
Abstract
Over the last two decades, red ochre has played a pivotal role in discussions about the cognitive and cultural evolution of early modern humans during the African Middle Stone Age. Given the importance of ochre for the scholarly debate about the emergence of ‘behavioral modernity’, the lack of long-term spatio-temporal analyses spanning large geographical areas represents a significant gap in knowledge. Here we take a continent-wide approach, rather than focusing on specific sites, regions or technocomplexes. We report the most comprehensive meta-analysis of ochre use to date, spanning Africa between 500 and 40 thousand years ago, to examine data from more than a hundred archaeological sites. Using methods based on time averaging, we identified three distinct phases of ochre use: the initial phase occurred from 500,000 to 330,000; the emergent phase from 330,000 to 160,000; and the habitual phase from 160,000 to 40,000 years ago. The number of sites with ochre increased with each subsequent phase. More importantly, the ratio of sites with ochre compared to those with only stone artifacts also followed this trend, indicating the increasing intensity of ochre use during the Middle Stone Age. While the geographical distribution expanded with time, the absolute number of ochre finds grew significantly as well, underlining the intensification of ochre use. We determine that ochre use established itself as a habitual cultural practice in southern, eastern and northern Africa starting about 160,000 years ago, when a third of archaeological sites contain ochre. We argue that this pattern is a likely material manifestation of intensifying ritual activity in early populations of Homo sapiens. Such ritual behavior may have facilitated the demographic expansion of early modern humans, first within and eventually beyond the African continent. We discuss the implications of our findings on two models of ritual evolution, the Female Cosmetic Coalitions Hypothesis and the Ecological Stress Hypothesis, as well as a model about the emergence of complex cultural capacities, the Eight-Grade Model for the Evolution and Expansion of Cultural Capacities.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ochre is a generic term used by archaeologists to describe earth pigments, usually of reddish color, containing different geochemical elements including iron oxides, such as hematite (Fe2O3). More recently, some researchers have expanded the term to include rocks containing hydrated iron oxides such as goethite (FeO(OH)), which produce yellowish and brownish colors (Henshilwood et al., 2009; Hodgskiss, 2012; Wadley, 2010a). At archaeological sites attributed to the Middle Stone Age (MSA) of Africa, ochre is typically found as red, yellow or brown-colored lumps or nodules. It can also be found in the form of powder in discrete features or as a residue adhering to stone artifacts, bones, shells and rocks (d’Errico et al., 2009; Henshilwood et al., 2001b; Rosso et al., 2016; Wadley, 2010b).
Ochre plays an important role in contemporary theoretical discussions about the emergence of modern human behavior (d’Errico & Stringer, 2011; Henshilwood & Dubreuil, 2011; Henshilwood & Marean, 2003; McBrearty & Brooks, 2000; Nowell, 2010; Pettitt, 2011b; Sterelny, 2011; Watts, 2015). However, our knowledge about the broad spatio-temporal patterning of ochre use during the MSA remains limited. Here we present a meta-analysis of ochre use on the African continent, beginning with the first occurrences of ochre associated with transitional industries dated to the end of the Early Stone Age (ESA) and follow the trend throughout the subsequent MSA. Our study is not predicated on a specific site, region or technocomplex, nor a specific hominin taxon. Rather, this large-scale meta-analysis provides an understanding of the spatio-temporal patterning of ochre use from a long-term evolutionary perspective, that is to say, the longue durée. Within this framework, we try to answer the question of when and where habitual ochre use emerged and what significance this had for the development of ritual behavior during the MSA.
Current State of Archaeological Research
Ochre is a frequent archaeological find documented at numerous MSA sites in Africa. A considerable number of ochre pieces show evidence of anthropogenic modification, including faceted surfaces with striations that indicate grinding for the production of powder. Some pieces also show evidence of rubbing on soft materials (e.g., human or animal skin), random scoring with a sharp tool, and even abstract (quasi-) geometrical engraving (Dayet et al., 2013; Henshilwood et al., 2009; Hodgskiss & Wadley, 2017; Hodgskiss, 2013a; Watts, 2010). Researchers often refer to intensively ground pieces with three or more facets converging to a point as crayons (d’Errico et al., 2003, p. 19; Henshilwood et al., 2001b, p. 433; Watts, 2002, p. 5). However, experiments conducted by Wadley (2005b) indicate that most crayons could represent waste products resulting from intensive grinding. Nonetheless, some specimens from Blombos Cave in South Africa exhibit specific patterns of use wear on their rounded tips which suggest they were ‘…used to produce defined areas of color, consistent with a design (like a ‘crayon’), on a coarse hard surface’ (Rifkin, 2012b, p. 190).
More or less neglected as a meaningful archaeological class for much of the twentieth century, finds of ochre from the MSA have increasingly attracted attention from archaeologists. The last two decades have witnessed an abundance of newly found assemblages containing from hundreds to thousands of individual ochre pieces, weighing several kilograms. During this period, the reevaluation of old collections also increased. We discuss some of the more important finds in greater detail below.
Ochre has garnered the attention of a number of early career researchers who dedicated their doctoral theses to this topic (Bernatchez, 2012; Dayet, 2012; Hodgskiss, 2013b; Rifkin, 2012a; Velliky, 2018; Watts, 1998). In addition to carefully analyzing the remains of Paleolithic ochre use, some of these researchers conducted experiments to better detect and categorize use wear typically found on archaeological specimens and assess the expenditure of human labor needed to produce powder.
Current research into the MSA recognizes ochre, especially red ochre, as a valuable archaeological material with a long evolutionary history. In addition, ochre has become a key focus in theoretical discussions about the emergence of modern cognitive abilities as well as the evolution of ritual in the lineage Homo (d’Errico, 2008; Hodgskiss, 2014; Kuhn, 2014; Mithen, 2014; Rossano, 2015; Wadley, 2006; Watts, 1999, 2002, 2009, 2014, 2015; Watts et al., 2016).
The Utilitarian/Functional—Symbolic/Ritualistic Divide
Some scientists see ochre use during the MSA as a functional application, while others interpret it as a symbolically mediated behavior associated with ritual display. Researchers tested applications of ochre experimentally to see whether it could be used in compound adhesives (Wadley et al., 2009; Wadley, 2005b; Zipkin et al., 2014); as an ingredient in hide-tanning (Rifkin, 2011); as mosquito repellent (Rifkin, 2015; or as sunscreen (Rifkin et al., 2015a, 2015b). The experiments showed that ochre (and other materials in which characteristics such as grain size were similar) conferred benefits to some of these functional applications, although some tests produced conflicting results (Kozowyk et al., 2016; Zipkin et al., 2014). In addition, a medical usage of ochre is reported in some ethnographic accounts (Velo, 1984), but this would be very difficult to verify archaeologically. Practical applications are mirrored in the archaeological record through evidence for the use of ochre powder in compound adhesives for hafting technology (Gibson et al., 2004; Lombard, 2006a, 2006b, 2007; Wadley et al., 2004; Wojcieszak & Wadley, 2018) and ochre nodules used as soft knapping tools and abraders (Soriano et al., 2009). The presence of ochre, animal fat and muscle tissue on the edge of three scrapers from Sibudu (South Africa) could be the result of processing hides and/or coloring them (Wadley & Langejans, 2014). Ochre residues on polished bone awls from Blombos Cave also associate ochre with processing and/or coloring hides and skins (d’Errico & Henshilwood, 2007; Henshilwood et al., 2001b).
On the other hand, it is also possible to offer a number of persuasive arguments for the ritual use of ochre. First, archaeologists should notice an important development in the field of ritual studies during the last decades: it has proven to be very useful for research to separate the term ritual from religious belief (Grimes, 2014; Kreinath et al., 2006). Rituals may or may not be religious. Rather, the phenomenon should be conceptualized as a special mode of universal human psychosocial behavior, which appears cross-culturally in many different areas of life. Ritual can be broken down into individual components, which in turn are rooted in our evolved psychology (Hobson et al., 2018; Legare & Watson-Jones, 2016). Whether and what symbolic content is communicated through this mode of behavior constitutes a separate question (McCauley & Lawson, 2002, pp. 8–10). This opens up the possibility of identifying ritualistic behavior in the archaeological record without having to speculate about any specific symbolic meaning of the material or what people might have believed in.
The most obvious characteristic of ritual is the repetition of formal behavior (Hobson et al., 2018; McCauley & Lawson, 2002; Rappaport, 1999; Rossano, 2012; Whitehouse, 2013a). The frequency of red ochre at many MSA sites indicates a repetitive, formalized behavior seen over long periods. Another factor evinced in the archaeological record is the intentional selection of color. Compared to other easily available pigments such as yellow ochre from weathered shales or black manganese, red ochre dominates both on a continental scale and at the site level (Watts, 2002; Watts et al., 2016). In rigorously analyzed large MSA assemblages from several African sites, authors show that ancient people preferentially sourced and selected fine-grained raw materials with saturated blood-red or bright red hues for intensive processing. Such sites include Blombos (Watts, 2009), Pinnacle Point 13B (Watts, 2010), Pinnacle Point 5–6 (Bernatchez, 2012, pp. 300–310), Rose Cottage (Hodgskiss & Wadley, 2017) and Sibudu (Hodgskiss, 2012) in South Africa, and Porc-Epic (Rosso et al., 2017) in Ethiopia. The preference for these specific attention-grabbing hues over long periods cannot be explained in purely utilitarian terms. Rather this preference is likely a manifestation of non-instrumental, rule-bound, goal-demoted and causally opaque behavior. Goal demotion and causal opacity are terms used in the cognitive science of religion to describe central properties of ritual action: ‘that is, rituals either lack overt instrumental purpose, or their constitutive actions themselves are not immediately causally linked to the stated goal of ritual.’ (Hobson et al., 2018, p. 261).
Furthermore, the ochre found at two Pinnacle Point sites (13B and 5–6) stems from one preferred source, despite the availability of ochre from other sources in the region. Based on comparable ethnographic studies, Bernatchez (2012, pp. 212–229) suggests that the chosen source had a symbolic meaning for the inhabitants of the caves and that procurement of ochre was ritualized. Dayet et al., (2013, 2016) also documented similar behavioral patterns for the procurement of red ochre at another South African site, Diepkloof Rock Shelter. Geochemical analyses at some sites show that hunter-gatherers procured ochre from regional and supra-regional sources up to 80 km away when they occupied the site (Bernatchez, 2012, p. 388; Dayet et al., 2016; Salomon et al., 2012). People may have even transported some pieces from as far as 170 km to their place of utilization (Watts et al., 2016). The latter specimens from Stratum 2a in Canteen Kopje (South Africa), dated to more than 300,000 years ago, consisted of specularite—a type of ochre with a sparkling quality for which no utilitarian application is so far known archaeologically or ethnographically.
Dayet (2012, tables 28, 29) and Rifkin (2012b) documented the great effort needed to produce powder from high quality specimens of red shale, although the effort is highly dependent on the hardness of the material. In addition, preparing ochre powder requires suitable grinding equipment, which must also be procured. Persuasive evidence for the preparation of ochre-rich coloring material with (semi-) liquid binders made of high-calorie animal fat, bone marrow or milkfat was discovered at two MSA sites, Blombos (Henshilwood et al., 2011) and Sibudu (Villa et al., 2015). While those resources are nutritionally valuable, especially in circumstances of chronic caloric uncertainty, people used them for non-utilitarian purposes. The combination of these factors—long distance transport, the effort of powder production and the use of otherwise highly valuable nutritional substances as binders for the production of coloring materials—indicates that, at least in some cases, ochre use implies costly signaling. This constitutes another important aspect of ritual display (Henrich, 2009; Knight et al., 1995; Power et al., 2013; Rossano, 2015; Ruffle & Sosis, 2007; Soler, 2012; Sosis, 2019; Sosis & Alcorta, 2003).
If we summarize all the evidence thus far, it can be said that large parts of the MSA ochre finds reflect costly behavior that was repetitive, formalized and non-instrumental—hallmarks of ritual. But we can go one step further. Some archaeological evidence gives us clues to what the material was actually used for. The overall dominance of grinding use-wear on archaeological specimens from the MSA indicates the primary production of powder (Dayet et al., 2013; Henshilwood et al., 2009; Hodgskiss & Wadley, 2017; Hodgskiss, 2012, 2013a; Rifkin, 2012b; Rosso et al., 2017; Watts, 2002, 2010). This observation is supported by several concentrations of ochre powder. The most impressive evidence comes from the Howiesons Poort layers of Sibudu, where people produced large quantities of powder, which was then deposited on the cemented crusts of hearths (Wadley, 2010b). The evidence for intentional selection of color and fine-grained raw materials, as well as the use of binders, indicates the preparation of mostly blood-red colorants. The question is, for what purpose were these colorants produced? Importantly, red ochre residues have been found on many MSA shell beads (Bouzouggar et al., 2007; d’Errico & Backwell, 2016; d’Errico et al., 2005, 2008, 2009; Henshilwood et al., 2004; Vanhaeren et al., 2013). These perforated mollusks do not derive from ochred layers, and post-depositional processes do not account for the presence of iron oxide pigments on the shells. The authors conclude that the shell beads rubbed against an ochred material such as human skin, hair or an animal hide, and/or they were deliberately colored. Furthermore, the detection of microscopic ochre residues in some microcracks located on the smoothed edges of perforations suggests that the thread stringing the beads was impregnated with ochre. These detailed observations show that there was a link between ochre use and the self-decoration of MSA people, including body, face, hair and hide painting, and/or the deliberate coloration of personal ornaments. If we look at contemporary humans for comparison, self-adornment almost always communicates certain aspects of the wearer’s social identity (Davies, 2020; Dubin, 1999; Fisher, 1984). One could say that personal ornamentation adds a second skin to the body—a social skin (Power, 2010; Turner, 2012). Social identity in turn is often constructed through a series of ritual practices (Hermanowicz & Morgan, 1999, p. 198), especially in traditional societies (Belier, 1995; Guenther, 1999; Henrich, 2016, pp. 159–164; Lewis, 2015; Marshall, 1999; Tuzin, 2001). Moreover, in most cultures around the world body and face adornment in its endless variety is a significant part of the material culture during ritual performances themselves (Beckwith & Fisher, 1999, 2012; Ebin, 1979; Eisenhofer, 2005; Gröning, 2001; O’Hanlon 1989).
Furthermore, several pieces of ochre found at Blombos, Klasies River Cave 1, Klein Kliphuis, Rose Cottage, Sibudu and Pinnacle Point 13B—South African MSA sites dated between about 160,000 and 40,000 years—exhibit geometrical or quasi-geometrical engravings (d’Errico et al., 2012; Henshilwood et al., 2009; Hodgskiss & Wadley, 2017; Hodgskiss, 2013a; Mackay & Welz, 2008; Watts, 2010). To these finds we add a cross-hatched pattern drawn on a ground silcrete flake with an ochre ‘crayon’ from a 73,000-year-old Still Bay layer at Blombos (Henshilwood et al., 2018). Such artifacts provide a turning point in the discussion about the emergence of symbolically mediated behavior (d’Errico, 2003; d’Errico & Stringer, 2011; Henshilwood & d’Errico, 2011b; Hodgson, 2017; Tylén et al., 2020).
Ochre is also associated with some human burials (d’Errico & Backwell, 2016; Hovers et al., 2003) and a possible animal interment (Solecki, 1982). In addition, the ochre assemblage from Rhino Cave in the Tsodilo Hills of Botswana comes from a unique context. The hills are visible from across the Kalahari, but the site is hidden from view and difficult to access because of a steep climb and narrow entries. Ochre was deposited together with more than one hundred deliberately burned, abandoned or intentionally smashed, well-made MSA bifacial points, most of them made from a colorful non-local siliceous rock. The ochre and the stone tools were found together with tabular grinding slabs directly beneath a massive, freestanding rock face grooved with hundreds of vertically aligned cupules of varying sizes and shapes (Coulson et al., 2011). It is not an overstatement to interpret these findings as the material manifestation of costly signaling in multiple ritual performances within a special sequestered place.
When looking at the numerous ethnographic reports about ochre use in traditional hunter-gatherer and pastoralist societies worldwide, we see that contemporary humans used and still use this material in a variety of ways. It is striking that the ritual context of ochre use—primarily in the form of body and face painting but also for rock art, ritual objects and burials—predominates in the ethnographic record (Basedow, 1925; Beckwith & Fisher, 1999, 2012; Bernatchez, 2012, pp. 17–34; Horton, 1994; Knight et al., 1995; Lydall & Strecker 1979a, b, c; Rifkin, 2012a, p. 12, 2015; Peabody, 1927; Rudner, 1982; Sagona, 1994; Stafford et al., 2003, pp. 84–86; Taçon, 2004; Tönjes, 1911; Watts, 2010, suppl. pp. 7–9).
Considering all of the evidence, arguments and comparisons, we view a large proportion of ochre finds from the MSA as the material remains of past ritual activity. This does not negate the possibility that sometimes people also used this material for practical purposes, simple aesthetic pleasure or perhaps a combination of these. In this regard, ochre would have been a truly multifunctional raw material.
Color Psychology and Symbolic Culture
To examine the evolutionary significance of red ochre in the context of early ritual activity, we draw attention to the growing literature on color psychology (Elliot, 2015; Elliot & Maier, 2014; Elliot et al., 2015). A number of recent experiments applying rigorous empirical methods assessed the perceptional and behavioral impact of red stimuli in different psychological and cultural contexts. A considerable amount of data showing that the perception of red generally attracts more attention than other colors has accumulated over the last years (Buechner et al., 2014; Folk, 2015; Lindsey et al., 2010; Pomerleau et al., 2014; Sokolik et al., 2014; Tchernikov & Fallah, 2010). Red stimuli have nontrivial effects on specific human emotions and (mostly unconscious) influence on behavior in competition, achievement, danger and especially mating contexts (Beall & Tracy 2013; Elliot & Niesta, 2008; Elliot et al., 2013a, 2013b; Maier et al., 2015; Pazda & Greitemeyer, 2015; Schwarz & Singer, 2013; Shi et al., 2015; Stephen & Perrett, 2015; Tanaka & Tokuno, 2011; Velden et al., 2012; Williams et al., 2017; Wu et al., 2018). We point out that the specific symbolic meaning of the color red varies across cultures and historical epochs. The measured effects of red stimuli manifest themselves as statistically significant cross-cultural tendencies operating at deeper psychological levels, namely in basal, mostly unconscious, emotional and motivational reactions. Thus, these measured reactions presumably have a neurobiological basis shaped by natural and sexual selection.
In addition to cross-cultural experiments on humans, primatological studies indicate a deep evolutionary basis for at least some (non-symbolic) emotional and motivational effects of red stimuli. Researchers observed and experimentally tested the role of reddened skin in the realm of primate social and sexual signaling (Bielert et al., 1989; Gerald et al., 2007; Higham & Winters, 2015; Stallmann & Froehlich, 2000; Waitt et al., 2003, 2006; Zinner et al., 2004). Interestingly, where reddened skin plays a role, signaling often occurs in the form of ritualized displays (Bergman et al., 2009; de Waal, 2007, pp. 151–162; Dixson, 2012; Goodall, 1986, pp. 443–465; Higham et al., 2012; Petersdorf et al., 2017; Setchell, 2015; Setchell & Wickings, 2005; Setchell & Dixson, 2001). Such deep-seated evolutionary reactions to the color red constitute a psychological starting point upon which colorful and attention-grabbing ritual performances with symbolic meaning could later be built—with the help of material culture and through cultural evolution. Therefore, it is possible that red ochre applied to the body, face, hair or clothes initially played a role as an artificial amplifier of sexual signals in mating contexts, dominance in cases of competition, or warning in contexts of danger or death, thus exploiting ancestral cognitive biases in primates. If we continue this line of reasoning, it seems likely that with red ochre, these artificially amplified signals were used ever more strategically in ritualized displays as the ‘social brain’ (cf. Gowlett et al., 2012) evolved during the Pleistocene.
Even when red ochre is used for utilitarian purposes, it brings forth an inherent signaling quality the moment it is applied to any visible surface. As noted, the psychological impact of red stimuli is profound. Therefore, it hardly seems possible to distinguish between purely functional and symbolic use. In fact, we believe that the divide between the functional and the symbolic is a false dichotomy. In traditional, but also Western societies, functionality and symbolic meaning of material culture are inseparably intertwined, as Homo sapiens is ultimately a symbolic species (Deacon, 1997; Henshilwood & d’Errico, 2011a). This is not to say that there cannot be a heuristic distinction between these two categories. Rather, we view them as endpoints on a continuous spectrum, and not as an implicit binary divide of material culture.
From an evolutionary perspective, costly signaling precedes tool production and is observed in many biological organisms (Maynard-Smith & Harper, 2003; Zahavi & Zahavi, 1997). Therefore, early use of red ochre was more likely embedded in signaling than utilitarian contexts, also because the material’s color represents its most perceptually salient property. On the other hand, we cannot assume a priori that late Acheulean and early MSA material culture was permeated with symbolism right from the start, as it appears in ethnographic and modern cultures. It is very likely that the capacity for full-fledged symbolic communication did not appear in modern humans as a sudden miracle, but instead evolved over a considerable timespan with a multimodal and multicausal origin (Prieur et al., 2020). Critical pre-adaptions, such as the capacity for shared intentionality (Tomasello et al., 2005) and higher orders of theory of mind (Dunbar, 2003, 2009; Gowlett et al., 2012), first had to be set in place. That there was likely a prolonged transitional process in which it would be very difficult to distinguish between proto-symbolic and fully symbolic communication is also indicated by the fact that a plethora of animal species use iconic and indexical signals organized by complex ‘proto-grammars’ (Reznikova, 2007).
In some primate and non-primate species it is even possible to teach a limited number of true symbolic references (sensu Peirce, 1998), insofar as human teachers establish a stable relationship of trust with the individuals in question (Goodall, 1986, pp. 33–34; Lyn & Chenkin, 2018; Pepperberg, 2012). Based on his long-term observations over three decades in the Taï National Park in the Ivory Coast and elsewhere, Boesch (2012, pp. 108–127) even argued that some basic aspects of symbolic culture are already present in wild chimpanzees. Several theorists hypothesize that group ritualization played a key role in the emergence of fully symbolic culture by establishing and stabilizing the necessary relationships of trust between signalers and receivers and generating a shared domain of meaning (Deacon, 1997; Durkheim, 1912; Henrich, 2009; Knight, 1999, 2014; Power, 2009, 2014; Rappaport, 1999). If a large proportion of the red ochre from the African MSA does indeed manifest past ritual behavior, then it might represent a material remain of this very critical transitional process in human evolution, one which allows us to glimpse the gradual emergence of symbolic material culture.
What Does Habitual Ochre Use Mean and Why is it Important?
The term habitual often appears in the archaeological literature, for example, in association with the evolution of fire use by hominins (Roebroeks & Villa, 2011; Sandgathe, 2017; Shahack-Gross et al., 2014; Shimelmitz et al., 2014). These authors define the term as a repeated, regular, systematic, consistent, successive, long-term behavior in specific sites and/or regions. Similarly, the Oxford English Dictionary (https://www.oed.com; Accessed 26 May 2020) defines habitual as: ‘Of the nature of a habit; fixed by habit; existing as a settled practice or condition; constantly repeated or continued; customary.’ An additional perspective comes from primatology. Whiten et al. (1999) systematically compared the cultural behavior of chimpanzees (Pan troglodytes) across Africa from seven long-term field studies, which together accumulated 151 years of direct observation. The authors distinguished customary from habitual behavior: in their view, a behavior is habitual if it occurs repeatedly in several individuals, consistent with some degree of social transmission. It only becomes customary when the behavior occurs in all or most able-bodied members of at least one age-sex class (such as adult males).
We concur with these definitions and apply this meaning in our study. Therefore, we consider a feature of hominin behavior as habitual if it reflects a constantly repeated or continued part of the behavioral repertoire of a substantial part of a given population (but not necessarily of every group member or every age-sex class), which is socially transmitted. Archaeologically this should manifest itself across many broadly contemporaneous sites as compared to other commonplace archaeological materials such as lithic artifacts. We view the emergence of habitual ochre use as a proxy for the emergence of regular collective rituals.
The Evolutionary Origins of Ritual
From an evolutionary perspective, the human collective ritual is a special mode of behavior composed of different psychologically active building blocks, some older, some younger. Like other theorists (Deacon, 1997; Dissanayake, 2018; Power, 2009, 2014; Rossano, 2012, 2016), we assume that the oldest components are traceable to non-symbolic ritualization and costly signaling. These behaviors are observable in many non-human species today (Huxley, 1966; Krebs & Dawkins, 1991; Maynard-Smith & Harper, 2003; Zahavi & Zahavi, 1997). When looking at human evolution, we must relate these behaviors to the special problem of encephalization.
Ritualization is an evolutionary process in which an accidentally informative part (e.g., gestures, vocal expressions) of an otherwise longer, functional behavioral sequence is decoupled, isolated, formalized, exaggerated and repeated in order to function purely as a communicative signal (Huxley, 1914; Tinbergen, 1952; Zahavi, 1980). The selection pressure is driven by the observer (e.g., prospective mate, predator, prey, offspring) who is interested in assessing the behavior of the signaler. The observer is reacting to these signals in a manner that vitally affects the fitness of the signaler (Krebs & Dawkins, 1991). In socially complex primates, ritualization serves a function in greeting, social bonding, trust enhancement, conflict mitigation, displays of dominance, mating, warning signals, and coordination of group spacing and travel (Arcadi et al., 1998; Crockford et al., 2017; Call & Tomasello, 2007; Dal Pesco & Fischer, 2018, 2020; de Waal, 1989, 2007; Goodall, 1986; van Leeuwen et al., 2012; Pollick & de Waal, 2007; Smuts & Watanabe, 1990; Tomasello & Call 2019; Whitham & Maestripieri, 2003). In short, ritualized behavior is an important mechanism for the regulation of social life in primates. Especially remarkable are the behavioral complexity and rich cultural repertoires of our closest primate relatives (Pan troglodytes and paniscus). Long-term observations of wild populations and in controlled environments revealed cultural traditions common in some communities, but not in others (Boesch, 2012; Horner & de Waal, 2009; Luncz & Boesch, 2014; Roffman et al., 2015; Whiten et al., 1999). Not only do these studies show how different chimpanzee communities have different socially transmitted traditions of tool use, they also demonstrate arbitrary behavioral conventions in the domain of ritualization, such as greeting and social grooming behaviors (Bonnie et al., 2007; McGrew, 2017; van Leeuwen et al., 2012, 2014). Recently, a large group of authors reported a previously unknown ritualized behavior in four different chimpanzee populations in West Africa, which includes both special ritual places and material objects (Kühl et al., 2016). Individuals hurl or hit stones against certain trees, throwing them into specific hollow tree stumps or cavities between the rib-like buttress roots of a certain tree. This action forms cairn-like stone accumulations that can consist of up to thirty stones. During handling of the stones or directly after throwing, the adult individuals perform typical powerful calls (‘pant hoots’) while the other members of the troop watch the spectacle. The groups visit the same place regularly, using the same trees and stones to perform the same sequence of behaviors. This specific cultural display occurs without any recognizable practical function and includes some important building blocks of human ritual behavior, such as the repetition of stereotypical, rule-bound, non-utilitarian, socially transmitted behavior, performed in front of an audience in special places incorporating the use of special material objects.
Equally complex are the behavioral reactions of chimpanzees confronted with dead group members. Such responses include testing for reaction by inspecting, shaking or covering the corpse with leaves; emotional calls; guarding or carrying the dead body for hours (and sometimes days); ritualized renegotiation of the dominance hierarchy and social relations through aggressive displays, submission gestures, and mutual social grooming near the corpse (Anderson, 2011; Anderson et al., 2010; Boesch, 2012; Goodall, 1986; Matsuzawa, 2011; Stewart et al., 2012; Teleki, 1973; van Leeuwen et al., 2016, 2017). A number of researchers see these emotional responses and behaviors in the domain of death as a basis for the development of more complex burial ritual in hominins during the Pleistocene (Anderson, 2011; Boesch, 2012; Pettitt, 2011a).
In addition, several studies report emotionally charged and fairly elaborated ritualized displays performed by individual primates. These displays are not directed at other individuals, but rather at overwhelming natural phenomena such as bushfires, storms, heavy rain and big waterfalls (Goodall, 2005; McGrew, 2004; Pruetz & LaDuke, 2010; Tennie & van Schaik, 2020). Whether or not these displays are a sign of awe is speculative, but the observations suggest that these ritualized behaviors lead to an internal regulation of emotion in situations of uncertainty and fear, just as they do in humans (cf. Boyer & Liénard, 2006; Hobson et al., 2018; Lang et al., 2019, 2020).
Costly signaling evolves if there is a conflict of interest between signaler and observer, or at least uncertainty about the honesty of the transmitted information. In such contexts, a signal must be costly and difficult to fake in order to be reliable for the observer, otherwise the observer will ignore the signal. A well-known example of this mechanism is the peacock’s tail. Because of its enormous size and colorfulness, the tail exacts energetically high costs for the peacock. Yet it becomes a handicap when a peacock must escape from a predator. Only a healthy, well-fed individual, resistant to parasites is able to develop a particularly large and magnificent tail and survive long enough. It is not possible to fake this kind of signal with a cheap trick. The display of a very costly and impractical tail is therefore an honest phenotypical signal to the peahen of the male carrier’s otherwise invisible true genetic fitness (Zahavi & Zahavi, 1997). In the process of sexual selection, peahens selected mates with ever more expensive tails over many generations. Evolutionarily informed psychologists and anthropologists successfully transferred this basic principle—that costly signals are honest and difficult to fake—into the realm of human collective rituals. As many researchers have noted and further investigated, most rituals are very costly for the participants in one way or another, because they can involve a great quantity of material resources as well as time, repetition, physical effort, risk taking and physical suffering. Although the nature and severity of ritual costliness varies greatly between different ritual types and societies (Kapitány et al., 2020; Atkinson & Whitehouse, 2011; Sosis et al., 2007), a consensus is emerging. The mechanism of costly signaling represents an effective psychological technique to signal and test true social, emotional and moralistic commitment to the group, and thus deters free-riders (Fischer & Xygalatas, 2014; Henrich, 2009; Irons, 2001; Knight et al., 1995; Power et al., 2013; Purzycki & Arakchaa, 2013; Rossano, 2015, 2016; Ruffle & Sosis, 2007; Soler, 2012; Sosis, 2019; Sosis & Alcorta, 2003; Sosis & Bressler, 2003; Whitehouse & Lanman, 2014).
Encephalization in hominin evolution, driven by increasing social (Gowlett et al., 2012) and cultural complexity (Muthukrishna et al., 2018), caused increased maternal energetic requirements in pregnancy and parenting, which in turn advanced the need for cooperative breeding (Burkart et al., 2009, 2014; Hrdy, 2009). In addition, the evolutionary trend towards larger social networks (probably one of the main driving factors of encephalization), whose number exceeded the local residence group and incorporated more genetically unrelated individuals with competing interests, led to ever more complex social environments (Gamble et al., 2014; Gowlett et al., 2012; Sterelny, 2014). Because these enlarged social networks were no longer composed purely of genetic relatives, and manual grooming among all members of the group to establish individual reciprocal relationships became impossible, it is not natural for such entities to behave as a cohesive unit (Dunbar, 2009). Thus, mechanisms of kin selection and reciprocal altruism on their own became insufficient. The evolution of new psychological adaptations may have been one consequence of the challenges of an enlarged and more complex social structure, as was the need for alloparenting (e.g., norm-psychology: Chudek & Henrich, 2011; group-mindedness/tribal social instincts: Richerson & Henrich, 2012; Tomasello et al., 2012; prosocial disposition for cooperative breeding: Burkart et al., 2009). Another result was the advent of powerful new cultural strategies to maintain cooperation, prosociality and group cohesion. One of those new cultural strategies was collective ritual (Whitehouse & Lanman, 2014). As Pinker (2016, p. 874) states, wasteful ritualizing in the context of human sociality is necessary because the group as such is not an elementary cognitive intuition that automatically triggers instinctive loyalty. The phenomenon of human collective ritual in turn developed from the aforementioned much older capacities for ritualized behavior and costly signaling in conjunction with evolutionarily younger adaptations that evolved some time after the split of our lineage from the chimpanzees. Younger psychological adaptations include the capacity for shared intentionality (Tomasello et al., 2005); group synchronization (Launay et al., 2016; Reddish et al., 2013); an innate proclivity for overimitation of non-instrumental behavior in early childhood (Keupp et al., 2013; Lyons et al., 2007; Nielsen & Tomaselli, 2010); and the capacity for symbolic communication (Botha & Knight, 2009a, b; Cole, 2017; Dor et al., 2014; Sterelny & Hiscock, 2014).
Furthermore, the evolution of all symbolic communication systems is initially impeded by a cooperative dilemma, as deceptive signaling is abundant in nature (Bergstrom, 2009). In order to communicate with a symbolic code that is not visually, physically or temporally bound to a real-world object, agent, behavior or quality, the signaler and the receiver have to trust each other. Symbols make lying, tactical deception and exaggeration very easy (Henrich, 2009, 2016, pp. 256–258; Lachmann & Bergstrom, 2004). Thus, some kind of mechanism to secure trust and generate a shared domain of meaning between signalers and receivers has to be in place before symbolic communication can be established. If it is true that trust-enhancing and meaning-generating collective rituals were needed to establish the first jointly shared fictions (i.e., symbolism) by solving the cooperative dilemma, then the emergence of habitual collective rituals was one important prerequisite for the evolution of symbolic communication (Deacon, 1997, pp. 402–407; Durkheim, 1912; Power, 2009; Rappaport, 1999, pp. 54–56; Rossano, 2016; Watts, 2009). Hence, it may be that the emergence of habitual ochre use during the MSA of Africa marks an important transition in the cognitive evolution of H. sapiens.
Testing the Predictions of Different Models
Our study examines two conceptual frameworks related to the evolution of ritual behavior and their specific predictions for the emergence of habitual ochre use. The first one is known as the Female Cosmetic Coalitions Hypothesis, while we call the second the Ecological Stress Hypothesis. In addition, our data have likely implications for broad scenarios about the emergence of complex cultural capacities during the evolution of hominins, such as the Eight-Grade Model for the Evolution and Expansion of Cultural Capacities.
Female Cosmetic Coalitions (FCC) Hypothesis
The FCC hypothesis was initially proposed by Knight, Power and Watts in 1995 and later updated with new data from primatology, paleoanthropology and archaeology (Power et al., 2013). The model links early ochre use to processes of encephalization, sexual selection and mechanisms of culture–gene coevolution. The evolution of increased cranial capacities with higher energetic costs of gestation and prolonged periods of lactation and childhood development during the Middle Pleistocene placed females under greater pressure to recruit reliable male support for parenting and nutrition (Kuzawa et al., 2014). Through their concealed ovulation, human females do not exhibit overt signs of estrus. Therefore, the only available cue to the female fertility status for males would be menstrual blood. Although menstruation occurs at a non-fertile period, it signals sexual maturity, non-pregnancy and an imminent ability to conceive.
According to the FCC hypothesis, the earliest use of ochre emerged as female coalitions ritualistically painted their bodies to signal a deceptive ‘sham menstruation’ in order to manipulate and control male parental investment and productive labor to meet the aforementioned increased energy requirements caused by encephalization (Knight et al., 1995). By painting themselves with red ochre (and/or other red cosmetics), non-fertile females pretended imminent fertility and triggered male attention. While it is unlikely that males would be fooled by this signal, the menstruation ritual would have constructed the first symbolic reality understood by both the signaling female coalition and male observers. The symbolic message was that without reliable male investment there would be no sexual access to any female member of the coalition. According to the model, initial use of ochre should not pre-date the encephalization associated with H. heidelbergensis (~ 800–700,000 years) (Power et al., 2013, p. 44; Watts, 2014, p. 217, 2015, p. 324).
In its early phase, ochre use should be irregular, sporadic and local as menstruation rituals would only be performed ad hoc when a female in the coalition was menstruating. The emergence of habitual ochre use should correlate with the last major encephalization event associated with H. sapiens and the emergence of modern cranial volumes. When the updated version of the FCC hypothesis was published, this was estimated to occur about 200,000 to 150,000 years ago (Power et al., 2013, p. 44). The FCC model posits that at this point, ritual performances were regularly planned and organized to motivate male investment regardless of whether a local female was actually menstruating (Knight et al., 1995, p. 81; Power et al., 2013, pp. 43–44; Watts, 2014, pp. 218–222). Since the model is predicated on a runaway process of sexual selection, it predicts an abrupt rather than gradual emergence of habitual ochre use (Power et al., 2013, p. 44; Watts, 2014, p. 217; Watts et al., 2016, p. 299).
Ecological Stress Hypothesis (ESH)
According to Rossano (2015), increased ochre use is a proxy for costly rituals that emerged specifically during periods of ecological stress and competitive situations when drought-related migrations were probable. Variations in the primary driving factors of insolation and global temperature caused dramatic fluctuations in climate, starting about 150,000 years ago, leading to displacements of resources and local scarcities. Especially severe mega-droughts that occurred periodically between 135,000 and 75,000 years ago in the tropical zone of Africa might have brought about genetic bottlenecks and migration events of populations of H. sapiens (Blome et al., 2012; Cohen et al., 2007; Kim et al., 2014; Scholz et al., 2007). Those migratory events were either from the tropical zone into adjacent subtropical regions, or, as precipitation increased, back into the tropical zone, and may have caused stress and conflict between formerly isolated groups. Costly rituals are especially important to maintain group cohesion during times of difficult circumstances and intergroup conflict (Sosis et al., 2007). According to this model, which we name the Ecological Stress Hypothesis (ESH), ochre use should fluctuate in the aforementioned regions as a function of migratory events and reach several different peaks between 150,000 and 70,000 years ago.
Eight-Grade Model for The Evolution and Expansion of Cultural Capacities (EECC)
The eight-grade model for the evolution and expansion of cultural capacities (EECC) is a broad framework that illustrates the expanding complexity of cultural capacities during the evolution of hominins (Haidle et al., 2015). The model is based on the extension of problem–solution distances in tool behavior observed from the field of ethology and the archaeological record (Haidle et al., 2015, fig. 3), with each grade showing evidence for increased cultural capacity. Based on ethological observations, the first four grades herald the beginnings of cultural capacities starting with (1) social information and (2) social learning, which then grade into (3) traditions and (4) basic culture. Based on archaeological evidence, the last four grades of cultural capacity include (5) modular, (6) composite, (7) complementary, and (8) notional phases.
Depending on whether ochre use was functional or symbolic, evidence from the archaeological record indicates two possible interpretations within the model (Haidle et al., 2015, table 1). If ochre was an ingredient in (semi-) liquid compounds (Henshilwood et al., 2011; Villa et al., 2015) or adhesives (Lombard, 2007; Wadley et al., 2004, 2009; Zipkin et al., 2014), we can infer the sixth grade, composite cultural capacity. Haidle et al. (2015) estimate that the sixth grade was reached between 500,000 and 200,000 years ago and is associated with critical cognitive developments: ‘Composite tools and materials expand cultural capacities by welding independently existing ideas and solutions into new concepts. …The resulting product possesses new qualities that go beyond the qualities of the parts, … that may be obtained at different times and in different places’ (Haidle et al., 2015, pp. 57–58).
But if the emergence of habitual ochre use is interpreted as indicating regular collective rituals that established socially shared fictional entities, attributes and relationships (i.e., symbolism), then it can be understood to indicate the eighth grade, notional cultural capacity. Based on current evidence, Haidle et al. (2015) estimate that the eighth grade was achieved by at least 40,000 years ago, and possibly as early as 130,000 years ago. The eighth grade includes socially transmitted information which incorporates non-physical concepts that can only be manipulated within the mind. While those symbolic concepts do not necessarily need a link to a physical object, they are often combined with one in order ‘… to form a composite with new functional qualities emerging out of the basic physical qualities and [conveying] a certain meaning …’ (Haidle et al., 2015, p. 59). Their full potential only unfolds within the social domain of a given group that shares the same mental templates. If the symbolic interpretation of habitual ochre use holds true, our findings could refine the dating of the emergence of notional cultural capacity in H. sapiens, and with it, a level of cognitive complexity that can only be considered as strictly modern.
Climate, Environment and Geography
Our study examines sites from a broad geographic context spread across Africa from the time between 500,000 and 40,000 years ago. By default, this approach reduces the bias of a specific environment, region or time in exerting undue influence on the results. This strategy is important because the sites included in the analysis are located in different ecological settings, including coastal, near-coastal, inland, highland and montane zones. Their vegetational settings are also highly variable, with sites in ecozones where fynbos, savannah, grassland, woodland, forest or desert predominate.
Africa is climatically diverse, showing great geographic variation in temperature and precipitation. Temperatures can range from well below freezing in the Highveld of South Africa to the extremes of the Sahara, while variable amounts of rainfall across the seasons can provide their own challenges. The varying elevation of the sites further drives the differentiation of climate and environment. While local glaciation played only a minor role, eustatic responses to global glacial cycles affected sea level. This fluctuation had an impact on coastal and near-coastal zones, opening up new landscapes, or erasing them. The same holds true for tectonic events such as earthquakes and volcanic eruptions.
We recognize that these factors contributed to the establishment of substantial variation across the African continent in different ways, at different times and in different places. However, many of these appear to follow the rhythm of orbital motion (Fig. 10g), which influences the African macroclimate through two main forcing factors, insolation and global temperature. Direct summer-season insolation is the primary driver determining hydroclimate, which includes rainfall and seasonality. It is strongest during phases with a high amplitude of precession, as indicated by a large number of paleoclimatic data (Nash & Meadows, 2012). The second major driver is global temperature, represented by the benthic δ18O proxy (Lisiecki & Raymo, 2005; Fig. 10f), with its feedback on sea surface temperatures, latent heat flux and ice-related processes affecting marine and terrestrial ecosystems of Africa. Especially during phases of low amplitude of precession, factors other than insolation, such as high-latitude forcing and greenhouse gases, gain importance in their impact on the African climate. The effect of these factors varies within the three macroclimatic zones of Africa: the tropics between + 23.5° and − 23.5° latitude and both northern and southern subtropical latitudes up to about 35°. Two hypotheses can be derived from climatic data and modeling results: (i) an asymmetric relationship with humid conditions in one hemisphere and arid conditions in the other, caused by millennial-scale, anti-phased variations of insolation; and (ii) a symmetric expansion and contraction of the tropical rain belt. Singarayer and Burrough (2015) conclude that both hypotheses operated mutually and influenced African paleoclimate on orbital scales. In addition to the Northern–Southern hemisphere antiphase signal modulated by precession, Kaboth-Bahr et al. (2021) identified alternating humid and arid phases between East and West Africa (Fig. 10e). These are likely driven by orbital eccentricity affecting Walker Circulation and El Niño Southern Ocean oscillation (ENSO) variability.
In sum, the sites analyzed in this study are spatially and temporally widespread. We therefore acknowledge the existence of significant variability in many factors, such as temperature, precipitation and seasonality. However, this heterogeneity balances the effect of climatic and environmental change on a continental scale and levels the results of this study (cf. Kandel et al., 2016; Steele & Klein, 2009).
Materials and Methods
Data Sources
We reviewed the literature to establish which archaeological sites in Africa document the presence of ochre between 500 and 40 thousand years ago. We limited the study to Africa in order to focus on behavioral changes in H. sapiens and its immediate predecessors, selecting the period of study to encompass the entire MSA and the preceding transitional industries of the ESA. Recent research has shown that some archaic hominin populations still existed at the beginning of the MSA on the African continent (Dirks et al., 2017; Grün et al., 2020; Hammer et al., 2011). Hypothetically, they also could have contributed to the archaeological record. But given the current state of research, it seems rather unlikely that they were responsible for the bulk of the MSA ochre finds. We examined ochre because of its status as a proxy for ritual and because of its durability and frequent co-occurrence with assemblages of stone artifacts. We also documented archaeological sites lacking ochre, but with stone artifacts, to place the sites with ochre into a broader context.
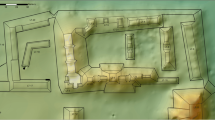
Our research focused on primary literature, most of which is written in English or French, and less frequently German or Italian. We also integrated new studies of previously documented ochre assemblages and contacted researchers directly to clarify piece counts or other details about the finds. In all, we collected information on more than a hundred ochre-bearing sites and entered 86 (Table 1, Fig. 1) into a database managed by the research project ‘The Role of Culture in Early Expansions of Humans’ (ROCEEH: http://www.roceeh.net). Together, these sites yield more than 25,000 individual ochre finds, representing a time span of nearly half a million years.

Map of Africa showing distribution of sites with lithics and those with ochre, keyed to Table 1
ROAD (ROCEEH Out-of-Africa Database) is a web-based platform uniting spatial information with data about assemblages from archaeological, paleoanthropological, paleontological and botanical localities plus bibliographic information (Haidle & Kandel, 2018; Haidle et al., 2010; Märker et al., 2011). Every site entered into ROAD is structured through its geological and archaeological profiles. The profiles contain layers which are stratigraphically ordered. Each layer is ascribed to a chronological framework based on an age model which incorporates both radiometric and relative dating as well as cultural chronostratigraphy defined through regional technocomplexes. An assemblage may consist of stone artifacts, organic tools, ochre, ‘symbolic’ finds and features along with human, animal and plant remains. Each assemblage is assigned to a specific geological and, as appropriate, archaeological layer. ROAD integrates this data and provides a means to query and analyze the spatial and chronological distribution of assemblages. The database includes a Map Module offering basic cartographic tools, allowing sites to be plotted on a wide array of topographic, climatic, ecological or vegetational maps. ROAD also offers other specially developed analytical tools, such as the ‘scenario tool’ to define a preferred hominin species, a ‘time slider’ to examine the distribution of finds during adjustable windows of time, and ‘time slice’, described below.
We evaluated the published data from each locality based on the character and quality of the ochre artifacts reported, the nature of the excavation, the stratigraphy of the site and the reliability of its dating. Due to the way we enter and link data in ROAD, all of these aspects are prerequisites for building a robust and reliable framework for analyzing data. As a result, we eliminated datasets judged to be from questionable contexts. Consequently, some sites frequently mentioned in the literature are not part of this analysis. We excluded sites with mixed, doubtful or poorly described assemblages and poor (or no) dating. In the end, we removed 18 sites from our analysis for reasons cited in Table 2.
Time Averaging
To overcome problems associated with variable data inherent in our age model, we employed the statistical concept of time averaging. We understand time averaging as a broad concept based on the calculation of moving averages which applies to: (i) aggregated time series; (ii) dating of stratified sequences; and (iii) the likelihood that an assemblage dates to a specific time. In the first case, moving averages provide an approximation of long-term trends in such disparate areas as paleoclimate, weather, the stock market, census analysis, currency exchange and infection rates (e.g., National Institute of Standards and Technology, 2012). To illustrate the second example, we draw attention to Lyman (2003), who uses time averaging to compare disparate datasets in zooarchaeology, whereas Kowalewski (2014) applies a similar concept to paleontology with the understanding that the accumulation of individual fossils in a geological formation rarely represents a synchronous event. For the third example, we point to Sadr’s (2009) interpretation of settlement intensity based on radiocarbon dates from different archaeological sites. Robinson et al. (2019) also follow this view, using radiocarbon ages in models of population ecology. Finally, Bluhm and Surovell (2019) examine the distribution of radiocarbon dates from archaeological sites, using them as proxies for paleodemography.
We consider time averaging to be a useful tool when a degree of variability is intrinsic in a series of data. The method is particularly helpful when examining trends over longer periods because it filters out noise in a signal. To resolve issues related to cases (ii) and (iii), we apply the concept of time averaging to analyze the age models entered into ROAD. Using the age models, we conducted a time series analysis (case i) with a tool called time slice and crosschecked the results with a statistical method called finite mixture distribution. While these methods have their limitations, in that they dissolve the fine-grained nature of a series of data, their advantage is that they provide an overview of general trends through time.
Time Slice Tool
To perform time averaging, we developed an analytical tool in ROAD called time slice. This tool tallies the numbers of localities and assemblages meeting certain criteria as specified in a predefined query. The user selects a query and defines the range of time to analyze by choosing a minimum and maximum age. Next the user picks different windows of analysis (time slices), defined in years. The method is a binning technique using variable bandwidths applied to a moving window. Thus, the tool allows the user to slide the analytical window across the scope of analysis at a given interval, which we call time step, also defined in years. In this way the analytical window (= time slice) is moved in an overlapping manner to calculate mean values (Fig. 2). The time slice tool yields a graphical representation of the results and allows downloading of the queried data (Fig. 3).

Schematic diagram explaining the terms ‘time slice’ and ‘time step’. We tested 13 different combinations with respect to the ochre data in ROAD (see Table 3)

Screenshot of the time slice tool in ROAD
For this analysis we developed two queries, one that generated a list of African localities with assemblages of ochre, and another for stone artifacts. We selected 13 combinations (Table 3), applying time slices of 10, 20, 30 and 40 thousand years and varying the time steps at intervals of 5, 10, 15, 20, 30 and 40 thousand years. Our aim was to examine the entire chronological range from 500 to 40 thousand years ago without losing too much resolution. Since radiocarbon dating covers only a small portion of the entire chronology, other radiometric methods provide ages for the majority. Analysis at an interval finer than 5000 years seemed overly optimistic, as this was often considerably more precise than the standard deviations of the dating methods. On the other hand, a resolution coarser than 40,000 years seemed unlikely to yield meaningful results.
One important consideration is that assemblages in ROAD belong to distinct geological and archaeological layers. Those layers are not only dated through radiometric methods, but also using relative means (e.g., marine isotope stage, paleomagnetic stratigraphy, biostratigraphy) as well as cultural stratigraphy. The layers may even be of unknown age. Several factors can contribute to increased uncertainty in ages, for example, large standard deviations in radiometric dating and broadly defined chronological frames for correlating biological or cultural entities. Thus, factors such as overlapping age ranges and dates with wide spreads affect the results of our queries (see the lithics plot in Fig. 7). Nonetheless, we relied on the information presented in the literature as the basis of our dataset.
Finite Mixture Distribution
To crosscheck the results of our time slice analysis, we applied a probability density function to estimate finite mixtures from the data (cf. Baxter, 2017). Using this method, we simplified the dates to a number of Gaussian distributions and aggregated them to an overall density of dates over time. It allowed us to examine some of the issues of uncertainty mentioned above, such as variable temporal accuracies explained through different dating methods (Fig. 4, Type A and B) and the existence of multiple dates assigned to the same layer (Fig. 4, Type C). This method is based on two assumptions. First, we assume that an assemblage in our record represents one event in time, expected to have occurred within a given time range. The latter represents the normally distributed measurement error of the dating method. Second, we assume that an event falls within a time range with a probability of p = 99%. Consequently, we expect the time range to represent Z = 2.58 standard deviations of the assumed Gaussian probability distribution. The presence of an assemblage in the timeline can be expressed as a probability density function
where μ is the mean of the upper and lower boundary of its time range and σ is the time range divided by Z. Based on these assumptions, a record with dated assemblages can be considered as a finite set of mixture components p1(x),…,pn(x) that are elements of a univariate and finite mixture distribution
where pi(x) is the probability density function of the mixture components and wi is a weighting factor. The weighting factor mitigates the overemphasis of assemblages with multiple dates. Finally, to compare the results with the time slice tool (see above), we examined the effect of smoothing by using moving averages equal to 5, 10, 15, 20, 30 and 40 thousand years. We performed calculations in the statistical computing language R v.4.0.5 (R Core Team, 2021) and the CRAN package ‘distr’ (Ruckdeschel et al., 2006).

Theoretical examples of the mixture distribution method. The figure displays three types of assemblages, their dates represented by the age range (horizontal lines at the bottom) and probability density function f(x). The hypothetical assemblages are precisely dated (Type A = red), imprecisely dated (Type B = green) or comprise multiple dates (Type C = blue). The mixture distribution p(x) is the result of the combination of the three. The black line represents the mixed density (Color figure online)
We see two advantages inherent in this method. Since the area under the curve of every p(x) equals one, precisely dated assemblages with narrow time ranges produce more pronounced peaks (Fig. 4, Type A), while less precisely dated assemblages with broad time ranges have dampened peaks (Fig. 4, Type B). The weighting function also allows us to compensate for overestimation, where one assemblage is associated with several dates because it ranges across a number of archaeological or geological layers (Fig. 4, Type C).
Additional Limitations
For transparency, we summarize the limitations inherent in our research. We mention these wide-reaching factors not to diminish our results, but for the sake of fairness, while acknowledging that they are an intrinsic part of the dataset that cannot be changed. First, we completed our review of published African literature towards the end of 2018. Data from publications appearing afterwards are not incorporated in the study. We referred to published books, journal articles and field reports, but in some cases, relied on personal communications.
We recognize that a certain research bias exists among African countries, so that spatial coverage in South Africa or Kenya, for example, is considerably better than in Nigeria or Mali. This reflects longer traditions of research as well as different institutional considerations, such as infrastructure and long-term political stability. Another factor is reporting bias on historical and methodological grounds, for example, differences in research traditions over time, variable excavation techniques resulting in disparate recovery of finds, and different analytical goals. Especially in older reports, the presence of ochre is often presented without further detail about its color, weight or count. Fortunately, many of these older collections have been reassessed using current methods. Still, there is no way to confirm that ochre pieces were collected in the past (or for that matter, in the present), or whether fine screening for small pieces of ochre was conducted.
A key physical factor is the preservation bias observed between cave and open-air sites. Not only does the presence of caves depend upon regional geology, caves often serve as focal points for systematic exploration, contributing to a greater likelihood of documentation. On the other hand, open air sites are often associated with featureless landscapes and may be neglected, unless they happen to be associated with spring deposits or are uncovered through chance activities.
As previously mentioned, bias at the continental scale cannot be ignored. However, it is our contention that the large geographical and chronological scales of this analysis tend to neutralize those differences. The same holds true for ecological bias related to differences in soil types, habitats and vegetation. Nor can we overlook the chronological bias that results from differences in radiometric dating methods, the ages reported and their standard deviations. Finally, we recognize the possibility that MSA people used other colorants made of organic materials besides mineral ochre, which are no longer archaeologically visible.
Results
After running the time slice tool for each of the 13 combinations of time slice and time step, we gave the resulting plots to ten observers (including the authors) to test inter-observer reliability (Fig. S1–S13). Each was asked to pick the times where they recognized inflection points in the ochre data. The observers noted two distinct chronological breaks, resulting in three main phases of ochre use, regardless of the combination selected. Based on the results of all ten observers, the mean values of the breaks occurred at 326 ± 14 and 157 ± 14 thousand years (Table S1). The overall similarity in the observers’ results for all 13 combinations suggests that the trend is robust and reproducible. For practicality, we rounded these values to 330 and 160 thousand years to define the breaks in our subsequent analytical units.
The results of the finite mixture distribution (Fig. 5) seem more gradual and less stepped when compared to those of the time slice tool analysis. The distinct break observed at 330 thousand years with the time slice tool appears less pronounced using the finite mixture distribution, which we attribute to the higher dating uncertainty of the older assemblages. Based on Fig. 6, the majority of older assemblages have age ranges greater than 100 thousand years. Furthermore, we do not see dramatic differences generated by the smoothing procedure, which we applied to make this analysis comparable with the time slice analysis. However, starting between about 160 and 130 thousand years, we begin to see a dramatic increase in ochre use. This varies with the smoothing interval, where larger intervals generate slightly older estimates. All distributions show a maximum around 60–70 thousand years and decrease towards the younger end of this study’s scope, likely a border effect induced through the sampling age boundary at 40 thousand years. The distributions with few or no smoothing intervals (≤ 10 thousand years) are characterized by a higher variability. We see their expression in the minor peaks postdating 120 thousand years, and attribute this to the increasing number of more precise dates (compare Fig. 6).

Results of the finite mixture method applied to the ochre dataset. Multiple smoothing bandwidths between 5000 and 40,000 years as well as an unsmoothed reference distribution illustrate the effect of time averaging (Color figure online)

Dating precision of ochre and lithic assemblages compared to the major orbital cycles affecting African macroclimate. Assemblages with higher age ranges than the respective orbital cycle disallow a correlation with climate proxies. Since older assemblages have large age ranges, they cannot reliably be correlated to climate cycles. A substantial number of assemblages younger than 150,000 years allow correlation with climate effects related to eccentricity or even precession. Scatterplot jitter was applied to the figure to enhance the visibility of overlapping points
Concerning the interaction of ochre use and climatic variation, we note a log–log linear increase of age uncertainty with greater mean age (Fig. 6). Thus, older assemblages often exceed the cycles of the primary driving factors, namely the 20–24,000 year cycle of precession, and even the 100,000 year cycle of eccentricity. For this reason, we cannot reach a conclusion about the period before 150 thousand years with regard to what impact climatic conditions had on the ESH model. For younger assemblages between 150 and 40 thousand years, there are many assemblages dated with a precision appropriate for examining the effect of the orbital driving factors on the ESH model.
Based on the time slice data, we defined the first phase of ochre use from 500 to 330, with the second phase from 330 to 160 thousand years ago. As discussed above, the results from the finite mixture distribution were not fine-grained enough to warrant their use, owing mainly to broad age ranges. However, the finite mixture distribution appears to confirm the time slice data for the last phase from 160 to 40 thousand years ago (compare Fig. 5 and 7).

Plot of the total number of localities with ochre (below, orange/red) and lithics (above, gray/black) over time. Each line denotes different time slices (10, 20, 30 and 40,000 years) using a constant time step of 5000 years, as shown in the legend. In the data analysis we used additional time steps of 10, 15, 20, 30 and 40,000 years which are not included in this plot (see Table 3, Figs. S1–S13). The ochre plot shows a generally level trend starting at 500,000 years, with a step-wise increase occurring at approximately 330,000 years, corresponding to the change from initial to emergent ochre use. Ochre use stays level during the emergent phase until 160,000 years, when it accelerates noticeably, denoting the beginning of the habitual phase. The two ‘bumps’ in the lithics graph at 300,000 and 200,000 years result from the way we estimate the age of undated assemblages. In ROAD, cultural periods are predefined and follow generally accepted archaeological practice. Specifically, the MSA begins at 300,000 years, and several transitional technocomplexes (e.g. Sangoan and Lupemban) begin at 300,000 and end at 200,000 years. The width of the ‘bumps’ varies in accordance with the time slice selected. Note that this issue does not affect the graph of the localities with ochre, because these are more precisely dated (Color figure online)
We also calculated the ratio of ochre versus stone artifacts to establish the frequency of ochre use over time, using the presence of stone artifacts as a minimum baseline to define an archaeological site (compare Tables S2 and S3). Thus all recognized sites have either lithics without ochre, or lithics plus ochre. No sites with ochre but without lithics have been documented so far. Our results show that each phase is followed not only by a stepwise increase in the number of localities with ochre, but also by a general increase in the percentage of sites yielding ochre with respect to stone artifacts (Fig. 8).

Graph based on Table S2 and S3 showing the general increase in the percentage of sites yielding ochre with respect to stone artifacts over the three chronological phases (Color figure online)
We divided the sites with ochre into three classes of intensity for low (n = 1–100), medium (n = 101–1000) and high (n > 1000) piece counts and used spatial information to plot their location for each of the three phases of ochre use (Fig. 9; interactive online map: https://www.roceeh.uni-tuebingen.de/maps/ochre-africa/). We note that count is the most consistently reported parameter in the literature, although weight would be a more informative indicator for the intensity of ochre use.

Geographical distribution of the three phases of ochre use showing ochre sites and the number of ochre pieces per site for each phase. Note that inclusion in a specific phase does not imply that the site yields ochre for the entire duration of the phase. Sites close to each other were displaced from their actual position following the rules of cartographic generalization to make them more recognizable. See interactive map at https://www.roceeh.uni-tuebingen.de/maps/ochre-africa/
Discussion
The Development of Ochre Use Between 500 and 40 Thousand Years Ago
Based on our data analysis we define three phases of increasing ochre use on the African continent during the MSA, which we name as follows: initial, emergent and habitual. The number of sites with ochre, the ratio of sites with versus without ochre, the geographical distribution of ochre assemblages and the intensity of ochre use at sites all increase significantly with each phase. In the following section we discuss important sites, as well as general paleoanthropological and cultural developments.
Initial Phase of Ochre Use
The initial phase of ochre use occurs from 500 to 330 thousand years ago (Fig. 7, Table S1). Ochre is documented at five sites scattered across southern and eastern Africa (Fig. 9, Table S2). Based on a total of 69 sites with lithics (Table S3), ochre is present at 7% of sites (Fig. 8). The number of localities remains stable throughout this period. Ochre is generally present in small quantities at these isolated localities. Still, 74 pieces are reported from member K3 at Kapthurin GnJh-15 (Kenya) (McBrearty & Brooks, 2000, p. 528). The earliest known grinding implements for powder production come from the same site (Tryon & Faith, 2013, p. S244). Forty pieces of pigment (excluding all doubtful assessments) are documented at Kathu Pan 1 (South Africa), five of which bear traces of utilization (Watts et al., 2016, table 1). In the cases of Wonderwerk and Canteen Kopje (both in South Africa), clear traces of anthropogenic modifications are also present (Watts et al., 2016). At one of the sites, Canteen Kopje, transport distance for the material (in this case, sparkling specularite) is estimated to be 170 km (but without geochemical analyses), a surprising distance indeed. It is also worth noting that the ochre pieces from Wonderwerk Cave were found along with manuports in the rearmost, darkest part of the 140 m deep cave, indicating non-utilitarian activities possibly illuminated by firelight (Chazan & Horwitz, 2009; Watts et al., 2016, p. 299). Traces of intentional abrasion and grinding tools represent the earliest reliable evidence for the production of ochre powder in the archaeological record.
Contrary to assertions made by Beaumont and Bednarik (2013), we find the archaeological evidence for ochre use during the Acheulean insufficient. To date, the literature adequately describes only one piece of ochre with possible anthropogenic use wear from a secure Acheulean context, the minute specimen (S29) from Wonderwerk Cave, Excavation 1, Stratum 8a (1.1 cm, 0.4 g; Watts et al., 2016, fig. S3). Other allegedly introduced pieces come from the South African sites of Kathu Pan 1, Stratum 4b (Beaumont & Bednarik, 2013, p. 43) and Duinefontein 2, Horizon 3 (Cruz-Uribe et al., 2003, p. 565); Melka Kunture, Garba 1, Layer B in Ethiopia (Chavaillon & Berthelet, 2004, p. 61); and the much older site of Upper Bed II at Olduvai in Tanzania (Leakey, 1958, p. 1100). The evidence suggests that these examples are either not demonstrably of anthropogenic origin, represent intrusions from younger layers, or have not yet been adequately examined and published (cf. Chase, 2006, p. 156; Harrod, 2014, p. 147; Oakley, 1981, p. 207; Watts et al., 2016, p. 299).
During this phase we observe cultural changes as Acheulean technologies in Africa are gradually replaced by transitional industries such as the Fauresmith in non-tropical regions of southern Africa as well as the Lupemban and the Sangoan in the forested areas of West and Central Africa (Barham & Mitchell, 2008; Herries, 2011; Mercader, 2002). The hominin taxa usually associated with this period are archaic H. sapiens and H. heidelbergensis (Rightmire, 2009; Roksandic et al., 2018; Stringer, 2016), but much debate exists, as few fossil remains are reported.
Emergent Phase of Ochre Use
During the emergent phase from 330 to 160 thousand years ago, ochre use becomes increasingly visible (Fig. 7). Documented at 16 sites (Table S2), ochre use starts to become more integrated into the human behavioral repertoire (Fig. 9). The emergent phase is associated with cultural changes as early MSA lithic technologies appear throughout Africa (Deino et al., 2018; Lombard, 2012; Scerri, 2017)—although in some regions this transition may have started earlier (Johnson & McBrearty, 2010; Wilkins & Chazan, 2012). The distribution of ochre expands to include northeastern Africa, while an increase in the number of sites and assemblages with ochre occurs in eastern and especially southern Africa. Based on 175 sites with lithics (Table S3), ochre is present at 9% of them (Fig. 8). Similar to the initial phase, the number of localities with ochre remains relatively stable throughout this period, with a slight increase towards the end of the phase.
The number of sites with significantly larger ochre assemblages also increases. At Twin Rivers in Zambia, more than 400 pieces of ochre were reported (Barham, 1998, 2002; Clark & Brown, 2001), including residues of ochre adhering to a quartzite cobble which Clark and Brown (2001, fig. 20:22) interpret as a grinding implement. Other possible grinding tools with ochre residues come from Sai Island 8-B-11 in Sudan (van Peer et al., 2004). Sai Island is exceptional in other ways too, because it is the only site of the entire MSA where yellow ochre dominates. At Olorgesailie, southern Kenya, an ochre block from Locality G (GOK-1) has two opposing holes of anthropogenic origin, the oldest evidence of a possible attempted perforation (Brooks et al., 2018, p. 93). Another important site belonging to the tail end of this phase is Pinnacle Point 13B. According to a recount of the ochre assemblage by Bernatchez (2012, table 4.52) and the dating provided by Marean et al. (2010, table 1), 64 pieces which could be classified as pigment come from layers that are likely 160,000 years or older. The ochre in these layers is associated with early evidence of shellfishing, bladelet production and possibly heat treatment of lithic artifacts to improve their knapping properties (Brown et al., 2009; Marean, 2010; Marean et al., 2007). These characteristics are associated with an expansion of cultural capacities, often referred to as cognitive complexity or behavioral modernity (e.g., d’Errico & Stringer, 2011; McBrearty & Brooks, 2000).
While there is no direct evidence relating stone artifacts and ochre to a specific hominin taxon, species associated with this period include at least archaic H. sapiens and H. naledi (Berger et al., 2015; Dirks et al., 2017; Hublin et al., 2017). Since the specific cultural attributes of these candidates remain unclear, either one could have produced the ochre remains of the African archaeological record. While we view modern humans as the most likely suspect (e.g., Clark et al., 2003; McDougall et al., 2005; White et al., 2003), in the rapidly changing landscape of African paleoanthropology, we can no longer rule out other possibilities.
Habitual Phase of Ochre Use
Finally, from 160 to 40 thousand years ago we see that ochre use has become habitual, incorporated as a regular, repeated and socially transmitted aspect in most people’s lives on the African continent. Ochre use is well documented at 74 sites across Africa (Table S2), with about two-thirds located in the southern part of the continent (Fig. 9). Based on 239 sites with lithics (Table S3), ochre is present at 31% of sites (Fig. 8). During the habitual phase, ochre becomes the most frequent archaeological find after lithics and faunal remains. In contrast to the stability seen in the previous phases, the number of archaeological sites further increases steadily from 160 until about 60 thousand years ago (Figs. 5, 7, and Figs. S1–S13).
During the phase of habitual ochre use new MSA industries appear all over the continent. These include the Aterian in northwestern Africa (Bouzouggar & Barton, 2012; Dibble et al., 2013), the Still Bay and Howiesons Poort in southern Africa (Henshilwood, 2012) and innovative MSA technologies in eastern Africa (Lombard, 2012; Tryon & Faith, 2013). The species commonly associated with this time period is H. sapiens, although informative fossils from the African continent remain rare (Stringer, 2016).
Six sites yield ochre assemblages containing more than 1000 pieces and weighing a total of several kilograms. Ordered by weight, the first of these is Porc-Epic (Ethiopia) with more than 4500 ochre finds (> 40 kg) (Rosso et al., 2017). The rest come from South Africa and include Sibudu Cave with c. 5500 ochre pieces > 8 mm collected by Wadley’s team until 2011 (> 15 kg) (Hodgskiss, 2013a, 2013b); Blombos Cave with more than 1500 pieces > 10 mm (5.6 kg) (Henshilwood et al., 2001b, 2009); Klein Kliphuis with c. 3500 pieces (4 kg) (Mackay, 2011, 2016 personal communication); Hollow Rock Shelter with more than 1000 pieces (1.3 kg) (Watts, 1998, site app. 2.11.1–2); and Howiesons Poort with more than 3000 ochre finds (weight unknown: Deacon, 1995; Table 1; Watts, 1998, app. 7p). Two further South African sites report more than 1000 ochre pieces, but we excluded these from the large class (> 1000 pieces) for the following reasons. While Watts (1998, app. 7 h) counted more than 1700 ochre finds (3 kg) from Umhlatuzana, most come from the transitional MSA/LSA and LSA deposits (Layers 10–18) and are thus younger than the range of our study. As a result, we count 722 pieces coming from the MSA and include this site in the medium class (n = 101–1000) on the map of habitual ochre use (Fig. 9). In addition, Watts (2010) reports 1032 potential ochre pieces from Pinnacle Point 13B weighing approximately 2 kg, but categorizes only 380 as pigment (1.08 kg). In Bernatchez’s (2012) re-counting of the collection, she identified 637 pieces (1.71 kg) as ochre. By excluding all pieces that she classified as ‘doubtful’ or ‘not pigment’, as well as those left unspecified or coming from an uncertain stratigraphic context, 356 pieces remain in the habitual phase. Regardless of which analysis is correct, Pinnacle Point 13B belongs in the medium class.
The overall abundance of ochre during the habitual phase is remarkable. It indicates two major regions where ochre use predominates, in southern and eastern Africa, and two smaller clusters visible in northwestern and northeastern Africa (Fig. 9). The lack of ochre finds in central and western Africa is striking, although not surprising, given that Paleolithic research there is still in its infancy due to issues of research priority, logistics, infrastructure, inaccessibility, political instability, and poor preservation in tropical regions (Mercader, 2002; Scerri, 2018). During the habitual phase, the highest concentrations of ochre are reported from sites in southern Africa. This spatial pattern suggests a center of ritual activity associated with greater population densities along the southern coast (cf. Marean, 2014; Will et al., 2016).
The habitual phase of ochre use is further characterized by several exceptional findings that allow us detailed glimpses into the varied ways in which hunter-gatherers of the African MSA utilized ochre. These include combining ochre with other materials, ochre pieces with abstract engravings, red pigment residues on personal ornaments, and ochre associated with burials and other ritual settings.
In 2008 excavators at Blombos Cave identified an ‘ochre-processing workshop’ within a 100,000-year-old MSA layer containing two abalone shells, bone, charcoal, grindstones and hammerstones, as well as ochre residues adhering to those artifacts. People used these toolkits to produce and store a liquefied, ochre-rich mixture (Henshilwood et al., 2011). Similarly, residue on a Late MSA stone flake from a 49,000-year-old layer at Sibudu Cave contained a composite of powdered ochre mixed with milk as a binder, which Villa et al. (2015) attribute to the killing of a lactating wild bovid.
Several pieces of ochre from Blombos Cave (Henshilwood et al., 2009), Klasies River (d’Errico et al., 2012), Klein Kliphuis (Mackay & Welz, 2008) and Pinnacle Point 13B (Watts, 2010) exhibit geometrical or quasi-geometrical engravings associated with the Still Bay and Howiesons Poort. These are comparable to deliberate engravings of abstract designs on stone pebbles (Anderson, 2012, pp. 189–192; Henshilwood & d’Errico, 2011b, pp. 77–78), animal bone (d’Errico & Henshilwood, 2007; d’Errico et al., 2001) and ostrich eggshells from southern Africa (Assefa et al., 2018; Henshilwood et al., 2014; Texier et al., 2010, 2013). Recently a crosshatched pattern likely drawn with an ochre crayon on a ground silcrete flake was reported from a 73,000-year-old Still Bay layer at Blombos (Henshilwood et al., 2018). The pattern shows remarkable similarities to engraved ochre pieces from the same site. Thus, the habitual phase correlates with the emergence of the earliest engraving and painting tradition on the African continent, although only a limited degree of standardization is recognizable—a crucial prerequisite for structured symbolic systems (Kuhn & Stiner, 2007; Pinker, 1994, pp. 83–125; Tylén et al., 2020).
Red ochre was also discovered as residues attached to the oldest personal ornaments known from the African continent. The Moroccan sites Ifri n’Ammar, Grotte des Pigeons, Rhafas and Grotte des Contrebandiers yielded nearly thirty perforated shells with ochre residues associated with the Aterian technocomplex, stemming from small marine gastropods primarily of the subfamily Nassariinae (d’Errico et al., 2009; Steele et al., 2019). Recently, the oldest perforated shell beads yet recovered, from Bizmoune Cave (Morocco), were reported to be about 142,000 years old (Sehasseh et al., 2021). While red ochre residues are also found on some shells from Bizmoune Cave, we did not include them in our meta-analysis because of our cut-off date of 2018. Similar to the northwestern African finds, several southern African sites yielded shell beads. Four perforated Nassarius shells with ochre residues come from a Still Bay layer at Blombos Cave (Vanhaeren et al., 2013). In addition, one Afrolittorina shell from a hearth associated with the Still Bay technocomplex in RGS layer at Sibudu Cave showed residues of red pigment covering its surface (d’Errico et al., 2008). At Border Cave, a perforated Conus shell with red residue was found associated with the burial of a four- to six-month-old infant, dated to about 74,000 years (d’Errico & Backwell, 2016). Furthermore, the bones of the child exhibit an unusual reddish stain (de Villiers, 1973, p. 239) which might indicate the use of ochre as part of a funerary ritual. However, the geochemical origin of the stain has not yet been investigated. Finally, the context of the ochre assemblage from the aforementioned Rhino Cave in the Tsodilo Hills of Botswana is quite unique. Here ochre was used in connection with the repeated ritualistic destruction of valuable and colorful tools made from non-local materials and the successive engraving of a massive quartzite outcrop (Coulson et al., 2011).
Direct archaeological evidence suggests that during the habitual phase some of the ochre was also used for functional purposes, for example, as an ingredient in compound adhesives (Gibson et al., 2004; Lombard, 2006a, b, 2007, 2008; Wadley et al., 2004), soft knapping tools and abrasion (Soriano et al., 2009), and possibly hide tanning (Wadley & Langejans, 2014). Yet it is unlikely that such functional uses of ochre are responsible for the main part of the archaeological record, especially considering that the aforementioned large assemblages of ochre exhibit deliberate color choice.
Implications for the Female Cosmetic Coalitions (FCC) Hypothesis
Our data do not contradict the first chronological prediction made by the FCC hypothesis, which stated that initial ochre use should not pre-date the encephalization associated with H. heidelbergensis between approximately 800,000 and 500,000 years ago. However, it is important to point out that the status of H. heidelbergensis as a species, its defining criteria and its cladistic position within the evolutionary tree are highly debated topics (Roksandic et al., 2018; Stringer, 2016).
Based partly on our findings and partly on new paleontological data, we point out several problems with the second chronological claim made by the FCC hypothesis that habitual ochre use should arise abruptly with the speciation of H. sapiens about 200,000 to 150,000 years ago:
-
1.
Recent genetic (Schlebusch et al., 2017) and fossil (Hublin et al., 2017) evidence has pushed the first appearance of the H. sapiens clade to an earlier date than previously recognized, now approximately 315,000 years ago. Specimens from Jebel Irhoud, Morocco are now considered to be the oldest reliably dated examples of H. sapiens (Richter et al., 2017), with modern cranial volumes of 1467 ± 6 and 1375 ± 6 cm3 (Neubauer et al., 2018). While the gracile face exhibits modern features, the braincase is described as having an archaic, elongated shape that is less globular than modern H. sapiens (Hublin et al., 2017; Neubauer et al., 2018). As far as we know, the globular shape of the brain of anatomically modern humans evolved only during the past 100,000 years (Neubauer et al., 2018). Still, these details would have made no significant difference with regard to the challenges of childbirth and child rearing. What is now clearer is that the evolution of H. sapiens was a gradual, complex and mosaic phenomenon, not a linear and abrupt event. Expansions, contractions, patterns of temporary population isolation and aggregation as well as interbreeding of morphologically different populations occurred on a continental scale (Scerri et al., 2018).
-
2.
Our habitual phase of ochre use starts at 160,000 and peaks between 80,000 and 60,000 years ago. Considering this information along with the evidence for an earlier emergence of the H. sapiens clade, we conclude that it is not possible to correlate the emergence of habitual ochre use with the speciation of H. sapiens.
Nonetheless, these issues do not repudiate some of the basic assumptions of the FCC hypothesis. We agree that encephalization and the accompanying problems of cooperative child rearing, sexual selection and costly signaling as well as perceptional and psychological biases towards the color red all played important roles in the evolution of collective ritual. But our data question the chronological predictions made by the FCC with regard to the emergence of habitual ochre use.
Implications for the Ecological Stress Hypothesis (ESH)
We confirm that changes in ochre use occur, but these seem to follow the general distribution of stone artifacts (Fig. 10d). We recognize a subtle increase in ochre use around 330 thousand years ago at the introduction of the emergent phase, which is contemporaneous with globally warmer conditions (Fig. 10f) during MIS 9 and rather arid conditions in Eastern Africa (Fig. 10e). However, the high age uncertainties of the assemblages in question make it difficult to draw conclusions about causal relationships. The habitual phase postdating 160 thousand years tends to have lower age uncertainties and is therefore more suitable for comparisons with climatic variables. For the main regions of Africa, we make the following observations:
-
Northern Africa The ochre distribution (Fig. 10a) is bimodal and has its highest peak at roughly 110 thousand years, which is more pronounced than the peak in lithic assemblages from the same region. This time range corresponds with one of the Green Sahara Periods, which created favorable conditions for human occupation around 100–110 thousand years ago (Ehrmann et al., 2017; Larrasoaña et al., 2013; Tjallingii et al., 2008). Furthermore, Jones et al., (2016) mention the onset of symbolic behavior, including ochre use, with the MIS 5e climatic optimum. Consequently, we suggest that the increase in ochre coincides with an ecological window of opportunity for early humans, rather than a period of ecological stress. We emphasize, however, that this signal is weak due to the generally low find density of this region.
-
Central and Eastern Africa We recognize an increase of ochre use correlated with more arid conditions (Fig. 10e) in East Africa and proposed mega-droughts between 135 and 75 thousand years ago (Scholz et al., 2007). However, the ochre distribution follows the trend of the lithic assemblages through the time span considered, which we interpret as an increase in population density affecting lithic and ochre abundances alike.
-
Southern Africa The highest abundance of both ochre and lithic assemblages is found in this region, representing 75% of all ochre assemblages from the continent. Throughout the emergent phase, ochre and lithic assemblages are equally distributed. During the habitual phase, the majority of ochre assemblages are related to relevant technocomplexes as explained in the Sect. 3 Habitual Phase of Ochre Use.

Regional occurrence of ochre and lithics in a northern Africa; b central and eastern Africa; c southern Africa; and d the overall distribution, each compared to climate proxies: e the Pan-African Hydroclimate represented by the first Principal Component (PC1) of a climate reconstruction by Kaboth-Bahr et al. (2021), which indicates alternating humid and arid conditions in eastern and western Africa; f the benthic δ18O record from Lisiecki and Raymo (2005), which serves as a proxy for the global temperature and sea ice volume; g macroscale climate drivers are represented by eccentricity and precession and were calculated with the equations of Berger (1978). The mixture distributions allow us to compare the development of ochre and lithic assemblages (n) over time in their respective region. However, because their abundances vary, comparisons between regions and classes (ochre, lithics) should be viewed with caution
We attribute the major peak of ochre use during the habitual phase (Fig. 5 and 7) to (i) narrower dating ranges, (ii) a larger proportion of assemblages with ochre (Fig. 8) and (iii) the presence of more archaeological sites in general (Table S3), the latter two possibly being related to an overall increase in population, especially in southern Africa. We would argue that this dynamic was mainly created by a positive feedback loop between collective rituals—with their positive effect on the size, density and stability of social networks (see below)—and growing populations.
Furthermore, the decreasing values in our diagrams between 60,000 and 40,000 years at the end of the timespan analyzed is likely an artifact of our methods, as we did not examine sites younger than 40,000 years. Consequently, there are no overlapping time slices for younger assemblages. It is possible that time averaging, with its dissolution of fine-grained details, smoothed away subtle fluctuations associated with rapid climate change. However, we also see no evidence for long-term fluctuation associated with global climatic shifts caused by the major drivers of African climate in the dataset.
Overall, we would argue that our data do not demonstrate an observable influence caused by stress-inducing climatic fluctuations. Consequently, we are unable to confirm the ESH proposed by Rossano (2015), who suggested that ecological stress contributed to the change observed in ochre distributions over time. Moreover, we recall that ochre is found at all latitudes and altitudes in radically different environments from the northern to the southern coasts of Africa across a distance of more than 8000 km.
Implications for Eight-Grade Model for The Evolution and Expansion of Cultural Capacities (EECC)
Earlier we discussed the role of ochre within the EECC model. The model estimates that the sixth grade, composite cultural capacity, would have been achieved between 500,000 and 200,000 years ago, depending on the interpretation of the laminar stone tool technology from Kathu Pan 1, South Africa (Wilkins & Chazan, 2012; Wilkins et al., 2012) and the lower Kapthurin Formation, Kenya (Johnson & McBrearty, 2010). If it could be clearly demonstrated that these artifacts were hafted to wooden spears, these occurrences would push the emergence of composite cultural capacity back to around 500,000 years ago (Haidle et al., 2015, p. 58). Our initial phase of ochre use also begins 500,000 years ago with the earliest reliable evidence from the African archaeological record (Watts et al., 2016). If ochre powder was mixed with a binder such as water, animal fat, milk or blood, regardless of whether for a functional or ritualistic purpose, then the initial phase of ochre use would fit well into the sixth grade of the EECC model. Lyn Wadley showed in her experimental analysis that complex cognition based on advanced working memory capacities is necessary to produce compound-adhesives for hafting tools (Wadley et al., 2009; Wadley, 2010c). However, the oldest direct evidence for the use of binders is currently about 100,000 (Henshilwood et al., 2011), and for compound adhesives with ochre as an ingredient about 70,000 years old (Lombard, 2006b; Wadley et al., 2009; Wojcieszak & Wadley, 2018).
An additional contribution of our findings concerns the timing of notional cultural capacity, the eighth grade within the EECC model. We interpret the emergence of habitual ochre use as an indicator of regular collective rituals. It is plausible to assume that these performances were already imbued in symbolism. During the habitual phase, ochre is associated with personal ornamentation (d’Errico et al., 2009; Vanhaeren et al., 2013), mortuary behavior (d’Errico & Backwell, 2016) and the ritualistic destruction and burial of hundreds of stone points (Coulson et al., 2011). Furthermore, as discussed above, collective ritual could have been a crucial prerequisite for the emergence of symbolic communication by solving the cooperative dilemma. Thus, the emergence of habitual ochre use 160,000 years ago, which we interpret as the materialization of regular collective rituals, might be one additional indication of the origin of symbolically mediated behavior around this time. This would push back the beginning of notional cultural capacity from the Upper Paleolithic of Eurasia into the early MSA of Africa (Haidle et al., 2015, p. 59).
Implications for the Development of Ritual Behavior and the Demographic Expansion of Homo sapiens During the MSA
If we interpret ochre finds mainly as a material manifestation of past ritual behavior, what does it mean for the evolution of ritual? What implications for the demographic expansion of H. sapiens can we deduce? According to our data, the spread of ritual activity involving ochre starts about 500,000 years ago slowly and gradually. About 330,000 years ago the development moderately accelerates; starting at 160,000 years ago, further acceleration is observable, as some kind of self-reinforcing feedback loops seem to kick in. Ritual then becomes a habitual element of the behavioral repertoire of our species.
Our findings might be important for the cultural evolution of cooperation and the emergence of H. sapiens as an ultra-social species (Tomasello, 2014). What anthropologists already observed a century ago (e.g., Durkheim, 1912; Malinowski, 1948; Radcliffe-Brown, 1922) is now empirically established by a plethora of psychological experiments in the laboratory as well as in the field. Ritual is a powerful social glue, creating ties that bind cultural groups together beyond the mechanisms of kinship, physical grooming and reciprocity (Hill et al., 2014; Hobson et al., 2018; Whitehouse & Lanman, 2014). Thanks to substantial progress over the last 20 years in the fields of cognitive science of religion and evolutionary psychology, the adaptive function of ritual for group cohesion, cooperation, prosociality and the transmission of cultural norms is well established within an evolutionary framework (Bulbulia & Sosis, 2011; Henrich, 2016, pp. 140–165; Legare & Nielsen, 2020; Legare & Watson-Jones, 2016; Rossano, 2016; Sosis & Alcorta, 2003; Watson-Jones & Legare, 2016; Whitehouse, 2012, 2013a). Ritual is a ‘psychologically prepared and culturally inherited behavioural hallmark of our species’ (Legare & Nielsen, 2020), which exploits various aspects of our evolved psychology, some of which are mentioned above. Regarding the demographic expansion of our species and the acceleration of cultural evolution during the second half of the MSA, the following psychologically active ingredients of ritual are crucial:
-
Costly signaling Substantial time, energy and resources, as well as risk, pain, and other personal sacrifices, are devoted to an attention-grabbing collective ritual display to demonstrate a group commitment that cannot be faked by free-riders, thus stabilizing group cohesion and cooperative behavior (Bulbulia & Sosis, 2011; Henrich, 2009; Irons, 2001; Sosis, 2003; Sosis & Alcorta, 2003).
-
Synchronization Repetitive vocal expressions (e.g., singing, chanting), body movements (e.g., dancing, jumping) and emotional states (mirroring) are often synchronized in collective rituals. Through synchronization the distinction between the group and the self is attenuated and thus the feeling of oneness, group affiliation and social bonding enhanced (Jackson et al., 2018; Konvalinka et al., 2011; Lang et al., 2017; Mogan et al., 2017; Tarr et al., 2016; Wiltermuth & Heath, 2009). In other words, ‘the group that chants and dances together hunts well together’ (von Zimmermann & Richardson, 2016, p. 1).
-
High emotional arousal In many rituals, multisensory stimulation, combined with behavioral synchronization and extreme physical exhaustion, leads to the release of endogenous opioids and monoamine neurotransmitters which induce euphoria and a positive feeling towards other group members (Dunbar et al., 2012; Fischer et al., 2014; Tarr et al., 2015; Xygalatas, 2012, pp. 167-184).
-
Goal demotion and overimitation Ritual actions are causally opaque and serve no direct instrumental purpose but will nevertheless be imitated exactly by other group members. They exploit our evolved proclivity for overimitation (copying causally irrelevant actions from others despite the presence of clear causal information) and implicit interpretation of such actions as highly normative (Kapitány & Nielsen, 2015, 2016, 2019; Keupp et al., 2013; Nielsen & Tomaselli, 2010; Nielsen et al., 2015, 2018; Over & Carpenter, 2012). This fosters the transmission of cultural norms, symbols and shared fictions (Humphrey & Laidlaw, 1994; Legare & Souza, 2012; Schjoedt et al., 2013; Whitehouse, 2012).
-
Transmission of cultural content Cultural norms, symbols and stories are shared, transmitted and internalized through the dramatization and continual repetition, rhythmicity, (over-)imitation, and synchronization on the basis of trust, feelings of oneness and a shared identity created through ritual action (Rossano, 2012; Whitehouse, 2004, 2012).
-
Creation of group identity Rituals generate group identity, as well as emotional and symbolically marked boundaries between groups (Watson-Jones & Legare, 2016). Consequently, they also foster cognitive and emotional in-group vs. out-group biases, intergroup competition and tribalism (Hobson & Inzlicht, 2016; Sosis et al., 2007; Wen et al., 2016; Whitehouse, 2012, 2013b), which in turn might have further increased the tendency towards in-group cooperation (Henrich, 2016, pp. 166–184; Richerson et al., 2016).
The key point here is that with these additional psychological mechanisms—originally starting from the primate base discussed above—ritual became a powerful social institution in the course of human evolution, one which was able to extend the social fabric of early H. sapiens populations.
The long-term development of ochre use beginning 500,000 years ago gradually grew into a habitual behavior starting 160,000 years ago. This pattern can be interpreted as a material manifestation of increasing ritual activity as H. sapiens evolved and demographically expanded throughout the African continent before the species’ permanent and successful dispersal into Eurasia, Australia and the New World. Beside technological innovation, collective ritual could have been an important cultural prerequisite for the successful expansion of our species as the ‘ultra-social animal’ (Tomasello, 2014) across the globe.
Increasing ritual activity may have given expanding populations of H. sapiens a crucial competitive and demographic advantage over other contemporary hominin groups consisting of Neanderthals or Denisovans, for example. Collective rituals could have instilled a variety of beneficial behaviors, including the effective deterrence of free-riders and the successful transmission of cultural norms. Most importantly, these also include the establishment of bigger and more stable cooperation and information networks which transcend narrow kinship boundaries by a large margin through trust, increased group cohesion and shared identity—even in situations when parts of the social network are physically separated from time to time. As mathematical models (Kobayashi & Aoki, 2012; Powell et al., 2009; Shennan, 2001), experimental evidence (Derex & Boyd, 2015; Derex et al., 2013; Muthukrishna et al., 2014) and ethnographic data (Collard et al., 2013; Henrich, 2004; Kline & Boyd, 2010) show, larger, denser and more stable social networks accelerate and intensify cumulative cultural evolution, leading to more (and more varied) technological innovations (cf. Henrich et al., 2016, Appendix). The establishment of trust in ritually constructed extended family groups and fictive kin could have also improved cooperative alloparenting for increasingly dependent and energy-hungry children, with a positive impact on birth rates (Cronk et al., 2019; Shaver et al., 2019, 2020). This may also have contributed to the demographic expansion of our species.
It is noteworthy that, according to our data, long-term changes in ochre use cannot be directly correlated with climatic fluctuations. If this finding is not an artifact of our analytical method, it could indicate that social institutions like collective ritual became more important for demographic and geographical expansions than external ecological factors by themselves during the late evolution of our species. In sum, collective rituals are a powerful social institution that makes use of different aspects of our evolved psychology. Rituals benefit human cooperation, promote social bonding and increase the sharing of information in larger social networks, thus fostering cumulative cultural evolution and demographic expansion. They are part of the ‘secret of our success’ as a species (Henrich, 2016).
The intentional exploitation of blood-red and bright red hues documented in the large assemblages of Blombos Cave (Watts, 2009), Sibudu Cave (Hodgskiss, 2012), Pinnacle Point 13B (Watts, 2010), Pinnacle Point 5–6 (Bernatchez, 2012), Rose Cottage Cave (Hodgskiss & Wadley, 2017), and Porc Epic (Rosso et al., 2017), also shown in an earlier meta-analysis of 11 MSA sites south of the Limpopo River (Watts, 2002), is most likely not a coincidence. As discussed above, the color red is highly attention-grabbing for the human visual perception system. The capacity for trichromatic vision evolved in primates about 35 million years ago (Jacobs, 2015) as a result of the benefits for recognizing ripe fruits in front of a background of green foliage (Regan et al., 2001). Another explanation is to observe the subtle changes in blood flow beneath hairless parts of the skin, which carry relevant information about the emotional state of conspecifics (Changizi et al., 2006). The relationship of the color red to bleeding might also have evolutionary significance because it is associated with wounds caused by predators, fatal accidents or fights with conspecifics, as well as menstruation and fertility.
From an evolutionary perspective, the perception of the color red may thus tap into primordial mental systems associated with the recognition of blood and ripe fruit. These two things are tightly associated with the primary driving factors of natural and sexual selection in terms of survival (food acquisition, danger, pain, death), mating (menstruation, sexual arousal, health) and social group living (emotional arousal, aggression, dominance). While the FCC hypothesis offers an explanation of why the color red was used first as a signaling device in female ritual displays (sham menstruation), we remain cautious and rather orient ourselves towards the literature of color psychology and primatology, independent of the reasoning of any particular model. Based on this literature, it is clear not only that red stimuli exert perceptual biases and psychological effects on male observers of female individuals in mating contexts, but that the effects of red stimuli are also measurable in other domains. The reason that these psychological effects evolved may well be that in all vertebrates, blood appears red due to the iron-containing protein complex hemoglobin. Red blood carries signaling qualities not only in the context of menstruation, but also through reddened skin areas in the context of dominance and aggression, especially in males (Mehta et al., 2008; Setchell, 2015; Stephen et al., 2012). Red also portrays sexual arousal (Elliot et al., 2008; Pazda & Greitemeyer, 2015), marks physical health and attractiveness in both women and men (Stephen & Perrett, 2015; Stephen et al., 2009a, b; 2012), and signals potentially life-threatening open wounds (Ritz et al., 2010).
In considering this body of literature, we cannot link the signaling qualities of the color red exclusively to menstruation and mating. Furthermore, from the perspective of evolutionary psychology, it is not unusual that relatively young cultural innovations tend to exploit ancient adaptations that evolved millions of years ago. Nor do these cultural inventions necessarily increase the genetic fitness of the respective user. Modern examples include industrially produced candy and internet pornography, which respectively exploit our evolved preferences for high caloric food and visual sexual stimuli (Huppin & Malamuth, 2017; Rozin & Todd, 2015). We propose that a similar ‘hijacking’ process of evolved perceptual biases, aesthetic preferences and emotional responses took place when red pigments were used in early rituals. Hence, it is no wonder that red pigments were used as early collective rituals emerged. Red exploits deep-rooted psychological biases in hominins to create highly attention-grabbing social performances. That early ritual ochre use was primarily about capturing attention and stimulating the senses is also indicated by the fact that, in some cases, a sparkling quality might have played an extra role when ancient people used a type of ochre known as specularite to produce powder with a metallic luster (Coulson et al., 2011; Watts et al., 2016).
It seems likely that archaic hominins initially experimented with red ochre because of its attention-grabbing quality in occasional but emotionally charged ritualized displays, without necessarily attaching a sophisticated symbolic meaning to it. But during the emergent phase of ochre use, the ritualized behavior became increasingly regular and organized. The artificially amplified signals were used ever more strategically as our social skills became more complex. According to the current state of archaeological research, ochre use becomes a habitual cultural practice starting 160,000 years ago. Now that ochre is clearly associated with personal ornaments, geometrical engravings, drawings, and mortuary behavior, we can assume a deep embedding of regular collective rituals in full-fledged symbolism.
Conclusion
In review, the results of our study show the following developments of ochre use in Africa:
-
1.
The number of sites with ochre increases together with the total number of archaeological sites over the period from 500,000 to 40,000 years ago.
-
2.
Three phases of ochre use were identified, which we named initial, emergent and habitual.
-
3.
Each phase witnesses an increase in the ratio of sites with versus without ochre, from 7% during the initial phase to 31% during the habitual phase.
-
4.
An increase in the prevalence of ochre occurs during the emergent phase, starting about 330,000 years ago.
-
5.
As ochre use intensifies during the habitual phase, this behavior spreads geographically across most of the African continent, with the exception of central and western areas.
-
6.
We identify centers of ochre use in southern and eastern Africa, with smaller centers in northwestern and northeastern Africa.
-
7.
Southern Africa seems to represent a major focal point for the proliferation of ochre use between 160,000 and 40,000 years ago—although this picture might be partially affected by the strong research history of this region.
Irrespective of the ochre finds, the total number of archaeological sites with lithics also increases substantially through the period studied (Fig. 7 and Figs. S1–S13, Table S3). This trend indicates that despite fluctuations within semi-isolated populations (Scerri et al., 2018) a demographic expansion occurred from the perspective of the longue durée, at least in southern, eastern and to a lesser extent also in northern Africa. Beside the increase in site numbers during the MSA, an overall demographic expansion is indicated by other archaeological observations, even though this development is interrupted and even reversed from time to time. Such evidence includes the occupation of new habitats hitherto not used during the Acheulean period, the intensified use of a wider variety of natural resources and an increase in transport distances of raw material, indicating larger exchange networks (Brooks et al., 2018; McBrearty & Brooks, 2000; Nash et al., 2013, 2016).
Collective rituals enabled the expansion of social networks and increased the number and reliability of internal connections in those networks. Thus, they were crucial for the transmission and cumulative evolution of cultural knowledge (Henrich, 2016, p. 224). Multiple lines of evidence suggest that red ochre was in large part used for ritual purposes during the MSA. If this is true, then the long-term pattern of ochre use indicates that collective ritual first evolved slowly and gradually, starting about 500,000 years ago, but later accelerated to become a habitual cultural phenomenon between 160,000 and 40,000 years ago spanning most of the African continent. This new and powerful social institution fostered group cohesion, prosociality, alloparenting and stable information flow in larger and denser social networks. It might have allowed ancient humans to expand into new biomes, cope with different climatic constraints and eventually expand demographically.
Our results suggest that as ritual activity intensified, the population density increased. Strikingly, the peak of the habitual phase of ochre use 80,000 to 60,000 years ago coincides with the last large dispersal of H. sapiens during the Pleistocene across Eurasia and into Australia (Bolus, 2015; Clarkson et al., 2017; Liu et al., 2015), although one has to admit that many uncertainties about the timing and character of this dispersal remain (Groucutt et al., 2015; Harvati et al., 2019; Hershkovitz et al., 2018). Thus, red ochre and ritual might have played an important role in the long-term development of demographic expansion, as material culture, human action and cognition are tightly intertwined in a process of culture-gene coevolution within our lineage (Dennett, 2017; Henrich, 2016).
Our arguments for ritual use of red ochre do not exclude functional applications of the material. Simultaneously ochre may also have been a crucial ingredient in various technological innovations such as sunscreen (Rifkin et al., 2015a, 2015b), insect repellent (Rifkin, 2015), hide tanning (Rifkin, 2011) and compound adhesives (Wadley et al., 2009; Zipkin et al., 2014). These functional applications of the material may also have helped expand the cultural capacities of H. sapiens, which in turn aided their dispersal across the globe. Furthermore, we understand that ochre (and its association with personal ornaments, engravings and drawings) is not the only archaeological material that allows us to draw conclusions about ritual evolution. Other evidence for example about mortuary activity (e.g., Pettitt, 2011a) or ritualized behavior in relation to stone tools (e.g., Coulson et al., 2011) has to be taken into account as well, if we want to paint a more complete picture. Our three phases of ochre use may provide a basic chronological framework for further research on the timing and geography of early ritual evolution on the African continent.
Suggestions for Further Research
Besides new excavations, future research should concentrate on the reanalysis and adequate presentation of the materials from older campaigns. For example, we excluded 18 sites from our analyses, mainly because of insufficient information (Table 2). During data collection we often faced the problem that ochre finds were described using different details and parameters. It would be beneficial if researchers agreed upon standardized documentation and terminologies, which more recent studies have achieved. The presentation of ochre finds should include piece counts, weights and evidence for anthropogenic modifications (e.g., grinding, scoring, scraping, engraving) presented separately for each stratigraphic layer. In-depth analyses should include the assessment of deliberate color choice and the identification of possible geological sources to infer transport distances, preferably using geochemical methods (e.g., Dayet et al., 2016).
Following the excellent examples of Bernatchez (2012), Dayet (2012), Hodgskiss (2013b), Rifkin (2012a), Velliky (2018) and Watts (1998), young researchers should be encouraged to analyze ochre assemblages systematically. Detailed scientific investigations of ochre have the potential to yield important findings about the evolution of ritual behavior. Another possible direction of research could compare African and European use of ochre (and other types of pigments) within the timeframe of this study so we can better understand differences and similarities between the behavior of Neanderthals and early modern humans and test predictions of various models in this regard.
References
Ambrose, S. H. (1998). Chronology of the Later Stone Age and food production in East Africa. Journal of Archaeological Science, 25, 377–392.
Anderson, H. (2012). Crossing the line: The early expression of pattern in Middle Stone Age Africa. Journal of World Prehistory, 25, 183–204.
Anderson, J. R. (2011). A primatological perspective on death. American Journal of Primatology, 73(5), 410–414.
Anderson, J. R., Gillies, A., & Lock, L. C. (2010). Pan thanatology. Current Biology, 20(8), R349–R351.
Arcadi, A. C., Robert, D., & Boesch, C. (1998). Buttress drumming by wild chimpanzees: Temporal patterning, phrase integration into loud calls, and preliminary evidence for individual distinctiveness. Primates, 39(4), 505–518.
Armstrong, A. L. (1931). Rhodesian archaeological expedition (1929): Excavations in Bambata Cave and researches on prehistoric sites in Southern Rhodesia. The Journal of the Royal Anthropological Institute of Great Britain and Ireland, 61, 239–276.
Assefa, Z., Asrat, A., Hovers, E., Lam, Y., Pearson, O., & Pleurdeau, D. (2018). Engraved ostrich eggshell from the Middle Stone Age contexts of Goda Buticha, Ethiopia. Journal of Archaeological Science Reports, 17, 723–729.
Atkinson, Q. D., & Whitehouse, H. (2011). The cultural morphospace of ritual form. Evolution and Human Behavior, 32(1) 50–62, S1090513810001029. https://doi.org/10.1016/j.evolhumbehav.2010.09.002.
Avery, G., Cruz-Uribe, K., Goldberg, P., Grine, F. E., Klein, R. G., Lenardi, M. J., Marean, C. W., Rink, W. J., Schwarcz, H. P., Thackeray, A. I., & Wilson, M. L. (1997). The 1992–1993 excavations at the Die Kelders Middle and Later Stone Age cave site South Africa. ournal of Field Archaeology, 24(3), 263–291.
Avery, G., Halkett, D., Orton, J., Steele, T. E., Tusenius, M., & Klein, R. (2008). The Ysterfontein 1 Middle Stone Age rock shelter and the evolution of coastal foraging. South African Archaeological Society Goodwin Series, 10, 66–89.
Backwell, L. R., McCarthy, T. S., Wadley, L., Henderson, Z., Steininger, C. M., deKlerk, B., Barré, M., Lamothe, M., Chase, B. M., Woodborne, S., Susino, G. J., Bamford, M. K., Sievers, C., Brink, J. S., Rossouw, L., Pollarolo, L., Trower, G., Scott, L., & d’Errico, F. (2014). Multiproxy record of late quaternary climate change and Middle Stone Age human occupation at Wonderkrater, South Africa. Quaternary Science Reviews, 99(6), 42–59. https://doi.org/10.1016/j.quascirev.2014.06.017
Barham, L. S. (1995). The Mumbwa Caves project, Zambia, 1993–94. Nyame Akuma, 43, 66–72.
Barham, L. S. (1998). Possible early pigment use in south-central Africa. Current Anthropology, 39(5), 703–710.
Barham, L. S. (2002). Systematic pigment use in the Middle Pleistocene of South-Central Africa. Current Anthropology, 43(1), 181–190.
Barham, L. S., & Mitchell, P. D. (2008). The first Africans: African archaeology from the earliest toolmakers to most recent foragers. Cambridge University Press.
Barham, L. S., Pinto, A. C., & Andrews, P. (2000a). The Mumbwa Caves behavioural record. In L. S. Barham (Ed.), The Middle Stone Age of Zambia, South Central Africa (pp. 81–148). Western Academic & Specialist Press.
Barham, L. S., Simms, M. J., Gilmour, M., & Debenhan, N. (2000b). Twin Rivers, excavation and behavioural record. In L. S. Barham (Ed.), The Middle Stone Age of Zambia, South Central Africa (pp. 165–216). Western Academic & Specialist Press.
Barham, L. S., Tooth, S., Duller, G. A. T., Plater, A. J., & Turner, S. (2015). Excavations at site C north, Kalambo Falls, Zambia: New insights into the mode 2/3 transition in South-Central Africa. Journal of African Archaeology, 13(2), 10.
Basedow, H. (1925). The Australian aboriginal. F. W. Preece.
Baxter, R. A. (2017). Mixture model. In C. Sammut & G. I. Webb (Eds.), Encyclopedia of machine learning and data mining (2nd ed., pp. 841–844). Springer. https://doi.org/10.1007/978-1-4899-7687-1
Beall, A. T., & Tracy, J. L. (2013). Women are more likely to wear red or pink at peak fertility. Psychological Science, 24(9), 1837–1841.
Beaumont, P. B. (1973). The ancient pigment mines of southern Africa. South African Journal of Science, 69, 140–146.
Beaumont, P. B. (1990a). Nooitgedacht 2 and Roseberry Plain 1. In P. B. Beaumont & D. Morris, (Eds.), Guide to archaeological sites in the Northern Cape. Prepared for the Southern African association of archaeologists post-conference excursion, 9th–13th September 1990a (pp. 4–6).
Beaumont, P. B. (1990b). Kathu Pan. In P. B. Beaumont & D. Morris (Eds.), Guide to archaeological sites in the Northern Cape. Prepared for the Southern African association of archaeologists post-conference excursion, 9th–13th September 1990b (pp. 75–100).
Beaumont, P. B. (2004a). Canteen Kopje. In D. Morris & P. B. Beaumont (Eds.), Archaeology in the Northern Cape: Some key sites. Paper prepared for the Southern African Association of Archaeologists conference (pp. 26–30). McGregor Museum.
Beaumont, P. B. (2004b). Kathu Pan and Kathu Townlands/Uitkoms. In D. Morris & P. B. Beaumont (Eds.), Archaeology in the Northern Cape: Some key sites. Paper prepared for the Southern African Association of Archaeologists conference (pp. 50–53). McGregor Museum.
Beaumont, P. B. (2004c). Pniel 6 (The Bend). In D. Morris & P. B. Beaumont (Eds.), Archaeology in the Northern Cape: Some key sites. Paper prepared for the Southern African Association of Archaeologists conference (pp. 61–63). McGregor Museum.
Beaumont, P. B., & Bednarik, R. G. (2013). Tracing the emergence of palaeoart in sub-Saharan Africa. Rock Art Research, 30(1), 33–54.
Beaumont, P. B., & Richardt, F. (2004). On the margins of Karee Vloer: Rooidam 1 & 2 and Biesiesput. In D. Morris & P. B. Beaumont (Eds.), Archaeology in the Northern Cape: Some key sites. Paper prepared for the Southern African Association of Archaeologists conference (pp. 22–25). McGregor Museum.
Beckwith, C., & Fisher, A. (1999). African ceremonies. Abrams.
Beckwith, C., & Fisher, A. (2012). Painted bodies: African body painting, tattoos and scarification. Rizzoli.
Bednarik, R. G. (1992). Palaeoart and archaeological myths. Cambridge Archaeological Journal, 2(1), 27–57.
Belier, W. (1995). Ritual and identity in Aboriginal Australia. In J. Platvoet & K. van der Toorn (Eds.), Studies in the history of religions: Pluralism and identity: Studies in ritual behavior (Vol. 67, pp. 69–90). Brill.
Berger, A. L. (1978). Long-term variations of daily insolation and quaternary climatic changes. Journal of the Atmospheric Sciences, 35(12), 2362–2367.
Berger, L. R., Hawks, J., de Ruiter, D. J., Churchill, S. E., Schmid, P., Delezene, L. K., Kivell, T. L., Garvin, H. M., Williams, S. A., DeSilva, J. M., Skinner, M. M., Musiba, C. M., Cameron, N., Holliday, T. W., Harcourt-Smith, W., Ackermann, R. R., Bastir, M., Bogin, B., Bolter, D. ... Zipfel, B. (2015). Homo naledi, a new species of the genus Homo from the dinaledi chamber South Africa. eLife, 4, e09560. https://doi.org/10.7554/eLife.09560
Bergman, T. J., Ho, L., & Beehner, J. C. (2009). Chest color and social status in male geladas (Theropithecus gelada). International Journal of Primatology, 30(6), 791–806.
Bergstrom, C. T. (2009). Dealing with deception in biology. In B. Harrington (Ed.), Deception: From ancient empires to internet dating (pp. 19–37). Stanford University Press.
Bernatchez, J. (2008). Geochemical characterization of archaeological ochre at Nelson Bay Cave (Western Cape Province), South Africa. The South African Archaeological Bulletin, 63(187), 3–11.
Bernatchez, J. (2012). The role of ochre in the development of modern human behavior: A case study from South Africa. Doctoral dissertation, Arizona State University.
Bielert, C., Girolami, L., & Jowell, S. (1989). An experimental examination of the colour component in visually mediated sexual arousal of the male chacma baboon (Papio ursinus). Journal of Zoology, 219(4), 569–579.
Blome, M. W., Cohen, A. S., Tryon, C. A., Brooks, A. S., & Russell, J. (2012). The environmental context for the origins of modern human diversity: A synthesis of regional variability in African climate 150,000–30,000 years ago. Journal of Human Evolution, 62(5), 563–592. https://doi.org/10.1016/j.jhevol.2012.01.011
Bluhm, L. E., & Surovell, T. A. (2019). Validation of a global model of taphonomic bias using geologic radiocarbon ages. Quaternary Research, 91(1), 325–328.
Boesch, C. (2012). Wild cultures: A comparison between chimpanzee and human cultures. Cambridge University Press.
Bolus, M. (2015). Dispersals of early humans: Adaptations, frontiers, and new territories. In W. Henke & I. Tattersall (Eds.), Handbook of paleoanthropology (2nd ed., pp. 2371–2400). Springer.
Bonnie, K. E., Horner, V., Whiten, A., & de Waal, F. B. M. (2007). Spread of arbitrary conventions among chimpanzees: A controlled experiment. Proceedings of the Royal Society B, 274, 367–372. https://doi.org/10.1098/rspb.2006.3733
Boshier, A., & Beaumont, P. B. (1972). Mining in southern Africa and the emergence of modern man. Optima, 22, 2–12.
Botha, R., & Knight, C. (Eds.). (2009a). The cradle of language. Oxford University Press.
Botha, R., & Knight, C. (Eds.). (2009b). The prehistory of language. Oxford University Press.
Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J. L., Stringer, C., Turner, E., Ward, S., Moutmir, A., & Stambouli, A. (2007). 82,000-year-old shell beads from North Africa and implications for the origins of modern human behaviour. Proceedings of the National Academy of Sciences of the USA, 104(24), 9964–9969.
Bouzouggar, A., & Barton, R. N. E. (2012). The identity and timing of the Aterian in Morocco. In J.-J. Hublin & S. J. P. McPherron (Eds.), Modern origins: A North African perspective (pp. 93–105). Springer.
Bower, J. R. F., Mabulla, A. Z. P., & Kobusiewicz, M. (2012). Loiyangalani: A cultural isolate in the Middle Stone Age of northern Tanzania. In J. Kabaciński, M. Chłodnicki, & M. Kobusiewicz (Eds.), Studies in African archaeology: Prehistory of northeastern Africa: New Ideas and Discoveries (Vol. 11, pp. 479–491). Poznan Archaeological Museum.
Boyer, P., & Liénard, P. (2006). Why ritualized behavior? Precaution systems and action parsing in developmental, pathological and cultural rituals. Behavioral and Brain Sciences, 29(6), 595–650. https://doi.org/10.1017/S0140525X06009332
Brandt, S. A., Fisher, E. C., Hildebrand, E. A., Vogelsang, R., Ambrose, S. H., Lesur, J., & Wang, H. (2012). Early MIS 3 occupation of Mochena Borago rockshelter, southwest Ethiopian highlands: Implications for Late Pleistocene archaeology, paleoenvironments and modern human dispersals. Quaternary International, 274(1), 38–54. https://doi.org/10.1016/j.quaint.2012.03.047
Brooks, A. S., Yellen, J. E., Potts, R., Behrensmeyer, A. K., Deino, A. L., Leslie, D. E., Ambrose, S. H., Ferguson, J. R., d’Errico, F., Zipkin, A. M., Whittaker, S., Post, J., Veatch, G. E., Foecke, K., & Clark, J. B. (2018). Long-distance stone transport and pigment use in the earliest Middle Stone Age. Science, 360, 90–94.
Brown, K. S., Marean, C. W., Herries, A. I. R., Jacobs, Z., Tribolo, C., Braun, D., Roberts, D. L., Meyer, M. C., & Bernatchez, J. (2009). Fire as an engineering tool of early modern humans. Science, 325, 859–862.
Buechner, V. L., Maier, M. A., Lichtenfeld, S., & Schwarz, S. (2014). Red—take a closer look. PLoS ONE, 9(9), e108111. https://doi.org/10.1371/journal.pone.0108111
Bulbulia, J. A., & Sosis, R. (2011). Signalling theory and the evolution of religious cooperation. Religion, 41(3), 363–388.
Burkart, J. M., Allon, O., Amici, F., Fichtel, C., Finkenwirth, C., Heschl, A., Isler, K., Kosonen, Z. K., Martins, E., Meulman, E. J., Richiger, R., Rueth, K., Spillmann, B., Wiesendanger, S., & van Schaik, C. P. (2014). The evolutionary origin of human hyper-cooperation. Nature Communications, 5, 4747.
Burkart, J. M., Hrdy, S. B., & van Schaik, C. P. (2009). Cooperative breeding and human cognitive evolution. Evolutionary Anthropology, 18(5), 175–186. https://doi.org/10.1002/evan.20222
Call, J., & Tomasello, M. (Eds.). (2007). The gestural communication of apes and monkeys. Erlbaum.
Carter, P. L., Mitchell, P. J., & Vinnicombe, P. (1988). Sehonghong: The Middle And Later Stone Age industrial sequence at a Lesotho rock-shelter (Vol. 406). BAR International Series.
Changizi, M. A., Zhang, Q., & Shimojo, S. (2006). Bare skin, blood and the evolution of primate colour vision. Biology Letters, 2(2), 217–221.
Chase, P. G. (2006). The emergence of culture: The evolution of a uniquely human way of life. Springer.
Chavaillon, J., & Berthelet, A. (2004). The archaeological sites of Melka Kunture. In J. Chavaillon & M. Piperno (Eds.), Studies on the Early Paleolithic site of Melka Kunture, Ethiopia (pp. 25–80). Istituto Italiano di Preistoria e Protostoria.
Chazan, M., & Horwitz, L. K. (2009). Milestones in the development of symbolic behaviour: A case study from Wonderwerk Cave, South Africa. World Archaeology, 41(4), 521–539.
Chudek, M., & Henrich, J. (2011). Culture-gene coevolution, norm-psychology and the emergence of human prosociality. Trends in Cognitive Sciences, 15(5), 218–226.
Clark, J. D. (1974). Kalambo Falls prehistoric site. Volume II: The later prehistoric cultures. Cambridge University Press.
Clark, J. D. (1988). The Middle Stone Age of East Africa and the beginnings of regional identity. Journal of World Prehistory, 2(3), 235–305.
Clark, J. D., Beyene, Y., WoldeGabriel, G., Hart, W., Renne, P. R., Gilbert, H., Defleur, A., Suwa, G., Katoh, S., Ludwig, K. R., Boisserie, J. R., Asfaw, B., & White, T. D. (2003). Stratigraphic, chronological and behavioral contexts of Pleistocene Homo sapiens from the Middle Awash, Ethiopia. Nature, 423, 747–752.
Clark, J. D., & Brown, K. S. (2001). The twin rivers Kopje, Zambia: Stratigraphy, fauna, and artifact assemblages from the 1954 and 1956 excavations. Journal of Archaeological Science, 28, 305–330.
Clark, J. D., Oakley, K. P., Wells, L. H., & McClelland, J. A. C. (1947). New studies on Rhodesian Man. Journal of the Royal Anthropological Society, 77, 4–33.
Clarkson, C., Jacobs, Z., Marwick, B., Fullagar, R., Wallis, L., Smith, M., Roberts, R. G., Hayes, E., Lowe, K., Carah, X., Florin, A. S., McNeil, J., Cox, D., Arnold, J. L., Hua, Q., Huntley, J., Brand, H. E. A., Manne, T., Fairbairn, A. … Pardoe, C. (2017). Human occupation of northern Australia by 65,000 years ago. Nature, 547, 306–310.
Close, A. E. (1993). BT-14 main excavation: The archaeological sequence of the East Lake (1986 and 1987 seasons). In F. Wendorf, R. Schild, & A. E. Close (Eds.), Egypt during the last interglacial: The middle Paleolithic of Bir Tarfawi and Bir Sahara East (pp. 288–344). Plenum Press.
Cohen, A. S., Stone, J. R., Beuning, K. R. M., Park, L. E., Reinthal, P. N., Dettman, D., & Ivory, S. J. (2007). Ecological consequences of early Late Pleistocene megadroughts in tropical Africa. Proceedings of the National Academy of Sciences of the USA, 104(42), 16422–16427.
Cole, J. (2017). Accessing hominin cognition: Language and social signaling in Lower to Middle Palaeolithic. In T. Wynn & F. L. Coolidge (Eds.), Cognitive models in Palaeolithic archaeology (pp. 157–196). Oxford University Press.
Collard, M., Ruttle, A., Buchanan, B., & O’Brien, M. J. (2013). Population size and cultural evolution in nonindustrial food-producing societies. PLoS ONE, 8(9), e72628. https://doi.org/10.1371/journal.pone.0072628
Cooke, C. K. (1963). Report on excavations at Pomongwe and Tshangula Caves, Matopo Hills, Southern Rhodesia. The South African Archaeological Bulletin, 18(71), 73–151.
Cooke, C. K. (1966). The archaeology of the Mafungabusi area, Gokwe, Rhodesia. Proceedings and Transactions of the Rhodesian Scientific Association, 51, 51–78.
Cooke, C. K. (1971). Excavations in Zombepata Cave, Sipolilo District, Mashonaland, Rhodesia. The South African Archaeological Bulletin, 26(103/104), 104–126.
Coulson, S., Staurset, S., & Walker, N. (2011). Ritualized behavior in the Middle Stone Age: Evidence from Rhino Cave, Tsodilo Hills, Botswana. PaleoAnthropology, 2011, 18–61. https://doi.org/10.4207/PA.2011.ART42
Crockford, C., Wittig, R. M., & Zuberbühler, K. (2017). Vocalizing in chimpanzees is influenced by social-cognitive processes. Science Advances, 3(11), e1701742. https://doi.org/10.1126/sciadv.1701742
Cronk, L., Steklis, D., Steklis, N., van den Akker, O. R., & Aktipis, A. (2019). Kin terms and fitness interdependence. Evolution and Human Behavior, 40(3), 281–291. https://doi.org/10.1016/j.evolhumbehav.2018.12.004
Cruz-Uribe, K., Klein, R. G., Avery, G., Avery, M., Halkett, D., Hart, T., Milo, R. G., Sampson, C. G., & Volman, T. P. (2003). Excavation of buried late Acheulean (mid-Quaternary) land surfaces at Duinefontein 2, Western Cape Province, South Africa. Journal of Archaeological Science, 30(5), 559–575.
Dal Pesco, F., & Fischer, J. (2018). Greetings in male Guinea baboons and the function of rituals in complex social groups. Journal of Human Evolution, 125, 87–98. https://doi.org/10.1016/j.jhevol.2018.10.007
Dal Pesco, F., & Fischer, J. (2020). On the evolution of baboon greeting rituals. Philosophical Transactions of the Royal Society B, 375(1805), 20190420. https://doi.org/10.1098/rstb.2019.0420
Dart, R. A., & Beaumont, P. B. (1967). Amazing antiquity of mining in southern Africa. Nature, 216, 407–408.
Dart, R. A., & Beaumont, P. B. (1969). Evidence of iron ore mining in Southern Africa in the Middle Stone Age. Current Anthropology, 10(1), 127–128.
Dart, R. A., & Beaumont, P. B. (1971). On a further radiocarbon date for ancient mining in Southern Africa. South African Journal of Science, 67, 10–11.
Dart, R. A., & del Grande, N. (1931). The ancient iron smelting cavern at Mumbwa. Transactions of the Royal Society of South Africa, 19(4), 379–427.
Davies, S. (2020). Adornment: What self-decoration tells us about who we are. Bloomsbury Academic.
Dayet, L. (2012). Matériaux, transformations et fonctions de l’ocre au Middle Stone Age: Le cas de Diepkloof Rock Shelter dans le contexte de l’Afrique australe. Doctoral dissertation, Université Bordeaux.
Dayet, L., Le Bourdonnec, F.-X., Daniel, F., Porraz, G., & Texier, J.-P. (2016). Ochre provenance and procurement strategies during the Middle Stone Age at Diepkloof Rock shelter South Africa. Archaeometry, 58(5), 807–829. https://doi.org/10.1111/arcm.12202
Dayet, L., Texier, J.-P., Daniel, F., & Porraz, G. (2013). Ochre resources from the Middle Stone Age sequence of Diepkloof Rock shelter, Western Cape, South Africa. Journal of Archaeological Science, 40(9), 3492–3505.
De la Peña, P., Val, A., Stratford, D. J., Colino, F., Esteban, I., Fitchett, J. M., HodgskissMatembo, T. J., & Moll, R. (2019). Revisiting Mwulu’s Cave: New insights into the Middle Stone Age in the southern African savanna biome. Archaeological and Anthropological Sciences, 11, 3239–3266.
De Villiers, H. (1973). Human skeletal remains from Border Cave, Ingwavuma district, Kwazulu, South Afrika. Annals of the Transvaal Museum, 28(13), 229–256.
De Waal, F. B. M. (1989). Peacemaking among primates. Harvard University Press.
De Waal, F.B.M. (2007). Chimpanzee politics: Power and sex among apes (25th anniversary edition). Johns Hopkins University Press.
Deacon, J. (1995). An unsolved mystery at the Howieson’s Poort name site. The South African Archaeological Bulletin, 50(162), 110–120.
Deacon, T. W. (1997). The symbolic species: The co-evolution of language and the brain. W. W. Norton.
Deino, A. L., Behrensmeyer, A. K., Brooks, A. S., Yellen, J. E., Sharp, W. D., & Potts, R. (2018). Chronology of the Acheulean to Middle Stone Age transition in eastern Africa. Science, 360, 95–98.
Deino, A. L., & McBrearty, S. (2002). 40Ar/39Ar dating of the Kapthurin formation, Baringo Kenya. Journal of Human Evolution, 42(1–2), 185–210.
Dennett, D. C. (2017). From bacteria to Bach and back: The evolution of minds. Norton.
Derex, M., Beugin, M.-P., Godelle, B., & Raymond, M. (2013). Experimental evidence for the influence of group size on cultural complexity. Nature, 503, 389–391.
Derex, M., & Boyd, R. (2015). The foundations of the human cultural niche. Nature Communications, 6, 8398. https://doi.org/10.1038/ncomms9398
D’Errico, F. (2003). The invisible frontier: A multiple species model for the origin of behavioural modernity. Evolutionary Anthropology, 12, 188–202.
D’Errico, F. (2008). Le rouge et le noir: Implications of early pigment use in Africa, the Near East and Europe for the origin of cultural modernity. South African Archaeological Society Goodwin Series, 10, 168–174.
D’Errico, F., & Backwell, L. (2016). Earliest evidence of personal ornaments associated with burial: The Conus shells from Border Cave. Journal of Human Evolution, 93, 91–108.
D’Errico, F., García Moreno, R., & Rifkin, R. F. (2012). Technological, elemental and colorimetric analysis of an engraved ochre fragment from the Middle Stone Age levels of Klasies River Cave 1, South Africa. Journal of Archaeological Science, 39, 942–952.
D’Errico, F., & Henshilwood, C. S. (2007). Additional evidence for bone technology in the southern African Middle Stone Age. Journal of Human Evolution, 52, 142–163.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A. M., Soressi, M., Bresson, F., Maureille, B., Nowell, A., Lakarra, J., Backwell, L., & Julien, M. (2003). Archaeological evidence for the emergence of language, symbolism, and music: An alternative multidisciplinary perspective. Journal of World Prehistory, 17, 1–70.
D’Errico, F., Henshilwood, C. S., & Nilssen, P. J. (2001). An engraved bone fragment from c. 70,000-year-old Middle Stone Age levels at Blombos Cave, South Africa: Implications for the origins of symbolism and language. Antiquity, 75, 309–318.
D’Errico, F., Henshilwood, C. S., Vanhaeren, M., & van Niekerk, K. L. (2005). Nassarius kraussianus shell beads from Blombos Cave: Evidence for symbolic behaviour in the Middle Stone Age. Journal of Human Evolution, 48, 3–24.
D’Errico, F., & Stringer, C. B. (2011). Evolution, revolution or saltation scenario for the emergence of modern cultures? Philosophical Transactions of the Royal Society B, 366, 1060–1069. https://doi.org/10.1098/rstb.2010.0340
D’Errico, F., Vanhaeren, M., Barton, N., Bouzouggar, A., Mienis, H. K., Richter, D., Hublin, J. J., McPherron, S. P., & Lozoueth, P. (2009). Additional evidence on the use of personal ornaments in the Middle Paleolithic of North Africa. Proceedings of the National Academy of Sciences of the USA, 106(38), 16051–16056. https://doi.org/10.1073/pnas.0903532106
D’Errico, F., Vanhaeren, M., & Wadley, L. (2008). Possible shell beads from the Middle Stone Age layers of Sibudu Cave, South Africa. Journal of Archaeological Science, 35(10), 2675–2685.
Dibble, H. L., Aldeias, V., Jacobs, Z., Olszewski, D. I., Rezek, Z., Lin, S. C., Alvarez-Fernández, E., Barshay-Szmidt, C. C., Hallett-Desguez, E., Reed, D., Reed, K., Richter, D., Steele, T. E., Skinner, A., Blackwell, B., Doronicheva, E., & El-Hajraoui, M. (2013). On the industrial attributions of the Aterian and Mousterian of the Maghreb. Journal of Human Evolution., 64(3), 194–210.
Dirks, P. H. G. M., Roberts, E. M., Hilbert-Wolf, H., Kramers, J. D., Hawks, J., Dosseto, A., Duval, M., Elliott, M., Evans, M., Grün, R., Hellstrom, J., Herries, A. I. R., Joannes-Boyau, R., Makhubela, T. V., Placzek, C. J., Robbins, J., Spandler, C., Wiersma, J., Woodhead, J., & Berger, L. R. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave South Africa. eLife, 6, e24231. https://doi.org/10.7554/eLife.24231
Dissanayake, E. (2018). From play and ritualisation to ritual and its arts: Sources of Upper Pleistocene ritual practices in Lower Middle Pleistocene ritualised and play behaviours in ancestral hominins. In C. Renfrew, I. Morley, & M. J. Boyd (Eds.), Ritual, play and belief, in evolution and early human societies (pp. 87–100). Cambridge University Press.
Dixson, A. F. (2012). Primate sexuality: Comparative studies of the prosimians, monkeys, apes, and human beings (2nd ed.). Oxford University Press.
Dor, D., Knight, C., & Lewis, J. (Eds.). (2014). The social origins of language. Oxford University Press.
Douka, K., Jacobs, Z., Lane, C., Grün, R., Farr, L., Hunt, C., Inglis, R. H., Reynolds, T., Albert, P., Aubert, M., Cullen, V., Hill, E., Kinsley, L., Roberts, R. G., Tomlinson, E. L., Wulf, S., & Barker, G. (2014). The chronostratigraphy of the Haua Fteah cave (Cyrenaica, northeast Libya). Journal of Human Evolution., 66, 39–63. https://doi.org/10.1016/j.jhevol.2013.10.001
Dubin, L. S. (1999). North American Indian jewelry and adornment: From prehistory to the present. Harry N. Abrams.
Dunbar, R. I. M. (2003). The social brain: Mind, language, and society in evolutionary perspective. Annual Review of Anthropology, 32(1), 163–181. https://doi.org/10.1146/annurev.anthro.32.061002.093158
Dunbar, R. I. M. (2009). Mind the bonding gap: Constraints on the evolution of hominin societies. In S. Shennan (Ed.), Pattern and process in cultural evolution (pp. 223–234). University of California Press.
Dunbar, R. I. M., Kaskatis, K., MacDonald, I., & Barra, V. (2012). Performance of music elevates pain threshold and positive affect: Implications for the evolutionary function of music. Evolutionary Psychology, 10(4), 688–702.
Durkheim, É. (1912). Les formes élémentaires de la vie religieuse: Le système totémique en Australie. Alcan.
Ebin, V. (1979). The body decorated. Thames & Hudson.
Ehrmann, W., Schmiedl, G., Beuscher, S., & Krüger, S. (2017). Intensity of African humid periods estimated from Saharan dust fluxes. PLoS ONE, 12(1), e0170989. https://doi.org/10.1371/journal.pone.0170989
Eisenhofer, S. (Ed.). (2005). Painted people: Die Faszination des zweiten Gesichts. Kunth.
Eiwanger, J. (2013). Farbgebrauch im Paläolithikum der Ifri n'Ammar (Nordost-Marokko). In H. Meller Ed., Tagungen des Landesmuseums für Vorgeschichte Halle Vol. 10. Rot - Die Archäologie bekennt Farbe: 5. Mitteldeutscher Archäologentag vom 04. bis 06. Oktober 2012 in Halle (Saale) (pp. 79–87). Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt.
Elliot, A. J. (2015). Color and psychological functioning: A review of theoretical and empirical work. Frontiers in Psychology, 6, 368. https://doi.org/10.3389/fpsyg.2015.00368
Elliot, A. J., Fairchild, M. D., & Franklin, A. (Eds.). (2015). Handbook of color psychology. Cambridge University Press.
Elliot, A. J., Greitemeyer, T., & Pazda, A. D. (2013a). Women’s use of red clothing as a sexual signal in intersexual interaction. Journal of Experimental Social Psychology, 49(3), 599–602.
Elliot, A. J., & Maier, M. A. (2014). Color psychology: Effects of perceiving color on psychological functioning in humans. Annual Review of Psychology, 65, 95–120.
Elliot, A. J., & Niesta, D. (2008). Romantic red: Red enhances men’s attraction to women. Journal of Personality and Social Psychology, 95(5), 1150–1164. https://doi.org/10.1037/0022-3514.95.5.1150
Elliot, A. J., Tracy, J. L., Pazda, A. D., & Beall, A. T. (2013b). Red enhances women’s attractiveness to men: First evidence suggesting universality. Journal of Experimental Social Psychology, 49(1), 165–168. https://doi.org/10.1016/j.jesp.2012.07.017
Evans, U. (1994). Hollow Rock Shelter: A Middle Stone Age site in the Cederberg. Southern African Field Archaeology, 3(2), 63–73.
Fisher, A. (1984). Africa adorned. William Collins.
Fischer, R., & Xygalatas, D. (2014). Extreme rituals as social technologies. Journal of Cognition and Culture, 14, 345–355.
Fischer, R., Xygalatas, D., Mitkidis, P., Reddish, P., Tok, P., Konvalinka, I., & Bulbulia, J. A. (2014). The fire-walker’s high: Affect and physiological responses in an extreme collective ritual. PLoS ONE, 9(2), e88355. https://doi.org/10.1371/journal.pone.0088355
Folk, C. L. (2015). The role of color in the voluntary and involuntary guidance of selective attention. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 481–501). Cambridge University Press.
Gamble, C., Gowlett, J. A. J., & Dunbar, R. I. M. (2014). Thinking big: How the evolution of social life shaped the human mind. Thames & Hudson.
Gerald, M. S., Waitt, C., Little, A. C., & Kraiselburd, E. (2007). Females pay attention to female secondary sexual color: An experimental study in Macaca mulatta. International Journal of Primatology, 28(1), 1–7.
Gibson, N. E., Wadley, L., & Williamson, B. S. (2004). Microscopic residues as evidence of hafting on backed tools from the 60000 to 68000 year-old Howiesons Poort layers of Rose Cottage Cave, South Africa. Southern African Humanities, 16, 1–11.
Gliganic, L. A., Jacobs, Z., Roberts, R. G., Domínguez-Rodrigo, M., & Mabulla, A. Z. (2012). New ages for Middle and Later Stone Age deposits at Mumba rockshelter, Tanzania: Optically stimulated luminescence dating of quartz and feldspar grains. Journal of Human Evolution, 62(4), 533–547. https://doi.org/10.1016/j.jhevol.2012.02.004
Goodall, J. (1986). The chimpanzees of Gombe: Patterns of behavior. Belknap.
Goodall, J. (2005). Primate spirituality. In B. Taylor (Ed.), Encyclopedia of religion and nature (pp. 1303–1306). Bloomsbury.
Gowlett, J. A. J., Gamble, C., & Dunbar, R. I. M. (2012). Human evolution and the archaeology of the social brain. Current Anthropology, 53(6), 693–722. https://doi.org/10.1086/667994
Grimes, R. L. (2014). The craft of ritual studies. Oxford University Press.
Gröning, K. (Ed.). (2001). Geschmückte Haut: Eine Kulturgeschichte der Körperkunst (2nd ed.). Frederking & Thaler.
Groucutt, H. S., Petraglia, M. D., Bailey, G., Scerri, E. M. L., Parton, A., Clark-Balzan, L., & Scally, A. (2015). Rethinking the dispersal of Homo sapiens out of Africa. Evolutionary Anthropology, 24(4), 149–164. https://doi.org/10.1002/evan.21455
Grün, R., & Beaumont, P. B. (2001). Border Cave revisited: A revised ESR chronology. Journal of Human Evolution, 40, 467–482.
Grün, R., Beaumont, P. B., Tobias, P. V., & Eggins, S. (2003). On the age of Border Cave 5 human mandible. Journal of Human Evolution, 45, 155–167.
Grün, R., Pike, A., McDermott, F., Eggins, S., Mortimer, G., Aubert, M., Kinsley, L., Joannes-Boyau, R., Rumsey, M., Denys, C., Brink, J., Clark, T., & Stringer, C. (2020). Dating the skull from Broken Hill, Zambia, and its position in human evolution. Nature, 580, 372–375. https://doi.org/10.1038/s41586-020-2165-4
Guenther, M. G. (1999). Tricksters and trancers: Bushman religion and society. Indiana University Press.
Haidle, M. N., Bolus, M., Bruch, A. A., Hertler, C., Kandel, A. W., Märker, M., & Mosbrugger, V. (2010). The role of culture in early expansions of humans—A new research center. Quaternary International, 223–224, 429–430. https://doi.org/10.1016/j.quaint.2009.07.011
Haidle, M. N., Bolus, M., Collard, M., Conard, N. J., Garofoli, D., Lombard, M., Nowell, A., Tennie, C., & Whiten, A. (2015). The nature of culture: An eight-grade model for the evolution and expansion of cultural capacities in hominins and other animals. Journal of Anthropological Sciences., 93, 43–70. https://doi.org/10.4436/jass.93011
Haidle, M. N., & Kandel, A. W. (2018). ROCEEH Out of Africa Database (ROAD)—Applications. https://www.hadw-bw.de/sites/default/files/documents/ROCEEH-ROAD%20Applications%202018.pdf
Halkett, D., Hart, T., Yates, R., Volman, T. P., Parkington, J. E., Orton, J., Klein, R. G., Cruz-Uribe, K., & Avery, G. (2003). First excavation of intact Middle Stone Age layers at Ysterfontein, Western Cape Province, South Africa: Implications for Middle Stone Age ecology. Journal of Archaeological Science, 30(8), 955–971. https://doi.org/10.1016/S0305-4403(02)00273-X
Hammer, V. M. F., Woerner, A. E., Mendez, F. L., Watkins, J. C., & Wall, J. D. (2011). Genetic evidence for archaic admixture in Africa. Proceedings of the National Academy of Sciences of the USA, 108(37), 15123–15128. https://doi.org/10.1073/pnas.1109300108
Harrod, J. B. (2014). Palaeoart at two million years ago? A review of the evidence. Arts, 3(1), 135–155. https://doi.org/10.3390/arts3010135
Harvati, K., Röding, C., Bosman, A. M., Karakostis, F. A., Grün, R., Stringer, C., Karkanas, P., Thompson, N. C., Koutoulidis, V., Moulopoulos, L. A., Gorgoulis, V. G., & Kouloukoussa, M. (2019). Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature, 571, 500–504. https://doi.org/10.1038/s41586-019-1376-z
Henrich, J. (2004). Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses: The Tasmanian case. American Antiquity, 69(2), 197–214.
Henrich, J. (2009). The evolution of costly displays, cooperation and religion: Credibility, enhancing display and their implications for cultural evolution. Evolution and Human Behavior, 30, 244–260.
Henrich, J. (2016). The secret of our success: How culture is driving human evolution, domesticating our species, and making us smarter. Princeton University Press.
Henrich, J., Boyd, R., Derex, M., Kline, M. A., Mesoudi, A., Muthukrishna, M., Powell Stephen, A. T., Shennan, J., & Thomas, M. G. (2016). Understanding cumulative cultural evolution. Proceedings of the National Academy of Sciences of the USA, 113(44), E6724–E6725. https://doi.org/10.1073/pnas.1610005113
Henshilwood, C. S. (2012). Late Pleistocene techno-traditions in southern Africa: A review of the Still Bay and Howiesons Poort, c. 75–59 ka. Journal of World Prehistory, 25(3–4), 205–237.
Henshilwood, C. S., & d’Errico, F. (Eds.). (2011a). Homo symbolicus: The dawn of language, imagination and spirituality. John Benjamins.
Henshilwood, C. S., & d’Errico, F. (2011b). Middle Stone Age engravings and their significance to the debate on the emergence of symbolic material culture. In C. S. Henshilwood & F. d’Errico (Eds.), Homo symbolicus: The dawn of language, imagination and spirituality (pp. 75–96). John Benjamins.
Henshilwood, C. S., d’Errico, F., Marean, C. W., Milo, R. G., & Yates, R. J. (2001b). An early bone tool industry from the Middle Stone Age at Blombos Cave, South Africa: Implications for the origins of modern human behaviour, symbolism and language. Journal of Human Evolution, 41(6), 631–678.
Henshilwood, C. S., d’Errico, F., van Niekerk, K. L., Coquinot, Y., Jacobs, Z., Lauritzen, S.-E., Menu, M., & García Moreno, R. (2011). A 100,000-year-old ochre-processing workshop at Blombos Cave, South Africa. Science, 334, 219–222. https://doi.org/10.1126/science.1211535
Henshilwood, C. S., d’Errico, F., van Niekerk, K. L., Dayet, L., Queffelec, A., & Pollarolo, L. (2018). An abstract drawing from the 73,000-year-old levels at Blombos Cave, South Africa. Nature, 562, 115–118. https://doi.org/10.1038/s41586-018-0514-3
Henshilwood, C. S., d’Errico, F., Vanhaeren, M., van Niekerk, K. L., & Jacobs, Z. (2004). Middle Stone Age shell beads from South Africa. Science, 304, 404.
Henshilwood, C. S., d’Errico, F., & Watts, I. (2009). Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa. Journal of Human Evolution, 57(1), 27–47. https://doi.org/10.1016/j.jhevol.2009.01.005
Henshilwood, C. S., d’Errico, F., Yates, R. J., Jacobs, Z., Tribolo, C., Duller, G. A. T., & Wintle, A. G. (2002). Emergence of modern human behavior: Middle Stone Age engravings from South Africa. Science, 295, 1278–1280.
Henshilwood, C. S., & Dubreuil, B. (2011). The Still Bay and Howiesons Poort, 77–59 ka: Symbolic material culture and the evolution of the mind during the African Middle Stone Age. Current Anthropology, 52(3), 361–400.
Henshilwood, C. S., & Marean, C. W. (2003). The origin of modern human behaviour: Critique of the models and their test implications. Current Anthropology, 44(5), 627–651.
Henshilwood, C. S., Sealy, J. C., Yates, R. J., Cruz-Uribe, K., Goldberg, P., Grine, F. E., Kleinh, R. G., Poggenpoeli, C., van Niekerkj, K., Wattsk, I., & Watts, I. (2001a). Blombos Cave, southern Cape, South Africa: Preliminary report on the 1992–1999 excavations of the Middle Stone Age levels. Journal of Archaeological Science, 28, 421–448.
Henshilwood, C. S., van Niekerk, K. L., Wurz, S., Delagnes, A., Armitage, S. J., Rifkin, R. F., Douze, K., Keene, P., Haaland, M. M., Reynard, J., Discamps, E., & Mienies, S. S. (2014). Klipdrift Shelter, southern Cape, South Africa: Preliminary report on the Howiesons Poort layers. Journal of Archaeological Science., 45, 284–303. https://doi.org/10.1016/j.jas.2014.01.033
Hermanowicz, J. C., & Morgan, H. P. (1999). Ritualizing the routine: Collective identity affirmation. Sociological Forum, 14(2), 197–214.
Herries, A. I. R. (2011). A chronological perspective on the Acheulian and its transition to the Middle Stone Age in southern Africa: The question of the Fauresmith. International Journal of Evolutionary Biology, 2011, 230156. https://doi.org/10.4061/2011/961401
Hershkovitz, I., Weber, G. W., Quam, R., Duval, M., Grün, R., Kinsley, L., & Weinstein-Evron, M. (2018). The earliest modern humans outside Africa. Science, 359(6374), 456–459.
Higham, J. P., Heistermann, M., Saggau, C., Agil, M., Perwitasari-Farajallah, D., & Engelhardt, A. (2012). Sexual signalling in female crested macaques and the evolution of primate fertility signals. BMC Evolutionary Biology, 12(1), 89. https://doi.org/10.1186/1471-2148-12-89.
Higham, J. P., & Winters, S. (2015). Color and mate choice in non-human animals. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 502–530). Cambridge University Press.
Hill, K. R., Wood, B. M., Baggio, J., Hurtado, A. M., & Boyd, R. (2014). Hunter-gatherer inter-band interaction rates: Implications for cumulative culture. PLoS ONE, 9(7), e102806. https://doi.org/10.1371/journal.pone.0102806
Hobson, N. M., & Inzlicht, M. (2016). Recognizing religion’s dark side: Religious ritual increases antisociality and hinders self-control. Behavioral and Brain Sciences, 39, 30–31.
Hobson, N. M., Schroeder, J., Risen, J. L., Xygalatas, D., & Inzlicht, M. (2018). The psychology of rituals: An integrative review and process-based framework. Personality and Social Psychology Review, 22(3), 260–284. https://doi.org/10.1177/1088868317734944
Hodgskiss, T. P. (2012). An investigation into the properties of the ochre from Sibudu, KwaZulu-Natal South Africa. Southern African Humanities, 24(1), 99–120.
Hodgskiss, T. P. (2013a). Ochre use in the Middle Stone Age at Sibudu, South Africa: Grinding, rubbing, scoring and engraving. Journal of African Archaeology, 11(1), 75–95.
Hodgskiss, T. P. (2013b). Ochre use at Sibudu Cave and its link to complex cognition in the Middle Stone Age. Doctoral dissertation, University of the Witwatersrand.
Hodgskiss, T. P. (2014). Cognitive requirements for ochre use in the Middle Stone Age at Sibudu South Africa. Cambridge Archaeological Journal, 24(03), 405–428. https://doi.org/10.1017/S0959774314000663
Hodgskiss, T. P., & Wadley, L. (2017). How people used ochre at Rose Cottage Cave, South Africa: Sixty thousand years of evidence from the Middle Stone Age. PLoS ONE, 12(4), e0176317. https://doi.org/10.1371/journal.pone.0176317
Hodgson, D. (2017). Deciphering patterns in the archaeology of South Africa: The neurovisual resonance theory. In T. Wynn & F. L. Coolidge (Eds.), Cognitive models in Palaeolithic archaeology (pp. 133–156). Oxford University Press.
Horner, V., & de Waal, F. B. M. (2009). Controlled studies of chimpanzee cultural transmission. In J. Y. Chiao (Ed.), Progress in brain research: Cultural neuroscience: Cultural influences on brain function (Vol. 178, pp. 3–15). Elsevier.
Horton, D. (Ed.) (1994). The encyclopaedia of Aboriginal Australia: Aboriginal and Torres Strait Islander history, society and culture (Vol. 2). Aboriginal Studies Press.
Hovers, E., Ilani, S., Bar-Yosef, O., & Vandermeersch, B. (2003). An early case of color symbolism: Ochre use by modern humans in Qafzeh Cave. Current Anthropology, 44(4), 491–522.
Hrdy, S. B. (2009). Mothers and others: The evolutionary origins of mutual understanding. Belknap.
Hublin, J.-J., Ben-Ncer, A., Bailey, S. E., Freidline, S. E., Neubauer, S., Skinner, M. M., Bergmann, I., Le Cabec, A., Benazzi, S., Harvati, K., & Gunz, P. (2017). New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature, 546, 289–292.
Humphrey, C., & Laidlaw, J. (1994). The archetypal actions of ritual: A theory of ritual illustrated by the Jain rite of worship. Clarendon Press.
Huppin, M., Malamuth, N. M. (2017). Pornography as a human product and source of data. In: T. K. Shackelford, V. A. Weekes-Shackelford (Eds.), Encyclopedia of evolutionary psychological science. Springer. https://doi.org/10.1007/978-3-319-16999-6_3581-2.
Huxley, J. S. (1914). The courtship habits of the great crested grebe (Podiceps cristatus) with an addition to the theory of sexual selection. Proceedings of the Zoological Society of London, 84(3), 491–562. https://doi.org/10.1111/j.1469-7998.1914.tb07052.x
Huxley, J. S. (1966). A discussion on ritualization of behaviour in animals and man: Introduction. Philosophical Transactions of the Royal Society B, 251(772), 249–271.
Inskeep, R. (1962). The age of the Kondoa rock paintings in the light of recent excavations at Kisese II rock shelter. In G. Mortelmans & J. Nenquin (Eds.), Actes du IVe Congrès panafricain de préhistoire et de l’étude du quaternaire (pp. 249–256). Museé Royal de l’Afrique Centrale.
Irons, W. (2001). Religion as a hard-to-fake sign of commitment. In R. M. Nesse (Ed.), Evolution and the capacity for commitment (pp. 292–309).
Jackson, J. C., Jong, J., Bilkey, D., Whitehouse, H., Zollmann, S., McNaughton, C., & Halberstadt, J. (2018). Synchrony and physiological arousal increase cohesion and cooperation in large naturalistic groups. Scientific Reports, 8(1), 127. https://doi.org/10.1038/s41598-017-18023-4
Jacobs, G. H. (2015). Evolution of color vision and its reflections in contemporary mammals. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 110–130). Cambridge University Press.
Johnson, C. R., & McBrearty, S. (2010). 500,000 year old blades from the Kapthurin Formation Kenya. Journal of Human Evolution, 58(2), 193–200.
Jones, N. (1940). Bambata Cave: A reorientation. Occasional Papers of the National Museum of Southern Rhodesia, 9, 11–28.
Jones, S., Antoniadou, A., Barton, H., Drake, N. A., Farr, L., Hunt, C., Inglis, R., Reynolds, T., White, K., & Barker, G. (2016). Patterns of hominin occupation and cultural diversity across the Gebel Akhdar of northern Libya over the last ~200 kyr. In S. C. Jones & B. A. Stewart (Eds.), Africa from MIS 6–2: Population dynamics and paleoenvironments (pp. 77–99). Springer. https://doi.org/10.1007/978-94-017-7520-5_5.
Kaboth-Bahr, S., Gosling, W. D., Vogelsang, R., Bahr, A., Scerri, E. M. L., Asrat, A., Cohen, A. S., Düsing, W., Foerster, V., Lamb, H. F., Maslin, M. A., Roberts, H. M., Schäbitz, F., & Trauth, M. H. (2021). Paleo-ENSO influence on African environments and early modern humans. Proceedings of the National Academy of Sciences of the USA, 118(23), e2018277118. https://doi.org/10.1073/pnas.2018277118
Kandel, A. W., Bolus, M., Bretzke, K., Bruch, A. A., Haidle, M. N., Hertler, C., & Märker, M. (2016). Increasing behavioral flexibility? An integrative macro-scale approach to understanding the Middle Stone Age of Southern Africa. Journal of Archaeological Method and Theory, 23(2), 623–668. https://doi.org/10.1007/s10816-015-9254-y
Kapitány, R., Kavanagh, C., & Whitehouse, H. (2020) Ritual morphospace revisited: The form, function and factor structure of ritual practice. Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1805). 20190436, https://doi.org/10.1098/rstb.2019.0436.
Kapitány, R., & Nielsen, M. (2015). Adopting the ritual stance: The role of opacity and context in ritual and everyday actions. Cognition, 145, 13–29.
Kapitány, R., & Nielsen, M. (2016). The ritual stance and the precaution system: The role of goal-demotion and opacity in ritual and everyday actions. Religion, Brain & Behavior, 7(1), 1–16.
Kapitány, R., & Nielsen, M. (2019). Ritualized objects: How we perceive and respond to causally opaque and goal demoted action. Journal of Cognition and Culture, 19(1–2), 170–194. https://doi.org/10.1163/15685373-12340053
Keller, C. M. (1973). Montagu Cave in prehistory: A descriptive analysis. University of California Press.
Kelsey, B., Brandt, S., Zipkin, A., & Wilson, E. (2019). Provenance of ochre at Mochena Borago Rockshelter, SW Ethiopia: An interdisciplinary approach [Presentation]. EAAPP, Nairobi.
Keupp, S., Behne, T., & Rakoczy, H. (2013). Why do children overimitate? Normativity is crucial. Journal of Experimental Child Psychology, 116(2), 392–406. https://doi.org/10.1016/j.jecp.2013.07.002
Kim, H. L., Ratan, A., Perry, G. H., Montenegro, A., Miller, W., & Schuster, S. C. (2014). Khoisan hunter-gatherers have been the largest population throughout most of modern-human demographic history. Nature Communications, 5, 5692. https://doi.org/10.1038/ncomms6692
Klein, R. G., Avery, G., Cruz-Uribe, K., Halkett, D., Parkington, J. E., Steele, T. E., Volman, T. P., & Yates, R. (2004). The Ysterfontein 1 Middle Stone Age site, South Africa, and early human exploitation of coastal resources. Proceedings of the National Academy of Sciences of the USA, 101(16), 5708–5715.
Kline, M. A., & Boyd, R. (2010). Population size predicts technological complexity in Oceania. Proceedings of the Royal Society B, 277, 2559–2564.
Knight, C. (1999). Sex and language as pretend-play. In R. I. M. Dunbar, C. Knight, & C. Power (Eds.), The evolution of culture: An interdisciplinary view (pp. 228–250). Rutgers University Press.
Knight, C. (2014). Language and symbolic culture: An outcome of hunter-gatherer reverse dominance. In D. Dor, C. Knight, & J. Lewis (Eds.), Oxford studies in the evolution of language: The social origins of language (Vol. 19, pp. 228–248). Oxford University Press.
Knight, C., Power, C., & Watts, I. (1995). The human symbolic revolution: A Darwinian account. Cambridge Archaeological Journal, 5(1), 75–114.
Kobayashi, Y., & Aoki, K. (2012). Innovativeness, population size and cumulative cultural evolution. Theoretical Population Biology, 82(1), 38–47.
Kohl-Larsen, L., Rafalski, S., & Tomsky, J. (1985). Funde aus Issanzu, Yambi und Iramba. In H. Müller-Beck (Ed.), Die archäologischen und anthropologischen Ergebnisse der Kohl-Larsen-Expeditionen in Nord-Tanzania 1933–1939 (Vol. 1). Archaeologica Venatoria.
Konvalinka, I., Xygalatas, D., Bulbulia, J. A., Schjoedt, U., Jegindø, E.-M., Wallot, S., & Roepstorff, A. (2011). Synchronized arousal between performers and related spectators in a fire-walking ritual. Proceedings of the National Academy of Sciences of the USA, 108(20), 8514–8519.
Kowalewski, M. (2014). Time-averaging (paleontology). AccessScience. Retrieved July 4, 2017, from https://doi.org/10.1036/1097-8542.802610
Kozowyk, P. R. B., Langejans, G. H. J., & Poulis, J. A. (2016). Lap shear and impact testing of ochre and beeswax in experimental Middle Stone Age compound adhesives. PLoS ONE, 11(3), e0150436. https://doi.org/10.1371/journal.pone.0150436
Krebs, J. R., & Dawkins, R. (1991). Animal signals: Mind-reading and manipulation. In J. R. Krebs & N. B. Davies (Eds.), Behavioural ecology: An evolutionary approach (3rd ed., pp. 380–402). Blackwell Scientific.
Kreinath, J., Snoek, J., & Stausberg, M. (2006). Ritual studies, ritual theory, theorizing rituals: An introductory essay. In J. Kreinath, J. Snoek, & M. Stausberg (Eds.), Theorizing rituals: Issues, topics, approaches, concepts (pp. xiii–xxv). Brill.
Kühl, H. S., Kalan, A. K., Arandjelovic, M., Aubert, F., D’Auvergne, L., Goedmakers, A., Jones, S., Kehoe, L., Regnaut, S., Tickle, A., Ton, E., van Schijndel, J., Abwe, E. E., Angedakin, S., Agbor, A., Ayimisin, E. A., Bailey, E., Bessone, M., Bonnet, M. ... Boesch, C. (2016). Chimpanzee accumulative stone throwing. Scientific Reports, 6, 22219. https://doi.org/10.1038/srep22219
Kuhn, S. L. (2014). Signaling theory and technologies of communication in the Paleolithic. Biological Theory, 9(1), 42–50. https://doi.org/10.1007/s13752-013-0156-5
Kuhn, S. L., & Stiner, M. C. (2007). Body ornamentation as information technology: Towards an understanding of the significance of early beads. In P. Mellars, K. Boyle, O. Bar-Yosef, & C. B. Stringer (Eds.), Rethinking the human revolution: New behavioural and biological perspectives on the origin and dispersal of modern humans (pp. 45–54). Oxbow.
Kuzawa, C. W., Chugani, H. T., Grossman, L. I., Lipovich, L., Muzik, O., Hof, P. R., & Lange, N. (2014). Metabolic costs and evolutionary implications of human brain development. Proceedings of the National Academy of Sciences of the USA, 111(36), 13010–13015. https://doi.org/10.1073/pnas.1323099111
Lachmann, M., & Bergstrom, C. T. (2004). The disadvantage of combinatorial communication. Proceedings of the Royal Society B, 271(1555), 2337–2343.
Lang, M., Bahna, V., Shaver, J. H., Reddish, P., & Xygalatas, D. (2017). Sync to link: Endorphin-mediated synchrony effects on cooperation. Biological Psychology, 127, 191–197.
Lang, M., Krátký, J., Shaver, J. H., Jerotijević, D., & Xygalatas, D. (2019). Is ritual behavior a response to anxiety? In D. J. Slone & W. W. McCorkle (Eds.), The cognitive science of religion: A methodological introduction to key empirical studies (pp. 181–192). Bloomsbury Academic.
Lang, M., Krátký, J., & Xygalatas, D. (2020). The role of ritual behaviour in anxiety reduction: An investigation of Marathi religious practices in Mauritius. Philosophical Transactions of the Royal Society B, 375(1805), 20190431. https://doi.org/10.1098/rstb.2019.0431
Larrasoaña, J. C., Roberts, A. P., & Rohling, E. J. (2013). Dynamics of green Sahara periods and their role in hominin evolution. PLoS ONE, 8(10), e76514. https://doi.org/10.1371/journal.pone.0076514
Launay, J., Tarr, B., Dunbar, R. I. M., & Bshary, R. (2016). Synchrony as an adaptive mechanism for large-scale human social bonding. Ethology, 122(10), 779–789.
Leakey, L. S. B. (1943). The industries of the Gorgora Rockshelter, Lake Tana. Journal of the East African and Uganda Natural History Society, 17, 199–203.
Leakey, L. S. B. (1958). Recent discoveries at Oldovai Gorge, Tanganyika. Nature, 181, 1099–1103.
Legare, C. H., & Nielsen, M. (2020). Ritual explained: Interdisciplinary answers to Tinbergen’s four questions. Philosophical Transactions of the Royal Society B, 375(1805), 20190419. https://doi.org/10.1098/rstb.2019.0419
Legare, C. H., & Souza, A. L. (2012). Evaluating ritual efficacy: Evidence from the supernatural. Cognition, 124, 1–15.
Legare, C. H., & Watson-Jones, R. E. (2016). The evolution and ontogeny of ritual. In D. M. Buss (Ed.), The handbook of evolutionary psychology: Integrations (2nd ed., Vol. 2, pp. 829–847). Wiley.
Lewis, J. (2015). Where goods are free but knowledge costs: Hunter-gatherer ritual economics in western Central Africa. Hunter Gatherer Research, 1(1), 1–27. https://doi.org/10.3828/hgr.2015.2
Lindsey, D. T., Brown, A. M., Reijnen, E., Rich, A. N., Kuzmova, Y. I., & Wolfe, J. M. (2010). Color channels, not color appearance or color categories, guide visual search for desaturated color targets. Psychological Science, 21(9), 1208–1214.
Lisiecki, L. E., & Raymo, M. E. (2005). A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography, 20, PA1003. https://doi.org/10.1029/2004PA001071
Liu, W., Martinón-Torres, M., Cai, Y., Xing, S., Tong, H., Pei, S., & Wu, X.-J. (2015). The earliest unequivocally modern humans in southern China. Nature, 526, 696–699. https://doi.org/10.1038/nature15696
Lombard, M. (2006a). Direct evidence for the use of ochre in the hafting technology of Middle Stone Age tools from Sibudu Cave. Southern African Humanities, 18(1), 57–67.
Lombard, M. (2006b). First impressions of the functions and hafting technology of Still Bay pointed artefacts from Sibudu Cave. Southern African Humanities, 18(1), 27–41.
Lombard, M. (2007). The gripping nature of ochre: The association of ochre with Howiesons Poort adhesives and Later Stone Age mastics from South Africa. Journal of Human Evolution, 53, 406–419.
Lombard, M. (2008). Finding resolution for the Howiesons Poort through the microscope: Micro-residue analysis of segments from Sibudu Cave, South Africa. Journal of Archaeological Science, 35, 26–41.
Lombard, M. (2012). Thinking through the Middle Stone Age of sub-Saharan Africa. Quaternary International, 270, 140–155.
Louw, A. W. (1969). Bushman Rock Shelter, Ohrigstad, eastern Transvaal: A preliminary investigation, 1965. The South African Archaeological Bulletin, 24(94), 39–51.
Luncz, L. V., & Boesch, C. (2014). Tradition over trend: Neighboring chimpanzee communities maintain differences in cultural behavior despite frequent immigration of adult females. American Journal of Primatology, 76(7), 649–657. https://doi.org/10.1002/ajp.22259
Lydall, J., & Strecker, I. (1979a). The Hamar of southern Ethiopia: I. Work journal. Arbeiten aus dem Institut für Völkerkunde der Universität Göttingen, Vol. 12. Klaus Renner Verlag.
Lydall, J., & Strecker, I. (1979b). The Hamar of Southern Ethiopia: II. Baldambe Explains. Arbeiten aus dem Institut für Völkerkunde der Universität Göttingen, Vol. 13. Klaus Renner Verlag.
Lydall, J., & Strecker, I. (1979c). The Hamar of Southern Ethiopia: III. Conservations in Dambaiti. Arbeiten aus dem Institut für Völkerkunde der Universität Göttingen, Vol. 14. Klaus Renner Verlag.
Lyman, R. L. (2003). The influence of time averaging and space averaging on the application of foraging theory in zooarchaeology. Journal of Archaeological Science, 30(5), 595–610.
Lyn, H., & Chenkin, B. (2018). Kanzi. In J. Vonk & T. K. Shackelford (Eds.), Encyclopedia of animal cognition and behavior. Springer. https://doi.org/10.1007/978-3-319-47829-6_1043-1
Lyons, D. E., Young, A. G., & Keil, F. C. (2007). The hidden structure of overimitation. Proceedings of the National Academy of Sciences of the USA, 104(50), 19751–19756. https://doi.org/10.1073/pnas.0704452104
Mackay, A. (2006). A characterization of the MSA stone artefact assemblage from the 1984 excavations at Klein Kliphuis, Western Cape. The South African Archaeological Bulletin, 61(184), 181–188.
Mackay, A. (2011). The Late Pleistocene archaeology of Klein Kliphuis rock shelter, Western Cape, South Africa: 2006 excavation. The South African Archaeological Bulletin, 65(192), 132–147.
Mackay, A., Sumner, A., Jacobs, Z., Marwick, B., Bluff, K., & Shaw, M. (2014). Putslaagte 1 (PL1), the Doring River, and the later Middle Stone Age in southern Africa’s winter rainfall zone. Quaternary International, 350, 43–58. https://doi.org/10.1016/j.quaint.2014.05.007
Mackay, A., & Welz, A. (2008). Engraved ochre from a Middle Stone Age context at Klein Kliphuis in the Western Cape of South Africa. Journal of Archaeological Science, 35, 1521–1532.
Maier, M. A., Hill, R. A., Elliot, A. J., & Barton, R. A. (2015). Color in achievement contexts in humans. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 568–584). Cambridge University Press.
Malan, B. D. (1955). The archaeology of the Tunnel Cave and Skildergat Kop, Fish Hoek, Cape of Good Hope. The South African Archaeological Bulletin, 10(37), 3–9.
Malinowski, B. (1948). Magic, science and religion and other essays: Selected, and with an introduction by Robert Redfield. The Free Press of Glencoe.
Marean, C. W. (2010). Pinnacle Point Cave 13B (Western Cape Province, South Africa) in context: The Cape Floral Kingdom, shellfish, and modern human origins. Journal of Human Evolution, 59(3–4), 425–443.
Marean, C. W. (2014). The origins and significance of coastal resource use in Africa and western Eurasia. Journal of Human Evolution, 77, 17–40.
Marean, C. W., Bar-Matthews, M., Bernatchez, J., Fisher, E., Goldberg, P., Herries, A. I. R., Jacobs, Z., Jerardino, A., Karkanas, P., Minichillo, T., Nilssen, P. J., Thompson, E., Watts, I., & Williams, H. M. (2007). Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature, 449, 905–908.
Märker, M., Schmidt, P., Hochschild, V., & Kanaeva, Z. (2011). Spatial assessment of early human expansions using GIS and database techniques: Examples from Southern Africa. In R. Macchiarelli & G.-C. Weniger (Eds.), Wissenschaftliche Schriften des Neanderthal Museums: Pleistocene databases: Acquisition, storing, sharing (Vol. 4, pp. 69–75). Neanderthal Museum.
Marks, A. E. (1968). The Khormusan: An Upper Pleistocene industry in Sudanese Nubia. In F. Wendorf (Ed.), The prehistory of Nubia (pp. 315–391). Fort Burgwin Research Center and Southern Methodist University Press.
Marshall, L. (1999). Nyae Nyae !Kung beliefs and rites. Peabody Museum monographs (Vol. 8). Peabody Museum of Archaeology and Ethnology, Harvard University.
Mason, R. J. (1962). Prehistory of the Transvaal: A record of human activity. Witwatersrand University Press.
Matsuzawa, T. (2011). Log doll: Pretence in wild chimpanzees. In T. Matsuzawa, T. Humle, & Y. Sugiyama (Eds.), Primatology monographs. The chimpanzees of Bossou and Nimba (pp. 131–135). Springer.
Maynard-Smith, J., & Harper, D. (2003). Animal signals. Oxford University Press.
Mazel, A. D. (1996). Maqonqo shelter: The excavation of Holocene deposits in the eastern Biggarsberg, Thukela Basin, South Africa. Natal Museum Journal of Humanities, 8, 1–39.
McBrearty, S., & Brooks, A. (2000). The revolution that wasn’t: A new interpretation of the origin of modern human behavior. Journal of Human Evolution, 39, 453–563.
McBurney, C. B. M. (1967). The Haua Fteah (Cyrenaica) and the Stone Age of the South-East Mediterranean. Cambridge University Press.
McCauley, R. N., & Lawson, E. T. (2002). Bringing ritual to mind: Psychological foundations of cultural forms. Cambridge University Press.
McDougall, I., Brown, F. H., & Fleagle, J. C. (2005). Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature, 433, 733–736.
McGrew, W. C. (2004). The cultured chimpanzee: Reflections on cultural primatology. Cambridge University Press.
McGrew, W. C. (2017). Grooming hand clasp. In A. Fuentes (Ed.), The international encyclopedia of primatology: Volume I: A-G (pp. 486–488). Wiley Blackwell.
Mehlman, M. J. (1979). Mumba-Höhle revisited: The relevance of a forgotten excavation to some current issues in East African prehistory. World Archaeology, 11(1), 80–94.
Mehlman, M. J. (1989). Late Quaternary archaeological sequences in northern Tanzania. Doctoral dissertation, University of Illinois.
Mehta, P. H., Jones, A. C., & Josephs, R. A. (2008). The social endocrinology of dominance: Basal testosterone predicts cortisol changes and behavior following victory and defeat. Journal of Personality and Social Psychology, 94(6), 1078–1093. https://doi.org/10.1037/0022-3514.94.6.1078
Mercader, J. (2002). Forest people: The role of African rainforests in human evolution and dispersal. Evolutionary Anthropology, 11, 117–124.
Mercader, J., Asmerom, Y., Bennett, T., Raja, M., & Skinner, A. (2009). Initial excavation and dating of Ngalue Cave: A Middle Stone Age site along the Niassa Rift Mozambique. Journal of Human Evolution, 57(1), 63–74. https://doi.org/10.1016/j.jhevol.2009.03.005
Mitchell, P. J. (1994). Understanding the MSA/LSA transition: The pre-20,000 BP assemblages from new excavations at Sehonghong rock-shelter. Southern African Field Archaeology, 3, 15–25.
Mitchell, P. J. (1995). Revisiting the Robberg: New results and a revision of old ideas at Sehonghong Rock Shelter Lesotho. The South African Archaeological Bulletin, 50(161), 28–38.
Mitchell, P. J., & Steinberg, J. M. (1992). Ntloana Tsoana: A Middle Stone Age sequence from western Lesotho. The South African Archaeological Bulletin, 47(155), 26–33.
Mithen, S. J. (2014). The cognition of Homo neanderthalensis and H. sapiens: Does the use of pigment necessarily imply symbolic thought? In T. Akazawa, Y. Nishiaki, & K. Aoki (Eds.), Dynamics of learning in Neanderthals and modern humans. Volume 2: Cognitive and physical perspectives (pp. 7–16). Springer.
Mogan, R., Fischer, R., & Bulbulia, J. A. (2017). To be in synchrony or not? A meta-analysis of synchrony’s effects on behavior, perception, cognition and affect. Journal of Experimental Social Psychology, 72, 13–20. https://doi.org/10.1016/j.jesp.2017.03.009
Morel, J. (1974). La station éponyme de l’Oued Djebbana à Bir-el-Ater (est algérien): Contribution à la connaissance de son industrie et de sa faune. L’anthropologie, 78(1), 53–80.
Murray-Wallace, C. V., Richter, J., & Vogelsang, R. (2015). Aminostratigraphy and taphonomy of ostrich eggshell in the sedimentary infill of Apollo 11 Rockshelter Namibia. Journal of Archaeological Science: Reports, 4(4951), 143–151. https://doi.org/10.1016/j.jasrep.2015.09.006
Muthukrishna, M., Doebeli, M., Chudek, M., & Henrich, J. (2018). The cultural brain hypothesis: How culture drives brain expansion, sociality, and life history. PLoS Computational Biology, 14(11), e1006504. https://doi.org/10.1371/journal.pcbi.1006504
Muthukrishna, M., Shulman, B. W., Vasilescu, V., & Henrich, J. (2014). Sociality influences cultural complexity. Proceedings of the Royal Society B, 281, 20132511. https://doi.org/10.1098/rspb.2013.2511
Nami, M., & Moser, J. (2010). La Grotte d'Ifri n'Ammar. Tome 2: Le Paléolithique moyen. Forschungen zur Archäologie Außereuropäischer Kulturen (Vol. 9). Reichert.
Nash, D. J., Coulson, S., Staurset, S., Ullyott, J. S., Babutsi, M., Hopkinson, L., & Smith, M. P. (2013). Provenancing of silcrete raw materials indicates long-distance transport to Tsodilo Hills, Botswana, during the Middle Stone Age. Journal of Human Evolution, 64(4), 280–288.
Nash, D. J., Coulson, S., Staurset, S., Ullyott, J. S., Babutsi, M., & Smith, M. P. (2016). Going the distance: Mapping mobility in the Kalahari Desert during the Middle Stone Age through multi-site geochemical provenancing of silcrete artefacts. Journal of Human Evolution, 96, 113–133. https://doi.org/10.1016/j.jhevol.2016.05.004
Nash, D. J., & Meadows, M. E. (2012). Africa. In D. J. Nash & S. E. Metcalfe (Eds.), Quaternary environmental change in the Tropics (pp. 79–150). Wiley.
Nespoulet, R., Debénath, A., El Hajraoui, M. A., Michel, P., Campmas, E., Oujaa, A., Ben-Ncer, A., Lacombe, J., Amani, F., Stoetzel, E., Boudad, L. (2008b). Le contexte archéologique des restes humains Atériens de la région de Rabat-Témara (Maroc): Apport des fouilles des Grottes d’el Mnasra et d’el Harhoura 2. In H. Aouraghe, H. Haddoumi, & K. El Hammouti (Eds.), Le quaternaire marocain dans son contexte méditerranéen: Actes de la quatrième rencontre des quaternaristes marocains (rqm4), Oujda, 15–17 novembre 2007. Volume 2: Paléontologie, taphonomie et préhistoire (pp. 356–375).
Nespoulet, R., El Hajraoui, M. A., Amani, F., Ben Ncer, A., Debénath, A., El Idrissi, A., Lacombe, J.-P., Michel, P., Oujaa, A., & Stoetzel, E. (2008a). Palaeolithic and Neolithic occupations in the Témara region (Rabat, Morocco): Recent data on hominin contexts and behavior. African Archaeological Review, 25(1–2), 21–39. https://doi.org/10.1007/s10437-008-9025-1
Neubauer, S., Hublin, J.-J., & Gunz, P. (2018). The evolution of modern human brain shape. Science Advances, 4(1), eaao5961. https://doi.org/10.1126/sciadv.aao5961
Nielsen, M., Kapitány, R., & Elkins, R. (2015). The perpetuation of ritualistic actions as revealed by young children’s transmission of normative behavior. Evolution and Human Behavior, 36(3), 191–198. https://doi.org/10.1016/j.evolhumbehav.2014.11.002
Nielsen, M., & Tomaselli, K. (2010). Overimitation in Kalahari Bushman children and the origins of human cultural cognition. Psychological Science, 21(5), 729–736. https://doi.org/10.1177/0956797610368808
Nielsen, M., Tomaselli, K., & Kapitány, R. (2018). The influence of goal demotion on children’s reproduction of ritual behavior. Evolution and Human Behavior, 39(3), 343–348. https://doi.org/10.1016/j.evolhumbehav.2018.02.006
Nowell, A. (2010). Defining behavioral modernity in the context of Neandertal and Anatomically Modern Human populations. Annual Review of Anthropology, 39(1), 437–452.
Oakley, K. P. (1981). The emergence of higher thought 3.0–0.2 Ma BP. Philosophical Transactions of the Royal Society B, 292, 205–211.
O’Hanlon, M. (1989). Reading the skin: Adornment, display, and society among the Wahgi. British Museum Publications.
Over, H., & Carpenter, M. (2012). Putting the social into social learning: Explaining both selectivity and fidelity in children’s copying behavior. Journal of Comparative Psychology, 126(2), 182–192.
Partridge, T. C. (1964). A Middle Stone Age and Iron Age site at Waterval, North West of Johannesburg. The South African Archaeological Bulletin, 19(76), 102–110.
Pazda, A. D., & Greitemeyer, T. (2015). Color in romantic contexts in humans. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 531–545). Cambridge University Press.
Peabody, C. (1927). Red paint. Journal de la Société des Américanistes de Paris, 19, 207–244.
Peirce, C. S. (1998). The essential Peirce: Selected philosophical writings. Indiana University Press.
Pepperberg, I. M. (2012). Symbolic communication in the grey parrot. In J. Vonk & T. K. Shackelford (Eds.), The Oxford handbook of comparative evolutionary psychology. Oxford University Press. https://doi.org/10.1093/oxfordhb/9780199738182.013.0016
Petersdorf, M., Dubuc, C., Georgiev, A. V., Winters, S., & Higham, J. P. (2017). Is male rhesus macaque facial coloration under intrasexual selection? Behavioral Ecology, 28(6), 1472–1481. https://doi.org/10.1093/beheco/arx110
Pettitt, P. (2011a). The Palaeolithic origins of human burial. Routledge.
Pettitt, P. (2011b). The living as symbols, the dead as symbols: Problematising the scale and pace of hominin symbolic evolution. In C. S. Henshilwood & F. d’Errico (Eds.), Homo symbolicus: The dawn of language, imagination and spirituality (pp. 141–162). John Benjamins.
Pinker, S. (1994). The language instinct. William Morrow.
Pinker, S. (2016). The false allure of group selection. In D. M. Buss (Ed.), The handbook of evolutionary psychology Volume 2. Integrations (2nd ed., pp. 867–884). Wiley.
Pollick, A. S., & de Waal, F. B. M. (2007). Ape gestures and language evolution. Proceedings of the National Academy of Sciences of the USA, 104(19), 8184–8189. https://doi.org/10.1073/pnas.0702624104
Pomerleau, V. J., Fortier-Gauthier, U., Corriveau, I., Dell’Acqua, R., & Jolicoeur, P. (2014). Colour-specific differences in attentional deployment for equiluminant pop-out colours: Evidence from lateralised potentials. International Journal of Psychophysiology, 91(3), 194–205.
Porraz, G., Val, A., Tribolo, C., Mercier, N., de la Peña, P., Haaland, M. M., Igreja, M., Miller, C. E., & Schmid, V. C. (2018). The MIS5 Pietersburg at ‘28’ Bushman Rock Shelter, Limpopo Province, South Africa. PLoS ONE, 13(10), e0202853. https://doi.org/10.1371/journal.pone.0202853
Powell, A., Shennan, S. J., & Thomas, M. G. (2009). Late Pleistocene demography and the appearance of modern human behavior. Science, 324, 1298–1301. https://doi.org/10.1126/science.1170165
Power, C. (2009). Sexual selection models for the emergence of symbolic communication: Why they should be reversed. In R. Botha & C. Knight (Eds.), Oxford studies in the evolution of language: The cradle of language (Vol. 12, pp. 257–280). Oxford University Press.
Power, C. (2010). Cosmetics, identity and consciousness. Journal of Consciousness Studies, 17(7–8), 73–94.
Power, C. (2014). The evolution of ritual as a process of sexual selection. In D. Dor, C. Knight, & J. Lewis (Eds.), Oxford studies in the evolution of language: The social origins of language (Vol. 19, pp. 196–207). Oxford University Press.
Power, C., Sommer, V., & Watts, I. (2013). The seasonality thermostat: Female reproductive synchrony and male behavior in monkeys, Neanderthals, and modern humans. PaleoAnthropology, 2013, 33–60.
Prieur, J., Barbu, S., Blois-Heulin, C., & Lemasson, A. (2020). The origins of gestures and language: History, current advances and proposed theories. Biological Reviews of the Cambridge Philosophical Society, 95(3), 531–554. https://doi.org/10.1111/brv.12576
Pruetz, J. D., & LaDuke, T. C. (2010). Reaction to fire by savanna chimpanzees (Pan troglodytes verus) at Fongoli, Senegal: Conceptualization of ‘fire behavior’ and the case for a chimpanzee model. American Journal of Physical Anthropology, 141(4), 646–650.
Purzycki, B. G., & Arakchaa, T. (2013). Ritual behavior and trust in the Tyva Republic. Current Anthropology, 54(3), 381–388.
R Core Team (2021). R: A language and environment for statistical computing (4.0.5) [Computer software]. R Foundation for Statistical Computing. https://www.r-project.org/
Radcliffe-Brown, A. R. (1922). The Andaman Islanders: A study in social anthropology. Cambridge University Press.
Rappaport, R. A. (1999). Ritual and religion in the making of humanity. Cambridge University Press.
Reddish, P., Fischer, R., Bulbulia, J. A., & Szolnoki, A. (2013). Let’s dance together: Synchrony, shared intentionality and cooperation. PLoS ONE, 8(8), e71182. https://doi.org/10.1371/journal.pone.0071182
Regan, B. C., Julliot, C., Simmen, B., Vienot, F., Charles-Dominique, P., & Mollon, J. D. (2001). Fruits, foliage and the evolution of primate colour vision. Philosophical Transactions of the Royal Society B, 356, 229–283.
Reznikova, Z. (2007). Dialog with black box: Using information theory to study animal language behaviour. Acta Ethologica, 10(1), 1–12. https://doi.org/10.1007/s10211-007-0026-x
Richerson, P., & Henrich, J. (2012). Tribal social instincts and the cultural evolution of institutions to solve collective action problems. Cliodynamics. The Journal of Quantitative History and Cultural Evolution, 3(1), 38–80.
Richerson, P. J., Baldini, R., Bell, A. V., Demps, K., Frost, K., Hillis, V., Mathew, S., Newton, E. K., Naar, N., Newson, L., Ross, C., Smaldino, P. E., Waring, T. M., & Zefferman, M. (2016). Cultural group selection plays an essential role in explaining human cooperation: A sketch of the evidence. Behavioral and Brain Sciences, 39, e30. https://doi.org/10.1017/S0140525X1400106X
Richter, D., Grün, R., Joannes-Boyau, R., Steele, T. E., Amani, F., Rué, M., & McPherron, S. J. P. (2017). The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature, 546, 293–296.
Rifkin, R. F. (2011). Assessing the efficacy of red ochre as a prehistoric hide-tanning ingredient. Journal of African Archaeology, 9(2), 131–158.
Rifkin, R. F. (2012a). The symbolic and functional exploitation of ochre during the South African Middle Stone Age. Doctoral dissertation, University of the Witwatersrand.
Rifkin, R. F. (2012b). Processing ochre in the Middle Stone Age: Testing the inference of prehistoric behaviours from actualistically derived experimental data. Journal of Anthropological Archaeology, 31(2), 174–195.
Rifkin, R. F. (2015). Ethnographic and experimental perspectives on the efficacy of red ochre as a mosquito repellent. The South African Archaeological Bulletin, 70(201), 64–75.
Rifkin, R. F., Dayet, L., Queffelec, A., Summers, B., Lategan, M., d’Errico, F., & Petraglia, M. D. (2015a). Evaluating the photoprotective effects of ochre on human skin by in vivo SPF assessment: Implications for human evolution, adaptation and dispersal. PLoS ONE, 10(9), e0136090. https://doi.org/10.1371/journal.pone.0136090
Rifkin, R. F., d’Errico, F., Dayet-Boulliot, L., & Summers, B. (2015b). Assessing the photoprotective effects of red ochre on human skin by in vitro laboratory experiments South African. Journal of Science, 111(34), 1–8. https://doi.org/10.17159/sajs.2015/20140202
Rightmire, G. P. (2009). Middle and later Pleistocene hominins in Africa and Southwest Asia. Proceedings of the National Academy of Sciences of the USA, 106(38), 16046–16050. https://doi.org/10.1073/pnas.0903930106
Ritz, T., Meuret, A. E., & Ayala, E. S. (2010). The psychophysiology of blood-injection-injury phobia: Looking beyond the diphasic response paradigm. International Journal of Psychophysiology, 78(1), 50–67. https://doi.org/10.1016/j.ijpsycho.2010.05.007
Robbins, L. H., Brook, G. A., Murphy, M. L., Campbell, A. C., Melear, N., & Downey, W. S. (2000). Late Quaternary archaeological and palaeoenvironmental data from sediments at Rhino Cave, Tsodilo Hills, Botswana. South African Field Archaeology, 9, 17–31.
Robbins, L. H., Murphy, M. L., Campbell, A. C., & Brook, G. A. (1996). Excavations at the Tsodilo Hills Rhino Cave. Botswana Notes and Records, 28, 23–45.
Robinson, E., Zahid, H. J., Codding, B. F., Haas, R., & Kelly, R. L. (2019). Spatiotemporal dynamics of prehistoric human population growth: Radiocarbon ‘dates as data’ and population ecology models. Journal of Archaeological Science, 101, 63–71.
Roebroeks, W., & Villa, P. (2011). On the earliest evidence for habitual use of fire in Europe. Proceedings of the National Academy of Sciences of the USA, 108(13), 5209–5214.
Roffman, I., Savage-Rumbaugh, S., Rubert-Pugh, E., Stadler, A., Ronen, A., & Nevo, E. (2015). Preparation and use of varied natural tools for extractive foraging by bonobos (Pan Paniscus). American Journal of Physical Anthropology, 158(1), 78–91. https://doi.org/10.1002/ajpa.22778
Roksandic, M., Radović, P., & Lindal, J. (2018). Revising the hypodigm of Homo heidelbergensis: A view from the Eastern Mediterranean. Quaternary International., 466/A, 66–81.
Rossano, M. J. (2012). The essential role of ritual in the transmission and reinforcement of social norms. Psychological Bulletin, 138(3), 529–549.
Rossano, M. J. (2015). The evolutionary emergence of costly rituals. PaleoAnthropology, 2015, 78–100.
Rossano, M. J. (2016). The ritual origins of humanity. In G. Hartung & M. Herrgen (Eds.), Interdisziplinäre Anthropologie Jahrbuch 3/2015: Religion und Ritual (pp. 3–128). Springer Fachmedien.
Rosso, D. E., d’Errico, F., & Queffelec, A. (2017). Patterns of change and continuity in ochre use during the late Middle Stone Age of the Horn of Africa: The Porc-Epic Cave record. PLoS ONE, 12(5), e0177298. https://doi.org/10.1371/journal.pone.0177298
Rosso, D. E., d’Errico, F., & Zilhão, J. (2014). Stratigraphic and spatial distribution of ochre and ochre processing tools at Porc-Epic Cave, Dire Dawa, Ethiopia. Quaternary International, 343, 85–99. https://doi.org/10.1016/j.quaint.2013.10.019
Rosso, D. E., Martí, A. P., & d’Errico, F. (2016). Middle Stone Age ochre processing and behavioural complexity in the Horn of Africa: Evidence from Porc-Epic Cave, Dire Dawa. Ethiopia. Plos One, 11(11), e0164793. https://doi.org/10.1371/journal.pone.0164793
Rots, V., van Peer, P., & Vermeersch, P. M. (2011). Aspects of tool production, use, and hafting in Palaeolithic assemblages from Northeast Africa. Journal of Human Evolution, 60(5), 637–664. https://doi.org/10.1016/j.jhevol.2011.01.001
Rozin, P., & Todd, P. M. (2015). The evolutionary psychology of food intake and choice. In D. M. Buss (Ed.), The handbook of evolutionary psychology: Volume 2. Foundations (2nd ed., pp. 183–205). Wiley.
Ruckdeschel, P., Kohl, M., Stabla, T., & Camphausen, F. (2006). S4 classes for distributions. R News, 6(2), 2–6.
Rudner, I. (1982). Khoisan pigments and paints and their relationship to rock paintings. Annals of the South African Museum (Vol. 87). South African Museum.
Ruffle, B. J., & Sosis, R. (2007). Does it pay to pray? Costly ritual and cooperation. The B.E. Journal of Economic Analysis Policy, 7(1), 18. https://doi.org/10.2202/1935-1682.1629
Sadr, K. (2009). Marine shell dates and surface lithic assemblages on the west coast of South Africa. Journal of Archaeological Science, 36(12), 2713–2729.
Sagona, A. (Ed.). (1994). Bruising the red earth: Ochre mining and ritual in Aboriginal Tasmania. Melbourne University Press.
Salomon, H., Vignaud, C., Coquinot, Y., Beck, L., Stringer, C. B., Strivay, D., & d’Errico, F. (2012). Selection and heating of colouring materials in the Mousterian levels of Es-Skhul (c. 100 000 years BP, Mount Carmel, Israel). Archaeometry, 54(4), 698–722.
Sandgathe, D. M. (2017). Identifying and describing pattern and process in the evolution of hominin use of fire. Current Anthropology, 58(Supplement 16), S360–S370. https://doi.org/10.1086/69145
Scerri, E. (2018). The Stone Age archaeology of West Africa. In T. Spear (Ed.), Oxford research encyclopaedia of African history. Oxford University Press. https://doi.org/10.1093/acrefore/9780190277734.013.137
Scerri, E. M. L. (2017). The North African Middle Stone Age and its place in recent human evolution. Evolutionary Anthropology, 26(3), 119–135.
Scerri, E. M. L., Thomas, M. G., Manica, A., Gunz, P., Stock, J. T., Stringer, C. B., & Chikhi, L. (2018). Did our species evolve in subdivided populations across Africa, and why does it matter? Trends in Ecology & Evolution, 33(8), 582–594. https://doi.org/10.1016/j.tree.2018.05.005
Schjoedt, U., Sørensen, J., Nielbo, K. L., Xygalatas, D., Mitkidis, P., & Bulbulia, J. A. (2013). Cognitive resource depletion in religious interactions. Religion, Brain & Behavior, 3(1), 39–55. https://doi.org/10.1080/2153599X.2012.736714
Schlebusch, C. M., Malmström, H., Günther, T., Sjödin, P., Coutinho, A., Edlund, H., & Jakobsson, M. (2017). Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago. Science, 358, 652–655.
Scholz, C. A., Johnson, T. C., Cohen, A. S., King, J. W., Peck, J. A., Overpeck, J. T., Talbot, M. R., Brown, E. T., Kalindekafe, L., Amoako, P. Y. O., Lyons, R. P., Shanahan, T. M., Castañeda, I. S., Heil, C. W., Forman, S. L., McHargue, L. R., Beuning, K. R., Gomez, J., & Pierson, J. (2007). East African megadroughts between 135 and 75 thousand years ago and bearing on early-modern human origins. Proceedings of the National Academy of Sciences of the USA, 104(42), 16416–16421.
Schwarz, S., & Singer, M. (2013). Romantic red revisited: Red enhances men’s attraction to young, but not menopausal women. Journal of Experimental Social Psychology, 49(1), 161–164.
Sehasseh, E. M., Fernandez, P., Kuhn, S., Stiner, M., Mentzer, S., Colarossi, D., Clark, A., Lanoe, F., Pailes, M., Hoffmann, D., Benson, A., Rhodes, E., Benmansour, M., Laissaoui, A., Ziani, I., Vidal-Matutano, P., Morales, J., Djellal, Y., Longet, B., & Bouzouggar, A. (2021). Early Middle Stone Age personal ornaments from Bizmoune Cave, Essaouira, Morocco. Science Advances, 7(39), eabi8620. https://doi.org/10.1126/sciadv.abi8620
Setchell, J. M. (2015). Color in competition contexts in non-human animals. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 546–567). Cambridge University Press.
Setchell, J. M., & Dixson, A. F. (2001). Changes in the secondary sexual adornments of male mandrills (Mandrillus sphinx) are associated with gain and loss of alpha status. Hormones and Behavior, 39(3), 177–184.
Setchell, J. M., & Jean Wickings, E. (2005). Dominance, status signals and coloration in male mandrills (Mandrillus sphinx). Ethology, 111(1), 25–50. https://doi.org/10.1111/j.1439-0310.2004.01054.x
Shahack-Gross, R., Berna, F., Karkanas, P., Lemorini, C., Gopher, A., & Barkai, R. (2014). Evidence for the repeated use of a central hearth at Middle Pleistocene (300 ky ago) Qesem Cave, Israel. Journal of Archaeological Science, 44, 12–21.
Shaver, J. H., Power, E. A., Purzycki, B. G., Watts, J., Sear, R., Shenk, M. K., Sosis, R., & Bulbulia, J. A. (2020). Church attendance and alloparenting: An analysis of fertility, social support and child development among English mothers. Philosophical Transactions of the Royal Society B, 375(1805), 20190428. https://doi.org/10.1098/rstb.2019.0428
Shaver, J. H., Sibley, C. G., Sosis, R., Galbraith, D., & Bulbulia, J. (2019). Alloparenting and religious fertility: A test of the religious alloparenting hypothesis. Evolution and Human Behavior, 40(3), 315–324. https://doi.org/10.1016/j.evolhumbehav.2019.01.004
Shennan, S. J. (2001). Demography and cultural innovation: A model and its implications for the emergence of modern human culture. Cambridge Archaeological Journal, 11(1), 5–16. https://doi.org/10.1017/S0959774301000014
Shi, J., Zhang, C., & Jiang, F. (2015). Does red undermine individuals’ intellectual performance? A test in China. International Journal of Psychology, 50(1), 81–84.
Shimelmitz, R., Kuhn, S. L., Jelinek, A. J., Ronen, A., Clark, A. E., & Weinstein-Evron, M. (2014). ‘Fire at will’: The emergence of habitual fire use 350,000 years ago. Journal of Human Evolution, 77, 196–203. https://doi.org/10.1016/j.jhevol.2014.07.005
Singarayer, J. S., & Burrough, S. L. (2015). Interhemispheric dynamics of the African rainbelt during the late Quaternary. Quaternary Science Reviews, 124, 48–67. https://doi.org/10.1016/j.quascirev.2015.06.021
Singer, R., & Wymer, J. J. (1982). The Middle Stone Age at Klasies River Mouth in South Africa. University of Chicago Press.
Smuts, B. B., & Watanabe, J. M. (1990). Social relationships and ritualized greetings in adult male baboons (Papio cynocephalus anubis). International Journal of Primatology, 11(2), 147–172.
Sokolik, K., Magee, R. G., & Ivory, J. D. (2014). Red-hot and ice-cold web ads: The influence of web ads’ warm and cool colors on click-through rates. Journal of Interactive Advertising, 14(1), 31–37.
Solecki, R. S. (1982). A ritual Middle Palaeolithic deer burial at Nahr Ibrahim Cave, Lebanon. In R. Saidah (Ed.), Archéologie au Levant: Recueil à la mémoire de Roger Saidah (pp. 47–56). Maison de l’Orient.
Soler, M. (2012). Costly signaling, ritual and cooperation: Evidence from Candomblé, an Afro-Brazilian religion. Evolution and Human Behavior, 33(4), 346–356.
Soriano, S., Villa, P., & Wadley, L. (2009). Ochre for the toolmaker: Shaping the Still Bay points at Sibudu (KwaZulu-Natal, South Africa). Journal of African Archaeology, 7(1), 41–54.
Sosis, R. (2003). Why aren’t we all Hutterites? Costly signaling theory and religious behavior. Human Nature: An Interdisciplinary Biosocial Perspective, 14(2), 91–127.
Sosis, R. (2019). Do religions promote cooperation? Testing signaling theories of religion. In D. J. Slone & W. W. McCorkle Jr. (Eds.), Scientific studies of religion: Inquiry and explanation. The cognitive science of religion: A methodological introduction to key empirical studies (pp. 155–162). Bloomsbury Academic.
Sosis, R., & Alcorta, C. S. (2003). Signaling, solidarity, and the sacred: The evolution of religious behavior. Evolutionary Anthropology, 12, 264–274.
Sosis, R., & Bressler, E. (2003). Cooperation and commune longevity: A test of the costly signaling theory of religion. Cross-Cultural Research, 37(2), 211–239.
Sosis, R., Kress, H. C., & Boster, J. S. (2007). Scars for war: Evaluating alternative signaling explanations for cross-cultural variance in ritual costs. Evolution and Human Behavior, 28(4), 234–247. https://doi.org/10.1016/j.evolhumbehav.2007.02.007
Stafford, M. D., Frison, G. C., Stanford, D., & Zeimans, G. (2003). Digging for the color of life: Paleoindian red ochre mining at the Powars II site Platte County, Wyoming, USA. Geoarchaeology, 18(1), 71–90.
Stallmann, R. R., & Froehlich, J. W. (2000). Primate sexual swellings as coevolved signal systems. Primates, 41, 1–16.
Steele, T. E., Álvarez Fernández, E., & Hallett-Desguez, E. (2019). A review of shells as personal ornamentation during the African Middle Stone Age. PaleoAnthropology, 24, 24–51.
Steele, T. E., & Klein, R. G. (2009). Late Pleistocene subsistence strategies and resource intensification in Africa. In J.-J. Hublin & M. P. Richards (Eds.), The evolution of hominin diets: Integrating approaches to the study of palaeolithic subsistence (pp. 113–126). Springer.
Steele, T. E., Mackay, A., Orton, J., & Schwortz, S. (2012). Varsche Rivier 003, a new Middle Stone Age site in southern Namaqualand, South Africa. The South African Archaeological Bulletin, 67, 108–119.
Stephen, I. D., Coetzee, V., Law Smith, M., Perrett, D. I., & Sear, R. (2009a). Skin blood perfusion and oxygenation colour affect perceived human health. PLoS ONE, 4(4), e5083. https://doi.org/10.1371/journal.pone.0005083
Stephen, I. D., Law Smith, M. J., Stirrat, M. R., & Perrett, D. I. (2009b). Facial skin coloration affects perceived health of human faces. International Journal of Primatology, 30(6), 845–857.
Stephen, I. D., Oldham, F. H., Perrett, D. I., & Barton, R. A. (2012). Redness enhances perceived aggression, dominance and attractiveness in men’s faces. Evolutionary Psychology, 10(3), 562–572.
Stephen, I. D., & Perrett, D. I. (2015). Color and face perception. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 585–602). Cambridge University Press.
Sterelny, K. (2011). From hominins to humans: How sapiens became behaviourally modern. Philosophical Transactions of the Royal Society B, 366, 809–822.
Sterelny, K. (2014). A Paleolithic reciprocation crisis: Symbols, signals, and norms. Biological Theory, 9(1), 65–77.
Sterelny, K., & Hiscock, P. (2014). Symbols, signals, and the archaeological record. Biological Theory, 9(1), 1–3.
Stewart, F. A., Piel, A. K., & O’Malley, R. C. (2012). Responses of chimpanzees to a recently dead community member at Gombe National Park, Tanzania. American Journal of Primatology, 74, 1–7. https://doi.org/10.1002/ajp.20994
Stringer, C. B. (2016). The origin and evolution of Homo sapiens. Philosophical Transactions of the Royal Society B, 371, 20150237. https://doi.org/10.1098/rstb.2015.0237
Taçon, P. S. C. (2004). Ochre, clay, stone and art: The symbolic importance of minerals as life force among Aboriginal peoples of northern and central Australia. In N. Boivin & M. A. Owoc (Eds.), Soil, stones and symbols: Cultural perceptions of the mineral world (pp. 30–42)
Tanaka, A., & Tokuno, Y. (2011). The effect of the color red on avoidance motivation. Social Behavior and Personality, 39(2), 287–288.
Tarr, B., Launay, J., Cohen, E., & Dunbar, R. I. M. (2015). Synchrony and exertion during dance independently raise pain threshold and encourage social bonding. Biology Letters, 11, 20150767. https://doi.org/10.1098/rsbl.2015.0767
Tarr, B., Launay, J., & Dunbar, R. I. M. (2016). Silent disco: Dancing in synchrony leads to elevated pain thresholds and social closeness. Evolution and Human Behavior, 37(5), 343–349.
Tchernikov, I., & Fallah, M. (2010). A color hierarchy for automatic target selection. PLoS ONE, 5(2), e9338. https://doi.org/10.1371/journal.pone.0009338
Teleki, G. (1973). Group response to the accidental death of a chimpanzee in Gombe National Park Tanzania. Folia Primatologica, 20(2), 81–94. https://doi.org/10.1159/000155569
ten Velden, F. S., Baas, M., Shalvi, S., Preenen, P. T. Y., & de Dreu, C. K. W. (2012). In competitive interaction displays of red increase actors’ competitive approach and perceivers’ withdrawal. Journal of Experimental Social Psychology, 48(5), 1205–1208.
Tennie, C., & van Schaik, C. P. (2020). Spontaneous (minimal) ritual in non-human great apes? Philosophical Transactions of the Royal Society B, 375(1805), 20190423. https://doi.org/10.1098/rstb.2019.0423
Texier, P.-J., Porraz, G., Parkington, J., Rigaud, J.-P., Poggenpoel, C., Miller, C., Tribolo, C., Cartwright, C., Coudenneau, A., Klein, R., Steele, T., & Verna, C. (2010). A Howiesons Poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at Diepkloof Rock Shelter, South Africa. Proceedings of the National Academy of Sciences of the USA, 107(14), 6180–6185.
Texier, P.-J., Porraz, G., Parkington, J., Rigaud, J.-P., Poggenpoel, C., & Tribolo, C. (2013). The context, form and significance of the MSA engraved ostrich eggshell collection from Diepkloof Rock Shelter, Western Cape, South Africa. Journal of Archaeological Science, 40(9), 3412–3431.
Thackeray, A. I. (2000). Middle Stone Age artefacts from the 1993 and 1995 excavations of Die Kelders Cave 1. Journal of Human Evolution, 38, 147–168.
Thompson, E., Williams, H. M., & Minichillo, T. (2010). Middle and late Pleistocene Middle Stone Age lithic technology from Pinnacle Point 13B (Mossel Bay, Western Cape Province, South Africa). Journal of Human Evolution, 59(3–4), 358–377.
Tinbergen, N. (1952). ‘Derived’ activities: Their causation, biological significance, origin, and emancipation during evolution. The Quarterly Review of Biology, 27(1), 1–32.
Tjallingii, R., Claussen, M., Stuut, J.-B.W., Fohlmeister, J., Jahn, A., Bickert, T., & Röhl, U. (2008). Coherent high- and low-latitude control of the northwest African hydrological balance. Nature Geoscience, 1(10), 670–675. https://doi.org/10.1038/ngeo289
Tobias, P. V. (1949). The excavations of Mwulu’s Cave, Potgietersrust District. The South African Archaeological Bulletin, 4, 2–13.
Tomasello, M. (2014). The ultra-social animal. European Journal of Social Psychology, 44(3), 187–194. https://doi.org/10.1002/ejsp.2015
Tomasello, M., & Call, J. (2019). Thirty years of great ape gestures. Animal Cognition, 22(4), 461–469. https://doi.org/10.1007/s10071-018-1167-1
Tomasello, M., Carpenter, M., Call, J., Behne, T., & Moll, H. (2005). Understanding and sharing intentions: The origins of cultural cognition. Behavioral and Brain Sciences, 28(5), 675–691; discussion 691–735. https://doi.org/10.1017/S0140525X05000129
Tomasello, M., Melis, A. P., Tennie, C., Wyman, E., & Herrmann, E. (2012). Two key steps in the evolution of human cooperation. Current Anthropology, 53(6), 673–692.
Tönjes, H. (1911). Ovamboland: Land, Leute, Mission. Mit besonderer Berücksichtigung seines größten Stammes Onkuanjama. Martin Warneck.
Tryon, C. A., & Faith, J. T. (2013). Variability in the Middle Stone Age of eastern Africa. Current Anthropology, 54(S8), S234–S254.
Tryon, C. A., Lewis, J. E., Ranhorn, K. L., Kwekason, A., Alex, B., Laird, M. F., Marean, C. W., Niespolo, E., Nivens, J., & Mabulla, A. Z. P. (2018). Middle and Later Stone Age chronology of Kisese II rockshelter (UNESCO World Heritage Kondoa Rock-Art Sites), Tanzania. Plos One, 13(2), e0192029. https://doi.org/10.1371/journal.pone.0192029
Turner, T. S. (2012). The social skin (Reprint). Journal of Ethnographic Theory, 2(2), 486–504.
Tuzin, D. F. (2001). Social complexity in the making: A case study among the Arapesh of New Guinea. Routledge.
Tylén, K., Fusaroli, R., Rojo, S., Heimann, K., Fay, N., Johannsen, N. N., & Lombard, M. (2020). The evolution of early symbolic behavior in Homo sapiens. Proceedings of the National Academy of Sciences of the USA, 117(9), 4578–4584. https://doi.org/10.1073/pnas.1910880117
Underhill, D. (2012). The Fauresmith: The transition from the Earlier to Middle Stone Ages in northern South Africa. Doctoral dissertation, University of Southampton.
Van Leeuwen, E. J. C., Cronin, K. A., & Haun, D. B. M. (2014). A group-specific arbitrary tradition in chimpanzees (Pan troglodytes). Animal Cognition, 17(6), 1421–1425. https://doi.org/10.1007/s10071-014-0766-8
Van Leeuwen, E. J. C., Cronin, K. A., & Haun, D. B. M. (2017). Tool use for corpse cleaning in chimpanzees. Scientific Reports, 7, 44091. https://doi.org/10.1038/srep44091
Van Leeuwen, E. J. C., Cronin, K. A., Haun, D. B. M., Mundry, R., & Bodamer, M. D. (2012). Neighbouring chimpanzee communities show different preferences in social grooming behaviour. Proceedings of the Royal Society B, 279, 4362–4367. https://doi.org/10.1098/rspb.2012.1543
Van Leeuwen, E. J. C., Mulenga, I. C., Bodamer, M. D., & Cronin, K. A. (2016). Chimpanzees’ responses to the dead body of a 9-year-old group member. American Journal of Primatology, 78(9), 914–922. https://doi.org/10.1002/ajp.22560
Van Noten, F., Cornelissen, E., Gysels, J., Moeyersons, J., Nijs, K., & Uytterschaut, H. (1987). The Kapthurin project, Baringo: The 1984 season. Nyame Akuma, 28, 20–27.
Van Peer, P., Fullagar, R., Stokes, S., Bailey, R. M., Moeyersons, J., Steenhoudt, F., Geerts, A., Vanderbeken, T., De Dapper, M., & Geus, F. (2003). The Early to Middle Stone Age transition and the emergence of modern human behaviour at site 8-B-11, Sai Island Sudan. Journal of Human Evolution., 45(2), 187–193. https://doi.org/10.1016/S0047-2484(03)00103-9
Van Peer, P., Rots, V., & Vroomans, J.-M. (2004). A story of colourful diggers and grinders: The Sangoan and Lupemban at site 8-B-11, Sai Island, northern Sudan. Before Farming, 2004(3), 1–27.
Van Peer, P., Rots, V., & Vermeersch, P. M. (2008). A wasted effort at the quarry: Wear analysis and interpretation of an MSA lanceolate point from Taramsa-8, Egypt. PaleoAnthropology, 234–250.
Van Peer, P., & Vermeersch, P. M. (2000). The Nubian complex and the dispersal of modern humans in North Africa. In K. L. Kroeper & M. Kobusiewicz (Eds.), Studies in African Archaeology: Recent research into the Stone Age of northeastern Africa (Vol. 7, pp. 47–60). Poznan Archaeological Museum.
Vanhaeren, M., d’Errico, F., van Niekerk, K. L., Henshilwood, C. S., & Erasmus, R. M. (2013). Thinking strings: Additional evidence for personal ornament use in the Middle Stone Age at Blombos Cave, South Africa. Journal of Human Evolution, 64(6), 500–517.
Velliky, E. C. (2018). Ochre pigments and symbolic artefacts in the Swabian Jura: A study of the relationships between hominins, ochre materials, and early symbolic expression in the Middle and Upper Palaeolithic of Europe. Doctoral dissertation, University of Western Australia.
Velo, J. (1984). Ochre as medicine: A suggestion for the interpretation of the archaeological record. Current Anthropology, 25(5), 674.
Villa, P., Pollarolo, L., Degano, I., Birolo, L., Pasero, M., Biagioni, C., Douka, K., Vinciguerra, R., Lucejko, J. J., & Wadley, L. (2015). A milk and ochre paint mixture used 49,000 years ago at Sibudu South Africa. PLoS ONE, 10(6), e0131273. https://doi.org/10.1371/journal.pone.0131273
Villa, P., Soriano, S., Tsanova, T., Degano, I., Higham, T. F. G., d’Errico, F., Backwell, L., Lucejko, J. J., Colombini, M. P., & Beaumont, P. B. (2012). Border Cave and the beginning of the Later Stone Age in South Africa. Proceedings of the National Academy of Sciences of the USA, 109(33), 13208–13213. https://doi.org/10.1073/pnas.1202629109
Vogelsang, R. (1998). Middle-Stone-Age-Fundstellen in Südwest-Namibia. Africa Praehistorica (Vol. 11). Heinrich-Barth-Institut.
Volman, T. P. (1978). Early archeological evidence for shellfish collecting. Science, 201, 911–913.
Volman, T. P. (1981). The Middle Stone Age in the southern Cape. Doctoral dissertation, University of Chicago.
von Zimmermann, J., & Richardson, D. C. (2016). Verbal synchrony and action dynamics in large groups. Frontiers in Psychology, 7, 2034. https://doi.org/10.3389/fpsyg.2016.02034
Wadley, L. (2005b). Ochre crayons or waste products? Replications compared with MSA ‘crayons’ from Sibudu Cave, South Africa. Before Farming, 3, 1–12.
Wadley, L. (2005a). Putting ochre to the test: Replication studies of adhesives that may have been used for hafting tools in the Middle Stone Age. Journal of Human Evolution, 49, 587–601.
Wadley, L. (2006). Revisiting cultural modernity and the role of ochre in the Middle Stone Age. In H. Soodyall (Ed.), The prehistory of Africa: Tracing the lineage of modern man.
Wadley, L. (2010a). A taphonomic study of ochre demonstrates post-depositional color transformations. Journal of Taphonomy, 8, 243–254.
Wadley, L. (2010b). Cemented ash as a receptacle or work surface for ochre powder production at Sibudu, South Africa, 58,000 years ago. Journal of Archaeological Science, 37(10), 2397–2406. https://doi.org/10.1016/j.jas.2010.04.012
Wadley, L. (2010c). Compound-adhesive manufacture as a behavioral proxy for complex cognition in the middle stone age. Current Anthropology, 51(S1), S111–S119. https://doi.org/10.1086/649836
Wadley, L., Hodgskiss, T. P., & Grant, M. (2009). Implications for complex cognition from the hafting of tools with compound adhesives in the Middle Stone Age, South Africa. Proceedings of the National Academy of Sciences of the USA, 106, 9590–9594.
Wadley, L., & Langejans, G. H. J. (2014). preliminary study of scrapers around combustion features in layer SS, Sibudu, 58 000 years ago. The South African Archaeological Bulletin, 69(199), 19–33.
Wadley, L., Williamson, B., & Lombard, M. (2004). Ochre in hafting in middle stone age southern Africa: A practical role. Antiquity, 78(301), 661–675.
Waitt, C., Gerald, M. S., Little, A. C., & Kraiselburd, E. (2006). Selective attention toward female secondary sexual color in male rhesus macaques. American Journal of Primatology, 68(7), 738–744.
Waitt, C., Little, A. C., Wolfensohn, S., Honess, P., Brown, A. P., Buchanan-Smith, H. M., & Perrett, D. I. (2003). Evidence from rhesus macaques suggests that male coloration plays a role in female primate mate choice. Proceedings of the Royal Society B, 270(Suppl. 2), S144–S146. https://doi.org/10.1098/rsbl.2003.0065
Walker, N. J. (1987). The dating of Zimbabwean rock art. Rock Art Research, 4(2), 137–149.
Walker, N. J. (1995). Late Pleistocene and Holocene hunter-gatherers of the Matopos: An archaeological study of change and continuity in Zimbabwe. Studies in African Archaeology (Vol. 10). Societas Archaeologica Upsaliensis.
Watson-Jones, R. E., & Legare, C. H. (2016). The social functions of group rituals. Current Directions in Psychological Science, 25(1), 42–46.
Watts, I. (1998). The origin of symbolic culture: The Middle Stone Age of southern Africa and Khoisan ethnography. Doctoral dissertation, University of London.
Watts, I. (1999). The origin of symbolic culture. In R. I. M. Dunbar, C. Knight, & C. Power (Eds.), The evolution of culture: An interdisciplinary view (pp. 113–146). Rutgers University Press.
Watts, I. (2002). Ochre in the Middle Stone Age of southern Africa: Ritualized display or hide preservative? The South African Archaeological Bulletin, 57(175), 1–14.
Watts, I. (2009). Red ochre, body painting, and language: Interpreting the Blombos ochre. In R. Botha & C. Knight (Eds.), Oxford studies in the evolution of language: The cradle of language (Vol. 12, pp. 62–92). Oxford University Press.
Watts, I. (2010). The pigments from Pinnacle Point Cave 13B, Western Cape, South Africa. Journal of Human Evolution, 59(3–4), 392–411.
Watts, I. (2014). The red thread: Pigment use and the evolution of collective ritual. In D. Dor, C. Knight, & J. Lewis (Eds.), The social origins of language. Oxford studies in the evolution of language (Vol. 19, pp. 208–227). Oxford University Press.
Watts, I. (2015). Early color symbolism. In A. J. Elliot, M. D. Fairchild, & A. Franklin (Eds.), Handbook of color psychology (pp. 319–339). Cambridge University Press.
Watts, I., Chazan, M., & Wilkins, J. (2016). Early evidence for brilliant ritualized display: Specularite use in the northern Cape (South Africa) between ∼500 and ∼300 Ka. Current Anthropology, 57(3), 287–310. https://doi.org/10.1086/686484
Wen, N. J., Herrmann, P. A., & Legare, C. H. (2016). Ritual increases children’s affiliation with in-group members. Evolution and Human Behavior, 37(1), 54–60.
Wendorf, F., et al. (1993). E-87–5: Occupations dating to the grey phases. In F. Wendorf, R. Schild, & A. E. Close (Eds.), Egypt during the last interglacial: The middle Paleolithic of Bir Tarfawi and Bir Sahara East (pp. 345–355). Plenum Press.
Wendorf, F., & Close, A. E., et al. (1993). BS-11: A site in the recessional beach of West Lake 1. In F. Wendorf, R. Schild, & A. E. Close (Eds.), Egypt during the last interglacial: The middle Paleolithic of Bir Tarfawi and Bir Sahara East (pp. 488–511). Plenum Press.
Wendt, W. E. (1974). ‘Art mobilier’ aus der Apollo-11-Grotte in Südwestafrika. Acta Praehistorica et Archaeologica, 5(6), 1–42.
White, T. D., Asfaw, B., DeGusta, D., Gilbert, H., Richards, G. D., Suwa, G., & Howell, F. C. (2003). Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 742–747.
Whitehouse, H. (2004). Modes of religiosity: A cognitive theory of religious transmission. AltaMira Press.
Whitehouse, H. (2012). Ritual, cognition, and evolution. In R. Sun (Ed.), Grounding social sciences in cognitive sciences (pp. 265–284). MIT Press.
Whitehouse, H. (2013a). Explaining ritual. In G. W. Dawes & J. Maclaurin (Eds.), Routledge studies in religion: A new science of religion (Vol. 23, pp. 81–99). Routledge.
Whitehouse, H. (2013b). Religion, cohesion, and hostility. In S. Clarke, R. Powell, & J. Savulescu (Eds.), Religion, intolerance, and conflict: A scientific and conceptual investigation (pp. 36–47). Oxford University Press.
Whitehouse, H., & Lanman, J. A. (2014). The ties that bind us: Ritual, fusion, and identification. Current Anthropology, 55(6), 674–695.
Whiten, A., Goodall, J., McGrew, W. C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C. E. G., Wrangham, R. W., & Boesch, C. (1999). Cultures in chimpanzees. Nature, 399, 682–685. https://doi.org/10.1038/21415
Whitham, J. C., & Maestripieri, D. (2003). Primate rituals: The function of greetings between male Guinea baboons. Ethology, 109(10), 847–859. https://doi.org/10.1046/j.0179-1613.2003.00922.x
Wilkins, J., & Chazan, M. (2012). Blade production ~500 thousand years ago at Kathu Pan 1, South Africa: Support for a multiple origins hypothesis for early Middle Pleistocene blade technologies. Journal of Archaeological Science, 39, 1883–1900.
Wilkins, J., Schoville, B. J., Brown, K. S., & Chazan, M. (2012). Evidence for early hafted hunting technology. Science, 338, 942–946.
Will, M., Kandel, A. W., Kyriacou, K., & Conard, N. J. (2016). An evolutionary perspective on coastal adaptations by modern humans during the Middle Stone Age of Africa. Quaternary International, 404, 68–86. https://doi.org/10.1016/j.quaint.2015.10.021
Will, M., Parkington, J. E., Kandel, A. W., & Conard, N. J. (2013). Coastal adaptations and the Middle Stone Age lithic assemblages from Hoedjiespunt 1 in the Western Cape, South Africa. Journal of Human Evolution, 64(6), 518–537. https://doi.org/10.1016/j.jhevol.2013.02.012
Williams, L. A., Schofield, T. P., & Whitford, T. J. (2017). Romantic red: Testing the characteristics of color-attraction effects in a novel paradigm. Collabra Psychology, 3(1), 10. https://doi.org/10.1525/collabra.54
Wiltermuth, S. S., & Heath, C. (2009). Synchrony and cooperation. Psychological Science, 20(1), 1–5.
Wojcieszak, M., & Wadley, L. (2018). Raman spectroscopy and scanning electron microscopy confirm ochre residues on 71 000-year-old bifacial tools from Sibudu South Africa. Archaeometry, 60(5), 1062–1076. https://doi.org/10.1111/arcm.12369
Wu, Y., Lu, J., van Dijk, E., Li, H., & Schnall, S. (2018). The color red is implicitly associated with social status in the United Kingdom and China. Frontiers in Psychology, 9, 1902. https://doi.org/10.3389/fpsyg.2018.01902
Wurz, S. (2000). The Middle Stone Age at Klasies River. Doctoral dissertation. University of Stellenbosch.
Xygalatas, D. (2012). The burning saints: Cognition and culture in the firewalking rituals of the Anastenaria. Routledge.
Zahavi, A. (1980). Ritualization and the evolution of movement signals. Behaviour, 72(1/2), 77–81.
Zahavi, A., & Zahavi, A. (1997). The handicap principle: A missing piece of Darwin’s puzzle. Oxford University Press.
Zinner, D., Nunn, C. L., van Schaik, C. P., & Kappeler, P. M. (2004). Sexual selection and exaggerated sexual swellings of female primates. In P. M. Kappeler & C. van Schaik (Eds.), Sexual selection in primates: New and comparative perspectives (pp. 71–89). Cambridge University Press.
Zipkin, A. M., Wagner, M., McGrath, K., Brooks, A. S., & Lucas, P. W. (2014). An experimental study of hafting adhesives and the implications for compound tool technology. PLoS ONE, 9(11), e112560. https://doi.org/10.1371/journal.pone.0112560
Acknowledgements
We are thankful to our sponsors, the Gerda Henkel Foundation (RD), the Heidelberg University Graduate Academy (RD), and the Heidelberg Academy of Sciences and Humanities (ROCEEH). The following people contributed to the research: Zara Kanaeva programmed the time slice tool, formulated queries and maintained ROAD; Alex Mackay and Laure Dayet provided corrected and unpublished data. We thank Robert L. Kelly for reading earlier versions of this manuscript and his insightful criticism. We would also like to thank Sheila Coulson for her useful comments and literature recommendations. Most of all, we are grateful to the reviewers who provided detailed guidance on how to improve this manuscript. For any omissions or errors, the authors take full responsibility.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
10963_2022_9170_MOESM4_ESM.xlsx
All sites with lithics dating between 500 and 40 thousand years recorded in the ROAD database, listed by phase of ochre use
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dapschauskas, R., Göden, M.B., Sommer, C. et al. The Emergence of Habitual Ochre Use in Africa and its Significance for The Development of Ritual Behavior During The Middle Stone Age. J World Prehist 35, 233–319 (2022). https://doi.org/10.1007/s10963-022-09170-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10963-022-09170-2