
Abstract
The advances in bioprocess design, genetic engineering, and media optimization have enabled enhanced bacterial cellulose (BC) production and its application for diverse purposes. Following the requirements of a bioeconomy, numerous approaches were conducted to investigate alternative carbon or nitrogen sources from industrial by-products for BC biosynthesis. They can, however, not only affect the BC production but also its properties. Beet molasses, vinasse, and waste beer fermentation broth (WBFB) have thus been investigated in single and combined approaches for their BC production potential and effects on structural properties using Komagataeibacter xylinus DSM 2325. Therefore, the composition of each complex component was initially analyzed for total organic carbon (TOC), total bound nitrogen (TNb), sugars, organic acids, and alcohols. The polymer properties were characterized via gel permeation chromatography and X-ray diffraction. In dynamic shake flask cultivations, the exchange of Hestrin-Schramm (HS) medium components for a combination of all three complex substrates on a TOC- or TNb-based quantity resulted in the highest BC concentration (8.2 g L−1). Comparable concentrations were achieved when combining molasses and WBFB (8.1 g L−1). Each investigated complex component led to differing degrees of polymerization (DPn: 2751−4601) and BC crystallinities (26−58%) in comparison to HS medium. Beet molasses and vinasse were found to decrease the polymer crystallinity but induce higher DPn whereas the opposite occurred for WBFB. This study thus highlights beneficial effects of food industry by-products for BC biosynthesis and elucidates concomitantly occurring structural polymer alterations to enable further studies dealing with alternative substrates for structurally tailored BC production.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The production of cellulose with bacterial origin is long known and its advantages over plant cellulose in terms of purity, crystallinity, or degree of polymerization are acknowledged by its extensive research during the past two decades (Klemm et al. 2018). Hence, bacterial cellulose (BC) has evolved as high-tech biopolymer that finds wide applications such as in electrochemistry, medicine, or sustainable textile development, whereas especially the latter two reflect its biocompatibility and environmental friendliness (Lei et al. 2020; Provin et al. 2021; Choi et al. 2022). At least for comparison, the biosynthesis of BC is frequently conducted in the majorly glucose, yeast extract, and peptone containing Hestrin-Schramm medium (HS medium) to investigate new BC producing strains or cultivation conditions although further minimal and complex media were found suitable for this purpose (Hestrin and Schramm 1954; Sperotto et al. 2021). Following bioeconomic attempts to obtain a holistically sustainable material, numerous by-product streams primary from the food industry have been investigated as substrate for BC production since they are generally low-cost, readily available, and can thus provide the requirements for a commercially competitive biopolymer product—an ambition that dates back to the first scientific studies of Teodula K. Africa (1949) who replaced seasonal pineapple juice for year-round available coconut water as substrate for the nowadays popular BC-based Philippine delicacy nata de coco.
During the past three decades, molasses and corn steep liquor (CSL) represent some of the most often investigated alternative complex carbon and nitrogen sources, respectively, and have since then been part of several studies related to BC production (Toyosaki et al. 1995; Bae and Shoda 2004). The former can be derived from sugar cane or beets and both were successfully applied as carbon sources throughout different investigations leading to often increased BC production when pre-treated or diluted sufficiently (Sperotto et al. 2021). However, due to differing pre-treatments of the molasses and non-uniform characterization of the BC, it is not clear from the available studies, how and to which extent molasses as carbon source has an impact on the cellulose structure.
The investigations dealing with vinasse as complex medium component, another sugar industry-related but molasses fermentation-derived by-product, are only scarce and ambiguous for BC bioprocesses (Sperotto et al. 2021). Velásquez-Riaño et al. (2013) found an improved BC production under static but worsened under dynamic cultivation conditions which was attributed to the induced formation of non-producing strains due to the applied shear force during shaking. The obtained BC was not further characterized. Barshan et al. (2019) determined a decreased crystallinity and less BC in vinasse-based media which was explained by an unfavorable carbon/nitrogen ratio. This accentuates the importance of a sufficient complex component analysis prior to its application followed by systematic pre-treatments or supplementation with lacking components. This was, for example, thoroughly investigated by Ha et al. (2008) and Lin et al. (2014) for waste beer fermentation broth (WBFB) where the lack of carbon source was compensated by glucose addition or more intense pre-treatment methods to obtain sufficient amount of reducing sugars. Both studies demonstrated enhanced BC production and the structural analysis of Shezad et al. (2010) additionally suggested a positive effect of WBFB on cellulose crystallinity in fed-batch cultivation mode. Based on the promising results towards BC production of these previous studies and due to the local availability of certain types of by-product streams, we selected beet molasses, vinasse, and WBFB as carbon or nitrogen source for the following investigations.
However, the natural variability of complex sources from food industry between differing biological origins and the often scarcely characterized components make it difficult to directly compare the obtained results from previous studies. Thus, a more standardized approach was chosen to obtain comparable results between cultivations and to provide a basis for future studies. Total organic carbon (TOC) and total bound nitrogen (TNb) were selected as a measure to adjust the share of complex sources added to the cultivation medium. Hence, the amount of added complex organic carbon or bound nitrogen replaced the amount of carbonaceous or nitrogenous media components that would originally occur in HS medium. This allows, for instance, for the consideration of different organic acids in molasses which are known to positively effect BC production but are usually not considered when investigating molasses in terms of a sugar equivalent level. Since the structural properties of BC are also partially dependent on the production process and medium composition, the influential parameters need to be known precisely to obtain the desired product (Campano et al. 2016). For that reason, product characterization in terms of gel permeation chromatography (GPC) and X-ray diffraction (XRD) was carried out to survey the product properties derived from different complex resources. The crystallinity index determined according to Segal et al. (1959) via XRD measurement has often been regarded as a BC quality indicator although this method is known to overestimate the crystallinity of BC samples (Park et al. 2010a; French 2020). For that reason, the Segal and peak deconvolution method using a previously modelled amorphous cellulose diffraction pattern were applied for crystallinity determination of HS-type and by-product stream media-derived BC to give more reasonable and comparable results beyond this study (Yao et al. 2020). Differences in the degree of polymerization (DP) were additionally quantified via GPC measurement with multi angle laser light scattering (MALLS) which allowed a more detailed view in the BC formation process using state-of-the art protocols for molecular weight distribution (MWD) determination.
Hence, the objective of this study was to initially characterize the composition of beet molasses, vinasse, and WBFB and prepare comparable media for BC biosynthesis. Effects on the production process such as the formation of BC or accumulation of catabolic degradation products by Komagataeibacter xylinus DSM 2325 were determined. The structural product properties were then evaluated by GPC and XRD analysis and comprehensively discussed to obtain more detailed information on the biopolymer formation process and its influencing parameters.
Materials and methods
Materials
The chemicals for media preparation were obtained from Carl Roth (Karlsruhe, Germany) or Thermo Fisher Scientific (Waltham, USA) with purities ≥ 99.0%. Nitrogen sources for cultivation media were yeast extract (Carl Roth) and bacto peptone (Life Technologies, Carlsbad, USA). Molasses and vinasse utilized during this study were provided by Nordzucker (Braunschweig, Germany) and originate from sugar beet processing with molasses representing the remaining non-crystallizable sugar residue of the sucrose purification step and vinasse being the developing yeast fermentation broth during bioethanol production with molasses as main carbon source after the ethanol distillation. Waster beer fermentation broth was obtained from Hofbrauhaus Wolters (Braunschweig, Germany). The selected bacterial strain Komagataeibacter xylinus DSM 2325 was purchased from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany) as a freeze-dried sample and revitalized according to the enclosed protocol.
Pre-treatment and characterization of complex media
Prior to analysis and application in cultivation experiments, all complex media solutions were diluted with an equal volume of distilled water and centrifuged at 5346 × g for 20 min to remove suspended solids. Molasses and vinasse were additionally autoclaved at 121 °C for 15 min. Autoclaving the WBFB was avoided as it was described that intense heating or autoclaving at acidic conditions lowers its applicability for BC production (Ha et al. 2008). Since no remaining microbial activity was detected after 7 days incubation at 30 °C when WBFB was spread on HS medium agar plates, the WBFB was applied as obtained after its centrifugation. After the respective pre-treatment, all complex solutions were stored at 4 °C until further use. Total organic carbon (TOC, Spectroquant, TOC kit 50–800 mg L−1, Merck, Darmstadt, Germany) and total bound nitrogen (TNb, Nanocolor TNb 220 cuvette kit, Machery-Nagel, Düren, Germany) were quantified according to the cuvette test kit protocols. Finally, the pH of all complex solutions was measured at 20 °C (CG 843 pH meter, Schott, Mainz, Germany with BlueLine 16 electrode, Xylem Analytics Germany Sales, Weilheim, Germany).
Media composition
HS medium was prepared as described by Hestrin and Schramm (1954) with the main carbon source being autoclaved separately from the remaining components. The media composition of HS-type media solely differed in terms of the supplementation of ethanol (HS_EtOH) or exchange of glucose for sucrose (HS_S) from the original HS medium. The cultivation media with complex industrial by-products were prepared by adding sufficient volumes of molasses (M), vinasse (V) or WBFB to meet equal TOC or TNb values as the carbon (C) or nitrogen (N) source would have in HS medium. Complex HS-type media with a single complex component (SCC) were termed HS_M(C), HS_V(N), and HS_WBFB(N), whereas the media with combined complex components (CCC) were named HS_M(C)/V(N), HS_M(C)/WBFB(N), and HS_M(C)/V(N)/WBFB(N). The HS_M(C)/V(N)/WBFB(N) medium, for example, consisted of the TOC-equal replacement of glucose for molasses and the TNb-equal substitution of peptone and yeast extract for vinasse and WBFB on a 1:1-TNb-based ratio. The final compositions can be derived from Table 1. The pH of all media was adjusted to 5.0 with NaOH or HCl. Our previous investigations in HS medium revealed this pH with K. xylinus DSM 2325 under the chosen conditions as most suitable for BC production which is in agreement with its utilization in other studies (Gwon et al. 2019; Jang et al. 2019).
Preparation of stock cultures
Glycerol stock cultures of K. xylinus DSM 2325 were prepared from vital cells after 5 days of cultivation in 250 mL shake flasks without baffles at 30 °C and shaking frequency of 150 min−1 at a shaking amplitude of 50 mm. Developed BC agglomerates were dissected with sterile spatulas and the leaking bacterial suspension was thoroughly mixed with an equal volume of 60% v/v sterile glycerol solution. After an incubation time of 15 min, the resulting suspension was frozen in liquid nitrogen and stored at − 80 °C until further use.
Cultivation conditions and sampling procedure
Komagataeibacter xylinus DSM 2325 was revitalized by inoculating freshly thawed glycerol stock cultures in HS medium with an initial pH value of 5.0. After 5 days of cultivation, the formed BC agglomerate was dissected to inoculate pre-cultures using 1 L shake flasks without baffles and with a filling volume of 100 mL. Cultivation conditions were set as described for the preparation of glycerol stock cultures. After 7 days, BC agglomerates were dissected, and the leaked bacterial suspension was pooled to equally inoculate main cultures to an initial optical density of 0.05 at 600 nm while simultaneously avoiding the transfer of BC pieces. The cultivation was carried out in 250 mL shake flasks without baffles and 10% filling volume at the same conditions described earlier. Entire biological triplicates were harvested at four defined cultivation times for BC determination and the supernatant of each shake flask was used for further analysis. The mean value with the uncorrected standard deviation was calculated from the determined results. The starting condition represents the freshly inoculated medium. In addition to the dynamic BC production, a static cultivation was additionally conducted for 7 days under otherwise equal conditions to obtain reference material for GPC and XRD analysis.
Analysis of cultivation supernatants
Samples from main cultures were filtered using 0.2 µm filters made of regenerated cellulose (RC) and subjected to further analysis. At first, the pH was determined which was followed by freezing of the supernatants at − 20 °C until HPLC analysis. Sucrose, fructose, and glycerol concentration were measured using a Chromaster HPLC System (Hitachi, Tokyo, Japan) with a MetaCarb 87C column and guard column (300 × 7.8 mm and 50 × 4.6 mm, Agilent Technologies, Santa Clara, USA) using ultrapure water (Milli-Q®, Merck, Darmstadt, Germany) as mobile phase at 85 °C and a flow rate of 0.6 mL min−1. Eluents were detected using a refractive index (RI) detector (5450, Hitachi, Tokyo, Japan) and self-prepared standards served as reference for quantification. Due to the co-elution of gluconic acid and glucose under these conditions, the latter was quantified enzymatically (Biochemistry Analyzer YSI 2950D, YSI, Yellow Springs, USA) with appropriate sample dilution to at least 9 g L−1. Organic acids and ethanol in the supernatant were separated by using a LaChrom Elite HPLC System with an Aminex HPX-87H column and a micro-guard column (300 × 7.8 mm and 30 × 4.6 mm, Bio-Rad, Hercules, USA) applying 12 mM H2SO4 as mobile phase at a flow rate of 0.5 mL min−1 and 40 °C. The equipped RI detector (L-2490, Hitachi, Tokyo, Japan) served for quantification of citric acid, lactic acid, formic acid, acetic acid, and ethanol whereas a diode array detector (L-2455, Hitachi, Tokyo, Japan) at 210 nm allowed for gluconic acid and pyruvic acid quantification using self-prepared standards as reference.
Purification and determination of BC concentration
Harvested cellulose agglomerates were first disrupted and centrifuged at 7500 × g to separate the supernatant from the solids. This was followed by a washing step with distilled water and pelleting of the contained solids. The resulting pellets were suspended in 0.1 M NaOH solution and incubated at 90 °C for 70 min to lyse and detach the biomass from the cellulose (Mohite et al. 2012). After at least two more washing and centrifugation steps with distilled water, the bacterial cellulose was dried at 105 °C for at least 48 h and cooled in a desiccator for 30 min to gravimetrically determine its dry weight.
GPC analysis
Sample preparation for GPC analysis started with several washing and activation steps prior to the final dissolution in N,N-dimethylacetamide/lithium chloride (DMAc/LiCl) (9% w/v) and was realized according to the direct dissolution protocol proposed by Potthast et al. (2015) with some minor changes. Briefly, 20 mg dried and cut cellulose pieces were washed with distilled water multiple times and filtered off through a 0.45 µm RC filter. Subsequently, several solvent exchange steps were performed at about 22 °C starting with the incubation in acetone for 2 min which was then filtered off. This step was repeated and then equally performed with dimethyl sulfoxide (DMSO) followed by overnight incubation in DMSO at low agitation. Afterwards, solvent exchange with DMAc was carried out twice which was then once again followed by overnight incubation in DMAc at low agitation. After filtering off, the sample was kept for dissolution in DMAc/LiCl (9% w/v) at a final concentration of 10 mg cellulose per g solvent which took 2–10 days until no solids or turbidity were visible. Deviating from the original protocol and embedding the additional DMSO activation step as recommended by Siller et al. (2014) for hardly soluble rayon fibers was crucial for our BC to dissolve as otherwise the final dissolution step did not result clear cellulose solutions. For GPC measurements, the polymer solutions were further diluted with pure DMAc to a final DMAc/LiCl concentration of 0.9% w/v, equilibrated for one day at room temperature and filtered through a 0.2 µm PTFE syringe filter prior to injection. Polymer separation was then achieved with a guard and a set of three separation columns (PSS GRAM, 8 × 300 mm, 10 µm particle size, porosities of 30 Å, 100 Å, and 3000 Å) at 70 °C and a flow rate of 1 mL min−1. A PSS SLD 7100 served as MALLS detector (PSS, Mainz, Germany) using a refractive index increment of 0.136 mL g−1 which was derived from Bohrn et al. (2006) and Potthast et al. (2015).
X-ray diffraction
Dried samples of BC were cut into small pieces and shortly mortared using a ceramic mortar and pistil for less than 2 min to avoid excessive amorphization due to BC processing but reduce the effect of preferred crystal orientation (Thygesen et al. 2005). The prepared samples were then kept dry and at room temperature until XRD analysis. The measurements were carried out in reflection mode using a PANalytical Empyrean diffractometer series 2 (Malvern Panalytical, Almelo, The Netherlands) with Cu Kα radiation (λ = 1.5419 Å, Kα2: Kα1 = 0.5, Cu LFF HR goniometer) on a silicon sample holder. The intensity was recorded with a PIXcel-3D detector from 10 to 40° 2θ with a step size of 0.053°. Determination of cellulose crystallinity was realized in accordance with Yao et al. (2020) which included the pre-treatment of the raw data in terms of three steps: background subtraction of the empty silicon sample holder, polarization correction of the intensities by multiplication with 2/(1 + cos22θ) (Buerger 1940), and data smoothing using the FFT filtering tool of PeakFit software (Version 4.12) (see SI Fig. S1). Voigt functions (Voigt Amp) were used for crystalline signal assignment according to the structure of cellulose Iα determined by Nishiyama et al. (2003) and an 8th order real form Fourier series model proposed by Yao et al. (2020) was selected for representation of the amorphous cellulose content. The automatic PeakFit scaling of crystalline and amorphous signals was done so to achieve high coefficients of determination (R2)–although manual rescaling was necessary to prevent the software from unrealistic (amorphous) signal broadening due to apparently higher R2 but lesser crystal structure validity. Finally, crystallinity was calculated as the share of the crystalline peak area from the total assigned area based on the crystalline and amorphous content multiplied by 100. The crystallinity determination according to Segal et al. (1959) was conducted for comparison only since the majority of available crystallinity values of BC in literature were evaluated this way due to the ease of the method. The calculation of the d-spacings d1 and d2, representing the hkl planes (100) and (010) of the cellulose phase Iα, using Bragg’s law was applied to differentiate between the crystalline phases of the cellulose allomorph. This allows for estimating the phase content of the cellulose phases Iα and Iβ in the cellulose structure (Ruan et al. 2016). Wada et al. (2001) found the deviations of the experimental diffraction angles from the theoretical Iα phase to be linked to the occurrence of the Iβ phase with the diffraction planes (1–10) and (110).
Results and discussion
Cellulose production in HS-type media
Three different types of HS media (HS, HS_EtOH, HS_S) were initially investigated with a focus on the media’s ability to facilitate BC production with K. xylinus DSM 2325. Additionally, the application of these basic media was conducted to provide insight into the dominating catabolic processes which might be affected upon the addition of complex industrial by-product components.
In HS medium, glucose was almost completely converted into organic acids in less than 40 h leading to a drastic pH decrease and a final BC concentration of about 1.7 g L−1 (Fig. 1a). This metabolization of glucose to predominantly gluconic acid (see SI Table S1) represents one of the main difficulties in BC production as it reduces the yield and leads to pH values lower than 3.5 which reportedly lowers or stops cellulose synthesis (Masaoka et al. 1993; Embuscado et al. 1994; Gwon et al. 2019). Despite the initial disadvantageous production of gluconic acid in the HS medium, it is slowly consumed afterwards and partially converted into BC (Fig. 1a). The extent of this by-product formation can be reduced by the selection of other main carbon sources (e.g., sucrose or glycerol) or additives such as ethanol which was shown to act as an additional ATP source and regulator of the transcriptome towards BC production (Naritomi et al. 1998; Yunoki et al. 2004; Ryngajłło et al. 2019a). Gwon et al. (2019) have demonstrated that this redirection of glucose metabolization pathways can also be directed more actively towards BC synthesis by applying metabolic engineering methods.

Cellulose concentration, pH, and carbon sources over the time course of dynamic shake flask cultivations in triplicate using a HS, b HS-type with 0.5% v/v ethanol supplementation (HS_EtOH), and c HS-type medium with sucrose (HS_S)
The effects of ethanol supplementation on the cultivation course were investigated by adding 0.5% v/v ethanol to the HS medium. Preliminary experiments have indicated that this concentration of ethanol is the most favorable one for BC synthesis in HS medium under the chosen cultivation conditions (see SI Fig. S2). Figure 1b highlights the increased amount of BC up to ~ 4 g L−1 after 168 h in comparison to the HS medium reference. Glucose consumption (0.2 g L−1 h−1) and organic acid formation rate were slowed down during the initial phase when ethanol was available which was also reported for several other strains and medium compositions (Park et al. 2003; Hyun et al. 2014; Souza et al. 2021). Analytics further confirmed a switch in the type of produced organic acids: Instead of an accumulation of citric acid up to 5.5 g L−1 in the HS medium after 42 h, acetic acid was formed in HS_EtOH media up to 2.6 g L−1 whereas the citric acid concentration was only 1.5-fold higher than the initial level of 1.3 g L−1 (see SI Table S1). This switch is due to the membrane-bound or intracellular ethanol oxidation to acetic acid which was previously confirmed by transcriptome analysis (Ryngajłło et al. 2019a). Interestingly, the following uptake rate of the initially produced organic acids is twice as high in the ethanol supplemented cultures as in the reference. In detail, acetic acid and gluconic acid are consumed in parallel whereas in HS medium gluconic acid is the only consumed organic acid of significance. The concomitant availability of gluconic acid for cellulose build-up and acetate as ATP generator via the tricarboxylic cycle (TCA) in HS_EtOH medium seem to allow for an improved BC synthesis despite a consistently low pH around 3.2 which proves the beneficial effects of ethanol as supplement in glucose-based cultivation media (Naritomi et al. 1998; Son et al. 2001; Liu et al. 2018; Ryngajłło et al. 2019a).
The exchange of the main carbon source glucose for sucrose in Fig. 1c resulted in a 1.2-fold increase in BC after 168 h compared to HS medium and an overall higher pH during the cultivation. This is most likely due to the 5.1-fold lower gluconic acid formation when comparing its peak concentration (2.8 g L−1) to the HS medium (see SI Table S1). The pH change and improved BC synthesis with sucrose is thereby in accordance with other cellulose producing strains (Embuscado et al. 1994; Jung et al. 2010b). The findings from these HS-type media experiments suggest that food-industry by-product streams like molasses as sucrose containing carbon source and waste beer fermentation (WBFB) as ethanol-containing complex media sources could have especially high potential for enhanced BC production media because they would combine the positive aspects of sucrose and ethanol.
Analysis of raw complex media components
Prior to its application, molasses, vinasse, and WBFB were investigated for their TOC and TNb content and a more detailed composition of the raw complex components was obtained from sugar, organic acid, and alcohol analysis. Molasses from sugar beets showed the highest TOC concentration with sucrose representing the greatest share of it (Table 2). Additionally, sucrose hydrolysis products like glucose and fructose or organic acids such as lactic acid and acetic acid are accumulated therein which is in agreement with previous reports. This high, sugar-based TOC content makes molasses a suitable carbon source for BC production whereas nitrogenous components such as amino acids, betaine, small amounts of nitrates and proteins contribute to an considerable TNb of the undiluted beet molasses (Stark 1961; Parnaudeau et al. 2008; Palmonari et al. 2020; Volova et al. 2022).
Beet vinasse was selected as potential nitrogen source as it contained the highest TNb of the investigated complex substrates. Although the TNb assay detects organically bound nitrogen in parallel with nitrate and ammonium ions, the share of the latter two was assumed to be low since previous studies determined that the majority of nitrogen is derived from proteins, betaine and a small share of free amino acids (Parnaudeau et al. 2008; Deutsche Melasse Handelsgesellschaft 2015; Hernández et al. 2019). Glycerol and lactic acid were found to constitute the major carbonaceous components of vinasse and could thereby add additional value to cultivation media (Table 2). The resolved analytes and concentrations confirm analyses from other researchers within the expectable variability due to already differing beet molasses as starting material for ethanol production (Parnaudeau et al. 2008; Harirchi et al. 2020; Bernardes et al. 2021).
WBFB was selected as second nitrogen source which reportedly contains proteins (e. g., plant storage proteins such as hordein, enzymes, peptides) and amino acids as nitrogenous components resulting from the malting, hopping, and brewing process (Gribkova et al. 2021). Small peptides and amino acids were, however, found to constitute the greatest share of the organically bound nitrogen (Khan et al. 2007; Abernathy et al. 2009). In addition, utilization of WBFB allows ethanol supplementation to the production medium which was found to be beneficial for BC synthesis in terms of yield and especially genetic stability of the organism as it can slow down the occurrence of cellulose deficient cells (Liu et al. 2018; Ryngajłło et al. 2019a). Besides from ethanol, lactic acid, and acetic acid, no further carbon sources were detected within the range of selected analytes.
Cellulose production in HS-type media with SCC
The next experimental approach was designed to investigate the BC production while applying single complex components (SCC) from industrial by-product streams in exchange for HS medium components on a TOC- or TNb-based quantity. The replacement of glucose by the addition of an equal TOC amount of molasses to the cultivation media resulted in an initial sugar concentration of about 17.3 g L−1 with citric and acetic acid covering for some of the lacking sugar-based carbon content (Fig. 2a and SI Table 1). The following sugar consumption rate (0.15 g L−1 h−1) until 84 h after inoculation is reduced in comparison to the HS-type media (0.24 g L−1 h−1) but resembles the initially reserved sucrose degradation in HS_S (Fig. 1b). This delay might be a result of the required time of the glucose adapted pre-culture for the biosynthesis of sucrose degrading enzymes which were found to be an intracellular sucrose phosphorylase and extracellular invertase for Acetobacter xylinum (Balasubramaniam and Kannangara 1982). Additionally, a delay could be due to residual (poly-)phenolic compounds from molasses. These by-products can have inhibitory impacts on bacterial growth (Palla 1983; Chen et al. 2017a) although their role in the metabolism of cellulose producing bacteria is ambiguous and might be compound-specific because lignosulfonates were previously found to increase BC production (Premjet et al. 1996; Keshk and Sameshima 2006). Interestingly, the organic acid formation was clearly reduced and citric and gluconic acid consumption was initiated after 42 and 84 h, respectively, which resulted in a pH increase. Potentially, the metabolization of amino acids from molasses such as aspartic acid or threonine could lead to ammonia release by putative aspartate or threonine ammonia lyases and thus concomitantly cause a pH increase (McMurry and Begley 2006; Jang et al. 2019). A catabolic pathway for betaine (trimethylglycine), a major nitrogenous compound in beet molasses, via dimethylglycine was not found in the KEGG database for related strains (K. xylinus E25, K. nataicola RZS01, K. europaeus SRCM101446) nor as putative gene annotation for K. xylinus DSM 2325 (Stark 1961; Parnaudeau et al. 2008; Zou et al. 2016; Jang et al. 2019). Overall, the utilization of molasses as carbon source led to a slightly lower BC concentration (1.8 g L−1) than in the sucrose containing HS-type medium which could be attributed to an unfavorable pH switch (pH > 6.5) at the later stage of cultivation or a diverging protein and energy metabolism since most of the initially available sugars were not converted in any of the quantified analytes and might have dominantly promoted biomass growth as described in previous studies (Park et al. 2010b; Çakar et al. 2014; Salari et al. 2019). However, the HS_M(C) resulted in a slightly higher (1.1-fold) BC concentration than HS medium which agrees with other studies where beet molasses was applied in terms of equal amounts of sugars in static cultivations which additionally caused a lower pH drop (Salari et al. 2019; Volova et al. 2022). Further investigations even determined enhanced biomass formation than in HS medium which was ascribed to molasses-derived beneficial nutrients such as vitamins (Park et al. 2010b; Çakar et al. 2014) which were found to improve cultivations under specific conditions (Premjet et al. 2007).

Cellulose concentration, pH, and carbon sources over the time course of dynamic shake flask cultivations in triplicate using a HS-type medium with a TOC equivalent amount of molasses as carbon source (HS_M(C)), b HS-type medium with TNb equivalent amount of vinasse as nitrogen source (HS_V(N)), and c HS-type medium with TNb equivalent amount of waste beer fermentation broth (WBFB) as nitrogen source (HS_WBFB(N))
The exchange of peptone and yeast extract for vinasse as nitrogen source based on an equal TNb value resulted in a considerable (4.6-fold) increase of the final BC concentration to 7.8 g L−1 (Fig. 2b). In contrary to all previous cultivations, the BC synthesis followed an exponential instead of a linear production behavior. This is of high similarity to the static and dynamic cultivations by Velásquez-Riaño et al. (2013) with Gluconacetobacter kakiaceti GM5 as organism using pure vinasse as production medium. Since glucose served as main carbon source in HS_V(N) medium, the accumulation of gluconic acid up to 9.6 g L−1 after 42 h and 4.4 g L−1 citric acid after 84 h is not surprising but considerably lower (1.5-fold, each) than in HS medium (see SI Table S1). The overall reduced organic acid formation in comparison to the reference medium is most likely related to the additional glycerol which originates from the vinasse solution and can serve as a parallel energy source which does not necessitate complete glucose oxidation to gluconic acid for energy supply. Glycerol is often regarded as an applicable carbon source like glucose or sucrose for BC production although the grade of suitability differs between cellulose producing strains (Masaoka et al. 1993; Mohite et al. 2013; Zhong et al. 2013; Tsouko et al. 2015; Wang et al. 2018). Recent investigations have shown differences in the genetic repertoire regarding glycerol metabolization and its cellular uptake mechanisms between strains although the lacking adaption during pre-cultures might have placed glycerol in disadvantage to sugar-based carbon sources in terms of space–time-yield in some studies (Ryngajłło et al. 2019b). Apparently, K. xylinus DSM 2325 shows a large pH range tolerance for BC synthesis since high cellulose concentrations occurred between pH 3.2 (HS_EtOH) and 7.5. Overall, vinasse exhibits high potential for enhancing BC production, but it seems essential to dilute and add other components to vinasse-based media since previous studies using pure or diluted forms of vinasse only resulted in small increases or even lower cellulose concentrations compared to the HS medium-like reference (Velásquez-Riaño and Lombana-Sánchez 2009; Velásquez-Riaño et al. 2013; Barshan et al. 2019).
The third complex by-product WBFB was also investigated as nitrogen-replacement in HS medium resulting in a final BC concentration of 4.2 g L−1 (Fig. 2c). As in HS_EtOH, a great increase of gluconic acid formation up to 11.1 g L−1 was detected within 42 h although it was still 1.3-fold less than in HS medium (see SI Table S1). The initial accumulation of organic acids also explains the drop of the pH value to about 3.5 during cultivation which increases slowly afterwards due to the metabolization of gluconic and acetic acid. However, citric acid is continuously secreted until the end of the cultivation and has only been found to be considerably reduced in sucrose containing cultivations. The applicability of WBFB as supplement agrees well with previous findings in shaken or static conditions were similar or slightly increased BC concentration have been achieved when glucose was added to WBFB (Ha et al. 2008; Shezad et al. 2009; Fleury et al. 2020). Moreover, the beneficial effect of ethanolic and nitrogen rich by-products on BC production has also been pointed out for makgeolli sludge filtrate that was supplemented to HS medium which highlights the potential of this type of food wastes (Hyun et al. 2014). The overall results of these experiments using HS-type media as base for the TOC or TNb equivalent addition of molasses, vinasse, or WBFB prove the high potential of the supplementation of different food-industry by-product streams to increase BC production in dynamic cultivations.
Cellulose production in HS-type media with CCC
The improvements due to the applications of SCCs encouraged us for an approach with combined complex components (CCC) using a carbon and nitrogen source from food industry by-products. Herein, the TOC content was covered by molasses whereas vinasse, WBFB, or a combination of both served solely as nitrogen source. At first, molasses was combined with vinasse giving high initial concentrations of lactic and citric acid resulting from both complex components (Fig. 3a). Sucrose as main carbon source was hydrolyzed more slowly in comparison to HS_S and HS_M(C) which might be a result of the availability of organic acids from the vinasse. Citric acid and lactic acid were consumed until 84 h, whereas the concentration of the latter increased slightly afterwards together with gluconic acid. Especially the consumption of citric acid could have led to a reduced buffering capacity of the cultivation medium which might explain the strong increase of the pH > 8.0 due to amino acid catabolism and despite the gluconic acid formation. Overall, the BC production (4.3 g L−1) is worsened in comparison to the HS_V(N) medium (7.8 g L−1) but improved comparing it with the HS_M(C) medium (1.8 g L−1). The unfavorable pH increase could have lowered biomass formation and BC production at the end of the experiment and thus might show the pH limit of this organism.

Cellulose concentration, pH, and carbon sources over the time course of dynamic shake flask cultivations in triplicate using a HS-type medium with TOC equivalent amount of molasses as carbon and TNb equivalent amount of vinasse as nitrogen source (HS_M(C)/V(N)), b HS-type medium with TOC equivalent amount of molasses as carbon and TNb equivalent amount of waste beer fermentation broth (WBFB) as nitrogen source (HS_M(C)/WBFB(N)), and c HS-type medium with TOC equivalent amount of molasses as carbon and TNb equivalent amount of vinasse and WBFB as nitrogen source (HS_M(C)/V(N)/WBFB(N))
In the next step, WBFB was added instead of vinasse as nitrogen source resulting in a final BC concentration of 8.1 g L−1 (Fig. 3b). This increase is partly a result of the piling effects of sucrose as carbon source and ethanol supplementation via the WBFB. However, gluconic acid is produced up to 5.6 g L−1 which was contradictory as it was half this amount in HS_S and even less with molasses as main carbon source (HS_M(C)). Lower gluconic acid formation when using sucrose instead of glucose as carbon source in an equimolar amount is mostly due to the lower molarity, slower release, and thus slower conversion of glucose as it only becomes enzymatically accessible during the cultivation course. The latter advantage was eliminated using thermally untreated WBFB as it contains remaining enzymes from the previous brewing process from where the complex by-product originates. These enzymes are mainly maltases, invertases, or other enzymes that are capable of hydrolyzing the molasses-derived sucrose into glucose and fructose (Marques et al. 2016). The initial availability of higher glucose concentrations thus led to the gluconic acid accumulation which was consumed quickly thereafter together with all other organic acids. The fructose concentration decreased much slower until the end of the cultivation course which is typical when glucose and fructose are available in parallel (Zhong et al. 2013). From 126 h on, the effects of a carbon limitation became visible since most of all carbon sources were exhausted and BC production clearly slowed down. The organic acid consumption was accompanied by a drastic pH increase up to 7.6 which could have additionally reduced the BC synthesis. The combination of these two substrates proves to the best of our knowledge for the first time the beneficial effects of the combination of molasses and WBFB leading to the highest BC concentration of our investigated media.
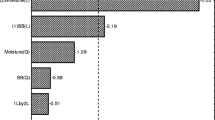
In a final step, molasses was used as TOC equivalent carbon source whereas vinasse and WBFB served as TNb equivalent nitrogen source in a 1:1 ratio (Fig. 3c). The cultivation course is very similar to the HS_M(C)/WBFB(N) medium with the initial ethanol availability causing a temporary acetic acid accumulation up to 2.1 g L−1 (see SI Table S1). The pH value reached up until 8.7 during the cultivation which is assumed to be one reason for the stagnating BC production after 126 h. The other is related to the exhaustion of the main carbon source and only small remaining amounts of lactic, gluconic, and citric acid. Interestingly, the shared application of all three complex resources led to the highest space–time yield of 1.57 g L−1 d−1 after 126 h which is a significant increase over 0.24 g L−1 d−1 in the HS medium. The successful combination of complex food industry by-products for BC production has recently been reported by Souza et al. (2021) who thereby reached their highest BC concentration of 11.1 g L−1. Locally available by-products such as sugar cane molasses and coffee grounds were combined with ethanol to a fruitful composition that reflects the potential of bioeconomic approaches and equals our own findings for sugar beet molasses, vinasse, and WBFB as the ethanol containing fraction. In both cases, the combinations resulted in higher BC concentrations and proved the strategy to utilize complex by-products from an economic and ecologic point of view successful. The overall results of the investigated BC production media are highlighted in Fig. 4 which concludes the improvements made based on the addition of complex by-products as replacement for HS medium components.

Cellulose concentration after 7 days of dynamic cultivation of K. xylinus DSM 2325 in HS and HS-type media applying single complex components (SCC) or combined complex components (CCC)
Molecular weight analysis
The MWDs of the produced BCs were investigated using GPC analysis with a MALLS detector. The resulting weight- (Mw) and number-average molecular weights (Mn) with their respective degree of polymerization (DP) are shown in Table 3. The comparison of absolute values concerning these MWD indicators with other authors should always be conducted carefully as they strongly depend on the selected protocol for sample preparation and the overall metrological approach (Potthast et al. 2015). Additionally, parameters such as the chosen bacterial strain, medium, cultivation conditions, and duration have an impact on the final results (Einfeldt and Klemm 1997; Strobin et al. 2004; Moon et al. 2006). This is initially reflected by the calculated DPw of 3478 for the dynamic HS medium which is 2.1-fold lower than the value determined by Hur et al. (2020) for K. xylinus DSM 2325 using a GPC measurement with polystyrene standards. This discrepancy of the DPw could be related to still slightly differing cultivation conditions such as a higher shaking frequency of 230 min−1, for example, or due to the indirect GPC calibration method which we found to result in a 3.3-fold overestimation of the DPw compared to the direct measurement via MALLS detection.
As ambiguous as the absolute values is the question of the effect of static or dynamic cultivation on the MWD or its indicative parameters. Some authors found a decrease of the DPw for agitated and aerated or solely aerated cultivation conditions which was attributed to cellulose degrading enzymes, the interference of water-soluble polysaccharide by-products, or the hydrodynamic disturbance of the biosynthetic procedure (Tahara et al. 1997; Watanabe et al. 1998; Moon et al. 2006; Shavyrkina et al. 2021). In contrast, Shi et al. (2013) found a slightly increased, viscometrically determined DPv and Hur et al. (2020) even distinctly greater DPw for cells incubated in dynamic cultivations which is in contrary to our own findings of a 2.3-fold higher DPw after 7 days of static cultivation and is included here for comparison. In terms of the course of the polymer molecular weight during the cultivation process, most of the studies concluded with a more or less continuous increase of the degree of polymerization (DPw and DPv) during static and dynamic cultivations which can be followed by decreases during the later stages of the cultivation (Husemann and Werner 1963; Marx-Figini and Pion 1974; Okajima et al. 1991; Einfeldt and Klemm 1997; Strobin et al. 2004; Skiba et al. 2022). However, a few studies found a continuous decrease of the DPw and DPv for different strains during bioreactor or static cultivation from the early stages on which was explained by cellulose degrading enzymes but could especially in case of DPv determinations also be biased by other viscosity inducing polymers such as acetan (Tahara et al. 1997; Fijałkowski et al. 2016; Ryngajłło et al. 2020).
In regard of the investigated cultivation media in this study, the HS medium has induced the lowest DPw with the greatest polydispersity index (PDI) because of a greater share of low molecular weight BC fibers. Despite this, the weight-average MWD still indicates a clear unimodal distribution as for all other samples which agrees with detailed previous studies using static cultivation conditions (Einfeldt and Klemm 1997; Strobin et al. 2004; Ono et al. 2016). The ethanol supplementation (HS_EtOH) and exchange of glucose for sucrose (HS_S) both increased DPw which coincides with a higher BC production than in the HS medium. Fructose instead of glucose as carbon source during static cultivations was shown to increase the fraction of low molecular weight cellulose which caused a reduced DPw (Strobin et al. 2004). This correlation could not be confirmed for our HS_S medium and might thus be specific for certain strains and cultivation conditions or only occur in complete glucose absence when there is no direct glucose precursor for cellulose biosynthesis available.
The utilization of a single complex substrate overall increased the respective DPw in relation to the HS medium. For instance, the selection of the HS_V(N) medium resulted in the highest DPw of all dynamic cultivations which could be related to the remaining glycerol content that was found before to induce higher DPv when exchanged for glucose in HS medium and to the overall higher TOC content that might have enabled prolonged usage of glucose for BC biosynthesis (Park et al. 2010b; Tsouko et al. 2015). However, a reduced DPv and thus the opposite has also been reported before and was explained by a lack of precursor supply for cellulose synthesis when glucose was replaced by glycerol (Shi et al. 2013). This explanation is not applicable to HS_V(N) due to the availability of glucose until 82 h which might explain the increased DPw here (Fig. 2b). Molasses in HS_M(C) medium as single complex carbon source caused similar DPw like HS_S although the share of low molecular weight cellulose fibers was higher in HS_S which is indicated by its respectively lower DPn. The higher DPw of HS_M(C)-derived BC thereby contradicts the results of four out of five investigated BC producing strains for which the DPv in statically grown cultures decreased when using diluted sugar cane molasses in comparison to HS medium (Keshk and Sameshima 2006).
However, multiple factors affect the final DP as discussed before in this section and one of those could be related to substrate limitations. For example, Fig. 1b shows a sugar exhaustion in HS_S after 82 h whereas this occurred 44 h later in HS_M(C) (Fig. 3a). The production of new cellulose fibers in HS_S might have been interrupted more often due to a lower precursor supply as also suggested by Skiba et al. (2022) for static kombucha cultures. Concomitant oxygen and glucose (diffusion) limitations have been pointed out by Hornung et al. (2006) to restrict the increase of the BC layer in static fermentations. Considering this and the previous findings by the aforementioned studies that the DP starts to decrease after a certain cultivation period, we assume that the DP reduction and PDI increase towards the end of a carbon limited BC production process is linked to lower precursor availability in the BC agglomerate due to e.g., low diffusion gradients and ending sugar supply (see SI Fig. 3a and 3b). Despite this proposed linkage, it cannot be clarified whether the occurrence of the rising low molecular weight fraction is caused by recently for K. xylinus DSM 2325 annotated cellulases (cmcax, bglaX (Jang et al. 2019)) as indicated by Tahara et al. (1997) or solely due to a more frequently interrupted BC synthesis process because of precursor-limitation as hypothesized by Skiba et al. (2022).
The cultivation media with CCC overall confirm the findings for the SCC media: Vinasse as nitrogen source in combination with molasses lead to the highest DPn within the investigated sample range and have had positive effects on the medium with all three complex substrates (HS_M(C)/V(N)/WBFB(N)). This effect is attributed to the additional glycerol and thus TOC which in these cases could have served as energy source in parallel to the sugar-based carbon sources. This allowed a more effective and prolonged conversion of the sugar-based carbon source in cellulose precursors instead of gluconic acid.
XRD analysis
For further characterization of the produced BC, XRD analysis was conducted to investigate the effects of complex media components on the BC structure and crystallinity. All obtained diffractograms showed crystalline diffraction patterns that were assigned to the cellulose Iα phase. However, slight shifts of the signal at 14.6° (Iα = (100)) towards higher and the signal at 16.8° (Iα = (010)) towards lower diffraction angles were regarded as a result of the underlying monoclinic Iβ phase (Iβ = (1 −10) and (110), respectively) that can coexist and is dealt with later (Atalla and VanderHart 1984; Wada et al. 2001; French 2014). Two representative diffractograms from BC that were generated in HS and HS_V(N) medium with their assigned crystalline signals after peak deconvolution are shown in Fig. 5. The main diffraction signals correspond to the triclinic unit cell with the Miller indices (100), (010) and (110) (Nishiyama et al. 2003). The small diffraction signal at 20.5° could not be further discriminated and was assigned to multiple overlapping Iα crystalline signals ((11−2), (01−2), (10−2)) as reported by others for BC samples (Iwata et al. 1998; Fang and Catchmark 2014; Aleshina et al. 2019; Skiba et al. 2022). A similar proceeding was applied to the signals at 28.8° and 29.8° or 33.9° and 34.7° where no precise signal assignment but consideration for peak integration was found reasonable as these occurred throughout many BC publications (Czaja et al. 2004; Fang and Catchmark 2014; Ruan et al. 2016; Aleshina et al. 2019; Yao et al. 2020). It is clearly visible from Fig. 5 that the dynamic cultivation in HS medium resulted in much more crystalline BC than the HS_V(N) since the amorphous content is much more prevalent there.

X-ray diffraction patterns of BC produced in a HS and b HS_V(N) medium with their respective assigned crystalline and amorphous signals after peak deconvolution
The calculated crystallinities of all investigated BC samples are listed in Table 4. As for the DPw in Table 3, the highest crystallinity of the HS-type media was achieved with the BC produced under static conditions although the difference to the dynamic cultivation is rather small. A discrepancy is typical and described in many previous studies (Iwata et al. 1998; Watanabe et al. 1998; Czaja et al. 2004; Moon et al. 2006; Cheng et al. 2009; Guo and Catchmark 2012). It has been deduced that occurring shear stress interferes with the formation and crystallization process of the cellulose ribbon which is corroborated here by the higher DPw of the statically produced BC (Table 3). The hampered BC production at shearing conditions was additionally assumed to induce smaller crystallite sizes that could induce the Iβ phase of the cellulose allomorph (Watanabe et al. 1998). A severe decrease of the cellulose Iα phase content compared to static conditions was found here, too (Table 4). The crystallinity determined via the Segal method in HS medium agrees well with previous studies where values from 70.5 to 89.7% (Moon et al. 2006; Choi et al. 2009; Chen et al. 2011; Tsouko et al. 2015) or 78% (Zhou et al. 2007; Jung et al. 2010a) have been reported for static or shaken conditions, respectively. However, the Segal method has been shown to overestimate the sample crystallinity of celluloses by about 25% in total in comparison to the peak deconvolution or 13C-NMR C4 peak separation method (Park et al. 2010a). This confirms our results obtained via the peak deconvolution method that are about 32% lower which is mainly attributed to a more reasonable consideration of the amorphous content. Still, a crystallinity of 56% for a static cultivation is low in comparison to the little available BC data determined via deconvolution or Rietveld method which ranged from 71.0 to 82.2% in HS medium (Watanabe et al. 1998; Park et al. 2010a; Guo and Catchmark 2012; Ruan et al. 2016; Chen et al. 2017b). The herein applied deconvolution method proposed by Yao et al. (2020) resulted for them in 33.4% crystallinity for a presumably static cultivation. This low value highlights that the therein performed XRD raw data correction for background (sample holder) and polarization effects, choice of peak function (Voigt-type), or applied amorphous background subtraction has incorporated effects that were not considered before. The Rietveld or 13C-NMR method was not applied there which would have allowed an even more comprehensive view on the available analysis methods.
The crystallinities obtained in our study support the findings of previous investigations that the cultivation medium and certain additives can affect the cellulose crystal formation (Iwata et al. 1998; Khattak et al. 2015; Wang et al. 2018). Our complex media with WBFB yielded higher crystallinities than those with molasses or vinasse which might be related to the additional ethanol content (Table 4). Slight enhancements of the BC crystallinity due to ethanol supplementation were also determined by Volova et al. (2018) although this increase is most likely due to a more efficient BC biosynthesis from glucose since ethanol as sole carbon source did rather decrease the crystallinity (Wang et al. 2018). A static fed batch process involving WBFB yielded 1.07-fold higher crystallinities in comparison to a HS-like medium (Shezad et al. 2010) and thus supports our findings of only small positive effects. For different media compositions based on ethanol-containing makgeolli sludge filtrate, the authors solely showed the respective diffraction patterns but lacked the crystallinity calculations (Hyun et al. 2014). Nonetheless, the assigned cellulose Iα diffraction signals were clearly more prevalent in all media containing ethanol which could indicate an increased structural order for ethanol supplemented cultivations.
The lower crystallinities for beet molasses and vinasse containing media are most likely linked to beet related compounds that interfered with the crystal formation. Several studies have investigated the effects of plant-derived hemicelluloses on the formation of the BC structure and found e.g., glucomannan and xyloglucan to be incorporated in the cellulose structure whereas arabinoxylan seemed to interact differently and was majorly found surface associated (Uhlin et al. 1995; Iwata et al. 1998; Tokoh et al. 2002; Martínez-Sanz et al. 2015). The incorporation of glucomannan and xyloglucan in the cellulose ribbon was accompanied by reduced crystallinities of the BC and a shift of the allomorph towards the Iβ phase. Additionally, it has been pointed out that BC production in presence of xyloglucans changed the cellulose structure from twisted ribbons to bundled fibrils which had lower diameters than the control (Uhlin et al. 1995). These findings suggest that the reduced crystallinity and increased Iβ phase content could be due to the interaction of remaining hemicelluloses in molasses and vinasse media with the cellulose polymer (Wohryzek 1928; Palmonari et al. 2020). A lower crystallinity and differing fibril arrangement has been reported before for BC produced in sugar cane molasses-based media and was referred to by Vazquez et al. (2013) as a lacking self-assembly of the glucan chain by the hydroxyl groups (Keshk and Sameshima 2006; Khattak et al. 2015). A reduction of the Iα content was, however, not found despite a double-check via FTIR and 13C-NMR measurement in one of these studies (Keshk and Sameshima 2006). This coincides with a constant Iα content and even slightly increased BC crystallinity with sugar cane molasses as HS medium supplement in another study (Tyagi and Suresh 2016). However, these seemingly deviating results can be explained by a more intense sulfuric acid treatment of the raw molasses. The applied high temperature and longer incubation time of the molasses might have sufficiently accelerated the degradation of remaining hemicelluloses and allowed the consumption of the released sugars by the microorganisms.
A reduced crystallinity in the HS_V(N) medium due to the glycerol component is less likely since glycerol as replacement for glucose in HS medium has yielded celluloses with similar or higher values (Jung et al. 2010a; Vazquez et al. 2013; Tsouko et al. 2015; Wang et al. 2018). The effect of hemicelluloses or crystallinity-affecting sugar beet compounds is also supported by the greater amount of vinasse that was added to HS_V(N) to meet the TNb requirement compared to HS_M(C). Hence, the crystallinity is the lowest in HS_V(N) which agrees with an investigation where vinasse-based media resulted in slightly lower crystallinities than HS medium under static conditions (Barshan et al. 2019).
In conclusion, our investigations, and comparisons with available data for BC crystallinities from complex media and industrial by-products corroborate that the composition of media can distinctly affect the crystallinity of BC in a positive or negative manner. Since the high crystallinity of BC is often regarded as one of its most remarkable properties, complex media pre-treatments and additives should be selected accurately before the production process to sustain this rare structural feature.
Conclusion
This study aimed at a deeper understanding of the applicability of beet molasses, vinasse, and WBFB for BC production and its effect on structural polymer properties in terms of DP, crystallinity, and dominating Iα or Iβ phase content of the cellulose allomorph. Therefore, the industrial by-products were initially characterized for their TOC, TNb, sugar, organic acid, and alcohol content to enable a profound comparison of the single components as carbon or nitrogen source based on HS medium equivalents. It was shown that molasses as sole TOC equivalent glucose replacement yielded similar BC concentrations (1.8 g L−1) compared to the HS medium (1.7 g L−1) but remarkably lowered its crystallinity along with an increase of the DPn. The structural changes were even more prevalent when vinasse was supplemented to HS medium on a TNb-based level. However, the BC production was increased 4.6-fold in HS_V(N) medium (7.8 g L−1) which makes vinasse a suitable nitrogen source. This was most likely due to the remaining glycerol content and organic acids that can serve as ATP source and allowed a more efficient utilization of the supplied carbon source. WBFB enhanced the BC production only 2.5-fold but sustained a high DPn and crystallinity of the polymer. The latter properties should be considered to obtain high quality BC despite low-cost complex substrates. For that reason, molasses as main carbon source and a mixture of vinasse and WBFB as nitrogen source were combined which yielded the highest BC concentration (8.2 g L−1) and one of the highest crystallinities (58%) obtained in this study while maintaining a moderate DPn (3343). This combination highlights the beneficial aspects of combined complex substrates for an upscaled BC production with the knowingly important ethanolic share within WBFB for genetic stability of the organism. Furthermore, the obtained results for DP and crystallinity of BC from different complex media will support to gain a deeper understanding of influential parameters on the BC formation process. Future studies might take a closer look at resolving remaining components in complex media such as the hemicelluloses and the contribution of amino acids or fed-batch processes on the DP and XRD analyses to increase the knowledge towards the BC biosynthesis. This could uncover the full potential of BC in terms of productivity, structural properties, and an environmentally more sustainable production chain.
Data availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
References
Abernathy DG, Spedding G, Starcher B (2009) Analysis of protein and total usable nitrogen in beer and wine using a microwell ninhydrin assay. J Inst Brew 115:122–127. https://doi.org/10.1002/j.2050-0416.2009.tb00356.x
Africa TK (1949) The production of nata from coconut water. Unitas 22:60–100
Aleshina LA, Gladysheva EK, Budaeva VV, Golubev DS, Skiba EA, Sakovich GV (2019) X-ray diffraction study of bacterial nanocellulose produced by Medusomyces Gisevii Sa-12 cultured in enzymatic hydrolysates of miscanthus. Crystallogr Reports 64:914–919. https://doi.org/10.1134/S1063774519060026
Atalla RH, VanderHart DL (1984) Native cellulose: a composite of two distinct crystalline forms. Science 223:283–285. https://doi.org/10.1126/science.223.4633.283
Bae S, Shoda M (2004) Bacterial cellulose production by fed-batch fermentation in molasses medium. Biotechnol Prog 20:1366–1371. https://doi.org/10.1021/bp0498490
Balasubramaniam K, Kannangara PN (1982) Sucrose phosphorylase and invertase activities in bacteria. J Natl Sci Found Sri Lanka 10:169. https://doi.org/10.4038/jnsfsr.v10i2.8426
Barshan S, Rezazadeh-Bari M, Almasi H, Amiri S (2019) Optimization and characterization of bacterial cellulose produced by Komagatacibacter xylinus PTCC 1734 using vinasse as a cheap cultivation medium. Int J Biol Macromol 136:1188–1195. https://doi.org/10.1016/j.ijbiomac.2019.06.192
Bernardes AN, Godoi LAG, Camiloti PR, da Gomes AC, Sanchez BLS, Torres APR, Botta LS (2021) Organic acids and alcohols quantification by HPLC/RID in sugarcane vinasse: analytical method validation and matrix effect assessment. Int J Environ Anal Chem 101:325–336. https://doi.org/10.1080/03067319.2019.1666833
Bohrn R, Potthast A, Schiehser S, Rosenau T, Sixta H, Kosma P (2006) The FDAM method: determination of carboxyl profiles in cellulosic materials by combining group-selective fluorescence labelling with GPC. Biomacromol 7:1743–1750. https://doi.org/10.1021/bm060039h
Buerger MJ (1940) The correction of X-Ray diffraction intensities for lorentz and polarization factors. Proc Natl Acad Sci 26:637–642. https://doi.org/10.1073/pnas.26.11.637
Çakar F, Özer I, Aytekin AÖ, Şahin F (2014) Improvement production of bacterial cellulose by semi-continuous process in molasses medium. Carbohydr Polym 106:7–13. https://doi.org/10.1016/j.carbpol.2014.01.103
Campano C, Balea A, Blanco A, Negro C (2016) Enhancement of the fermentation process and properties of bacterial cellulose: a review. Cellulose 23:57–91. https://doi.org/10.1007/s10570-015-0802-0
Chen HH, Chen LC, Huang HC, Bin LS (2011) In situ modification of bacterial cellulose nanostructure by adding CMC during the growth of Gluconacetobacter xylinus. Cellulose 18:1573–1583. https://doi.org/10.1007/s10570-011-9594-z
Chen M, Zhao Z, Meng H, Yu S (2017a) The antibiotic activity and mechanisms of sugar beet (Beta vulgaris) molasses polyphenols against selected food-borne pathogens. LWT - Food Sci Technol 82:354–360. https://doi.org/10.1016/j.lwt.2017.04.063
Chen S-Q, Mikkelsen D, Lopez-Sanchez P, Wang D, Martinez-Sanz M, Gilbert EP, Flanagan BM, Gidley MJ (2017b) Characterisation of bacterial cellulose from diverse Komagataeibacter strains and their application to construct plant cell wall analogues. Cellulose 24:1211–1226. https://doi.org/10.1007/s10570-017-1203-3
Cheng KC, Catchmark JM, Demirci A (2009) Enhanced production of bacterial cellulose by using a biofilm reactor and its material property analysis. J Biol Eng 3:1–10. https://doi.org/10.1186/1754-1611-3-12
Choi CN, Song HJ, Kim MJ, Chang MH, Kim SJ (2009) Properties of bacterial cellulose produced in a pilot-scale spherical type bubble column bioreactor. Korean J Chem Eng 26:136–140. https://doi.org/10.1007/s11814-009-0021-1
Choi SM, Rao KM, Zo SM, Shin EJ, Han SS (2022) Bacterial cellulose and its applications. Polymers 14:1080. https://doi.org/10.3390/polym14061080
Czaja W, Romanovicz D, Brown RM (2004) Structural investigations of microbial cellulose produced in stationary and agitated culture. Cellulose 11:403–411. https://doi.org/10.1023/B:CELL.0000046412.11983.61
de Souza KC, Trindade NM, de Amorim JDP, do Nascimento HA, Costa AFS, Henrique MA, Caetano VF, Sarubbo LA, Vinhas GM (2021) Kinetic study of a bacterial cellulose production by Komagataeibacter Rhaeticus using coffee grounds and sugarcane molasses. Mater Res 24(3). https://doi.org/10.1590/1980-5373-mr-2020-0454
Deutsche Melasse Handelsgesellschaft (2015) Vinasse. https://www.deutsche-melasse.de/wp/wp-content/uploads/produkte/vinasse/2-Vinasse-EN-DMH-ES2SG.pdf. Accessed 20 Oct 2022
Einfeldt L, Klemm D (1997) The control of cellulose biosynthesis by Acetobacter Xylinum in view of molecular weight and molecular weight distribution part I: change of molecular weight of bacterial cellulose by simple variation of culture conditions 1. J Carbohydr Chem 16:635–646. https://doi.org/10.1080/07328309708007341
Embuscado ME, Marks JS, BeMiller JN (1994) Bacterial cellulose. I. Factors affecting the production of cellulose by Acetobacter xylinum. Food Hydrocoll 8:407–418. https://doi.org/10.1016/S0268-005X(09)80084-2
Fang L, Catchmark JM (2014) Structure characterization of native cellulose during dehydration and rehydration. Cellulose 21:3951–3963. https://doi.org/10.1007/s10570-014-0435-8
Fijałkowski K, Żywicka A, Drozd R, Kordas M, Rakoczy R (2016) Effect of Gluconacetobacter xylinus cultivation conditions on the selected properties of bacterial cellulose. Polish J Chem Technol 18:117–123. https://doi.org/10.1515/pjct-2016-0080
Fleury B, Abraham E, De La Cruz JA, Chandrasekar VS, Senyuk B, Liu Q, Cherpak V, Park S, ten Hove JB, Smalyukh II (2020) Aerogel from sustainably grown bacterial cellulose pellicles as a thermally insulative film for building envelopes. ACS Appl Mater Interfaces 12:34115–34121. https://doi.org/10.1021/acsami.0c08879
French AD (2014) Idealized powder diffraction patterns for cellulose polymorphs. Cellulose 21:885–896. https://doi.org/10.1007/s10570-013-0030-4
French AD (2020) Increment in evolution of cellulose crystallinity analysis. Cellulose 27:5445–5448. https://doi.org/10.1007/s10570-020-03172-z
Gribkova IN, Eliseev MN, Belkin YD, Zakharov MA, Kosareva OA (2021) The influence of biomolecule composition on colloidal beer structure. Biomolecules 12:24. https://doi.org/10.3390/biom12010024
Guo J, Catchmark JM (2012) Surface area and porosity of acid hydrolyzed cellulose nanowhiskers and cellulose produced by Gluconacetobacter xylinus. Carbohydr Polym 87:1026–1037. https://doi.org/10.1016/j.carbpol.2011.07.060
Gwon H, Park K, Chung S-C, Kim R-H, Kang JK, Ji SM, Kim N-J, Lee S, Ku J-H, Do EC, Park S, Kim M, Shim WY, Rhee HS, Kim J-Y, Kim J, Kim TY, Yamaguchi Y, Iwamuro R, Saito S, Kim G, Jung I-S, Park H, Lee C, Lee S, Jeon WS, Jang WD, Kim HU, Lee SY, Im D, Doo S-G, Lee SY, Lee HC, Park JH (2019) A safe and sustainable bacterial cellulose nanofiber separator for lithium rechargeable batteries. Proc Natl Acad Sci USA 116:19288–19293. https://doi.org/10.1073/pnas.1905527116
Ha JH, Shehzad O, Khan S, Lee SY, Park JW, Khan T, Park JK (2008) Production of bacterial cellulose by a static cultivation using the waste from beer culture broth. Korean J Chem Eng 25:812–815. https://doi.org/10.1007/s11814-008-0134-y
Harirchi S, Etemadifar Z, Yazdian F, Taherzadeh MJ (2020) Efficacy of polyextremophilic Aeribacillus pallidus on bioprocessing of beet vinasse derived from ethanol industries. Bioresour Technol 313:123662. https://doi.org/10.1016/j.biortech.2020.123662
Hernández J, Viana MT, Lastra M, Matus de la Parra A, Toledo-Agüero P (2019) The possible use of beet-vinasse as carbohydrate replacer in formulated diets for the juvenile abalone, Haliotis tuberculata. J Appl Aquac 31:271–288. https://doi.org/10.1080/10454438.2018.1547672
Hestrin S, Schramm M (1954) Synthesis of cellulose by Acetobacter xylinum. 2. Preparation of freeze-dried cells capable of polymerizing glucose to cellulose. Biochem J 58:345–352. https://doi.org/10.1042/bj0580345
Hornung M, Ludwig M, Gerrard AM, Schmauder H-P (2006) Optimizing the production of bacterial cellulose in surface culture: evaluation of substrate and mass transfer influences on the bioreaction (Part I). Eng Life Sci 6:537–545. https://doi.org/10.1002/elsc.200620162
Hur DH, Rhee H-S, Lee JH, Shim WY, Kim TY, Lee SY, Park JH, Jeong KJ (2020) Enhanced production of cellulose in Komagataeibacter xylinus by preventing insertion of IS element into cellulose synthesis gene. Biochem Eng J 156:107527. https://doi.org/10.1016/j.bej.2020.107527
Husemann VE, Werner R (1963) Cellulosesynthese durch Acetobacter xylinum. I. Über Molekulargewicht und Molekulargewichtsverteilung von Bakteriencellulose in Abhängigkeit von der Synthesedauer. Die Makromol Chemie 59:43–60. https://doi.org/10.1002/macp.1963.020590104
Hyun JY, Mahanty B, Kim CG (2014) Utilization of makgeolli sludge filtrate (MSF) as Low-cost substrate for bacterial cellulose production by Gluconacetobacter xylinus. Appl Biochem Biotechnol 172:3748–3760. https://doi.org/10.1007/s12010-014-0810-9
Iwata T, Indrarti L, Azuma JI (1998) Affinity of hemicellulose for cellulose produced by Acetobacter xylinum. Cellulose 5:215–228. https://doi.org/10.1023/A:1009237401548
Jang WD, Kim TY, Kim HU, Shim WY, Ryu JY, Park JH, Lee SY (2019) Genomic and metabolic analysis of Komagataeibacter xylinus DSM 2325 producing bacterial cellulose nanofiber. Biotechnol Bioeng 116:3372–3381. https://doi.org/10.1002/bit.27150
Jung H-I, Jeong J-H, Lee O-M, Park G-T, Kim K-K, Park H-C, Lee S-M, Kim Y-G, Son H-J (2010a) Influence of glycerol on production and structural–physical properties of cellulose from Acetobacter sp. V6 cultured in shake flasks. Bioresour Technol 101:3602–3608. https://doi.org/10.1016/j.biortech.2009.12.111
Jung H-I, Lee O-M, Jeong J-H, Jeon Y-D, Park K-H, Kim H-S, An W-G, Son H-J (2010b) Production and characterization of cellulose by Acetobacter sp. V6 using a cost-effective molasses-corn steep liquor medium. Appl Biochem Biotechnol 162:486–497. https://doi.org/10.1007/s12010-009-8759-9
Keshk S, Sameshima K (2006) The utilization of sugar cane molasses with/without the presence of lignosulfonate for the production of bacterial cellulose. Appl Microbiol Biotechnol 72:291–296. https://doi.org/10.1007/s00253-005-0265-6
Khan T, Hyun SH, Park JK (2007) Production of glucuronan oligosaccharides using the waste of beer fermentation broth as a basal medium. Enzyme Microb Technol 42:89–92. https://doi.org/10.1016/j.enzmictec.2007.08.007
Khattak WA, Khan T, Ul-Islam M, Wahid F, Park JK (2015) Production, characterization and physico-mechanical properties of bacterial cellulose from industrial wastes. J Polym Environ 23:45–53. https://doi.org/10.1007/s10924-014-0663-x
Klemm D, Cranston ED, Fischer D, Gama M, Kedzior SA, Kralisch D, Kramer F, Kondo T, Lindström T, Nietzsche S, Petzold-Welcke K, Rauchfuß F (2018) Nanocellulose as a natural source for groundbreaking applications in materials science: today’s state. Mater Today 21:720–748. https://doi.org/10.1016/j.mattod.2018.02.001
Lei W, Jin D, Liu H, Tong Z, Zhang H (2020) An Overview of bacterial cellulose in flexible electrochemical energy storage. ChemSusChem 13:3731–3753. https://doi.org/10.1002/cssc.202001019
Lin D, Lopez-Sanchez P, Li R, Li Z (2014) Production of bacterial cellulose by Gluconacetobacter hansenii CGMCC 3917 using only waste beer yeast as nutrient source. Bioresour Technol 151:113–119. https://doi.org/10.1016/j.biortech.2013.10.052
Liu M, Liu L, Jia S, Li S, Zou Y, Zhong C (2018) Complete genome analysis of Gluconacetobacter xylinus CGMCC 2955 for elucidating bacterial cellulose biosynthesis and metabolic regulation. Sci Rep 8:6266. https://doi.org/10.1038/s41598-018-24559-w
Marques WL, Raghavendran V, Stambuk BU, Gombert AK (2016) Sucrose and Saccharomyces cerevisiae: a relationship most sweet. FEMS Yeast Res 16:fov107. https://doi.org/10.1093/femsyr/fov107
Martínez-Sanz M, Lopez-Sanchez P, Gidley MJ, Gilbert EP (2015) Evidence for differential interaction mechanism of plant cell wall matrix polysaccharides in hierarchically-structured bacterial cellulose. Cellulose 22:1541–1563. https://doi.org/10.1007/s10570-015-0614-2
Marx-Figini M, Pion BG (1974) Kinetic invetigations of biosynthesis of cellulose by Actobacter xylinum. Biochim Biophys Acta Gen Subj 338:382–393. https://doi.org/10.1016/0304-4165(74)90299-2
Masaoka S, Ohe T, Sakota N (1993) Production of cellulose from glucose by Acetobacter xylinum. J Ferment Bioeng 75:18–22. https://doi.org/10.1016/0922-338X(93)90171-4
McMurry J, Begley T (2006) Organische Chemie der biologischen Stoffwechselwege, 1st edn. Spektrum Akademischer Verlag, Munich
Mohite BV, Kamalja KK, Patil SV (2012) Statistical optimization of culture conditions for enhanced bacterial cellulose production by Gluconoacetobacter hansenii NCIM 2529. Cellulose 19:1655–1666. https://doi.org/10.1007/s10570-012-9760-y
Mohite BV, Salunke BK, Patil SV (2013) Enhanced production of bacterial cellulose by using Gluconacetobacter hansenii NCIM 2529 strain under shaking conditions. Appl Biochem Biotechnol 169:1497–1511. https://doi.org/10.1007/s12010-013-0092-7
Moon S-H, Park J-M, Chun H-Y, Kim S-J (2006) Comparisons of physical properties of bacterial celluloses produced in different culture conditions using saccharified food wastes. Biotechnol Bioprocess Eng 11:26–31. https://doi.org/10.1007/BF02931864
Naritomi T, Kouda T, Yano H, Yoshinaga F (1998) Effect of ethanol on bacterial cellulose production from fructose in continuous culture. J Ferment Bioeng 85:598–603. https://doi.org/10.1016/S0922-338X(98)80012-3
Nishiyama Y, Sugiyama J, Chanzy H, Langan P (2003) Crystal structure and hydrogen bonding system in cellulose Iα from synchrotron X-ray and neutron fiber diffraction. J Am Chem Soc 125:14300–14306. https://doi.org/10.1021/ja037055w
Okajima K, Matsuda Y, Kamide K (1991) Study on change in the degree of polymerisation of bacterial cellulose produced by Acetobacter xylinum during its cultivation. Polym Int 25:145–151. https://doi.org/10.1002/pi.4990250303
Ono Y, Tanaka R, Funahashi R, Takeuchi M, Saito T, Isogai A (2016) SEC–MALLS analysis of ethylenediamine-pretreated native celluloses in LiCl/N, N-dimethylacetamide: softwood kraft pulp and highly crystalline bacterial, tunicate, and algal celluloses. Cellulose 23:1639–1647. https://doi.org/10.1007/s10570-016-0948-4
Palla G (1983) Characterization of the main secondary components of the liquid sugars from cane molasses. J Agric Food Chem 31:545–548. https://doi.org/10.1021/jf00117a020
Palmonari A, Cavallini D, Sniffen CJ, Fernandes L, Holder P, Fagioli L, Fusaro I, Biagi G, Formigoni A, Mammi L (2020) Short communication: characterization of molasses chemical composition. J Dairy Sci 103:6244–6249. https://doi.org/10.3168/jds.2019-17644
Park JK, Jung JY, Park YH (2003) Cellulose production by Gluconacetobacter hansenii in a medium containing ethanol. Biotechnol Lett 25:2055–2059. https://doi.org/10.1023/B:BILE.0000007065.63682.18
Park S, Baker JO, Himmel ME, Parilla PA, Johnson DK (2010) Cellulose crystallinity index: measurement techniques and their impact on interpreting cellulase performance. Biotechnol Biofuels 3:10. https://doi.org/10.1186/1754-6834-3-10
Park SM, Yoon SJ, Son HJ, Lee CY, Kim HS (2010) Properties of bacterial cellulose cultured in different carbon sources. Polymer (Korea) 34:522–526
Parnaudeau V, Condom N, Oliver R, Cazevieille P, Recous S (2008) Vinasse organic matter quality and mineralization potential, as influenced by raw material, fermentation and concentration processes. Bioresour Technol 99:1553–1562. https://doi.org/10.1016/j.biortech.2007.04.012
Potthast A, Radosta S, Saake B, Lebioda S, Heinze T, Henniges U, Isogai A, Koschella A, Kosma P, Rosenau T, Schiehser S, Sixta H, Strlič M, Strobin G, Vorwerg W, Wetzel H (2015) Comparison testing of methods for gel permeation chromatography of cellulose: coming closer to a standard protocol. Cellulose 22:1591–1613. https://doi.org/10.1007/s10570-015-0586-2
Premjet S, Ohtani Y, Sameshima K (1996) X-ray diffraction diagram of the bacterial cellulose membrane produced by Acetobacter xylinum in the medium with lignosulfonate. J Fiber Sci Technol 52:169–174. https://doi.org/10.2115/fiber.52.169
Premjet S, Premjet D, Ohtani Y (2007) The effect of ingredients of sugar cane molasses on bacterial cellulose production by Acetobacter xylinum ATCC 10245. J Fiber Sci Technol 63:193–199. https://doi.org/10.2115/fiber.63.193
Provin AP, dos Reis VO, Hilesheim SE, Bianchet RT, de Aguiar Dutra AR, Cubas ALV (2021) Use of bacterial cellulose in the textile industry and the wettability challenge—a review. Cellulose 28:8255–8274. https://doi.org/10.1007/s10570-021-04059-3
Ruan C, Zhu Y, Zhou X, Abidi N, Hu Y, Catchmark JM (2016) Effect of cellulose crystallinity on bacterial cellulose assembly. Cellulose 23:3417–3427. https://doi.org/10.1007/s10570-016-1065-0
Ryngajłło M, Jacek P, Cielecka I, Kalinowska H, Bielecki S (2019a) Effect of ethanol supplementation on the transcriptional landscape of bionanocellulose producer Komagataeibacter xylinus E25. Appl Microbiol Biotechnol 103:6673–6688. https://doi.org/10.1007/s00253-019-09904-x
Ryngajłło M, Kubiak K, Jędrzejczak-Krzepkowska M, Jacek P, Bielecki S (2019b) Comparative genomics of the Komagataeibacter strains—efficient bionanocellulose producers. MicrobiologyOpen 8:1–25. https://doi.org/10.1002/mbo3.731
Ryngajłło M, Jędrzejczak-Krzepkowska M, Kubiak K, Ludwicka K, Bielecki S (2020) Towards control of cellulose biosynthesis by Komagataeibacter using systems-level and strain engineering strategies: current progress and perspectives. Appl Microbiol Biotechnol 104:6565–6585. https://doi.org/10.1007/s00253-020-10671-3
Salari M, Sowti Khiabani M, Rezaei Mokarram R, Ghanbarzadeh B, Samadi Kafil H (2019) Preparation and characterization of cellulose nanocrystals from bacterial cellulose produced in sugar beet molasses and cheese whey media. Int J Biol Macromol 122:280–288. https://doi.org/10.1016/j.ijbiomac.2018.10.136
Segal L, Creely JJ, Martin AE, Conrad CM (1959) An empirical method for estimating the degree of crystallinity of native cellulose using the X-Ray diffractometer. Text Res J 29:786–794. https://doi.org/10.1177/004051755902901003
Shavyrkina NA, Skiba EA, Kazantseva AE, Gladysheva EK, Budaeva VV, Bychin NV, Gismatulina YA, Kashcheyeva EI, Mironova GF, Korchagina AA, Pavlov IN, Sakovich GV (2021) Static culture combined with aeration in biosynthesis of bacterial cellulose. Polymers 13:1–12. https://doi.org/10.3390/polym13234241
Shezad O, Khan S, Khan T, Park JK (2009) Production of bacterial cellulose in static conditions by a simple fed-batch cultivation strategy. Korean J Chem Eng 26:1689–1692. https://doi.org/10.1007/s11814-009-0232-5
Shezad O, Khan S, Khan T, Park JK (2010) Physicochemical and mechanical characterization of bacterial cellulose produced with an excellent productivity in static conditions using a simple fed-batch cultivation strategy. Carbohydr Polym 82:173–180. https://doi.org/10.1016/j.carbpol.2010.04.052
Shi QS, Feng J, Li WRU, Zhou G, Chen AIM, Ouyang YS, Chen YIB (2013) Effect of different conditions on the average degree of polymerization of bacterial cellulose produced by Gluconacetobacter intermedius BC-41. Cellul Chem Technol 47:503–508
Siller M, Ahn K, Pircher N, Rosenau T, Potthast A (2014) Dissolution of rayon fibers for size exclusion chromatography: a challenge. Cellulose 21:3291–3301. https://doi.org/10.1007/s10570-014-0356-6
Skiba EA, Gladysheva EK, Budaeva VV, Aleshina LA, Sakovich GV (2022) Yield and quality of bacterial cellulose from agricultural waste. Cellulose 29:1543–1555. https://doi.org/10.1007/s10570-021-04372-x
Son H-J, Heo M-S, Kim Y-G, Lee S-J (2001) Optimization of fermentation conditions for the production of bacterial cellulose by a newly isolated Acetobacter sp. A9 in shaking cultures. Biotechnol Appl Biochem 33:1. https://doi.org/10.1042/ba20000065
Sperotto G, Stasiak LG, Godoi JPMG, Gabiatti NC, De Souza SS (2021) A review of culture media for bacterial cellulose production: complex, chemically defined and minimal media modulations. Cellulose 28:2649–2673. https://doi.org/10.1007/s10570-021-03754-5
Stark JB (1961) The composition of beet molasses with particular reference to nitrogenous compounds. J Sugarbeet Res 11:507–514. https://doi.org/10.5274/jsbr.11.6.507
Strobin G, Wlochowicz A, Ciechanska D, Boryniec S, Struszczyk H, Sobczak S (2004) GPC studies on bacterial cellulose. Int J Polym Mater 53:889–900. https://doi.org/10.1080/009114030490502418
Tahara N, Tabuchi M, Watanabe K, Yano H, Morinaga Y, Yoshinaga F (1997) Degree of polymerization of cellulose from Acetobacter xylinum BPR2001 decreased by cellulase produced by the strain. Biosci Biotechnol Biochem 61:1862–1865. https://doi.org/10.1271/bbb.61.1862
Thygesen A, Oddershede J, Lilholt H, Thomsen AB, Ståhl K (2005) On the determination of crystallinity and cellulose content in plant fibres. Cellulose 12:563–576. https://doi.org/10.1007/s10570-005-9001-8
Tokoh C, Takabe K, Sugiyama J, Fujita M (2002) CP/MAS 13C NMR and electron diffraction study of bacterial cellulose structure affected by cell wall polysaccharides. Cellulose 9:351–360. https://doi.org/10.1023/A:1021150520953
Toyosaki H, Naritomi T, Seto A, Matsuoka M, Tsuchida T, Yoshinaga F (1995) Screening of bacterial cellulose-producing Acetobacter strains suitable for agitated culture. Biosci Biotechnol Biochem 59:1498–1502. https://doi.org/10.1271/bbb.59.1498
Tsouko E, Kourmentza C, Ladakis D, Kopsahelis N, Mandala I, Papanikolaou S, Paloukis F, Alves V, Koutinas A (2015) Bacterial cellulose production from industrial waste and by-product streams. Int J Mol Sci 16:14832–14849. https://doi.org/10.3390/ijms160714832
Tyagi N, Suresh S (2016) Production of cellulose from sugarcane molasses using Gluconacetobacter intermedius SNT-1: optimization & characterization. J Clean Prod 112:71–80. https://doi.org/10.1016/j.jclepro.2015.07.054
Uhlin KI, Atalla RH, Thompson NS (1995) Influence of hemicelluloses on the aggregation patterns of bacterial cellulose. Cellulose 2:129–144. https://doi.org/10.1007/BF00816385
Vazquez A, Foresti ML, Cerrutti P, Galvagno M (2013) Bacterial cellulose from simple and low cost production media by Gluconacetobacter xylinus. J Polym Environ 21:545–554. https://doi.org/10.1007/s10924-012-0541-3
Velásquez-Riaño M, Lombana-Sánchez N (2009) Cellulose production by Gluconacetobacter sp. GM5 in a static semi-continuous fermentation process using vinasse as culture media. Water Sci Technol 59:1195–1200. https://doi.org/10.2166/wst.2009.039
Velásquez-Riaño M, Lombana-Sánchez N, Villa-Restrepo AF, Fernández-Calle EP (2013) Cellulose production by Gluconacetobacter kakiaceti GM5 in two batch process using vinasse as culture media. Water Sci Technol 68:1079–1084. https://doi.org/10.2166/wst.2013.347
Volova TG, Prudnikova SV, Sukovatyi AG, Shishatskaya EI (2018) Production and properties of bacterial cellulose by the strain Komagataeibacter xylinus B-12068. Appl Microbiol Biotechnol 102:7417–7428. https://doi.org/10.1007/s00253-018-9198-8
Volova TG, Prudnikova SV, Kiselev EG, Nemtsev IV, Vasiliev AD, Kuzmin AP, Shishatskaya EI (2022) Bacterial cellulose (BC) and BC composites: production and properties. Nanomaterials 12:192. https://doi.org/10.3390/nano12020192
Wada M, Okano T, Sugiyama J (2001) Allomorphs of native crystalline cellulose I evaluated by two equatoriald-spacings. J Wood Sci 47:124–128. https://doi.org/10.1007/BF00780560
Wang S-S, Han Y-H, Chen J-L, Zhang D-C, Shi X-X, Ye Y-X, Chen D-L, Li M (2018) Insights into bacterial cellulose biosynthesis from different carbon sources and the associated biochemical transformation pathways in Komagataeibacter sp. W1. Polymers 10:963. https://doi.org/10.3390/polym10090963
Watanabe K, Tabuchi M, Morinaga Y, Yoshinaga F (1998) Structural features and properties of bacterial cellulose produced in agitated culture. Cellulose 5:187–200. https://doi.org/10.1023/A:1009272904582
Wohryzek O (1928) Chemie der Zuckerindustrie. Springer
Yao W, Weng Y, Catchmark JM (2020) Improved cellulose X-ray diffraction analysis using Fourier series modeling. Cellulose 27:5563–5579. https://doi.org/10.1007/s10570-020-03177-8
Yunoki S, Osada Y, Kono H, Takai M (2004) Role of ethanol in improvement of bacterial cellulose production: analysis using 13C-Labeled carbon sources. Food Sci Technol Res 10:307–313. https://doi.org/10.3136/fstr.10.307
Zhong C, Zhang G-C, Liu M, Zheng X-T, Han P-P, Jia S-R (2013) Metabolic flux analysis of Gluconacetobacter xylinus for bacterial cellulose production. Appl Microbiol Biotechnol 97:6189–6199. https://doi.org/10.1007/s00253-013-4908-8
Zhou LL, Sun DP, Hu LY, Li YW, Yang JZ (2007) Effect of addition of sodium alginate on bacterial cellulose production by Acetobacter xylinum. J Ind Microbiol Biotechnol 34:483–489. https://doi.org/10.1007/s10295-007-0218-4
Zou H, Chen N, Shi M, Xian M, Song Y, Liu J (2016) The metabolism and biotechnological application of betaine in microorganism. Appl Microbiol Biotechnol 100:3865–3876. https://doi.org/10.1007/s00253-016-7462-3
Acknowledgments
The authors are grateful to Marion Görke (Institute for Particle Technology, TU Braunschweig) for methodological discussion and execution of XRD measurements. We would also like to thank Nordzucker AG for providing molasses and vinasse and Hofbrauhaus Wolters for providing WBFB.
Funding
Open Access funding enabled and organized by Projekt DEAL. R. L. Heydorn, D. Lammers, and K. Dohnt received financial research support from Volkswagen AG.
Author information
Authors and Affiliations
Contributions
Conceptualization: RLH, KD, MG; Methodology: RLH, DL; Formal analysis and investigation: RLH, DL; Visualization: RLH; Writing—original draft preparation: RLH; Writing—review and editing: RLH, KD, DL, MG; Funding acquisition: KD; Supervision: KD, MG.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors have no financial or non-financial interest to disclose.
Ethical approval
Not applicable.
Consent for publication
All authors have approved the manuscript and agree with its submission.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Heydorn, R.L., Lammers, D., Gottschling, M. et al. Effect of food industry by-products on bacterial cellulose production and its structural properties. Cellulose 30, 4159–4179 (2023). https://doi.org/10.1007/s10570-023-05097-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10570-023-05097-9