
Abstract
Potassium is an essential macronutrient, where its availability regulates numerous biochemical, phenological, and physiological responses in plants. Synchronizing potassium supply with plant demand is a key factor to enhance growth and grain production of wheat grown in cadmium-contaminated saline soils. Field experiments were conducted in El Fayoum province, Egypt, between latitudes 29° 02′ and 29° 35′ N and longitudes 30° 23′ and 31° 05′ E, during the cropping seasons of 2017–2018 and 2018–2019 to determine the influence of different applied potassium rates and times on nutrient uptake and wheat yield grown under Cd-contaminated saline soil (ECe = 8.53 dS m−1 and Cd = 18 mg kg−1 soil). Four K levels (K0, K40, K80, and K120 representing 0, 40, 80, and 120 kg ha−1) were applied at different application times [full dose (basal) at sowing (100% S), two equal split doses at sowing and flowering stage (50% S + 50% F), and full dose at flowering stage (100% F)]. The experimental treatments were arranged in a randomized split complete block design and replicated three times. The applied K rates, times, and their interaction induced significant differences in nutrient uptake and physiological responses which in turn improved the growth and yield of the wheat crop. Potassium addition with 120 kg ha−1 at two equal split doses (50% S + 50% F) resulted in the highest values of plant height (97 cm), Fv/Fm (0.83), PI (5.49), SPAD (58.63), MSI (34.57), seed yield (5.04 t ha−1), straw yield (9.04 t ha−1), and water productivity (0.99 kg m−3). Similarly, the uptake of N, P, K, Ca, Mg, Fe, Mn, and Zn was increased, while the uptake of Na and Cd decreased as the K supply increased under the split application. The addition of potassium by 120 kg ha−1 in two equal split doses at the sowing and flowering stage could be a valuable approach to improve yield and yield quality of wheat crop grown under cadmium-contaminated saline soils.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Worldwide, cereal crops are the main staple food, which directly supplies more than 50% of total human calorie input (FAO 2019). Among them, wheat (Triticum aestivum L.) holds progressive authority as one of the main sources of calories and dietary protein for the permanently burgeoning population of the world (FAO 2019). Potassium is an essential macronutrient, where its availability regulates numerous biochemical, phenological, and physiological responses in plants (Hasanuzzaman et al. 2018; Johnson et al. 2022) such as water uptake (Sardans and Peñuelas 2015); nutrient translocation (Xu et al. 2020), enzyme activation (Hasanuzzaman et al. 2020), photosynthesis (Siddiqui et al. 2021), protein synthesis (Sahi et al. 2021), osmoregulation (Wu and Wei-Hua 2013), energy transfer (Sardans and Peñuelas 2021), stomatal opening mechanism (Anokye et al. 2021), and stress resistance, i.e., salinity (Adhikari et al. 2020), drought (Abd El–Mageed et al. 2017), heat (Singh and Singh 2020), and heavy metals (Yasin et al. 2018). Potassium is a key factor involved in several metabolic processes, affecting yield and yield quality. Compared with nitrogen and phosphorus uptake, potassium is rapidly absorbed by plant roots (Ravichandran and Sriramachandrasekharan 2011). Like nitrogen and phosphorus, potassium has a greater significant effect on yield. Many research studies strongly confirmed the evidence about the direct and/or indirect role of K respecting to increased yield of wheat crop (Szczepanek and Grzybowski 2016; Wang et al. 2020). Sedri et al. (2022) found that the usage of potassium increased growth, yield components, nutrient uptake, straw yield, and grain yield of wheat crop. Instead, the insufficient K levels lead to a decrease in yield though other nutrients are available. Thornburg et al. (2020) concluded that K+ deficiency caused a significant reduction in tillers number, grain filling, and yield development in wheat. Plants that experience potassium deficiency, in addition to their low yield production, became more vulnerable to salinity, drought, and disease stresses (Thornburg et al. 2020).
In Egypt, soils are deficient in K content; Abdelfattah et al. (2021) indicates deterioration in soil fertility especially after building of the Aswan High Dam and stopping the sedimentation. Furthermore, maintaining an intensified cropping system with high yielding genotypes following an imbalanced fertilizer application scheme could have resulted in further depletion of soil K reserves (Raj et al. 2021). K deficiency induces negative effects on the growth and productivity of wheat crop; this reduction may be magnified under adverse conditions such as salinity and soil contamination with heavy metals like cadmium. Salinity is a major abiotic stress, diminishing the development and yield of several crops worldwide (Abd El-Mageed et al. 2022; Semida et al. 2014; Shaaban et al. 2022). Increasing osmotic potential due to salt stress restricts the availability and uptake of water by plants (Rady et al. 2020). It also inhibits the nutrient availability for plants induced by a weak structure of saline soils (Abd El-Mageed et al. 2021).
Additionally, recent research indicated remarkable retardation in plant growth and physiological processes induced by salinity stress owing to osmotic and/or ionic imbalance, (Abd El-Mageed et al. 2018; Semida et al. 2021; Abdou et al. 2022). Furthermore, under conditions of saline soil, the excess of Na+ concentration in soil solution leads plants to suffer from K-deficiency, initiated due to the antagonist effect, hence decreases in K-availability.
Cadmium (Cd) is one of the main deleterious environmental pollutants for living organisms. The existence of Cd in soils may generate due to the usage of phosphate fertilizers, and use of agricultural drainage water, and sewage sludge (Abd El-Mageed et al. 2020; Murtaza et al. 2015; Nagajyoti et al. 2010). However, results reported by Rizwan et al. (2016) showed that cadmium reduced germination, growth, photosynthesis activates, and grain yield of wheat crop. Also Abd El-Mageed et al. (2019) indicated photo-inhibition and reduction of mineral assimilation in sugar beet plants caused by cadmium toxicity.
Potassium has a vital protection function against abiotic stresses. These mechanisms reflect its responsibility in ion homeostasis as well as the integrity of the cellular membrane (Farooq et al. 2018; Johnson et al. 2022). Many reports have suggested that under salinity stress, sustaining a sufficient K supply alleviates the detrimental effects on saline-stressed strawberry (Kaya et al. 2003); wheat, soybean, cotton, and maize (Pettigrew 2008); and pepper (Rubio et al. 2011). Consequently, enhancing the K status of grown plants could be proposed as a helpful tool to decrease the oxidative damage of salt stress. Therefore, to compensate for the lack of potassium in soil and to reduce the injurious effects induced by salinity stress and high Cd levels on the growth and yield of wheat, appropriate K management strategies in wheat cropping systems are required. Rate and time of K application are key factors affecting the use efficiency of applied K. These factors would be considered in formulating a high-efficient K management scheme. In most annual crops, K supply as a basal addition with its whole dose is a dominant K fertilization technique, though it might contribute to diminish K—use efficiency. Crop demand of K varies distinctly according to growth stage and productivity level. Split K application may enhance the use efficiency of K by reducing K losses via fixation and/or leaching. However, the available research studies performed to evaluate the split K application method on the growth and yield of wheat crop grown in saline soil are still insufficient. In addition, the role of potassium in reducing the uptake of Cd by plants is still unclear and needs further studies. Thus, owing to the significant evidence of the negative effects of salinity and cadmium stress on plant growth, the current experiment was conducted to investigate the effects of potassium fertilization rate and application time on morpho-physiological responses, plant water status, and yield of wheat grown under Cd-contaminated saline soil. We hypothesized that K different rates and timing can overcome the injurious effects of Cd stress under saline soil conditions. The results could prove useful for developing sustainable management strategies for crop production under Cd-contaminated saline soil.
2 Materials and Methods
2.1 Experimental Location
Field trials were done from October to April months (2017–2018 and 2018–2019 located in El-Fayoum Governorate, Egypt, at latitudes 29°02 and 29°35_N and longitudes 30°23 and 31°05_E. The altitude of experimental site is − 17 m. The given data in Tables 1 and 2 represent that the soil used in the experimental site is sandy loam in texture (77.53% sand, 10.10% silt, and 12.37% clay), with a bulk density of 1.55 g cm−3 and retains available water by 12.13%. Soil salinity (ECe) was 8.53 dS m−1 and CaCO3% = 4.86, ESP = 15.62, and pH = 7.69. The content of organic matter in the soil was 1.12%, and the total N was 0.03%. Cadmium content (18 mg kg−1) indicates Cd-contaminated soil (Chen 2003). All physiochemical analysis of the studied soil was carried out according to Klute and Dirksen (1986) and Page et al. (1982).
2.2 Treatments and Experimental Design
The experimental treatments were arranged in a split-plot design and replicated three times. Potassium fertilizer (i.e., 0, 40, 80, and 120 kg ha−1) rates were fallen in the main plots, while time application (i.e., 100% at sowing (100% S), 50% at sowing and 50% at flowering stage (50% S + 50% F), and 100% at flowering stage (100% F)) were allocated in the subplots, with a net plot area of subplots of 20 m2.
2.3 Plant Management Practices
Healthy wheat grains (Triticum aestivum L. Cv. Giza 168) prepared by the Crop Research Institute, Agricultural Research Center, Egypt, were sown on 20 and 22 November of the 2017–2018 and 2018–2019 seasons (by the conventional method, the row-to-row distance was 20 cm) and harvested on 20 and 25 April of the 2017–2018 and 2018–2019 seasons, respectively. The area of each main plot was 50 m × 4 m. However, thirty-six exponential plots were used, and the area of the experimental plot was 20 m2 (5 m × 4 m). Based on the local agronomical practices recommendations particularizing for bread wheat cultivars in Egypt, the investigated soil was supplemented by 62 kg P2O5 ha−1 (i.e., 400 kg calcium monophosphate; 15.5% P2O5) during land preparation. In addition, 200 kg N ha−1 (i.e., 600 kg ammonium-nitrate; 33.5% N) was implemented by broadcasting in 3 equal doses (1/3 at sowing, however, the second and third doses were performed with the 1st and 2nd irrigation events, respectively). In this study, wheat plants were irrigated every 15 days utilizing the surface watering method.
2.4 Physiological Measurements
Sixty days after sowing, six fully expanded leaves of 10 plants plot−1 were selected randomly from each treatment to determine leaf relative chlorophyll content and chlorophyll fluorescence. Leaf relative chlorophyll content was estimated by a chlorophyll meter (SPAD-502 Minolta, Japan) on two fully expanded leaves. Chlorophyll fluorescence was determined on two different sunny days using a portable Handy PEA fluorometer (Hansatech Instruments Ltd, Kings Lynn, UK). The maximum quantum yield of PS II (Fv/Fm) was described by Maxwell and Johnson (2000). Fv/F0 reflects the efficiency of electron donation to the PSII RCs and the rate of photosynthetic quantum conversion at PSII RCs (Spoustová et al. 2013). PI reflects the performance index and was calculated according to Clark et al. (2000).
Membrane stability index (MSI) % was measured using the method of Premachandra et al. (1990) and then calculated using the following equation.
where MSI%: membrane stability index, C1: EC of the solution at 40 °C, and C2: EC of the solution at 100 °C.
2.5 Macro and Micronutrient Determination
Macronutrient concentrations in wheat leaves (i.e., nitrogen; N, phosphorus; P, and potassium; K+, magnesium; Mg++, calcium; Ca++, and sodium; Na+), as well as micronutrient (i.e., iron; Fe++, manganese; Mn, zinc; Zn, and copper; Cu) and cadmium (Cd2+), were assessed. Plant tissue samples for the analysis of the nutrient status of wheat plants were collected at the beginning of grain development according to the Biologische Bundesanstalt, Bundessortenamt, and CHemical industry (BBCH) scale (Meier 2001) (BBCH stage 71). A leaf area equivalent of 0.5 m2 from each plot was randomly selected one time at BBCH 71 stage (after about 20 days from the last potassium application). All of these plant materials were dried at 70 °C until constant weight and ground to pass a 1-mm mesh. Representative subsamples (200 mg) were taken and digested according to the standard method for plant nutrient determination. N content using a micro-Kjeldahl apparatus (Ningbo Medical Instruments Co., Ningbo, China) was determined according to the method of AOAC (2000). P content was assessed as described by Jackson (1973) using standard reagents of H2MoO7S, molybdenum blue, diluted H2MoO7S, and 8% (w/v) NaHSO3–H2SO4. The Ca2+, Mg2+, Cd2+, and micronutrient contents in leaves were evaluated by atomic absorption spectrophotometer (Perkin-Elmer, Model 3300) following the methods introduced by Chapman and Pratt. (1961). The ratio of K:Na was computed using the following equation:
Also the Ca:Na ratio was computed as follows:
2.6 Measurements of Growth Traits, Yield, and WP
To estimate growth traits of wheat plants, 10–12-week-old plants (10 plants plot−1) were randomly chosen from each experimental area to measure plant height (cm) with a meter scale, tillers number plant−1, and productive tillers plant−1. Also, the shoot fresh weight (FW) of each plant was weighed and then dried at 70 °C by an electric oven and the dry weight (DW) was recorded. At harvest, 10 plants plot−1 were randomly selected to determine yield and its components. Number of spikes plant−1, number of grains spike−1, grain weight spike−1 (g), 100-grain weight (g), and number of spikes m−2 were estimated in 1 m2 from each plot. All plants of each plot were removed to estimate 100-grain weight (g), grain yield (t ha−1), straw yield (t ha−1), and biological yield. Harvest index (HI) was calculated on a dry mass basis by using the following formula:
HI = gran yield (t ha−1)/biological yield (t ha−1).
The WP values as kg grain yield per m3 of applied water were calculated according to Jensen (1983):
2.7 Statistical Analysis
The obtained data were statistically analyzed and the significant differences between means were compared at p ≤ 0.05 using Duncan’s multiple range test. The statistical analysis was carried out using COSTAT computer software (CoHort Software version 6.303, Berkeley, CA, USA).
3 Results
3.1 Growth Parameters
Table 3 displays the effect of different applied K rates, time, and their interaction on the growth attributes of wheat plants. Plant height (PLH), tillers No (TN), productive tillers No (PTN), shoot fresh weight (SFW), and shoot dry weight (SDW) were significantly affected by the applied rates, time of potassium application, and their interaction. Results indicated a significant increase in plant height as well as K application dose increased, compared with control. Meanwhile, the increase in K application dose from 40 to 80 kg ha−1 did not cause significant differences in TN, SFW, and SDW. The tillers No showed a non-significant response between K80 and K120. Compared with control (K0), wheat growth parameters such as PLH, TN, PTN, SFW, and SDW increased by 0.83, 44.50, 42.33, 61.21, and 64.64%, respectively, for K40, and by 8.86, 83.33, 80.83, 67.42, and 72.00%, respectively, for K80 and by 13.50, 72.33, 68.50, 85.48, and 102.78%, respectively, for K120. In addition, the obtained results exhibited remarkable evidence that the wheat vegetative traits differed significantly in correspondence to the timing of potassium addition. The best results were recorded when potassium fertilizer was applied as a split treatment (50% S + 50% F) with a significant increase in PLH (5.53 and 6.09%), TN (14.36 and 6.59%), PTN (14.36 and 12.32%), SFW (16.26 and 27.00%), and SDW (19.96 and 27.22%) compared with other application methods (100% S and 100% F), respectively. Though, enriching the soil with the full dose of K via sowing (100% S) and flowering (100% F) treatments was associated with a notable decrease in growth of wheat plants compared with the split application (50% S + 50% F) but still significantly higher than control (non-amended soil plots with K).
Interestingly the differences in wheat growth due to the applied K in various levels at different times were found to be significant. The greatest PLH (97 cm), SFW (36.73 g), and SDW (13.10 g) were produced with K120 × 50% S + 50% F; however, the highest values of TN (4) and PTN (4) were recorded for split K application at dose 80 kg ha−1. Meanwhile, the lowest values of PLH, TN, PTN, SFW, and SDW were 78.67 (cm), 2.67, 2.54, 23.99 (g), and 8.38 (g) which were recorded in control plants.
3.2 Physiology
The effects induced by different applied K levels, time of application, and their interaction on wheat physiological responses are given in Table 4. Chlorophyll fluorescence parameters (Fv/Fm, Fv/Fo, PI, and SPAD) and membrane stability index (MSI) of wheat plants were influenced positively as the K application rate increased. Increasing the K application dose to 40, 80, and 120 kg ha−1 resulted significant increase in Fv/Fm (25.13, 26.15, and 26.67%), Fv/Fo (51.01, 95.98, and 103.74%), PI (47.28, 105.44, and 198.35%), SPAD (41.94, 51.36, and 93.15%), and MSI (41.81, 61.47, and 76.25%), respectively, relative to control (K0). A significant increase was observed in Fv/Fm, Fv/Fo, PI, SPAD, and MSI due to applied potassium (K120) as compared to control and K40. However, differences were insignificant for Fv/Fm and Fv/Fo between K120 and K80. Noteworthy, split application (50% S + 50% F) resulted in better physiological responses of wheat plants than whole application at basal (100% S) and flowering stage (100% F) treatments. Moreover, statistical results showed non-significant differences in Fv/Fo and PI between 50% S + 50% F and 100% F treatments. Similarly, Fv/Fm and SPAD also recorded non-significant differences between 100% S and 100% F treatments. Split K application (50% S + 50% F) induced increases in Fv/Fm (0.81 and 2.06%), Fv/Fo (0.39 and 10.29%), PI (28.49 and 36.15%), SPAD (20.34 and 24.19%), and MSI (8.98 and 25.08%) compared with 100% S and 100% F, respectively. The greatest mean values as an average of both seasons of Fv/Fm, Fv/Fo, PI, SPAD, and MSI by 0.83, 4.75, 5.49, 58.63, and 34.57 were observed for K120 × 50% S + 50% F treatment. Hence, split application of K with 120 kg ha−1 was favorable to achieve the best physiological responses of wheat plants.
3.3 Macronutrient Uptake
Data presented in Figs. 1 and 2 showed significant effects induced by applied K rates, the timing of K application, and their interactions on the nutrient status of wheat plants. The content of nitrogen (N) (Fig. 1A), phosphorus (P) (Fig. 1B), potassium (K) (Fig. 1C), calcium (Ca) (Fig. 1D), and Mg (Fig. 2A) increased and differed significantly as the K application level increased compared with the control (K0). Respecting to K time application effect, the highest levels from N (27.71 mg g DW−1), P (6.15 mg g DW−1), K (32.23 mg g DW−1), Ca (6.59 mg g DW−1), and Mg (6.79 mg g DW−1) were noted for split K addition (50% S + 50% F) with K application rate of 120 kg ha−1, and the minimum recorded for control (K0). Interestingly, the increase of K application rate from K80 to K120 did not cause significant differences in plant nutrient status. Estimated K:Na ratio and Ca:Na ratio showed a significant increase over control as the K application dose increased. The preferable records for K:Na (2.41) and Ca:Na (0.62) were observed for plants amended with K120 application rate. The differences in K:Na and Ca:Na percentages were insignificant between K40 and K80 treatments. Potassium application time resulted in significant effects on K:Na ratio, while the applied K at sowing and split application did not cause significant differences in the Ca:Na ratio. Additionally, in response to K application rate × K application time, except Ca it was observed that a significant increase in N, P, K, and Ca contents due to applied split K with 120 kg ha−1 as compared with other treatments.

Interactive effect of potassium application rates interacted with potassium application time on N: nitrogen (A), P: phosphorus (B), K: potassium (C), and Ca: calcium (D) contents (mg g DW−1) of wheat plants grown under Cd-contaminated saline soil (data pooled over both seasons). K0, K40, K80, and K120 are potassium application rates by 0, 40, 80, and 120 kg ha−1, respectively. 100% S is full applied K dose at sowing, 50% S + 50% F is split applied K by two equal doses at sowing and flowering stage, and 100% F is full applied K dose at flowering stage. Different letters on the bars refer to significant differences among means based on Duncan least-significant difference test at the p < 0.05 level

Interactive effect of potassium application rates interacted with potassium application time on Mg: magnesium (A), Na: sodium (B) content (mg g DW−1), K:Na potassium:sodium (C), and Ca:Na calcium:sodium ratio (D) of wheat plants grown under Cd-contaminated saline soil (data pooled over both seasons). K0, K40, K80, and K120 are potassium application rates by 0, 40, 80, and 120 kg ha−1, respectively. 100% S is full applied K dose at sowing, 50% S + 50% F is split applied K by two equal doses at sowing and flowering stage, and 100% F is full applied K dose at flowering stage. Different letters on the bars refer to significant differences among means based on Duncan least-significant difference test at the p < 0.05 level
3.4 Micronutrients and Cadmium Content
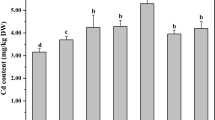
Micronutrient contents in response to K application time and rates Fe (Fig. 3A), Mn (Fig. 3B), Zn (Fig. 3C), and Cu (Fig. 3D) were increased with increasing K application level. The increase in Mn and Zn was non-significant between K40 and K80 treatments. Fe content of leaves showed non-significant differences between K80 and K120 treatments. Similarly, no significant differences were observed for Cu content in correspondence to K application levels. Split K application resulted in a significant increase for Fe, Mn, and Zn concentrations by 10.30, 34.23, and 24.02% compared with full K applied at flowering. The differences for Fe and Zn were found to be non-significant between whole K application treatment at sowing and split application (50% S + 50% F) application. Also, the effect of basal K treatment (100% S) was non-significant on Mn and Cu compared with the total applied K dose at flowering (100% F). Fe, Mn, Zn, and Cu recorded their maximum values of 1545, 149, 120, and 53 mg kg DW, respectively, under split K application in combination with a high K application rate (K120). Conversely, Na (Fig. 2B) and Cd (Fig. 3E) showed a significant decrease as the K application dose increased compared with the control (K0). The reduction in Na and Cd were 20.41, 24.94, and 48.75% and 87.74, 83.91, and 80.84%, respectively, due to applied K levels by K40, K80, and K120, respectively, as compared to K0. Results also exhibited non-significant effects for K application time on cadmium content in wheat plants.

Interactive effect of potassium application rates interacted with potassium application time on Fe: ferric (A), Mn: manganese (B), Zn: zinc (C), Cu: copper (D), and cadmium (E) content of wheat plants grown under Cd-contaminated saline soil (data pooled over both seasons). K0, K40, K80, and K120 are potassium application rates by 0, 40, 80, and 120 kg ha−1, respectively. 100% S is full applied K dose at sowing, 50% S + 50% F is split applied K by two equal doses at sowing and flowering stage, and 100% F is full applied K dose at flowering stage. Different letters on the bars refer to significant differences among means based on Duncan least-significant difference test at the p < 0.05 level
3.5 Yield and Yield Components
Yield and yield characters of wheat plants significantly differed due to the different applied rates and time of K fertilization (Table 5). The increases in wheat yield parameters (i.e., weight of 100 grains (W100G), seed yield (SY), straw yield (STY) as well as biological yield (BY), and harvest index (HI)) were parallel with the increments of applied K dose. The applied K levels by 40, 80, and 120 kg ha−1 resulted in a significant increase in W100G (8.57, 23.81, and 29.52%), SY (17.90, 29.57, and 42.73%), STY (10.74, 17.10, and 22.69%), BY (13.21, 21.33, and 15.63%), and HI (7.29, 10.42, and 15.63%) than that of control (K0). Interestingly, it is worth mentioning that, though the addition of K at its high rate (K120) was superior to achieve the greatest values of W100G, SY, STY, and BY compared with other K application rates, its effect on W100G, STY, and BY was non-significant compared with K80. Concerning the effect of K application time, W100G and HI showed non- significant responses due to the applied K application time. No significant differences were observed in SY, STY, and BY when whole K dose was applied at sowing or flowering stage. Split application of potassium achieved the highest yield traits of wheat crop over full K addition at sowing and flowering treatments. Results also indicated a favorable effect due to split applied K with application dose (K120) on grain yield and yield traits of the wheat crop compared with other treatments.
Interestingly also, the highest value (0.99 kg m−3) of water productivity (WP) was in line with the increase in seed yield of wheat crop in response to split applied potassium with its high rate. WP recorded increases by 18.18, 29.80, and 45.45% as K application dose increased up to 40, 80, and 120 kg ha−1, respectively, over control (K0). The effect of different applied rates and timing of K fertilization was significant on WP. Split addition of K resulted in a significant increase in WP by 6.59 and 11.79% over applied K at sowing (100% S) and flowering (100% F) treatments, respectively. However, the lowest value of WP (0.66 kg m−3) was recorded for non-amended plots with potassium (K0).
3.5.1 Discussion
Potassium (K) is an essential macronutrient regulating many physiological responses in grown plants like water uptake, nutrient translocation, synthesis of protein, an activator of dozens of important enzymes, photosynthesis, transport of starch, and sugar (Johnson et al. 2022; Sardans and Peñuelas 2015; Siddiqui et al. 2021). In Egypt, soils are deficient in K content (Abdelfattah et al. 2021). K deficiency also may increase as a result of following an imbalanced fertilizer scheme (Li et al. 2021) under intensified cropping system (Raj et al. 2021). Hence, to deal with the lack of potassium in soil appropriate K management strategies in wheat cropping systems are required. Potassium supply as a basal application has been widely discussed by many scientific reports (Abd El–Mageed et al. 2017; Khan et al. 2007), while the experimental work detailed with the influence of split applied K fertilizers on yield and yield quality of wheat crop are very little. In addition, the relation between K availability and cadmium uptake by wheat plants grown in Cd-contaminated soils is still unclear. Therefore, adopting new K management practices could be useful for improving soil fertility and wheat productivity and mitigating adverse effects induced by Cd under saline soil conditions.
Our results indicated positive responses in vegetative growth of wheat plants due to applied K at different rates and times. Plant height, tillers No, productive tillers No, shoot fresh weight, and shoot dry weight were influenced significantly and increased as well as K application dose increased. Achieved in soil sufficient K levels with K80 and K120 treatments may be enhanced nutritional element uptake and improve physiological responses of wheat plants which in turn positively affected wheat growth. Induced growth by K could be due to K regulating the role of the stomatal opening system, cell turgidity, and cell elongation (Bang et al. 2021; Sardans and Peñuelas 2021; Xu et al. 2020). K has a stimulating effect on ATPase production in the plasma membrane, resulting in better plant growth (Siddiqui et al. 2021; Tränkner et al. 2018). Increases in plant growth with increasing K supply were also reported by Hussain et al. (2011) on mung bean, Tang et al. (2015) and Zelelew et al. (2016) on potatoes, and Gerardeaux et al. (2010) on cotton. The timing of the K application also caused significant differences in wheat growth parameters. Split applied K treatment resulted in the highest growth compared with other application methods. Hence, K application in two equal split doses (50% S + 50% F) may be associated with greater K release for plant uptake, and consequently higher growth obtained. Singh et al. (2021) pointed out that the availability of K in soil solution increases with split K addition. Improvement in nutrient use efficiency with the split application may refer to the reduction in K losses via transformation into non-exchangeable form and/or leaching losses. A similar finding was reported by Islam et al. (2014) who reported that plant height, tillers m−2, and accumulation of dry matter were superior at two equal split doses of K at sowing and tiller stages than a single basal application. Hossain et al. (2015) found that the growth and yield of spring wheat gradually increased as well as K2O rates increased up to 72 kg ha−1 for both conventional basal and split applications.
Chlorophyll fluorescence aspects are widely used to estimate the activity of photosynthetic apparatus and the rate of carbon assimilation (Desoky et al. 2021; Rady et al. 2017; Semida 2016; Semida et al. 2019). In the present work, chlorophyll fluorescence traits such as Fv/Fm, Fv/F0, PI, and SPAD were determined in response to potassium application rates. Our results exhibited a significant increase in these traits with increasing K application levels. Leaf photosynthesis comprises of the several physiological activities, that is, CO2 assimilation, PSII photochemistry, and light harvesting (Etsushi et al. 2011). K deficiency inhibits photosynthetic functions, including PSII photochemistry, which immediately influence the photosynthetic apparatus (Thornburg et al. 2020). The function of K in photosynthetic activity is regulating the light energy of PSII (Tränkner et al. 2018), controlling ATP synthesis, and activation of enzymes that are involved in photosynthesis (Chen et al. 2016). K has been reported to play a key role in CO2 fixation, transportation, and consumption of photo-assimilates (Tränkner et al. 2018) and acting as an antagonist to light-caused due to H+ flux within the thylakoid membranes (Anschütz et al. 2014). Therefore, the highest values of wheat physiological aspects were noted for K application at 120 kg ha−1 and significantly decreased with increasing K deficit level. K deficiency inhibits the structure and function of the reaction center contributing to a reduction in chlorophyll fluorescence kinetics. Römheld and Kirkby (2010) stated that potassium deficiency disrupted transpiration rates and resulted in a decrease in chlorophyll fluorescence. Marschner (2011) noted that photosynthetic rate, electron transfer energy, and stomatal conductance were significantly decreased for K-deficiency stressed maize plants. Results also showed a better response in wheat chlorophyll fluorescence parameters due to applied K as a split application. Induced K availability by split addition could be contributed to higher K uptake and growth of wheat plants, resulting in better plant physiological responses as compared with other applied K techniques.
Potassium plays a fundamental role in the osmotic adjustment ability; an appropriate K level may promote osmotic adjustment, which retains higher turgor pressure, relative water content in plant cells. Therefore, values of membrane stability index (MSI%) showed a considerable increase under the high K application level. Johnson et al. (2022) stated that K regulates water influx to guard cells, hence maintaining turgor pressure and enhancing the stability of cellular membranes. Aksu and Altay (2020) reported that the leaf relative water content and membrane stability increased significantly of amended sugar beet with K under drought stress. K plays a critical function in turgor regulation within the guard cells during stomatal movement. Some studies also reported that potassium deficiency may cause stomatal closure and disrupt photosynthetic activities in several crop plants (Houmani et al. 2022), resulting negative effects on plant physiological attributes, leading to lower growth and yield of wheat plants.
K application increased the uptake of macro and micro nutritional elements. Nutrient uptake stimulated with K indicates improvement in water movement and solvent transport. This finding could be a result of the K-regulating role of trans-membrane potential and osmotic pressure in xylem tubes which is intensively influenced by the K status in roots (Abd El–Mageed et al. 2017). Schwartzkopf (2018) noted that K improved water flow and mineral translocation through the xylem, whereas the uptake of nitrates NO3−, PO4−3, Ca, Mg, and amino acid under non-optimal K supply is reduced. Murtaza et al. (2015) observed that nutrient uptake and growth of maize plants were significantly improved as the K application rate increased. Misskire et al. (2019) reported that the uptake of N, P, K, Ca, and Mg was increased in response to applied K rates. Ali et al. (2019) observed an increase in contents of N, P, and K due to increased K addition relative to control. The decrease in Na with K increased application levels as compared with control is due to the competitive effect between ions. Under saline soil conditions with low or without K supply, Na+ exhibits a competitive inhibition for K uptake, resulting in lower plant metabolism and disrupting plant growth. Meanwhile, when K supply is sufficient for instance under K supply levels like K80 and K120, it leads to higher K accumulation in plants, resulting in a higher K/Na ratio, which consequently leads to a further decrease in Na concentration. Chakraborty et al. (2016) noted that the application of K stimulated plant growth and reduced the uptake of Na.
Results also showed a significant reduction in leaf Cd concentration of wheat plants in response to applied K fertilization relative to control. Decreases in Cd uptake induced by K indicate a decrease in the activity of free Cd ions. The release of SO4 ions from KSO4 (applied form of K fertilizer) in soil solution may enhance the formation of CdSO4 complexes, resulting in lower free Cd activity, accordingly decreases in Cd uptake. Our results are similar to those reported by Sterckeman and Thomine (2020) who stated that the extraction of cadmium cation by roots greatly responded to the concentration of other elements in the soil solution. Cd diffusion and uptake were restricted by the presence of SO4 in the soil solution, due to created Cd-SO4= complexes. Sahi et al. (2021) noted reduction in the bio-Cd content in the soil and Cd accumulation in Panax notoginseng as a result of improved Bacteroidetes Proteobacteria, and Mortierellomycota activity stimulated by K application. Yang and Juang (2015) revealed that K fertilizer mitigated Cd toxicity in soybean roots through detoxification mechanisms. A similar trend of disrupting the oxidative stress induced by heavy metals involved Cd under adequate K supply has been introduced by some authors (Naciri et al. 2021; Yasin et al. 2018).
Yield and yield-contributing characteristics of wheat plants showed a significant increase with increasing K applied rates. Potassium addition significantly improved chlorophyll fluorescence responses, nutrient uptake, and plant water status, which are key factors for getting optimal growth and appropriate grain yield. In this study, the cumulative effect of yield-contributing traits, including tillers No (TN), productive tillers No (PTN), and 100-GW were reasonable for obtaining higher GY with K addition at 120 kg ha−1. Therefore, supplying a sufficient K level under K120 was superior to achieve the highest yield of wheat compared with other application rates. Grain yield produced under K120 (50% S + 50% F) treatment was higher due to enhanced K uptake from the soil, consequently increased the translocation of metabolites which is essential for grain filling and plant development. The plant potassium status is highly linked with the process of translocation of photosynthesis and sugars in the plant system that may be responsible for the higher grain yield. Additionally, potassium is a co-factor for several enzymes and its effect on starch synthesis is well established. In a K-sufficient plant, the synthesis proteins, starches, and cellulose were markedly increased, Hence, the availability of K might be induced a positive effect on grain development. As compared with other interactive combinations, the use of K application dose K120 with application technique (50% S + 50% F) may resulted in greater availability of K and lower transformation of potassium into non-exchangeable pool, which regulated the continuous growth of cells and tissues, enhanced N uptake and protein synthesis, improved many physiological growth processes, and delayed plant leaf senescence, hence increased the growth and yield of the crop. This finding was confirmed with previous studies conducted by Islam et al. (2014) and Sharma and Singh (2020). Oosterhuis et al. (2014) reported that the improvement of growth and yield induced by potassium is a result of a beneficial role K in modulating water and nutrient uptake, biosynthesis, and metabolite processes. Raza et al. (2014) noted an increase in spike length and grain yield of wheat under the deficit-water condition with K addition. Results also exhibited improvement in yield and yield aspects of wheat in response to split K supply. Potassium is an essential nutrient during flowering and pollen development stages as well as in seed germination. The split application method may be achieved a constant and homogeneous K supply, resulting in a better reproductive-vegetative ratio, which is a key factor for getting a higher yield. Also, as mentioned by Wani et al. (2014) the positive response in K split application might be contributed to higher buffering capacity of soil associated with lower potassium fixation. Wheat plants need a huge amount of K, and a constant supply is necessary until the reproductive stage is the maximum (Ravichandran and Sriramachandrasekharan 2011). Therefore, the decrease in yield and yield component with 100% S treatment could be referred to as the lower availability of K during the reproductive stages of wheat crop. K applied at sowing might be got exhausted from the soil through its further transformations to other K pools, resulting in lower K availability. In addition, the increase recorded in biological yield may be owing to the integrated effect and positive contribution of yield and better vegetative growth characters produced with the K supply in two equal splits. Split K application induced potassium availability during the growth period which achieved in better plant metabolic activities resulting in more grain and biological yields of wheat crop. Hence, an un-synchronized K supply adversely affected the yield of wheat crop. Zou Tie (2007) stated that adequate K supply contributed to a significant increase in photosynthesis during the grain filling stage of wheat, resulting increase in the grain number and grain yield.
4 Conclusion
The results of the 2-year experiment indicated that the growth and yield was significantly affected by the amount and time of applied potassium. Among the conducted treatments, the use of potassium application dose 120 kg per hectare with application technique (50% at sowing stage + 50 at flowering stage) may resulted in greater availability of potassium and lower transformation of potassium into non-exchangeable pool, which regulated the continuous growth of cells and tissues, resulting in better growth, physiological attributes nutrient uptake, yield, and yield quality of wheat crop grown in cadmium-contaminated saline soils. Therefore, the split application was found superior to the single basal dose of potassium fertilizer. Also, the accumulation of cadmium in leaves of wheat plants was significantly reduced with increasing potassium application, indicating a crucial role of potassium in alleviating cadmium hazardous effects on yield and yield quality of wheat crop.
References
Abd El–Mageed TA, El-Sherif AMA, Ali MM ABD, El-Wahed MH (2017) Combined effect of deficit irrigation and potassium fertilizer on physiological response, plant water status and yield of soybean in calcareous soil. Arch Agron Soil Sci 63:827–840. https://doi.org/10.1080/03650340.2016.1240363
Abd El-Mageed TA, Semida WM, Taha RS, Rady MM (2018) Effect of summer-fall deficit irrigation on morpho-physiological, anatomical responses, fruit yield and water use efficiency of cucumber under salt affected soil. Sci Hortic (amsterdam) 237:148–155. https://doi.org/10.1016/j.scienta.2018.04.014
Abd El-Mageed TA, El-Sherif AMA, Abd El-Mageed SA, Abdou NM (2019) A novel compost alleviate drought stress for sugar beet production grown in Cd-contaminated saline soil. Agric Water Manag 226:105831. https://doi.org/10.1016/j.agwat.2019.105831
Abd El-Mageed TA, Abdurrahman HA, Abd El-Mageed SA (2020) Residual acidified biochar modulates growth, physiological responses, and water relations of maize (Zea mays) under heavy metal–contaminated irrigation water. Environ Sci Pollut Res 27:22956–22966
Abd El-Mageed TA, Rady MOA, Semida WM, Shaaban A, Mekdad AAA (2021) Exogenous micronutrients modulate morpho-physiological attributes, yield, and sugar quality in two salt-stressed sugar beet cultivars. J Soil Sci Plant Nutr 21:1421–1436. https://doi.org/10.1007/s42729-021-00450-y
Abd El-Mageed TA, Rady MOA, Abd El-Wahed MH, Abd El-Mageed SA, Omran WM, Aljuaid BS, El-Shehawi AM, El-Tahan AM, El-Saadony MT, Abdou NM (2022) Consecutive seasonal effect on yield and water productivity of drip deficit irrigated sorghum in saline soils. Saudi J Biol Sci. https://doi.org/10.1016/j.sjbs.2021.12.045
Abdelfattah MA, Rady MM, Belal HEE, Belal EE, Al-Qthanin R, Al-Yasi HM, Ali EF (2021) Revitalizing fertility of nutrient-deficient virgin sandy soil using leguminous biocompost boosts Phaseolus vulgaris performance. Plants 10;10(8):1637. https://doi.org/10.3390/plants10081637
Abdou NM, El-Saadony FMA, Roby MHH, Mahdy HAA, El-Shehawi AM, Elseehy MM, El-Tahan AM, Abdalla H, Saad AM, AbouSreea AIB (2022) Foliar spray of potassium silicate, aloe extract composite and their effect on growth and yielding capacity of roselle (Hibiscus sabdariffa L.) under water deficit stress conditions. Saudi J of Bio Sci 1–12. https://doi.org/10.1016/j.sjbs.2022.02.033
Adhikari B, Dhungana SK, Kim ID, Shin DH (2020) Effect of foliar application of potassium fertilizers on soybean plants under salinity stress. J Saudi Soc Agric Sci 19:261–269. https://doi.org/10.1016/j.jssas.2019.02.001
Aksu G, Altay H (2020) The effects of potassium applications on drought stress in sugar beet. Sugar Tech 22:1092–1102. https://doi.org/10.1007/s12355-020-00851-w
Ali I, Khan AA, Munsif F, He L, Khan A, Ullah S, Saeed W, Iqbal A, Adnan M, Ligeng J (2019) Optimizing rates and application time of potassium fertilizer for improving growth, grain nutrients content and yield of wheat crop. Open Agric 4:500–508. https://doi.org/10.1515/opag-2019-0049
Anschütz U, Becker D, Shabala S (2014) Going beyond nutrition: regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. J Plant Physiol 171:670–687. https://doi.org/10.1016/j.jplph.2014.01.009
Anokye E, Lowor ST, Dogbatse JA, Padi FK (2021) Potassium application positively modulates physiological responses of cocoa seedlings to drought stress. Agronomy 11(3):563. https://doi.org/10.3390/agronomy11030563
AOAC (2000) Official methods of analysis, 17th edn. The association of official analytical chemists. Gaithersburg, MD. USA. Methods 925.10, 65.17, 974.24, 992.16
Bang TC, Husted S, Laursen KH, Persson DP, Schjoerring JK (2021) The molecular–physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol 229:2446–2469. https://doi.org/10.1111/nph.17074
Chakraborty K, Bhaduri D, Meena HN, Kalariya K (2016) External potassium (K+) application improves salinity tolerance by promoting Na+-exclusion, K+-accumulation and osmotic adjustment in contrasting peanut cultivars. Plant Physiol Biochem 103:143–153. https://doi.org/10.1016/j.plaphy.2016.02.039
Chapman HD, Pratt PF (1961) Methods of analysis for soils, plants and waters. University of California, Los Angeles
Chen, ZS. (2003) Relationship between heavy metal concentrations in soils of Taiwan and uptake by crops. Technical Bulletin - Food and Fertilizer Technology Center 2000 No.149 pp.16 pp. ref.52
Chen D, Cao B, Wang S, Liu P, Deng X, Yin L, Zhang S (2016) Silicon moderated the K deficiency by improving the plant-water status in sorghum. Sci Rep 6:1–14. https://doi.org/10.1038/srep22882
Clark AJJ, Landolt W, Bucher JBB, Strasser RJJ (2000) Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll a fluorescence performance index. Environ Pollut 109:501–507
Desoky EM, Elrys AS, Mansour E, Eid RSM, Selem E, Rady MM, Ali EF, Mersal GAM, Semida WM (2021) Application of biostimulants promotes growth and productivity by fortifying the antioxidant machinery and suppressing oxidative stress in faba bean under various abiotic stresses. Sci Hortic (amsterdam) 288:110340. https://doi.org/10.1016/j.scienta.2021.110340
Etsushi K, Ken JI, Satoshi U, Keiichi O, Cevayir C, Shizuo A (2011) Silica crystals and aluminum salts regulate the production of prostaglandin in macrophages via NALP3 inflammasome-independent mechanisms. Immunity 34:514–526
FAO (2019) The state of the world’s biodiversity for food and agriculture. In: Bélanger J, Pilling D (eds) FAO Commission on Genetic Resources for Food and Agriculture Assessments. Rome. 572 pp. http://www.fao.org/3/CA3129EN/ca3129en.pdf . Accessed 6 Dec 2020
Farooq S, Ahmad S, Hussain S, Hussain M (2018) Plant nutrients and abiotic stress tolerance. In: Hasanuzzaman M, Fujita M, Oku H, Nahar K, Hawrylak-Nowak B (eds) Plant Nutrients and Abiotic Stress Tolerance, Springer 391–413. https://doi.org/10.1007/978-981-10-9044-8
Gerardeaux E, Jordan-Meille L, Constantin J, Pellerin S, Dingkuhn M (2010) Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ Exp Bot 67:451–459. https://doi.org/10.1016/j.envexpbot.2009.09.008
Hasanuzzaman M, MHMB B, Nahar K, Hossain MS, Mahmud JA, Hossen MS, Masud AAC, Moumita FM (2018) Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 8 (3):31. https://doi.org/10.3390/agronomy8030031
Hasanuzzaman M, Bhuyan MHMB, Zulfiqar F, Raza A, Mohsin SM, Al Mahmud J, Fujita M, Fotopoulos V (2020) Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants 9:1–52. https://doi.org/10.3390/antiox9080681
Hossain A, da Silva JAT, Bodruzzaman M (2015) Rate and application methods of potassium in light soil for irrigated spring wheat. Songklanakarin J Sci Technol 6:635–642
Houmani H, Debez A, Freitas-Silva L, Abdelly C, Palma JM, Corpas FJ (2022) Potassium (K+) starvation-induced oxidative stress triggers a general boost of antioxidant and NADPH generating systems in the halophyte Cakile maritima. Antioxidants 11(2):401. https://doi.org/10.3390/antiox11020401
Hussain F, Malik AU, Haji MA, Malghani AL (2011) Growth and yield response of two cultivars of mungbean (Vigna radiata L.) to different potassium levels. J Anim Plant Sci 21:622–625
Islam MR, Haque KS, Akter N, Karim MA (2014) Leaf chlorophyll dynamics in wheat based on SPAD meter readingand its relationship with grain yield. Sci Agric 8(1);13–18. https://doi.org/10.15192/pscp.sa.2014.4.1.1318
Jackson ML (1973) Soil chemical analysis. Prentice Hall of India Private Limited, New Delhi, 498
Jensen ME (1983) Design and Operation of Farm Irrigation Systems. American Society of Agricultural Engineers, Michigan, USA, pp. 827, Michigan, USA
Johnson R, Vishwakarma K, Hossen MS, Kumar V, Shackira AM, Puthur JT, Abdi G, Sarraf M, Hasanuzzaman M (2022) Potassium in plants: growth regulation, signaling, and environmental stress tolerance. Plant Physiol Biochem 172:56–69. https://doi.org/10.1016/j.plaphy.2022.01.001
Kaya C, Ak BE, Higgs D (2003) Response of salt-stressed strawberry plants to supplementary calcium nitrate and/or potassium nitrate. J Plant Nutr 26:543–560. https://doi.org/10.1081/PLN-120017664
Khan R, Gurmani AR, Gurmani AH, Zia MS (2007) Effect of potassium application on crop yields under wheat rice system. Sarhad J Agric 23:277–299
Klute A, Dirksen C (1986) Hydraulic conductivity and diffusivity: laboratory methods. Methods of soil analysis: part 1-physical and mineralogical methods. Soil Science Society of America Amer Soc Agron, Madison, pp 687–734. https://doi.org/10.2136/sssabookser5.1.2ed.c28
Li J, Hu W, Lu Z, Meng F, Cong R, Li X, Ren T, Lu J (2021) Imbalance between nitrogen and potassium fertilization influences potassium deficiency symptoms in winter oilseed rape (Brassica napus L.) leaves. Crop J 10(2);565–576. https://doi.org/10.1016/j.cj.2021.06.001
Marschner H (Ed. ). (2011) Marschner’s mineral nutrition of higher plants. Academic press
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence - a practical guide. J Exp Bot 51:659–668. https://doi.org/10.1093/jxb/51.345.659
Meier U (2001) BBCH-monograph: Growth stages of mono-and dicotyledonous plants, 2th edn. Federal Biological Research Centre for Agriculture and Forestry. https://doi.org/10.5073/20180906-074619
Misskire Y, Mamo T, Taddesse AM, Yermiyahu U (2019) The effect of potassium on yield, nutrient uptake and efficiency of teff (Eragrostis tef Zucc. Trotter) on vertisols of north Western Ethiopian Highlands. J Plant Nutr 42:307–322. https://doi.org/10.1080/01904167.2018.1554681
Murtaza G, Javed W, Hussain A, Wahid A, Murtaza B, Owens G (2015) Metal uptake via phosphate fertilizer and city sewage in cereal and legume crops in Pakistan. Environ Sci Pollut Res 22:9136–9147. https://doi.org/10.1007/s11356-015-4073-y
Naciri R, Lahrir M, Benadis C, Chtouki M, Oukarroum A (2021) Interactive effect of potassium and cadmium on growth, root morphology and chlorophyll a fluorescence in tomato plant. Sci Rep 11:1–10. https://doi.org/10.1038/s41598-021-84990-4
Nagajyoti PC, Lee KD, Sreekanth TVM (2010) Heavy metals, occurrence and toxicity for plants: a review. Environ Chem Lett 8:199–216. https://doi.org/10.1007/s10311-010-0297-8
Oosterhuis DM, Dimitra LA, Eduardo M, Pettigrew TW (2014) Chapter three - the physiology of potassium in crop production. Adv Agron 126:203–233
Page AL, Miller RH, Keeney DR (1982) Methods of soil analysis part 2. American Society of Agronomy Inc, Madison (WI), Chemical and microbiological properties
Pettigrew WT (2008) Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol Plant 133:670–681. https://doi.org/10.1111/j.1399-3054.2008.01073.x
Premachandra GS, Saneoka H, Ogata S (1990) Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. J Agric Sci 115:63–66
Rady MM, Taha RS, Semida WM, Alharby HF (2017) Modulation of salt stress effects on Vicia faba L. plants grown on a reclaimed-saline soil by salicylic acid application. Rom Agric Res 34:175–185
Rady MOA, Semida WM, Abd El-Mageed TA, Howladar SM, Shaaban A (2020) Foliage applied selenium improves photosynthetic efficiency, antioxidant potential and wheat productivity under drought stress. Int J Agric Biol 24:1293–1300
Raj D, Antil RS, Garg R, Dahiya DS, Arora VK (2021) Impact of Intensive cropping systems on crop productive and change in soil fertility Impact of Intensive Cropping Systems on Crop Productivity and Changes in Soil Fertility : A Case Study on Benchmark Sites of Haryana. Indian J Fertilisers 16(10);988–996
Ravichandran M, Sriramachandrasekharan MV (2011) Optimizing timing of potassium application in productivity enhancement of crops. Karnataka J Agric Sci 24:75–80
Raza MAS, Saleem MF, Shah GM, Khan IH, Raza A (2014) Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. J Soil Sci Plant Nutr 14:348–364. https://doi.org/10.4067/S0718-95162014005000028
Rizwan M, Ali S, Abbas T, Muhammad Zia-ur-Rehman FH, Keller C, Al-Wabel MI, Ok YS (2016) Cadmium minimization in wheat: a critical review. Ecotoxicol Environ Saf 130:43–53
Römheld V, Kirkby EA (2010) Research on potassium in agriculture: needs and prospects. Plant Soil 335:155–180. https://doi.org/10.1007/s11104-010-0520-1
Rubio JS, Pereira WE, Garcia-Sanchez F, Murillo L, García AL, Martínez V (2011) Sweet pepper production in substrate in response to salinity, nutrient solution management and training system. Hortic Bras 29:275–281. https://doi.org/10.1590/s0102-05362011000300003
Sahi N, Mostajeran A, Ghanadian M (2021) Altering amino acid profile in Catharanthus roseus (L.) G. Don using potassium and ascorbic acid treatments. Open Biochem J 15:53–60. https://doi.org/10.2174/1874091x02115010053
Sardans J, Peñuelas J (2015) Potassium: a neglected nutrient in global change. Glob Ecol Biogeogr 24:261–275. https://doi.org/10.1111/geb.12259
Sardans J, Peñuelas J (2021) Potassium control of plant functions: Ecological and Agricultural Implications. Plants 10(2):419. https://doi.org/10.3390/plants10020419
Semida WM (2016) Hydrogen peroxide alleviates salt-stress in two onion (Allium cepa L.) cultivars. Am J Agric Environtal Sci 16:294–301
Semida WMM, Abd El-Mageed TAA, Howladar SMM (2014) A novel organo-mineral fertilizer can alleviate negative effects of salinity stress for eggplant production on reclaimed saline calcareous soil. Acta Hortic 1034:493–500. https://doi.org/10.17660/ActaHortic.2014.1034.61
Semida WM, Abd El-Mageed TA, Hemida K, Rady MM (2019) Natural bee-honey based biostimulants confer salt tolerance in onion via modulation of the antioxidant defence system. J Hortic Sci Biotechnol 94:632–642. https://doi.org/10.1080/14620316.2019.1592711
Semida WM, El-Mageed TAA, Abdalla RM, Hemida KA, Howladar SM, Leilah AAA, Rady MOA (2021) Sequential antioxidants foliar application can alleviate negative consequences of salinity stress in Vicia faba L. Plants 10(5):914. https://doi.org/10.3390/plants10050914
Shaaban A, Al OAAI, Nasr E, Khaulood MA, Ahmed AH, El SMA, Abdel MA, Wael R, Gamal MS, Abd El Mageed TA (2022) Filter mud enhanced yield and soil properties of water - stressed Lupinus termis L in saline calcareous soil. J Soil Sci Plant Nutr. https://doi.org/10.1007/s42729-021-00755-y
Sharma S, Singh J (2020) Evaluation of split application of potassium for improving yield and potassium uptake in wheat. Int J Chem Stud 8:459–464. https://doi.org/10.22271/chemi.2020.v8.i3e.9254
Schwartzkopf CARL (2018) Potassium, calcium, magnesium-how they relate to plant growth
Sedri MH, Roohi E, Niazian M, Niedbała G (2022) Interactive effects of nitrogen and potassium fertilizers on quantitative-qualitative traits and drought tolerance indices of rainfed wheat cultivar. Agronomy 12(1):30. https://doi.org/10.3390/agronomy12010030
Singh VK, Dwivedi BS, Rathore SS, Mishra RP, K. STM (2021) Timing potassium applications to synchronize with plant demand. In: Murrell TS, Mikkelsen RL, Sulewski G, Norton R, Thompson ML (eds) Improving Potassium Recommendations for Agricultural Crops. Springer, Cham. https://doi.org/10.1007/978-3-030-59197-7_13
Siddiqui MH, Khan MN, Mukherjee S, Alamri S, Basahi RA, Al-Amri AA, Alsubaie QD, Al-Munqedhi BMA, Ali HM, Almohisen IAA (2021) Hydrogen sulfide (H2S) and potassium (K+) synergistically induce drought stress tolerance through regulation of H+-ATPase activity, sugar metabolism, and antioxidative defense in tomato seedlings. Plant Cell Rep 40:1543–1564. https://doi.org/10.1007/s00299-021-02731-3
Singh KM, Singh HK (2020) Effect of foliar application of potassium nitrate on late sown wheat (Triticum aestivum L.) in mitigating terminal heat stress. J Pharmacogn Phytochem 9(6):492–495
Spoustová P, Synková H, Valcke R, Čevrovská N (2013) Chlorophyll a fluorescence as a tool for a study of the potato virus Y effects on photosynthesis of nontransgenic and transgenic Pssu-ipt tobacco. Photosynthetica 51:191–201
Sterckeman T, Thomine S (2020) Mechanisms of cadmium accumulation in plants. CRC Crit Rev Plant Sci 39:322–359. https://doi.org/10.1080/07352689.2020.1792179
Szczepanek M, Grzybowski K (2016) Yield and macronutrient accumulation in grain of spring wheat (Triticum aestivum ssp. vulgare L.) as affected by biostimulant application. Adv Crop Sci Tech 4:22.. https://doi.org/10.4172/2329-8863.1000228
Tang ZH, Zhang AJ, Wei M, Chen XG, Liu ZH, Li HM, Ding YF (2015) Physiological response to potassium deficiency in three sweet potato (Ipomoea batatas [L.] Lam.) genotypes differing in potassium utilization efficiency. Acta Physiol Plant 37:1–10. https://doi.org/10.1007/s11738-015-1901-0
Thornburg TE, Liu J, Li Q, Xue H, Wang G, Li L, Fontana JE, Davis KE, Liu W, Zhang B, Zhang Z, Liu M, Pan X (2020) Potassium deficiency significantly affected plant growth and development as well as microRNA-mediated mechanism in wheat (Triticum aestivum L.). Front Plant Sci 11:1–10. https://doi.org/10.3389/fpls.2020.01219
Tie Z (2007) Effects of nitrogen and potassium application levels on flag leaf photosynthetic characteristics after anthesis in winter wheat. Acta Agron Sin 33:1667–1673
Tränkner M, Tavakol E, Jákli B (2018) Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol Plant 163:414–431. https://doi.org/10.1111/ppl.12747
Wani JA, Malik MA, Dar MA, Akhter F, Raina SK (2014) Impact of method of application and concentration of potassium on yield of wheat. J Environ Biol 35:623–626
Wang Y, Wu W-H (2013) Potassium transport and signaling in higher plants. Annu Rev Plant Biol 64:451–476
Wang Y, Zhang Z, Liang Y, Han Y, Han Y, Tan J (2020) High potassium application rate increased grain yield of shading-stressed winter wheat by improving photosynthesis and photosynthate translocation. Front Plant Sci 11:1–10. https://doi.org/10.3389/fpls.2020.00134
Xu X, Du X, Wang F, Sha J, Chen Q, Tian G, Zhu Z, Ge S, Jiang Y (2020) Effects of potassium levels on plant growth, accumulation and distribution of carbon, and nitrate metabolism in apple dwarf rootstock reedlings. Front Plant Sci 11:1–13. https://doi.org/10.3389/fpls.2020.00904
Yang CM, Juang KW (2015) Alleviation effects of calcium and potassium on cadmium rhizotoxicity and absorption by soybean and wheat roots. J Plant Nutr Soil Sci 178:748–754. https://doi.org/10.1002/jpln.201500002
Yasin NA, Zaheer MM, Khan WU, Ahmad SR, Ahmad A, Ali A, Akram W (2018) The beneficial role of potassium in Cd-induced stress alleviation and growth improvement in Gladiolus grandiflora L. Int J Phytoremediation 20:274–283. https://doi.org/10.1080/15226514.2017.1374337
Zelelew DZ, Lal S, Tesfai Tsegai Kidane BMG (2016) Effect of potassium levels on growth and productivity of potato varieties. Am J Plant Sci 7:1629–1638
Acknowledgements
The authors thank the agriculture experimental center (Faculty of Agriculture, Fayoum University) for performing the field experiments.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
El-Mageed, T.A.A., Semida, W.M., Abdou, N.M. et al. Coupling Effects of Potassium Fertilization Rate and Application Time on Growth and Grain Yield of Wheat (Triticum aestivum L.) Plants Grown Under Cd-Contaminated Saline Soil. J Soil Sci Plant Nutr 23, 1070–1084 (2023). https://doi.org/10.1007/s42729-022-01104-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-022-01104-3