarbons (PAHs), chlorinated paraffin phthalates, and others, such as the so-called emerging pollutants, which may include pharmaceuticals (i.e. pain relievers, antibiotics, hormones, endocrine disruptors), plasticizers, and compounds contained in self-care products, among others [2,3,4,5,6,7,8]. Different treatment approaches have been explored, ranging from physical and chemical methods to biotechnological strategies (such as the use of laccase enzymes), to retain or transform these molecules into less harmful ones [9, 10]. The pollution of water bodies is a technical, social, and environmental challenge, attributable to continuous population increase and limited waste elimination strategies coupled with poor public management of water contaminants [4, 5, 11]. In fact, a variety of molecules originating from home and industry are released into water bodies without regulation; including emerging pollutants suspected to have effects on the environment and health (Fig. 1) [1, 12,13,14].

Schematic illustration of the potential sources of water contaminants and their bioremediation by laccases. Emerging contaminants such as antibiotics, endocrine disruptors, dye-based pollutants and pharmaceutical drugs are often released into the environment causing harmful impacts and health problems to humans and other animals, water treatment with laccases and their biotechnological approaches generate less-toxic, inert or fully degraded compounds
In order to eliminate pollutants from contaminated water, the identification, study, and implementation of laccase-mediated processes form an intensive research area aimed at generating ecofriendly and effective tools for treating and improving water quality (Fig. 1). Laccases, which belong to the enzyme family of multi-copper oxidases (MCOs), are classified as benzenediol oxygen reductases (EC 1.10.3.2) and are also known as urushiol oxidases and p-diphenol oxidases [15, 16]. They are considered versatile enzymes capable of oxidizing a large number of phenolic and non-phenolic molecules due to their low substrate specificity, using oxygen as electron acceptor and generating water as a by-product [17,18,19]. Laccases are widely expressed in nature; they can be obtained from various fungi, plants, bacteria, lichen, and insects (Fig. 2), with laccases from each species exhibiting particular catalytic characteristics and sequences [20,21,22]. UniProtKB search results for “laccase” with sequence sizes between 220 and 800 amino acids, revealed approximately 7300 cellular-organism sources, with 1026 bacteria, 6258 eukaryotes, and 16 halobacteria (archaea). Hence, it can be predicted that this large number of enzymes produced by different organisms could have a wide range of applications in water bioremediation (Fig. 2). To date, many of these enzymes have been applied in processes like electrocatalysis, delignification, and ethanol production [23]. In this review, we aim to describe laccases from different organisms, used in water bioremediation, their varying properties based on their origin, their biotechnological prospectives for pollutant degradation (fabric discoloration, herbicide degradation, and emerging pollutants transformation), and the different strategies that have been explored to increase their activity and application.

Phylogenetic tree constructed with some of the different organism sources of laccases, as well as some of their applications in bioremediation. According to their bacterial, insect, plant or fungal origin, they are colored with blue, red, green or orange, respectively. The alignments and phylogenetic relationships were done using the MEGA X suite
Sources of laccases that are useful in water bioremediation
Fungal laccases
The first fungal laccase was reported by Bertrand [24], who observed that this enzyme was responsible for the color change in mushrooms of the Boletus genus when in contact with air. A large number of fungi have been confirmed as laccase producers, with white rot fungi being the most recognized. Among fungal species, the basidiomycetes, specifically Agaricus bisporus, Pleurotus ostreatus, Trametes versicolor, Phanerochaete chrysosporium, and Coprinus cinereus, produce various laccase isoforms (Table 1) [23, 25, 26].
Fungal laccases are involved in sporulation, pigment production, fruiting body formation, stress defense, plant pathogenesis, and lignin degradation [27, 28]. Although most purified laccases are extracellular enzymes, wood-rotting fungi also contain intracellular laccases. It has been suggested that the localization of laccase is probably connected with its physiological function and determines the range of available substrates [29]. Laccases exist in a variety of structures; most of them are monomeric, but some are also present in homodimeric, heterodimeric, and multimeric forms. Their molecular mass ranges from 50 to 140 kDa, depending on the organism, although a typical fungal laccase will range from 60 to 70 kDa with an isoelectric point around pH 4.0 [29, 30]. Fungal laccases are usually glycosylated, with a 10–25% increase in mass, although some laccases present with a > 30% increase. The carbohydrate portion of laccases has been demonstrated to ensure their conformational stability and to protect the enzyme from proteolysis and inactivation by radicals [31, 32].
The redox potential (E°) of laccases has a direct relationship with the energy required to remove an electron from the reducing substrate, constituting one of the fundamental characteristics of these enzymes [33]. Therefore, laccases with a high E°, like fungal laccases, are of special interest in biotechnology cause they are capable of oxidizing substrates with high E° (E° > 400 mV) [33,34,35,36]. For example, the E° of bisphenol A (BPA), p-nonylphenol and azo dye BR114 are above 600 mV [37, 38]. Fungal laccases aid bioremediation through the oxidation of polycyclic aromatic hydrocarbons (PAHs) [39, 40], plastics and phenolic compounds [41,42,43,44], dyes [44,45,46,47,48], and the degradation of pharmaceutically active compounds [49,50,51,52], among others (Table 1). Given that laccases from white-rot fungi have the potential for phenolic compound degradation, different studies have involved the immobilization of microorganisms, such as T. versicolor, into silica-alginate and loofa sponges as supports for phenol removal [43]. While crude extract from Trametes pubescens has been used for the degradation of chlorophenols (Table 1) [44]. Also, crude extract from the white-rot fungus Trametes hirsuta, proved capable of degrading chloramphenicol (one of the most persistent micro-pollutants in pharmaceutical wastes), with or without mediators (Table 1) [44, 53]. Fukuda et al. [54] used a free purified laccase from Trametes villosa to degrade BPA, another hazardous pollutant discharged into rivers and seas, without the requirement of mediators. Meanwhile, Barrios-Estrada et al. [42] reported that the degradation of BPA (20 mg/L) occurred within the first 24 h when using Pycnoporus sanguineus (CS43) and T. versicolor laccases immobilized onto ceramic membranes (Table 1). Different steroidal estrogens can be removed or degraded from aqueous systems by the free laccases from P. sanguineus or laccases from T. versicolor or Myceliophthora thermophila that have been immobilized onto ceramic membranes (Table 1) [55, 56]. Other problematic compounds in effluents of textile and paper industries include synthetic dyes, of which, many are toxic for mammals. Therefore, efforts have been made towards their elimination from industrial wastewaters using laccases from T. versicolor and Trametes trogii (Table 1) [49, 58].
The laccase yield from native fungal sources fails to meet the industrial need, as natural hosts often produce several laccase isozymes making it challenging to isolate the laccase of interest, more so when the enzyme is silent or not abundantly expressed. Therefore, heterologous laccase expression has become a promising alternative [23, 59,60,61,62,63,64,65,66,67,68,69,70,71]. Heterologous expression of many fungal laccases has been reported in bacteria such as E. coli [60], yeasts like Pichia pastoris and Yarrowia lipolytica [43, 61,62,63,64], filamentous fungi such as Aspergillus oryzae, A. niger, and Trichoderma atroviride [65,66,67], and plants like Arabidopsis thaliana and Zea mays (Table 1) [68, 69]. Yeasts and filamentous fungi are usually more attractive hosts for heterologous protein production owing to their faster microbial growth, ease of gene manipulation, their ability to secrete large amounts of proteins into the growth medium, as well as the ability to perform post-translational modifications [30, 49].
Recombinant fungal laccases have also been widely applied for bioremediation purposes. For instance, the recombinant proteins Lcc1 and Lcc3 from T. trogii, produced in P. pastoris proved to be a useful biocatalyst for the oxidative degradation of several polluting dyes, such as indigo carmine, the most important dye used for manufacturing blue jeans [62, 70]. Moreover, Darvishi et al. [71] expressed and produced a recombinant laccase (Lcc IIIb) from T. versicolor in Y. lipolytica, proving its capability of decolorizing five phenolic azo dyes with > 40% efficiency after 4 h (Table 1). Similarly, Wang et al. [68] expressed a laccase from the ectomycorrhizal fungus Laccaria bicolor in P. pastoris and A. thaliana, which proved capable of decolorizing > 80% of the crystal violet dye, tested using laboratory-scale studies, providing an alternative to the decolorization of industrial wastes. In another study, Balcázar-López et al. [67] expressed a laccase from P. sanguineus in the filamentous fungus T. atroviride; the heterologously expressed laccase maintained similar properties to those of the native enzyme, although the recombinant showed the potential to remove > 90% of the phenanthrene and benzo[α]pyrene present in wastewater from a biofuel industry plant using laboratory-scale studies [67].
Plant and insect laccases
The first identified and reported laccase from plants was from the Japanese lacquer tree Toxicodendron vernicifluum (Rhus vernicifera) [72]. However, studies on plant laccases are rare. Plant laccases share their molecular architecture and reaction mechanisms with fungal laccases. In general, they have a lower E° like bacterial laccases (0.41 V for R. vernicifera and a pI between 7.0 and 9.6) [22, 31, 73]. These proteins show a higher glycosylation pattern (22–45%) [74, 75], consist of 500–600 amino acids, and weigh approximately 60–130 kDa [31]. Plant laccases have been described and associated with biosynthesis and polymerization of lignin [76, 77], elongation [78,79,80], and the stress response [81,82,83].
Although plant laccases have not been largely involved in bioremediation, some applied cases have been reported. Wang et al. [84] presented a system of phytoremediation ex planta based on the overproduction in A. thaliana of a secretory laccase (LAC1), which was natively expressed in the roots of Gossypium arboreum. LAC1 expression in A. thaliana conferred resistance to several toxic phenolic compounds, probably attributable to LAC1-induced transformation. Recombinant LAC1 plants were resistance to phenolic compounds under greenhouse conditions, helping to detoxify their growth environment [84]. Watharkar et al. [85] showed that laccases and other enzymes from Asparagus densiflorus could be applied in the treatment of industrial textile effluents. For lab scale studies, they used a vertical subsurface flow phytoreactor based on vertical percolation of wastewater through layers of soil, root zone and a netted bottom. For large scale studies, they planted beds of A. densiflorus on a high rate transpiration system (HRTS), which has been used successfully for some industries. In both cases, A. densiflorus showed the ability to degrade dyes and reduced levels of toxic heavy metals.
Laccases from other plants have been proposed and successfully tested for dye degradation using suspension cells and purified laccases [86,87,88,89,90]. Huang et al. [91] identified laccases in rice (Oryza sativa), possibly involved in atrazine and isoproturon (herbicides) catabolism or detoxification. The two Oryza sativa laccases expressed heterologously in P. pastoris, led to the increased resistance of cells to atrazine and isoproturon, suggesting that some of the laccases could be involved in detoxification or degradation of these herbicides [91].
Plants have been successfully used as recombinant expression systems of fungal and plants laccases. Chiaiese et al. [92] expressed a laccase from P. ostreatus in Nicotiana tabacum, capable of reducing 70% of the total phenol content from olive mill wastewaters (Table 1). Other authors have expressed fungal laccases with industrial applications in rice-based [93] and tobacco plants [94, 95], as well as maize seeds [69]. Conversely, insect laccases have been reported to play an important role in cuticle sclerotization and pigmentation, as well as other processes such as wound healing and immune system development and maintenance [96, 97]. To the best of our knowledge, no insect laccase has been reported for bioremediation processes.
Bacterial laccases
Laccase activity in bacteria was detected for the first time in Azospirillum lipoferum, isolated from a rice rhizosphere in 1993 [98]. Several laccases were then gradually discovered in bacteria from different genera, such as Bacillus, Streptomyces, Klebsiella, Pseudomonas, Yersinia, Proteobacterium, and Marinomonas, among others (Table 3) [99, 100]. Moreover, these enzymes have also been found in microorganisms of the Archaea domain such as Haloferax volcanii [101].
Under native conditions, bacterial laccases are involved in pigmentation processes, morphogenesis, toxin oxidation, and protection against oxidizing agents and UV light [100, 102]. The molecular weight of these enzymes is in the range of 50–70 kDa, with a majority being monomeric intracellular proteins, except those from bacteria in the Streptomyces genera and some other examples [103,104,105,106], such as the laccase produced by Bacillus tequilensis SN4, an extracellular enzyme [104].
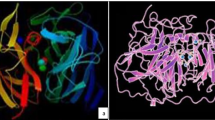
One of the most well-known bacterial laccases is the outer endospore coat protein CotA from Bacillus subtilis, which has three cupredoxin domains (Fig. 3) [107]. Other similar bacterial MCOs include the copper homeostasis protein CueO from E. coli [108]. Bacterial laccases with three-dimensional structures of two-domain laccases have been found in Streptomyces, Amycolatopsis, and Nitrosomonas, belonging to the group denoted as SLACs (small laccases). The implication of the absence of this domain is the need to form a homotrimer to be catalytically active (Fig. 3) [109,110,111].

Cartoon structures of the three-domain laccase from Bacillus subtilis (PDB 1GSK) and the homotrimeric two-domain laccase from Streptomyces coelicolor (PDB 3CG8). The domain assignations were made using the SWORD partition algorithm
The most significant biochemical properties of bacterial laccases are their stability under various conditions of pH, temperature, organic solvents, and salt concentrations [105]. Usually, bacterial laccases are highly stable at elevated temperatures, as seen in the B. subtilis laccase at 70 °C, with a thermal half-life (t1/2) of 250 min, or the t1/2 of 30 min at 80 °C of the Streptomyces viridochromeogenes laccase, compared with the 10 min t1/2 of Cerrena unicolor fungal laccase at the same temperature [112,113,114,115]. With respect to media pH, bacterial laccases usually work better in neutral to alkaline pH, similar to plant laccases, but unlike fungal laccases, which have optimum activities in acidic pH. Nevertheless, its optimal pH is dependent on the substrate. For instance, for phenols, such as 2,3-dimethoxyphenol, the optimal pH for B. subtilis, B. clausii, and Streptomyces coelicolor are pH 7, 8 and 9, respectively, while for ABTS, all three enzymes require a pH of 4 [112, 116]. However, bacterial laccases have shown greater tolerability to high concentrations of sodium chloride, being active in 1 M or higher concentrations, as seen with the laccases of Marinomonas mediterranea and Bacillus halodurans, among others [115, 117]. Some bacterial laccases have exhibited high tolerance to different solvents, including ethanol, methanol, dimethylformamide, acetonitrile, acetone, and dimethylsulfoxide, as observed in the Bacillus pumilus W3 laccase, which generally retains > 50% of its activity in solvent–water mixtures [118].
Although bacterial laccases are generally more robust and stable enzymes in comparison to fungal laccases, their application has been restricted by their low E° (E° T1 < +460 mV) [22, 112]. Nevertheless, bacterial laccases represent a good option for the treatment of contaminated wastes such as textile effluents, which usually have high salt concentrations (40–100 g/L) and alkaline pH [119].
Heterologous overexpression of bacterial laccases has been reported in E. coli [99, 112, 115, 118, 120,121,122,123,124,125], P. pastoris [126,127,128], and Streptomyces coelicolor [116]. Although E. coli is the most used expression system for bacterial laccases, the production of MCOs in its cytoplasm has a major drawback as its copper homeostasis systems maintain a cellular copper concentration around 10 µM under aerobic conditions [129,130,131], which is insufficient to achieve fully loaded copper laccases [132]. Copper-depleted laccases are incapable of reaching their maximum catalytic activity [122, 132]. This limitation can be overcome by changing the oxygen concentration when cultivating recombinant E. coli expressing laccases, because under anaerobic (or microaerobic) conditions, the intracellular copper accumulation is 80-fold higher, compared with that attained under aerobic conditions [103, 122, 132, 133].
Bacterial laccases have been used in bioremediation, mainly for the degradation of synthetic dyes. Liu et al. [99] reported a thermostable and pH-stable Klebsiella pneumoniae laccase which degrades diverse dyes used in industrial processes (such as reactive brilliant blue X-BR, reactive dark blue M-2GE, congo red, bromophenol blue, and malachite green, among others) in short reaction times (90 min) under diverse pH values at 70 °C (Table 3). Another case is the B. pumilus CotA-laccase mutant WLF, obtained by Luo et al. [125], which has an improved expression in E. coli and has been tested for the degradation of diverse dyes, obtaining higher decoloration yields with anthraquinonic and triphenylmethane dyes, compared with aromatic heterocyclic dyes (Table 3). Meanwhile, high decolorization of toluidine, malachite green, and reactive black 5 by the azide-resistant spore laccase from halotolerant Bacillus safensis, has also been reported [134]. Recombinant laccases from E. coli [116] or Thermus thermophiles [126] expressed in P. pastoris, efficiently decolorized congo red and remazol brilliant blue R (Table 3). Recently, the recombinant Streptomyces ipomoea SilA laccase expressed in E. coli, in the presence of mediators such as acetosyringone and methyl syringate, enhanced the decolorization and detoxification of a variety of textile dyes, like reactive black 5, orange II, and indigo carmine, also diminishing the toxicity of acid orange 63, tartrazine and its products [135].
Outside these pollutants, other contaminant compounds have been degraded with laccases. Singh et al. [136] used recombinant Yersinia enterocolitica laccase to biotransform two nonsteroidal anti-inflammatory drugs (diclofenac (DF) and aspirin), obtaining complete transformation of these molecules in 24 h (Table 3). Furthermore, DF and mefenamic acid were also transformed by laccases produced by Streptomyces cyaneus [137], and Streptomyces mutabilis laccases transformed antibiotics like sulfadiazine and sulfathiazole [138]. Similarly, the recombinant S. ipomoea SilA laccase expressed in E. coli, has shown a high percent conversion of ciprofloxacin and norfloxacin [139]. Interestingly, PAHs such as anthracene, pyrene benzo[α]pyrene, phenanthrene, and fluoranthene, have been oxidized by the recombinant laccase CotA from B. subtilis produced in E. coli [123]. Moreover, laccases from S. cyaneus have demonstrated full BPA degradation after 2 days [137].
Structure of laccases and comparative structure analyses
Laccases as MCOs, have four copper atoms in remarkably special oxidation states: one type-1, one type-2, and two type-3s, all forming their catalytic site (Fig. 4). Laccases are members of the cupredoxin superfamily, particularly the family of multi-domain cupredoxins. This family is characterized by the cupredoxin fold, which consists of two β-sheets arranged into a Greek-key barrel. The Greek-key motif has at least seven antiparallel β-strands twisted to form a closed barrel structure, in which some β-strands are adjacent in space but not in sequence [109].

Representation of the different amino acids of the catalytic site that coordinates the catalytic coppers in Trametes versicolor laccase (PDB 1KYA). The amino acids of the histidine-cysteine pathway are in green
The classification of the copper atoms is based on the environment of the metal ion and its spectroscopic characteristics; T1: paramagnetic ‘blue’ copper, with an absorbance at 610 nm, T2: paramagnetic ‘non-blue’ copper, and T3: a diamagnetic spin-coupled copper–copper pair, with an absorbance at 330 nm [140]. T1 copper has the highest E° and is the substrate oxidation site. This ion has a trigonal orientation, with two conserved histidines and one cysteine as equatorial ligands, and an axial ligand of variable nature, usually methionine in bacteria and leucine or phenylalanine in fungal laccases. Type-2 and the two type-3 coppers form a cluster, where molecular oxygen is reduced, and water is released. Types-2 and 3 copper atoms are coordinated by histidine side chains (T2 by two of them and T3 by six) (Fig. 4). A hydroxyl bridge maintains the antiferromagnetic coupling between T3 copper atoms [141].
Common laccases contain three homologous cupredoxin domains. Their mononuclear copper site exists in domain 3 and their trinuclear cluster is formed at the interface between domain 1 and 3 [109]. In laccases with this topology, the function of domain 2 is to join and position domains 1 and 3, enabling the formation of the trinuclear cluster [106]. In contrast, in two-domain laccases, which are from bacteria and are so called small laccases, their mononuclear copper site exists in domain 1 or 2, but for the formation of their trinuclear cluster they need to oligomerize as homotrimers, generating this catalytic site at the interface between the domain 1 of one monomer and the domain 2 of the other monomer [106, 142].
In both cases, the distance and relative position between the copper sites are conserved (the distance between T1 copper and the cluster); about 12 Å in all laccases [109].
There are several hypotheses on the evolution of laccases; all of them consider that the cupredoxin domain, with one copper atom in its structure, developed in different forms of MCOs, including dicyanin, ascorbate oxidase, nitrite reductase, ceruloplasmin, SLACs, and three-domain laccases [140, 143]. These hypotheses postulate different pathways and intermediate species that led to the development of the trinuclear cluster and the origin of the different MCOs. These structures maintain the original cupredoxin domain but are associated in dimers of independent chains or form longer chains by gene fusion [111]. Some of these domains maintain the copper-binding site, and different forms of interdomain association were evolutionarily explored by independent divergence to develop at least two cluster types of three copper atoms [142]. Other interesting schemes of this hypothesis have also been reported [144, 145].
Three-domain laccases are mainly studied in fungi, but have also been observed in some bacteria, archaea, plants, and insects [22, 146]. The database containing information on different laccases and MCOs is BioCatNet [147]. Laccases are considered “moonlighting” proteins, owing to their multiple biological activities [108]. PDB structures of > 70 fungal and a few bacterial laccases have been reported, crystallized in their wild-type, mutant, and derivative forms, as well as complexed to a variety of substrate-like ligands and oxygen reactive species [109, 148]. This set of structures has shed light on their stabilities and functional characteristics, as described above.
Nevertheless, no structures from other species have been reported, except a plant (zucchini) ascorbate-oxidase closely homologous to laccases [149, 150]. The general three-domain structure of laccases is maintained in different species, with the loops protruding the cupredoxin domains being the most conspicuous difference [111, 151], as well as the form and by consequence, the selectivity at the substrate binding site [152]. More subtle differences are situated in the axial position of the T1 copper atom, causing the span of E°s from 400 mV in plant and bacterial laccases to approximately 800 mV in the majority of fungal types [22, 139, 153]. In Fig. 5, we compare the structures of representative fungal, bacterial, plant, and insect laccases. This latter structure was homology modeled from its amino acid sequence. The conserved orientation of the coppers and the cavities for substrates and products are also presented.

Laccase structure conservation and function. a Structure of Trametes versicolor (PDB ID 1GYC), and Bacillus subtilis (PDB ID 1GSK) laccases compared to Cucurbita pepo (zucchini) ascorbate oxidase (PDB ID 1AOZ) from left to right. Domain 1 (D1) is at the front and right of the structure, domain 2 (D2) is behind and in the upper portion, domain 3 (D3) is at the left. Brown spheres symbolize the position of copper atoms, T1 above the trinuclear cluster. b The molecular surface shows protruding chemical groups, in red, and concave or cavity regions, in green. Some of these latter regions correspond to the ligand-binding site (LB) along with the dioxygen molecule entrance (O2) and the water exit (H2O) channels. Central and right images were created from that on the left by rotating it 30° over the horizontal axis, or 30° over the vertical axis, respectively
Several review papers have been published discussing the structure of laccase and its implications on function. For instance, the description of the molecular mechanism of substrate oxidation in the T1 site, the intramolecular electron transfer to the trinuclear cluster located about 12 Å away, and the oxygen reduction to water, can be understood in the scheme of Hakulinen and Rouvinen [109], and the detailed descriptions of Mot and Silaghi-Dumitrescu [140], Pardo and Camareno [154], and Sitarz et al. [155]. These references show the complexity and subtlety of the reaction pathway through enzyme structure. This mechanism involves a substrate binding pocket [154, 156], which confers selectivity by proper docking, and also affects the E° by induced fitting to the active site. There, the T1 copper atom extracts an electron from the substrate, followed by a relay of protein functional groups, namely thiol, carbonyl, and imidazole groups, which transfer that electron through the trinuclear cluster, where they are gathered until four electrons are collected. T3 copper atoms transfer such electrons to the T2 copper, and an oxygen channel allows an oxygen molecule to reach this buried metal ion and be reduced [140].
The reduced oxygen atoms are then converted to water following assistance from two carboxylate groups from aspartic and glutamic acids, which transfer the required hydrogen atoms. At least two structural water molecules also contribute to the electron transfer process. The generated water molecules finally go out a second channel formed by polar residues of the protein. Copper atoms undergo a series of at least five stages during this process [109, 154, 157]. This depiction of the process exposes the number of chemical species involved during oxidative catalysis, and the essential participation of the molecular structure [156].
Characteristics of the biological activity of laccases enable the name “green catalysts,” as they oxidize different substrates, only require oxygen molecules as reactants, and only produce water molecules as byproducts [22]. The structure of the molecular system is complex, involving its protein structure as well as its carbohydrate moiety as a stabilizing fastener [140, 156] and functional coadjuvant, along with structural water molecules, a C-terminus rearrangement, the coordination state of copper atoms, electron transfer through main and side-chains, and mediators [158]. Moreover, solvent composition is also a determinant in laccase stability, for example the presence of polyhydroxyl compounds [159].
Different approaches have been employed to handle such complexity for the development of laccases tailored to specific industrial and bioremediation processes [22, 152, 156]. These approaches can be classified as rational (computer prediction based on molecular modeling, quantum mechanics, and molecular dynamics simulations) [152, 160], semi-rational (experimental assays of trial and error mutants on a structural position identified by knowledge-based analyses or calculation), directed evolution screenings, assays of chimeric structures and laccase immobilization [152, 154, 161], and recent synthetic biology schemes [162]. These approaches have successfully produced laccase mutants or derivatives with enhanced temperature or organic solvent stability; activities tailored to develop specificity to certain substrates; higher E° in the T1 site, enhanced heterologous expression, the shift of pH-activity profiles, and tolerance to chemical inhibitors. In structural terms, these improvements were achieved by modifying the functional groups in the substrate binding site and T1 copper coordination [22], as well as introducing stabilizing mutants in the domain interface [163]. Nevertheless, the precise prediction of the effect of a specific mutation remains elusive [152, 155].
Mechanism of action of laccases
The potential application of laccases in numerous and different biocatalytic processes for industry and environmental solutions has increased the interest in understanding their mechanism of action. In general, laccases oxidize a wide range of substrates; typically substituted phenols and aromatic amines, which are transformed into free radicals (Fig. 6a) [164, 165]. Unstable chemical products and primarily generated free radicals commonly start domino reactions (Fig. 6b), leading to complex chemical transformations of biological relevance such as lignin synthesis and degradation [166].

Mechanism, kinetic model and structural elements involved in laccase functional properties and reaction. a Representation of the laccase mechanism of action in the active site of Trametes versicolor laccase (PDB 1KYA). In orange are represented those aminoacids involved in the binding, stabilization and orientation of the substrate, in grey and green those that are involved in the coordination of catalytic coppers and the electron transfer and in yellow those that transfer protons for the oxygen assisted reduction. b Laccase action on a lignin-model illustrating the domino effect [198]. c Complex two-site ping-pong bi-bi kinetic model proposed for the laccase reaction [184, 185]. d Structural and functional elements involved in different steps of the laccase reaction
The overall laccase reaction involves one electron (1e−), sequential oxidations of four molecules of reducing substrates, concurrently with two double electron (2 × 2e−) reductions of oxygen atoms into their respective H2O molecules. This process is accompanied by a catalytic exchange of 4 H+ equivalents [167]. From the structural, mechanistic, and kinetical points of view, a laccase reaction is approached as two half-reactions connected by an internal electron transfer (IET) step, assisted by the catalytic copper ions located at the T1 Cu and T2 Cu/T3 Cuα/T3 Cuβ trinuclear cluster (TNC) sites [157, 167, 168].
The fully conserved nature of the eleven (one Cys and ten His) residues forming the T1 copper and TNC laccase sites, and in general all MCOs, explain their essential role in the catalytic action. This relationship has been experimentally demonstrated by the comparison of sequences and mutagenic approaches in many studies [168,169,170]. Similarly, other fully or highly conserved residues achieving important roles in different catalytic steps involved in laccase action have been identified and include the recognition and docking of reducing substrates, IET from the T1 copper ion into the TNC site, and reduction of oxygen atoms at the TNC site. As a rule, these residues are located in the vicinity of their respective sites of action, where they appear as second sphere residues [171].
Despite these advances in the understanding of the action of laccases in terms of structure–function, a complete picture relating their molecular properties and mechanisms with their kinetic performance remains unclear. This condition could be understood based on the evolutionarily adjusted broad range of organic molecules capable of being oxidized by a laccase, and the relative ability of diverse laccases to drag substrates into recognition sites and favorably orientate them to be oxidized, limiting an integrated scheme [172,173,174,175]. A brief review of how these structural elements are mechanistically linked to the function of the catalytic copper center, and the manner in which their kinetic performance is influenced, is outlined below.
In the first half semi-reaction, 1e− substrate oxidation takes place at the T1 copper site located at the bottom of the substrate binding pocket. In T. versicolor laccase 2 (TvL), this substrate interaction region is delimited by several highly conserved hydrophobic residues; Phe 162, Leu 164, Phe 265, Phe 332, and Pro 391, that form a favorable environment for the docking of hydrophobic molecules, such as the typical aromatic phenol/amine substrates to be oxidized by laccase. In addition, the fully conserved residue Asp 206 (near His 458 of the T1 copper site), located at the bottom of the substrate binding pocket, contributes to substrate stabilization and orientation (through O–H interactions) at the catalytic T1 copper site, through the participation of a fully conserved His 458 [174, 176]. This last residue is exposed to the solvent at the interface of the substrate binding cavity. In this manner, Asp 206 acts as an essential mechanistic element by promoting electron subtraction and transfer from substrate donor molecules into the T1 copper ion (Cu2+ → Cu1+) through a direct interaction with His 458, in the T1 copper site. Moreover, the high E°′ observed on this TvL has been directly related to the presence of the non-ligating semi-conserved hydrophobic residue Phe 463 at the axial position in this center [177]. Based on this, laccases are commonly classified into three classes: low E°′ laccases (< +460 mV), typically found in plants and bacteria, with the axial position at the T1 copper site occupied by a 4th ligating Met residue; medium E°′ laccases (from 460 to 710 mV), typically reported in ascomycetes and other lignin-degrading fungi sharing similar ecophysiological niches, with a non-ligating Leu residue at the axial position of the T1 copper; and high E°′ laccases (> +710 mV), typically reported in white-rot basidiomycetes, with the non-ligating Phe residue at this position [22, 153, 174, 178].
In the IET linking step, electrons from the reduced T1 copper ion are rapidly transferred into Cu3α at the TNC site. This step is a result of the interaction between conserved Cys 453 (linked to the T1 Cu1+ ion) and His 454 (linked to Cu3α), mediated by an N–H backbone group of the semi-conserved Leu 455. This structural redox-arrangement is known as the Cys-His electron transfer (ET) bridge. The second half-reaction of oxygen reduction takes place in two consecutive two-electron steps at the TNC in a redox reaction, which requires the synchronized action of the Cys-His IET bridge to complete one catalytic cycle. In this step, oxygen molecules diffuse into the TNC through the entry solvent accessible channel [179]. The first two electrons are donated by T3 copper ions of the fully reduced enzyme to oxygen, resulting in the formation of the laccase peroxide intermediate (PI). The second two electrons are then transferred from the T1 and T2 copper ions to reductively cleave the O–O bond. This step occurs in a concerted way with a catalytic transfer of H+ from the carboxylic Glu 424 residue, in an action that involves the mediation of the semi-conserved Asp 77 exposed to solvent at the interface of the TNC site. The reductive O–O cleavage results in the formation of the laccase native intermediate (NI). This mechanism of electrons/H+ transfer avoids the generation of reactive oxygen radicals at the TNC site, and the first H2O molecule produced, diffuses out of the TNC site through the exit solvent accessible channel [109, 179, 180].
Kinetic properties of laccases, Km (apparent affinity), kcat (catalytic rate constant), and kcat/Km have been commonly determined in steady-state studies (Fig. 6c, d). A complex two-site ping-pong bi-bi mechanism has been suggested from a study with the pioneering model Rhus vernicifera plant laccase (oxygen reduction with different organic molecules as donor substrates). In addition, the rate-limiting step was associated with the organic substrate oxidation half-reaction (step 1), as the IET (step 2) rate constant (kIET) is higher, compared to the catalytic rate constant (kcat), and oxygen reduction (step 3) is controlled by diffusion [181]. Results of subsequent research on diverse laccases with several substrates are in accordance with this initial finding, attributing the rate-limiting step to the substrate oxidation reaction. A linear relationship between the laccase catalytic performance, referred to as log(kcat/Km) and the E° difference between the T1 copper site and substrate (ΔE°′) have also been derived from experimental data [182].
Given the catalytic versatility of laccase, novel technological tools have recently been introduced to the theoretical analyses of the molecular basis of laccase action. In recent work using several theoretical approaches, the importance of the fully conserved residues, Asp 206 and His 458, as well as the hydrophobic nature of the substrate recognition site, were confirmed by modelling with two fungal laccases (TvL and CuL). Likewise, the physicochemical properties that influence the Km (the ionization potential, shape, and binding affinity of the substrate) were defined. General results highlight that this catalytic constant depends on substrate binding, as well as enzyme molecular characteristics [152].
Engineering laccases: the quest for a better biocatalyst
As mentioned in other sections of this review, laccases have a broad spectrum of applications in different fields [102]. However, in some cases, the improvement of certain properties is desirable to achieve commercially attractive laccase-based applications or enhance catalyst performance. In this section, modern tools and strategies to achieve stability, optimum operation conditions, and inhibition by reaction medium components, are reviewed.
Laccase activity is lost after a certain number of reaction cycles. For instance, in the presence of free radicals produced from 1e- subtraction of phenols, aromatic amines, or mediators, the enzyme has been reported to eventually inactivate [183,184,185]. Although the mechanism of inactivation by free radicals is not well understood, it may involve oxidation of important residues near the active site that are essential for maintaining protein structure, as demonstrated for other enzymes catalyzing similar reactions [186]. Another important issue is the activity and stability in the presence of organic solvents; due to the hydrophobic nature of most substrates, the presence of co-solvents is necessary to increase substrate availability. Most enzymes are susceptible to activity loss in the presence of organic solvents, owing to denaturation and other mechanisms [187]. Laccases are not exempted, given their instability in the presence of methanol, ethanol, acetone, acetonitrile, as well as other water-soluble solvents.
Mesophilic proteins from organisms living in normal environmental conditions tend to be stable in aqueous media at 25–35 °C and neutral pH. Commercially available laccases come from such organisms, for example laccases from T. versicolor, A. bisporus, P. ostreatus and Rhus vernicifera can be purchased from international companies. A recent review of patents related to laccase applicability can be consulted in Ref. [188]. One strategy to overcome the limitations of mesophilic proteins is to search for those with desired properties, usually focusing on samples from extreme environments. For instance, a bacterial laccase (CotA) from B. clausii, was active in the presence of up to 1 M NaCl over a broader pH range, compared to CotA from B. subtilis [112]. Further, a novel laccase-like protein obtained through metagenome mining of samples from the South China Sea displayed halide- and alkali-tolerance, as well as the ability to decolorize dyes [189], which are properties of interest for industrial applications. Conversely, one may introduce or enhance the desired property by protein engineering. A powerful strategy is laboratory evolution and rational design, usually aided by computational tools [22, 176, 190, 191]. An example of laboratory evolution is the quadruple mutant L386W/G417L/G57F/K317N of the CotA-laccase from B. pumilus W3, where the mutations enhanced thermal stability and improved dye degradation [125].
The optimum pH for enzyme activity may be modified following these strategies. Based on biotechnological requirements, fungal laccases are the most attractive given their high E°. For most of these enzymes, the optimal pH is acidic (pH 3–5); however, some applications require the optimum to be closer to physiological conditions such as neutral pH or even alkaline conditions [102]. For instance, laccase-initiated precursor cross-coupling has been used to generate polymeric compounds for cotton dyeing; this application is limited by low precursor solubility at acidic pH, which requires alkaline conditions to be solubilized. Other uses of laccases that may benefit from enzyme function at neutral conditions are biofuel cells, biosensors, and medical applications, in which the operation conditions (such as sample pH) are close to neutrality. Examples of the modification of optimum pH of laccases are available in the literature. For instance, a laccase from Pycnoporus cinnabarinus was made to evolve during six cycles, using a multiscreening assay with three different substrates, at pH 5 [192]. The resulting variant, α*-3PO, contained five mutations in the mature protein; N208S, R280H, N331D, D341N, and P394H. Compared with the parent enzyme at pH 5, the variant showed a similar or higher catalytic efficiency (kcat/KM) towards model substrates: an 18-fold increase for ABTS (nonphenolic substrate), a 5.7-fold increase for sinapic acid (phenolic substrate), and a 1.6-fold increase for 2,6-dimethoxyphenol (DMP, phenolic substrate). Interestingly, the optimum pH shifted from 2 to 4.5 for ABTS, and from 3 to 4.5 for DMP. Another example is the in vitro evolution of the thermostable laccase mutant MtLT2 from the ascomycete fungus Myceliophthora thermophila; after five cycles, the optimum pH of the resulting variant shifted from 4 to 6.5 [193]. With respect to catalytic efficiency, the variant showed an increase of 31-fold for ABTS and a ninefold increase for DMP. This variant contained one conservative mutation (D530E) away from the active site on the protein surface that may be unrelated to the shift in optimum pH. Another mutation; N109S, occurred in the vicinity of the T2/T3 site, and according to the authors, may have remodeled the configuration of the site, thus conferring the trinuclear cluster a certain resistance to inhibition by OH− [194]. Following a different approach, a shift of 1.4 in optimum pH was obtained after site-directed mutagenesis of a laccase from T. versicolor. The mutagenic site was decided following X-ray structure analyses of a complex between the enzyme and xylidine, a nonphenolic substrate [195]. Residue Asp 206 was observed to strongly interact with the substrate, and when replaced by Asn, the mutant showed an optimum pH of 4.8, compared to 3.4 for the parent enzyme. According to the authors, this shift may result from a lower deprotonation of the substrate at acidic pH in the absence of the carboxylate group, thus, requiring a higher pH in bulk water.
With respect to organic solvent stability, the thermostable laccase mutant MtTL2, from M. thermophila, was made to evolve using a screening assay in the presence of increasing concentrations of organic co-solvent; the activity and stability increased from 20 to 60% v/v [196]. After five cycles, a variant (R2) showing enhanced properties was selected for detailed characterization. Variant R2 displayed improved activity in the presence of co-solvents such as 50% v/v ethanol (19-fold) and 30% v/v acetonitrile (12.6-fold), compared with the parent enzyme, as well as improved tolerance to the presence of organic co-solvents. Impressively, it retained > 80% activity after a 24 h incubation period in the presence of as high as 50% v/v organic co-solvents of different natures, such as acetonitrile, dimethylacetamide, dimethylformamide, dimethylsulfoxide, acetone, methanol, and ethanol. Furthermore, it was more active in the presence of 30% v/v of acetonitrile or ethanol, compared to several native fungal enzymes, such as laccases from T. versicolor, P. cinnabarinus, Coriolopsis gallica, and P. ostreatus. Interestingly, the optimum pH shifted from 4 to 5, with variant R2 retaining 70% activity at pH 6. These improved properties could not be attributed to a higher intrinsic reactivity of the variant, as the E°s of the T1 and T2/T3 sites were similar to the parent enzyme. This variant accumulated four beneficial mutations; one was a conservative mutation of a residue buried in the protein matrix (L429V) and three mutations (E182K, S280N, and N552H) occurred on the protein surface, away from the catalytic site, probably establishing new interactions that could reduce the susceptibility of the protein to denaturation in the presence of organic co-solvents. In a completely different approach, a chimeric laccase was obtained through domain swapping of two laccase mutants from Coriolopsis sp. (OB1), and the above mentioned α*-3PO from P. cinnabarinus [63]. Domains 1 and 3 contain residues coordinating the copper atoms in laccase, while domain 2 bridges domains 1 and 3, and shapes the active site cavity of the T1 copper. The chimeric laccase contained domains 1 and 3 from OB1 and domain 2 from α*-3PO. The resulting enzyme was functional and displayed better tolerance to organic solvents such as 50% v/v ethanol or methanol, compared with the parent variants. Thermal stability was also enhanced in the chimeric laccase. The half-life at 50 °C was 13.8 h, compared with 8.1 h for OB1 and 3.6 h for α*-3PO.
Another property particularly relevant for commercial applications is total turnover (TTN). Total turnover is defined as the amount of converted substrate or generated product per amount of enzyme consumed in the reaction. As mentioned above, enzymes inactivate after a certain number of catalytic cycles or turnovers. Thus, TTN may be interpreted as yield, reflecting the activity and stability of the catalyst during operation, and it is independent of time. In the case of free-radical generating reactions such as phenol oxidation, laccases may inactivate due to oxidation of relevant residues. Using a fungal laccase from C. gallica as a model, quantum mechanics/molecular mechanics (QM/MM) calculations were performed to identify which residues are susceptible to oxidation within the active site cavity of the T1 copper; where free radicals are generated after substrate-electron subtraction [197]. According to theoretical predictions, phenylalanine residues, particularly those exposed to the solvent, are prone to oxidation. Upon replacement by site-directed mutagenesis with nonaromatic apolar residues, single mutants F357L and F413A showed similar kinetic constants with syringaldazine, but higher TTN (2- to 2.6-fold) during 4-methoxyphenol oxidation; a reaction that generates phenoxy free radicals.
In summary, manipulating the intrinsic multifactorial properties of enzymes in general and laccases in particular, is feasible, using modern protein engineering tools. Properties such as optimum pH, organic solvent tolerance, thermotolerance, and operational stability may be targeted to modify desired biocatalyst properties according to water treatment requirements.
Potential application of laccases in water bioremediation
There are more than 15 laccase-based commercial products used in the textile, food and paper industry, which reflects the viability of their industrial use [199]. The most successful and versatile applications have been developed for the textile industry, particularly for the bleaching of indigo-stained denim, such as Denilitel, DeniliteITM, Zylite, and Bleach-cut 3S. Furthermore, the commercialization of the laccase Metzyme, which has high ligninolytic properties, opens the possibility of commercial use of other laccases in the near future [199]. Unfortunately, no commercial laccase products for water treatment are available yet.
Nevertheless, there are experiments that clearly demonstrate the efficacy of laccases to remove pollutants in real wastewater. A laccase from Trametes versicolor has proven to be effective at reducing estrogen concentrations from a municipal wastewater treatment plant in Rolla, Missouri (USA), tested at laboratory-scale. Because of cost effectiveness, laccases may present important advantages over other degrading enzymes in municipal wastewater treatment [200]. The fungal crude extracts from T. pubescens MUT 2400 transformed target molecules and decreased up to 70% of the initial concentration of 2-hydroxybiphenyl, Naproxen, DF, and diethyl phthalate under real municipal wastewater conditions in north-western Italy (Turin) [11]. Also, the positive effect of removing phenol from aqueous refinery samples was demonstrated using laccase SP504 from Novozymes (Franklinton, NC, USA) [201].
The efficacy of immobilized laccase from Coriolopsis gallica was demonstrated for the elimination of pollutants such as BPA, DF and 17-a-ethinylestradiol (EE2) in real samples from the AQUIRIS wastewater treatment plant [202]. While the laccase of the white-rot fungus Coriolopsis polyzona was immobilized onto spherical nanoparticles and tested in effluent water from the wastewater treatment plant Birsfelden, Switzerland, demonstrating that in real wastewater, immobilized laccase can retain its activity over 1 month [203]. In addition, laccase and tyrosinase as combined crosslinked aggregates showed high transformation of acetaminophen to its oligomers in real municipal wastewaters [204]. Even with the presence of colloids and certain ions or other molecules that induce the formation of precipitates that affect enzyme stability, the removal of BPA by laccase was demonstrated in a continuous enzymatic membrane reactor operated with real wastewater [205]. While, Jahangiri et al. [206] demonstrated that laccase from Phoma sp. UHH 5-1-03, cross-linked to polyvinylidene fluoride membranes, removed with high efficiency pharmaceutical compounds such as acetaminophen, bezafibrate, indometacin, ketoprofen, mefenamic acid, and naproxen under the harsh conditions of real wastewater [206]. Furthermore, laccase from Trametes versicolor that was encapsulated in core–shell magnetic copper alginate beads, allowing bead recycling with enhanced field feasibility, efficiently degraded triclosan and recalcitrant dyes in real wastewater from a chemical factory [207].
Along with the positive results however, there are inconveniences, which should be remedied to achieve commercial success. These include the improvement of the search for enzymes using omics tools for the application of microorganisms that present enhanced production of laccases, improvement in the processes of enzyme production taking into account the cost and the environmental impact, the development of scalable and economically viable processes that do not affect the environment, and in which the stability and reuse of enzymes is ensured. Added to this, as in the textile industry, customized formulations may be developed according to the treatment conditions.
In summary, fungal, plant and bacterial laccases are currently used in many bioremediation applications, removing pharmaceutical to industrial pollutants as summarized in Tables 1, 2 and 3. Manipulating the intrinsic multifactorial properties of enzymes in general and laccases in particular, is feasible using modern protein engineering tools. Thus, the generation and/or discovery of laccase variants able to function under a wide range of conditions (pH, temperature, suspended solids, mechanical stress, among others) will help in the development of robust enzymes for commercial products in the pressing issue of using water in a sustainable fashion.
Concluding remarks
There is currently an urgent need to reverse the pollution of global water bodies caused by humans. Despite the diversity of pollutants discharged into waterbodies, such as plastics, herbicides, fertilizers, synthetic dyes, phthalates, and pharmaceuticals, laccases appear to be an efficient biocatalytic tool with the ability to oxidize these molecules, yielding less-toxic and harmful inactive compounds. Some limiting factors, such as the complex composition of contaminated water (high concentrations of salts and/or high pH values), can be overcome by engineering laccases using modern techniques like in vitro evolution and site-directed mutagenesis, enhanced by theoretical tools such as molecular modeling, and dynamic simulations, among others. Not only engineering well-known enzymes, but also searching for new enzymes, with new properties, can provide guidance on the use or design of new structural and biochemical mechanisms of action of the biocatalysts, improving their application in the bioremediation and biotransformation of contaminated waterbodies.
Existing literature suggests that the degradation of emerging pollutants can be achieved (including that of hard-to-degrade molecules, such as nonsteroidal anti-inflammatory drugs or PAHs), but the practical implementation of the degrading bioprocesses is always cost-dependent. Conventional approaches to immobilize laccases, as well as those to extract and purify them, are time-consuming and can be expensive. However, the high-yield production of recombinant laccases, with elevated activity and stability and a decrease in the number of steps for enzyme pre-purification would be advantageous in terms of cost-effectiveness.
Availability of data and materials
All information is included in this published article.
References
Daughton CG. Non-regulated water contaminants: emerging research. Environ Impact Assess Rev. 2004;24:711–32. https://doi.org/10.1016/j.eiar.2004.06.003.
Cabana H, Jiwan JLH, Rozenberg R, Elisashvili V, Penninckx M, Agathos SN, Jones JP. Elimination of endocrine disrupting chemicals nonylphenol and bisphenol A and personal care product ingredient triclosan using enzyme preparation from the white rot fungus Coriolopsis polyzona. Chemosphere. 2007;67:770–8. https://doi.org/10.1016/j.chemosphere.2006.10.037.
Chakroun H, Mechichi T, Martinez MJ, Dhouib A, Sayadi S. Purification and characterization of a novel laccase from the ascomycete Trichoderma atroviride: application on bioremediation of phenolic compounds. Process Biochem. 2010;45:507–13. https://doi.org/10.1016/j.procbio.2009.11.009.
Corcoran E, Nellemann C, Baker E, Bos R, Osborn D, Savelli H. Sick water? The central role of wastewater management in sustainable development. A rapid response assessment. United Nations Environment Programme: Nairobi; 2010. ISBN 978-82-7701-075-5.
Bilal M, Asgher M, Parra-Saldivar R, Hu H, Wang W, Zhang X, Iqbal HMN. Immobilized ligninolytic enzymes: an innovative and environmental responsive technology to tackle dye-based industrial pollutants—a review. Sci Total Environ. 2017;576:646–59. https://doi.org/10.1016/j.scitotenv.2016.10.137.
Klaper R, Welch LC. Emerging contaminant threats and the Great Lakes: existing science, estimating relative risk and determining policies. Chicago: Alliance for the Great Lakes; 2011.
Garcia-Morales R, Rodríguez-Delgado M, Gomez-Mariscal K, Orona-Navar C, Hernandez-Luna C, Torres E, Parra R, Cárdenas-Chávez D, Mahlknecht J, Ornelas-Soto N. Biotransformation of endocrine-disrupting compounds in groundwater: bisphenol A, nonylphenol, ethynylestradiol and triclosan by a laccase cocktail from Pycnoporus sanguineus CS43. Water Air Soil Pollut. 2015;226:251. https://doi.org/10.1007/s11270-015-2514-3.
Nguyen LN, Hai FI, Price WE, Leusch FD, Roddick F, McAdam EJ, Magram SF, Nghiem LD. Continuous biotransformation of bisphenol A and diclofenac by laccase in an enzymatic membrane reactor. Int Biodeter Biodegr. 2014;95:25–32. https://doi.org/10.1016/j.ibiod.2014.05.017.
Gasser CA, Ammann EM, Shahgaldian P, Corvini PFX. Laccases to take on the challenge of emerging organic contaminants in wastewater. Appl Microbiol Biotechnol. 2014;98:9931–52. https://doi.org/10.1007/s00253-014-6177-6.
Rodríguez-Delgado M, Orona-Navar C, García-Morales R, Hernandez-Luna C, Parra R, Mahlknecht J, Ornelas-Soto N. Biotransformation kinetics of pharmaceutical and industrial micropollutants in groundwaters by a laccase cocktail from Pycnoporus sanguineus CS43 fungi. Int Biodeter Biodegr. 2016;108:34–41. https://doi.org/10.1016/j.ibiod.2015.12.003.
Spina F, Cordero C, Schilirò T, Sgorbini B, Pignata C, Gilli G, Bicci C, Varese GC. Removal of micropollutants by fungal laccases in model solution and municipal wastewater: evaluation of estrogenic activity and ecotoxicity. J Clean Prod. 2015;100:185–94. https://doi.org/10.1016/j.jclepro.2015.03.047.
Deblonde T, Cossu-Leguille C, Hartemann P. Emerging pollutants in wastewater: a review of the literature. Int J Hyg Environ Health. 2011;214:442–8. https://doi.org/10.1016/j.ijheh.2011.08.002.
Chatha SAS, Asgher M, Iqbal HMN. Enzyme-based solutions for textile processing and dye contaminant biodegradation—a review. Environ Sci Pollut Res Int. 2017;24:14005–18. https://doi.org/10.1007/s11356-017-8998-1.
Barber LB, Loyo-Rosales JE, Rice CP, Minarik TA, Oskouie AK. Endocrine disrupting alkylphenolic chemicals and other contaminants in wastewater treatment plant effluents, urban streams, and fish in the Great Lakes and upper Mississippi River Regions. Sci Total Environ. 2015;517:195–206. https://doi.org/10.1016/j.scitotenv.2015.02.035.
Giardina P, Faraco V, Pezzella C, Piscitelli A, Vanhulle S, Sannia G. Laccases: a never-ending story. Cell Mol Life Sci. 2010;67:69–385. https://doi.org/10.1007/s00018-009-0169-1.
Riva S. Laccases: blue enzymes for green chemistry. Trends Biotechnol. 2016;24:219–26. https://doi.org/10.1016/j.tibtech.2006.03.006.
Kim C, Lorenz WW, Hoopes JT, Dean JF. Oxidation of phenolate siderophores by the multicopper oxidase encoded by the Escherichia coli yacK gene. J Bacteriol. 2001;183:4866–75. https://doi.org/10.1128/JB.183.16.4866-4875.2001.
Morozova OV, Shumakovich GP, Shleev SV, Yaropolov YI. Laccase-mediator systems and their applications: a review. Appl Biochem Microbiol. 2007;43:523–35. https://doi.org/10.1134/S0003683807050055.
Upadhyay P, Shrivastava R, Agrawal PK. Bioprospecting and biotechnological applications of fungal laccase. 3 Biotech. 2016;6:15. https://doi.org/10.1007/s13205-015-0316-3.
Mayer AM, Staples RC. Laccase: new functions for an old enzyme. Phytochemistry. 2002;60:551–65. https://doi.org/10.1016/S0031-9422(02)00171-1.
Santhanam N, Vivanco JM, Decker SR, Reardon KF. Expression of industrially relevant laccases: prokaryotic style. Trends Biotechnol. 2011;29:480–9. https://doi.org/10.1016/j.tibtech.2011.04.005.
Mate DM, Alcalde M. Laccase engineering: from rational design to directed evolution. Biotechnol Adv. 2015;33:25–40. https://doi.org/10.1016/j.biotechadv.2014.12.007.
Viswanath B, Rajesh B, Janardhan A, Kumar AP, Narasimha G. Fungal laccases and their applications in bioremediation. Enzyme Res. 2014;2014:1–21. https://doi.org/10.1155/2014/163242.
Bertrand G. Sur la presence simultanee de la laccase et de la tyrosinase dans le suc de quelques champignons. CR Hebd Seances Acad Sci. 1896;123:463–5.
Strong P, Claus H. Laccase: a review of its past and its future in bioremediation. Crit Rev Environ Sci Technol. 2011;41:373–434. https://doi.org/10.1080/10643380902945706.
Kiliç N, Nasiri F, Cansaran-Duman D. Fungal laccase enzyme applications in bioremediation of polluted wastewater. Phytoremediation. New York: Springer; 2016. p. 201–9.
Thurston CF. The structure and function of fungal laccases. Microbiology. 1994;140:19–26. https://doi.org/10.1099/13500872-140-1-19.
Alcalde M. Laccases: biological functions, molecular structure and industrial applications. In: Polaina J, MacCabe A, editors. Industrial enzymes: structure, function and applications. Dordrecht: Springer; 2007. p. 461–76. ISBN 978-1-4020-5377-1.
Baldrian P. Fungal laccases–occurrence and properties. FEMS Microbiol Rev. 2006;30:215–42. https://doi.org/10.1111/j.1574-4976.2005.00010.x.
Rivera-Hoyos CM, Morales-Álvarez ED, Poutou-Piñales RA, Pedroza-Rodríguez AM, RodrÍguez-Vázquez R, Delgado-Boada JM. Fungal laccases. Fungal Biol Rev. 2013;27:67–82. https://doi.org/10.1016/j.fbr.2013.07.001.
Morozova O, Shumakovich G, Gorbacheva M, Shleev S, Yaropolov A. “Blue” laccases. Biochemistry (Mosc). 2007;72:1136–50. https://doi.org/10.1134/S0006297907100112.
Maestre-Reyna M, Liu WC, Jeng WY, Lee CC, Hsu CA, Wen TN, Wang AHJ, Shyur LF. Structural and functional roles of glycosylation in fungal laccase from Lentinus sp. PLoS ONE. 2015;10:e0120601. https://doi.org/10.1371/journal.pone.0120601.
Shleev SV, Morozova OV, Nikitina OV, Gorshina ES, Rusinova TV, Serezhenkov VA, Burbaev DS, Gazaryan IG, Yaropolov AI. Comparison of physico-chemical characteristics of four laccases from different basidiomycetes. Biochimie. 2004;86:693–710. https://doi.org/10.1016/j.biochi.2004.08.005.
Rodgers CJ, Blanford CF, Giddens SR, Skamnioti P, Armstrong FA, Gurr SJ. Designer laccases: a vogue for high-potential fungal enzymes? Trends Biotechnol. 2010;28:63–72. https://doi.org/10.1016/j.tibtech.2009.11.001.
Maté D, García-Ruiz E, Camarero S, Alcalde M. Directed evolution of fungal laccases. Curr Genom. 2011;12:113–22. https://doi.org/10.2174/138920211795564322.
Agrawal K, Chaturvedi V, Verma P. Fungal laccase discovered but yet undiscovered. Bioresour Bioprocess. 2018;5:4. https://doi.org/10.1186/s40643-018-0190-z.
Kuramitz H, Saitoh J, Hattori T, Tanaka S. Electrochemical removal of p-nonylphenol form dilute solutions using a carbon fiber anode. Water Res. 2002;36:3323–9. https://doi.org/10.1016/S0043-1354(02)00040-4.
Tavares APM, Cristóvão RO, Gamelas JAF, Loureiro JM, Boaventura RAR, Macedo EA. Sequential decolourization of reactive textile dyes by laccaase mediator system. J Chem Technol Biotechnol. 2009;84:442–6. https://doi.org/10.1002/jctb.2060.
Acevedo F, Pizzul L, del Pilar Castillo M, Cuevas R, Diez MC. Degradation of polycyclic aromatic hydrocarbons by the Chilean white-rot fungus Anthracophyllum discolor. J Hazard Mater. 2011;185:212–9. https://doi.org/10.1016/j.jhazmat.2010.09.020.
Uribe-Alvarez C, Ayala M, Perezgasga L, Naranjo L, Urbina H, Vazquez-Duhalt R. First evidence of mineralization of petroleum asphaltenes by a strain of Neosartorya fischeri. Microb Biotechnol. 2011;4:663–72. https://doi.org/10.1111/j.1751-7915.2011.00269.x.
Ramírez-Cavazos LI, Junghanns C, Ornelas-Soto N, Cárdenas-Chávez DL, Hernández-Luna C, Demarche P, Enaud E, García-Morales R, Agathos SN, Parra R. Purification and characterization of two thermostable laccases from Pycnoporus sanguineus and potential role in degradation of endocrine disrupting chemicals. J Mol Cat B Enzym. 2014;108:32–42. https://doi.org/10.1016/j.molcatb.2014.06.006.
Barrios-Estrada C, de Jesús Rostro-Alanis M, Parra AL, Belleville MP, Sanchez-Marcano J, Iqbal HM, Parra-Saldívar R. Potentialities of active membranes with immobilized laccase for bisphenol A degradation. Int J Biol Macromol. 2018;108:837–44. https://doi.org/10.1016/j.ijbiomac.2017.10.177.
Carabajal M, Perullini M, Jobbágy M, Ullrich R, Hofrichter M, Levin L. Removal of phenol by immobilization of Trametes versicolor in silica–alginate–fungus biocomposites and loofa sponge. CLEAN Soil Air Water. 2016;44:180–8. https://doi.org/10.1002/clen.201400366.
Gaitan IJ, Medina SC, González JC, Rodríguez A, Espejo ÁJ, Osma JF, Sarria V, Alméciga-Díaz CJ, Sánchez OF. Evaluation of toxicity and degradation of a chlorophenol mixture by the laccase produced by Trametes pubescens. Bioresour Technol. 2011;102:3632–5. https://doi.org/10.1016/j.biortech.2010.11.040.
Kunamneni A, Ghazi I, Camarero S, Ballesteros A, Plou FJ, Alcalde M. Decolorization of synthetic dyes by laccase immobilized on epoxy-activated carriers. Process Biochem. 2008;43:169–78. https://doi.org/10.1016/j.procbio.2007.11.009.
Simões MF, Maiorano AE, dos Santos JG, Peixoto L, de Souza RFB, Neto AO, Brito AG, Ottoni CA. Microbial fuel cell-induced production of fungal laccase to degrade the anthraquinone dye remazol brilliant blue R. Environ Chem Lett. 2019. https://doi.org/10.1007/s10311-019-00876-y.
Palazzolo MA, Postemsky PD, Kurina-Sanz M. From agro-waste to tool: biotechnological characterization and application of Ganoderma lucidum E47 laccase in dye decolorization. 3 Biotech. 2019;9(6):213. https://doi.org/10.1007/s13205-019-1744-2.
Iark D, dos Reis Buzzo AJ, Garcia JAA, Côrrea VG, Helm CV, Corrêa RCG, Peralta RA, Moreira RFPM, Bracht A, Peralta RM. Enzymatic degradation and detoxification of azo dye Congo red by a new laccase from Oudemansiella canarii. Bioresour Technol. 2019. https://doi.org/10.1016/j.biortech.2019.121655.
Garcia LF, Lacerda MF, Thomaz DV, de Souza JC, Pereira M, de Souza G, Schimidt F, Santiago MF. Optimization of laccase-alginate-chitosan-based matrix toward 17 α-ethinylestradiol removal. Prep Biochem Biotechnol. 2019;49:375–83. https://doi.org/10.1080/10826068.2019.1573195.
Alharbi SK, Nghiem LD, van de Merwe JP, Leusch FD, Asif MB, Hai FI, Price WE. Degradation of diclofenac, trimethoprim, carbamazepine, and sulfamethoxazole by laccase from Trametes versicolor: transformation products and toxicity of treated effluent. Biocatal Biotransform. 2019. https://doi.org/10.1080/10242422.2019.1580268.
Pulicharla R, Das RK, Brar SK, Drogui P, Surampalli RY. Degradation kinetics of chlortetracycline in wastewater using ultrasonication assisted laccase. Chem Eng J. 2018;347:828–35. https://doi.org/10.1016/j.cej.2018.04.162.
Singh SK, Khajuria R, Kaur L. Biodegradation of ciprofloxacin by white rot fungus Pleurotus ostreatus. 3 Biotech. 2017;7(1):69. https://doi.org/10.1007/s13205-017-0684-y.
Navada KK, Kulal A. Enzymatic degradation of chloramphenicol by laccase from Trametes hirsuta and comparison among mediators. Int Biodeter Biodegr. 2019;138:63–9. https://doi.org/10.1016/j.ibiod.2018.12.012.
Fukuda T, Uchida H, Takashima Y, Uwajima T, Kawabata T, Suzuki M. Degradation of bisphenol A by purified laccase from Trametes villosa. Biochem Biophys Res Commun. 2001;284:704–6. https://doi.org/10.1006/bbrc.2001.502.
Becker D, Rodriguez-Mozaz S, Insa S, Schoevaart R, Barcelo D, de Cazes M, Belleville MP, Sanchez-Marcano J, Misovic A, Oehlmann J, Wagner M. Removal of endocrine disrupting chemicals in wastewater by enzymatic treatment with fungal laccases. Org Process Res Dev. 2017;21(4):480–91. https://doi.org/10.1021/acs.oprd.6b00361.
Golveia JCS, Santiago MF, Sales PTF, Sartoratto A, Ponezi AN, Thomaz DV, Gil ES, Bara MTF. Cupuaçu (Theobroma grandiflorum) residue and its potential application in the bioremediation of 17-Α-ethinylestradiol as a Pycnoporus sanguineus laccase inducer. Prep Biochem Biotechnol. 2018;48(6):541–8. https://doi.org/10.1080/10826068.2018.1466161.
Casas N, Parella T, Vicent T, Caminal G, Sarrà M. Metabolites from the biodegradation of triphenylmethane dyes by Trametes versicolor or laccase. Chemosphere. 2009;75(10):1344–9. https://doi.org/10.1016/j.chemosphere.2009.02.029.
Grassi E, Scodeller P, Filiel N, Carballo R, Levin L. Potential of Trametes trogii culture fluids and its purified laccase for the decolorization of different types of recalcitrant dyes without the addition of redox mediators. Int Biodeter Biodegr. 2011;65:635–43. https://doi.org/10.1016/j.ibiod.2011.03.007.
Piscitelli A, Pezzella C, Giardina P, Faraco V, Sannia G. Heterologous laccase production and its role in industrial applications. Bioeng Bugs. 2010;1:254–64. https://doi.org/10.4161/bbug.1.4.11438.
Salony GN, Baranwal R, Chhabra M, Mishra S, Chaudhuri TK, Bisaria VS. Laccase of Cyathus bulleri: structural, catalytic characterization and expression in Escherichia coli. Biochim Biophys Acta. 2008;1784:259–68. https://doi.org/10.1016/j.bbapap.2007.11.006.
Klonowska A, Gaudin C, Asso M, Fournel A, Reglier M, Tron T. LAC3, a new low redox potential laccase from Trametes sp. strain C30 obtained as a recombinant protein in yeast. Enzyme Microb Technol. 2005;36:34–41. https://doi.org/10.1016/j.enzmictec.2004.03.022.
Colao MC, Lupino S, Garzillo AM, Buonocore V, Ruzzi M. Heterologous expression of lcc1 gene from Trametes trogii in Pichia pastoris and characterization of the recombinant enzyme. Microb Cell Fact. 2006;5:31. https://doi.org/10.1186/1475-2859-5-31.
Madzak C, Otterbein L, Chamkha M, Moukha S, Asther M, Gaillardin C, Beckerich JM. Heterologous production of a laccase from the basidiomycete Pycnoporus cinnabarinus in the dimorphic yeast Yarrowia lipolytica. FEMS Yeast Res. 2005;5:635–46. https://doi.org/10.1016/j.femsyr.2004.10.009.
Pardo I, Rodríguez-Escribano D, Aza P, De Salas F, Martínez AT, Camarero S. A highly stable laccase obtained by swapping the second cupredoxin domain. Sci Rep. 2018;8:15669. https://doi.org/10.1038/s41598-018-34008-3.
Yaver DS, Xu F, Golightly EJ, Brown KM, Brown SH, Rey MW, Schneider P, Halkier T, Mondorf K, Dalboge H. Purification, characterization, molecular cloning, and expression of two laccase genes from the white rot basidiomycete Trametes villosa. Appl Environ Microbiol. 1996;62:834–41.
Bohlin C, Jönsson LJ, Roth R, Van Zyl WH. Heterologous expression of Trametes versicolor laccase in Pichia pastoris and Aspergillus niger. Appl Biochem Biotechnol. 2006;129–132:195–214. https://doi.org/10.1007/978-1-59745-268-7_15.
Balcázar-López E, Méndez-Lorenzo LH, Batista-García RA, Esquivel-Naranjo U, Ayala M, Kumar VV, Savary O, Cabana H, Herrera-Estrella A, Folch-Mallol JL. Xenobiotic compounds degradation by heterologous expression of a Trametes sanguineus laccase in Trichoderma atroviride. PLoS ONE. 2016;11:e0147997. https://doi.org/10.1371/journal.pone.0147997.
Wang B, Yan Y, Xu J, Fu X, Han H, Gao J, Li Z, Wang L, Tian Y, Peng R. Heterologous expression and characterization of a laccase from Laccaria bicolor in Pichia pastoris and Arabidopsis thaliana. J Microbiol Biotechnol. 2018;28:2057–63. https://doi.org/10.1080/13102818.2015.1104261.
Bailey MR, Woodard SL, Callaway E, Beifuss K, Magallanes-Lundback M, Lane JR, Horn ME, Mallubhotla H, Delaney DD, Ward M, Van Gastel F, Howard JA, Hood EE. Improved recovery of active recombinant laccase from maize seed. Appl Microb Biotechnol. 2004;63:390–7. https://doi.org/10.1007/s00253-003-1362-z.
Campos PA, Levin LN, Wirth SA. Heterologous production, characterization and dye decolorization ability of a novel thermostable laccase isoenzyme from Trametes trogii BAFC 463. Process Biochem. 2016;51:895–903. https://doi.org/10.1016/j.procbio.2016.03.015.
Darvishi F, Moradi M, Jolivalt C, Madzak C. Laccase production from sucrose by recombinant Yarrowia lipolytica and its application to decolorization of environmental pollutant dyes. Ecotox Environ Safe. 2018;165:278–83. https://doi.org/10.1016/j.ecoenv.2018.09.026.
Yoshida H. Chemistry of lacquer (Urushi) part 1. J Chem Soc. 1883;43:472–86. https://doi.org/10.1039/ct8834300472.
Frasconi M, Favero G, Boer H, Koivula A, Mazzei F. Kinetic and biochemical properties of high and low redox potential laccases from fungal and plant origin. Biochim Biophys Acta. 2010;1804:899–908. https://doi.org/10.1016/j.bbapap.2009.12.018.
Sterjiades R, Jeffrey FDD, Eriksson KEL. Laccase from sycamore maple (Acer pseudoplatanus) polymerizes monolignols. Plant Physiol. 1992;99:1162–8. https://doi.org/10.1104/pp.99.3.1162.
Dwivedi UN, Singh P, Pandey VP, Kumar A. Structure-function relationship among bacterial, fungal and plant laccases. J Mol Catal B Enzym. 2011;68:117–28. https://doi.org/10.1016/j.molcatb.2010.11.002.
Zhao Q, Nakashima J, Chen F, Yin YB, Fu CX, Yun JF, Shao H, Wang XQ, Wang ZY, Dixon RA. Laccase is necessary and nonredundant with peroxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell. 2013;25:3976–87. https://doi.org/10.1105/tpc.113.117770.
Tobimatsu Y, Schuetz M. Lignin polymerization: how do plants manage the chemistry so well? Curr Opin Biotechnol. 2019;56:75–81. https://doi.org/10.1016/j.copbio.2018.10.001.
Liang M, Davis E, Gardner D, Cai X, Wu Y. Involvement of AtLAC15 in lignin synthesis in seeds and in root elongation of Arabidopsis. Planta. 2006;224:1185–96. https://doi.org/10.1007/s00425-006-0300.
Nigam D, Kavita P, Tripathi RK, Ranjan A, Goel R, Asif M, Shukla A, Singh G, Rana D, Sawant SV. Transcriptome dynamics during fibre development in contrasting genotypes of Gossypium hirsutum L. Plant Biotechnol J. 2014;12:204–18. https://doi.org/10.1111/pbi.12129.
Balasubramanian VK, Rai KM, Thu SW, Hii MM, Mendu V. Genome-wide identification of multifunctional laccase gene family in cotton (Gossypium spp.); expression and biochemical analysis during fiber development. Sci Rep. 2016;6:34309. https://doi.org/10.1038/srep34309.
Cai X, Davis EJ, Ballif J, Liang M, Bushman E, Haroldsen V, Torabinejad J, Wu Y. Mutant identification and characterization of the laccase gene family in Arabidopsis. J Exp Bot. 2006;57:2563–9. https://doi.org/10.1093/jxb/erl022.
Cho HY, Lee C, Hwang SG, Park YC, Lim HL, Jang CS. Overexpression of the OsChI1 gene, encoding a putative laccase precursor, increases tolerance to drought and salinity stress in transgenic Arabidopsis. Gene. 2014;552:98–105. https://doi.org/10.1016/j.gene.2014.09.018.
Liu Q, Luo L, Wang X, Shen Z, Zheng L. Comprehensive analysis of rice laccase gene (OsLAC) family and ectopic expression of OsLAC10 enhances tolerance to copper stress in Arabidopsis. Int J Mol Sci. 2017;18:209. https://doi.org/10.3390/ijms18020209.
Wang GD, Li QJ, Luo B, Chen XY. Ex planta phytoremediation of trichlorophenol and phenolic allelo chemicals via an engineered secretory laccase. Nat Biotechnol. 2004;22:893–7. https://doi.org/10.1038/nbt982.
Watharkar AD, Kadam SK, Khandare RV, Kolekar PD, Jeon BH, Jadhav JP, Govindwar SP. Asparagus densiflorus in a vertical subsurface flow phytoreactor for treatment of real textile effluent: a lab to land approach for in situ soil remediation. Ecotoxicol Environ Saf. 2018;161:70–7. https://doi.org/10.1016/j.ecoenv.2018.05.078.
Kagalkar AN, Khandare RV, Govindwar SP. Textile dye degradation potential of plant laccase significantly enhances upon augmentation with redox mediators. RSC Adv. 2015;5:80505–17. https://doi.org/10.1039/c5ra12454a.
Telke AA, Kagalkar AN, Jagtap UB, Desai NS, Bapat VA, Govidwar SP. Biochemical characterization of laccase from hairy root culture of Brassica juncea L. and role of redox mediators to enhance its potential for the decolorization of textile dyes. Planta. 2011;234:1137–49. https://doi.org/10.1007/s00425-011-1469-x.
Kabra AN, Khandare RV, Kurade MB, Govindwar SP. Phytoremediation of a sulphonated azo dye Green HE4B by Glandularia pulchella (Sweet) Tronc. (Moss Verbena). Environ Sci Pollut Res Int. 2011;18:1360–73. https://doi.org/10.1007/s11356-011-0491-7.
Rane NR, Chandanshive VV, Watharkar AD, Khandare RV, Patil TS, Pawar PK, Govindwar SP. Phytoremediation of sulfonated Remazol Red dye and textile effluents by Alternanthera philoxeroides: an anatomical, enzymatic and pilot scale study. Water Res. 2015;83:271–81. https://doi.org/10.1016/j.watres.2015.06.046.
Chandanshive VV, Kadam SK, Khandare RV, Kurade MB, Jeon BH, Jadhav JP, Govindwar SP. In situ phytoremediation of dyes from textile wastewater using garden ornamental plants, effect on soil quality and plant growth. Chemosphere. 2018;210:968–76. https://doi.org/10.1016/j.chemosphere.2018.07.064.
Huang MT, Lu YC, Zhang S, Luo F, Yang H. Rice (Oryza sativa) laccases involved in modification and detoxification of herbicides atrazine and isoproturon residues in plants. J Agric Food Chem. 2016;64:6397–406. https://doi.org/10.1021/acs.jafc.6b02187.
Chiaiese P, Palomba F, Galante C, Esposito S, de Biasi MG, Filippone E. Transgenic tobacco plants expressing a fungal laccase are able to reduce phenol content from olive mill wastewaters. Int J Phytoremediat. 2012;14:835–44. https://doi.org/10.1080/15226514.2011.628715.
de Wilde C, Uzan E, Zhou Z, Kruus K, Andberg M, Buchert J, Record E, Asther M, Lomascolo A. Transgenic rice as a novel production system for Melanocarpus and Pycnoporus laccases. Transgenic Res. 2008;17:515–27. https://doi.org/10.1007/s11248-007-9124-9.
Sonoki T, Kajita S, Ikeda S, Uesugi M, Tatsumi K, Katayama Y, Iimura Y. Transgenic tobacco expressing fungal laccase promotes the detoxification of environmental pollutants. Appl Microb Biotechnol. 2005;67:138–42. https://doi.org/10.1007/s00253-004-1770-8.
De-Jesús-García R, Folch-Mallol JL, Dubrovsky JG. Transgenic callus of Nicotiana glauca stably expressing a fungal laccase maintains its growth in presence of organic contaminants. Plant Cell Tissue Organ Cult. 2019;138(2):311–24. https://doi.org/10.1007/s11240-019-01626-2.
Du MH, Yan ZW, Hao YJ, Yan ZT, Si FL, Chen B, Qiao L. Suppression of Laccase 2 severely impairs cuticle tanning and pathogen resistance during the pupal metamorphosis of Anopheles sinensis (Diptera: Culicidae). Parasites Vectors. 2017;10(1):171. https://doi.org/10.1186/s13071-017-2118-4.
Dittmer NT, Kanost MR. Insect multicopper oxidases: diversity, properties, and physiological roles. Insect Biochem Mol Biol. 2010;40:179–88. https://doi.org/10.1016/j.ibmb.2010.02.006.
Givaudan A, Effosse A, Faure D, Bouillant M-L, Bally R, Potier P. Polyphenol oxidase in Azospirillum lipoferum isolated from rice rhizosphere: evidence for laccase activity in non-motile strains of Azospirillum lipoferum. FEMS Microbiol Lett. 1993;108:205–10. https://doi.org/10.1111/j.1574-6968.1993.tb06100.x.
Liu Y, Huang L, Guo W, Jia L, Fu Y, Gui S, Lu F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochem. 2017;53:125–34. https://doi.org/10.1016/j.procbio.2016.11.015.
Singh G, Bhalla A, Kaur P, Capalash N, Sharma P. Laccase from prokaryotes: a new source for an old enzyme. Rev Environ Sci Biotechnol. 2011;10:309–26. https://doi.org/10.1007/s11157-011-9257-4.
Uthandi S, Saad B, Humbard MA, Maupin-Furlow JA. LccA, an archaeal laccase secreted as a highly stable glycoprotein into the extracellular medium by Haloferax volcanii. Appl Environ Microbiol. 2010;76:733–43. https://doi.org/10.1128/AEM.01757-09.
Mate DM, Alcalde M. Laccase: a multi-purpose biocatalyst at the forefront of biotechnology. Microb Biotechnol. 2017;10:1457–67. https://doi.org/10.1111/1751-7915.12422.
Martins LO, Durão P, Brissos V, Lindley PF. Laccases of prokaryotic origin: enzymes at the interface of protein science and protein technology. Cell Mol Life Sci. 2015;72:911–22. https://doi.org/10.1007/s00018-014-1822-x.
Sondhi S, Sharma P, Saini S, Puri N, Gupta N. Purification and characterization of an extracellular, thermo-alkali-stable, metal tolerant laccase from Bacillus tequilensis SN4. PLoS ONE. 2014;9:1–10. https://doi.org/10.1371/journal.pone.0096951.
Guan ZB, Liao Q, Wang HR, Chen Y, Liao XR. Bacterial laccases: promising biological green tools for industrial applications. Cell Mol Life Sci. 2018;75:3569–92. https://doi.org/10.1007/s00018-018-2883-z.
Chauhan PS, Goradia B, Saxena A. Bacterial laccase: recent update on production, properties and industrial applications. 3 Biotech. 2017;7:323. https://doi.org/10.1007/s13205-017-0955-7.
Enguita FJ, Martins LO, Henriques AO, Carrondo MA. Crystal structure of a bacterial endospore coat component. J Biol Chem. 2003;278:19416–25. https://doi.org/10.1074/jbc.M301251200.
Roberts SA, Weichsel A, Grass G, Thakali K, Hazzard JT, Tollin G, Rensing C, Montfort WR. Crystal structure and electron transfer kinetics of CueO, a multicopper oxidase required for copper homeostasis in Escherichia coli. Proc Natl Acad Sci. 2002;99:2766–71. https://doi.org/10.1073/pnas.052710499.
Hakulinen N, Rouvinen J. Three-dimensional structures of laccases. Cell Mol Life Sci. 2015;72:857–68. https://doi.org/10.1007/s00018-014-1827-5.
Majumdar S, Lukk T, Solbiati JO, Bauer S, Nair SK, Gronan JE, Gerlt JA. Roles of small laccases from Streptomyces in lignin degradation. Biochemistry. 2014;53:4047–58. https://doi.org/10.1021/bi500285t.
Lawton TJ, Sayavedra-Soto LA, Arp DJ, Rosenzweig AC. Crystal structure of a two-domain multicopper oxidase implications for the evolution of multicopper blue proteins. J Biol Chem. 2009;284:10174–80. https://doi.org/10.1074/jbc.M900179200.
Brander S, Mikkelsen JD, Kepp KP. Characterization of an alkali-and halide-resistant laccase expressed in E. coli: CotA from Bacillus clausii. PLoS ONE. 2014;9:e99402. https://doi.org/10.1371/journal.pone.0099402.
Songulashvili G, Jimenéz-Tobón GA, Jaspers C, Penninckx MJ. Immobilized laccase of Cerrena unicolor for elimination of endocrine disruptor micropollutants. Fungal Biol. 2012;116:883–9. https://doi.org/10.1016/j.funbio.2012.05.005.
Trubitsina LI, Tishchenko SV, Gabdulkhakov AG, Lisov AV, Zakharova MV, Leontievsky AA. Structural and functional characterization of two-domain laccase from Streptomyces viridochromogenes. Biochimie. 2015;112:151–9. https://doi.org/10.1016/j.biochi.2015.03.005.
Jimenez-Juarez N, Baeza A, Roman-Miranda R, Vazquez-Duhalt R, Valderrama B, Sánchez-Amat A. Alkali and halide-resistant catalysis by the multipotent oxidase from Marinomonas mediterranea. J Biotechnol. 2005;117:73–82. https://doi.org/10.1016/j.jbiotec.2005.01.002.
Dubé E, Shareck F, Hurtubise Y, Daneault C, Beauregard M. Homologous cloning, expression, and characterization of a laccase from Streptomyces coelicolor and enzymatic decolourisation of an indigo dye. Appl Microbiol Biotechnol. 2008;79:597–603. https://doi.org/10.1007/s00253-008-1475-5.
Ruijssenaars HJ, Hartmans S. A cloned Bacillus halodurans multicopper oxidase exhibiting alkaline laccase activity. Appl Microbiol Biotechnol. 2004;65:177–82. https://doi.org/10.1007/s00253-004-1571-0.
Guan Z, Song C, Zhang N, Zhou W. Overexpression, characterization, and dye-decolorizing ability of a thermostable, pH-stable, and organic solvent-tolerant laccase from Bacillus pumilus W3. J Mol Catal B Enzym. 2014;101:1–6. https://doi.org/10.1016/j.molcatb.2013.11.009.
Theerachat M, Guieysse D, Morel S, Remaud-Siméon M, Chulalaksananukul W. Laccases from marine organisms and their applications in the biodegradation of toxic and environmental pollutants: a review. Appl Biochem Biotechnol. 2019;187:583–611. https://doi.org/10.1007/s12010-018-2829-9.
Callejón S, Sendra R, Ferrer S, Pardo I. Recombinant laccase from Pediococcus acidilactici CECT 5930 with ability to degrade tyramine. PLoS ONE. 2017;12:1–19. https://doi.org/10.1371/journal.pone.0186019.
Guan Z, Zhang N, Song C. Molecular cloning, characterization and dye-decolorizing ability of a temperature- and pH-stable laccase from Bacillus subtilis X1. Appl Biochem Biotechnol. 2014;172:1147–57. https://doi.org/10.1007/s12010-013-0614-3.
Ihssen J, Reiss R, Luchsinger R, Thöny-Meyer L, Richter M. Biochemical properties and yields of diverse bacterial laccase-like multicopper oxidases expressed in Escherichia coli. Sci Rep. 2015;5:1–13. https://doi.org/10.1038/srep10465.
Tonin F, Rosini E, Piubelli L, Sanchez-Amat A, Pollegioni L. Different recombinant forms of polyphenol oxidase A, a laccase from Marinomonas mediterranea. Protein Expr Purif. 2016;123:60–9. https://doi.org/10.1016/j.pep.2016.03.011.
Zeng J, Zhu Q, Wu Y, Lin X. Oxidation of polycyclic aromatic hydrocarbons using Bacillus subtilis CotA with high laccase activity and copper independence. Chemosphere. 2016;148:1–7. https://doi.org/10.1016/j.chemosphere.2016.01.019.
Luo Q, Wang KQ, Liao XR, Guan ZB, Chen Y, Cai YJ, Xia J. Functional expression enhancement of Bacillus pumilus CotA-laccase mutant WLF through site-directed mutagenesis. Enzyme Microb Technol. 2018;109:11–9. https://doi.org/10.1016/j.enzmictec.2017.07.013.
Liu H, Cheng Y, Du B, Tong C, Liang S, Han S, Zheng S, Lin Y. Overexpression of a novel thermostable and chloride-tolerant laccase from Thermus thermophilus SG0.5JP17-16 in Pichia pastoris and its application in synthetic dye decolorization. PLoS ONE. 2015;10:1–14. https://doi.org/10.1371/journal.pone.0119833.
Lu L, Wang TN, Xu TF, Wang JY, Wang CL, Zhao M. Cloning and expression of thermo-alkali-stable laccase of Bacillus licheniformis in Pichia pastoris and its characterization. Bioresour Technol. 2013;134:81–6. https://doi.org/10.1016/j.biortech.2013.02.015.
Ma X, Liu L, Li Q, Liu Y, Yi L, Ma L, Zhai C. High-level expression of a bacterial laccase, CueO from Escherichia coli K12 in Pichia pastoris GS115 and its application on the decolorization of synthetic dyes. Enzyme Microb Technol. 2017;103:34–41. https://doi.org/10.1016/j.enzmictec.2017.04.004.
Argüello JM, Raimunda D, Padilla-Benavides T. Mechanisms of copper homeostasis in bacteria. Front Cell Infect Microbiol. 2013;3:1–14. https://doi.org/10.3389/fcimb.2013.00073.
Outten FW, Huffman DL, Hale JA, O’Halloran TV. The independent cue and cus systems confer copper tolerance during aerobic and anaerobic growth in Escherichia coli. J Biol Chem. 2001;276:30670–7. https://doi.org/10.1074/jbc.M104122200.
Rensing C, Grass G. Escherichia coli mechanisms of copper homeostasis in a changing environment. FEMS Microbiol Rev. 2003;27:197–213. https://doi.org/10.1016/S0168-6445(03)00049-4.
Durão P, Chen Z, Fernandes AT, Hildebrandt P, Murgida DH, Todorovic S, Pereira MM, Melo EP, Martins LO. Copper incorporation into recombinant CotA laccase from Bacillus subtilis: characterization of fully copper loaded enzymes. J Biol Inorg Chem. 2008;13:183–93. https://doi.org/10.1007/s00775-007-0312-0.
Mohammadian M, Fathi-Roudsari M, Mollania N, Badoei-Dalfard A, Khajeh K. Enhanced expression of a recombinant bacterial laccase at low temperature and microaerobic conditions: purification and biochemical characterization. J Ind Microbiol Biotechnol. 2010;37:863–9. https://doi.org/10.1007/s10295-010-0734-5.
Siroosi M, Amoozegar MA, Khajeh K, Dabirmanesh B. Decolorization of dyes by a novel sodium azide-resistant spore laccase from a halotolerant bacterium, Bacillus safensis sp. strain S31. Water Sci Technol. 2018;77(12):2867–75. https://doi.org/10.2166/wst.2018.281.
Blánquez A, Rodríguez J, Brissos V, Mendes S, Martins LO, Ball AS, Arias ME, Hernández M. Decolorization and detoxification of textile dyes using a versatile Streptomyces laccase-natural mediator system. Saudi J Biol Sci. 2019;26:913–20. https://doi.org/10.1016/j.sjbs.2018.05.020.
Singh D, Rawat S, Waseem M, Gupta S, Lynn A, Nitin M, Ramchiary N, Sharma KK. Molecular modeling and simulation studies of recombinant laccase from Yersinia enterocolitica suggests significant role in the biotransformation of non-steroidal anti-inflammatory drugs. Biochem Biophys Res Commun. 2016;469:306–12. https://doi.org/10.1016/j.bbrc.2015.11.096.
Margot J, Bennati-Granier C, Maillard J, Blánquez P, Barry DA, Holliger C. Bacterial versus fungal laccase: potential for micropollutant degradation. AMB Express. 2013;3(1):63. https://doi.org/10.1186/2191-0855-3-63.
Reda FM, El-Mekkawy RM, Hassan NS. Detoxification and bioremediation of sulfa drugs and synthetic dyes by Streptomyces mutabilis A17 laccase produced in solid state fermentation. J Pure Appl Microbiol. 2019;13(1):85–96. https://doi.org/10.22207/JPAM.13.1.09.
Blánquez A, Guillén F, Rodríguez J, Arias ME, Hernández M. The degradation of two fluoroquinolone based antimicrobials by SilA, an alkaline laccase from Streptomyces ipomoeae. World J Microbiol Biotechnol. 2016;32(3):52. https://doi.org/10.1007/s11274-016-2032-5.
Mot A, Silaghi-Dumitrescu R. Laccases: complex architectures for one-electron oxidations. Biochemistry. 2012;77:1395–407. https://doi.org/10.1134/S0006297912120085.
Claus H. Laccases: structure, reactions, distribution. Micron. 2004;35:93–6. https://doi.org/10.1016/j.micron.2003.10.029.
Komori H, Miyazaki K, Higuchi Y. X-ray structure of a two-domain type laccase: a missing link in the evolution of multi-copper proteins. FEBS Lett. 2009;583:1189–95. https://doi.org/10.1016/j.febslet.2009.03.008.
Nakamura K, Kawabata T, Yura K, Go N. Novel types of two-domain multi-copper oxidases: possible missing links in the evolution. FEBS Lett. 2003;553:239–44. https://doi.org/10.1016/S0014-5793(03)01000-7.
Nakamura K, Go N. Function and molecular evolution of multicopper blue proteins. Cell Mol Life Sci. 2005;62:2050–66. https://doi.org/10.1007/s00018-004-5076-x.
Vasin A, Klotchenko S, Puchkova L. Phylogenetic analysis of six-domain multi-copper blue proteins. PLoS Curr. 2013;5:1–18. https://doi.org/10.1371/currents.tol.574bcb0f133fe52835911abc4e296141.
Geng A, Wu J, Xie RR, Li X, Chang FX, Sun JZ. Characterization of a laccase from a wood-feeding termite, Coptotermes formosanus. Insect Sci. 2016;25:251–8. https://doi.org/10.1111/1744-7917.12415.
Sirim D, Wagner F, Wang L, Schmid RD, Pleiss J. The Laccase Engineering Database: a classification and analysis system for laccases and related multicopper oxidases. Database. 2011. https://doi.org/10.1093/database/bar006.
Liu Z, Xie T, Zhong Q, Wang G. Crystal structure of CotA laccase complexed with 2, 2-azinobis-(3-ethylbenzothiazoline-6-sulfonate) at a novel binding site. Acta Crystallogr Sect F Struct Biol Commun. 2016;72:328–35. https://doi.org/10.1107/S2053230X1600426X.
Messerschmidt A, Ladenstein R, Huber R, Bolognesi M, Avigliano L, Petruzzelli R, Rossi A, Finazzi-Agró A. Refined crystal structure of ascorbate oxidase at 1.9 Å resolution. J Mol Biol. 1992;224:179–205. https://doi.org/10.1016/0022-2836(92)90583-6.
Messerschmidt A, Luecke H, Huber R. X-ray structures and mechanistic implications of three functional derivatives of ascorbate oxidase from zucchini: reduced, peroxide and azide forms. J Mol Biol. 1993;230:997–1014. https://doi.org/10.1006/jmbi.1993.1215.
Kumar SS, Phale PS, Durani S, Wangikar PP. Combined sequence and structure analysis of the fungal laccase family. Biotechnol Bioeng. 2003;83:386–94. https://doi.org/10.1002/bit.10681.
Mehra R, Muschiol J, Meyer AS, Kepp KP. A structural-chemical explanation of fungal laccase activity. Sci Rep. 2018;8:17285. https://doi.org/10.1038/s41598-018-35633-8.
Glazunova O, Trushkin N, Moiseenko K, Filimonov I, Fedorova T. Catalytic efficiency of basidiomycete laccases: redox potential versus substrate-binding pocket structure. Catalysts. 2018;8:152. https://doi.org/10.3390/catal8040152.
Pardo I, Camarero S. Laccase engineering by rational and evolutionary design. Cell Mol Life Sci. 2015;72:897–910. https://doi.org/10.1007/s00018-014-1824-8.
Sitarz AK, Mikkelsen JD, Meyer AS. Structure, functionality and tuning up of laccases for lignocellulose and other industrial applications. Crit Rev Biotechnol. 2016;36:70–86. https://doi.org/10.3109/07388551.2014.949617.
Ernst HA, Jørgensen LJ, Bukh C, Piontek K, Plattner DA, Østergaard LH, Larsen S, Bjerrum MJ. A comparative structural analysis of the surface properties of asco-laccases. PLoS ONE. 2018;13:e0206589. https://doi.org/10.1371/journal.pone.0206589.
Jones SM, Solomon EI. Electron transfer and reaction mechanism of laccases. Cell Mol Life Sci. 2015;72:869–83. https://doi.org/10.1007/s00018-014-1826-6.
Azimi M, Nafissi-Varcheh N, Mogharabi M, Faramarzi MA, Aboofazeli R. Study of laccase activity and stability in the presence of ionic and non-ionic surfactants and the bioconversion of indole in laccase-TX-100 system. J Mol Catal B Enzym. 2016;126:69–75. https://doi.org/10.1016/j.molcatb.2016.02.001.
Kang C, Ren D, Zhang S, Zhang X, He X, Deng Z, Huang C, Guo H. Effect of polyhydroxyl compounds on the thermal stability and structure of laccase. Pol J Environ Stud. 2019;28:3253–9. https://doi.org/10.15244/pjoes/94839.
Díaz R, Díaz-Godínez G, Anducho-Reyes MA, Mercado-Flores Y, Herrera-Zúñiga LD. In silico design of laccase thermostable mutants from Lacc 6 of Pleurotus ostreatus. Front Microbiol. 2018;9:1–12. https://doi.org/10.3389/fmicb.2018.02743.
Drozd R, Rakoczy R, Wasak A, Junka A, Fijałkowski K. The application of magnetically modified bacterial cellulose for immobilization of laccase. Int J Biol Macromol. 2018;108:462–70. https://doi.org/10.1016/j.ijbiomac.2017.12.031.
Mateljak I, Rice A, Yang K, Tron T, Alcalde M. The generation of thermostable fungal laccase chimeras by SCHEMA-RASPP structure-guided recombination in vivo. ACS Synth Biol. 2019;8:833–43. https://doi.org/10.1021/acssynbio.8b00509.
Herrera-Zúñiga LD, Millán-Pacheco C, Viniegra-González G, Villegas E, Arregui L, Rojo-Domínguez A. Molecular dynamics on laccase from Trametes versicolor to examine thermal stability induced by salt bridges. Chem Phys. 2019;517:253–64. https://doi.org/10.1016/j.chemphys.2018.10.019.
Yang J, Yang X, Lin Y, Ng TB, Lin J, Ye X. Laccase-catalyzed decolorization of malachite green: performance optimization and degradation mechanism. PLoS ONE. 2015;10:e0127714. https://doi.org/10.1371/journal.pone.0127714.
Lassouane F, Aït-Amar H, Amrani S, Rodriguez-Couto S. A promising laccase immobilization approach for bisphenol A removal from aqueous solutions. Bioresour Technol. 2019;271:360–7. https://doi.org/10.1016/j.biortech.2018.09.129.
Llevot A, Grau E, Carlotti S, Grelier S, Cramail H. Selective laccase-catalyzed dimerization of phenolic compounds derived from lignin: towards original symmetrical bio-based (bis) aromatic monomers. J Mol Catal B Enzym. 2016;125:34–41. https://doi.org/10.1016/j.molcatb.2015.12.006.
Polyakov K, Gavryushov S, Ivanova S, Fedorova T, Glazunova O, Popov A, Koroleva O. Structural study of the X-ray-induced enzymatic reduction of molecular oxygen to water by Steccherinum murashkinskyi laccase: insights into the reaction mechanism. Acta Crystallogr D Struct Biol. 2017;73(Pt. 5):388–401. https://doi.org/10.1107/s2059798317003667.
Gabdulkhakov A, Kostareva O, Kolyadenko I, Mikhaylina A, Trubitsina L, Tishchenko S. Incorporation of copper ions into T2/T3 centers of two-domain laccases. J Mol Biol. 2018;52:23–9. https://doi.org/10.1134/S0026893318010041.
Hoegger PJ, Kilaru S, James TY, Thacker JR, Kües U. Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J. 2006;273:2308–26. https://doi.org/10.1111/j.1742-4658.2006.05247.x.
Awasthi M, Jaiswal N, Singh S, Pandey VP, Dwivedi UN. Molecular docking and dynamics simulation analyses unraveling the differential enzymatic catalysis by plant and fungal laccases with respect to lignin biosynthesis and degradation. J Biomol Struct Dyn. 2015;33:1835–49. https://doi.org/10.1080/07391102.2014.975282.
Kjaergaard CH, Jones SM, Gounel S, Mano N, Solomon EI. Two-electron reduction versus one-electron oxidation of the type 3 pair in the multicopper oxidases. J Am Chem Soc. 2015;137:8783–94. https://doi.org/10.1021/jacs.5b04136.
Patel H, Gupte S, Gahlout M, Gupte A. Purification and characterization of an extracellular laccase from solid-state culture of Pleurotus ostreatus HP-1. 3 Biotech. 2014;4:77–84. https://doi.org/10.1007/s13205-013-0129-1.
Pardo I, Santiago G, Gentili P, Lucas F, Monza E, Medrano FJ, Galli C, Martínez AT, Guallar V, Camarero S. Re-designing the substrate binding pocket of laccase for enhanced oxidation of sinapic acid. Catal Sci Technol. 2016;6:3900–10. https://doi.org/10.1039/C5CY01725D.
Lucas MF, Monza E, Jørgensen LJ, Ernst HA, Piontek K, Bjerrum MJ, Martinez AT, Camarero S, Guallar V. Simulating substrate recognition and oxidation in laccases: from description to design. J Chem Theory Comput. 2017;13:1462–7. https://doi.org/10.1021/acs.jctc.6b01158.
Wu MH, Lee CC, Hsiao AS, Yu SM, Wang AHJ, Ho THD. Kinetic analysis and structural studies of a high-efficiency laccase from Cerrena sp. RSD 1. FEBS Open Bio. 2018;8:1230–46. https://doi.org/10.1002/2211-5463.12459.
Bertrand T, Jolivalt C, Briozzo P, Caminade E, Joly N, Madzak C, Mougin C. Crystal structure of a four-copper laccase complexed with an arylamine: insights into substrate recognition and correlation with kinetics. Biochemistry. 2002;41:7325–33. https://doi.org/10.1021/bi0201318.
Pardo I, Camarero S. Exploring the oxidation of lignin-derived phenols by a library of laccase mutants. Molecules. 2015;20:15929–43. https://doi.org/10.3390/molecules200915929.
Kameshwar AKS, Barber R, Qin W. Comparative modeling and molecular docking analysis of white, brown and soft rot fungal laccases using lignin model compounds for understanding the structural and functional properties of laccases. J Mol Graph Model. 2018;79:15–26. https://doi.org/10.1016/j.jmgm.2017.10.019.
Damas JM, Batptista AM, Soares CM. The pathway for O2 diffusion inside CotA laccase and possible implications on the multicopper oxidases family. J Chem Theory Comput. 2014;10:3525–31. https://doi.org/10.1021/ct500196e.
Solomon EI, Augustine AJ, Yoon J. O2 reduction to H2O by the multicopper oxidases. Dalton Trans. 2008;30:3921–32. https://doi.org/10.1039/b800799c.
Petersen LC, Degn H. Steady-state kinetics of laccase from Rhus vernicifera. Biochim Biophys Acta. 1978;526:85–92. https://doi.org/10.1016/0005-2744(78)90292-9.
Stines-Chaumeil C, Roussarie E, Mano N. The nature of the rate-limiting step of blue multicopper oxidases: homogeneous studies versus heterogeneous. Biochim Open. 2017;4:36–40. https://doi.org/10.1016/j.biopen.2017.01.001.
Li K, Xu F, Eriksson KEL. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl Environ Microbiol. 1999;65:2654–60.
Kulys J, Krikstopaitis K, Ziemys A, Schneider P. Laccase-catalyzed oxidation of syringates in presence of albumins. J Mol Cat B Enzym. 2002;18:99–108. https://doi.org/10.1016/S1381-1177(02)00073-5.
Kurniawati S, Nicell JA. A comprehensive kinetic model of laccase-catalyzed oxidation of aqueous phenol. Biotechnol Prog. 2009;25:763–73. https://doi.org/10.1002/btpr.111.
Kim SJ, Joo JC, Song BK, Yoo YJ, Kim YH. Engineering a horseradish peroxidase C stable to radical attacks by mutating multiple radical coupling sites. Biotechnol Bioeng. 2015;112:668–76. https://doi.org/10.1002/bit.25483.
Dordick JS. Designing enzymes for use in organic solvents. Biotechnol Prog. 1992;8:259–67. https://doi.org/10.1021/bp00016a001.
Patel N, Shahane S, Shivam, Majumdar R, Mishra U. Mode of action, properties, production, and application of laccase: a review. Recent Pat Biotechnol. 2019;13:19–32. https://doi.org/10.2174/1872208312666180821161015.
Fang Z, Li T, Wang Q, Zhang X, Peng H, Fang W, Hong Y, Ge H, Xiao Y. A bacterial laccase from marine microbial metagenome exhibiting chloride tolerance and dye decolorization ability. Appl Microbiol Biotechnol. 2011;89:1103–10. https://doi.org/10.1007/s00253-010-2934-3.
Giacobelli VG, Monza E, Lucas MF, Pezzella C, Piscitelli A, Guallar V, Sannia G. Repurposing designed mutants: a valuable strategy for computer-aided laccase engineering—the case of POXA1b. Catal Sci Technol. 2017;7:515–23. https://doi.org/10.1039/c6cy032410f.
Santiago G, De Salas F, Lucas MF, Monza E, Acebes S, Martinez AT, Camarero S, Guallar V. Computer-aided laccase engineering: toward biological oxidation of arylamines. ACS Catal. 2016;6:5415–23. https://doi.org/10.1021/acscatal.6b01460.
Camarero S, Pardo I, Cañas AI, Molina P, Record E, Martínez AT, Martínez MJ, Alcalde M. Engineering platforms for directed evolution of Laccase from Pycnoporus cinnabarinus. Appl Environ Microbiol. 2012;78:1370–84. https://doi.org/10.1128/AEM.07530-11.
Torres-Salas P, Mate D, Ghazi I, Alcalde M. Widening the pH activity profile of a fungal laccase bye directed evolution. ChemBioChem. 2013;14(8):934–7. https://doi.org/10.1002/cbic.201300102.
Xu F. Effects of redox potential and hydroxide inhibition on the pH activity profile of fungal laccases. J Biol Chem. 1997;272:924–8. https://doi.org/10.1074/jbc.272.2.924.
Madzak C, Mimmi M, Caminade E, Brault A, Baumberger S, Briozzo P, Mougin C, Jolivalt C. Shifting the optimal pH of activity for a laccase from the fungus Trametes versicolor by structure-based mutagenesis. Protein Eng Des Sel. 2006;19:77–84. https://doi.org/10.1093/protein/gzj004.
Zumárraga M, Bulter T, Shleev S, Polaina J, Martínez-Arias A, Plou FJ, Ballesteros A, Alcalde M. In vitro evolution of a fungal laccase in high concentrations of organic cosolvents. Chem Biol. 2007;14:1052–64. https://doi.org/10.1016/j.chembiol.2007.08.010.
Avelar M, Pastor N, Ramirez-Ramirez J, Ayala M. Replacement of oxidizable residues predicted by QM-MM simulation of a fungal laccase generates variants with higher operational stability. J Inorg Biochem. 2018;178:125–33. https://doi.org/10.1016/j.jinorgbio.2017.10.007.
Won DW. Structure and action mechanism of ligninolytic enzymes. Appl Biochem Biotechnol. 2009;157:174–209. https://doi.org/10.1007/s12010-008-8279-z.
Rodríguez-Couto S. Fungal laccase: a versatile enzyme for biotechnological applications. In: Yadav A, Mishra S, Singh S, Gupta A, editors. Recent advancement in white biotechnology through fungi. Fungal biology. Cham: Springer; 2019. p. 429–57.
Auriol M, Filali-Meknassi Y, Adams CD, Tyagi RD, Noguerol TN, Pina B. Removal of estrogenic activity of natural and synthetic hormones from a municipal wastewater: efficiency of horseradish peroxidase and laccase from Trametes versicolor. Chemosphere. 2008;70(3):445–52.
Steevensz A, Al-Ansari MM, Taylor KE, Bewtra JK, Biswas N. Comparison of soybean peroxidase with laccase in the removal of phenol from synthetic and refinery wastewater samples. J Chem Technol Biotechnol. 2009;84(5):761–9. https://doi.org/10.1002/jctb.2109.
Nair RR, Demarche P, Agathos SN. Formulation and characterization of an immobilized laccase biocatalyst and its application to eliminate organic micropollutants in wastewater. New Biotechnol. 2013;30(6):814–23. https://doi.org/10.1016/j.nbt.2012.12.004.
Zimmermann YS, Shahgaldian P, Corvini PFX, Hommes G. Sorption-assisted surface conjugation: a way to stabilize laccase enzyme. Appl Microbiol Biotechnol. 2011;92:169–78. https://doi.org/10.1007/s00253-011-3534-6.
Ba S, Haroune L, Cruz-Morató C, Jacquet C, Touahar IE, Bellenger J-P, Legault CY, Jones JP, Cabana H. Synthesis and characterization of combined cross-linked laccase and tyrosinase aggregates transforming acetaminophen as a model phenolic compound in wastewaters. Sci Total Environ. 2014;487:748–55.
Arca-Ramos A, Eibes G, Feijoo G, Lema JM, Moreira MT. Potentiality of a ceramic membrane reactor for the laccase-catalyzed removal of bisphenol A from secondary effluents. Appl Microbiol Biotechnol. 2015;99(21):9299–308. https://doi.org/10.1007/s00253-015-6826-4.
Jahangiri E, Thomas I, Schulze A, Seiwert B, Cabana H, Schlosser D. Characterisation of electron beam irradiation-immobilised laccase for application in wastewater treatment. Sci Total Environ. 2018;624:309–22. https://doi.org/10.1016/j.scitotenv.2017.12.127.
Le TT, Murugesan K, Lee CS, Vu CH, Chang YS, Jeon JR. Degradation of synthetic pollutants in real wastewater using laccase encapsulated in core–shell magnetic copper alginate beads. Bioresour Technol. 2016;216:203–10. https://doi.org/10.1016/j.biortech.2016.05.077.
Saparrat MC, Jurado M, Díaz R, Romera IG, Martínez MJ. Transformation of the water soluble fraction from “alpeorujo” by Coriolopsis rigida: the role of laccase in the process and its impact on Azospirillum brasiliense survival. Chemosphere. 2010;78(1):72–6. https://doi.org/10.1016/j.chemosphere.2009.09.050.
Acknowledgements
We would like to thank “Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo” in the Latin American development network “Lacasas Inmovilizadas para la Degradación de Compuestos Aromáticos en Aguas Residuales” (LIDA, project 318RT0552), as well as the Institutional Program of the “Instituto de Investigaciones Biomédicas-UNAM”: “La producción de biomoléculas de interés biomédico en bacterias y hongos.” We would like to thank Editage (www.editage.com) for English language editing. XGG, MHS and DRM thank the scholarships from CONACYT-México. XGG, MHS and DRM are students from Programa de Maestría en Ciencias Bioquímicas, Universidad Nacional Autónoma de México (UNAM).
Funding
This review was developed under the collaboration sponsored by the Institutional Program of the “Instituto de Investigaciones Biomédicas-UNAM”: “La producción de biomoléculas de interés biomédico en bacterias y hongos.” This work was partially financed by “Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica, Universidad Nacional Autónoma de México” (PAPIIT-UNAM IT-200719, IN-210419).
Author information
Authors and Affiliations
Contributions
LA, MA, XGG, GGS, CEHL, MHS, LL, ARD, DRM, MCNS, MATR and NAVC contributed to analyzing published data, discussing the previously published results and preparing the manuscript. XGG, MHS, DRM, MATR and NAVC prepared Figs. 1, 2, 3, 4. Figure 5 was made by LA and ARD. Figure 6 was made by GGS, CEHL and DRM. The data in the tables were compiled by MA, XGG, MHS, LL, ARD, DRM, MCNS, MATR and NAVC. MATR and NAVC prepared the final version of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Arregui, L., Ayala, M., Gómez-Gil, X. et al. Laccases: structure, function, and potential application in water bioremediation. Microb Cell Fact 18, 200 (2019). https://doi.org/10.1186/s12934-019-1248-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-019-1248-0