
Abstract
Human disturbance and seawater intrusion have unpredictable effects on coastal species. Dune systems are crucial coastal habitats because they provide natural front protection against seawater influence. Adult plant stands in dunes are important since they maintain dune structure. This study evaluated salinity stress responses in adult plants of two key dune-building grasses differing in salt tolerance: Thinopyrum junceum (tolerant) and Calamagrostis arenaria (less tolerant). Cultivated plants were exposed to various seawater (SW) dilutions (Tap-Water, 6.25%, 12.5%, 25%, 50%, and 100%) and gas exchange measurements and oxidative stress biomarkers were determined after two months of treatment. Stress conditions were maintained until flowering to assess effects on the reproductive effort. The reproductive response showed high plasticity at various inflorescence stages in T. junceum, minimizing abortion processes. C. arenaria showed lower plasticity and higher abortion rates. Physiological traits responded similarly to salinity in both species, with greater effect on Fv/Fm decrease and non-photochemical quenching increase for T. junceum. Biochemical patterns of response were also similar, with increasing enzymatic activities at 25% SW treatment, mainly for glutathione reductase and peroxidase (GSH and GPx), and stress damage occurring at 50% and 100% SW treatments for increased malonaldehyde (MDA) levels. These findings provide insight into the salinity tolerance mechanisms of dune-building grasses. Higher salinity tolerance is suggested for T. junceum based on better reproductive effort. Higher juvenile tolerance, faster growth, and plant cycle adjustments are indicated as key traits for higher salinity tolerance of T. junceum in contrast to C. arenaria.
Similar content being viewed by others


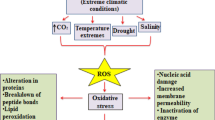
Introduction
Salinity is an important threat to global coastal ecosystems, as the increasing salinity impact significantly exerts its effects (Ondrasek and Rengel 2020). Dune systems stand out among coastal habitats, holding particular importance by serving as a buffer zone between marine waters and inland regions (van Puijenbroek et al. 2017). Various abiotic factors play pivotal roles in shaping the intricate plant communities that extend from the shoreline to the inland areas (Acosta et al. 2007; Maun 2009; Ciccarelli 2015). Within these communities, salinity is a significant factor influencing front plant communities such as embryonic dunes and dune ridges (Maun 2009; van Puijenbroek et al. 2017). This influence primarily stems from salt spray but also encompasses soil salinity due to events like water surges (direct seawater impact) and seawater intrusion (Maun 2009; Pauw et al. 2012; Ruocco et al. 2014; Cozzolino et al. 2017; Du and Hesp 2020). Future climate projections suggest that the impact of salinity may intensify due to an increased occurrence of storm events, rising sea levels, and anthropogenic pressures (Cozzolino et al. 2017; IPCC 2014). In Mediterranean dune systems, anthropogenic activities such as the removal of Posidonia oceanica L. "banquettes" have the potential to exacerbate the influence of seawater (Boudouresque et al. 2016; Del Vecchio et al. 2017).
Salinity significantly impacts plant development (Munns 2002). This influence primarily stems from two key effects: the osmotic effect, which limits water uptake, and the ionic effect, causing Na+ and Cl− accumulation and subsequent toxicity, inducing water stress and hampering photosynthesis (Munns and Tester 2008; Koyro et al. 2013). To alleviate the osmotic effect, plants employ stomatal closure and photosynthetic adaptations (Hameed et al. 2010; Sharma et al. 2012). However, excessive reductive power generated under limited photosynthesis and high irradiation can lead to overheating. This phenomenon is partially decreased through mechanisms such as non-photochemical quenching (NPQ) and structural leaf modifications. These processes are often accompanied by increased production of reactive oxygen species (ROS), potentially causing oxidative damage (Galmés et al. 2007; Arora et al. 2016). Plants activate physiological and biochemical adjustments to counteract ROS-induced damage and salinity-related stress. When ROS production escalates, enzymatic pathways like the ascorbate–glutathione (AsA-GSH) cycle, glutathione peroxidase (GPx) cycle, superoxide dismutase (SOD), and catalase (CAT) activity are the primary responders (Hasanuzzaman et al. 2012; Gupta et al. 2016). These mechanisms neutralize and reduce ROS, enhancing plant resilience to salinity-induced challenges (Munns 2011).
The physiological and biochemical repercussions of salt stress manifest as reduced carbon assimilation, modified carbon allocation, and perturbed regulatory processes (Guo et al. 2019). Reproduction is a pivotal phase in the plant life cycle, with salinity stress exerting a direct and often adverse impact (Boscaiu et al. 2005). Enhanced salt tolerance has primarily been associated with the capacity to maintain a low fruit abortion rate and minimize yield decline (Tang et al. 2022). However, reproductive plasticity, defined as the adjustment of flowering resource allocation under stress conditions, has also been identified as a tolerance trait (Yuan et al. 2019), even when flowering itself is inhibited (Naidoo and Mundree 1993). Overall, salinity tends to cause an adverse effect since carbon allocation can be disrupted, and ionic toxicity can cause functionality loss (i.e., pollen and ovule sterility), with the degree of this effect dependent on each species (Guo et al. 2019).
Coastal dunes progress from unpredictable strand conditions with front-line annual species to less mobile sand environments, including mobile dunes, semifixed dunes, and fixed dunes (Acosta et al. 2007). Embryonic dunes and dune ridge are crucial to dune formation and stability, especially along the Mediterranean and Atlantic coasts. For this geographical areas, Thinopyrum junceum (L.) Á.Löve. initiates embryonic dune formation, while Calamagrostis arenaria (L.) Roth. takes over as sand accumulates, forming dune ridges. With their intricate root systems these grasses aid in dune construction (Maun 2009; Fenu et al. 2013; van Puijenbroek et al. 2017; Bazzichetto et al. 2020). These dynamic areas also safeguard semifixed and fixed dune plant communities, making them critical for backdune preservation (Hesp 2002).
The impact of salt stress on key dune species C. arenaria and T. junceum remains relatively understudied, with limited insights into their tolerance (Sykes and Wilson 1989; Hilton et al. 2006; van Puijenbroek et al. 2017). Both species are recognized for their high resistance to salt spray (van Puijenbroek et al. 2017). For soil salinity, T. junceum has generally exhibited greater tolerance for vegetative growth than C. arenaria (Adriani and Terwindt 1974; Bruls et al. 2016; van Puijenbroek et al. 2017). Many studies focusing on salinity have focused on juvenile plants, with early establishment being a primary concern and experimental focus for both T. junceum and C. arenaria. The successful establishment of these species is paramount for proper dune development, representing a crucial phase for dune revegetation efforts (Huiskes 1979; Bruls et al. 2016). This significance is evident in numerous endeavors to restore dunes across coastal regions in the British Isles and the Netherlands, including the introduction of C. arenaria into dune systems worldwide, where it has become invasive (Konlechner et al. 2013; Pickart 2021). However, the ability to thrive at the adult stage and achieve reproductive success also constitutes essential objectives for plant restoration (Godefroid et al. 2011). Reproductive success through seed production serves as an indicator of potential self-sustaining population development, representing the final goal of plant restoration (Menges 2008; Godefroid et al. 2011). These latter aspects have received comparatively less attention in studies on dune-forming species.
For both species used in the present study, adult stage tolerance and reproductive response to salinity are overall unknown. Much of the plant response to salinity for C. arenaria and T. junceum is inferred from basic physiological response and biomass production, without further insight into other mechanisms. Considering the importance of these species in dune restoration and given the possible increasing salinity influence, the aims of the present study were (1) to analyze the effect of seawater salinity on the physiological and biochemical response of mature (capable to reproduce) dune-building grasses C. arenaria and T. junceum; (2) to assess how salinity stress affects the reproductive effort of C. arenaria and T. junceum; (3) to compare the overall response of both species to seawater stress, combining their reproductive, physiological, and biochemical response.
Materials and Methods
Plant Material and Experimental Design
Calamagrostis arenaria and Thinopyrum junceum plants were cultivated using seeds collected from Son Serra de Marina (Mallorca, Balearic Islands, Spain, UTM: ETRS89 31N EE2097). Several seeds were sown in each pot and only one individual was allowed to develop per pot. Since both species display different growth rates (slower in C. arenaria) species were cultivated differently to match the maturity stage in the same experimental set-up. For C. arenaria, 120 plants were cultivated in 2020 and maintained for one year, with two different approaches: 60 plants were maintained in forestry trays with 0.3 L capacity alveoli (Small Calamagrostis), to limit growth until the salinity experiment, and 60 were cultivated in 3 L capacity pots to favor plant development (Big Calamagrostis). One month prior to the salinity experiment, Small Calamagrostis were transplanted in 3 L capacity pots, and Big Calamagrostis were transplanted in 5 L capacity pots. By this means, both approaches ensured similar plant maturity (plants capable of flowering, mainly Big Calamagrostis), and the same plant size as compared to T. junceum (Small Calamagrostis). For T. junceum, only one set of 60 plants were germinated directly in 3 L capacity pots 5 months prior to the salinity experiment since reproductive maturity is achieved before one year due to fast growth under cultivated conditions (per. obs.). By this means, T. junceum plants were used for physiological-antioxidant measurements and then maintained for flowering assessment.
Individuals were randomly allocated to six seawater treatments (10 plants/treatment) (electric conductivity (EC in mS/cm)): Control-Tap Water (1.05), 6.25% SW (5.07), 12,5% SW (9.30), 25% SW (16.34), 50% SW (30.30), 100% SW (55.69), and each treatment was applied as described in Cerrato et al. (2022). Flowering time for both species begins between April and May, varying between localities. Based on previous trials in the experimental facility, salinity stress was applied to ensure two months of full treatment before measurements. Treatments were maintained during subsequent flowering to assess reproductive effort until the end of the experiment, when flowering and fruiting ended in August. For C. arenaria, full treatment began in February 2021, and for T. junceum, it began in March 2021.
Growing Conditions and Field Conductivity Measurements
Plants were cultivated as described in Cerrato et al. (2022) with 3 and 5 L pots with culture substrate composed of 61.50% coconut fiber, 33.00% white peat moss, and 5.50% of expanded perlite, fertilized with 4.40 mg/L of Osmocote NPK 19-10-19, a slow-release fertilizer. These conditions allowed proper plant development until the flowering stage for both species under the experimental conditions (smaller substrate volume compared to natural conditions) and allowed to mimic nutrient input from P. oceanica wracks in front communities (del Vecchio et al. 2020). Plants were maintained through all the growing and experimentation periods under a shade cloth (50% light exclusion) outdoors at the University of the Balearic Islands (Mallorca, Spain). Temperature and humidity conditions were not controlled, and individual plants were randomized within the facility. Shade cloth covered the plants but did not enclose the edges, allowing the wind to pass evenly throughout all the plants, permitting wind pollination (anemophily) as it naturally occurs for both species.
Seawater treatments were established by combining the corresponding proportion of seawater (collected in Sa Ràpita locality; Mallorca, Balearic Islands, Spain, UTM: ETRS89 31 N DD9657) with tap water from the experimental field facility. Unfiltered seawater stored in dark conditions for 2 months as indicated in Hanley et al. (2020) was used for salinity treatments. Watering was done until field capacity with a frequency that varied from weekly to 3 times per week based on soil moisture. Soil conductivity was periodically measured (XS Instruments Cond 51+) to ensure that soil salinity was maintained at the corresponding treatment conductivity value. If conductivity varied, all treatments were watered with tap water to remove excess salts, and then the treatment was subsequently applied to prevent recovery.
Conductivity measurements were conducted to understand salinity variations in the superficial root systems of the studied species and to determine if there were natural differences in salt exposure. Soil samples were collected at 17 m (for T. junceum) and 25 m (for C. arenaria) from the sea, and 15 cm depth, with a 15-day frequency during 2019, on natural populations of both species in the dunes of Sa Ràpita (Mallorca, Balearic Islands, Spain, UTM: ETRS89 31 N DD9657). Soil conductivity was measured by diluting soil samples in distilled water in a 1:5 ratio with a magnetic shaker for 2 h. Samples were then filtered, and conductivity measured (XS Instruments Cond 51+).
Reproductive Measurements
Reproductive traits were evaluated in T. junceum and C. arenaria (Big Calamagrostis). The total number of panicles produced per plant was recorded for each treatment and species, and specific measurements recorded by selecting blindly random panicles. In T. junceum, 4 random panicles were blindly selected per plant, and total length and fertile length (length comprising spikelets) were recorded. In C. arenaria, 9 panicles were selected to cover greater heterogeneity. Ratio Fertile/total length described decreased pedicel development. Since fertile length precedes panicle stem elongation, the ratio does not imply greater fertile length but rather delayed or insufficient nutrient supply for panicle stem growth. Thus, the ratio allows for describing general miniaturization (constant values) or uneven miniaturization during panicle development (increased ratio).
The number of spikelets per panicle was measured in 4 random panicles per plant in each species and standardized as the number of spikelets per cm of fertile inflorescence length. The same procedure was applied for the number of flowers in T. junceum (counting the number of flowers per spikelet). This measurement was not applied to C. arenaria since spikelets are uniflowered. For spikelet, glumes were measured in all spikelets of each of the 4 panicles randomly selected for T. junceum and 10 spikelets were randomly selected from each of 4 randomly selected panicles per plant in C. arenaria (due to a high number of spikelets). Seed-set was calculated for both species as the ratio of the number of fruits to the number of flowers per panicle in 4 random panicles per plant.
Gas Exchange and Fluorescence Measurements
Gas exchange measurements were conducted after two months of salt exposure for each species (including both sets of C. arenaria plants), to evaluate possible size effect on stress tolerance. Plants in each treatment were measured (n = 10) using an open gas exchange system with a coupled fluorescence chamber of 2 cm2 (Li-6400. Li-Cor Inc., Lincoln, USA). Measurements were conducted at light saturation of ~ 1500 µmol/m/s providing 400 µmol/mol CO2 with a flow rate of 300 µmol/s. Measurements were performed between 10:00 and 14:00 with humidity and temperature set to match environmental conditions. One young newly developed leaf was used in each measurement, and after each measurement, digital images were taken and measured by image analysis (Fiji software; Schindelin et al. 2012) to correct for leaf area. For each measured plant, the following parameters were measured: Net assimilation (An), stomatal conductance (gs), intercellular CO2 (Ci), and water use efficiency (WUE).
Fluorescence-related parameters were measured as indicated in Flexas et al. (2002). The PSII photochemical efficiency (PhiPS2) and electron transport rate (ETR) were calculated simultaneously with gas exchange measurements. Maximum quantum efficiency of PSII (Fv/Fm) was measured after plants were maintained under dark conditions for at least 4 h. Non-photochemical quenching (NPQ) and ratio between electron transport rate and assimilation ratio (ETR/An) were also calculated as indicated in Flexas et al. (2002). The latter ratio was used to check, in Control-Tap water plants, for both proper well-functioning of the equipment and optimal photosynthetic status of control plants at the onset of the experiment, as described in Perera-Castro and Flexas (2023).
Sample Processing for Biochemical Assays
After two months of salinity stress, leaf samples (n = 10) from T. junceum and C. arenaria (Small Calamagrostis) were collected and stored at – 80 °C until further processing. Leaves were homogenized in 50 mM Tris HCL buffer and 1 mM ethylenediaminetetraacetic acid (EDTA) at pH 7.5, in a proportion of 1:5 (weight: volume). Samples were then centrifuged for 10 min at 10000×g and 4 °C to remove cell debris, nuclei, and mitochondria from the supernatant, which was maintained at − 80 °C until biochemical analysis. Enzymatic activities were determined using a Shimadzu UV-2100 spectrophotometer at 25 °C, and lipid peroxidation was determined in a Bio-Tek PowerWave XS microplate spectrophotometer. All biochemical results were normalized per mg of protein using the colorimetric kit Biorad®, using Bovine Serum albumin (BSA) as a standard.
Antioxidant Enzyme Activities
Catalase (CAT) (EC 1.11.1.6) activity was determined according to the method described by Aebi (1984); results are expressed as mK(s−1)/mg protein. Superoxide dismutase (SOD) (EC 1.15.1.1) was determined according to Flohé and Otting (1984); results are presented as pKat/mg protein. Glutathione reductase (GRd) (EC 1.8.1.7) as indicated in Goldberg and Spooner (1984); results are presented as pKat/mg protein. Glutathione peroxidase (GPx) (EC 1.11.1.9) activity was determined by adapting the method described by Flohé and Gunzler (1984); results are presented as nKat/mg protein.
Lipid Peroxidation Assay
Malondialdehyde (MDA) levels were used as a lipid peroxidation indicator. MDA concentration was measured by a colorimetric assay based on the specific reaction of MDA with a reagent to produce a stable chromophore with a maximum absorbance at 586 nm. N-methyl-2-phenindole (10.3 mM) was added to samples in acetonitrile: methanol (3:1). After that, HCl 12 N was added, and the samples were incubated for 1 h at 45 °C. MDA concentration was calculated using a standard curve of known concentration at 586 nm.
Statistical Analysis
Data analysis was conducted using the statistical software R (R Core Team 2021). Continuous data, such as physiological and biochemical parameters, were analyzed using linear models (LM), while count and proportional data were modeled using generalized linear models (GLM, with quasipoisson and Binomial families, respectively). For reproductive parameters, mixed models were implemented with plant as a random effect using the lmer function (package: lme4, Bates et al. 2015) and models were evaluated using the lmertest function (package: lmerTest, Kuznetsova et al. 2017). Variability explained by simple models was evaluated using qqplots (package mgcviz; Fasiolo et al. 2019) and R-squared. When R-squared was not possible to compute, MacFadden pseudoR2 was used (package pscl; Jackman 2020). The effect of the salinity treatment among the response variables was evaluated using the analysis of variance (ANOVA). Two-way ANOVAs were carried out with treatment and species as main factors, and additionally, when interaction was not significative, one-way ANOVAs were carried out for each species to test treatment effect. When ANOVA conditions were not fulfilled, differences were evaluated using the Kruskal–Wallis test (McKight and Najab 2010) or Scheirer Ray Hare (package: rcompanion, Mangiafico 2022). Differences among treatments were evaluated using the Tukey Honest significance test (Abdi and Williams 2010) or Dunn-test when required (Dinno 2017).
Results
The mean field salinity measurements (Supplementary data: Table 1) ranged from 0.039 to 1.46 mS/cm, which were far below the lowest experimental treatment of 5.07 mS/cm for 6.25% seawater. The C. arenaria site displayed more constant values throughout the year, with maximum values in September–October and minimum values in November–December. T. junceum site displayed a similar pattern with maximum and minimum values during the same periods; however, conductivity values were overall more variable and 2 to 6 times higher than in the C. arenaria site.
Reproductive Measurements
All reproductive parameters showed significant interaction between both factors implying different response patterns on C. arenaria and T. junceum regarding treatment effect (Supplementary data: Table 2). The number of panicles was mainly affected in T. junceum, where a strong reduction occurred at 12.5% SW (40% decrease) and 25% SW and 50% SW (60–70% decrease), while C. arenaria showed constant production (less than 30% reduction until 50%SW treatment). Both species showed a high decrease at 100% SW with almost null panicle production (reduction: 80% in C. arenaria and 97% in T. junceum) (Fig. 1). Panicle total length decreased progressively in T. junceum starting at 25% SW (22% decrease), while fertile-to-total length ratio showed similar results (less than 15% ratio increase), with a lack of differences between treatments (excluding 100% SW which was excluded from the analysis since number of panicles was insufficient for statistical treatment and resulting measurements yielded undefined patterns) (Fig. 1). In contrast, C. arenaria was negatively affected in both total length and ratio length, with abrupt decreases for total length at 25% SW (~ 20% reduction) and the remaining treatments (60–70% decrease), and a significant ratio increase at 50% SW and 100% SW (60% increase) (Fig. 1).

Seawater effect on panicle production for Calamagrostis arenaria and Thinopyrum junceum. a Total number of panicles per plant (T. junceum n = 10; C. arenaria n = 12). b Panicle total length (T. junceum n = 40; C. arenaria n = 95–75; 100% SW: 34). c Fertile length/Total length relation (T. junceum n = 40; C. arenaria n = 95–75; 100% SW: 34)
Panicle reproductive effort was moderately affected in T. junceum with the smallest spikes occurring at 50% SW (maximum 25% decrease regarding control) for both spike length (Fig. 2a) and spikes per cm of fertile length (Supplementary data: Table 2). C. arenaria maintained a similar spikelet length and production efficiency (spikes per cm) remained stable until 12.5% SW (~ 20% decrease) and significantly decreased at a 25% SW and 50% SW (~ 40 decrease) (Fig. 2b). In both T. junceum and C. arenaria, 100% SW could be discarded for negligible spikelet production and quality. Fertility measured as seed-set was affected in both species. Both taxa showed a modest decrease until 12.5% SW (~ 20% decrease), with the further decreases occurring at 25% SW and 50% SW. For C. arenaria, this decrease implied strong reduction at a 25% SW (70% seed-set reduction) and almost absence of seed production at 50% SW, while T. junceum decreased less abruptly at 25% SW (33% reduction) and strongly at 50% SW (~ 70% reduction) maintaining some seed production (Fig. 2c).

Seawater effect on reproductive traits for Calamagrostis arenaria and Thinopyrum junceum. a Spikelet length (T. junceum n = 418–371; C. arenaria n = 180–205; 50% SW:110). b Number of Spikelets/cm (T. junceum n = 10; C. arenaria n = 12). c Seed-set (T. junceum n = 40; C. arenaria n = 40; 50% SW: 22)
Physiological Measurements
A comparison between T. junceum and C. arenaria showed similar patterns for all physiological parameters (Supplementary data: Table 3). There was a strong effect on gas exchange (Fig. 3). Overall assimilation rate and conductance were affected in both C. arenaria (LM: F = 99.919, df. = 1, p-value ≤ 0.001) and T. junceum (LM: F = 133.37, df. = 1, p-value = < 0.001) (gs: Table 3). Patterns were similar with a low (20–30% reduction) effect until 12.5% SW and an abrupt decrease at 25% SW (50–60% decrease) until null values at 100% SW. Intercellular CO2 was also affected by salinity treatment in both C. arenaria (LM: F = 28.876, df. = 1, p-value = < 0.001) and T. junceum (LM: F = 14.259, df. = 1, p-value ≤ 0.001) with a slight decrease until 12.5% SW and progressive increase for the following treatments. Ci results were similar between both species; however, increases at 50% SW (50% increase C. arenaria, 20% T. junceum) and 100% SW (80% increase C. arenaria, 30% T. junceum) differed in intensity between species. WUE showed the inverse pattern of Ci, mimicking species differences.

Seawater effect on gas exchange measurements for Calamagrostis arenaria (3l) and Thinopyrum junceum. Assimilation rate (An), Intercellular CO2 (Ci), Stomatal conductance (gs), electron transport rate (ETR), ETR/Assimilation rate (ETR/An) (log scaled), and WUEi (An/gs). (n = 10)
For fluorescence, (Figs. 3 and 4) results showed similar patterns for all variables in each species. ETR for T. junceum showed a significant reduction with 50% SW (30% reduction), and an abrupt decrease occurred at 100% SW (~ 60% reduction). In contrast, C. arenaria displayed a more progressive decrease maintaining high similarity between treatments (maximum decrease at 100% SW with 30% reduction). ETR/A showed only a treatment effect (Supplementary data: Table 3, Fig. 3), with significant effect for C. arenaria (LM: F = 5.879, df. = 1, p-value = 0.002) and T. junceum (Kruskal–Wallis: X2 = 49.066, p-value ≤ 0.001). Differences among treatments showed similar patterns in both species, with a significant increase occurring at 25% SW with the increase being higher in C. arenaria (60% increase) than T. junceum (40%). Further increase occurred similarly for both species. Fv/Fm results showed significant effect of both treatment and species factor (Supplementary data: Table 3, Fig. 4) with C. arenaria displaying higher Fv/Fm values (steady 5% higher values in all treatments). For C. arenaria (LM: F = 13.635, df. = 1, p-value ≤ 0.001), the decrease started at 50% SW, but significant reduction occurred at 100% SW treatment implying overall a small 5% reduction. In T. junceum (LM: F = 54.537, df. = 1, p-value ≤ 0.001), the decrease followed a similar pattern but a significant decrease at 50% SW and 100% SW implying a small reduction. NPQ as Fv/Fm displayed equal patterns for both C. arenaria (LM: F = 17.188, df. = 1, p-value ≤ 0.001) and T. junceum (LM: F = 60.356, df. = 1, p-value ≤ 0.001), being overall higher in T. junceum (30–50% higher depending on treatment) and increasing for both taxa over 30% at 25% SW and further 60% at 100% SW (Supplementary data: Table 3, Fig. 4).

Seawater effect on Fluorescence-related measurements for Calamagrostis arenaria (3l) and Thinopyrum junceum. Fv/Fm and non-photochemical quenching (NPQ). (n = 10)
Regarding Big and Small Calamagrostis comparison, a lack of interaction was apparent for all parameters and species factor has equally no effect in all but ETR and PhiPS2 parameters (Supplementary data: Table 4). For gas exchange parameters when Big Calamagrostis was modeled alone, assimilation rate (LM: F = 34.337, df. = 1, p-value ≤ 0.001), gs (Kruskal–Wallis: X2 = 38.844, df. = 1, p-value ≤ 0.001), and Ci (LM: F = 8.02, df. = 1, p-value ≤ 0.001) showed similar patterns as Small Calamagrostis. For ETR (LM: F = 8.4, df. = 1, p-value = < 0.001), the pattern remained similar even though a greater effect seemed to occur in Big Calamagrostis. Similar tendencies occurred for Fv/Fm and NPQ parameters, but statistically both were not significantly affected by treatment factor (Fv/Fm: (Kruskal–Wallis: X2 = 3.350, df. = 1, p-value = 0.650; NPQ: Kruskal–Wallis: X2 = 6.517, df. = 1, p-value = 0.26).
Biochemical Measurements
Lack of interaction in both species showed similar patterns regarding seawater stress treatments (Supplementary data: Table 5, Fig. 5). For SOD activity, both species displayed significant effect with displaying C. arenaria higher activity (17.04% higher) than T. junceum, and treatment displayed a general effect, but no specific effect was observed for both C. arenaria (LM: F = 3.721, df. = 1, p-values = 0.058), and T. junceum (LM: F = 1.962, df. = 1, p-values = 0.167). GRd and GPx activities were constitutively higher in T. junceum (28 and 38% higher respectively), but similar patterns were observed in GRd for C. arenaria (LM: F = 18.970, df. = 1, p-value ≤ 0.001) and T. junceum (LM: F = 14.820, df. = 1, p-value ≤ 0.001), with similar values at low salinity treatments and an increase of 30–40% activity at 25% seawater treatment which was maintained at higher seawater levels. For GPx, a significant effect was observed for C. arenaria (LM: F = 5.717, df. = 1, p-value ≤ 0.001) increasing progressively (40–50% activity increase regarding unstressed plants) until 25% SW treated plants, and subsequently decreasing until basal levels at 100% SW treatment. A similar but weaker non-significant pattern occurred in T. junceum (LM: F = 2.226, df. = 1, p-value = 0.065). Regarding CAT activity, similar levels of activity were observed for both species, but C. arenaria showed lack of significant variation with salinity treatment (LM: F = 0.854, df. = 1, p-value = 0.518) in contrast to T. junceum (LM: F = 3.143, df. = 1, p-value = 0.015) with a peak increase of 40% activity at 25% seawater which is maintained at further levels (30–40% higher than controls). Regarding MDA levels (Fig. 6; Supplementary data: Table 5), both species display similar levels, with seawater displaying a similar effect (T. junceum: LM: F = 18.610, df. = 1, p-value = < 0.001; C. arenaria: LM: F = 32.747, df. = 1, p-value ≤ 0.001). A small increase (~ 10%) occurred at low salinity levels (until 12.5% SW) followed by a 30% increase at moderate to high levels (until 50% SW) and increasing over 50% (60% for C. arenaria) at 100%SW.

Seawater effect on antioxidant enzymatic activities for Calamagrostis arenaria and Thinopyrum junceum. Catalase (CAT), Superoxide dismutase (SOD), Glutathione peroxidase (GPx), and glutathione reductase (GRd). (n = 10)

Effect of seawater stress on malondialdehyde (MDA) levels of Calamagrostis arenaria and Thinopyrum junceum. (n = 10)
Discussion
Both species displayed similar physiological responses to salinity stress, showing little effect at low levels and partial effects at higher levels. These findings contrasted with previous results which reported differing salt tolerance between the two species (Bruls et al. 2016; van Puijenbroek et al. 2017). However, the biochemical results of the present experiment revealed distinct patterns, suggesting variations in their underlying antioxidant mechanisms. These differences in biochemical responses along with variations in reproductive plasticity and resource allocation strategies are considered key traits contributing to T. junceum success in colonizing more saline embryonic dunes.
Conductivity values observed in this study suggest that salinity is not a limiting factor under field conditions. Low conductivity values have been previously reported for dune systems (Kearney 1904; Maun 2009), especially where C. arenaria are established (Rozema et al. 1985; Bruls et al. 2016). However, differences in conductivity observed between the experimental set-up and field conditions could also be explained by other factors, such as sporadic flooding and increasing salinity influence at lower depths (Sykes and Wilson 1989; Cozzolino et al. 2017; van Puijenbroek et al. 2017). Even though the data in this study show low conductivity values, the complexity of directly comparing water and soil conductivity may result in an underestimation of the actual water salinity conductivity for the studied samples. Since seawater influence is expected to occur more frequently through surges and water intrusion (Pauw et al. 2012; Ruocco et al. 2014; Cozzolino et al. 2017), further in-depth studies are required to elucidate the true exposure to soil salinity in dune habitats and its evolution. The results do provide evidence suggesting that T. junceum stands are more saline than C. arenaria.
Similar physiological responses were measured in both species, with little effect at low salinity (12.5% SW) and a partial effect at mid-high salinity, and being unable to cope with 100% SW. Previous studies have suggested higher tolerance for T. junceum, both during plant growth (van Puijenbroek et al. 2017) and the germination and seedling stages (El-katony et al. 2015; Cardona 2019; del Vecchio et al. 2020). In contrast, C. arenaria has been indicated to display high salt sensitivity during germination and early development (Chergui et al. 2018) but tolerance to several stressful conditions in the adult stage (Konlechner et al. 2013). Since plant age can influence salt tolerance (Singh et al. 2021), results from this study may differ from previous research due to the resilience of adult plants. T. junceum early-stage high tolerance might explain its superior colonization of saline sites, particularly embryonic dunes, whereas adult C. arenaria plants struggle to establish in these areas (Pavlik 1983; El-katony et al. 2015).
Gas exchange measurements revealed that both taxa had high and similar water use efficiency (WUE) which is in accordance with previous studies under natural conditions (Alessio et al. 2004; Gratani et al. 2009). Long-term WUE differ, being higher in C. arenaria (Alessio et al. 2004). These results suggest distinct strategies during the adult stage: T. junceum adopts a stress-escaping strategy with summer deciduousness, and C. arenaria maintains an evergreen, stress-tolerant approach (Alessio et al. 2004). T. junceum fast growth and deciduous nature minimizes long-term stress-related issues, while C. arenaria leaves remain functional longer, implying the need for different strategies to deal with issues such as photoinhibition.
Fluorescence parameters exhibit similar patterns, but overall C. arenaria showed a smaller decrease in Fv/Fm and lower NPQ values, indicating fewer heat dissipation mechanisms. This suggested that T. junceum was more affected by salinity stress, as reflected in the decreasing Fv/Fm values with increasing salinity. Increased NPQ in T. junceum implied the possible activation of the xanthophyll cycle for thermal dissipation of excess energy (Galmés et al. 2007). In C. arenaria, the lower NPQ and stable Fv/Fm values can be explained by alternative anatomical and physiological adjustment strategies (Chergui et al. 2017; Aráujo et al. 2021), such as reduced chlorophyll content (Sorce et al. 2019) or leaf rolling (Kadioglu et al. 2012).
Excessive reactive oxygen species (ROS) production, commonly induced during salinity-triggered photoinhibition, initiates the antioxidant response (Arora et al. 2016). Superoxide dismutase (SOD), a pivotal enzyme in ROS defense, catalyzes the conversion of superoxide anions (O−2) into hydrogen peroxide (H2O2), subsequently detoxified to water by catalase (Bose et al. 2014; Leung 2018). Low SOD variation under stressful conditions has been related to several causes, such as isoenzyme variation, or long-term decrease in favor to more stable adjustments such as proline accumulation (Kartashov et al. 2008; Qureshi et al. 2013; Houmani et al. 2016). This occurs with T. junceum since catalase activation suggested elevated H2O2 formation, beginning at the same salinity levels as the decrease in flowering production. The absence of SOD and catalase activation in C. arenaria may be attributed to either the simultaneous loss of activity in both enzymes (Modarresi et al. 2013; Ghaderi et al. 2018) or a limited ROS production, as indicated by fluorescence parameters revealing reduced non-photochemical quenching (NPQ) and Fv/Fm variation. Previously documented leaf characteristics, such as rolling and reduced chlorophyll content, suggest a mechanism of photoinhibition and H2O2 avoidance, likely accomplished through physical protective adaptations and peroxisome-related processes (Abogadallah et al. 2010; Sorce et al. 2019).
Activation of GRd in both species suggested the possible initiation of the GRd-ASA cycle, a critical ROS detoxification pathway in plants (Hasanuzzaman et al. 2012). GPx serves dual roles in plants, acting as an antioxidant and regulatory enzyme (Bela et al. 2015). It catalyzes GSH oxidation, enabling H2O2 detoxification (Leng et al. 2015) and, along with GRd, regulates the vital GSH/GSSG ratio for cellular redox balance (Hasanuzzaman et al. 2017). The absence of GPx activation in T. junceum may be due to prolonged reduced enzyme activity or regulatory processes to increase glutathione levels. Conversely, in C. arenaria, concurrent activation of both GPx and GRd may respond to ROS production and elevated GSH levels, serving as a non-enzymatic antioxidant mechanism against stress. GSH enzymes seem associated with regulatory processes and ROS detoxification for both species, unrelated to photoinhibitory H2O2 (Cavalcanti et al. 2004).
Both species exhibited enzymatic mechanisms to mitigate salinity stress. Previous studies have emphasized the significance of other physiological and biochemical (non-enzymatic) mechanisms in preventing or addressing oxidative damage (Mouri et al. 2012; Bouchemal et al. 2022). Despite their specific mechanisms, both species appeared to experience ROS overproduction, as indicated by elevated levels of MDA at 100% SW, coinciding with reproductive failure and physiological stress (high ETR/An levels). MDA is a marker of oxidative damage, primarily associated with the failure to control excessive ROS production (Hernández and Almansa 2002). Elevated MDA levels occurred under natural conditions for both species and suggested a relatively weak antioxidant system (Sorce et al. 2019). In T. junceum, being a geophyte, stress may accelerate stasis through leaf senescence, with MDA serving as an indicator of this process and ROS overproduction due to salinity (Taulavuori et al. 2010; Liu et al. 2011). Conversely, for C. arenaria, high MDA levels may primarily relate to increased ROS levels.
The physiological and antioxidant responses indicated stress starting at 25% SW, with increasing photodamage related to salinity observed at 50% SW and 100% SW. Stress appeared to be influenced by the contrasting plant cycles of summer deciduous T. junceum and evergreen C. arenaria. Regarding reproductive structures, the effects of salinity result in decreased resource allocation, potentially leading to miniaturization (e.g., smaller panicles in grasses), a reduction in the number of flowers (spikelets and spikes), and/or inhibition (a decreased number of inflorescences) (Abdullah et al. 2001; Boscaiu et al. 2005; Najla et al. 2009; Mazhar et al. 2012; Ma et al. 2020). T. junceum exhibited both a decreased number of panicles and reduced size with increasing salinity, displaying high plasticity at early growth stages. However, inflorescences maintained efficient spikelet production relative to panicle size. In C. arenaria, panicle production exhibited low plasticity (minimal variation among treatments), but spikelet reduction was more pronounced concerning panicle size and less efficient compared to T. junceum.
Allometric relations under stress have traditionally considered that sensitive plants tend to display higher plasticity, while tolerant (halophytes) maintain steady reproductive effort (Aronson et al. 1993; Tang et al. 2022). Equally, better relocation of resources to flowers and seeds, avoiding or minimizing abortion process is a main difference between halophytes and non-halophytes (Yuan et al. 2019). For halophytic dune species such as Cakile maritima, higher salt tolerance can be defined considering a trade-off between efficient resource investment and low degree of abortion (Davy et al. 2006; Debez et al. 2008). This idea suggests that higher tolerance could be granted to T. junceum, since inflorescence production is better adjusted to stress status with abortion occurring at late stages of inflorescence production. In contrast, C. arenaria displayed early and late abortion (panicle, spikelet, and seed abortion) suggesting lower reproductive adaptation to stress. Reproductive plasticity differences between both taxa imply difference in the capability to adjust reproductive response to stress conditions. Active stress during flowering period in contrast to vegetative growth causes increased abortion rate and lower plasticity because of lower reproductive investment prediction (Wankhade et al. 2013). For salinity, water absorption during flowering implies higher salt accumulation which can increase flower abortion and sterility (Abdulah et al. 2001).
For T. junceum and C. arenaria, two key differences can explain their reproductive response. T. junceum summer deciduousness implies low or absent overlap between growth and reproduction, implying better inflorescence investment prediction, while C. arenaria struggles to maintain both processes. The second key difference is related to growth rate differences and time to maturity. Since C. arenaria requires a longer period of growth until maturity, inflorescence production could be programmed long before flowering, thus being less receptive to recent stressful variations. This implies that T. junceum also benefits from a stress escape strategy during flowering, which allows further advantages not only for survival, but also for seed production and further recolonization of the embryonic and foredunes.
Conclusions
Both biochemical antioxidant and physiological gas exchange parameters, as well as flowering parameters, displayed high similarity between both species, with the stress effect becoming evident from 25% SW onwards. The traditionally observed higher salinity tolerance in T. junceum compared to C. arenaria, primarily in adult stages, can be attributed to a combination of plant cycle adjustments and a more rapid growth rate, leading to faster development to maturity. These traits, coupled with findings from previous studies on juvenile stages (indicating higher physiological tolerance in T. junceum), provide valuable insights into the colonization advantage of less stable environments. Specifically, the present results shed light on the success of T. junceum at the adult stage over C. arenaria, especially in the context of unstable and more seawater-influenced embryonic dunes. Therefore, it can be inferred that T. junceum ability to cope with salinity stress contributes to its competitive advantage and prevalence in such environments.
In conclusion, considering overall physiological and biochemical traits, both C. arenaria and T. junceum exhibit similar tolerance to seawater stress in the adult stage. However, the impact of salinity on reproductive effort differs significantly between these two species, which has implications for their respective colonization patterns in dune systems.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Abdi H, Williams LJ (2010) Tukey’s honestly significant difference (HSD) test. In: Salkind NJ, Dougherty DM, Frey B (eds) Encyclopedia of research design. Sage, Thouand Oaks, pp 583–585
Abdullah ZKMA, Khan MA, Flowers TJ (2001) Causes of sterility in seed set of rice under salinity stress. J Agron Crop Sci 187(1):25–32. https://doi.org/10.1046/j.1439-037X.2001.00500.x
Abogadallah GM, Serag MM, Quick WP (2010) Fine and coarse regulation of reactive oxygen species in the salt tolerant mutants of barnyard grass and their wild-type parents under salt stress. Physiol Plant 138(1):60–73. https://doi.org/10.1111/j.1399-3054.2009.01297.x
Acosta A, Ercole S, Stanisci A, Pillar VDP, Blasi C (2007) Coastal vegetation zonation and dune morphology in some Mediterranean ecosystems. J Coast Res 23(6):1518–1524. https://doi.org/10.2112/05-0589.1
Adriani MJ, Terwindt JH (1974) Sand stabilization and dune building. Rijkswaterstaat Commun 19:1
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Alessio GA, De Lillis M, Brugnoli E, Lauteri M (2004) Water sources and water-use efficiency in Mediterranean coastal dune vegetation. Plant Biol 6(03):350–357. https://doi.org/10.1055/s-2004-820882
Aráujo T, Beirão JNDC, Arruda AJVD, Cruz C (2021) The adaptive power of ammophila arenaria: biomimetic study, systematic observation, parametric design and experimental tests with bimetal. Polymers 13(15):2554. https://doi.org/10.3390/polym13152554
Aronson J, Kigel J, Shmida A (1993) Reproductive allocation strategies in desert and Mediterranean populations of annual plants grown with and without water stress. Oecologia 93(3):336–342. https://doi.org/10.1007/BF00317875
Arora D, Jain P, Singh N, Kaur H, Bhatla SC (2016) Mechanisms of nitric oxide crosstalk with reactive oxygen species scavenging enzymes during abiotic stress tolerance in plants. Free Radic Res 50(3):291–303. https://doi.org/10.3109/10715762.2015.1118473
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting Linear Mixed-Effects Models Using lme. J Stat Softw 67(1):1–48. https://doi.org/10.18637/jss.v067.i01
Bazzichetto M, Sperandii MG, Malavasi M, Carranza ML, Acosta ATR (2020) Disentangling the effect of coastal erosion and accretion on plant communities of Mediterranean dune ecosystems. Estuar Coast Shelf Sci 241:106758. https://doi.org/10.1016/j.ecss.2020.106758
Bela K, Horváth E, Gallé Á, Szabados L, Tari I, Csiszár J (2015) Plant glutathione peroxidases: emerging role of the antioxidant enzymes in plant development and stress responses. J Plant Physiol 176:192–201. https://doi.org/10.1016/j.jplph.2014.12.014
Boscaiu M, Estrelles E, Soriano P, Vicente O (2005) Effects of salt stress on the reproductive biology of the halophyte Plantago crassifolia. Biol Plant 49(1):141–143. https://doi.org/10.1007/s10535-005-1143-x
Bose J, Rodrigo-Moreno A, Shabala S (2014) ROS homeostasis in halophytes in the context of salinity stress tolerance. J Exp Bot 65(5):1241–1257. https://doi.org/10.1093/jxb/ert430
Bouchemal A, Slimani H, Djebbar R (2022) Antioxidant activities of plant species adapted to coastal dune of Zemmouri el Bahri (Algeria). Analele Universităţii Din Oradea 29(1):21–29
Boudouresque CF, Pergent G, Pergent-Martini C, Ruitton S, Thibaut T, Verlaque M (2016) The necromass of the Posidonia oceanica seagrass meadow: fate, role, ecosystem services and vulnerability. Hydrobiologia 781(1):25–42. https://doi.org/10.1007/s10750-015-2333-y
Bruls A, Hermens J, Huizing E, Tanson M, Verstraeten L (2016) Factors influencing dune growth and the vitality of dune building Ammophila arenaria. Ann Rev Mar Sci 1:117–141
Cardona C (2019) Tolerància germinativa a la salinitat d’espècies litorals de les Illes Balears. Phd dissertation. University of the Balearic Islands. Palma.
Cavalcanti FR, Oliveira JTA, Martins-Miranda AS, Viégas RA, Silveira JAG (2004) Superoxide dismutase, catalase and peroxidase activities do not confer protection against oxidative damage in salt-stressed cowpea leaves. New Phytol 163(3):563–571. https://doi.org/10.1111/j.1469-8137.2004.01139.x
Cerrato M, Ribas-Serra A, Cortés-Fernández I, Cardona Ametller C, Mir-Rosselló PM, Douthe C, Flexas J, Gil L, Sureda A (2022) Effect of seawater salinity stress on Sporobolus pungens (Schreb.) Kunth, a halophytic grass of the mediterranean embryonic dunes. Plant Growth Regul 98:191–204. https://doi.org/10.1007/s10725-022-00846-5
Chergui A, El Hafid L, Melhaoui M (2017) Characteristics of marram grass (Ammophila arenaria L.), plant of the coastal dunes of the Mediterranean Eastern Morocco: ecological, morpho-anatomical and physiological Aspects. J Mater Environ Sci 8:3759–3765
Chergui A, Zakariya I, Nejjari R, El Hafid L, Melhaoui M (2018) The effects of temperature, hydric and saline stress on the seedling growth of marram grass (Ammophila arenaria L.). Int J Appl Environ Sci 13(7):633–642
Ciccarelli D (2015) Mediterranean coastal dune vegetation: are disturbance and stress the key selective forces that drive the psammophilous succession? Estuar Coast Shelf Sci 165:247–253. https://doi.org/10.1016/j.ecss.2015.05.023
Cozzolino D, Greggio N, Antonellini M, Giambastiani BMS (2017) Natural and anthropogenic factors affecting freshwater lenses in coastal dunes of the Adriatic coast. J Hydrol 551:804–818. https://doi.org/10.1016/j.jhydrol.2017.04.039
Davy AJ, Scott R, Cordazzo CV (2006) Biological flora of the British isles: Cakile maritima Scop. J Ecol 94(3):695–711. https://doi.org/10.1111/j.1365-2745.2006.01131.x
Debez A, Koyro HW, Grignon C, Abdelly C, Huchzermeyer B (2008) Relationship between the photosynthetic activity and the performance of Cakile maritima after long-term salt treatment. Physiol Plant 133(2):373–385. https://doi.org/10.1111/j.1399-3054.2008.01086.x
Del Vecchio S, Jucker T, Carboni M, Acosta AT (2017) Linking plant communities on land and at sea: the effects of Posidonia oceanica wrack on the structure of dune vegetation. Estuar Coast Shelf Sci 184:30–36. https://doi.org/10.1016/j.ecss.2016.10.041
Del Vecchio S, Fantinato E, Roscini M, Acosta AT, Bacchetta G, Buffa G (2020) The germination niche of coastal dune species as related to their occurrence along a sea–inland gradient. J Veg Sci 31(6):1112–1121. https://doi.org/10.1111/jvs.12899
Dinno A (2017) Dunn.test: Dunn’s Test of Multiple Comparissons Using Rank Sums. R packages version 1.3.5. https://CRAN.Rproject.org/package=dunn.test
Du J, Hesp PA (2020) Salt spray distribution and its impact on vegetation zonation on coastal dunes: a review. Estuar Coast 43(8):1885–1907. https://doi.org/10.1007/s12237-020-00820-2
El-Katony TM, Khedr AHAF, Soliman NG (2015) Nutrients alleviate the deleterious effect of salinity on germination and early seedling growth of the psammophytic grass Elymus farctus. Botany 93(9):559–571. https://doi.org/10.1139/cjb-2015-0096
Fasiolo M, Nedellec R, Goude Y, Wood SN (2019) Scalable visualization methods for modern Generalized Additive Models. arXiv:1809.10632v2. https://doi.org/10.1080/10618600.2019.1629942
Fenu G, Carboni M, Acosta AT, Bacchetta G (2013) Environmental factors influencing coastal vegetation pattern: new insights from the Mediterranean Basin. Folia Geobot 48(4):493–508. https://doi.org/10.1007/s12224-012-9141-1
Flexas J, Bota J, Escalona JM, Sampol B, Medrano H (2002) Effects of drought on photosynthesis in grapevines under field conditions: an evaluation of stomatal and mesophyll limitations. Funt Plant Biol 29(4):461–471. https://doi.org/10.1071/PP01119
Flohé L, Günzler WA (1984) Assays of glutathione peroxidase. Methods in Enzymol 105:114–120. https://doi.org/10.1016/S0076-6879(84)05015-1
Flohé L, Otting F (1984) Superoxide dismutase assays. Methods Enzymol 105:93–104. https://doi.org/10.1016/S0076-6879(84)05013-8
Galmés J, Abadía A, Cifre J, Medrano H, Flexas J (2007) Photoprotection processes under water stress and recovery in Mediterranean plants with different growth forms and leaf habits. Physiol Plant 130(4):495–510. https://doi.org/10.1111/j.1399-3054.2007.00919.x
Ghaderi N, Mozafari A, Siosehmardeh A (2018) Change in antioxidant enzymes activity and some morpho-physiological characteristics of strawberry under long-term salt stress. Physiol Mol Biol Plants 24(5):833–843. https://doi.org/10.1007/s12298-018-0535-2
Godefroid S, Piazza C, Rossi G, Buord S, Stevens AD, Aguraiuja R, Cowelli C, Weekley CW, Vogg G, Iriondo M, Johnson I, Dixon B, Gordon D, Magnanon S, Valentino B, Bjureke K, Koopman R, Vicens M, Virevaire M, Vanderborght T (2011) How successful are plant species reintroductions? Biol Conserv 144(2):672–682. https://doi.org/10.1016/j.biocon.2010.10.003
Goldberg DM, Spooner RJ (1984) Glutathione reductase. In: Bergmeyer HU (ed) Methods in enzymatic analysis. Verlag Chemie, Basel, pp 258–265
Gratani L, Varone L, Crescente MF (2009) Photosynthetic activity and water use efficiency of dune species: the influence of air temperature on functioning. Photosynthetica 47(4):575–585. https://doi.org/10.1007/s11099-009-0083-7
Guo J, Dong X, Han G, Wang B (2019) Salt-enhanced reproductive development of Suaeda salsa L. coincided with ion transporter gene upregulation in flowers and increased pollen K+ content. Front Plant Sci 10:333. https://doi.org/10.3389/fpls.2019.00333
Gupta DK, Palma JM, Corpas FJ (eds) (2016) Redox state as a central regulator of plant-cell stress responses (p. 386). Springer International Publishing, Dordrecht
Hameed M, Ashraf M, Ahmad MSA, Naz N (2010) Structural and functional adaptations in plants for salinity tolerance. In Plant adaptation and phytoremediation (pp. 151–170). Springer, Dordrecht.
Hasanuzzaman M, Hossain MA, da Silva JAT, Fujita M (2012) Plant response and tolerance to abiotic oxidative stress: antioxidant defense is a key factor. In: Venkateswarlu B, Shanker AK, Shanker C, Maheswari M (eds) Crop stress and its management: perspectives and strategies,. Springer, Dordrecht, pp 261–315
Hasanuzzaman M, Nahar K, Anee TI, Fujita M (2017) Glutathione in plants: biosynthesis and physiological role in environmental stress tolerance. Physiol Mol Biol Pla 23(2):249–268. https://doi.org/10.1007/s12298-017-0422-2
Hernández JA, Almansa MS (2002) Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol Plant 115(2):251–257. https://doi.org/10.1034/j.1399-3054.2002.1150211.x
Hesp P (2002) Foredunes and blowouts: initiation, geomorphology and dynamics. Geomorphology 48(1–3):245–268. https://doi.org/10.1016/S0169-555X(02)00184-8
Hilton M, Harvey N, Hart A, James K, Arbuckle C (2006) The impact of exotic dune grass species on foredune development in Australia and New Zealand: a case study of Ammophila arenaria and Thinopyrum junceiforme. Aust Geogr 37(3):313–334. https://doi.org/10.1080/00049180600954765
Houmani H, Rodríguez-Ruiz M, Palma JM, Abdelly C, Corpas FJ (2016) Modulation of superoxide dismutase (SOD) isozymes by organ development and high long-term salinity in the halophyte Cakile maritima. Protoplasma 253(3):885–894. https://doi.org/10.1007/s00709-015-0850-1
Huiskes AHL (1979) Ammophila arenaria (L.) Link (Psamma arenaria (L.) Roem et Schult; Calamgrostis arenaria (L.) Roth). J Ecol 67(1):363–382. https://doi.org/10.2307/2259356
IPCC (2014) Climate change 2014: Synthesis report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva
Jackman S (2020) pscl: classes and Methods for R developed in the political science computational laboratory. United States Studies Centre University of Sydney, Sydney
Kadioglu A, Terzi R, Saruhan N, Saglam A (2012) Current advances in the investigation of leaf rolling caused by biotic and abiotic stress factors. Plant Sci 182:42–48. https://doi.org/10.1016/j.plantsci.2011.01.013
Kartashov AV, Radyukina NL, Ivanov YV, Pashkovskii PP, Shevyakova NI, Kuznetsov VV (2008) Role of antioxidant systems in wild plant adaptation to salt stress. Russ J Plant Physiol 55(4):463–468. https://doi.org/10.1134/S1021443708040055
Kearney TH (1904) Are plants of sea beaches and dunes true halophytes? Bot Gaz 37(6):424–436
Konlechner TM, Hilton MJ, Orlovich DA (2013) Accommodation space limits plant invasion: Ammophila arenaria survival on New Zealand beaches. J Coast Conserv 17(3):463–472. https://doi.org/10.1007/s11852-013-0244-5
Koyro HW, Hussain T, Huchzermeyer B, Khan MA (2013) Photosynthetic and growth responses of a perennial halophytic grass Panicum turgidum to increasing NaCl concentrations. Environ Exp Bot 91:22–29. https://doi.org/10.1016/j.envexpbot.2013.02.007
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 82(13):1–26. https://doi.org/10.18637/jss.v082.i13
Leng X, Jia H, Sun X, Shangguan L, Mu Q, Wang B, Fang J (2015) Comparative transcriptome analysis of grapevine in response to copper stress. Sci Rep 5(1):1–17. https://doi.org/10.1038/srep17749
Leung DW (2018) Studies of catalase in plants under abiotic stress. In Antioxidants and antioxidant enzymes in higher plants (pp. 27–39). Springer, Cham.
Liu C, Liu Y, Guo K, Fan D, Li G, Zheng Y, Yang Yu (2011) Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. EEB 71(2):174–183. https://doi.org/10.1016/j.envexpbot.2010.11.012
Ma X, Zhao Su, Ma H (2020) Molecular genetic analyses of abiotic stress responses during plant reproductive development. J Exp Bot 71(10):2870–2885. https://doi.org/10.1093/jxb/eraa089
Mangiafico S (2022) rcompanion: functions to Support. Extension Education Program Evaluation. R package version 2.4.15. https://CRAN.R-project.org/package=rcompanion
Maun MA (2009) The biology of coastal sand dunes. Oxford University Press, Oxford
Mazhar AA, Shedeed SI, Abdel-Aziz NG, Mahgoub MH (2012) Growth, flowering and chemical constituents of Chrysanthemum indicum L. plant in response to different levels of humic acid and salinity. J Appl Sci Res 8(7):3697–3706
McKight PE, Najab J (2010) Kruskal-wallis test. In: Weiner IB, Craighead WE (eds) The corsini encyclopedia of psychology. Wiley, New York
Menges ES (2008) Restoration demography and genetics of plants: when is a translocation successful? Aust J Bot 56(3):187–196. https://doi.org/10.1071/BT07173
Modarresi M, Nematzadeh GA, Moradian F (2013) Salinity response pattern and isolation of catalase gene from halophyte plant Aeluropus littoralis. Photosynthetica 51(4):621–629. https://doi.org/10.1007/s11099-013-0060-z
Mouri C, Benhassaini H, Bendimered FZ, Belkhodja M (2012) Variation saisonnière de la teneur en proline et en sucres solubles chez l’oyat (Ammophila arenaria (L.) Link) provenant du milieu naturel de la côte ouest de l’Algérie. Acta Botanica Gallica 159(1):127–135. https://doi.org/10.1080/12538078.2012.673822
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25(2):239–250. https://doi.org/10.1046/j.0016-8025.2001.00808.x
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Ann Rev Plant Biol 59:651–681. https://doi.org/10.1146/annurev.arplant.59.032607.092911
Munns R (2011) Plant adaptations to salt and water stress: differences and commonalities. Adv Bot Res 57:1–32. https://doi.org/10.1016/B978-0-12-387692-8.00001-1
Naidoo G, Mundree SG (1993) Relationship between morphological and physiological responses to waterlogging and salinity in Sporobolus virginicus (L.) Kunth. Oecologia 93(3):360–366. https://doi.org/10.1007/BF003178798y
Najla S, Vercambre G, Pages L, Grasselly D, Gautier H, Genard M (2009) Tomato plant architecture as affected by salinity: descriptive analysis and integration in a 3-D simulation model. Botany 87(10):893–904. https://doi.org/10.1139/B09-061
Ondrasek G, Rengel Z (2020) Environmental salinization processes: detection, implications & solutions. Sci of the Total Environ 754:142432. https://doi.org/10.1016/j.scitotenv.2020.142432
Pauw P, De Louw PG, Essink GO (2012) Groundwater salinisation in the Wadden Sea area of the Netherlands: quantifying the effects of climate change, sea-level rise and anthropogenic interferences. Neth J Geosci 91(3):373–383. https://doi.org/10.1017/S0016774600000500
Pavlik BM (1983) Nutrient and productivity relations of the dune grasses Ammophila arenaria and Elymus mollis. Oecologia 57(1):227–232
Perera-Castro A, Flexas J (2023) The ratio of electron transport to assimilation (ETR/AN): underutilized but essential for assessing both equipment’s proper performance and plant status. Planta 257(2):29. https://doi.org/10.1007/s00425-022-04063-2
Pickart AJ (2021) Ammophila invasion ecology and dune restoration on the West Coast of North America. Diversity 13(12):629. https://doi.org/10.3390/d13120629
Qureshi MI, Abdin MZ, Ahmad J, Iqbal M (2013) Effect of long-term salinity on cellular antioxidants, compatible solute and fatty acid profile of Sweet Annie (Artemisia annua L.). Phytochemistry 95:215–223. https://doi.org/10.1016/j.phytochem.2013.06.026
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/
Rozema J, Bijwaard P, Prast G, Broekman R (1985) Ecophysiological adaptations of coastal halophytes from foredunes and salt marshes. Vegetation 62(1):499–521. https://doi.org/10.1007/BF00044777
Ruocco M, Bertoni D, Sarti G, Ciccarelli D (2014) Mediterranean coastal dune systems: Which abiotic factors have the most influence on plant communities? Estuar Coast Shelf Sci 149:213–222. https://doi.org/10.1016/j.ecss.2014.08.019
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:217037. https://doi.org/10.1155/2012/217037
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A (2012) Fiji: an open-source platform for biological image analysis. Nat Methods 9:676–682. https://doi.org/10.1038/nmeth.2019
Singh RK, Kota S, Flowers TJ (2021) Salt tolerance in rice: seedling and reproductive stage QTL mapping come of age. Theor Appl Genet 134(11):3495–3533. https://doi.org/10.1007/s00122-021-03890-3
Sykes MT, Wilson JB (1989) The effect of salinity on the growth of some New Zealand sand dune species. Acta Bot Neerl 38(2):173–182
Sorce C, Bottega S, Spanò C (2019) Seasonal and microclimatic influences on the ecophysiology of Mediterranean coastal dune plants. Estuar Coast Shelf Sci 219:317–327. https://doi.org/10.1016/j.ecss.2019.02.031
Tang L, Zhou QS, Gao Y, Li P (2022) Biomass allocation in response to salinity and competition in native and invasive species. Ecosphere 13(1):e3900. https://doi.org/10.1002/ecs2.3900
Taulavuori E, Tahkokorpi M, Laine K, Taulavuori K (2010) Drought tolerance of juvenile and mature leaves of a deciduous dwarf shrub Vaccinium myrtillus in a boreal environment. Protoplasma 241(1):19–27. https://doi.org/10.1007/s00709-009-0096-x
van Puijenbroek ME, Teichmann C, Meijdam N, Oliveras I, Berendse F, Limpens J (2017) Does salt stress constrain spatial distribution of dune building grasses Ammophila arenaria and Elytrichia juncea on the beach? Ecol and Evol 7(18):7290–7303. https://doi.org/10.1002/ece3.3244
Wankhade SD, Cornejo MJ, Mateu-Andrés I, Sanz A (2013) Morpho-physiological variations in response to NaCl stress during vegetative and reproductive development of rice. Acta Physiol Plant 35(2):323–333. https://doi.org/10.1007/s11738-012-1075-y
Yuan F, Guo J, Shabala S, Wang B (2019) Reproductive physiology of halophytes: current standing. Front Plant Sci 9:1954. https://doi.org/10.3389/fpls.2018.01954
Acknowledgements
M.D. Cerrato was supported by a Beca de Formación de Profesorado Universitario (FPU) (Ministerio de Universidades, Gobierno de España). A. Sureda was granted by Instituto de Salud Carlos III through the Fondo de Investigación para la Salud (Project CIBEROBN CB12/03/30038).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. M.D. Cerrato was supported by a Beca de Formación de Profesorado Universitario (FPU) (Ministerio de Universidades, Gobierno de España). A. Sureda was granted by Instituto de Salud Carlos III through the Fondo de Investigación para la Salud (Project CIBEROBN CB12/03/30038).
Author information
Authors and Affiliations
Contributions
Marcello Cerrato, Jaume Flexas, Antoni Sureda, and Lorenzo Gil contributed to the study conception and design. The methodology was developed by Marcello Cerrato, Antoni Sureda, Jaume Flexas, and Lorenzo Gil. Material preparation, data collection, and analysis were performed by Marcello Cerrato, Arnau Ribas-Serra, Pere Miquel Mir-Rosselló, Cyril Douthe, Iván Cortés-Fernández, and Carles Cardona Ametller. The first draft of the manuscript was written by Marcello Cerrato, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Handling Editor: Wendy Stirk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cerrato, M.D., Ribas-Serra, A., Mir-Rosselló, P.M. et al. Response to Seawater Exposure During the Flowering Stage of Two Dune-Building Grasses: Calamagrostis arenaria (L.) Roth and Thinopyrum junceum (L.) Á.Löve. J Plant Growth Regul (2024). https://doi.org/10.1007/s00344-024-11274-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00344-024-11274-2