
Abstract
We summarized existing knowledge on Dreissena polymorpha (the zebra mussel) and D. r. bugensis (the quagga mussel), including data on their taxonomy, systematics, evolution, life cycle, reproduction, feeding, growth and longevity, population dynamics, interspecific competition, habitat requirements, and distribution within and among waterbodies. We analyzed the history of spread of both species and the major pathways and vectors of their spread in Europe and North America. Special consideration was given to their ecological and economic impacts and their natural enemies, like waterfowl, fishes, and parasites, as well as the prevention of their introduction, early detection, control, and containment. We also outlined the most salient ecosystem services provided by zebra and quagga mussels, including water purification, nutrient recycling, culling the effects of eutrophication, biomonitoring, and their role as a food resource for fish and waterfowl. Finally, we identified major knowledge gaps and key studies needed to better understand the biology, ecology, and impacts of these aggressive freshwater invaders. Our review indicates that much crucial information on the quagga mussel is still missing, including key life history parameters, like spawning cues, fecundity, and longevity, particularly for the profundal zone of deep lakes.
Similar content being viewed by others


Introduction
The dreissenid bivalves Dreissena polymorpha (Pallas), the zebra mussel, and D. rostriformis bugensis (Andrusov), the quagga mussel, belong to a family of sessile mussels originated in the marine environment, but later adapted to the brackish and fresh waters of the northern hemisphere (Starobogatov & Andreeva, 1994; Orlova, 2014). Their life history, which includes planktonic larvae and an attached adult stage, typical for marine mussels, represents a novel ecological type in freshwaters (Johnson & Carlton, 1996; Karatayev et al., 1997, 2007a).
D. polymorpha was found in 1769 in an oxbow lake in the lower Yaik River (later re-named as Ural River) by Peter Pallas (Pallas, 1771), a German zoologist and botanist who worked in Russia from 1767 to 1810. Pallas described D. polymorpha as Mytilus polymorphus because of its resemblance to the marine mussel Mytilus and reported that he found it only in the rivers Volga, Ural, and in the Caspian Sea (Pallas, 1773). For thousands of years, since the last glaciation, the geographic range of D. polymorpha was limited to the Ponto–Caspian basin (Zhadin, 1946; Mordukhai-Boltovskoi, 1960; Logvinenko & Starobogatov, 1968; Kinzelbach, 1992; Starobogatov & Andreeva, 1994; Son, 2007; bij de Vaate et al., 2014). However, after canals were built to connect shipping routes between the Black Sea and Baltic Sea basins, D. polymorpha quickly spread across eastern and western Europe (reviewed in Zhadin, 1946; Kerney & Morton, 1970; Kinzelbach, 1992; bij de Vaate et al., 2014; Karatayev et al., 2003, 2007b, 2008; Pollux et al., 2010). The unusual life cycle of D. polymorpha and their fouling of water supply systems have brought the attention of numerous scientists and engineers since the late 1700s (reviewed in Zhadin, 1946; Clarke, 1952), resulting in hundreds of publications in the nineteenth and twentieth centuries (Limanova, 1964, 1978; Schloesser et al., 1994). By the 1980s, when zebra mussels finally reached North America, over 2000 papers on their biology, ecology, spread, and control were published in Europe (reviewed in Karatayev et al., 2015a). The rapid spread of zebra mussels across North American freshwaters, with their significant ecological impacts and damages to industrial installations using raw water, triggered another strong spike in their study, both in the New and the Old Worlds (Nalepa & Schloesser, 1993, 2014; Starobogatov, 1994a; van der Velde et al., 2010a; Schloesser & Schmuckal, 2012; Karatayev et al., 2015a). The numbers of publications on various aspects of the biology, ecology, impact, and control of D. polymorpha are still growing and by the end of 2021 exceeded 77% of all papers published on dreissenids (Table 1).
Before the second half of the twentieth century, in contrast to the zebra mussel, relatively little was known about its congener the quagga mussel (Karatayev et al., 2015a). Dreissena r. bugensis was first recorded in the Bug Liman (a shallow productive estuary) and was described as D. rostriformis (Andrusov, 1890). Later, Andrusov (1897) redescribed the species as D. bugensis (for current taxonomy see Taxonomy, Systematics, and Evolution). The native range of D. r. bugensis was much smaller than that of D. polymorpha and included the Dnieper-Bug Liman, the Dnieper River delta, and the lower reaches of the South Bug and Ingulets rivers in Ukraine (Zhuravel, 1965, 1967; Mills et al., 1996; Orlova et al., 2005; Zhulidov et al., 2010; Karatayev et al., 2007b, 2011a; Orlova, 2014). Dreissena r. bugensis started spreading beyond its range only in the middle of the twentieth century (reviewed in Zhulidov et al., 2004, 2010; Karatayev et al., 2007b, 2011a, 2015a; van der Velde et al., 2010b). Its initial spread was slow, but it increased dramatically since the 1980s in both eastern and western Europe, as well as in North America (Zhulidov et al., 2004, 2010; Karatayev et al., 2007b, 2011a, 2015a; bij de Vaate et al., 2014; van der Velde et al., 2010b; Orlova, 2014; Benson, 2014), fostering intensive research in the Old and the New Worlds and resulting in a large number of publications in the last three decades (Table 1).
Both D. polymorpha and D. r. bugensis are among the most notorious freshwater invaders with tremendous ecological and economic impacts (Karatayev et al., 1997, 2002, 2007a, 2007b, 2015b; Beekey et al., 2004; Higgins & Vander Zanden, 2010; Kelley et al., 2010; Burlakova et al., 2012). Both species are considered ecosystem engineers that attach to the substrate with byssal threads and form three-dimensional reef-like structures altering the benthic environments colonized (reviewed in Karatayev et al., 2002, 2007a; Vanderploeg et al., 2002; Gutierrez et al., 2003; Zhu et al., 2006; Sousa et al., 2009) (Fig. 1).

Dreissenid aggregations on the bottom of Lake Erie, July 2019
Dreissenids also affect the planktonic community, trophic relationships, and nutrient cycling via their feeding and filtering activities (reviewed in Karatayev et al., 1997, 2002, 2007a, 2007b, 2015b; Beekey et al., 2004; Higgins & Vander Zanden, 2010; Kelley et al., 2010). Their economic impacts are estimated in hundreds of millions of dollars in damage and increased operating expenses in raw water-dependent infrastructures (O’Neill, 1996, 1997, 2008).
Although these two Dreissena species have similar life histories, they are not identical and have different histories of spread, different tolerances to environmental parameters, and different distributions within waterbodies, resulting in different ecosystem impacts (Nalepa et al., 2010; Karatayev et al., 2011a, 2015a, 2021a; Benson, 2014). The goal of this paper is to summarize current knowledge on various aspects of their biology, ecology, spread, and impacts, emphasizing similarities and differences between these species, as well as to identify the most significant areas where further research is necessary.
Research efforts: an overview of the literature
The section that follows is based on a search of the SCOPUS database performed in December 2021 using words “Dreissena” or “zebra mussel” or “quagga mussel” in the abstract, title, or keywords, yielding 2,853 publications (after eliminating duplicates and non-applicable titles). It should be stressed that the coverage of the earlier literature in SCOPUS is very sketchy; thus, most of the publications retrieved dated from 1980 to 2021 and only 19 from 1951 to1979 (compare with Table 1). All the publications were separated into eight major categories: (1) taxonomy, systematics, and evolution (phylogeny, phylogeography, genetic diversity); (2) general biology (including anatomy and morphology, physiology, life history traits, symbionts, parasites, and ecotoxicology); (3) ecology (ecological traits and habitat requirements); (4) dispersal (past and future dispersal, vectors and pathways); (5) impacts (studies that assess negative ecological and/or economic impacts on ecosystems or native communities/species, including competition, transfer of contaminants up the food chain, and others, see below); (6) ecosystem functions and services (including positive impacts on ecosystems/communities, such as water purification, reduction in phytoplankton, nutrient cycles (e.g., phosphorus reduction), food for other animals, positive effects on the native benthos consumed by fish and birds, macrophytes, etc., biomonitoring, bioremediation, various uses, mussel farming, etc.); (7) management (risk analysis, early detection, prevention, control, management, eradication, etc.); and (8) other studies that did not fit into the above categories. Publications were separated based on our interpretation of the results provided by authors. Documents often covered more than one subject but were tallied only once and assigned to the most relevant category. As ecosystem functions (i.e., “dynamic processes that determine the amount, forms, distribution, fluxes, import, and export of energy and various materials,” Strayer, 2012) are much more encompassing than ecosystem services (i.e., those that directly or indirectly contribute to human well-being), papers dealing with ecosystem functions resulting in predominantly negative ecological impact were included in the impact category, while publications dealing with predominantly positive ecological impacts were included in the ecosystem functions and services category.
The largest number of publications has been focused on ecosystem functions and services (34%) (Fig. 2). Within this group, 28% of publications examined the use of dreissenids as environmental bioindicators, 16% reported consumption of dreissenids by fishes, birds, and other animals, 14% reported reductions in phytoplankton, 9% described the positive effect of dreissenids on the benthos, and 5% and 7%, respectively, were on improvement in water clarity and their effects on nutrient cycles. The next most intensively studied subjects included general biology (19% of all publications), followed by ecology (17%), impact and management (9% each), dispersal (7%), and taxonomy, systematics, and evolution (2%, Fig. 2). The largest number of papers in the impact category were on the effects of dreissenids on unionids (32%), impacts on water utilities (10%), their negative effects on the deep-water amphipod Diporeia (11%), and fishes (8%), and on the transfer of contaminants (10%). The total share of all papers on toxicology (including studies across different subjects as the use of dreissenids as bioindicators, ecotoxicology, and accumulation and transfer of contaminants to fishes and birds) was 16%. Studies on natural enemies (including consumption by birds, fishes and other animals, and parasites) comprised 8% of all publications.

Thematic classification of publications on Dreissena spp. (according to SCOPUS, search performed in December 2021 using “Dreissena” or “zebra mussel” or “quagga mussel” in the abstract, title, or keywords). Documents often covered several subjects but were assigned to the most relevant category only
Taxonomy, systematics, and evolution
According to Turgeon et al. (1998), Dreissena polymorpha belongs to the phylum Mollusca; class Bivalvia Linnaeus, 1758; subclass Heterodonta Neumayr, 1884; order Veneroida H. Adams and A. Adams, 1856; superfamily Dreissenoidea Gray, 1840; family Dreissenidae Gray, 1840; and genus Dreissena Van Beneden, 1835. This taxonomy is currently accepted by the Integrated Taxonomic Information System (ITIS, www.itis.gov). Recently, however, the superfamily Dreissenoidea was transferred from the order Veneroida to the order Cardiida Ferussac, 1822 (Orlova, 2014; Araujo & de Jong, 2015).
Since the original discovery of D. polymorpha in 1769 (Pallas, 1771) the systematics and evolution of Dreissenidae have been revised multiple times with new genera and species being described, split, and lumped (reviewed in Andrusov, 1897; Mordukhai-Boltovskoi, 1960; Logvinenko & Starobogatov, 1968; Babak, 1983; Rosenberg & Ludyanskiy, 1994; Starobogatov, 1994b; van der Velde et al., 2010b; Orlova, 2014; Stepien et al., 2014). Originally, Pallas named the new species Mytilus polymorphus based on its resemblance to marine mussels Mytilus (family Mytilidae), but later Van Beneden (1835) discovered that Dreissenidae were different from Mytilidae in the anatomy and internal shell morphology and erected the genus Dreissena, renaming the species as Dreissena polymorpha. Gray (1840) placed this family in a separate superfamily—Dreissenacea. Gill (1871) excluded the Dreissenacea from the order Mytilidae and considered them closely related to the Corbiculacea and Veneracea (reviewed in van der Velde et al., 2010b). Subsequently, it was shown that Mytilidae and Dreissenidae are unrelated (Yonge & Campbell, 1968), and the similarities between them are the result of convergent evolution (reviewed in van der Velde et al., 2010b). While some authors suggested that the Dreissenoidea arose from corbiculoidean ancestors (Keen & Casey, 1969; Morton, 1970, 1993, 1996), others (McMahon, 1991; Starobogatov, 1994b) proposed that the three freshwater bivalve families, Dreissenidae, Sphaeriidae, and Corbiculidae, represent independent radiation processes, a notion which was later supported by molecular studies (Park & Ó Foighil, 2000).
According to Morton (1993), ancestral Dreissenidae originated in fresh and brackish waters of the Tethys Ocean during the Triassic period. Dreissenidae are the only extant representatives of the Superfamily Dreissenoidea that arose during the Eocene and flourished in the Late Miocene and Pleistocene (Andrusov, 1897; Keen & Casey, 1969; Morton, 1970, 1993, 1996; Babak, 1983; Nuttall, 1990; Starobogatov, 1994b; Geary et al., 2000; Bilandžija et al., 2013). The origin of the family was recently estimated at about 37.4 million years ago (MYA) (Bilandžija et al., 2013) and corresponds to the period of the formation of the Paratethys Sea and the occurrence of the first unequivocal dreissenid fossils (Benton, 1993; Bilandžija et al., 2013). Based on phylogenetic and molecular clock analyses using both nuclear and mitochondrial markers, Bilandžija et al. (2013) suggested that the split within Dreissena occurred 11.6 MYA, when the genus first appeared in the fossil record. Accordingly, the first split between D. rostriformis and the other three Dreissena spp. was estimated at 12.7 MYA and D. polymorpha branched off around 6.9 MYA. The timing of the first split is in agreement with the literature data (10 – 15 MYA; Stepien et al., 1999, 2001, 2003, but see Gelembiuk et al., 2006). These estimates of the appearance of D. polymorpha are more ancient than previously reported (Stepien et al., 2003), but Bilandžija et al. (2013) used newly developed methods that accounted for rate variations and assumed uncorrelated rates of evolution, thus allowing for more accurate estimates.
Although extant representatives of Dreissena are limited in number, their radiation about 10 – 11 MYA in giant brackish European lakes gave rise to over 200 species (albeit mostly subjective) and left massive fossil deposits (Kinzelbach, 1992; Starobogatov, 1994b). Over 130 species of Dreissenidae were described from the ancient Lake Pannon, located in central-eastern Europe 12 – 4 MYA, hosting a variety of habitats with different salinity levels, and numerous empty niches for colonization by survivors of the “Sarmatian Sea” that previously occupied the Pannonian basin (Geary et al., 2000). Most of these Dreissena species went extinct leaving large amounts of fossil deposits.
Only three genera of Dreissenidae have survived until the present day: Dreissena Van Beneden, 1835, Mytilopsis Conrad, 1857, and Congeria Partsch, 1835 (Nuttall, 1990; Rosenberg & Ludyanskiy, 1994; Bilandžija et al., 2013; Stepien et al., 2014). Two species of Dreissena became invasive both in Europe and in North America: D. polymorpha and D. rostriformis bugensis. Although several subspecies of D. polymorpha with the difference tolerance to salinity were reported from the Ponto–Caspian basin (see Habitat Requirements), only D. polymorpha polymorpha invaded Europe and North America.
While the validity of D. polymorpha was never questioned, the status of D. r. bugensis changed multiple times (Andrusov, 1890, 1897; Mordukhai-Boltovskoi, 1960; Starobogatov, 1994b; Rosenberg & Ludyanskiy, 1994; Stepien et al., 2014). Dreissena r. bugensis was first described as D. rostriformis by Andrusov (1890) and redescribed as a separate species by the same author a few years later as D. bugensis (Andrusov, 1897). Mordukhai-Boltovskoi (1960) classified it as a subspecies – D. r. bugensis, but Starobogatov (1994b) and Rosenberg & Ludyanskiy (1994) subsequently restored its species status. Finally, Stepien et al. (2014) suggested that D. r. bugensis is genetically very similar to four extant subspecies of D. rostriformis from the Caspian Sea, including D. r. grimmi, D. r. distincta, D. r. compressa, and D. r. pontocaspica. These four subspecies inhabit chiefly the middle and southern parts of the Caspian Sea at depths of 20 – 300 m, overlap in their distribution and shell morphology (Logvinenko & Starobogatov, 1968; Nuttall, 1990; Starobogatov, 1994b; Starobogatov & Andreeva, 1994), are genetically very similar, and thus were suggested to be a single taxonomic entity (Stepien et al., 2014). Stepien et al. (2014) also suggested that D. rostriformis from the Caspian Sea is genetically similar to D. r. bugensis and proposed to synonymize all subspecies of D. rostriformis.
Although genetically D. r. bugensis is only slightly different from D. rostriformis and thus likely does not qualify as a separate species, they occupy drastically different habitats. While all populations of D. rostriformis are brackish water organisms inhabiting the profundal areas of the Caspian Sea with salinities around 5 – 13‰, D. r. bugensis is typically freshwater and avoids salinities above 3 – 4‰ (Therriault et al., 2004, 2005; Orlova, 2014). Since D. rostriformis spawns at > 5 ‰, whereas D. r. bugensis does at < 3‰, they are unlikely to interbreed (Starobogatov, 1994b). Therriault et al. (2004, 2005) noted subtle but consistent genetic differences between D. r. bugensis from the Dnieper–Bug basin and D. rostriformis rostriformis (= distincta) from the Caspian Sea basin, suggesting that they represent two races: D. r. distincta – evolved in brackish water and D. r. bugensis – evolved in freshwater. It is likely that the divergence of D. r. bugensis from D. rostriformis occurred during the late Novoeuxinian period (late Pleistocene, ~ 0.1 – 0.01 MYA), as there are no fossil records of D. r. bugensis before this time (reviewed in Orlova, 2014), although according to Gelembiuk et al. (2006) the divergence of D. r. bugensis from D. rostriformis occurred about 0.5 MYA. The name D. r. bugensis is generally accepted by most scientists, and we suggest that this name should be used in future. Additional studies are needed, however, to better understand the differences between D. r. bugensis and D. rostriformis and their taxonomic status. Especially important are investigations of the salinity range of the populations of D. r. bugensis that recently colonized the Volga River, including the Volga Delta and freshened zones of the Caspian Sea (Orlova et al., 2004; Zhulidov et al., 2004, 2005; Son, 2007), and the Caspian Sea populations of D. rostriformis to confirm if the two subspecies are truly isolated by their salinity tolerance.
Biology and ecology
Life cycle
Both species of Dreissena have free swimming planktonic veliger larvae and an attached adult stage, a life history which is typical for marine mussels, but unique for freshwaters. The life history and reproductive characteristics of D. polymorpha have been studied much more extensively than those of D. r. bugensis. In recent decades, however, more data became available for the latter, especially due to comprehensive studies in the Laurentian Great Lakes (reviewed in Karatayev et al., 2014a; Karatayev & Burlakova, this issue). Although the life cycle of both dreissenids is similar, many important traits differ (Tables 2, 3).
In polymictic lakes, the gonads of D. polymorpha are fully developed by spring and spawning occurs in summer. Unspawned eggs may be resorbed in winter and then the cycle repeats during the next season (reviewed in Lvova & Makarova, 1994). The sizes of D. polymorpha eggs range around 40 – 100 µm, similar to egg sizes of D. r. bugensis (Table 2; Ackerman et al., 1994; Lvova & Makarova, 1994; Roe & MacIsaac, 1997). However, the sperm head of D. r. bugensis appears to be larger than that of D. polymorpha. According to Denson and Wang (1994), the sperm head of D. polymorpha is shorter, straight, and the acrosome is more bulbous and oval, while in the D. r. bugensis sperm cells are longer, curved, and the acrosome is wide at the point of attachment to the rest of the sperm head, but tapers more sharply toward the tip.
After the external fertilization of gametes released into the water column, a short-lived trochophore larva 57 – 121 µm in diameter develops (Ackerman et al., 1994). The trochophore transforms into a veliger stage that has a bilateral symmetric D-shape shell that feeds and moves with a ciliated velum (Ackerman et al., 1994; Lvova et al., 1994b; Verween et al., 2010). The size of D. polymorpha veliger ranges from 50 – 110 µm (Lvova et al., 1994b) to 70 – 160 µm (Ackerman et al., 1994). Subsequently, a veliconcha develops that has a more ornamented shell with a clam-like profile. This is the last free swimming stage of Dreissena larvae, whose average size is about 225 µm (range: 120 – 300 µm; Lvova et al., 1994b; Ackerman et al., 1994). The duration of the planktonic stage of D. polymorpha lasts 10 – 18 days (Hillbricht-Ilkowska & Stanczykowska, 1969; Lvova, 1980; Sprung, 1989; Borcherding & De Ruyter van Steveninck, 1992; Ackerman et al., 1994), but when water temperature decreases in autumn, the growth and development of larvae slow down, and the duration of the planktonic stage may last for over a month (Kirpichenko, 1964; Stanczykowska, 1977; Karatayev, 1981; Lvova et al., 1994b; Pollux et al., 2010).
After developing a foot and several other organs, including gills in the mantle cavity, the behavior of the larvae changes from free swimming to benthic and the shape of the shell becomes similar to that of adult individuals. These pediveliger larvae crawl on the bottom or swim near it. After selecting an appropriate surface, the pediveliger secretes byssal threads to attach to the substrate, undergoes metamorphosis, and becomes a postveliger (Kirpichenko, 1964; Ackerman et al., 1994; Verween et al., 2010).
Dreissena r. bugensis has a similar larval development, but the larvae are generally larger (Gerstenberger et al., 2011; Bowen et al., 2018) and, according to Martel et al. (2001), settle at significantly larger sizes than those of D. polymorpha (256 – 284 µm, vs. 236 – 249 µm, respectively). Further, the larvae of D. r. bugensis were reported to settle at a greater size in offshore hypolimnion habitats (314 µm), than in nearshore epilimnion ones (262 µm) (Martel et al., 2001). Laboratory trials also suggest that D. r. bugensis larvae may remain longer in the plankton than D. polymorpha (32 vs. 21 days) (Wright et al., 1996).
Gonad, oocyte development, and spawning are different for D. polymorpha from shallow, polymictic waters, as compared with deep, cold profundal waters (Walz, 1978a; Bacchetta et al., 2010). In warm polymictic habitats, the gonads of D. polymorpha are fully developed by spring and spawning occurs in late spring (May to June in the northern hemisphere). Temperature is the key factor that triggers spawning of zebra mussels in polymictic waters: D. polymorpha larvae first appear in the plankton when water temperatures reach 12 – 15 °C and continue to spawn until the end of summer (August or September) (Karatayev, 1981; Sprung, 1987; Borcherding, 1991; Lvova et al., 1994b; Burlakova, 1998; Bacchetta et al., 2010; Karatayev et al., 2010, 2014a; Pollux et al., 2010). According to Borcherding (1991), the increase in temperature above the 12 °C threshold is necessary for the simultaneous release of male and female gametes, and D. polymorpha cannot establish viable populations in waterbodies with maximum daily temperature below 12 °C throughout the year.
Veliger densities are characterized by 1 – 3 peaks in abundance (reviewed in Lvova et al., 1994b; Burlakova, 1998; Karatayev et al., 2010). Cold springs/early summers may greatly delay the reproduction of D. polymorpha, producing a single, short, and sharp peak in larval densities, while in warm years zebra mussel larvae may show several small peaks in abundance or one extended peak (Karatayev, 1981, 1983; Lvova et al., 1994b; Burlakova, 1998; Karatayev et al., 2010). Dreissena polymorpha larvae are typically present in the plankton from May through September/October at densities ranging from 1 to 500 ind./L, although they occasionally may reach local peaks of up to 9000 ind./L in areas with high adult densities (Hillbricht-Ilkowska & Stanczykowska, 1969; Stanczykowska, 1977; Karatayev, 1981, 1983; Borcherding & De Ruyter van Steveninck, 1992; Kern et al., 1994; Lvova et al., 1994b; Mackie & Schloesser, 1996; Burlakova, 1998; Karatayev et al., 2010; Pollux et al., 2010).
In contrast to warm polymictic areas, very few studies were conducted on D. polymorpha in cold, profundal regions, and their results are contradictory (Walz, 1978a; Bacchetta et al., 2010). According to Bacchetta et al. (2010), in cold deep (25 m) waters of Lake Iseo (Italy) where water temperature never reaches 12 °C, mussels have ripe gonads for extended periods of time and may spawn smaller numbers of eggs over many months. These results contradict with the 12 °C threshold for successful reproduction (Borcherding, 1991), as well as with earlier assessments by Walz (1978a) that “it is certain that these mussels (found at 50 m depth in Lake Constance) could not have completed their early developmental stages at this depth but must have landed there accidently from their original habitat after the gametes have been formed.” Thus, the reproduction dynamics of D. polymorpha in cold, deep waters needs further research.
In polymictic waterbodies, quagga mussels likely spawn at the same time as D. polymorpha and may have similar spawning cues, in particular increases in temperature. In shallow areas of Lake Erie, both dreissenid species were found to initiate spawning when the water reached 18 – 20 °C (Claxton & Mackie, 1998). In contrast, in the profundal zones of deep lakes, quagga mussels have been found to spawn at colder water temperatures (Roe & MacIsaac, 1997; Claxton & Mackie, 1998; Nalepa et al., 2010; Glyshaw et al., 2015; Pothoven & Elgin, 2019). For the profundal zone of the eastern basin of Lake Erie at 55 m depth, Roe and MacIsaac (1997) reported evidence of gonad development and spawning at a temperature of 4.8 °C, in July 1996. In Lake Michigan, mature mussels were found in April at 45 m depth at temperatures of 2.4–5.9 °C and even at 93 m depth at 2.5 to 4.5 °C (Glyshaw et al., 2015). More work is needed to understand the conditions that determine spawning in quagga mussels, especially as related to water depth.
The dynamics of quagga mussel larvae in the plankton differs dramatically from that of zebra mussels. For Lake Michigan, Glyshaw et al. (2015) reported that the density of quagga mussel veligers peaked at different times of the year depending on water depth: at 15 m in mid-summer and at 45 and 110 m in December. According to Pothoven and Elgin (2019), D. r. bugensis veligers were present at all sites monitored in Lake Michigan throughout the sampling season (March–December), with peaks of density in summer and fall. In several inland lakes in California colonized by quagga mussels only, with water temperature always > 12 °C, veligers were present year-round, but their densities were lowest during winter and highest when epilimnetic temperature exceeded 22 °C (Reid et al., 2010; Gerstenberger et al., 2011). Likewise, quagga mussel veligers in the Great Lakes were reported to be present in the plankton year-round (James Watkins, personal communication). There also are indications that quagga mussel larvae are present year-round in the plankton of deep European lakes (Piet Spaak, personal communication).
Only two studies investigated the fecundity of zebra mussels. Lvova (1977, 1980) reported 275,000 –300,000 eggs per female per season, whereas Sprung (1991) estimated that females may produce up to 1,000,000 eggs and males up to nearly 10,000,000,000 sperm per season, comprising more than 30% of their body weight prior to spawning. No data for the fecundity of quagga mussels are available, and although their fecundity has been assumed to be the same as that of zebra mussels (Keller et al., 2007), given the many differences in the biology and ecology of these species this assumption is speculative and, probably, unwarranted. Because the spread of quagga mussels is presently ongoing in both Europe and North America, and in deep stratified lakes they build up huge populations below the thermocline, it is critically important to study their fecundity, as well as the impact of environmental variables like temperature, depth, and food on their reproductive potential.
Growth
The growth and longevity of D. polymorpha were studied for over 70 years in Europe and later in North America using various methods, including counting annual growth rings, analysis of size-frequency distributions, assessing their growth under experimental conditions, and monitoring marked mussels in the wild, without removing them from the substrate (reviewed in Lvova et al., 1994a; Karatayev et al., 2006; Pollux et al., 2010). All methods have their pros and cons. Counting the annual rings that are formed on the mussels’ shells during the slow-growth winter periods was the most commonly used method in the past and probably the easiest (reviewed in Karatayev et al., 2006). However, this method was largely abandoned in the 1990s as it became clear that every year multiple “disturbance” rings may form on the shells and it is difficult to distinguish the annual rings from those formed due to other factors, resulting in a substantial overestimation of the mussels’ longevity (Morton, l969a; Lvova, 1980; Lvova et al., 1994a; Jantz, 1996).
Size-frequency distributions have been used in multiple studies of zebra mussels’ growth (Morton, 1969a; Jantz & Neumann, 1992; Martel, 1993, 1995; Smit et al., 1993; Dall & Hamburger, 1996; Chase & Bailey, 1999; Orlova & Panov, 2004), but this method requires that spawning and settlement are synchronized, and each generation forms distinct size (age) classes or cohorts (reviewed in Karatayev et al., 2006). However, yearly cohorts are often difficult to distinguish, as in many waterbodies zebra mussels spawn throughout the entire summer and settled young-of-the-year mussels do not form distinct size (age) groups (bij de Vaate, 1991; Jantz & Neumann, 1992; Lvova et al., 1994a). Nevertheless, when the population encompasses few and clearly distinguishable age classes, this method can be effective in time series analyses of population size distribution curves (Burlakova, 1998; Allen et al., 1999).
After the method of counting annual rings was largely discontinued, the most common approach to study the growth of D. polymorpha has been the monitoring of individual, often marked, specimens in cages deployed in the field (Lvova, 1980; Smit et al., 1992, 1993; Sprung, 1992, 1995a; Dorgelo, 1993; Dall & Hamburger, 1996; Bitterman et al., 1994; MacIsaac, 1994; Allen et al., 1999; Horvath & Lamberti, 1999; Yu & Culver, 1999; Garton & Johnson, 2000; Karatayev et al., 2006, 2018b; Beisel et al., 2010) or in the laboratory (Walz, 1978a, 1978b; Jantz & Neumann, 1992, 1998; Dorgelo, 1993; Neumann et al., 1993; Jantz, 1996; Stoeckmann & Garton, 1997; Baldwin et al., 2002; Karatayev et al., 2011b). However, the use of caged mussels may underestimate growth rates as the mesh on the cages may reduce water exchange, especially if cages become overgrown by periphyton (Kachanova, 1963; Karatayev, 1983; Stanczykowska & Lewandowski, 1995; Burlakova, 1998). For example, in 3–4 year experiments in the Uchinskoe Reservoir (Russia), mussels in cages not overgrown with periphyton exhibited high growth rates and reached high sizes (up to 36 mm, which coincides with the maximum size of mussels commonly found in the reservoir) (Lvova, 1980). In contrast, in Lake Lukomskoe (Belarus), cages made of the same material became covered with periphyton, resulting in much slower growth rates and noticeably smaller maximum sizes even after 10 years of experimentation, suggesting that impaired water exchange decreased food availability for the animals (Lvova et al., 1994a). Further, suspension of cages in the water column above the benthic layer has been suggested to yield unrealistically high growth rates compared to animals on the bottom, where food resources are often depleted due to their filtration by dense mussel beds (Yu & Culver, 1999; Karatayev et al., 2006, 2018b; Malkin et al., 2012).
Monitoring the growth of tagged mussels under natural conditions (i.e., on the bottom, attached to rocks) could conceivably provide the most realistic estimates of their growth, but this method has been used in very few studies (e.g., Stoeckman & Garton, 1997; Burlakova, 1998). Burlakova (1998) found that the growth rate of mussels on stones in the Svisloch River (Belarus) was up to 400% greater than that of caged mussels at the same sites due to clogging of the cages with macrophytes and periphyton. The disadvantage of this method is that mussels can displace, form druses, or be consumed by predators, thus requiring very large numbers of initially marked individuals in order to retrieve reasonable high numbers in subsequent observations.
Much fewer studies were conducted on the growth of D. r. bugensis. Stoeckmann & Garton (1997) used marked mussels attached to rocks in the western basin of Lake Erie. Caged mussels were used in the western (MacIsaac, 1994; Stoeckmann, 2003) and the eastern basins of Lake Erie (Karatayev et al., 2018b), in Lake Ontario (Malkin et al., 2012; Elgin et al., 2022), in the St. Lawrence River (Casper et al., 2014), and in the Meuse River (Marescaux et al., 2015). Growth in laboratory conditions was also assessed in a few studies (Baldwin et al., 2002; Karatayev et al., 2011b; Metz et al., 2018). The growth of quagga mussels was also investigated in Lake Mead, Arizona, which is much warmer (winter temperatures always > 12 °C) and was colonized by D. r. bugensis only (Wong et al., 2012).
Dreissena spp. growth rates depend on a wide array of environmental variables, including temperature, food, season of the year, depth, water velocity, and other factors (Table 3). Many of these variables covary (e.g., temperature declines in the fall and so does food availability), making it very difficult to tease apart their relative importance. All waterbodies where observations on the growth of D. r. bugensis were conducted (except for the western USA) were initially colonized by D. polymorpha and only later by D. r. bugensis, and all reported higher rates for D. r. bugensis than for D. polymorpha (Baldwin et al., 2002; Stoeckmann, 2003; Karatayev et al., 2011b; Casper et al., 2014; Marescaux et al., 2015; Metz et al., 2018). The only exception is the work of MacIsaac (1994), who concluded, after having used cages deployed in Lake Erie for 56 days that the growth and survival rates of the two species are very similar.
One of the most controversial environmental drivers of the growth of D. polymorpha is water temperature, especially its lowest threshold. Many studies based on seasonal observations in temperate regions showed that mussel growth stops in the winter and resumes in the spring after water temperatures reach > 10 °C, suggesting that 10 °C is the lower temperature threshold for growth (Morton, 1969a, 1969b; Karatayev, 1983; Mackie, 1991; Jantz & Neumann, 1992). However, other studies suggested that mussels still grow at 6 °C (bij de Vaate, 1991), and even at 3 °C (Smit et al., 1992). These lower thresholds were supported by occasional reports of D. polymorpha from depths far below the thermocline: 50 m in Garda Lake (Franchini, 1978), 55 m in Bodensee Lake (Walz, 1973), 80 m in Lake Huron (Nalepa et al., 2007), 109 m in Lake Ontario (Watkins et al., 2007), and 128 m in Lake Michigan (Nalepa et al., 2014). The near-bottom temperature at these depths is always below 10 °C, but D. polymorpha attained the adult stage there, even though the growth rates were likely very slow.
In the Great Lakes, observations of D. polymorpha at such depths are very unusual and could potentially be a result of misidentifications with D. r. bugensis (Karatayev & Burlakova, this issue). In an experiment conducted in Lake Constance, D. polymorpha were kept in two treatments with water pumped constantly from 1 to 60 m for two years (Walz, 1978b). In one treatment mussels were exposed to the natural seston concentrations and temperature regimes (5–20 °C) of surface water, while in the other mussels were kept at the conditions of profundal waters (4.5–5.5 °C and low food concentrations). Over the two years of the experiment, the length of D. polymorpha remained constant in the deep-water treatment, while the dry weight, protein, and carbon content of the soft body decreased exponentially (Walz, 1978b). This experiment suggests that the low food and temperature conditions of the profundal waters are unsuitable for D. polymorpha. Similar studies were conducted with D. polymorpha and D. r. bugensis kept in flow-through tanks for over 9 months using Lake Erie surface water at two temperatures: ambient temperature (that mimicked the littoral environment, range 4–25 °C) and chilled water (range 5–8 °C) to mimic the hypolimnion conditions (Karatayev et al., 2011b). While neither species grew during the winter at either treatment when cold water coincided with low food supply, during the growing season both species showed significant growth rates in both treatments, suggesting that the lack of food, rather than low temperatures, curtail the growth of both dreissenid species during the winter (Karatayev et al., 2011b). In these experiments, the growth rate of D. r. bugensis was significantly higher than that of D. polymorpha, which agrees with the fact that D. r. bugensis grows well in the cold profundal zones of the Great Lakes, where near-bottom temperatures are always below 6 °C. It is likely that D. r. bugensis has a lower temperature threshold than D. polymorpha, but the question whether temperature, food, or a combination of both usually (albeit with exceptions) limit the range of D. polymorpha to the well-mixed areas and prevent the establishment of sizable populations in the cold profundal zone, is still open.
Increased water temperature enhances the growth rate of D. polymorpha and D. r. bugensis in a wide range of temperatures (Table 3). In this respect, data from cooling water reservoirs for thermal and nuclear power plants are particularly interesting. The growth rates of D. polymorpha exposed to different temperature regimes, but with otherwise very similar environmental conditions allowed for direct estimates of the effects of temperature on the mussels (Yaroshenko & Naberezhnyi, 1971; Skalskaya, 1976a, 1976b; Elagina et al., 1978; Karatayev & Tishchikov, 1979; Karatayev, 1983; Kornobis, 1977; Karatayev et al., 2006). In all cases D. polymorpha grew faster in the areas heated by the power plant outflow, than in those unaffected by it. However, when temperatures exceeded 30 °C, the growth rate of D. polymorpha declined and at temperatures > 32 °C, most mussels died (Karatayev, 1983; Protasov et al., 1983; Vladimirov, 1983; Sinitsina & Protasov, 1993; Lyakhnovich et al., 1994). Because the upper temperature survival limit of D. r. bugensis is lower than that of D. polymorpha, it is likely that the upper temperature threshold for D. r. bugensis growth is also lower.
As reviewed above, food availability is another important factor that governs the growth of Dreissena spp. (Table 3). In eutrophic lakes, the growth rates of D. polymorpha are higher than in meso-oligotrophic lakes, most probably due to the higher chlorophyll content and better food conditions (Dorgelo, 1993; Burlakova, 1998; Jants & Neumann, 1998). According to Sprung (1995a), a positive correlation between D. polymorpha shell growth and seston concentration (i.e., availability of particulate organic matter) occurs when seston concentrations stay below those at which the intestine is filled to capacity when the animal filters at a maximum rate (Sprung, 1995b).
There are strong seasonal variations in the growth of Dreissena spp. (Table 3). In temperate regions, the maximum growth rates of both D. polymorpha and D. r. bugensis usually occur early in the growing season and coincide with peaks in phytoplankton abundance (Walz, 1978a; Smit et al., 1992; Lvova et al., 1994a; Burlakova, 1998; Garton & Johnson, 2000; Karatayev et al., 2006; Malkin et al., 2012). In mid-summer growth rates often decline, most likely due to spawning (Lvova, 1977; 1980; Allen et al., 1999), lower food concentrations (Walz, 1978a; Malkin et al., 2012), or blooms of unpalatable or toxic algae that hamper filter feeding (Stanczykowska & Lewandowski, 1995; Boegehold et al., 2019). Strong summer food depletion in the profundal zone of the Great Lakes slows down the growth of D. r. bugensis (Karatayev et al., 2018b). In these areas, mussels grow faster during periods of active vertical water mixing, when surface waters with the high phytoplankton concentrations reach the bottom. In contrast, in the southern edge of dreissenid distribution, mid-summer growth is inhibited by high water temperature (Mihuc et al., 1999; Allen et al., 1999).
The growth of Dreissena spp. may also depend on the time since mussel invasion. Recent studies conducted in Lake Erie suggest that early in the invasion, when food resources in the profundal zone were abundant, D. r. bugensis grew fast (Karatayev et al., 2018b; Karatayev & Burlakova, this issue). However, as mussel populations increased, they reduce the amount of food (particulate organic matter, including chlorophyll and plankton in general) available lake wide, resulting in much slower growth rates later in the invasion (reviewed in Karatayev et al., 1997; Higgins & Vander Zanden, 2010; Pothoven & Fahnenstiel, 2014; Rowe et al., 2015).
Longevity
Reported longevity of D. polymorpha ranges from 3 to 19 years, but maximum sizes are roughly similar (30–36 mm). This wide range of reported longevity may be due to natural variation but may also reflect biased assumptions in the method used (reviewed in Karatayev et al., 2006; Pollux et al., 2010). The two earliest life span estimates, reported in 1952, had a ninefold difference: Karpevich (1952) estimated the longevity at 18 years by counting the annual growth rings on the shells, while Clarke (1952), using size-frequency distributions, concluded that the animals typically live about three years. Later, Lvova (1980), based on observations of caged D. polymorpha, suggested that mussels may reach their maximum size in 4 years. These data are in line with several other reports from warm polymictic environments (Jantz & Neumann, 1992; Burlakova, 1998; Garton & Johnson, 2000), suggesting that the typical longevity of D. polymorpha is around 3 – 5 years (Table 3; reviewed in Lvova et al., 1994a; Karatayev et al., 2006, 2014a) depending on the water temperature, with shorter lifespan in warmer lakes (reviewed in Stanczykowska, 1977; Pollux et al., 2010). However, it is also known that if food is limited, mussels may live longer. The estimated longevity of D. polymorpha kept in cages in Lake Lukomskoe (Belarus) heavily overgrown with periphyton was about 10 years (Lvova et al., 1994a). Finally, we lack data on D. polymorpha longevity in the profundal zone of stratified lakes where their growth is limited by both low temperatures and low food concentrations.
In warm polymictic waterbodies, the longevity of D. r. bugensis is similar to that of D. polymorpha (Table 3). However, as in D. polymorpha, longevity may increase substantially in areas with low temperature and limited food resources. In addition, in the profundal zone of deep stratified lakes, the longevity of D. r. bugensis may depend on the time since initial colonization and may increase substantially later in the invasion due to the depletion of food resources (see above). Based on growth increments of caged mussels and size-frequency distributions, it was estimated that 30-mm-long individuals of D. r. bugensis from the profundal zone of eastern Lake Erie could be 15 or even 30 years old (Karatayev et al., 2018b; Karatayev & Burlakova, this issue). Similar results were also reported by Elgin et al. (2022) for the profundal zone of Lake Ontario.
Population dynamics
The fact that in many invasive species there often is a lag time between the first introduction and the development of a large population has been observed in many investigations. This lag is variable, ranging from less than a year to decades and probably depends on many intrinsic, species specific, and environmental variables (Kiritani & Yamamura, 2003; Simberloff & Gibbons, 2004; Daehler, 2009). In order to estimate this lag time, the date of the first introduction needs to be known. However, for many aquatic invaders that were introduced unintentionally, including Dreissena spp., these data are scare and therefore most assessments often rely on the date of the first record (which may or may not coincide with the actual introduction time; reviewed in Burlakova et al., 2006; Karatayev et al., 2011a).
Population explosions early in the invasion were documented frequently for both Dreissena species (Sebestyen, 1937; Lyakhov & Mikheev, 1964; Karatayev, 1983; Hebert et al., 1991; Dermott & Munawar, 1993; Marsden et al., 1993; Nalepa et al., 1995, 1996, 2003; Griffiths et al., 1991; Burlakova et al., 2006; Karatayev et al., 2011a, 2015a, 2021a; Hetherington et al., 2019; Strayer et al., 1996, 2019). The lag time between first detection and peak population size are much shorter for zebra (2.5 ± 0.2 SE years), than for quagga mussels (12.2 ± 1.5 SE years) (Karatayev et al., 2011a, 2015a; Ginn et al., 2018). Lake morphometry has been shown to largely govern the population dynamics of quagga mussels (Karatayev et al., 2021a, 2021b, 2022). In shallow polymictic lakes where quagga mussels grow fast and are short lived, the average time required to reach the population maximum is 6 – 19 years, whereas in the deep areas of stratified lakes (e.g., the Laurentian Great Lakes) where quagga mussels grow slowly and live longer (Elgin et al., 2022), their population densities are still increasing even 30 years after initial invasion (Nalepa et al., 2010, 2020; Karatayev et al., 2021a, 2022; Karatayev & Burlakova, this issue).
Later in the invasion the population size of dreissenids can fluctuate widely, from maintaining relatively stable levels for substantial periods of time, to declining strongly or even disappearing altogether if the environment becomes unsuitable (Kinzelbach, 1992; Ramcharan et al., 1992; Stanczykowska & Lewandowski, 1993; Burlakova et al., 2006; Strayer & Malcom, 2006; Noordhuis et al., 2016; Strayer et al., 2019). The environmental factors that control zebra mussel populations cover a wide array of conditions, including substrate and food limitation, eutrophication, pollution, oxygen depletion, competition, and predation (Stanczykowska & Lewandowski, 1993; Kinzelbach, 1992; Karatayev et al., 1997, 2002, 2003, 2011a; Molloy et al., 1997; Noordhuis et al., 2016; Wawrzyniak-Wydrowska et al., 2019; Rudstam & Gandino, 2020). It is likely that the environmental variables that govern the population dynamics of D. r. bugensis are similar to those listed above. Long-term systematic surveys of zebra and especially quagga mussel population dynamics, however, are rare, and findings are often inconclusive due to heterogeneous methodologies and/or system-specific environmental conditions (Strayer et al., 2019, but also see Hetherington et al., 2019). More lake-wide long-term studies are needed to understand the multiannual dynamics of these mussels under different environmental conditions.
Interspecific competition
Multiple studies conducted in inland waterbodies in Europe and in North America have shown that the introduction of quagga mussels into lakes and reservoirs previously colonized by zebra mussels often causes dramatic declines of zebra mussel populations (Orlova et al., 2004, 2005; Noordhuis et al., 2016; Balogh et al., 2018; Karatayev et al., 2011a, 2015a; Heiler et al., 2013; bij de Vaate et al., 2014; Hetherington et al., 2019; Strayer et al., 2019). The outcome of this competition, however, varies from almost complete extirpation of zebra mussels to the co-existence of both species and sometimes even reversals to a predominance of D. polymorpha (Strayer & Malcom, 2006; Zhulidov et al., 2010; Karatayev et al., 2011a, 2021c; Strayer et al., 2019; Rudstam & Gandino, 2020). The results largely depend on lake morphometry, prevalent substrate types, food availability, predation, etc. (Karatayev et al., 1997, 2002, 2011a, 2021a; Hunter & Simons, 2004; Hecky et al., 2004; Patterson et al., 2005; Burlakova et al., 2006; bij de Vaate et al., 2014; Rudstam & Gandino, 2020; Jackson et al., 2020).
In shallow polymictic lakes, quagga mussels become dominant 4 – 12 years after coexisting with previously established zebra mussels, but despite their dominance, even after 30 years they do not appear to fully replace zebra mussels (Orlova et al., 2004; Noordhuis et al., 2016; Balogh et al., 2018; Hetherington et al., 2019; Karatayev et al., 2014b, 2021a, 2021c).
Even in waterbodies initially dominated by quagga mussels, species-selective predation may favor zebra mussels, resulting in their population increase and occasional dominance (Zhulidov et al., 2006, 2010; Rudstam & Gandino, 2020). Quagga mussels are more vulnerable to predation because their shells are thinner, their attachment strength is lower, their aggregation behavior is weaker, and they are less prone to seek refuge (Mackie, 1991; Dermott & Munawar, 1993; Claxton & Mackie, 1998; Kobak & Kakareko, 2009; Peyer et al., 2009, 2010; Naddafi & Rudstam, 2013; Balogh et al., 2019; Czarnoleski & Müller, 2014). After large hypoxia-induced (or other factors) mortality events of mixed populations, zebra mussels can recolonize the site much faster due to their larger presence in inflowing rivers (Karatayev et al., 2021c).
All these factors affect the population dynamics of both species, and their combined densities in polymictic lakes may increase (Noordhuis et al., 2016; Hetherington et al., 2019; Wegner et al., 2019; Karatayev et al., 2021a), remain stable (Karatayev et al., 2021a), or decline (Rudstam & Gandino, 2020; Karatayev et al., 2021a) compared to when the waterbody was colonized by zebra mussels alone.
In contrast, in large deep stratified lakes, quagga mussels become dominant faster, attain much higher densities, and drive zebra mussels to virtual extirpation (Nalepa et al., 2009a, 2010; Madenjian et al., 2015; Karatayev et al., 2021a; Karatayev & Burlakova, this issue).
Habitat requirements
Salinity
While several subspecies of D. polymorpha exist (or existed) in the Ponto–Caspian basin, only D. polymorpha polymorpha invaded waterbodies in Europe and North America. In their native range, the northern part of the Caspian Sea (Karpevich, 1964; Shkorbatov et al., 1994) and the Taganrog Gulf of the Azov Sea (Karpevich, 1955; Kruglova, 1957; Shkorbatov et al., 1994), D. p. polymorpha thrives in the least saline areas, from freshwater and to up to 5‰. In the Hudson River, D. p. polymorpha successfully settles and survives at salinities between 5‰ and 6‰ but is absent from areas where salinity ranges from 6 to 9‰ (Walton, 1996). It is likely that different populations of D. p. polymorpha may have different upper salinity limits (reviewed in Shkorbatov et al., 1994; Karatayev et al., 1998). The tolerance to salinity of D. p. polymorpha from various regions of the Volga River decreases with increasing distance from the Caspian Sea, likely due to genetic differences (Antonov & Shkorbatov, 1983; Shkorbatov, 1986; Shkorbatov & Antonov, 1986; Shkorbatov et al., 1994). However, conclusive long-term experiments to determine if these local differences are due to acclimation or genetic adaptations are yet to be conducted (Karatayev et al., 1998). Overall, field observations in Europe and North America suggest that D. p. polymorpha can establish stable populations at salinities below 6‰ (Table 2). Higher salinity tolerance limits (up to 12 ‰) were suggested for D. p. polymorpha (Strayer & Smith, 1993; Pollux et al., 2010), but these values are likely based on the data from the Aral and Caspian seas subspecies of Dreissena with the higher than D. p. polymorpha salinity tolerances.
Three subspecies of D. polymorpha (D. p. andrusovi, D. p. aralensis, and D. p. obtusicarinata) are native to the Ponto–Caspian basin and have a higher salinity tolerance, none of them have ever spread outside of their native range (reviewed in Starobogatov & Andreeva, 1994; Karatayev et al., 1998). D. p. andrusovi, commonly found in the Northern Caspian Sea (Malinovskaja et al., 1998), is a brackish water mussel whose optimal salinity is 3–10‰, but it can survive between 2 and 14‰ and even tolerate up to 20‰ for short periods (Lyakhnovich et al., 1994; Shkorbatov et al., 1994; Karatayev et al., 1998). The two other subspecies, D. p. aralensis and D. p. obtusicarinata, that dominated the benthic communities of the Aral Sea in the 1950s (Zenkevich, 1963; Mordukhai-Boltovskoi, 1972; Yablonskaya et al., 1973; Starobogatov & Andreeva, 1994), tolerated salinities up to 17.6 – 18.4‰ (Khusainova, 1958; Lyakhnovich et al., 1994). The intensive irrigation-related diversion of water from the Amu Darya and Syr Darya rivers, which feed the Aral Sea, caused a dramatic decrease in the volume and increase in the salinity of the sea, likely triggering the extirpation of both subspecies from the sea in 1970 (reviewed in Karatayev et al., 1998; Micklin et al., 2020). While D. p. obtusecarinata likely became extinct, D. p. aralensis remained in the Syr Darya River and the lakes connected with it and recently has been successfully reintroduced in the freshened zones of the Aral Sea (reviewed in Aladin et al., 2019).
Data on the upper salinity tolerance of D. r. bugensis are scarce. In the Dnieper–Bug Liman, D. r. bugensis was recorded in areas with salinities up to 3.5‰ (Grigoryev, 1965). Similar limits were reported by Orlova (2014) and Mills et al. (1996) (Table 2). Thus, D. polymorpha is likely more salinity tolerant than D. r. bugensis, allowing it to invade more estuarine-type habitats.
We should mention that the above-reported salinity limits for both D. polymorpha and D. r. bugensis are for the chronic exposure. Both species can tolerate higher salinity for a short period (Shkorbatov et al. 1994; Ellis & MacIsaac, 2009). Thus, in experiments with D. polymorpha from the Volga River exposed to the gradual increase in salinity, first mussels start dying at 7–10‰, but some mussels’ survived exposure to 15‰ for up to 20 days. In contrast, all mussels exposed to abrupt salinity increase to 9‰ died within 12 days (Shkorbatov et al., 1994). According to Ellis & MacIsaac (2009), both dreissenid species exhibit little mortality after the exposure for 5 h to 30‰ and few individuals survived for 48 h.
Temperature
The lower temperature limit for D. polymorpha is > 0 °C as zebra mussels cannot survive freezing (Luferov, 1965). Dreissena r. bugensis probably has a similar lower temperature threshold (reviewed in Karatayev et al., 1998, 2007b, 2014a), although we are not aware of studies addressing this question (Garton et al., 2014). The exposure of D. polymorpha and D. r. bugensis to average daily temperatures from − 3.6 to − 7.2 °C for five days during a low-water event in the River Nederrijn, the Netherlands, resulted in the mortality of all mussels of both species (Leuven et al., 2014), suggesting that neither of these dreissenids can tolerate freezing. In laboratory experiments, however, two out of 16 zebra mussels survived temperatures ~ -3 °C for about 3 h (Paukstis et al., 1996), suggesting that at least a small percentage of the mussels can survive short-term freezing.
Laboratory studies suggest that the upper temperature limit for D. polymorpha ranges between 28 and 32 °C and depends on the climatic zone from which mussels were collected, acclimation time, and experimental design (reviewed in Shkorbatov et al., 1994; Aldridge et al., 1995; McMahon, 1996; Karatayev et al., 1998; Garton et al., 2014). The upper temperature limit observed in the field (rather than in laboratory conditions) is probably a better predictor of the geographic limits for the spread of D. polymorpha (reviewed in Karatayev et al., 1998). Multiple studies conducted in Europe in cooling reservoirs for thermal and nuclear power stations indicate that adult D. polymorpha can survive at temperatures up to 32–33 °C, depending on the exposure time (Table 4; Karatayev, 1983; Vladimirov, 1983; Lyakhnovich et al., 1994; Karatayev et al., 1998, 2007b). Similarly, in North America, zebra mussels have been shown to have upper thermal limits of around 31–32.5 °C (Aldridge et al., 1995; Allen et al., 1999; Garton et al., 2014). On the other hand, some studies suggest that D. polymorpha larvae appear to be less tolerant of high temperature than the adults. The upper temperature limit for larvae from the Dnieper–Krivoi–Rog Canal (Ukraine) is 30 °C (Shevtsova, 1968a) and 29 °C in the Konin Lakes (Poland) (Lewandowski & Ejsmont-Karabin, 1983). In general terms, D. r. bugensis appears to be less tolerant of high temperatures than D. polymorpha (Table 4).
Dissolved oxygen
While both dreissenid species are sensitive to the lack of oxygen, the zebra mussel is less tolerant to low oxygen than the quagga mussel (Table 4; reviewed in Shkorbatov et al., 1994; Karatayev et al., 1998, 2007b, 2015a; McMahon & Bogan, 2001). Shkorbatov et al. (1994) found that after 4 days of exposure to anoxic conditions at 20 °C, all zebra mussels died, while all quagga mussels survived. Birger et al. (1975) also showed that zebra mussels require higher oxygen concentrations than quagga mussels for survival. The higher tolerance of quagga than zebra mussels to lower oxygen conditions is perhaps one reason why quagga mussels can inhabit finer sediments and the profundal regions of lakes where zebra mussels are rarely found. Low oxygen, however, often prevents both dreissenid species from establishing stable populations in stratified lakes that experience periodic bottom hypoxia. For example, in the central basin of Lake Erie at depths > 20 m where bottom hypoxia routinely develops by the end of the growing season, only young-of-the-year mussels are found in small numbers, indicating that hypoxia eliminated the adult mussels settled during the previous season and restricted recruitment and survival of young-of-the-year Dreissena (Karatayev et al., 2018a, 2021d). Therefore, monitoring the distribution of Dreissena can be an effective tool for mapping the extent and frequency of hypoxia in freshwaters.
pH and calcium concentrations
Numerous studies established that pH and calcium are the most important chemical parameters that limit the establishment of sustainable D. polymorpha populations (Sprung, 1987; Ramcharan et al., 1992; Burlakova, 1998; Kraft et al., 2002; Drake & Bossenbroek, 2009; Hallstan et al., 2010; Karatayev et al., 2015c). Using data from 553 of the 1072 glacial lakes in the Republic of Belarus examined for the presence of D. polymorpha between 1971 and 1996, Burlakova (1998) found that of the 30 limnological parameters tested, the best predictors were pH (> 7.47) and calcium (> 24.7 mg/L, Burlakova, 1998; Karatayev et al., 2003, 2015c). Of the 80 Belarusian lakes free of D. polymorpha in 1996 and re-examined in 1997 – 2008, D. polymorpha had successfully invaded an additional 31 lakes previously classified as suitable, and none of the lakes previously classified as unsuitable were invaded (Karatayev et al., 2015c). The 31 lakes invaded in 1997–2008 confirmed the threshold successful invasion pH (7.47), which is similar to other estimates reported elsewhere (Table 4). On the other hand, calcium concentration limiting values yielded slightly lower values (23.1 mg/L) (Karatayev et al., 2015c). Lower calcium concentration limits (8–15 mg/L) have been reported for the St. Lawrence River (Jones & Ricciardi, 2005; Mellina & Rasmussen, 1994), but these lower values may reflect the limits for the survival of adult mussels, rather than the establishment of locally sustainable populations (Karatayev et al., 2015c). Sprung (1987) found that calcium concentrations of 24 mg/L are needed for the successful development of zebra mussel embryos and larvae. In lakes, population development is typically dependent upon local larval development and recruitment. In rivers, on the other hand, local populations may be supported by larvae produced upstream, where local colonization represents sink populations (Karatayev & Burlakova, 1995a; Horvath et al., 1996; Allen & Ramcharan, 2001; Schneider et al., 2003; Stoeckel et al., 2004; Karatayev et al., 2015c).
There are no published data on pH or calcium limits for quagga mussels (reviewed in Karatayev et al., 2007b, 2015c). The only exception is the report by Kirkendall et al. (2021), suggesting that, in Lake Huron, a threshold calcium concentration of around 22 mg/L is necessary for veligers to survive to larger sizes and produce juvenile benthic recruits. This threshold is similar to one found by Sprung (1987) for D. polymorpha veligers. Both in Europe and in North America, D. polymorpha have colonized many more waterbodies than have quagga mussels. With few exceptions, almost all habitats had already been colonized by zebra mussels when quagga mussels invaded, suggesting that the pH limits for both species of Dreissena largely overlap.
Waterbody types
Dreissenids can be found in a wide range of waterbodies; however, most work has been focused on factors affecting their presence and abundance in lakes and reservoirs (reviewed in Karatayev et al., 1998), and far more studies were performed on D. polymorpha than on D. r. bugensis (Karatayev et al., 2015a). Strayer (1991) suggested that small warm lakes are more suitable for D. polymorpha than deep and cold lakes. It was long noted that the waterbody’s trophic status affects the probability of colonization by D. polymorpha (Deksbakh, 1935; Lyakhnovich et al., 1994; Karatayev & Burlakova, 1995a, b; Karatayev et al., 2003). D. polymorpha is most often found in mesotrophic lakes, less often in oligotrophic and meso-oligotrophic lakes, least often in eutrophic lakes, and almost never in dystrophic lakes with low pH and calcium concentration (reviewed in Karatayev et al., 1998, 2003). In rivers, D. polymorpha is most limited by the suitable substrates for attachment, mobile bottom sediments, and high concentrations of suspended matter, especially during periodic flooding events (reviewed in Karatayev et al., 1998, 2010). Unidirectional water flow in rivers hinders local recruitment as larvae are swept downstream (Horvath et al., 1996; Allen & Ramcharan, 2001; Schneider et al., 2003; Stoeckel et al., 2004). Therefore, in rivers D. polymorpha seldom attain high densities, with the exception of outflow areas in the vicinity of invaded lakes or reservoirs which provide a source of larvae and juveniles (Lyakhnovich et al., 1994; Karatayev & Burlakova, 1995a; Horvath et al., 1996; Allen & Ramcharan, 2001; Schneider et al., 2003; Stoeckel et al., 2004). D. polymorpha can also form high densities in the lower courses of rivers and deltas due to reduced water flow and sediment remobilization (Kuchina, 1964; Grigoriev, 1965; Kharchenko & Shevtsova, 1983; Lyakhnovich et al., 1994). Finally, in canals dreissenids densities are much higher than in lakes and reservoirs owing to a constant unidirectional water current that consistently deliver food and oxygen and are higher than in rivers due to much stable bottom sediments and lower concentration of suspended inorganic matter (Lvova, 1977; Kornobis, 1977; Kharchenko & Shevtsova, 1983; Karatayev et al., 1998, 2010).
Distribution within waterbodies
Lake size and morphometry determine how deep zebra and quagga mussels can spread, the depth range where they thrive best, and their overall population sizes. In shallow polymictic lakes the distribution of D. polymorpha is driven by the availability of hard substrate for attachment and wave activity, and if appropriate substrates are available, mussels may colonize the whole bottom (Karatayev et al., 1998, 2021a; Nalepa et al., 1995, 1996; Hunter & Simons, 2004). In the littoral zone of small polymictic lakes, D. polymorpha usually attains maximum population densities between 1 and 6 m deep (Stanczykowska, 1977; Lyakhnovich et al., 1994; Karatayev et al., 1998; Burlakova et al., 2006; Goedkoop et al., 2011). D. polymorpha very rarely colonizes soft unconsolidated sediments, attaining highest densities on natural and artificial hard substrates, including rocks, shells, silty sand, and the submerged portions of macrophytes (Mordukhai-Boltovskoi, 1960; Lewandowski, 1982b; Draulans & Wouters, 1988; Lyakhnovich et al., 1994; Karatayev & Burlakova, 1995a, Nalepa et al., 1995; Burlakova et al., 2006; Hetherington et al., 2019). D. polymorpha can be extremely abundant on the submerged parts of aquatic vegetation, but here their populations are usually represented by young mussels because adults do not survive after macrophyte stems die back (Lewandowski, 1982b; Lyakhnovich et al., 1994; Burlakova et al., 2006). To our knowledge, the largest density of zebra mussels ever reported (9,169,000 ind./m2) was found on semi-submerged macrophytes in the South Bug Liman (Grigoriev, 1965). High densities of D. polymorpha may also be found in newly constructed reservoirs where the bottom is covered with a thin layer of silt, but as reservoirs age and silt builds up, zebra mussel densities decline (reviewed in Lyakhnovich et al., 1994; Karatayev et al., 1998).
In the shallowest areas of large, stratified lakes, strong wave activity inhibits the settlement of D. polymorpha, but their deeper thermocline allows oxygenated water and food to reach deeper bottom areas thus permitting mussels to spread down to 50 m with maximum densities at 18 m (Lake Garda, Italy, Franchini, 1978) and even down to 55 m with maximum densities at 5–15 m (Lake Constance, Germany-Switzerland-Austria, Walz, 1973). So far, the deepest occurrence of D. polymorpha was recorded in the Laurentian Great Lakes: in Lake Huron down to 80 m, peaking at 20–30 m (Nalepa et al., 2007), in Lake Ontario down to 109 m, peaking at 18–38 m (Watkins et al., 2007), and in Lake Michigan down to 128 m (Nalepa et al., 2014), peaking at 27–46 m (Fleischer et al., 2001). Overall, however, the bulk of D. polymorpha populations in the deep Great Lakes is always limited to a relatively narrow band in the nearshore zone with > 99% of lake-wide populations occurring at < 50 m depths in Lake Michigan and at < 30 m in lakes Ontario and Huron (Karatayev et al., 2021a; Karatayev & Burlakova, this issue). Likewise, in Lake Constance, maximum densities of newly settled juvenile mussels occur at 4 m depth, with deeper areas (15 m and 30 m) showing only isolated settlement clusters (Wacker & Von Elert, 2003). Thus, zebra mussels are largely limited to the well-mixed littoral zone and, even if suitable substrates are available, never form high densities below the thermocline due to the low temperatures for growth, scarce food, and, occasionally, low oxygen concentrations (Grim, 1971; Walz, 1973, 1978a, 1978b; Karatayev et al., 1998, 2015a, 2021a; Burlakova et al., 2006; Goedkoop et al., 2011; Hetherington et al., 2019).
Similar to zebra mussels, in areas exposed to currents and wave action in shallow polymictic lakes quagga mussels require hard substrate for attachment (Nalepa et al., 2010; Burlakova et al., 2012). Therefore, in the littoral zone both species have extremely patchy distributions and may form large, isolated multilayer druses separated by bare sediment with few or no mussels (Karatayev et al., 1983, 1997; Nalepa et al., 2010; Burlakova et al., 2012). Nevertheless, even in polymictic lakes and well-mixed littoral areas of deep lakes, the distribution of both species is not identical, with D. r. bugensis forming higher densities on soft sediments and zebra mussels dominating in most shallow areas exposed to strong wave activity and stronger currents, as well as on macrophytes (Diggins et al., 2004; Karatayev et al., 2013, 2021a; Hetherington et al., 2019).
A dramatic difference between the distribution patterns of both dreissenid species was found in the deep Great Lakes. In the profundal zone of lakes Ontario, Michigan, and Huron, quagga mussels colonize the entire bottom spreading down to the maximum depths (> 200 m), with peak densities at 30 – 100 m and attaining populations densities exceeding those of D. polymorpha by an order of magnitude (Watkins et al., 2007; French et al., 2009; Nalepa et al., 2010, 2014, 2020; Karatayev et al., 2021a, 2021b, 2022). The ability of D. r. bugensis to colonize the profundal zone of deep lakes and reservoirs suggests that quagga mussels inherited some features from the ancestral D. rostriformis which inhabited colder, oligotrophic, profundal waters in ancient large lakes, and currently occupies depths from 30 to 300 m in the profundal zone of the Caspian Sea (reviewed in Orlova et al., 2005).
Dispersal
Major pathways
The spread of exotic dreissenids has not been a continuous process, but rather punctuated by periods of intense activity associated with major geographic expansions (Karatayev et al., 2007b). Most of these “jumps” were associated with changes in the tempo of human activities that provided previously unavailable means of spread, such as the construction of shipping canals for trade, building of reservoirs for water storage and power production, political boundary changes and changes in political systems which affected the position or permeability of national borders, changes in the mode and volume of international trade, and recent industrial practices and environmental regulations (Karatayev et al., 2007b). In spite of similar life history traits, the two dreissenid species have very different invasion histories and exhibit different rates of spread on both global (country or a major region within a country in Europe and states and provinces in North America) and local (waterbodies including lakes, reservoirs, ponds, and quarries) scales (Zhadin, 1946; Kerney & Morton, 1970; Kinzelbach, 1992; Starobogatov & Andreeva, 1994; Karatayev et al., 2007b, 2011a, 2014a; Pollux et al., 2010; Bidwell, 2010; Benson, 2014; Benson et al., 2021a, b). Once established in a new system, dreissenid larvae can spread downstream and between hydrologically connected waterbodies over considerable distances with the aid of currents (Horvath et al., 1996; Johnson & Padilla, 1996; Jantz & Nauman, 1998; Kraft et al., 2002; Schneider et al., 2003). Other natural vectors of spread suggested in the literature, like waterfowl, amphibians, crayfishes, and turtles do not seem to play a significant role in their dispersal (Carlton, 1993; Johnson & Carlton, 1996; Karatayev et al., 2003; Pollux et al., 2010). Although both species can utilize the same vectors of spread, D. polymorpha can attach more strongly to substrates (Peyer et al., 2009, 2010; Collas et al., 2018), which favors a much more efficient dispersion pathway for the adult stage, even overland, between isolated waterbodies. Limited data suggest that the larvae of D. r. bugensis have a longer planktonic stage (Wright et al., 1996), which presumably may allow them to disperse with currents over longer distances than D. polymorpha.
Dreissena polymorpha
Since the last glaciation and until the eighteenth century, the geographic range of D. polymorpha was limited to the river basins of the Black, Azov, and Caspian seas, isolated and semi-isolated relic estuarine reservoirs along coasts, and in areas of these seas influenced by freshwater in Russia, Bulgaria, Hungary, Romania, Moldova, Ukraine, Kazakhstan, and Turkey (Zhadin, 1946; Mordukhai-Boltovskoi, 1960; Kinzelbach, 1992; Starobogatov & Andreeva, 1994; Son, 2007; Arslan et al., 2018; Fig. 3). It is also likely that the native range of D. polymorpha included Northern Iran, Turkmenistan, and Uzbekistan (Starobogatov, 1994c; Starobogatov & Andreeva, 1994; Andreeva & Andreev, 2003). We should also emphasize that, although the Aral Sea basin is included in the native range, D. polymorpha in this area is represented by the endemic subspecies D. p. obtusecarinata and D. p. aralensis (see Habitat Requirements: Salinity).

Distribution of Dreissena polymorpha in Eurasia. The presumed native range (the lower reaches of river basins of Black, Azov, and Caspian seas, as well as waters of these seas influenced by freshwater) is denoted with red hatching (the full extent of the rivers inhabited is not shown). Years indicate first records, followed by letters denoting the corresponding countries, as identified by their internet domain names. Countries where the mussel is almost certainly present, but we did not find records in the literature (mostly in its presumed native range), are also included. AL Albania, AM Armenia, AT Austria, AZ Azerbaijan, BA Bosnia and Herzegovina, BE Belgium, BG Bulgaria, BY Belarus, CH Switzerland, CZ Czech Republic, DE Germany, DK Denmark, EE Estonia, ES Spain, FR France, GE Georgia, GR Greece, HR Croatia, HU Hungary, IE Ireland, IR Iran, IT Italy, LT Lithuania, LU Luxembourg, MK Macedonia, NL Netherlands, PL Poland, PT Portugal, RO Romania, RS Serbia, RU Russian Federation, SE Sweden, SI Slovenia, SK Slovakia, TM Turkmenistan, TR Turkey, UA Ukraine, UK United Kingdom, UZ Uzbekistan. Green—countries where the species is present; faded green—the species is almost certainly present, but we did not find records in the literature; white—no records of the species
However, as the demand for Russian firewood and other goods grew considerably in western Europe, three navigation canals connecting the Black Sea and Baltic Sea basins were built in western Russia (now Belarus). These canals connected the Dnieper and Zapadnyi Bug rivers (early 1800s), the Dnieper and Neman rivers (1784), and the Dnieper and Zapadnaya Dvina rivers (1805) (Karatayev et al., 2008). After the start of regular commercial ship traffic, D. polymorpha spread quickly throughout eastern and western Europe (Zhadin, 1946; Deksbakh, 1935; Mordukhai-Boltovskoi, 1960; Starobogatov & Andreeva, 1994; Karatayev et al., 2007b, 2008, 2011a; Pollux et al., 2010; Bidwell, 2010; bij de Vaate et al., 2014; Orlova, 2014; Fig. 3). During the nineteenth century (1800 to 1867), D. polymorpha spread across Europe at an exponential rate (Figs. 3, 4; Karatayev et al., 2011a). By the beginning of the 1870s, most of the countries in Europe with extensive trade and habitats suitable for colonization were invaded; thus, further range expansion essentially slowed down, and over the next 90 years (1871 – 1961), D. polymorpha colonized only three additional regions of Europe (Fig. 4, reviewed in Kinzelbach, 1992; Karatayev et al., 2007b, 2011a) and expanded its range in Turkey after 1936 (Aksu et al., 2017).

Timeline of the spread of D. polymorpha and D. r. bugensis in Europe and North America. Circles: first records in Europe (countries or geographical provinces within large countries); squares: first records in the USA, Canadian provinces, and Mexico; triangles: first records in both Europe and North America in the same year
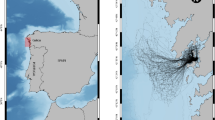
However, because shipping was the major vector of spread in the nineteenth century by mussels attached to boat hulls or to timber transported by ships (Zhadin, 1946; Kerney & Morton, 1970; Kinzelbach, 1992), mostly countries with intense ship traffic were colonized during that century. As a result, D. polymorpha was not recorded in Ireland until 1993 (Minchin, 2000) and was only found in Spain in 2001 (bij de Vaate et al., 2002) (Fig. 3). In addition, increased pollution associated with the Industrial Revolution, especially in rivers and ports, further prevented the spread of D. polymorpha (Kinzelbach, 1992; Karatayev et al., 2007b, 2011a; bij de Vaate et al., 2014).
Globalization of the economy, recreational activities, construction of new canals, and water quality improvement provided new means for spread and triggered the second exponential spread of zebra mussels both in Europe and in North America, which was significantly faster than the spread in the nineteenth century (Karatayev et al., 2011a; bij de Vaate et al., 2014) (Fig. 4). In the 1960s D. polymorpha was recorded in isolated high alpine lakes where they were transported attached to recreational boats trailered overland, leading to colonization of lakes Geneva, Zurich, and Constance in Switzerland (Kinzelbach, 1992; Pollux et al., 2010). According to Pollux et al. (2010), the movement of watercraft to and from high alpine lakes resulted in the colonization of Italy (1969), Yugoslavia (1970s), and Spain (2001; Araujo et al., 2010). The import of second-hand leisure boats was responsible for the colonization of Ireland (Pollux et al., 2003). In 2019, D. polymorpha was first found in Portugal (Catita et al., 2020). By 2019 virtually all European countries were colonized by this invader (Fig. 3). Finally in 2021, D. polymorpha was found for the first time in the Pyshma River in the West Siberian Plain (Babushkin et al., 2022). By 2021 zebra mussels were present in 31 European countries and 10 additional geographical provinces within large countries, including their native area (Fig. 3, 4). In addition, the distribution range of D. polymorpha increased in Turkey (Aksu et al., 2017) and, possibly, in a few other Asian countries adjacent to the Ponto–Caspian basin.
Increases in the wheat trade between North America and the former Soviet Union facilitated the introduction of D. polymorpha and D. r. bugensis into North America in the 1980s by ballast water discharges into the Great Lakes (Hebert et al., 1989; Mills et al., 1993; Karatayev et al., 2007b; Carlton, 2008). The Soviet Union suffered poor wheat harvests in the late 1970s and became dependent upon foreign sources of grain, resulting in the dramatic increase in wheat shipments to the Soviet Union (Karatayev et al., 2007b). As for many (or most) invasions by aquatic species, globalization of trade became the primary driver of the spread of these two mussels (reviewed in Ruiz & Carlton, 2003b). Within the USA, D. polymorpha initially spread across regions at an exponential rate and then slowed with the saturation of nearby regions after 1993 (reviewed in Karatayev et al., 2011a). Less than 40 years after its first sighting in North America, D. polymorpha was present in 31 states and 3 Canadian provinces (Fig. 5, 6; Benson et al., 2021a).


Spread at the local scale. Although the spread of D. polymorpha to waterbodies connected through shipping canals was rapid, subsequent dispersion to isolated waterbodies was slower and was constrained by various factors, including political borders, economic systems (socialism vs. capitalism), and recreational and commercial activities (Karatayev et al., 2003, 2007b). The establishment of a socialist political system in eastern Europe restricted trade with western Europe and led to the development of very different socio-economic regimes, which in turn resulted in different vectors responsible for the spread of D. polymorpha in these two parts of Europe (Karatayev et al., 2007b, 2010). For example, what is now Belarus was from 1922 to 1939 part of the USSR and Poland; here, from the early 1800s to the mid-1950s, the rate of spread of D. polymorpha was very low, averaging approximately 4 lakes every 50 years (Karatayev et al., 2010). After these borders changed and the Second World War ended, between 1960 and 2000 the rate of spread increased dramatically to an average of 2 lakes per year (Karatayev et al., 2010).
The transport mechanisms and vectors responsible for the spread of D. polymorpha to isolated lakes and lake systems were different in eastern and western Europe largely due to contrasting economies and social structures. In western Europe, leisure boat traffic was the major vector of spread to isolated lakes (Kinzelbach, 1992; De Ventura et al., 2016), while in the Soviet Union it was commercial fishing subsidized by the government (Deksbakh, 1935; Karatayev et al., 2003, 2010). When the Soviet Union dissolved, commercial fisheries subsidies were discontinued, resulting in sharp declines in commercial fishing activity (Karatayev et al., 1999, 2010). However, subsequently recreational activities increased and became responsible for the spread of D. polymorpha (Karatayev et al., 2010). Recreational boats are the primary vector for the spread of dreissenids among regions and waterbodies in the USA as well (Padilla et al., 1996; Buchan & Padilla, 1999; Johnson et al., 2001; Bossenbroek et al., 2007).
Dreissena rostriformis bugensis
Although data on the native range of D. r. bugensis are controversial (Mills et al., 1996; Son, 2007; Zhulidov et al., 2010), currently their native distribution is believed to include the whole Dnieper–Bug Liman system (a large coastal lake connected to the Black Sea estuary), the Dnieper River delta, and the lower reaches of the South Bug and Ingulets Rivers (Zhulidov et al., 2010). During the middle twentieth century when ship traffic between areas inhabited by D. r. bugensis and other regions of eastern and western Europe was intensive, D. r. bugensis did not follow the dispersion pattern of D. polymorpha and remained restricted to its native range until the 1940s. In 1941 D. r. bugensis was introduced into the Dnieprovskoe Reservoir on boats evacuated from ports of the Dnieper–Bug Liman system during the Second World War. With the construction of new reservoirs on the Dnieper River in the 1950s and 1960s and canals for commercial shipping and irrigation, D. r. bugensis began spreading throughout eastern and western Europe (Zhuravel, 1965, 1967; Mills et al., 1996; Orlova et al., 2005; Zhulidov et al., 2010; Karatayev et al., 2007b, 2011a; Orlova, 2014). During the first 40 years of invasion (1940s – 1980s) the rate of spread of D. r. bugensis was low (reviewed in Orlova et al., 2005) (Fig. 4, 7). It dispersed northward, upstream along the Dnieper River reservoirs, southward along irrigation canals on the Crimean Peninsula (Kharchenko, 1995; Mills et al., 1996; Son, 2007), and eastward to the lower Don River system, where it was first recorded in 1980 (Zhulidov et al., 2004; Son, 2007). In 1981, D. r. bugensis was reported in the Volga River basin (Zhulidov et al., 2004; Son, 2007) and by 2001, the species was present in a 3000-km stretch of the river from Rybinsk Reservoir in the north to the Caspian Sea Delta in the south (Antonov & Kozlovsky, 2001; Orlova & Shcherbina, 2001; Orlova et al., 2004; Orlova, 2014; Zhulidov et al., 2004, 2005; Son, 2007). In 1988, D. r. bugensis was found in the Dniester River in Ukraine (Shevtsova, 2000) and in 1992, in Moldova (Son, 2007).

Distribution of Dreissena rostriformis bugensis in Eurasia. The presumed native range (Dnieper–Bug Liman, Dnieper River delta, and the lower reaches of the South Bug and Ingulets rivers in Ukraine) is denoted with red hatching (the full extent of the rivers inhabited is not shown). Years indicate first records, followed by letters denoting the corresponding countries, as identified by their internet domain names. AT Austria; BE Belgium; BG Bulgaria; CZ Czech Republic; DE Germany; FR France; IE Ireland; MD Moldova; NL Netherlands; PL Poland; RO Romania; RS Serbia; RU Russian Federation; SI Slovenia; UA Ukraine; UK United Kingdom. Green—countries where the species is present; white—no records of the species
Starting from the 1990s, the rate of spread of D. r. bugensis increased greatly (Figs. 4, 7). In 2003, adult mussels were found in the Moscow River within the city of Moscow (Lvova, 2004). In the first decades of the twenty-first century, D. r. bugensis was present in the Danube River in Romania (2004, Micu & Telembici, 2004), Bulgaria (2006, Hubenov & Trichkova, 2007), and Serbia (2010, Rakovic et al., 2013). In 2008 it was found in Lake Balaton, Hungary (Balogh et al., 2018).
In 2006, the first record of this species for western Europe was in Hollandsch Diep (The Netherlands), in the Rhine delta (Molloy et al., 2007). The introduction likely occurred via the Main–Danube Canal, a southern corridor for movement of non-indigenous aquatic species between western and eastern Europe (bij de Vaate et al., 2002; Müller et al., 2002). This introduction in the Rhine River was most likely a result of a jump dispersal, either via ballast water transport of larvae from the Black Sea or North American port into the Port of Rotterdam or Hollandsch Diep or via the transport of adult mussels attached to inland ship hulls (van der Velde & Platvoet, 2007; bij de Vaate, 2010; Heiler et al., 2013; bij de Vaate et al., 2014).
Regardless of the vectors and the origins of the introductions into western Europe, D. r. bugensis spread quickly into other western European countries, including Germany (2007), Belgium and Austria (2009), France (2011), the UK (2014), and Switzerland (2014) (Van der Velde & Platvoet, 2007; Sablon et al., 2010; Heiler et al., 2013; Marescaux et al., 2012; Aldridge et al., 2014). In 2021, D. r. bugensis was also found in Ireland (Mainnín, 2021). By 2021, D. r. bugensis was present in 16 European countries and 4 additional geographical provinces within large countries (Fig. 4, 7).
D. r. bugensis expanded geographically in North America at a much slower rate than D. polymorpha during the first 8 years of invasion (1989 – 1996) (reviewed in Karatayev et al., 2011a), and this rate did not change in the next decades. In 2006, D. r. bugensis jump-spread to the western USA likely on trailered long-distance boats and are now found in 5 western states from Colorado to California (Benson et al., 2021b). Overall, by 2021 D. r. bugensis was recorded in 17 states and 2 provinces of three countries in North America (Canada, Mexico, and the USA) (Fig. 5; Benson et al., 2021b).
Factors affecting the rate of spread
The rate of spread of D. polymorpha depends upon the spatial resolution of the scale used and is much slower at a local (i.e., isolated lakes or lake systems), than on a global scale (e.g., countries) (Karatayev et al., 2011a). In the last two centuries, D. polymorpha colonized virtually all European countries, but at the local scale their spread in Europe is still far from complete. For example, only 16.8% of the 553 Belarusian lakes studied had been colonized in 1996, after 200 years of continuous expansion (Karatayev et al., 2003). In North America, D. polymorpha had largely exhausted its potential for further spread at the global scale by 2021 as the majority of suitable states and provinces had already been invaded. However, their spread on the local scale is much slower: for example, in the state of Wisconsin, after > 30 years only 168 (1.1%) of the > 15,000 lakes were invaded by 2021 (Benson et al., 2021a).
Nevertheless, the rate of spread was shown to be much faster for the zebra than for the quagga mussel at virtually all spatial scales (Orlova et al., 2005; Karatayev et al., 2011a, 2015a). Although both species of Dreissena were introduced into North America at about the same time (Mills et al., 1993; van der Velde & Platvoet, 2007; Carlton, 2008; Matthews et al., 2014), by 2021 D. polymorpha had colonized 1.7-fold more the USA states than D. r. bugensis (31 vs. 17) and > 17 times (1230 vs. 71, respectively) more waterbodies (Fig. 5; Benson et al., 2021a, b). Initially, D. polymorpha spread exponentially in the USA, invading 20 states during the first 8 years (1986 – 1993; 2.5 states/year), but in the following 27 years (1994 to 2021), the rate of spread slowed down, and only 11 additional states (0.41 states/year) were colonized. In contrast, during the first 8 years of invasion (1989 – 1996) D. r. bugensis expanded geographically at a much slower rate than D. polymorpha, and this rate did not change in 1997 – 2021 (0.63 vs. 0.50 states/year, respectively).
Although over the last 3 – 4 decades D. r. bugensis exhibited a relatively high rate of spread in both Europe and in North America, the timings of the geographic expansion of the two dreissenids differ widely, and the reasons for this difference are probably multiple. Adults of D. polymorpha were transported attached to boat hulls and rafts or to their cargo (e.g., timber) within the continent, attached to boat hulls of marine vessels or timber transported overseas by ships between ports (Zhadin, 1946; Kerney & Morton, 1970; Kinzelbach, 1992), and attached to recreational boats in North America (Johnson & Carlton, 1996; Johnson & Padilla, 1996; Johnson et al., 2001, 2006; Padilla et al., 1996; Karatayev et al., 2013). Because of its higher attachment capabilities, D. polymorpha is much more efficient in colonizing substrates suspended in the water column (e.g., macrophytes, navigation buoys, and boat hulls) than D. r. bugensis (Conn & Conn, 2007; Diggins et al., 2004; Karatayev et al., 2013) and due to their flattened ventral shell surface and their higher rate of byssal thread production, D. polymorpha are more resilient to accidental dislodgment than D. r. bugensis (Claxton et al., 1998; Dermott & Munawar, 1993; Mackie, 1991; Peyer et al., 2009, 2010; Collas et al., 2018). This ability is critical for the overland dispersal via macrophytes entangled in boats, trailers, and fishing gear or the direct attachment to these structures (primarily responsible for the short distance spread by the recreational boats). Although quagga mussels represent ~ 99% of the dreissenids of Lake Ontario and the eastern basin of Lake Erie, zebra mussels were more abundant on boat hulls and significantly larger than quagga mussels, making these waterbodies a significant source for zebra mussels spread, despite their comparatively low densities (Karatayev et al., 2013).
In addition, in the nineteenth century D. polymorpha also colonized the shallow areas of rivers and canals (Karatayev et al., 2011a), habitats not favorable for D. r. bugensis which prefer still-water areas of deep lakes and reservoirs (Orlova et al., 2005; Karatayev et al., 2021a). Many large, deep reservoirs constructed in the second half of the twentieth century became the steppingstones for D. r. bugensis (Orlova et al., 2005; Karatayev et al., 2007b, 2011a), where they can thrive due to their higher tolerance to siltation, low oxygen conditions, and low temperatures (Zhuravel, 1967; Pligin, 1979; Antonov & Kozlovsky, 2001; Orlova & Shcherbina, 2001). As a matter of fact, in the Laurentian Great Lakes (except Lake Superior where low calcium concentrations likely prevent population establishment, Vanderploeg et al., 2002) D. r. bugensis inhabits chiefly the cold, deep areas (Patterson et al., 2005; Watkins et al., 2007; Nalepa et al. 2009a, 2010, 2014, 2020; Karatayev et al., 2015a, 2021a, 2022). Over the last decades new vectors of spread, such as ballast water, facilitated the spread of the mussels’ planktonic larvae, while the increased rates of commercial and recreational boat traffic in general, including overland movement of pleasure boats, facilitated the spread of both species (Ricciardi & MacIsaac, 2000; Karatayev et al., 2007b; van der Velde & Platvoet, 2007; van der Velde et al., 2010b; bij de Vaate et al., 2014; Heiler et al., 2013).
Impacts and effects
Ecological impacts
Both Dreissena species are sessile suspension feeders that attach to the substrate with byssal threads and form three-dimensional reef-like structures altering the benthic environment (local impacts). They also affect the planktonic community, trophic relationships, and nutrient cycling via their feeding activity (system-wide impacts) (Karatayev et al., 1997, 2002, 2007a, 2007b, 2015b; Beekey et al., 2004; Higgins & Vander Zanden, 2010; Kelley et al., 2010; Burlakova et al., 2012). Historically, most of the information on their ecological impacts was based on the zebra mussel (reviewed in Karatayev et al., 1994a, 1997, 2007a, 2015a) or on dreissenids in general, as these two species are often considered to have very similar ecological impacts and are therefore treated jointly (Karatayev et al., 2007b; Ward & Ricciardi, 2007; Higgins & Vander Zanden, 2010; Kelley et al., 2010; Kissman et al., 2010). As a result, data on the impacts of quagga mussels only are comparatively few and virtually no data are available for lakes colonized solely by quagga mussels (Karatayev et al., 2007a, 2015a, 2015b; Fahnenstiel et al., 2010; Barbiero et al., 2018). Most of the studies were performed in polymictic lakes, although the lake type was often not reported. Of 2,853 publications on dreissenids in SCOPUS (see Research Efforts), more publications (25%) described positive ecosystem services (e.g., provision of food, water purification, changes in nutrient cycle, and positive effects on benthos and macrophytes, not including bioindication services described below in Uses) compared to studies describing negative ecological impact (9%). In this chapter we describe dreissenid ecosystem functions resulting in both positive and negative ecological impacts.
Local impacts
The local impacts of dreissenids on benthic communities are associated with their role as ecosystem engineers (reviewed in Karatayev et al., 2002, 2007a; Vanderploeg et al., 2002; Gutierrez et al., 2003; Zhu et al., 2006; Sousa et al., 2009), as they can physically change bottom substrates creating new three-dimensional structures that provide refuge from predation and other stressors (waves, currents, desiccation) for benthic organisms that would otherwise be absent from this environment (Karatayev & Lyakhnovich, 1990; Karatayev et al., 1994a, 1997, 2002, 2007a, 2007b; Gutierrez et al., 2003; Nalepa et al., 2003; Beekey et al., 2004; Burlakova et al., 2012; Rennie & Evans, 2012). Multiple studies conducted in the littoral zone showed that dreissenids have positive effects on most native epifaunal invertebrates, including amphipods, isopods, leeches, turbellarians, and hydrozoans, as well as some oligochaetes and chironomids, which take advantage of both the structural complexity and the food resources provided by the zebra mussels and associated organisms (Karatayev et al., 1983, 1994a, 1997, 2002, 2007a, 2007b; Mayer et al., 2002; Gutierrez et al., 2003; Beekey et al., 2004; Ward & Ricciardi, 2007; Higgins & Vander Zanden, 2010; Ozersky et al., 2011; Burlakova et al., 2012). The overall density and biomass of native invertebrates are always higher in Dreissena beds as compared with nearby bare sediments (reviewed in Karatayev et al., 1983, 1997, 2002, 2007b; Burlakova et al., 2012), resulting in significant increases in the abundance of native invertebrates in the littoral zone (reviewed in Karatayev et al., 1997, 2002; Ward & Ricciardi, 2007; Higgins & Vander Zanden, 2010).
These data were largely obtained from research conducted on zebra mussels in polymictic lakes or the littoral zone of large, stratified lakes (Karatayev et al., 2015a). Less information is available for quagga mussels, especially from the profundal zone of deep stratified lakes. In general, in the soft sediments of the Great Lakes profundal zone quagga mussels do not create large multilayered beds, often covering the entire bottom in a single layer where animals are spaced apart by the length of their siphons (at 30 – 90 m) or form small clusters that float on the surface of soft silt (at > 90 m) areas (Dermott & Kerec, 1997; Karatayev et al., 2021b; Karatayev & Burlakova, this issue). In addition, most profundal zone benthic invertebrates are less mobile (Merritt et al., 1984), and there are almost no epifaunal species that could take advantage from the dreissenid aggregations. Native filter feeders such as sphaeriids and the deep-water detritivorous amphipods Diporeia sp. are negatively affected by dreissenids due to the trophic competition in this food-limited deep area, especially during summer stratification (Dermott, 2001; Lozano et al., 2001; Dermott & Geminiuc, 2003; Watkins et al., 2007; Nalepa et al., 2006, 2009a, b, 2014; Burlakova et al., 2018; Bayba et al., 2022). However, with the recent proliferation of quagga mussels in the deep areas of lakes Michigan and Huron, the density and biomass of Oligochaeta were found to increase in dreissenid aggregations across all depth gradients, and the depth-related decline in native benthos density and biomass has decreased with increasing quagga mussel populations (Bayba et al., 2022).
System-wide impacts: D. polymorpha
The system-wide effects of zebra mussels are associated with their role as suspension feeders. Suspension feeding not only affects planktonic communities, it also transfers materials from the water column to the benthos, enhancing the coupling between the planktonic and the benthic components of the ecosystem, which can trigger a suite of changes that increase the relative importance of the benthic community – a process referred to as “benthification” (Mayer et al., 2014) resulting in the redistribution of invertebrate biomass and secondary production from the pelagic to the benthic environment (reviewed in Karatayev et al., 1997, 2002). Before zebra mussel invasion in Lake Lukomskoe (Belarus), the lake-wide average zooplankton wet biomass (7.6 g/m2) was almost 20-fold higher than that of the zoobenthos (0.4 g/m2), allowing planktonic filterers to filter the volume equivalent of that of the lake in 5 days, much faster than benthic filter feeders (15 years). Shortly after the invasion, the benthic biomass increased over 77-fold (mostly due to Dreissena), zooplankton abundance declined 30-fold, and the time required for the zooplankton to filter the equivalent of the volume of the lake increased to 17 days, while the filtering capacity of benthic invertebrates had increased 320 times, allowing to filter the volume of the lake in 17 days (Karatayev & Burlakova, 1992, 1995b; Karatayev et al., 1997).
The introduction of zebra mussels is associated with an increase in water transparency and a reduction in the concentration of seston, phosphorous, chlorophyll, and phytoplankton (reviewed in Karatayev et al., 1994a, 1997, 2002, 2021d; Ibelings et al., 2007; Kelly et al., 2010; Higgins & Vander Zanden, 2010; Goedkoop et al., 2011; Mayer et al., 2014; Noordhuis et al., 2016). Improved water transparency increases macrophyte coverage and biomass as macrophytes grow deeper in the lake (reviewed in Karatayev et al., 1997, 2002, 2021d; Higgins & Vander Zanden, 2010; Zhu et al., 2006; Ibelings et al., 2007; Mayer et al., 2014; Noordhuis et al., 2016; Wegner et al., 2019). Macrophytes may hinder water mixture which, together with their nutrient consumption, may decrease allochthonous nutrient availability for the phytoplankton. Increased light penetration also stimulates the growth of periphyton and benthic algae. These changes often trigger a shift from a turbid to a clear water state (Ibelings et al., 2007; Karatayev et al., 2014c; Mayer et al., 2014; Noordhuis et al., 2016). This ability of dreissenids to reduce phytoplankton biomass and increase water clarity was suggested by European scientists as a tool for using the mussels as biofilters in order to decrease the effects of anthropogenic eutrophication in lakes (Karatayev, 1983; Reeders et al., 1989, 1993; Reeders & bij de Vaate, 1990; Karatayev et al., 2010; Noordhuis et al., 1992, 2016; Stybel et al., 2009; Dionisio Pires et al., 2010; Goedkoop et al., 2011; McLaughlan & Aldridge, 2013; Reynolds & Aldridge, 2021). The decline in phytoplankton may also trigger declines in zooplankton but impacts on the zoobenthos are more heterogeneous (reviewed in Karatayev et al., 1997, 2002, 2021d; Mayer et al., 2002; Zhu et al., 2006; Kelly et al., 2010; Higgins & Vander Zanden, 2010; Kissman et al., 2010; see Local Impacts).
The magnitude and extent of system-wide effects depend on many factors, including mussel density and distribution in the waterbody, water mixing rates, retention time, lake morphology, and time since the initial invasion (Karatayev et al., 1997, 2002; Reed-Andersen et al., 2000; Kelly et al., 2010). Because zebra mussels are usually restricted to the littoral zone, their impacts may be significantly greater in shallow polymictic lakes than in deep dimictic lakes (Karatayev et al., 2014b, 2021a). In addition, the impacts of zebra mussels are time dependent and are the strongest early in the invasion when population levels are high and climbing (Karatayev et al., 1997, 2021d), a pattern observed for many invasive species (Strayer et al., 2006; Bradley et al., 2019).
Recently, long-term studies of six polymictic European and North American lakes showed that the most noticeable ecosystemic changes occur within 5 – 10 years of the invasion (Karatayev et al., 2021d). After 10 – 15 years, the trends in almost all ecosystem parameters affected by zebra mussels get stabilized or even reversed, suggesting at least a partial recovery to the original conditions. This could be due to a decline in zebra mussel biomass after their initial increase and/or due to the adaptation of native species to the altered environment. Substantial mussel declines or their extirpation could trigger a reverse trend in the ecosystem and a regime shift from clear back to turbid waters (Mitrakhovich et al., 2008; Karatayev et al., 2021d). In Lake Lukomskoe, a tenfold decline in zebra mussel biomass 30 years after their peak, most likely driven by the increase in nutrient loads and oxygen depletion caused by a fish hatchery, was followed by the almost complete return of the lake to pre-invasion conditions, including the decline in Secchi depth, macrophyte coverage, non-mussel benthic biomass, and increases in phytoplankton and zooplankton biomass (Karatayev et al., 2021d).
Fewer data are available on the impacts of zebra mussels in stratified lakes (but see Kissman et al., 2010; Jimenez et al., 2011; Halfman, 2016). The establishment of zebra mussels in Seneca Lake (New York) in 1992 led to a 4.5 m increase in Secchi depth and a decline in chlorophyll concentrations in the water column. After their populations crashed in 1998, the release of nutrients previously sequestered by the mussels back to the water column led to a decline in Secchi disk depths and an increase in chlorophyll back to pre-1992 levels (Halfman, 2016). In 25 thermally stratified lakes in Michigan colonized by dreissenids (authors did not report if lakes were colonized by both or one species), chlorophyll a concentration, phytoplankton, microzooplankton, and macrozooplankton biomass became significantly lower, while water clarity increased significantly compared to 25 uninvaded lakes (Kissman et al., 2010). No substantial changes were observed in lakes Michigan and Huron in the 1990s when they were colonized by zebra mussels alone (Fahnenstiel et al., 2010; Kerfoot et al., 2010; Mida et al., 2010; Pothoven & Fahnenstiel, 2014; Barbiero et al., 2018; Pothoven & Vanderploeg, 2020; Karatayev & Burlakova, this issue), suggesting that mussel populations limited to the nearshore areas were not sufficient to produce lake-wide impacts. However, zebra mussel-related impacts were evident in shallow embayments of the Great Lakes, like Saginaw Bay of Lake Huron (Fahnenstiel et al., 1995; Johengen et al., 1995; Skubinna et al., 1995; Vanderploeg et al., 2010).
System-wide impacts: D. r. bugensis
Virtually all studies on the impacts of quagga mussels are from lakes initially invaded by zebra mussels and only later by quagga mussels (but see Reynolds & Aldridge, 2021), making it very hard to tease apart the impacts of the two species. Since they both are suspension feeders, they have similar effects on the waterbodies invaded, suggesting that the magnitude of their impacts should be directly related to their biomass, and the impacts should be additive (Noordhuis et al., 2016). However, the ability of quagga mussels to survive and grow at lower food and temperature regimes than zebra mussels (Diggins, 2001; Baldwin et al., 2002; Stoeckmann, 2003; Nalepa et al., 2010) allow them to consume phytoplankton, reduce chlorophyll concentration, and increase Secchi depths more effectively than zebra mussels, both earlier and later in the growing season, resulting in stronger ecosystem impact per unit of mussel biomass, as was observed in Oneida Lake (Karatayev et al., 2021d). A common scenario of zebra and subsequently quagga mussel, invasions involve strong ecosystem responses during the first 5–10 years, some ecosystem recovery to pre-invasion conditions in the next 5–10 years and finally the disruption and reversal of the recovery after the introduction of quagga mussels, which amplifies the initial zebra-related impacts (Karatayev et al., 2021d).
The polymictic lakes Wolderwijd, Veluwe, and Eem (The Netherlands) initially harbored zebra mussels, which were extirpated in the late 1960s–early 1970s due to the increase in nutrient loadings, causing a shift from clear to turbid water states, along with sharp declines in macrophytes and Secchi depth and increase in planktonic chlorophyll (Ibelings et al., 2007). Reductions in phosphorous loads and removal of benthivorous fishes initiated a shift toward a clear water state, further enhanced by the deliberate re-introduction of zebra mussels in 1996, causing an increase in Secchi depth, macrophyte coverage, and declines in phosphorous concentrations and chlorophyll, including blue-green algae (Ibelings et al., 2007; Noordhuis et al., 2016). These effects were further amplified by the unintentional introduction of quagga mussels in 2009 (Noordhuis et al., 2016).
The impact of both dreissenid species on cyanobacteria is still controversial (reviewed in Kelly et al., 2010; Karatayev et al., 2015a, 2015b; Reynolds & Aldridge, 2021) and needs further investigation. Most European and some North American studies reported that dreissenids can effectively suppress cyanobacteria (often including toxic Microcystis) (Reeders & bij de Vaate, 1990; Strayer et al., 1999; Baker & Levinton, 2003; Dionisio Pires et al., 2005, 2010; McLaughlan & Aldridge, 2013; Noordhuis et al., 2016; Reynolds & Aldridge, 2021), while a number of North American authors suggested that zebra and quagga mussels can promote toxic blooms of Microcystis by selective grazing and rejection of toxic strains or by releasing nutrients to the water column (Vanderploeg et al., 2001, 2009; Conroy et al., 2005a, b).
Fewer data are available on the impacts of quagga mussels on stratified lakes (but see Halfman, 2016). It is reasonable to assume that, because in these waterbodies quagga mussels reach much higher lake-wide biomass values than zebra mussels, their ecological impact is stronger. It also is probable that in deep lakes (as compared with shallow lakes), impacts may take longer to manifest. However, more observations on the population dynamics and ecological impacts of quagga mussels in deep stratified lakes are necessary to test these suggestions.
The expansion of quagga mussels in the 2000s into deeper areas of dimictic lakes Ontario, Michigan, Huron, and the eastern basin of Lake Erie was associated with an almost complete extirpation of zebra mussels and a dramatic increase (22- to 45-fold) of the lake-wide mussel biomass (Karatayev et al., 2021a; Karatayev & Burlakova, this issue), causing strong and consistent system-wide changes, including increases in Secchi depth and declines in total phosphorus concentrations (Dobiesz & Negel, 2009; Dove, 2009; Mida et al., 2010; Barbiero et al., 2012, 2018; Pothoven & Fahnenstiel, 2014; Dove & Chapra, 2015; Pothoven & Vanderploeg, 2020). In lakes Michigan and Huron, strong declines in chlorophyll a were recorded (Barbiero et al., 2012, 2018; Pothoven & Fahnenstiel, 2014; Pothoven & Vanderploeg, 2020), as well as phytoplankton primary production and phytoplankton and zooplankton biomass (Barbiero et al., 2012, 2018; Pothoven & Fahnenstiel, 2014). Phytoplankton depletions derived in the virtually complete disappearance of the winter and spring diatom blooms (Fahnenstiel et al., 2010; Kerfoot et al., 2010; Barbiero et al., 2012), which in turn increased the concentrations of silica in the water column (Barbiero et al., 2012, 2018; Dove & Chapra, 2015). The significant decline in total phosphorus concentrations associated with the dreissenids resulted in the oligotrophication of all deep Great Lakes (Dove, 2009; Evans et al., 2011; Barbiero et al., 2011, 2012, 2018; Chapra & Dolan, 2012; Dove & Chapra, 2015; Karatayev et al., 2018c). Quagga mussels are now the primary driver of phosphorus cycling in the four lower Great Lakes, representing a dramatic example of a large-scale reorganization of the biogeochemical cycle caused by a single organism (Li et al., 2021).
Impacts on unionids
Unionid bivalves often provide the most abundant source of hard substrate for D. polymorpha, especially when other hard substrates are scarce (Sebestyen, 1937; Wiktor, 1963; Hebert et al., 1989; Hunter & Bailey, 1992; Lyakhnovich et al., 1994; Karatayev & Burlakova, 1995b; Nalepa et al., 1996; Burlakova et al., 2000; Hunter & Simons, 2004). Heavy shell infestation may cause mass mortality of the host unionids, and this impact is among the best documented negative ecological consequences of dreissenid invasions (reviewed in Karatayev et al., 1997, 2013; Burlakova et al., 2000, 2014; Strayer & Malcom, 2007; Lucy et al., 2014). Mass mortality of unionids in turn can lead to the decline of D. polymorpha populations as substrates for their attachment decrease (Hunter & Simons, 2004).
By attaching to unionids, D. polymorpha hamper host movement and burrowing, filter feeding, respiration, reproduction, and reduce glycogen reserves, and the added mass of D. polymorpha can weigh down unionids, resulting in burial in unconsolidated sediments (Mackie, 1991; Hunter & Bailey, 1992; Haag et al., 1993; Gillis & Mackie, 1994; Ricciardi et al., 1996; Karatayev et al., 1997; Strayer, 1999; Burlakova et al., 2000; Hallac & Marsden, 2000; Sousa et al., 2011; Lucy et al., 2014). In addition, mussel attachment to unionid valves can increase drag and the likelihood of dislodgment by water motion for species living near the shore (Karatayev, 1983; Karatayev et al., 1997). While the causal mechanisms of unionid mortality as a result of dreissenid infestation are complex, the strong link between the level of infestation and mortality suggests that infestation intensity directly relates to the impact, and a threshold of dreissenid to unionid mass ratio of ≥ 1.0 was suggested to be lethal to the clams (Ricciardi et al., 1995, 1996). Although direct attachment of D. polymorpha to unionids is not the only mechanism that can affect the latter, it was suggested to be a very important component of the overall negative effects (Baker & Hornbach, 2000, 2008).
The severity of D. polymorpha impacts depends on their density in the waterbody, time since invasion, and sediment type (Karatayev et al., 1997; Burlakova et al., 2000; Lucy et al., 2014). Extensive overgrowth of unionids resulting in mass mortality is typical for early invasion periods when populations of D. polymorpha grow rapidly (Sebestyen, 1937; Dussart, 1966; Karatayev & Burlakova, 1995a). Later in the invasion, D. polymorpha can co-exist with native bivalves harmlessly, and although overgrowth can cause some host mortality, unionids not only survive but can also maintain high densities (Karatayev, 1983; Miroshnichenko et al., 1984; Miroshnichenko, 1987; Ponyi, 1992; Burlakova et al., 2000; Lucy et al., 2014). The effect of D. polymorpha on unionids also depends on the type of bottom and is much more severe on hard substrates, where unionids cannot bury and are largely exposed for mussels’ attachment (Arter, 1989; Burlakova et al., 2000).
Although fewer studies were conducted on the impact of D. r. bugensis, it looks like their impact on unionids is by far less severe than that of D. polymorpha (Burlakova et al., 2014). In the lower Great Lakes, the replacement of D. polymorpha by D. r. bugensis coincided with significantly lower unionid infestation rates. In the 1990s, an average of > 200 dreissenids/unionid was reported (Haag et al., 1993; Gillis & Mackie, 1994), whereas in 2011 – 2012 only one-third of all unionids collected in lakes Erie and Ontario were infested (median: 3 dreissenids/unionid), although half of the unionids without Dreissena had evidence of past infestations. At the same time (2011 – 2012), in waterbodies still predominantly colonized by D. polymorpha (several Michigan inland lakes, the Detroit River, and Lake St. Clair), unionid infestation was almost three times higher, and the mean mass of attached dreissenids an order of magnitude higher than in lakes Erie and Ontario, although all these waterbodies were colonized by dreissenids at approximately the same time (Burlakova et al., 2014).
The lower infestation by D. r. bugensis may be a result of their lower attachment strength (Claxton et al., 1998; Peyer et al., 2009, 2010; Collas et al., 2018) and/or their higher species-specific predation by fish compared to D. polymorpha (Bowers et al., 2005; Casper & Johnson, 2010), as their thinner shells are more easily crushed and digested (Protasov et al., 1994; Diggins et al., 2004). Finally, more complex interactions may also contribute to these different infestation patterns, including substrate preferences, and colonization and post-settlement survivorship (reviewed in Burlakova et al., 2014).
Some authors suggested that unionid mortality also depends on the unionid life history and ecological traits (like burrowing activity, substrate preference, feeding behavior, brooding period, growth rate) or shell morphology (Arter, 1989; Haag et al., 1993; Nalepa, 1994; Nichols & Wilcox, 1997a, b; Allen & Vaughn, 2009; Sousa et al., 2011). Studies conducted in lakes Erie, Ontario, and St. Clair, the Detroit River, and three inland lakes in Michigan did not show significant infestation differences based on unionid taxonomy, morphology, or life history (Burlakova et al., 2014), calling for further investigations.
Impacts on fishes
The overall impacts of Dreissena spp. on fishes vary depending on the feeding mode of the consumer, the morphology of the waterbody invaded, time since mussel invasion, co-evolutionary history, and Dreissena species and is different in Europe and North America (Karatayev et al., 1997, 2002, 2015a; Molloy et al., 1997; Strayer et al., 2004; Higgins & Vander Zanden, 2010). Impact on fishes maybe direct, through the consumption of mussels, or indirect, via Dreissena’s impact on various traits of the aquatic ecosystem.
Since North American fishes did not co-evolve with dreissenids, early in the invasion of the Great Lakes, these mussels were assumed to be a major loss of energy and potential production due to the redirection of food resources from the pelagic to the benthos (Johnson et al., 2005) and the decline in sphaeriids and especially in the preferred fish prey item, the amphipod Diporeia (Dermott, 2001; Lozano et al., 2001; Dermott & Geminiuc, 2003; Watkins et al., 2007; Nalepa et al., 2006, 2009a, b, 2014). Indeed, the dramatic decline of Diporeia in the Great Lakes caused major losses in the abundance of commercially important species, including the whitefish (Coregonus clupeaformis), alewife (Alosa pseudoharengus), sculpin (Cottus cognatus), bloater (Coregonus hoyi), and others that are prey for larger piscivores, including salmon and trout (Hoyle et al., 1999; Pothoven et al., 2001; Nalepa et al., 2009a; Karatayev & Burlakova, this issue). The shift in the diet of whitefish from the amphipod to quagga mussels resulted in the decline of lake whitefish condition, growth, and abundance (Pothoven et al., 2001; Hoyle et al., 2008; Nalepa et al., 2009b; Rennie et al., 2009). However, with time dreissenids became an important component of diet for many commercially important native fishes in the invaded North American freshwaters (details in Natural Enemies: Fish below).
Another Ponto–Caspian invader, the round goby (Neogobius melanostomus), recorded in the Great Lakes since 1990 (Jude et al., 1992; Charlebois et al., 1997), feeds actively on benthic invertebrates, including dreissenids (Barton et al., 2005; Bunnell et al., 2005; Johnson et al., 2005; Brush et al., 2012; Kornis et al., 2012; Naddafi & Rudstam, 2014; Barrett et al., 2017; Foley et al., 2017). In turn, the round goby is consumed by a number of North American fishes, including the lake trout (Salvelinus namaycush), burbot (Lota lota), yellow perch (Perca flavescens), smallmouth bass (Micropterus dolomieu), lake sturgeon (Acipenser fulvescens), cisco (Coregonus artedi), and walleye (Sander vitreus) (Dietrich et al., 2006; Madenjian et al., 2011; Weber et al., 2011; Crane & Einhouse, 2016; Jacobs et al., 2017; Pothoven et al., 2017; Breaker et al., 2020). Although all these fish species, as well as 9 others, are known to consume dreissenids in North America (reviewed in Molloy et al., 1997), predation on the round goby substantially increased the transfer of energy accumulated by dreissenids in benthic environments back to the pelagic and increased fish productivity, including several commercially important species (reviewed in Karatayev & Burlakova, this issue).
Despite the many studies on the impact of dreissenids on fishes, broad generalizations of the ensuing effects are still contentious. Some authors reported an enhancement of all benthivorous fishes, even those that do not feed on dreissenids, due to the increased biomass of mussel-related invertebrates (Karatayev, 1983; Lyakhnovich et al., 1983, 1988; Karatayev & Burlakova, 1992, 1995b; Strayer et al., 2004; Karatayev et al., 1997, 2002; Molloy et al., 1997). Others, however, underscore the negative effects, including the declines of important fish prey items, such as Diporeia and sphaeriids (Dermott & Kerec, 1997; Hoyle et al., 1999; Lozano et al., 2001; Pothoven et al., 2001; Nalepa et al., 2009a), particularly in the Great Lakes. In riverine systems, like the Hudson River, mixed effects were found: the introduction of D. polymorpha was associated with a 28% decline in the abundance of open-water species, whereas littoral species increased 97% (Strayer et al., 2004).
Planktivorous fishes can be negatively affected as a result of lower phytoplankton abundance and associated decreases in zooplankton, competition with benthic species, and from higher larval fish predation due to increased water transparency (Francis et al., 1996; Lozano et al., 2001). Fouling by dreissenid mussels may reduce habitat quality for fish spawning by reducing egg deposition and increasing damage to eggs (Marsden & Chotkowski, 2001). On the other hand, positive effects have been suggested that could mitigate the negative impacts of dreissenids on planktivorous fish. The larvae of both dreissenid species became an important component of plankton, a prey item for planktivorous fish (details in Natural Enemies: Fish below), as well as in fish fry diets, helping to offset declines in zooplankton (Nack et al., 2015; Marin Jarrin et al., 2015; Withers et al., 2015).
Economic impacts
The first reports of zebra mussel impacts on water supply systems were published in the nineteenth century (in Budapest, Hamburg, Berlin, and Paris; reviewed in Zhadin, 1946; Clarke, 1952). With the development of water-related infrastructure, their negative effects were noticed in power generation stations and in irrigation water supply systems (Limanova, 1964; reviewed in Karatayev et al., 1994b). The main impacts are the reduction in the bore of pipes and water flow drops, corrosion of steel and cast-iron pipes, accumulation of mussel shells at the pipe outlets, and water contamination when mussels are killed in control operations (Mackie & Claudi, 2010). Mussel fouling interferes with the operation of pumps, forbays, holding tanks, trashracks, and condenser units (Kovalak et al., 1990; Minchin et al., 2002). In two years, zebra mussels can overgrow the walls of raw water intake structures with up to 10 – 12 kg of mussels per m2 and increase gradually to up to 19 kg/m2 with live mussels settling on top of a thick layer of dead shells. Eventually, both live molluscs and empty shells are flushed downstream clogging sieves, filters, and heat exchangers (reviewed in Karatayev et al., 1994b). In Moscow’s drinking water supply system, zebra mussel beds reached densities over 19,000/m2 and a biomass of 15 kg/m2 (Sokolova, 1959). In irrigation pipes, the thickness of mussel fouling can reach 15 cm. However, in Europe fouling problems were most common during the early invasion period and currently they are most frequent in the newly colonized regions (e.g., Ireland, Minchin et al., 2002; Spain, Binimelis et al., 2007). In their native range or countries invaded decades ago, preventive measures are usually implemented, and problems arise if adequate maintenance operations are not undertaken (e.g., Turkey, Bobat et al., 2004).
Likewise, in North America, dreissenids also have significant impacts on raw water-dependent infrastructure, including power generation stations, drinking water treatment plants, industrial facilities, navigation locks, dam structures, and other facilities (Mackie & Claudi, 2010). During the last decades, a comprehensive “toolbox” of mussel control approaches was developed to eradicate, contain, and monitor dreissenid mussels in various facilities and to prevent their spread to uninfested waters (reviewed in Mackie & Claudi, 2010; Chakraborti et al., 2016).
In the early years of Dreissena’s arrival in the USA, their ecological and economic impacts were predicted to be catastrophic. The first estimates of damage and associated control actions were as high as $1 billion per year (Pimentel, 2005), $1.8 – $3.4 billion by the end of the twentieth century (Office of Technology Assessment, 1993), and $4 billion over the last decade of the twentieth century (Roberts, 1990). Although precise data are lacking, these “guesstimates” later proved highly exaggerated, largely because the methods applied were based on scarce, scattered, and often anecdotal information, and no solid, systematic empirical methods were used (Lovell et al., 2006). O’Neill (1997) estimated the economic costs of zebra mussels across a range of industries that maintain infrastructure susceptible to mussel fouling (339 facilities) as $69 million between 1989 and 1995. The largest expenditures were recorded in electric power generation plants (51%), drinking water treatment plants (31%), and other industries and agencies (8 and 7%). Other affected categories included proactive measures by the National Scenic Riverway, shipping and navigation locks, hatcheries and aquaculture, impoundments and reservoirs, recreational facilities, and marinas (O’Neill, 1997). The largest part ascribed to expenses by industrial facilities was spent on prevention actions, followed by the costs of chemical control compounds, planning, monitoring, and others. For water treatment plants, the costliest categories were chemical treatments, planning, and retrofitting, while costs of prevention were the largest for hydroelectric and nuclear power generating plants. Connelly et al. (2007) updated the estimates of the total economic costs for electric generation and water treatment facilities from 1989 to 2004 at $267 million (range: $161 –$467 million). They concluded that during the early years of zebra mussel infestation, annual costs were greater ($44,000/facility) than in recent years ($30,000), likely leading, together with other factors, to large overestimates in the early predictions. These costs did not include those associated with other impacts on the industry and navigation, natural resources (e.g., fisheries), or the impacts related to recreational boating and tourism.
The most recent assessment of the global economic costs of dreissenids between 1980 and 2020 (using the InvaCost database, Diagne et al., 2020) yielded $51.1 billion (2017 US$) (Haubrock et al., 2022). Unfortunately, this estimate suffers from many shortcomings, such as overlapping costs extracted from different sources (as acknowledged by the authors; Diagne et al., 2021), the fact that 98% of the data collated are from North America and “cost” categories include several contentious expenditures, such as research, administration, detection, surveillance, monitoring, education, communication and information, and risk assessment. Although estimates of costs alone are useful, they are a biased indicator of the overall negative economic impacts because they ignore the many benefits of dreissenids, including those on drinking water processing plants (Wang et al., 2021), fishing, and many others (see Uses; Burlakova et al., 2022b; Boltovskoy et al., 2022).
Natural enemies
Natural enemies of dreissenids include fishes, birds, and other animals which feed on the mussels, as well as parasites that can affect mollusc densities and their population dynamics.
Fish
At least 58 species of benthivorous fishes on both continents feed on adult dreissenids (Vorobiev, 1949; Zhadin, 1952; reviewed in Karatayev et al., 1994b, 2015b; Molloy et al., 1997; Baer et al., 2022; Burlakova et al., 2022b). In Europe, dreissenids are readily consumed by a wide range of fishes, as many of them are evolutionarily adapted to feed on mussels (reviewed in Karatayev et al., 1994b; Molloy et al., 1997), and the introduction of zebra mussels to new lakes is often associated with increases in fish productivity and commercial catches (Lyagina & Spanowskaya, 1963; Poddubny, 1966; Lyakhnovich et al., 1988; Karatayev & Burlakova, 1995b; reviewed in Karatayev et al., 1994b; Molloy et al., 1997). In the northern Caspian Sea, the native area of several dreissenid species (see Taxonomy), about 90% of the annual production of mussels (13,000 tonnes wet weight) are consumed by fishes (Yablonskaya, 1985). Dreissenids are also very important in the diets of fishes from the Azov Sea (Vorobiev, 1949). In Uchinskoe Reservoir (Russia), benthivorous fishes consume approximately 80% of the yearly production of zebra mussels under 15 mm (Lvova, 1977). The roach (Rutilus rutilus) and its subspecies Caspian roach or vobla (Rutilus rutilus caspicus) are the most prominent consumers of several dreissenid species in fresh and brackish waters (Prejs, 1976; Karatayev et al., 1994b, 1997; Molloy et al., 1997). In Europe, the introduction of the zebra mussel led to the establishment of new subpopulations of mussel-eating roach with much higher individual growth rates, larger body size, and higher lipid content, compared to the pre-dreissenid period (Lyagina & Spanowskaya, 1963; Poddubny, 1966). In addition to roach, dreissenids in Europe are actively consumed by bream (Abramis brama), silver bream (Abramis bjoerkna), European eel (Anguilla anguilla), Russian sturgeon (Acipenser gueldenstaedtii), sterlet (Acipenser ruthenus), and beluga (Huso huso) (reviewed in Molloy et al., 1997). Increases in the abundance of not only benthic feeding fishes, but also species that do not feed on the mussels but consume other benthic invertebrates whose abundance is enhanced by the presence of Dreissena spp. were recorded (Karatayev, 1983; Lyakhnovich et al., 1983, 1988; Karatayev & Burlakova, 1992, 1995b; Strayer et al., 2004; Karatayev et al., 1997, 2002; Molloy et al., 1997).
Dreissenids became an important component of diet for many commercially important native fishes in the invaded North American freshwaters. In addition to whitefish (Pothoven & Madenjian, 2008; Madenjian et al., 2010), round whitefish (Prosopium cylindraceum) (Turschak & Bootsma, 2015), yellow perch (Perca flavescens) (Morrison et al., 1997; Watzin et al., 2008; Shields & Beckman, 2015), blue catfish (Ictalurus furcatus), channel catfish (I. punctatus) (Thorp et al., 1998), and freshwater drum (Aplodinotus grunniens) were documented to feed on dreissenids. Dreissena spp. comprise a major part of the diet of the endangered silver chub (Macrhybopsis storeriana) (Kocovsky, 2019), and a species of conservation concern – the lake sturgeon (Jackson et al., 2002). Several species of Lepomis were reported to shift from a pre-invasion diet of benthic littoral invertebrates to zebra mussels (Molloy et al., 1997; Mercer et al., 1999; Magoulick & Lewis, 2002; Colborne et al., 2015). Another Ponto–Caspian invader, the round goby, feeds actively on benthic invertebrates, including dreissenids (see in Impacts on fishes).
Dreissenids were recorded in stomachs of other fish (burbot, Lota lota; ruffe, Gymnocephalus cernua; brown trout, Salmo trutta; northern pike, Esox lucius; deep-water Arctic charr, Salvelinus profundus; normal Arctic charr, S. umbla; and chub, Squalius cephalus), but their occurrence in the diet was low, in addition to being seasonal, and mussels were possibly consumed as incidental by catch (Millane et al., 2012; Baer et al., 2022). For example, in the digestive tracts of omnivorous fish (chub, charr, burbot, or ruffe) in Lake Constance, quagga mussels were found alongside larger quantities of gammarids, likely swallowed unintentionally during the pursuit of amphipods in dreissenid aggregations (Baer et al., 2022).
The larvae of both dreissenid species became an important component of plankton and in lakes often dominate the zooplankton during the summer, comprising 18 to 73% of the total zooplankton density and from 6 to 40% of the zooplankton biomass and production (Karatayev, 1983; Mitrakhovich & Karatayev, 1986; Lvova et al., 1994b; David et al., 2009; Karatayev et al., 2010; Withers et al., 2015; Lazareva et al., 2016; Bowen et al., 2018). These larvae represent an important food resource for at least 17 species of fish in Europe and North America (Wiktor, 1958; Kornobis, 1977; Molloy et al., 1997; Withers et al., 2015). Although the importance of Dreissena spp. larvae as a prey item for fish varies widely (Wiktor, 1958; Kornobis, 1977; Limburg & Arend, 1994; Limburg et al., 1997; Mills et al., 1995; Chrisafi et al., 2007; Watzin et al., 2008; Nack et al., 2015; Turschak & Bootsma, 2015; Withers et al., 2015), under certain conditions they may become an important component of fish fry diets and may help offset decreases in the overall abundance of zooplankton (Nack et al., 2015; Marin Jarrin et al., 2015; Withers et al., 2015).
A major concern associated with mussel-consuming fishes and other animals is whether this trophic pathway facilitates the transfer of bioaccumulated contaminants up the food web by providing a novel route for the less accessible sediment-deposited toxicants (Bruner et al., 1994; Roper et al., 1996; Johns & Timmerman, 1998; Robertson & Lauenstein, 1998; Kwon et al., 2006; Schummer et al., 2010; Mezek et al., 2011; Wallace & Blersch, 2015). Dreissenids bioaccumulate many toxicants (reviewed in Binelli et al., 2015, Burlakova et al., 2022b), but bioaccumulation and trophic transfer of contaminants vary among species (Matthews et al., 2015; Evariste et al., 2018) and pollutant types (Perez-Fuentetaja et al., 2015), and we do not know whether invasive mussels are more hazardous in this respect than other prey, all of which bioaccumulate pollutants. Some contaminants were reported to bioaccumulate very significantly in the mussels’ tissue or shells (reviewed in Binelli et al., 2015), whereas bioaccumulation of others (e.g., PBDEs—polybrominated diphenyl ethers) have been found to be lower than for zooplankton and amphipods (Perez-Fuentetaja et al., 2015), which may decrease the levels of these compounds in mussel-feeding fishes (Hahm et al., 2009). Thus, further research is needed to assess whether dreissenids are effectively more harmful in transferring contaminants than other native or introduced prey species.
Waterfowl
Birds are among the best documented natural enemies of Dreissena spp. At least 36 species of waterfowl have been recorded to feed on Dreissena spp., including 21 species in Europe and 20 in North America. The most common and best researched consumers are the tufted duck (Aythya fuligula), pochard (A. ferina), greater scaup (A. marila), lesser scaup (A. affinis), goldeneye (Bucephala clangula), and coot (Fulica atra) (reviewed in Molloy et al., 1997). Migrating waterfowl can quickly locate areas with dense mussel populations (reviewed in Molloy et al., 1997) and forage on mussels between autumn and spring during their migrations (Hamilton et al., 1994; Mitchell & Carlson, 1993) or overwintering on site (bij de Vaate, 1991; Cleven & Frenzel, 1993). Birds are often the most aggressive predators on dreissenids, consuming up to 30% of the annual zebra mussel production in shallow areas (Smit et al., 1993) and up to 70 – 90% of their biomass (Mikulski et al., 1975; Stempniewicz, 1974; Werner et al., 2005). Their impact on dreissenid populations depends on several factors, including depth, substrate, and Dreissena species (Molloy et al., 1997; Werner et al., 2005). For example, in Lake Constance (Germany), while mussel biomass reduction is strongest in shallow areas, in deeper areas the impact is highly variable, depending on the substrate, and almost no impact occurs at 11 m (Werner et al., 2005). Therefore, waterfowl predation is likely to be more intense in shallow polymictic lakes than in deep dimictic lakes. Since zebra mussels are largely limited to littoral zone, they are probably more vulnerable to waterfowl predation than quagga mussels that are usually more abundant in the profundal than in the littoral zones.
Large increases in waterfowl flocks feeding on dreissenids were recorded both in Europe and in North America. Since zebra mussels invaded Lake Constance in the 1960s, the number of overwintering waterbirds increased fourfold (Werner et al., 2005). After dreissenids colonized Long Point Bay on Lake Erie, one of the most important waterfowl staging areas on the Great Lakes (Petrie & Knapton, 1999), a noticeable increase in waterfowl was recorded, including a 92-fold increase in scaups (Aythya spp.). The number of waterfowl including scaups, canvasback (Aythya valisineria), and redhead ducks (Aythya americana) using Lake St. Clair (USA-Canada) in USA waters during their fall migrations increased from 1.1 million use days before dreissenids were present to 2.1 million after it (Schummer et al., 2010). While scaups prey on dreissenids directly, other birds, such as canvasbacks, likely responded to increased submerged aquatic macrophyte food associated with the enhanced water clarity due to mussel colonization (Luukkonen et al., 2014). In the brackish lagoons of the Odra River Estuary (south-western Baltic Sea), an important area for greater scaup (A. marila) during the non-breeding season, the birds consume an average of 5,400 tonnes of zebra mussels annually (Marchowski et al., 2015). The declining European population of A. marila thus depends on the non-native zebra mussels that constitute > 90% of their food (in terms of biomass).
The introduction of zebra mussels was associated not only with dramatic increases in waterfowl numbers but also affected their migration patterns (reviewed in Molloy et al., 1997). Prior to the introduction of zebra mussels into Swiss lakes, waterfowl fed on aquatic macrophytes and migrated to the south in the fall after plant die back. In contrast, after the invasion by Dreissena, large numbers of birds overwinter locally (Leuzinger & Schuster, 1970). Ten to 50-fold increases in overwintering tufted ducks, pochards (Aythya spp.), and coots (Fulica spp.) were observed soon after zebra mussels colonized western Lake Constance in the late 1960s and goldeneye began to arrive in the area earlier (Suter, 1982b). Conversely, in areas where zebra mussel populations declined, diving birds showed a tendency to leave overwintering areas earlier, likely due to lower food availability (Suter, 1982a). Ice formation precludes foraging in winter, but in waterbodies where open waters are available year-round as in cooling water reservoirs for power plants, large flocks of mallards (Anas platyrhynchos) regularly overwinter and consume large quantities of zebra mussels in shallow, open waters (Karatayev et al., 1994b; Kozulin, 1995). Food availability, particularly Dreissena, were likely the major factor for the shifts of the geographical range of the tufted duck (Aythya fuligula) in England (Olney, 1963) and for overwintering diving ducks in Switzerland (Pedroli, 1981; Suter, 1994).
In addition to the direct consumption of dreissenids, waterfowl also prey on the invertebrates associated with the mussels, as well as on macrophytes and bottom algae that benefit from dreissenid-enhanced water clarity. Thus, the abundance of macroinvertebrates associated with Dreissena colonies (mostly Oligochaeta, Chironomidae, and Ephemeroptera) were significantly reduced in unprotected enclosures in shallow areas of Lake Constance, presumably due to waterfowl predation (Mörtl et al., 2010). Due to the recolonization of the lake by stoneworts (Characeae) after the introduction of zebra mussels in the 1980s, the bay of Lucerne (Switzerland) has become an internationally important wintering site for the red-crested pochard (Netta rufinadue) (Schwab et al., 2001). In some lakes, both Chara and zebra mussels are now considered keystone species which control ecosystem resilience, and careful management of these species has been suggested as important as the control of nutrients (Ibelings et al., 2007).
Parasites and commensals
A detailed review of dreissenid parasites and commensals was produced by Molloy et al. (1997). In brief, over 45 taxa are known to date to be associated with the mantle cavity and/or visceral mass of D. polymorpha, including protozoans, haplosporidians, nematodes, trematodes, oligochaetes, leaches, mites, and chironomids (Molloy et al., 1997, 2012; Molloy, 2002; Karatayev et al., 2000a; Mastitsky, 2004; Mastitsky & Gagarin, 2004; Mastitsky & Samoilenko, 2005). All parasites and commensals of D. polymorpha were reported from attached mussels (i.e., none from their planktonic larvae).
Five species of ciliates including Ancistrumina limnica, Conchophthirus acuminatus, Hypocomagalma dreissenae, Sphenophrya dreissenae, and Sphenophrya naumiana are known from the mantle cavity and at least one species (Ophryoglena hemophaga) from the digestive gland (Molloy et al., 1997, 2005). With the exception of A. limnica (a non-specific endosymbiont of freshwater lamellibranchs and gastropods), all ciliates are highly specific to Dreissena spp. that were found in Europe only. Among the ciliates, the most common endosymbiont is C. acuminatus, which was found in almost all European populations of D. polymorpha with an infection prevalence often reaching 100% and infection densities in excess of hundreds of ciliates per mussel (Molloy et al., 1997; Karatayev et al., 2000a, 2007c; Mastitsky, 2004). Because C. acuminatus has not been found in North America, Karatayev et al. (2000a, 2007c) hypothesized that (1) larval (rather than adult) individuals of Dreissena invaded North America and (2) the European waterbodies invaded by Dreissena where C. acuminatus-infestations are present were either colonized by adult mussels, or, alternatively, if the waterbody was colonized by planktonic larvae, it had an upstream source of veligers and C. acuminatus. In addition to D. polymorpha, C. acuminatus was also reported from D. r. bugensis (Karatayev et al., 2000b) and from D. stankovici from Lake Ohrid, Republic of Macedonia (Molloy et al., 2010). In contrast to C. acuminatus, the other ciliates reported from Dreissena spp. (H. dreissenae, S. dreissenae, S. naumiana, and O. hemophaga) are parasites, but they do not seem to cause significant host mortality (Molloy et al., 1997, 2010).
A lethal infection with haplosporidians has been reported from D. polymorpha populations in The Netherlands (reviewed in Molloy, 2002). An extensive study conducted on 43 waterbodies across 11 European countries showed that haplosporidian (described as H. raabei) infections in zebra mussels are extremely rare, as only 0.7% of all mussels were infected (Molloy et al., 2012). In contrast to the low prevalence, the disease intensity was quite high, with 79.5% of the infections showing signs of sporogenesis. Infection with multinucleate plasmodia and sporocysts were observed systemically in connective tissues, including gills, gonads, and digestive gland. However, no major signs of disease were evident upon dissection (Molloy et al., 2012).
Seven genera of trematodes have been reported as parasites of Dreissena spp. (Molloy et al., 1997). In their life cycles, dreissenids can serve as the first intermediate host (e.g., for Bucephalus polymorphus and Phyllodistomum spp.), second intermediate host (e.g., for Echinoparyphium recurvatum), or the only host (e.g., for Aspidogaster spp.) (Molloy et al., 1997; Molloy, 2002). Bucephalus polymorphus requires three hosts to complete its life cycle. Infection commences in dreissenids at the earliest parasite larval stage, the miracidium, which hatches from an egg, enters the mussel’s visceral mass, and gives rise to the sporocyst stage. Cercariae released into the surrounding water attach to fish fry, encyst in their tissues, and transform into metacercaria. The final hosts of B. polymorphus are predatory fish that have consumed fish infected with metacercariae. The prevalence of B. polymorphus infection varied from 0.4% to 28% (reviewed in Molloy et al., 1997; Karatayev et al., 2000a). It primarily affects the gonads, typically inducing sterility (Molloy et al., 1997; Laruelle et al., 2002). Although it was suggested that B. polymorphus is species specific to Dreissena (Molloy et al., 1997), additional studies are required to confirm this assumption.
Other organisms that were occasionally reported from the mantle cavity of dreissenids include nematodes, oligochaetes, leaches, mites, and chironomids. They are not obligate Dreissena symbionts and have little or no adverse impacts on their host (Molloy et al., 1997; Laruelle et al., 2002). Although data at hand suggest that D. r. bugensis and D. stankovici are less susceptible to infections by parasites and commensals than D. polymorpha (Karatayev et al., 2000b; Molloy et al., 2010), this assumption may reflect the fact that research on these species has been very uneven.
Uses: ecosystem services and disservices
While many negative biological invasion-driven effects of dreissenids on the structure and functioning of ecosystems have been identified, much less has been done on evaluating and/or monetizing their ecosystem services (Diagne et al., 2021). Further, many of the services are difficult to monetize and are regularly ignored (Pejchar & Mooney, 2009; Thompson, 2014; Boltovskoy et al., 2022). In this section we focus on the positive effects of these mussels and review their ecosystem services based on the assumption that if a particular effect of a native species is considered as a service (e.g., water clarity increases by native unionid mussels; Vaughn, 2017; Vaughn & Hoellein, 2018), the same effect is also a service when provided by exotic bivalves.
The three major recognized categories of services include regulating and maintenance services (e.g., water purification, climate regulation, pollination), provisioning services (e.g., provision of food, water, fuel), and cultural services (e.g., education, recreation, tourism, esthetic and spiritual values) (Millennium Ecosystem Assessment, 2005; Haines-Young & Potschin, 2011). A comprehensive review of dreissenid ecosystem services is provided in Burlakova et al. (2022b).
Of 2,853 publications on dreissenids in SCOPUS (see Research Efforts), the largest number was focused on ecosystem services (34%), while impact and management categories both yielded 9% of publications each. This signifies the recognition of positive effects of dreissenids and their wide usage as indicator species.
Regulating and supporting/maintenance services
Biofiltration
Water purification is perhaps the most important and one of the best documented services provided by dreissenids (see Impacts and Effects: System-Wide Impacts). Mussel filtration has dramatic impacts on lakes and rivers yielding particulate suspended matter reductions of up to over 60%, Secchi disc depth increases up to 200% and 50–250% increases in light penetration (reviewed in Karatayev et al., 1997, 2002; Higgins & Vander Zanden, 2010; Mayer et al., 2014). Improvements in lake water clarity can increase property prices (reviewed in Jakus et al., 2013), and the services conveyed by the mussels’ filtration have been assessed in economic terms based on the changes in waterfront property values in association with changes in the water clarity of the waterbodies involved (Limburg et al., 2010; see Cultural Services). In addition, economic benefits of mussel filtration can significantly alleviate the costs of water potabilization (Wang et al., 2021).
Nutrient cycling and storage
Dreissenids can alter the stoichiometry and the rates of nutrient recycling, as well as the spatial distribution of nutrients (Arnott & Vanni, 1996; Mellina et al., 1995; Naddafi et al., 2009; Katsev, 2017; Li et al., 2018). Filter-feeding dreissenids remove particulate phosphorus (P) from water at P deposition rates tenfold greater than passive P settling rates, which shortens P residence times in the water column and increases the rates of P transfer to the benthos (Mosley & Bootsma, 2015). This process induces long-term declines in total P concentrations in the water column of lakes colonized by dreissenids (Higgins, 2014; Mayer et al., 2014; Dove & Chapra, 2015). The Laurentian Great Lakes are a dramatic example of large-scale reorganization of biogeochemical cycles due to the impacts of quagga mussels, whose tissues and shells now contain nearly as much phosphorus as the entire water column (Li et al., 2021). The effects of these invasive mussels on nitrogen (N) compounds are complex and highly context dependent (Higgins & Vander Zanden, 2010). Quagga mussels living in deep areas of the Great Lakes are hotspots for N cycling, including denitrification. These processes have not yet been adequately explored, but they are likely responsible for very substantial pathways of N fluxes in many freshwater habitats (Katona et al., in review). In addition to their effect on P and N which are of particular importance for primary producers, Dreissena plays a major role in the recycling and sequestration of many other elements as well, including As, C, Ca, Cu, Fe, Ni, Ti, Mg, Si, V, and Al (Walz, 1978d; Karatayev et al., 1994a; Wojtal-Frankiewicz & Frankiewicz, 2010; Schaller & Planer-Friedrich, 2017).
Reduction of phytoplanktonic primary production
Another important service provided by dreissenids is the reduction of phytoplanktonic primary production (see Impacts and Effects: System-Wide Impacts). The use of zebra mussels for culling the effects of eutrophication was the object of many studies, particularly in The Netherlands, where many shallow freshwater lakes suffer from severe algal blooms (Reeders & bij de Vaate, 1990; Noordhuis et al., 1992; Smit et al., 1993; Waajen et al., 2016) (see below in Bioremediation).
Habitat creation/modification
By creating complex reef-like structures, dreissenids provide new habitats that are used by many invertebrates as refuge from predation and abiotic stress, and organic matter-rich feces and pseudofeces, together with increased organic matter and carbon fluxes to the benthos, enhance the food subsidy for benthic deposit feeders (Karatayev & Burlakova, 1992, 1995b; Karatayev et al., 1994a, 2002, 2007a, b; Botts & Patterson, 1996; Stewart et al., 1998; Burlakova et al., 2005, 2012; Gergs et al., 2009; Ozersky et al., 2015). Algal and bacterial communities that grow on the shells and in their aggregations represent another source of food for benthic invertebrates inhabiting the mussels’ beds (Lohner et al., 2007; Makarevich et al., 2008; Higgins & Vander Zanden, 2010). These habitat modifications usually result in significant increases in the abundance of many native invertebrates (see Ecological Impacts: Local Impacts). The increased water clarity caused by the filtration of invasive bivalves promotes the growth of submerged aquatic vegetation (also ecosystem engineers), further affecting water flow and providing resources, such as food and habitat for zooplankton, benthic invertebrates, birds, and larval and adult fishes (see Natural Enemies; Lyakhnovich et al., 1988; Reeders & bij de Vaate, 1990; Skubinna et al., 1995; Lowe & Pillsbury, 1995; MacIsaac, 1996; Strayer et al., 1999; Mayer et al., 2001; Luukkonen et al., 2014), as well as many other ecosystem services (Thomaz, 2021).
Wastewater treatment
Dreissenids can filter and clean water from organic pollution and toxic substances, including heavy metals (Selegean & Heidtke, 1994; Elliot et al., 2008; Binelli et al., 2014; Gomes et al., 2018). In 96-h trials, zebra mussels effectively removed and biodeposited nearly all seston and P from diluted activated sewage sludge, significantly decreasing the biochemical oxygen demand of the sludge and improving its clarity (Mackie & Wright, 1994). In Germany, biofilters consisting of zebra mussel-overgrown artificial modules were successfully tested for wastewater treatment (Kusserov et al., 2010). Mussels can also eliminate pathogenic organisms (e.g., Escherichia coli, enteric viruses, Toxoplasma gondii, Giardia duodenalis) from wastewaters (Mezzanotte et al., 2016; Géba et al., 2020).
Mussel shells can be used as a P-binding agent for the removal of P from wastewater effluents: in test trials, Dreissena shell fragments had the highest phosphorus adsorption capacity compared to other media (Van Weelden & Anderson, 2003). Zebra mussel shells can remove over 99% of the phosphorus from wastewater, showing a potential as an alternative source to mined calcium carbonate for the production of lime (McCorquodale-Bauer & Cicek, 2020).
Bioremediation
Zebra mussels have been used to mitigate the effects of eutrophication since the 1980s (Piesik, 1983). The first successful field experiments were carried out in The Netherlands in the early 1990s (Reeders & bij de Vaate, 1990; Noordhuis et al., 1992; Reeders et al., 1993; Smit et al., 1993; Waajen et al., 2016). The treatment pond with zebra mussels experienced a steady increase in Secchi disc depths, with a ~ 1-m difference (with respect to the control) remaining stable throughout the first year of the experiment. Further, both organic and inorganic suspended matter declined, light penetration increased, and no cyanobacterial blooms were observed (Noordhuis et al., 1992). Quagga mussels were also shown to reduce the phytoplankton (including Cyanobacteria) in a hypertrophic urban pond and induce a clear water state (Waajen et al., 2016).
Dreissenid mussel farming can be a potentially profitable industry in coastal waters for tackling eutrophication and facilitating macrophyte restoration (Stybel et al., 2009; Schernewski et al., 2012; Friedland et al., 2019). A single 0.5-ha mussel farm could compensate for the total annual input of phosphorus from 23 ha of the watershed or the biologically available P from 49 ha of agricultural land (Goedkoop et al., 2021). In the Baltic Sea, artificial reefs and substrates were used for the cultivation of the alga Furcellaria and the bivalves Mytilus and Dreissena to restore the spawning grounds of the Baltic herring (Korolevs & Kondratjeva, 2006).
Environmental monitoring
Zebra mussels have been widely used since the late 1970s as sentinel organisms to monitor chemical pollution in aquatic environments because they are filter feeding sessile bottom dwellers that bioaccumulate many contaminants with little metabolic transformation and provide time-integrated information of chemical contamination in the environment (reviewed in Binelli et al., 2015). A large number of studies on application of dreissenids in environmental monitoring (9.5% of the total 2,853 publications on dreissenids in SCOPUS) signifies their wide usage as indicator species, and D. polymorpha was suggested as promising freshwater counterpart of widely used in marine ecotoxicological studies sentinel organisms Mytilus spp. (Binelli et al., 2015). Because they accumulate large amounts of a wide range of pollutants in their soft tissues and shells, dreissenids are suitable for the biomonitoring of heavy metals (Cd, Co, Cr, Cu, Hg, Ni, Pb, Zn, Hg, Se), organic compounds such as methylmercury, dichlorodiphenyltrichloroethane, and several related pesticides (DDTs), polychlorinated biphenyls (PCBs), polycyclic aromatic hydrocarbons (PAHs), hexachlorobenzene (HCB), hexachlorocyclohexanes (HCHs), organophosphate insecticides, and even radioactive contamination (Neumann & Jenner, 1992; reviewed in Binelli et al., 2015). Bioaccumulated pesticides and metals in transplanted zebra mussels can give an insight not only into their bioavailability in the environment but also into the ecological responses of the benthic communities affected by these toxicants (Bashnin et al., 2019).
Dreissenids were selected as target organisms by the National Status and Trends Mussel Watch Project (National Oceanic and Atmospheric Administration—NOAA, USA) which examines contaminants at nearshore sites on a biennial basis, providing crucial information for the identification of the levels and distribution of toxicants (Apeti & Lauenstein, 2006). Invasive mussels are also intensively used as both in vivo and in vitro biomarkers and in transcriptomics and proteomics studies to provide information on the potential impact of pollutants on the health of other organisms (Binelli et al., 2015).
Dreissenid behavior, in particular the closing of valves in response to toxic substances, can be monitored automatically in ad hoc flow-through devices in water treatment plants, triggering an alarm when the proportion of closed shells are above predefined threshold values. These “Dreissena-Monitors” were used by German potabilization plants, providing an efficient early warning system for the intake of polluted water (Borcherding, 2006).
In addition to chemical pollutants, dreissenids are used to monitor aquatic pathogenic organisms, including viruses, such as the ones responsible for the COVID-19 pandemic (Le Guernic et al., 2022). The use of zebra mussels as indicators of contamination by fecal coliforms and Escherichia coli provide advantages when compared with traditional monitoring methods (Selegean et al., 2001; reviewed in Gomes et al., 2018). Zebra mussels were found to host > 100 times more E. coli and other intestinal enterococci than ambient water after two days of pulse exposures to the bacteria, thus providing a time integrating and much more sensitive indicator of bacterial contamination than water samples (Bighiu et al., 2019).
Monitoring of pathogenic protists (Cryptosporidium parvum, Giardia duodenalis, Giardia lamblia, Toxoplasma gondii, Cyclospora sp., Enterocytozoon intestinalis, E. hellem, E. bieneusi) in the water requires filtration of large volumes because their densities are often low. In the process of filtration, mussels concentrate pathogenic protists, often proportionally to their abundance in the water, allowing usage of the bivalve’s tissues as a good integrative matrix for biomonitoring (Lucy et al., 2008, 2010; Lucy, 2009; Conn et al., 2014; Ladeiro et al., 2014; Gomes et al., 2018; Géba et al., 2020). Due to the digestion of C. parvum and T. gondii oocysts, mussels can also be used as a bioremediation tool (Géba et al., 2021a, b). Zebra mussels were found useful in assessing viral contamination due to their accumulation of viral pollution F-specific RNA bacteriophages (Capizzi-Banas et al., 2021) and a low pathogenic form of the avian influenza virus H5N1 (Stumpf et al., 2010; reviewed in Gomes et al., 2018). A significant reduction in gastroenteritis-causing rotavirus in spiked treated municipal wastewater was found in treatments with Dreissena, and although the viruses remained in the soft zebra mussel tissues or in the liquid phase, they were not transmissible to other species (Mezzanotte et al., 2016). The ability to bioaccumulate microcystins by zebra mussels can potentially be used for biomonitoring of toxic cyanobacterial blooms (Paldavičienė et al., 2015).
Hypoxia
Oxygen deprivation is currently a key global stressor in freshwater, estuarine, and marine benthic ecosystems (Diaz & Rosenberg, 2008; Tellier et al., 2022). Both zebra and quagga mussels are intolerant of even moderate hypoxia, and, in contrast to motile pelagic organisms and some benthic species, dreissenids cannot migrate to escape hypoxia (see Habitat Requirements). Their extremely high fecundity and their planktonic larvae, however, allow them to recolonize substrates even after large-scale die-offs and therefore, monitoring the occurrence and length-frequency distribution of Dreissena spp. can be an effective tool for mapping the extent and frequency of hypoxia in freshwaters (Karatayev et al., 2018b, 2021b). Due to their large body size and high densities, in clear waters dreissenids can be surveyed using remote sensing techniques (e.g., underwater video) allowing for swift collection of information on their distribution over large areas (Karatayev et al., 2018a, 2021c; Burlakova et al., 2022a) and providing a record of the recent history, rather than instantaneous snapshots, of hypoxic events.
Provisioning services
Food for fishes, birds, and other animals
Both dreissenid species provide an abundant food resource for fishes, birds (see Natural Enemies), and other animals. Several species of crayfish (Cambarus affinis, C. robustus, Orconectes limosus, O. virilis, O. propinquus) and muskrats (Ondatra zibethicus) have been observed eating zebra mussels in Europe and North America (reviewed in Karatayev et al., 1994b; Molloy et al., 1997). Adult stages of the crayfish O. limosus can consume 6,000 young dreissenids per crayfish during the summer, potentially limiting their population growth (Piesik, 1974). Zebra mussel populations crashed in 1992 in the Hudson River near Catskill (New York, USA) due to predation by blue crabs (Callinectes sapidus) (Molloy et al., 1994). A declining Laurentian Great Lakes native species, the mudpuppy salamander (Necturus maculosus), was recorded feeding on Dreissena (Beattie et al., 2017). Mongooses (Mungos mungo) and the oriental small-clawed otters (Aonyx cinerea) were feeding on zebra mussels in trials in the ZooOsnabrück (Germany), and raccoons (Procyon lotor) prefer zebra mussels over other food items (Schernewski et al., 2019). The northern map turtle (Graptemys geographica) and the stinkpot turtle Sternotherus odoratus were found to prey heavily on invasive mussels in the Laurentian Great Lakes (Lindeman, 2006; Patterson & Lindeman, 2009), and zebra mussels constitute up to 36% of G. geographica diet in Lake Opinicon (Canada) where it can consume over 3000 kg of mussels per year (Bulté & Blouin-Demers, 2008). In trials involving multi-prey assemblages, two native mysid species from the St. Lawrence River middle estuary, Neomysis americana and Mysis stenolepis, exhibited high predation rates on zebra mussel veligers (Winkler et al., 2007).
Food for farm animals
In terms of fresh weight, zebra mussels contain 8.4% of protein, 0.8% of fat, 0.14% of P, 1.3% of raw ash, and 89.3% of water (Schernewski et al., 2019), which makes them a potential substitute or additive for protein-rich fodder for farm animals. Reviews of the Russian literature on the use of dreissenids as food for livestock and poultry in dry food blends and to extract vitamins have been published as early as the mid-1900s (Zhadin, 1952; reviewed in Karatayev et al., 1994b). The mussel biomass that can potentially be harvested in some European waterbodies has been estimated at 5,000 t of dreissenid tissue, containing ~ 500 t of protein (Miroshnichenko, 1990). From artificial ad hoc substrates, up to 500 kg per hectare and 20–30 kg of molluscs per square meter can be obtained annually (reviewed in Karatayev et al., 1994b). Zebra mussels were found to be a palatable food supplement for chickens (McLaughlan et al., 2014). Harvested and grinded mussels (raw or boiled) were used as a food supplement for several fish species, including carp, sterlet, sturgeon, bester, and salmon (Karatayev et al., 1994b).
Materials
Mussel shells are composed primarily (> 80%) of calcium carbonate (Immel et al., 2016) and thus can be used as an alternative source of calcium carbonate (McCorquodale-Bauer & Cicek, 2020). Zebra mussel shells washed ashore were used as a food supplement for chicken, as well as for ducks and pigs (reviewed in Karatayev et al., 1994b). In agriculture, dreissenids can be also used as a fertilizer, soil amendment, or mulch (Mackie & Claudi, 2010; McDonnell, 1996).
Cultural services
Esthetics, leisure, and property values
The effects of invasive species on cultural services (i.e., recreation, tourism, history, education, science, heritage, inspiration, spirituality, and esthetics) are difficult to assess because they are based on personal and local value systems (Pejchar & Mooney, 2009; Cassini, 2020).
Dreissenids significantly enhance water transparency, and improvements in water clarity increase its esthetical perception, reduce some negative impacts such as odor and toxic cyanobacterial blooms (but see Ecological Impacts: System-Wide Impacts), and facilitate most water-related recreational activities. In the North American Great Lakes, divers enjoy the increased visibility after dreissenid introduction, and the scuba diving industry has boomed around the lakes as a result (Williams, 2020). By clarifying the water, dreissenids can increase the property value of neighboring real estate, as the market value of lakefront properties is tightly associated with the lake’s water quality, which in turn is chiefly perceived through the lake’s water clarity (Jakus et al., 2013). A one-meter improvement in lake water clarity in USA lakes was estimated to increase property prices from $11 to $382 per foot of water frontage and an overall property increase of 0.9 to 6.6% (reviewed in Jakus et al., 2013). Several studies analyzed the relationships between property values and water clarity based on surveys of owners of waterfront property. In the vacation region of North Central Wisconsin (the USA), increased lake water clarity associated with the presence of dreissenids was suggested to increase property values by about 10% (Johnson & Meder, 2013). Limburg et al. (2010) surveyed business owners and homeowners in the State of New York (the USA) to calculate the effects of dreissenids on property values due to both increased water quality and abundance of nuisance filamentous algae and estimated that the net benefit would be in the multiple millions of dollars, indicating that mussels produced mixed, but dominantly beneficial, impacts.
Information and knowledge
The value of exotic mussels as a tool in the areas of intellectual and experiential (scientific and educational) services has received limited recognition, but our SCOPUS analysis (see above) revealed that publications describing the use of mussels as biomonitors and bioindicatiors comprised almost a third of the publications in the Ecosystem function and services category. Dreissenid byssus production and adhesion are investigated with the aim of developing new materials to prevent biofouling for underwater adhesive bonds, anticorrosives, metal-sequestering reagents, and novel biomimetic polymers, and mussel shells have been used as a proxy of paleoclimatological and palaeolimnological variations (reviewed in Burlakova et al., 2022b). The accumulated knowledge on the biology, ecology, and distribution of these exotic mussels, as well as the animals themselves, which are easily obtainable for experimentation and teaching purposes, are used widely in K-12 and higher education (Burlakova et al., 2022b).
Management options
Prevention
Preventing the introduction and spread of dreissenids is the first line of defense against their invasions. It is also the most efficient and, in most cases, cost-effective method. Selecting the best management options is extremely important to efficiently allocate limited resources for preventing their spread and largely depends on the main vectors of dispersal. For example, the introduction of both dreissenids into North America, along with many other freshwater invaders in the twentieth century, occurred with ballast water. In the last decades, strategies have been adopted at the national and international levels in order to minimize this introduction pathway for aquatic organisms (reviewed in Bailey et al., 2013; Sturtevant et al., 2019). Secondary range expansions rely on different vectors, including recreational watercraft, coastal navigation, commercial and recreational fishing, diving, and aquarium trade (Carlton, 1993) and therefore, control measures are more heterogeneous and more difficult to enforce. Guidelines are available for the inspection and cleaning of freshwater-related equipment before it is moved to a different waterbody in order to prevent the spread of these and other introduced species (e.g., Morse, 2009; Comeau et al., 2011; DiVittorio et al., 2012; https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev3_014876.pdf; http://dbw.parks.ca.gov/pages/28702/files/Boating-QuaggaGuide.pdf), but compliance is likely very variable, and judging from the timeline of the mussels’ spread (Figs. 4, 5) is probably low.
Early detection and eradication
For the early detection of dreissenids, monitoring programs should be established in waterbodies free of the mussels but likely to be colonized (Marsden, 1991; Mackie & Claudi, 2010). The early detection of dreissenids is critically important for their eradication (if practically feasible, see below), as well as for anticipating potential problems for the industrial and potabilization plants involved. Currently, there are no ecologically sound, species-specific procedures for the eradication of dreissenids from large waterbodies. In some cases, D. polymorpha disappeared from entire lakes or rivers, but this was usually due to heavy pollution (bij de Vaate et al., 1992; Jantz & Neumann, 1992; Karatayev et al., 2003). Purposeful eradication using chemical control and dewatering (Fernald & Watson, 2014; Hammond & Ferris, 2019) or manual removal (Wimbush et al., 2009) have been achieved in a few cases, but usually at the expense of large impacts on most non-target aquatic organisms or in waterbodies with marginal chemical properties for dreissenid survival (e.g., Lake George, NY; Wimbush et al., 2009). Further, in some cases the intensive use of toxicants (a copper-based algaecide/bactericide in Christmas Lake, Minnesota) proved unsuccessful for preventing the establishment of zebra mussels (Lund et al., 2018). Thus, the applicability of these methods is very limited, with the exception of small impoundments, like fishponds, quarries, or very small lakes (Fernald & Watson, 2014; Hammond & Ferris, 2019).
Monitoring usually involves sampling for Dreissena larvae in the water column during the summer and searching for adult mussels on the bottom, especially on hard substrates, including artificial constructions, rocks, submerged wood, and unionid shells, as well as the deployment of ad hoc artificial settling devices (Marsden, 1991). More recently, environmental DNA analyses have become a standard and very effective tool for the early detection of dreissenids. This method allows for distinguishing the two dreissenid species at both their larval and their adult stages, even early in the invasion when their densities are very low (Gingera et al., 2017; Ardura et al., 2017; Amberg et al., 2019).
After establishment, continuous monitoring is important to understand their population dynamics and especially their reproduction peaks, which determine the timing of mass settlements on water intake facilities and, therefore, the scheduling of biofouling control operations (reviewed in Mackie & Claudi, 2010). Because dreissenids have major effects on many aspects of ecosystem function and services, routine mussel monitoring programs should include tracking of mussel biomass and distributions and changes in mussel demographics, particularly size distributions and changes in abundance of veliger larvae and juvenile mussels (Pergl et al., 2020; Li et al., 2021).
Control and containment
Because the eradication of dreissenids from an entire waterbody is largely unfeasible, it has been long recognized that their control should be focused on the raw water components of human-made structures, where their negative impacts on human interests and the economy are highest, and efficient control methods are available (Zhadin, 1946). The problem of dreissenid fouling in a wide range of industrial and municipal water supply systems dates back to the nineteenth century and led to the development of a large array of control methods. These methods include mechanical, physical, chemical, and biological control, as well as a combination of the above (reviewed in Claudi & Mackie, 1994; Karatayev et al., 1994b; Mackie & Claudi, 2010; Passamaneck, 2018). Mechanical methods include scraping, scrubbing, pigging, and high-pressure water jetting of the components fouled. They are conceptually simple and ecologically sound, but often very labor-intensive, marginally efficient, and not always applicable. The physical methods proposed include the use of electric currents, magnetism, ultrasound, ultraviolet light, manipulation of water flow, turbulence, water temperature, desiccation, and hypoxia. Most of these methods are also ecologically acceptable since they only affect the organisms within the plant’s installations and do not release pollutants (except for dead plants and animals) into the waterbody. Hot water treatment is often used in thermal and nuclear power plants to clean their cooling systems (reviewed in Claudi & Mackie, 1994; Karatayev et al., 1994b; Rajagopal et al., 2010). The availability of large volumes of hot water in these facilities can make this method cheap and efficient if retrofitting the plant to allow for the backwash of hot water is feasible. A 100% mortality of the mussels can be achieved after 1 h of exposure to 40ºC (reviewed in Karatayev et al., 1994b; Mackie & Claudi, 2010; Rajagopal et al., 2010); at 55 ºC, all mussels not only die but also detach from the substrate (reviewed in Karatayev et al., 1994b).
Chemical control includes the use of various molluscicides, such as chlorine, bromine, ozone, aromatic hydrocarbon compounds, quaternary ammonium compounds, and several others, with chlorination being most common (reviewed in Karatayev et al., 1994b; Claudi & Mackie, 1994; Sprecher and Getsinger, 2000; Mackie & Claudi, 2010). The advantage of chemical treatments is their efficiency and ease of use, but they involve toxic compounds that affect all organisms, have stringent environmental regulations, and often require costly detoxification procedures at the outlet, despite which a fraction can end up discharged back to the environment where they cause serious negative impacts on numerous non-target species (reviewed in Claudi & Mackie, 1994; Mackie & Claudi, 2010). Encapsulating the active ingredient (KCl) in microscopic particles of edible material (the BioBullets) can potentially reduce the risk of environment pollution (Aldridge et al., 2006; Costa et al., 2011).
A biological control method based on a toxin produced by the bacteria Pseudomonas fluorescens (Zequanox) was recently registered by the U.S. EPA for use in the open water and does not affect non-target species (Molloy et al., 2013a, b; Meehan et al., 2014; Waller & Bertsh, 2018). However, it was designed for protection of industrial and municipal water intakes and can be costly and impractical for the eradication of mussels from large waterbodies (Molloy et al. 2013a, b; Lund et al., 2018; Waller & Bartsh, 2018).
Conclusions and future needs
Although the two Dreissena species have many similarities, they differ in many key aspects. Their histories of spread and invasion dynamics are different, as well as their tolerance to environmental parameters and distribution patterns within waterbodies, resulting in different ecosystem impacts. Much more information on the spread, biology, ecology, and impacts are available on zebra than on quagga mussels (Figs. 8–10), and most of these studies were conducted in polymictic lakes. As a result, data from deep dimictic lakes are scarcer and there is almost no information for waterbodies colonized by quagga mussels only.

State of knowledge of life history parameters of Dreissena polymorpha and D. r. bugensis. 0—no data found; 1—data very scarce; 2—some data available; 3—well covered. Bold font indicates topics that are treated in subsequent lines, and the state of knowledge is an average estimate for the topic. Data, measurement units, and references are provided in Table 2
While the name D. r. bugensis is the most widely accepted in the literature, additional studies are needed to better understand the biological differences between D. r. bugensis and D. rostriformis and to confirm if D. r. bugensis and the Caspian Sea populations of D. rostriformis are effectively isolated by their different salinity tolerances.
Although the life cycles of both dreissenids are similar, their biological traits are not identical, and even subtle differences can lead to large disparities in spread, competition outcomes, and ecosystem effects. Again, in contrast to the better-studied zebra mussels, data on the fecundity, duration of the planktonic stage, time to sexual maturity, and sex ratio of quagga mussels are very scarce or absent (Fig. 8). Due to these blanks, data on fecundity (and other traits) of quagga mussels are often derived from information on the zebra mussels, but these extrapolations may be speculative and unwarranted. Because of the ongoing spread of D. r. bugensis in both Europe and North America and their ability to form large populations below the thermocline in deep stratified lakes, it is critically important to quantify these life history parameters, as well as to identify the influence of environmental variables (e.g., temperature, depth, food availability) on their reproductive potential. Even for the intensively studied D. polymorpha, reproduction patterns have been adequately covered in shallow, temperate, and warm habitats only, whereas very little is known for their deep-water populations. More work is also needed to determine the spawning cues (e.g., temperature and food) of deep-water quagga mussels, as well as their longevity in the profundal zone of stratified lakes, where their growth is limited by both low temperature and low food concentrations.
The growth rates of Dreissena spp. depend on a variety of environmental factors (Fig. 9), and in the presence of both species D. r. bugensis usually grows faster than D. polymorpha. However, it is very difficult to tease apart the independent effects of these factors, especially in natural waterbodies, where many of them covary. For example, the decline in water temperature in the fall usually coincides with declines in food availability. Similarly, depth is associated with temperature, light penetration, and food availability. Several factors may have additive or synergistic effects, complicating interpretations of the dominant drivers involved, an essential requirement for the adequate understanding of the biology and ecology of these mussels.

State of knowledge on the effects of different variables on the growth and longevity of Dreissena polymorpha and D. r. bugensis. 0—no data found; 1—data very scarce; 2—some data available; 3—well covered. Bold font indicates topics that are treated in subsequent lines, and the state of knowledge is an average estimate for the topic. Data, measurement units, and references are provided in Table 3
The longevity of both dreissenid species is still an open question (Fig. 9). Although data on their longevity in polymictic lakes are abundant, their life span at low temperatures below the thermocline, especially when food resources are depleted, is practically unknown. We also do not know if early in the invasion, when food resources are usually sufficient, dreissenids growth faster and live shorter than later in the invasion.
Knowledge of the species’ physiological limits is imperative for predicting dreissenid spread and distribution. There are virtually no studies addressing the lower temperature limit for both D. polymorpha and D. r. bugensis (Fig. 10). Considering that temperature limits can depend on the climatic zone from which mussels were collected, acclimation time, and the design of the experiment, both upper and lower temperature limits observed in the field, rather than in the lab, are most probably more meaningful for the prediction of the potential geographic limits to their spread. Virtually all reports agree that pH and calcium are among the most important environmental variables limiting the spread of zebra and, likely also, quagga mussels. While data for zebra mussels are numerous, for quagga mussels such information on pH and calcium are lacking. Additional research is also needed to define whether there are significant differences to key environmental variables between the adults and their larvae.

State of knowledge of habitat characteristics and environmental limits of Dreissena polymorpha and D. r. bugensis. 0—no data found; 1—data very scarce; 2—some data available; 3—well covered. Bold font indicates topics that are treated in subsequent lines, and the state of knowledge is an average estimate for the topic. Data, measurement units, and references are provided in Table 4
During their history of invasion in the Old and New Worlds, zebra mussels dispersed faster and utilized more vectors of spread than quagga mussels. However, the recent fast expansion of quaggas in Europe and to a lower extent in North America, suggests that in the near future they may colonize most lakes and rivers already inhabited by zebra mussels.
For both polymictic and dimictic lakes, abundant data are available for the initial stages of the invasion when populations are growing; however, long-term trends surveys of the populations and their impacts are relatively scarce, and their results are often inconclusive, likely due to the different methods used but also to system-specific settings. Therefore, more lake-wide long-term studies of dreissenids are needed to understand their dynamics under different environmental conditions, as well as the potential outcomes of the long-term co-existence of both species in the same waterbody.
Among specific impacts, the impacts of Dreissena spp. on fish vary depending on the feeding mode of the fishes, morphology of the invaded waterbody, time since mussel invasion, co-evolution history, Dreissena species, and are different in Europe and North America. Early in the North American invasion, the degree of predation on dreissenid mussels was believed to be limited. However, with time, dreissenids become an important component of diet for many commercially important native fishes in invaded North American freshwaters, especially after the introduction of round gobies which serve as an important pass of energy accumulated by dreissenids in benthic environment back to the pelagic that may increase fish productivity. More studies on the effect of dreissenids on the flow of energy through food webs in different continents and waterbody types are needed to fully understand and predict their impact on recipient fish communities.
Dreissenid densities and population dynamics can be affected by their natural enemies, including fishes, birds, and other predators, as well as parasites. Although multiple cases of the local impact of predation were reported in the literature, these studies were usually limited in space and time. Additional research is needed to understand if predators and parasites can cause long-term, system-wide declines in mussel populations.
Many reports, chiefly from Europe, suggest that dreissenids are responsible for important ecosystem services, including water purification, nutrient recycling, provision of habitat for other benthic invertebrates, and food for fishes, waterfowl, and other animals. They have also been widely used as sentinel organisms for biomonitoring purposes. These benefits and their economically quantifiable effects should be included in the assessments of their economic impacts. Currently, damage caused by invasive species attracts more attention than their benefits (Guerin et al., 2018; Jernelöv, 2017), and assessments of damage only are dominant in the literature (Perrings et al., 2001; Pimentel, 2011; Diagne et al., 2021). While dreissenids do have sizable benefits, many of which involve major economic gains, their ecosystemic and economic benefits are usually ignored, minimized, or deemed “non-monetizable” (Haubrock et al., 2022; but see also Boltovskoy et al., 2022). Given the widespread distribution of invasive mussels and the long-term focus of scientists and managers on the negative aspects of their dispersal, it is important to assess quantitatively their positive ecological effects and economic benefits as an opportunity to provide an additional piece of information for scientists, managers, and policymakers (Vimercati et al., 2020).
During the last century effective methods for the early detection, prevention, and control of dreissenids were developed on both continents. Routine mussel monitoring programs are extremely important and should include tracking of mussel biomass and distributions and changes in mussel demographics. So far all control methods are applicable to the culling of biofouling of human-made facilities, but not in the wild. Genetic engineering approaches have been proposed to eradicate other invasive mussels at regional scales (e.g., Rebelo et al., 2018), but their viability has not been confirmed.
Finally, we should stress that in spite of our efforts, this review cannot cover every aspect of what we know about dreissenids, as the volume of published work to date is outstanding and is still growing. Summarizing this large body of literature, however, we tried to highlight the most important sources and review papers to guide the readers for future exploration. Most importantly, we have been able to pinpoint what we do not know about these mussels in order to encourage future research efforts.
Data availability
Data sharing not applicable to this review article, as only publicly available datasets were analyzed during the current study.
References
Ackerman, J. D., B. Sim, S. J. Nichols & R. Claudi, 1994. A review of the early life history of zebra mussels (Dreissena polymorpha): comparisons with marine bivalves. Canadian Journal of Zoology 72: 1169–1179.
Aksu, S., D. Yıldız & A. P. Güngör, 2017. How Zebra mussels threaten to water supply security and effects of preventive measures in Turkey. World Scientific News 64: 99–126.
Aladin, N. V., V. I. Gontar, L. V. Zhakova, I. S. Plotnikov, A. O. Smurov, P. Rzymski & P. Klimaszyk, 2019. The zoocenosis of the Aral Sea: six decades of fast-paced change. Environmental Science and Pollution Research 26: 2228–2237.
Aldridge, D. W., B. S. Payne & A. C. Miller, 1995. Oxygen consumption, nitrogenous excretion, and filtration rates of Dreissena polymorpha at acclimation temperatures between 20 and 32°C. Canadian Journal of Fisheries and Aquatic Sciences 52: 1761–1767.
Aldridge, D. C., P. Elliott & G. D. Moggridge, 2006. Microencapsulated BioBullets for the control of biofouling zebra mussels. Environmental Science & Technology 40: 975–979.
Aldridge, D. C., S. Ho & E. Froufe, 2014. The Ponto-Caspian quagga mussel, Dreissena rostriformis bugensis (Andrusov, 1897), invades Great Britain. Aquatic Invasions 9: 529–535.
Alexander, J. E., Jr., J. H. Thorp & R. D. Fell, 1994. Turbidity and temperature effects on oxygen consumption in zebra mussels (Dreissena polymorpha). Canadian Journal of Fisheries and Aquatic Sciences 51: 179–184.
Allen, Y. C. & C. W. Ramcharan, 2001. Dreissena distribution in commercial waterways of the U.S.: using failed invasions to identify limiting factors. Canadian Journal of Fisheries and Aquatic Sciences 58: 898–907.
Allen, D. C. & C. C. Vaughn, 2009. Burrowing behavior of freshwater mussels in experimentally manipulated communities. Journal of the North American Benthological Society 28: 93–100.
Allen, Y. C., B. A. Thompson & C. W. Ramcharan, 1999. Growth and mortality rates of the zebra mussel, Dreissena polymorpha, in the Lower Mississippi River. Canadian Journal of Fisheries and Aquatic Sciences 56: 748–759.
Amberg, J. J., C. M. Merkes, W. Stott, C. B. Rees & R. A. Erickson, 2019. Environmental DNA as a tool to help inform zebra mussel, Dreissena polymorpha, management in inland lakes. Management of Biological Invasions 10: 96–110.
Andreeva, S. I. & N. I. Andreev, 2003. Evolutionary transformations of bivalves of the Aral Sea under the conditions of the environmental crisis. Omsk, Omsk State Pedagogical University Press, Russia. 382 pp. (in Russian).
Andrusov, N., 1890. Dreissena rostriformis Desh. In the River Bug. Vestnik Estestvoznaniya I 6: 261–262 (in Russian).
Andrusov, N. I., 1897. Fossil and living Dreissensiidae of Eurasia. Trudy Sankt-Peterburgskago Obschestva Estestvoispitatelei. Department of Geology and Mineralogy. 25: 683 pp. (in Russian with German summary).
Antonov, P. I. & G. L. Shkorbatov, 1983. Eco-physiological and eco-morphological variability of the Volga River population of Dreissena polymorpha (Pallas). In Sokolov, N. E. (ed), Species and Their Productivity Within Their Range Nauka Press, Moscow: 135–141. (in Russian).
Antonov, P. I. & G. L. Shkorbatov, 1990. Eco-physiological characteristics of Dreissena of the lower reaches of the Dnieper River. In Khmeleva, N. N., et al., (eds), Species Within Their Range: Biology, Ecology and Productivity of Aquatic Invertebrates Navuka i Tekhnika Press, Minsk: 161–166. (in Russian).
Antonov, P. I. & S. V. Kozlovsky, 2001. Spontaneous areal expansion of some Ponto-Caspian species in reservoirs cascades. In William, P. G. et al. (eds), Abstracts of the U.S. – Russia invasive species workshop. Yaroslavl’ State Technical University Press, Yaroslavl, Russia: 16–18. (in Russian).
Apeti, D. A. & G. G. Lauenstein, 2006. An assessment of mirex concentrations along the southern shorelines of the Great Lakes, USA. American Journal of Environmental Sciences 2: 95–103.
Araujo, R. & Y. de Jong, 2015. Fauna Europaea: Mollusca – Bivalvia. Biodiversity Data Journal 3: e5211. https://doi.org/10.3897/BDJ.3.e5211.
Araujo, R., M. Valladolid & I. Gomez, 2010. Life cycle and density of a newcomer population of zebra mussels in the Ebro River, Spain. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 183–189.
Ardura, A., A. Zaiko, Y. J. Borrell, A. Samuiloviene & E. Garcie-Vazquez, 2017. Novel tools for early detection of a global aquatic invasive, the zebra mussel Dreissena polymorpha. Aquatic Conservation: Marine and Freshwater Ecosystems 27: 165–176.
Arnott, D. L. & M. J. Vanni, 1996. Nitrogen and phosphorus recycling by the zebra mussel (Dreissena polymorpha) in the western basin of Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 53: 646–659.
Arslan, N., S. Kökmen-Aras & D. Mercan, 2018. An indigenous species, Dreissena polymorpha (Pallas, 1771) (Mollusca, Bivalvia), as an invader in Lake Büyük Akgöl. Transylvanian Review of Systematical and Ecological Research 20: 39–50.
Arter, H. E., 1989. Effect of eutrophication on species composition and growth of freshwater mussels (Mollusca, Unionidae) in Lake Hallwil (Aargau, Switzerland). Aquatic Science 51: 87–99.
Babak, E. V., 1983. The Pliocene and Quarternary Dreissenidae of the Euxinian Basin. Trudy Paleontologicheskogo Instituta, Akademia Nauk SSSR 204: 1–104 (in Russian).
Babushkin, E. S., M. V. Vinarski, A. A. Gerasimova, S. N. Ivanov & T. A. Sharapova, 2022. First find of Dreissena polymorpha (Pallas, 1771) (Mollusca, Bivalvia) in Siberia. Russian Journal of Biological Invasions 1: 13–21.
Bacchetta, R., P. Mantecca & G. Vailati, 2010. Reproductive behaviour of zebra mussels living in shallow and deep water in the South Alps lakes. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 161–168.
Baer, J., C. Spiessl & A. Brinker, 2022. Size matters? Species- and size-specific fish predation on recently established invasive quagga mussels Dreissena rostriformis bugensis Andrusov 1897 in a large, deep oligotrophic lake. Journal of Fish Biology 100: 1272–1282.
Bailey, S. A., M. G. Deneau, L. Jean, C. J. Wiley, B. Leung & H. J. MacIsaac, 2013. Evaluating efficacy of an environmental policy to prevent biological invasions. Environmental Science and Technology 45: 2554–2561.
Baker, S. M. & D. J. Hornbach, 2000. Physiological status and biochemical composition of a natural population of unionid mussels (Amblema plicata) infested by zebra mussels (Dreissena polymorpha). American Midland Naturalist 143: 443–452.
Baker, S. M. & J. S. Levinton, 2003. Selective feeding by three native North American freshwater mussels implies food competition with zebra mussels. Hydrobiologia 505: 97–105.
Baker, S. M. & D. J. Hornbach, 2008. Zebra mussels (Dreissena polymorpha) attached to native mussels (Unionidae) or inanimate substrates: comparison of physiological rates and biochemical composition. American Midland Naturalist 160: 20–28.
Baldwin, B. S., M. S. Mayer, J. Dayton, N. Pau, J. Mendilla, M. Sullivan, A. Moore, A. Ma & E. L. Mills, 2002. Comparative growth and feeding in zebra and quagga mussels (Dreissena polymorpha and Dreissena bugensis): implications for North American lakes. Canadian Journal of Fisheries and Aquatic Sciences 59: 680–694.
Balogh, C., A. Vláčilová & L. G.-Tóth & Z. Serfőző, 2018. Dreissenid colonization during the initial invasion of the quagga mussel in the largest Central European shallow lake, Lake Balaton, Hungary. Journal of Great Lakes Research 44: 114–125.
Balogh, C., Z. Serfőző & A. bij de Vaate, R. Noordhuis & J. Kobak, 2019. Biometry, shell resistance and attachment of zebra and quagga mussels at the beginning of their co-existence in large European lakes. Journal of Great Lakes Research 45: 777–787.
Barbiero, R. P., B. M. Lesht & G. J. Warren, 2011. Evidence for bottom–up control of recent shifts in the pelagic food web of Lake Huron. Journal of Great Lakes Research 37: 78–85.
Barbiero, R. P., B. M. Lesht & G. J. Warren, 2012. Convergence of trophic state and the lower food web in Lakes Huron, Michigan and Superior. Journal of Great Lakes Research 38: 368–380.
Barbiero, R. P., B. M. Lesht, G. J. Warren, L. G. Rudstam, J. M. Watkins, E. D. Reavie, K. E. Kovalenko & A. Y. Karatayev, 2018. A comparative examination of recent changes in nutrients and lower food web structure in Lake Michigan and Lake Huron. Journal of Great Lakes Research 44: 573–589.
Barrett, K. B., J. M. Haynes & D. I. Warton, 2017. Thirty years of change in a benthic macroinvertebrate community of southwestern Lake Ontario after invasion by four Ponto-Caspian species. Freshwater Science 36: 90–102.
Barton, D. R., R. A. Johnson, L. Campbell, J. Petruniak & M. Patterson, 2005. Effects of round gobies (Neogobius melanostomus) on dreissenid mussels and other invertebrates in eastern Lake Erie, 2002–2004. Journal of Great Lakes Research 31: 252–261.
Bashnin, T., V. Verhaert, M. De Jonge, L. Vanhaecke, J. Teuchies & L. Bervoets, 2019. Relationship between pesticide accumulation in transplanted zebra mussel (Dreissena polymorpha) and community structure of aquatic macroinvertebrates. Environmental Pollution 252: 591–598.
Bayba, S., L. E. Burlakova, A. Y. Karatayev & R. J. Warren II., 2022. Non-native Dreissena associated with increased native benthic community abundance with greater lake depth. Journal of Great Lakes Research 48: 734–745.
Beattie, A. M., M. R. Whiles & P. W. Willink, 2017. Diets, population structure, and seasonal activity patterns of mudpuppies (Necturus maculosus) in an urban, Great Lakes coastal habitat. Journal of Great Lakes Research 43: 132–143.
Beekey, M. A., D. J. McCabe & J. E. Marsden, 2004. Zebra mussels affect benthic predator foraging success and habitat choice on soft sediments. Oecologia 141: 164–170.
Beisel, J.-N., V. Bachmann & J.-C. Moreteau, 2010. Growth-at-length model and related life-history traits of Dreissena polymorpha in lotic systems. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 191–197.
Benson, A. J., 2014. Chronological history of zebra and quagga mussels (Dreissenidae) in North America, 1988–2010. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, Florida, USA: 9–31.
Benson, A. J., D. Raikow, J. Larson, A. Fusaro, A. K. Bogdanoff & A. Elgin, 2021a. Dreissena polymorpha (Pallas, 1771): U.S. Geological Survey, Nonindigenous Aquatic Species Database, Gainesville, FL, https://nas.er.usgs.gov/queries/FactSheet.aspx?speciesID=5, Revision Date: 5/17/2021a, Access Date: 11/18/2021a.
Benson, A. J., M. M. Richerson, E. Maynard, J. Larson, A. Fusaro, A. K. Bogdanoff, M. E. Neilson & A. Elgin, 2021b. Dreissena rostriformis bugensis (Andrusov, 1897): U.S. Geological Survey, Nonindigenous Aquatic Species Database, Gainesville, FL, https://nas.er.usgs.gov/queries/FactSheet.aspx?speciesID=95, Revision Date: 9/13/2019, Access Date: 11/18/2021b.
Benton, M. J., 1993. Fossil Record 2. London: Chapman & Hall: 845 pp.
Bidwell, J. R., 2010. Range expansion of Dreissena polymorpha: a review of major dispersal vectors in Europe and North America. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 69–78.
Bighiu, M. A., A. Norman Haldén, W. Goedkoop & J. Ottoson, 2019. Assessing microbial contamination and antibiotic resistant bacteria using zebra mussels (Dreissena polymorpha). Science of the Total Environment 650: 2141–2149.
bij de Vaate, A., 1991. Distribution and aspects of population dynamics of the zebra mussel, Dreissena polymorpha (Pallas, 1771), in the Lake IJsselmeer area (The Netherlands). Oecologia 86: 40–50.
bij de Vaate, A., 2010. Some evidence for ballast water transport being the vector of the quagga mussel (Dreissena rostriformis bugensis Andrusov 1897) introduction into Western Europe and subsequent upstream dispersal in the River Rhine. Aquatic Invasions 5: 207–209.
bij de Vaate, A., M. Greijdanus-Klaas & H. Smit, 1992. Densities and biomass of zebra mussels in the Dutch part of the lower Rhine. In Neumann, D. & H. A. Jenner (eds), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management Gustav Fisher, Stuttgart: 67–77.
bij de Vaate, A., K. Jazdzewski, H. A. M. Ketelaars, S. Gollash & G. van der Velde, 2002. Geographical patterns in range extension of Ponto-Caspian macroinvertebrate species in Europe. Canadian Journal of Fisheries and Aquatic Sciences 59: 1159–1174.
bij de Vaate A., G. Van der Velde, R. S. E. W. Leuven & K. C. M. Heiler, 2014. Spread of the quagga mussel, Dreissena rostriformis bugensis, in Western Europe. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, Florida, USA: 83–92.
Bilandžija, H., B. Morton, M. Podnar & H. Ćetković, 2013. Evolutionary history of relict Congeria (Bivalvia: Dreissenidae): unearthing the subterranean biodiversity of the Dinaric Kars. Frontiers in Zoology 10: 5.
Binelli, A., S. Magni, C. Soave, F. Marazzi, E. Zuccato, S. Castiglion, M. Parolini & V. Mezzanotte, 2014. The biofiltration process by the bivalve D. polymorpha for the removal of some pharmaceuticals and drugs of abuse from civil wastewaters. Ecological Engineering 71: 710–721.
Binelli, A., C. Della Torre, S. Magni & M. Parolini, 2015. Does zebra mussel (Dreissena polymorpha) represent the freshwater counterpart of Mytilus in ecotoxicological studies? A critical review. Environmental Pollution 196: 386–403.
Binimelis, R., I. Monterroso & B. Rodriguez-Labajos, 2007. A social analysis of the bioinvasions of Dreissena polymorpha in Spain and Hydrilla verticillata in Guatemala. Environmental Management 40: 555–566.
Birger, T. I., A. Y. Malyarevskaya & O. M. Arsan, 1975. Physiological aspects of adaptations of molluscs to abiotic and biotic factors, due to blue-green algae. In Likharev, I. M. (ed), Molluscs: Systematic, Evolution and Signification in the Nature Nauka Press, Leningrad, Abstracts of the Fifth Meeting on the Investigation of Molluscs: 91–94. (in Russian).
Bitterman, A. M., R. D. Hunter & R. C. Haas, 1994. Allometry of shell growth of caged and uncaged zebra mussels (Dreissena polymorpha) in Lake St. Clair. American Malacological Bulletin 11: 41–49.
Bobat, A., M. O. Hengírmen & W. Zapletal, 2004. Zebra mussel and fouling problems in the Euphrates Basin. Turkish Journal of Zoology 28: 161–177.
Boegehold, A. G., N. S. Johnson & D. R. Kashian, 2019. Dreissenid (quagga and zebra mussel) veligers are adversely affected by bloom forming cyanobacteria. Ecotoxicology and Environmental Safety 182: 109426.
Boltovskoy, D., R. Guiaşu, L. Burlakova, A. Karatayev, M. Schlaepfer & N. Correa, 2022. Misleading estimates of economic impacts of biological invasions: including the costs but not the benefits. Ambio 51: 1786–1799.
Borcherding, J., 1991. The annual reproductive cycle of the freshwater mussel Dreissena polymorpha Pallas in lakes. Oecologia 87: 208–218.
Borcherding, J., E. D. De Ruyter & van Steveninck, 1992. Abundance and growth of Dreissena polymorpha larvae in the water column of the River Rhine during downstream transportation. In Neumann, D. & H. A. Jenner (eds), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management Gustav Fisher, Stuttgart: 29–44.
Borcherding, J., 2006. Ten years of practical experience with the Dreissena Monitor, a biological early warning system for continuous water quality monitoring. Hydrobiologia 556: 417–426.
Bossenbroek, J. M., L. E. Johnson, B. Peters & D. M. Lodge, 2007. Forecasting the expansion of zebra mussels in the United States. Conservation Biology 21: 800–810.
Botts, P. S. & B. A. Patterson, 1996. Zebra mussel effects on benthic invertebrates: physical or biotic? Journal of the North American Benthological Society 15: 179–184.
Bowen, K. L., A. J. Conway & W. J. S. Currie, 2018. Could dreissenid veligers be the lost biomass of invaded lakes? Freshwater Science 37: 315–329.
Bowers, R., J. C. Sudomir, M. W. Kershner & F. A. de Szalay, 2005. The effects of predation and unionid burrowing on bivalve communities in a Laurentian Great Lake coastal wetland. Hydrobiologia 545: 93–102.
Bradley, B. A., B. B. Laginhas, R. Whitlock, J. M. Allen, A. E. Bates, G. Bernatchez, J. M. Diez, R. Early, J. Lenoir, M. Vilà & C. J. B. Sorte, 2019. Disentangling the abundance-impact relationship for invasive species. Proceedings of the National Academy of Sciences of the United States of America 116: 9919–9924.
Breaker, B. S., K. L. Pangle, K. C. Donner, J. B. Smith, B. A. Turschak, R. M. Claramunt, D. B. Bunnell & J. L. Jonas, 2020. Piscivory in recovering Lake Michigan cisco (Coregonus artedi): The role of invasive species. Journal of Great Lakes Research 46: 1402–1411.
Bruner, K. A., S. W. Fisher & P. F. Landrum, 1994. The role of the zebra mussel, Dreissena polymorpha, in contaminant cycling: II. Zebra mussel contaminant accumulation from algae and suspended particles, and transfer to the benthic invertebrate, Gammarus fasciatus. Journal of Great Lakes Research 20: 735–750.
Brush, J. M., A. T. Fisk, N. E. Hussey & T. B. Johnson, 2012. Spatial and seasonal variability in the diet of round goby (Neogobius melanostomus): stable isotopes indicate that stomach contents overestimate the importance of dreissenids. Canadian Journal of Fisheries and Aquatic Sciences 69: 573–586.
Buchan, L. A. J. & D. K. Padilla, 1999. Estimating the probability of long-distance overland dispersal of invading aquatic species. Ecological Applications 9: 254–265.
Bulté, G. & G. Blouin-Demers, 2008. Northern map turtles (Graptemys geographica) derive energy from the pelagic pathway through predation on zebra mussels (Dreissena polymorpha). Freshwater Biology 53: 497–508.
Bunnell, D. B., T. B. Johnson & C. T. Knight, 2005. The impact of introduced round gobies (Neogobius melanostomus) on phosphorus cycling in central Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 62: 15–29.
Burlakova, L. E., 1998. Ecology of Dreissena polymorpha (Pallas) and its role in the structure and function of aquatic ecosystems. Candidate Dissertation, Zoology Institute of the Academy of Science Republic Belarus, Minsk (Belarus): 167 pp. (in Russian).
Burlakova, L. E., A. Y. Karatayev & D. K. Padilla, 2000. The impact of Dreissena polymorpha (Pallas) invasion on unionid bivalves. International Review of Hydrobiology 85: 529–541.
Burlakova, L. E., A. Y. Karatayev & D. K. Padilla, 2005. Functional changes in benthic freshwater communities after Dreissena polymorpha (Pallas) invasion and consequences for filtration. In: Dame, R. & S. Olenin (eds), The Comparative Roles of Suspension Feeders in Ecosystems. NATO Science Series: IV – Earth and Environmental Sciences. Springer: 263–275.
Burlakova, L. E., A. Y. Karatayev & D. K. Padilla, 2006. Changes in the distribution and abundance of Dreissena polymorpha within lakes through time. Hydrobiologia 517: 133–146.
Burlakova, L. E., A. Y. Karatayev & V. A. Karatayev, 2012. Invasive mussels induce community changes by increasing habitat complexity. Hydrobiologia 685: 121–134.
Burlakova, L. E., B. L. Tulumello, A. Y. Karatayev, R. A. Krebs, D. W. Schloesser, D. T. Zanatta, W. L. Patterson, T. A. Griffith, M. W. Scott & T. Crail, 2014. Competitive replacement of invasive congeners may relax impact on native species: interactions among zebra, quagga, and native unionid mussels. PLoS ONE 9(12): E114926. https://doi.org/10.1371/journal.pone.0114926.
Burlakova, L. E., R. P. Barbiero, A. Y. Karatayev, S. E. Daniel, E. K. Hinchey & G. J. Warren, 2018. The benthic community of the Laurentian Great Lakes: analysis of spatial gradients and temporal trends from 1998–2014. Journal of Great Lakes Research 44: 600–617.
Burlakova, L. E., A. Y. Karatayev, K. Mehler & E. K. Hinchey, 2022a. Exploring Great Lakes benthoscapes: can we visually delineate hypoxic habitats? Hydrobiologia. https://doi.org/10.1007/s10750-022-04821-z.
Burlakova, L. E., A. Y. Karatayev, D. Boltovskoy & N. M. Correa, 2022b. Ecosystem services provided by the exotic bivalves Dreissena polymorpha, D. rostriformis bugensis, and Limnoperna fortunei. Hydrobiologia. https://doi.org/10.1007/s10750-022-04935-4
Capizzi-Banas, S., M. P. Ladeiro, F. Bastien, I. Bonnard, N. Boudaud, C. Gantzer & A. Geffard, 2021. The utility of Dreissena polymorpha for assessing the viral contamination of rivers by measuring the accumulation of F-Specific RNA bacteriophages. Water (Switzerland) 13: 904.
Carlton, J. T., 1993. Dispersal mechanisms of the zebra mussel Dreissena polymorpha. In Nalepa, T. F. & D. W. Schloesser (ed), Zebra Mussels. Biology, Impacts, and Control. Lewis Publishers, Ann Arbor, Michigan, USA: 677–697.
Carlton, J. T., 2008. The zebra mussel Dreissena polymorpha found in North America in 1986 and 1987. Journal of Great Lakes Research 34: 770–773.
Casper, A. F. & L. E. Johnson, 2010. Contrasting shell/tissue characteristics of Dreissena polymorpha and Dreissena bugensis in relation to environmental heterogeneity in the St. Lawrence River. Journal of Great Lakes Research 36: 184–189.
Casper, A. F., L. E. Johnson & H. Glémet, 2014. In situ reciprocal transplants reveal species-specific growth pattern and geographic population differentiation among zebra and quagga mussels. Journal of Great Lakes Research 40: 705–711.
Cassini, M. H., 2020. A review of the critics of invasion biology. Biological Reviews 95: 1467–1478.
Catita, D., M. Gama, R. Azedo, F. Banha, J. Pinto, A. Ilhéu & P. M. Anastácio, 2020. Detection and possible elimination of the first recorded population of the zebra mussel (Dreissena polymorpha) in Portugal from a reservoir. Management of Biological Invasions 11: 406–414. https://doi.org/10.3391/mbi.2020.11.3.04.
Chakraborti, R. K., S. Madon & J. Kaur, 2016. Costs for controlling dreissenid mussels affecting drinking water infrastructure: Case studies. American Water Works Association 108: E442–E453.
Chapra, S. C. & D. M. Dolan, 2012. Great Lakes total phosphorus revisited: 2. Mass balance modeling. Journal of Great Lakes Research 38: 741–754.
Charlebois, P. M., J. E. Marsden, R. G. Goettel, R. K. Wolfe, D. J. Jude & S. Rudnicka, 1997. The round goby, Neogobius melanostomus (Pallas), a review of European and North American literature, Illinois Natural History Survey Special Publications, No:, 20.
Chase, M. E. & R. C. Bailey, 1999. The ecology of the zebra mussels (Dreissena polymorpha) in the lower Great Lakes of North America: I. Population dynamics and growth. Journal of Great Lakes Research 25: 107–124.
Chrisafi, E., P. Kaspiris & G. Katselis, 2007. Feeding habits of sand smelt (Atherina boyeri, Risso 1810) in Trichonis Lake (Western Greece). Journal of Applied Ichthyology 23: 209–214.
Clarke, K. B., 1952. The infestation of waterworks by Dreissena polymorpha, a freshwater mussel. Journal of the Institution of Water Engineers 6: 370–379.
Claudi, R. & G. Mackie, 1994. Zebra Mussels Monitoring and Control, Lewis Publishers, Inc., Boca Raton, FL:, 221.
Claxton, W. T. & G. L. Mackie, 1998. Seasonal and depth variations in gametogenesis and spawning of Dreissena polymorpha and Dreissena bugensis in eastern Lake Erie. Canadian Journal of Zoology 76: 2010–2019.
Claxton, W. T., A. B. Wilson, G. L. Mackie & E. G. Boulding, 1998. A genetic and morphological comparison of shallow- and deep-water populations of the introduced dreissenid bivalve Dreissena bugensis. Canadian Journal of Zoology 76: 1269–1276.
Cleven, E. J. & P. Frenzel, 1993. Population dynamics and production of Dreissena polymorpha (Pallas) in River Seerhein, the outlet of Lake Constance (Obersee). Archiv Für Hydrobiologie 127: 395–407.
Colborne, S. F., A. D. M. Clapp, F. J. Longstaffe & B. D. Neff, 2015. Foraging ecology of native pumpkinseed (Lepomis gibbosus) following the invasion of zebra mussels (Dreissena polymorpha). Canadian Journal of Fisheries and Aquatic Sciences 72: 983–990.
Collas, F. P. L., A. Y. Karatayev, L. E. Burlakova & R. S. E. W. Leuven, 2018. Detachment rates of dreissenid mussels after boat hull-mediated overland dispersal. Hydrobiologia 810: 77–84.
Comeau, S., S. Rainville, W. Baldwin, E. Austin, S. Gerstenberger, C. Cross & W. H. Wong, 2011. Susceptibility of quagga mussels (Dreissena rostriformis bugensis) to hot-water sprays as a means of watercraft decontamination. Biofouling 27: 267–274.
Conn, D. B. & D. A. Conn, 2007. Navigational buoy survey of invasive and native benthic invertebrates of the St. Lawrence River, Lake Ontario, and the Welland Canal. Abstracts of the 15th International Conference on Aquatic Invasive Species, Nijmegen, The Netherlands, p. 54.
Conn, D. B., F. E. Lucy & T. K. Graczyk, 2014. Dreissenid mussels as sentinel biomonitors for human and zoonotic pathogens. Chapter 17. In Nalepa, T. & D. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, Florida, USA: 265–272.
Connelly, N. A., C. R. O’Neill, B. A. Knuth & T. L. Brown, 2007. Economic impacts of zebra mussels on drinking water treatment and electric power generation facilities. Environmental Management 40: 105–112.
Conroy, J. D., D. D. Kane, D. M. Dolan, W. J. Edwards, M. N. Charlton & D. A. Culver, 2005a. Temporal trends in Lake Erie plankton biomass: roles of external phosphorus loading and dreissenid mussels. Journal of Great Lakes Research 31(Suppl. 2): 89–110.
Conroy, J. D., W. J. Edwards, R. A. Pontius, D. D. Kane, H. Zhang, J. F. Shea, J. N. Richey & D. A. Culver, 2005b. Soluble nitrogen and phosphorus excretion of exotic freshwater mussels (Dreissena spp.): potential impacts for nutrient remineralization in western Lake Erie. Freshwater Biology 50: 1146–1162.
Costa, R., D. C. Aldridge & G. D. Moggridge, 2011. Preparation and evaluation of biocide-loaded particles to control the biofouling zebra mussel, Dreissena polymorpha. Chemical Engineering Research and Design 89: 2322–2329.
Crane, D. P. & D. W. Einhouse, 2016. Changes in growth and diet of smallmouth bass following invasion of Lake Erie by the round goby. Journal of Great Lakes Research 42: 405–412.
Czarnoleski, M. & T. Müller, 2014. Antipredator strategy of zebra mussels (Dreissena polymorpha). From behavior to life history. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 345–357.
Daehler, C. C., 2009. Short lag times for invasive tropical plants: evidence from experimental plantings in Hawai’i. PLoS ONE 4: e4462.
Dall, P. C. & K. Hamburger, 1996. Recruitment and growth of Dreissena polymorpha in Lake Esrom, Denmark. Limnologica 26: 27–37.
David, K. A., B. M. Davis & R. D. Hunter, 2009. Lake St. Clair zooplankton: evidence for post-Dreissena changes. Journal of Freshwater Ecology 24: 199–209.
Deksbakh, N. K., 1935. Spread of Dreissena polymorpha Pallas (Mollusca) in the European part of the USSR and the factors responsible for their spread. Byulleten Moskovskogo Obschestva Ispytatelej Prirody, Otdelenie Biologii 44: 153–163 (in Russian).
Delmott, S. & D. Edds, 2014. Zebra mussel maturation and seasonal gametogenesis in Marion Reservoir, Kansas, USA. BioInvasions Records 3: 247–260.
Denef, V. J., H. J. Carrick, J. Cavaletto, E. Chiang, T. H. Johengen & H. A. Vanderploeg, 2017. Lake bacterial assemblage composition is sensitive to biological disturbance caused by an invasive filter feeder. mSphere 2(3): e0018917.
Denson, D. & S. Y. Wang, 1994. Morphological differences between zebra and quagga mussel spermatozoa. American Malacological Bulletin 11: 79–81.
Dermott, R., 2001. Sudden disappearance of the amphipod Diporeia from eastern Lake Ontario. Journal of Great Lakes Research 27: 423–433.
Dermott, R. & M. Munawar, 1993. Invasion of Lake Erie offshore sediments by Dreissena, and its ecological implications. Canadian Journal of Fisheries and Aquatic Sciences 50: 2298–2304.
Dermott, R. & D. Kerec, 1997. Changes to the deep water benthos of eastern Lake Erie since the invasion of Dreissena: 1979–1993. Canadian Journal of Fisheries and Aquatic Sciences 54: 922–930.
Dermott, R. & M. Geminiuc, 2003. Changes in the benthic fauna of Lake Ontario 1990–1995, with local trends after 1981. In Munawar, M. (ed), State of Lake Ontario (SOLO)-Past, Present, and Future Ecovision World Monograph Series, Backhaus Publishers, Leiden, The Netherlands: 323–345.
De Ventura, L., N. Weissert, R. Tobias, K. Kopp & J. Jokela, 2016. Overland transport of recreational boats as a spreading vector of zebra mussel Dreissena polymorpha. Biological Invasions 18: 1451–1466.
Diagne, C., B. Leroy, R. E. Gozlan, A. C. Vaissiere, C. Assailly, L. Nuninger, D. Roiz, F. Jourdain, I. Jaric & F. Courchamp, 2020. InvaCost, a public database of the economic costs of biological invasions worldwide. Scientific Data 7: 277.
Diagne, C., B. Leroy, A.-C. Vaissière, R. E. Gozlan, D. Roiz, I. Jarić, J.-M. Salles, C. J. A. Bradshaw & F. Courchamp, 2021. High and rising economic costs of biological invasions worldwide. Nature 592: 571–576.
Diaz, R. J. & R. Rosenberg, 2008. Spreading dead zones and consequences for marine ecosystems. Science 321: 926–929.
Dietrich, J. P., B. J. Morrison & J. A. Hoyle, 2006. Alternative ecological pathways in the eastern Lake Ontario food web—round goby in the diet of lake trout. Journal of Great Lakes Research 32: 395–400.
Diggins, T. P., 2001. A seasonal comparison of suspended sediment filtration by quagga (Dreissena bugensis) and zebra (D. polymorpha) mussels. Journal of Great Lakes Research 27: 457–466.
Diggins, T. P., M. Weimer, K. M. Stewart, R. E. Baier, A. E. Meyer, R. F. Forsberg & M. A. Goehle, 2004. Epiphytic refugium: are two species of invading freshwater bivalves partitioning spatial resources? Biological Invasions 6: 83–88.
Dionisio Pires, L. M., B. M. Bontes, E. Van Donk & B. W. Ibelings, 2005. Grazing on colonial and filamentous, toxic and non-toxic cyanobacteria by the zebra mussel Dreissena polymorpha. Journal of Plankton Research 27: 331–339.
Dionisio Pires, L. M, B. W. Ibelings & E. van Donk, 2010. Zebra mussels as a potential tool in the restoration of eutrophic shallow lakes, dominated by toxic cyanobacteria. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 331–342.
DiVittorio, J., M. Grodowitz & J. Snow, 2012. Inspection and Cleaning Manual for Equipment and Vehicles to Prevent the Spread of Invasive Species. U.S. Department of the Interior, Bureau of Reclamation, Technical Memorandum No. 86-68220-07-05. https://www.landcan.org/pdfs/USBREquipmentInspectionandCleaningManual2012(1).pdf
Dobiesz, N. E. & P. L. Negel, 2009. Changes in mid-summer water temperature and clarity across the Great Lakes between 1968 and 2002. Journal of Great Lakes Research 35: 371–384.
Domm, S., R. W. McCauley, E. Kott & J. D. Ackerman, 1993. Physiological and taxonomic separation of two dreissenid mussels in the Laurentian Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 50: 2294–2297.
Dorgelo, J., 1993. Growth and population structure of the zebra mussel (Dreissena polymorpha) in Dutch lakes different in trophic state. In Nalepa, T. F. & D. W. Schloesser (ed), Zebra Mussels. Biology, Impacts, and Control. Lewis Publishers, Ann Arbor, Michigan, USA: 79–94.
Dorgelo, J. & M. Gorter, 1984. Preliminary data on size composition and settlement of Dreissena polymorpha (Pallas) (Molluska: Bivalvia) in lakes differing in trophic state. Hydrobiological Bulletin 18: 159–163.
Dove, A., 2009. Long-term trends in major ions and nutrients in Lake Ontario. Aquatic Ecosystem Health & Management 12: 281–295.
Dove, A. & S. C. Chapra, 2015. Long-term trends of nutrients and trophic response variables for the Great Lakes. Limnology and Oceanography 60: 696–721.
Drake, J. M. & J. M. Bossenbroek, 2009. Profiling ecosystem vulnerability to invasion by zebra mussels with support vector machines. Theoretical Ecology 2: 189–198.
Draulans, D. & R. Wouters, 1988. Density, growth and calorific value of Dreissena polymorpha (Mollusca: Bivalvia) in a pond created by sand extraction, and its importance as food for fish. Annales de la Société Royale Zoologique de Belgique 118: 51–60.
Dussart, B., 1966. Limnologie, Gauthier-Villars, Paris:
Dyga, A. K. & V. I. Zolotareva, 1976. Biology of Dreissena bugensis Andrusov of Zaporozhskoe Reservoir and their role in water purification. In Andrushaitis, G. P., et al., (eds), Abstracts of the Third Congress of the All-Union Hydrobiological Society, Vol. 3. Zinatne Press, Riga: 237–239. (in Russian).
Elagina, T. S., T. G. Zyablova & E. M. Shubina, 1978. Distribution and density of Dreissena polymorpha Pallas in a heated bay, from Kostromskaya thermal power station, in the Shacha River. In: Terrestrial and Aquatic Ecosystems. Gorkiy, Russia: 141–145 (in Russian).
Elgin, A. K., P. W. Glyshaw & B. C. Weidel, 2022. Depth drives growth dynamics of dreissenid mussels in Lake Ontario. Journal of Great Lakes Research 48: 289–299.
Elliott, P., D. C. Aldridge & G. D. Moggridge, 2008. Zebra mussel filtration and its potential uses in industrial water treatment. Water Research 42: 1664–1674.
Ellis, S. & H. J. Macisaac, 2009. Salinity tolerance of Great Lakes invaders. Freshwater Biology 54: 77–89.
Evans, M. A., G. Fahnenstiel & D. Scavia, 2011. Incidental oligotrophication of North American Great Lakes. Environmental Science and Technology 45: 3297–3303.
Evariste, L., E. David, P.-L. Cloutier, P. Brousseau, M. Auffret, M. Desrosiers, P. E. Groleau, M. Fournier & S. Betoulle, 2018. Field biomonitoring using the zebra mussel Dreissena polymorpha and the quagga mussel Dreissena bugensis following immunotoxic reponses. Is there a need to separate the two species? Environmental Pollution 238: 706–716.
Fahnenstiel, G. L., G. A. Lang, T. F. Nalepa & T. H. Johengen, 1995. Effects of zebra mussel (Dreissena polymorpha) colonization on water quality parameters in Saginaw Bay, Lake Huron. Journal of Great Lakes Research 21: 435–448.
Fahnenstiel, G. L., S. A. Pothoven, T. F. Nalepa, H. A. Vanderploeg, D. M. Klarer & D. Scavia, 2010. Recent changes in primary production and phytoplankton in the offshore region of southeastern Lake Michigan. Journal of Great Lakes Research 36: 20–29.
Fernald, R. T. & B. T. Watson, 2014. Eradication of zebra mussels (Dreissena polymorpha) from Millbrook Quarry, Virginia. Rapid response in the real world. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, Florida, USA: 195–213.
Fleischer, G. W., T. J. Desorcie & J. D. Holuszko, 2001. Lake-wide distribution of Dreissena in Lake Michigan, 1999. Journal of Great Lakes Research 27: 252–257.
Foley, C. J., S. R. Andree, S. A. Pothoven, T. F. Nalepa & T. O. Höök, 2017. Quantifying the predatory effect of round goby on Saginaw Bay dreissenids. Journal of Great Lakes Research 43: 121–131.
Franchini, D. A., 1978. Vertical distribution of Dreissena polymorpha (Pallas) in Garda Lake. Bollettino Di Zoologia 45: 257–260.
Francis, J. T., S. R. Robillard & J. E. Marsden, 1996. Yellow perch management in Lake Michigan: a multi-jurisdictional challenge. Fisheries (Bethesda) 21: 18–20.
French, J. R. P., III., J. S. Schaeffer, E. F. Roseman, C. S. Kiley & A. Fouilleroux, 2009. Abundance and distribution of benthic invertebrates in offshore soft sediments in Western Lake Huron, 2001–2007. Journal of Great Lakes Research 35: 120–127.
Friedland R., A.-L. Buer, S. Dahlke & G. Schernewski, 2019. Spatial effects of different zebra mussel farming strategies in an eutrophic Baltic Lagoon. Frontiers in Environmental Science 6 doi:https://doi.org/10.3389/fenvs.2018.00158.
Garton, D. V. & L. E. Johnson, 2000. Variation in growth rates of the zebra mussel, Dreissena polymorpha, within Lake Wawasee. Freshwater Biology 45: 443–451.
Garton, D. W., R. McMahon & A. M. Stoeckmann, 2014. Limiting Environmental factors and competitive interactions between zebra and quagga mussels in North America. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 383–402.
Geary, D. H., I. Magyar & P. Müller, 2000. Ancient Lake Pannon and its endemic molluscan fauna (Central Europe; Mio-Pliocene). Advances in Ecological Research 31: 463–482.
Géba, E., D. Aubert, L. Durand, S. Escotte, S. La Carbona, C. Cazeaux, I. Bonnard, F. Bastien, M. Palos Ladeiro, J. P. Dubey, I. Villena, A. Geffard & A. Bigot-Clivot, 2020. Use of the bivalve Dreissena polymorpha as a biomonitoring tool to reflect the protozoan load in freshwater bodies. Water Research 170: 115297.
Géba, E., D. Rioult, O. Palluel, O. Dedourge-Geffard, S. Betoulle, D. Aubert & A. Bigot-Clivot, 2021a. Resilience of Dreissena polymorpha in wastewater effluent: Use as a bioremediation tool? Journal of Environmental Management 278: 111513.
Géba, E., A. Rousseau, A. Le Guernic, S. Escotte-Binet, L. Favennec, S. La Carbona, G. Gargala, J. P. Dubey, I. Villena, S. Betoulle, D. Aubert & A. Bigot-Clivot, 2021b. Survival and infectivity of Toxoplasma gondii and Cryptosporidium parvum oocysts bioaccumulated by Dreissena polymorpha. Journal of Applied Microbiology 130: 504–515.
Gelembiuk, G. W., G. E. May & C. E. Lee, 2006. Phylogeography and systematics of zebra mussels and related species. Molecular Ecology 15: 1033–1050.
George, S. D., B. T. Duffy, B. P. Baldigo, D. Skaros & A. J. Smith, 2022. Condition of macroinvertebrate communities in the Buffalo River Area of Concern following sediment remediation. Journal of Great Lakes Research 48: 183–194.
Gergs, R., K. Rinke & K.-O. Rothhaupt, 2009. Zebra mussels mediate benthic-pelagic coupling by biodeposition and changing detrital stoichiometry. Freshwater Biology 54: 1379–1391.
Gerstenberger, S. L., A. A. Mueting & W. H. Wong, 2011. Veligers of invasive quagga mussels (Dreissena rostriformis bugensis, Andrusov 1897) in Lake Mead, Nevada-Arizona. Journal of Shellfish Research 30: 933–938.
Gill, T., 1871. Arrangement of the families of mollusks. Smithsonian Miscellaneous Collections 227: xvi, 1–49.
Gillis, P. L. & G. L. Mackie, 1994. Impact of the zebra mussel, Dreissena polymorpha, on populations of Unionidae (Bivalvia) in Lake St. Clair. Canadian Journal of Zoology 72: 1260–1271.
Gingera, T. D., R. Bajno, M. F. Docker & J. D. Reist, 2017. Environmental DNA as a detection tool for zebra mussels Dreissena polymorpha (Pallas, 1771) at the forefront of an invasion event in Lake Winnipeg, Manitoba, Canada. Management of Biological Invasions 8: 287–300.
Ginn, B. K., R. Bolton, D. Coulombe, T. Fleischaker & G. Yerex, 2018. Quantifying a shift in benthic dominance from zebra (Dreissena polymorpha) to quagga (Dreissena rostriformis bugensis) mussels in a large, inland lake. Journal of Great Lakes Research 44: 271–282.
Glyshaw, P. W., C. M. Riseng, T. F. Nalepa & S. A. Pothoven, 2015. Temporal trends in condition and reproduction of quagga mussels (Dreissena rostriformis bugensis) in southern Lake Michigan. Journal of Great Lakes Research 41: 16–26.
Goedkoop, W., R. Naddafi & U. Grandin, 2011. Retention of N and P by zebra mussels (Dreissena polymorpha Pallas) and its quantitative role in the nutrient budget of eutrophic Lake Ekoln, Sweden. Biological Invasions 13: 1077–1086.
Goedkoop, W., M. I. Choudhury, D. C. P. Lau & U. Grandin, 2021. Inverting nutrient fluxes across the land-water interface – Exploring the potential of zebra mussel (Dreissena polymorpha) farming. Journal of Environmental Management 281: 111889.
Gomes, J., A. Matos, R. M. Quinta-Ferreira & R. C. Martins, 2018. Environmentally applications of invasive bivalves for water and wastewater decontamination. Science of the Total Environment 630: 1016–1027.
Gray, J. E., 1840. A manual of the land and fresh-water shells of the British Islands. Turton W. (ed), Longman, Orme, Brown, Green, and Longmans, London: ix + 324 pp.
Griffiths, R. W., D. W. Schloesser, J. H. Leach & W. P. Kovalak, 1991. Distribution and dispersal of the zebra mussel (Dreissena polymorpha) in the Great Lakes region. Canadian Journal of Fisheries and Aquatic Sciences 48: 1381–1388.
Grigoryev, B. F., 1965. On composition and distribution of molluscs in lower reaches of the South Bug River. In Likharev, I. M. (ed), Molluscs: Problems of Theoretical and Applied Malacology Nauka Press, Leningrad, Abstracts of the Second Meeting on the Investigation of Molluscs: 86–89. (in Russian).
Grim, J., 1971. Tiefenverteilung der Dreikantmuschel Dreissena polymorpha (Pallas) im Bodensee. Gas Wasserfach Wasser Abwasser 112: 437–441 (in German).
Guerin, G. R., I. Martín-Forés, B. Sparrow & A. J. Lowe, 2018. The biodiversity impacts of non-native species should not be extrapolated from biased single-species studies. Biodiversity and Conservation 27: 785–790.
Gutierrez, J. L., C. G. Jones, D. L. Strayer & O. O. Iribarne, 2003. Mollusks as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101: 79–90.
Haag, W. R., D. R. Berg, D. W. Garton & J. L. Farris, 1993. Reduced survival and fitness in native bivalves in response to fouling by the introduced zebra mussel (Dreissena polymorpha) in western Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 50: 13–19.
Haines-Young, R. & M. Potschin, 2011. Common international classification of ecosystem services (CICES): 2011 update. Expert Meet Ecosystem Accounts 1–17.
Hahm, J., J. B. Manchester-Neesvig, D. DeBord & W. C. Sonzogni, 2009. Polybrominated diphenyl ethers (PBDEs) in Lake Michigan forage fish. Journal of Great Lakes Research 35: 154–158.
Halfman, J. D., 2016. Water quality of the Finger Lakes, New York: 2005 – 2016. Finger Lakes Institute, Hobart & William Smith Colleges, Geneva, NY: 52 pp. Available at: http://people.hws.edu/halfman/Data/2016%20FL-WQ-Update.pdf
Hallac, D. E. & J. E. Marsden, 2000. Differences in tolerance to and recovery from zebra mussel (Dreissena polymorpha) fouling by Elliptio complanata and Lampsilis radiata. Canadian Journal of Zoology 78: 161–166.
Hallstan, S., U. Grandin & W. Goedkoop, 2010. Current and potential distribution of the zebra mussel (Dreissena polymorpha) in Sweden. Biological Invasions 12: 285–296.
Hamilton, D. J., C. D. Ankney & R. C. Bailey, 1994. Predation of zebra mussels by diving ducks: An exclosure study. Ecology 75: 521–531.
Hammond, D. & G. Ferris, 2019. Low doses of EarthTec QZ ionic copper used in effort to eradicate quagga mussels from an entire Pennsylvania lake. Management of Biological Invasions 10: 500–516.
Hasler, C. T., J. Leathers, A. Ducharme & N. J. Casson, 2019. Biological effects of water velocity and other hydrodynamic characteristics of flow on dreissenid mussels. Hydrobiologia 837: 1–14.
Haubrock, P. J., R. N. Cuthbert, A. Ricciardi, C. Diagne & F. Courchamp, 2022. Economic costs of invasive bivalves in freshwater ecosystems. Diversity and Distributions 28: 1010–1021.
Hebert, P. D. N., B. W. Muncaster & G. L. Mackie, 1989. Ecological and genetic studies on Dreissena polymorpha (Pallas): a new mollusc in the Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 46: 1587–1591.
Hecky, R. E., R. E. H. Smith, D. R. Barton, S. J. Guildford, W. D. Taylor, M. N. Charlton & E. T. Howell, 2004. The near shore phosphorus shunt: A consequence of ecosystem engineering by dreissenids in the Laurentian Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 61: 1285–1293.
Heiler, K. C. M., A. bij de Vaate, K. Ekschmitt, P. V. von Oheimb, C. Albrecht & T. Wilke, 2013. Reconstruction of the early invasion history of the quagga mussel (Dreissena rostriformis bugensis) in Western Europe. Aquatic Invasions 1: 53–57.
Hetherington, A., L. G. Rudstam, R. L. Schneider, K. T. Holeck, C. W. Hotaling, J. E. Cooper & J. R. Jackson, 2019. Invader invaded: population dynamics of zebra mussels (Dreissena polymorpha) and quagga mussels (Dreissena rostriformis bugensis) in polymictic Oneida Lake, NY, USA (1992–2013). Biological Invasions 21: 1529–1544.
Higgins, S. N., 2014. Meta-analysis of dreissenid effects on freshwater ecosystems. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 487–494.
Higgins, S. N. & M. J. Vander Zanden, 2010. What a difference a species makes: A meta-analysis of dreissenid mussel impacts on freshwater ecosystems. Ecological Monographs 80: 179–196.
Hillbricht-Ilkowska, A. & A. Stanczykowska, 1969. The production and standing crop of planktonic larvae of Dreissena polymorpha (Pall.) in two Mazurian lakes. Polish Archives of Hydrobiology 16: 193–203.
Horvath, T. G. & G. A. Lamberti, 1999. Recruitment and growth of zebra mussels (Dreissena polymorpha) in a coupled lake-stream system. Archiv Für Hydrobiologie 145: 197–217.
Horvath, T. G., G. A. Lamberti, D. M. Lodge & W. L. Perry, 1996. Zebra mussel dispersal in lake–stream systems: source–sink dynamics? Journal of the North American Benthological Society 15: 564–575.
Hoyle, J. A., T. Schaner, J. M. Casselman & R. Dermott, 1999. Changes in lake whitefish (Coregonus clupeaformis) stocks in eastern Lake Ontario following Dreissena mussel invasion. Great Lakes Research Review 4: 5–10.
Hoyle, J. A., J. N. Bowlby & B. J. Morrison, 2008. Lake whitefish and walleye population responses to dreissenid mussel invasion in eastern Lake Ontario. Aquatic Ecosystem Health and Management 11: 403–411.
Hubenov, Z. & T. Trichkova, 2007. Dreissena bugensis (Mollusca: Bivalvia: Dreissenidae): New invasive species to the Bulgarian malacofauna. Acta Zoologica Bulgarica 59: 203–209.
Hunter, R. D. & J. F. Bailey, 1992. Dreissena polymorpha (zebra mussel): colonization of soft substrata and some effects on unionid bivalves. Nautilus 106: 60–67.
Hunter, R. G. & K. A. Simons, 2004. Dreissenids in Lake St. Clair in 2001: Evidence for population regulation. Journal of Great Lakes Research 30: 528–537.
Ianniello, R. S., 2013. Effects of environmental variables on the reproduction of quagga mussels (Dreissena rostriformis bugensis) in Lake Mead, NV/AZ. UNLV Theses, Dissertations, Professional Papers, and Capstones. 1842. https://doi.org/10.34917/4478261.
Ibelings, B. W., R. Portielje, E. H. R. R. Lammens, R. Noordhuis, M. S. van den Berg, W. Joosse & M. L. Meijer, 2007. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study. Ecosystems 10: 4–16.
Immel, F., C. Broussard, B. Catherinet, L. Plasseraud, G. Alcaraz, I. Bundeleva & F. Marin, 2016. The shell of the invasive bivalve species Dreissena polymorpha: biochemical, elemental and textural investigations. PLoS ONE. https://doi.org/10.1371/journal.pone.0154264.
Jackson, J., A. VanDeValk, T. Brooking, O. VanKeeken & L. Rudstam, 2002. Growth and feeding dynamics of lake sturgeon, Acipenser fulvescens, in Oneida Lake, New York: results from the first five years of a restoration program. Journal of Applied Ichthyology 18: 439–443.
Jackson, J. R., A. J. VanDeValk, T. E. Brooking, K. T. Holeck, C. Hotaling & L. G. Rudstam, 2020. The fisheries and limnology of Oneida Lake 2019, New York State Department of Environmental Conservation, Albany, NY, USA: 25.
Jacobs, G. R., E. L. Bruestle, A. Hussey, D. Gorsky & A. T. Fisk, 2017. Invasive species alter ontogenetic shifts in the trophic ecology of Lake Sturgeon (Acipenser fulvescens) in the Niagara River and Lake Ontario. Biological Invasions 19: 1533–1546.
Jakus, P., M. J. Kealy, J. Loomis, N. Nelson, J. Ostermiller, C. Stanger & N. von Stackelberg, 2013. Economic Benefits of Nutrient Reductions in Utah’s Waters. Report prepared for the State of Utah, Utah Division of Water Quality. Salt Lake City, Utah: CH2M Hill. Available at: https://documents.deq.utah.gov/water-quality/standards-technical-services/DWQ-2020-000676.pdf.
Jantz, B, 1996. Wachstum, Reproduktion, Populationsentwicklung und Beeinträchtigung der Zebramuschel (Dreissena polymorpha) in einem großen Fließgewässer, dem Rhein. University of Cologne Ph.D. Thesis. Cologne, Germany: 141 pp. (in German with English summaries).
Jantz, B. & D. Neumann, 1992. Shell growth and population dynamics of Dreissena polymorpha in the River Rhine. In Neumann, D. & H. A. Jenner (eds), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management Gustav Fisher, Stuttgart: 49–66.
Jantz, B. & D. Neumann, 1998. Growth and reproductive cycle of the zebra mussel in the River Rhine as studied in a river bypass. Oecologia 114: 213–225.
Jernelöv, A., 2017. The long-term fate of invasive species: aliens forever or integrated immigrants with time? Springer, Cham (Switzerland).
Jimenez, A., M. D. Rennie, W. G. Sprules & J. La Rose, 2011. Temporal changes in the benthic invertebrate community of Lake Simcoe, 1983–2008. Journal of Great Lakes Research 37: 103–112.
Johengen, T. H., T. F. Nalepa, G. L. Fahnenstiel & G. Goudy, 1995. Nutrient changes in Saginaw Bay, Lake Huron after the establishment of the zebra mussel Dreissena polymorpha. Journal of Great Lakes Research 21: 449–464.
Johns, C. & B. E. Timmerman, 1998. Total cadmium, copper, and zinc in two dreissenid mussels, Dreissena polymorpha and Dreissena bugensis, at the outflow of Lake Ontario. Journal of Great Lakes Research 24: 55–64.
Johnson, L. E. & J. T. Carlton, 1996. Post-establishment spread in large-scale invasions: dispersal mechanisms of the zebra mussel Dreissena polymorpha. Ecology 77: 1686–1690.
Johnson, L. E. & D. K. Padilla, 1996. Geographic spread of exotic species: ecological lessons and opportunities from the invasion of the zebra mussel Dreissena polymorpha. Biological Conservation 78: 23–33.
Johnson, M. F. & M. E. Meder, 2013. Effects of aquatic invasive species on home prices: evidence from Wisconsin. Human Settlement & Movement eJournal. https://papers.ssrn.com/sol3/papers.cfm?abstract_id=2316911.
Johnson, L. E., A. Ricciadri & J. T. Carlton, 2001. Overland dispersal of aquatic invasive species: a risk assesment of transient recreational boating. Ecological Applications 11: 1789–1799.
Johnson, T. B., D. B. Bunnell & C. T. Knight, 2005. A potential new energy pathway in Central Lake Erie: the round goby connection. Journal of Great Lakes Research 31(Suppl. 2): 238–251.
Johnson, L. E., J. M. Bossenbroek & C. E. Kraft, 2006. Patterns and pathways in the postestablishment spread of non-indigenous aquatic species: the slowing invasion of North American inland lakes by the zebra mussel. Biological Invasions 8: 475–489.
Jones, L. A. & A. Ricciardi, 2005. Influence of physicochemical factors on the distribution and biomass of invasive mussels (Dreissena polymorpha and Dreissena bugensis) in the St. Lawrence River. Canadian Journal of Fisheries and Aquatic Sciences 62: 1953–1962.
Jude, D. J., R. H. Reider & G. R. Smith, 1992. Establishment of Gobiidae in the Great Lakes basin. Great Lakes Research Review 3: 27–34.
Kachanova, A. A., 1963. About growth of Dreissena polymorpha Pallas in the Uchinskoe Reservoir and in the channels of Moswater-supply. In Sokolova, N. Y. (ed), Uchinskoe and Mozhaiskoe Reservoirs Moscow University Press, Moscow: 226–234. (in Russian).
Karatayev, A. Y., 1981. Larval stage of Dreissena polymorpha Pallas in the Lake Lukomlskoe, a cooling reservoir of thermal power plant. Vestnik Belorusskogo Universiteta. Ser. 2(3): 54–59 (in Russian).
Karatayev, A. Y., 1983. Ecology of Dreissena polymorpha Pallas and its effects on macrozoobenthos of the thermal power plant’s cooling reservoir. Candidate Dissertation, Zoology Institute of Academy of Science Belorussian SSR, Minsk, Belarus:153 pp. (in Russian).
Karatayev, A. Y. & G. M. Tishchikov, 1979. Effect of heated water of thermal power plant on the growth of Dreissena polymorpha Pallas. Vestnik Belorusskogo Universiteta. Ser. 2(3): 39–43 (in Russian).
Karatayev, A. Y. & V. P. Lyakhnovich, 1990. Effect of Dreissena polymorpha Pallas on benthic crustaceans (Gammarus lacustris Sars, Pallasea quadrispinosa Sars and Asellus aquaticus L.) in Lukomskoe lake. In Khmeleva, N. N., et al., (eds), Species Within Their Range: Biology, Ecology, and Productivity of Aquatic Invertebrates Navuka i Tekhnika Press, Minsk, Belarus: 123–125. (in Russian).
Karatayev, A. Y. & L. E. Burlakova, 1992. Changes in tropic structure of macrozoobenthos of an eutrophic lake, after invasion of Dreissena polymorpha. Biologiya Vnutrennikh Vod. Informacionnyi Bulletin 93: 67–71 (in Russian).
Karatayev, A. Y. & L. E. Burlakova, 1995. Present and further patterns in Dreissena polymorpha (Pallas) population development in the Narochanskaya lakes system. Vestsi Akademii Navuk Belarusi. Seriya Biyalagichnikh Navuk 3: 95–99 (in Belarusian).
Karatayev, A. Y. & L. E. Burlakova, 1995b. The role of Dreissena in lake ecosystems. Russian Journal of Ecology 26(3): 207–211. Translated into English from Ecologiya 26(3): 232–236.
Karatayev, A. Y. & L. E. Burlakova, this issue. Dreissena in the Great Lakes: what have we learned in 30 years of invasion. Hydrobiologia. https://doi.org/10.1007/s10750-022-04990-x
Karatayev, A. Y., G. M. Tishchikov & I. V. Karatayeva, 1983. Dreissena polymorpha Pallas as a specific community of benthic animals. Biologiya Vnutrennikh Vod. Informatsionnyi Bulletin 61: 18–21 (in Russian).
Karatayev, A. Y., V. P. Lyakhnovich, S. A. Afanasiev, L. E. Burlakova, V. P. Zakutskiy, S. M. Lyakhov, M. P. Miroshnichenko, T. G. Moroz, M. Y. Nekrasova, S. P. Nechvalenko, I. A. Skalskaya, T. G. Kharchenko & A. A. Protasov, 1994a. The place of species in ecosystem. In Starobogatov, J. I. (ed), Freshwater zebra mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, ecology, practical meaning. Nauka Press, Moscow, USSR: 180–195 (in Russian).
Karatayev, A. Y., V. P. Mikheev, S. A. Afanasiev, M. Y. Kirpichenko, A. A. Protasov, L. V. Shevtsova & T. G. Kharchenko, 1994b. Practical uses and control on artificial structures. In Starobogatov, J. I. (ed), Freshwater zebra mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, ecology, practical meaning. Nauka Press, Moscow, USSR: 206–221 (in Russian).
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 1997. The effects of Dreissena polymorpha (Pallas) invasion on aquatic communities in eastern Europe. Journal of Shellfish Research 16: 187–203.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 1998. Physical factors that limit the distribution and abundance of Dreissena polymorpha (Pall.). Journal of Shellfish Research 17: 1219–1235.
Karatayev, A. Y., I. A. Rudakovskiy, P. G. Prischepov & L. E. Burlakova, 1999. The fish productivity of Belarusian lakes. In Proceedings of the international conference: the present status and perspectives of development of aquaculture. Belarusian Agricultural Academy Press, Gorky, Belarus: 22–23 (in Russian).
Karatayev, A. Y., L. E. Burlakova, D. P. Molloy & L. K. Volkova, 2000a. Endosymbionts of Dreissena polymorpha (Pallas) in Belarus. International Review of Hydrobiology 85: 543–559.
Karatayev, A. Y., L. E. Burlakova & D. P. Molloy, 2000b. Seasonal dynamics of Conchophthirus acuminatus (Ciliophora, Conchophthiridae) infection in Dreissena polymorpha and D. bugensis (Bivalvia, Dreissenidae). European Journal of Protistology 36: 397–404.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2002. Impacts of zebra mussels on aquatic communities and their role as ecosystem engineers. In Leppakoski, E., S. Gollach & S. Olenin (eds), Invasive Aquatic Species of Europe: Distribution, Impacts and Management Kluwer Academic Publishers, Dordreicht: 433–446.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2003. Patterns of spread of the zebra mussel (Dreissena polymorpha (Pallas)): the continuing invasion of Belarussian lakes. Biological Invasions 5: 213–221.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2006. Growth rate and longevity of Dreissena polymorpha (Pallas): a review and recommendations for future study. Journal of Shellfish Research 25: 23–32.
Karatayev, A. Y., D. Boltovskoy, D. K. Padilla & L. E. Burlakova, 2007a. The invasive bivalves Dreissena polymorpha and Limnoperna fortunei: parallels, contrasts, potential spread and invasion impacts. Journal of Shellfish Research 26: 205–213.
Karatayev, A. Y., D. K. Padilla, D. Minchin, D. Boltovskoy & L. E. Burlakova, 2007b. Changes in global economies and trade: the potential spread of exotic freshwater bivalves. Biological Invasions 9: 161–180.
Karatayev, A. Y., L. E. Burlakova, D. P. Molloy & S. E. Mastitsky, 2007c. Dreissena polymorpha and Conchophthirus acuminatus: What can we learn from host-commensal relationships. Journal of Shellfish Research 26: 1153–1160.
Karatayev, A. Y., L. E. Burlakova, S. E. Mastitsky & S. Olenin, 2008. Past, current, and future of the Central European Corridor for aquatic invasions in Belarus. Biological Invasions 10: 215–232.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2010. Dreissena polymorpha in Belarus: history of spread, population biology, and ecosystem impacts. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 101–112.
Karatayev, A. Y., L. E. Burlakova, S. E. Mastitsky, D. K. Padilla & E. L. Mills, 2011a. Contrasting rates of spread of two congeners, Dreissena polymorpha and Dreissena rostriformis bugensis, at different spatial scales. Journal of Shellfish Research 30: 923–931.
Karatayev, A. Y., S. E. Mastitsky, D. K. Padilla, L. E. Burlakova & M. M. Hajduk, 2011b. Differences in growth and survivorship of zebra and quagga mussels: size matters. Hydrobiologia 668: 183–194.
Karatayev, V. A., A. Y. Karatayev, L. E. Burlakova & D. K. Padilla, 2013. Lakewide dominance does not predict the potential for spread of dreissenids. Journal of Great Lakes Research 39: 622–629.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2014a. General overview of zebra and quagga mussels what we do and do not know. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 695–703.
Karatayev, A. Y., L. E. Burlakova, C. Pennuto, J. Ciborowski, V. A. Karatayev, P. Juette & M. Clapsadl, 2014b. Twenty five years of changes in Dreissena spp. populations in Lake Erie. Journal of Great Lakes Research 40: 550–559.
Karatayev, V. A., A. Y. Karatayev, L. E. Burlakova & L. D. Rudstam, 2014c. Eutrophication and Dreissena invasion as drivers of biodiversity: a century of change in the mollusc community of Oneida Lake. PLoS One 9(7): e101388.
Karatayev, A. Y., L. E. Burlakova & D. K. Padilla, 2015a. Zebra versus quagga mussels: a review of their spread, population dynamics, and ecosystem impacts. Hydrobiologia 746: 97–112.
Karatayev, A., D. Boltovskoy, L. Burlakova & D. Padilla, 2015b. Parallels and contrasts between Limnoperna fortunei and Dreissena species. In Boltovskoy, D. (ed) Limnoperna fortunei, The Ecology, Distribution and Control of a Swiftly Spreading Invasive Fouling Mussel. Invading Nature – Springer Series in Invasion Ecology 10: 261–297.
Karatayev, A. Y., L. E. Burlakova, S. E. Mastitsky & D. K. Padilla, 2015c. Predicting the spread of aquatic invaders: insight from 200 years of invasion by zebra mussels. Ecological Applications 25: 430–440.
Karatayev, A. Y., L. E. Burlakova, K. Mehler, S. A. Bocaniov, P. D. Collingsworth, G. Warren, R. T. Kraus & E. K. Hinchey, 2018a. Biomonitoring using invasive species in a large lake: Dreissena distribution maps hypoxic zones. Journal of Great Lakes Research 44: 639–649.
Karatayev, A. Y., V. A. Karatayev, L. E. Burlakova, M. D. Rowe, K. Mehler & M. D. Clapsadl, 2018b. Food depletion regulates the demography of invasive dreissenid mussels in a stratified lake. Limnology and Oceanography 63: 2065–2079.
Karatayev, A. Y., L. E. Burlakova, K. Mehler, R. P. Barbiero, E. K. Hinchey, P. D. Collingsworth, K. E. Kovalenko & G. Warren, 2018c. Life after Dreissena: The decline of exotic suspension feeder may have significant impacts on lake ecosystems. Journal of Great Lakes Research 44: 650–659.
Karatayev, A. Y., V. A. Karatayev, L. E. Burlakova, K. Mehler, M. D. Rowe, A. K. Elgin & T. F. Nalepa, 2021a. Lake morphometry determines Dreissena invasion dynamics. Biological Invasions 23: 2489–2514.
Karatayev, A. Y., L. E. Burlakova, K. Mehler, S. E. Daniel & A. R. Hrycik, 2021b. Lake Ontario Benthos Survey Cooperative Science and Monitoring Initiative 2018. Technical Report. USEPA-GLRI GL00E02254. Great Lakes Center, SUNY Buffalo State, Buffalo, New York, USA: 37 pp. Available at: https://greatlakescenter.buffalostate.edu/sites/greatlakescenter.buffalostate.edu/files/uploads/Documents/Publications/LakeOntarioBenthosSurveyCSMI2018FinalReport.pdf.
Karatayev, A. Y., L. E. Burlakova, K. Mehler, E. K. Hinchey, M. Wick, M. Bakowska & N. Mrozinska, 2021c. Rapid assessment of Dreissena population in Lake Erie using underwater videography. Hydrobiologia 848: 2421–2436.
Karatayev, V. A., L. G Rudstam, A. Y. Karatayev, L. E. Burlakova, B. V. Adamovich, H. A Zhukava, K. T. Holeck, A. L. Hetherington, J. R. Jackson, C. W. Hotaling, T. V. Zhukova, T. M. Mikheyeva, R. Z. Kovalevskaya, O. A. Makarevich & D. V. Kruk, 2021d. Serial invasions can disrupt the time course of ecosystem recovery, bioRxiv preprint doi: https://doi.org/10.1101/2021d.10.29.466526.
Karatayev, A. Y., L. E. Burlakova, K. Mehler, L. G. Rudstam, J. M. Watkins & M. Wick, 2022. Dreissena in Lake Ontario 30 years post-invasion. Journal of Great Lakes Research 48: 264–273.
Karpevich, A. F., 1952. Growth of some molluscs of Caspian and Aral seas. Doklady VNIRO on Biology, Taxonomy and Feeding of Fish, Sea Chemistry and Preservation of Nets. Pischepromizdat Press, Moscow, USSR 1: 116–120 (in Russian).
Karpevich, A. F., 1955. Response of invertebrates of the Azov Sea to changes in salinity. Trudy Atlanticheskogo Nauchno-Issledovatel’skogo Instituta Rybnogo Khozyaistva i Okeanografii 31: 240–275 (in Russian).
Karpevich, A. F., 1964. Distinctive features of reproduction and growth of bivalves in brackish seas of the USSR. In Zenkevich, L. A. (ed), The Ecology of Invertebrates in Southern Seas of the USSR Nauka Press, Moscow: 3–60. (in Russian).
Katona, L. R., L. E. Burlakova, A. K. Karatayev, Y. Vadeboncoeur, In review. Microbial N cycling may explain progressive enrichment of benthic primary producer and dreissenid 15N with depth in lakes. Ecosystems.
Katsev, S., 2017. When large lakes respond fast: A parsimonious model for phosphorus dynamics. Journal of Great Lakes Research 43: 199–204.
Keen, M. & H. Casey, 1969. Family Corbiculidae. In Moore, R. C. (ed), Treatise on Invertebrate Paleontology. Part N. Mollusca 6. Bivalvia 2. Geological Society of America, Boulder, and University of Kansas: 643–644.
Keller, R. P., J. M. Drake & D. M. Lodge, 2007. Fecundity as a basis for risk assessment of nonindigenous freshwater molluscs. Conservation Biology 21: 191–200.
Kelly, D. W., L.-M. Herborg & H. J. MacIsaak, 2010. Ecosystem changes associated with Dreissena invasions: recent developments and emerging issues. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 199–209.
Kerfoot, W. C., F. Yousef, S. A. Green, J. W. Budd, D. J. Schwab & H. A. Vanderploeg, 2010. Approaching storm: disappearing winter bloom in Lake Michigan. Journal of Great Lakes Research 36(Suppl. 3): 30–41.
Kern, R., J. Borcherding & D. Neumann, 1994. Recruitment of a freshwater mussel with a planktonic life-stage in running waters - Studies on Dreissena polymorpha in the River Rhine. Archiv Für Hydrobiologie 131: 385–400.
Kerney, M. P. & B. S. Morton, 1970. The distribution of Dreissena polymorpha (Pallas) in Britain. Journal of Conchology 27: 97–100.
Kharchenko, T. A., 1995. Dreissena: range, ecology, biological handicaps. Hydrobiological Journal 31(3): 3–21.
Kharchenko, T. A. & L. V. Shevtsova, 1983. Expansion of the range of Dreissena due to construction of canals for interbasin water transfer. In Likharev, I. M. (ed), Molluscs: Systematics, Ecology and Patterns of Occurrence Nauka Press, Leningrad, Abstracts of the Seventh Meeting on the Investigation of Molluscs: 218–220 (in Russian).
Khusainova, N. Z., 1958. The Biological Features of Some Abundant Bottom Invertebrates Used by Fishes in the Aral Sea. Kazakhstan State University Press, Alma-Ata, USSR, 116 pp. (in Russian).
Kinzelbach, R., 1992. The main features of the phylogeny and dispersal of the zebra mussel Dreissena polymorpha. In Neumann, D. & H. A. Jenner (eds), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management. Gustav Fisher, Stuttgart, Germany: 5–17.
Kiritani, K. & K. Yamamura, 2003. Exotic insects and their pathways for invasion. In Ruiz, G. M. & J. T. Carlton (eds), Invasive Species: Vectors and Management Island Press, Washington, DC: 44–67.
Kirkendall, D. S., D. B. Bunnell, P. M. Armenio, L. A. Eaton, A. S. Trebitz & N. M. Watson, 2021. Spatial and temporal distributions of Dreissena spp. veligers in Lake Huron: does calcium limit settling success? Journal of Great Lakes Research 47: 1040–1049.
Kirpichenko, M. Y., 1964. Phenology, population dynamics, and growth of Dreissena larvae in the Kuibyshev Reservoir. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 7(10): 15–24 (in Russian).
Kirpichenko, M. Y., 1971. On ecology of Dreissena polymorpha Pallas in Tsimlyanskoe Reservoir. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 21: 142–154 (in Russian).
Kissman, C. E. H., L. B. Knoll & O. Sarnelleb, 2010. Dreissenid mussels (Dreissena polymorpha and Dreissena bugensis) reduce microzooplankton and macrozooplankton biomass in thermally stratified lakes. Limnology and Oceanography 55: 1851–1859.
Kobak, J. & T. Kakareko, 2009. Attachment strength, aggregation and movement of the zebra mussel (Dreissena polymorpha, Bivalvia) in the presence of potential predators. Fundamental and Applied Limnology 174: 193–204.
Kocovsky, P. M., 2019. Diets of endangered silver chub (Macrhybopsis storeriana, Kirtland, 1844) in Lake Erie and implications for recovery. Ecology of Freshwater Fish 28: 33–40.
Kornis, M. S., N. Mercado-Silva & M. J. Vander Zanden, 2012. Twenty years of invasion: a review of round goby Neogobius melanostomus biology, spread and ecological implications. Journal of Fish Biology 80: 235–285.
Kornobis, S., 1977. Ecology of Dreissena polymorpha (Pall) (Dreissenidae, Bivalvia) in lakes receiving heated water discharges. Polish Archives of Hydrobiology 24(4): 531–545.
Korolevs A. & N. Kondratjeva, 2006. Artificial reefs and substrata in the coastal zone of the Baltic — The problems and decisions. 2006 IEEE US/EU Baltic International Symposium, pp. 1–14. https://doi.org/10.1109/BALTIC.2006.7266190.
Kovalak, W. P., G. D. Longton & R. D. Smithee, 1990. Infestation of Monroe power plant by the zebra mussel (Dreissena polymorpha). Proceedings of the American Power Conference 52: 998–1000.
Kozulin, A., 1995. Ecology of Mallards Anas platyrhynchos wintering in low temperature conditions in Belarus. Acta Ornithologica (Warsaw) 30: 125–134.
Kraft, C. E., P. J. Sullivan, A. Y. Karatayev, L. E. Burlakova, J. C. Nekola, L. E. Johnson & D. K. Padilla, 2002. Landscape patterns of an aquatic invader: assessing dispersal extent from spatial distributions. Ecological Applications 12: 749–759.
Kruglova, V. M., 1957. Production of food organisms as a means of increasing the fish productivity of waterbodies. Rybnoe Khozyaistvo 12: 21–27 (in Russian).
Kuchina, E. S., 1964. Distribution of the mollusc Dreissena polymorpha Pallas in the Northern Dvina River. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 7(10): 31–37 (in Russian).
Kusserov, R., M. Mörtl, J. Mählmann, D. Uhlmann & I. Röske, 2010. The design of a Zebra-Mussel-Biofilter. In van der Velde, G., S. Rajagopal, A. bij de Vaate (eds), The Zebra Mussel in Europe, Backhuys Publishers, Leiden (Netherlands): 323–330.
Kwon, T.-D., S. W. Fisher, G. W. Kim, H. Hwang & J.-E. Kim, 2006. Trophic transfer and biotransformation of polychlorinated biphenyls in zebra mussel, round goby, and smallmouth bass in Lake Erie, USA. Environmental Toxicology and Chemistry 25: 1068–1078.
Ladeiro, M. P., D. Aubert, I. Villena, A. Geffard & A. Bigot, 2014. Bioaccumulation of human waterborne protozoa by zebra mussel (Dreissena polymorpha): Interest for water biomonitoring. Water Research 48: 148–155.
Lancioni, T. & E. Gaino, 2006. The invasive zebra mussel Dreissena polymorpha in Lake Trasimeno (Central Italy): distribution and reproduction. Italian Journal of Zoology 73: 335–346.
Laruelle, F., D. P. Molloy & V. A. Roitman, 2002. Histological analysis of trematodes in Dreissena polymorpha: Their location, pathogenicity, and distinguishing morphological characteristics. Journal of Parasitology 88: 856–863.
Lavrentyev, P. J., H. A. Vanderploeg, G. Franze, D. H. Chacin, J. R. Liebig & T. H. Johengen, 2014. Microzooplankton distribution, dynamics, and trophic interactions relative to phytoplankton and quagga mussels in Saginaw Bay, Lake Huron. Journal of Great Lakes Research 40: 95–105.
Lazareva, V. I., A. I. Kopylov, E. A. Sokolova & E. G. Pryanichnikova, 2016. Veliger larvae of dreissenids (Bivalvia, Dreissenidae) in the plankton foodweb of Rybinsk Reservoir. Biology Bulletin 43(10): 1313–1321.
Le Guernic, A., M. Palos Ladeiro, N. Boudaud, J. Do Nascimento, C. Gantzer, J.-C. Inglard, J.-M. Mouchel, C. Pochet, L. Moulin, V. Rocher, P. Waldman, S. Wurtzer & A. Geffard, 2022. First evidence of SARS-CoV-2 genome detection in zebra mussel (Dreissena polymorpha). Journal of Environmental Management 301: 113866.
Leuven, R. S. E. W., F. P. L. Collas, K. R. Koopman, J. Matthews & G. van der Velde, 2014. Mass mortality of invasive zebra and quagga mussels by desiccation during severe winter conditions. Aquatic Invasions 9: 243–252.
Leuzinger, H. & S. Schuster, 1970. Auswirkungen der massenvermehrung de wandermuschel Dreissena polymorpha auf die Wasservögel des Bodensees. Der Ornithologische Beobachter 67: 269–274. (in German)
Lewandowski, K., 1982a. The role of early developmental stages in the dynamics of Dreissena polymorpha (Pall.) (Bivalvia) populations in Lakes. Occurrence of larvae in the plankton. Ekologia Polska 30: 81–109.
Lewandowski, K., 1982b. The role of early developmental stages in the dynamics of Dreissena polymorpha (Pall.) (Bivalvia) populations in lakes. 2. Settling of larvae and the dynamics of numbers of settled individuals. Ekologia Polska 30: 223–286.
Lewandowski, K. & J. Ejsmont-Karabin, 1983. Ecology of planktonic larvae of Dreissena polymorpha (Pall.) in lakes with different degree of heating. Polish Archives of Hydrobiology 30: 89–101.
Li, J., Y. Zhang & S. Katsev, 2018. Phosphorus recycling in deeply oxygenated sediments in Lake Superior controlled by organic matter mineralization. Limnology & Oceanography 63: 1372–1385.
Li, J., V. Ianaiev, A. Huff, J. Zalusky, T. Ozersky & S. Katsev, 2021. Benthic invaders control the phosphorus cycle in the world’s largest freshwater ecosystem. Proceedings of the National Academy of Sciences of the United States of America 118: e2008223118.
Limanova, N. A., 1964. Dreissena: Bibliography. Biology of Dreissena and its control. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 7: 83–136 (in Russian).
Limanova, N. A., 1978. Dreissena: Bibliographic Guide. Moscow: 115 pp. (in Russian).
Limburg, K. E. & K. K. Arend, 1994. Zebra Mussel veligers observed in larval fish guts. Dreissena! 5: 4.
Limburg, K. E., M. L. Pace, D. Fischer & K. K. Arend, 1997. Consumption, selectivity, and use of zooplankton by larval Striped Bass and White Perch in a seasonally pulsed estuary. Transactions of the American Fisheries Society 126: 607–621.
Limburg, K. E., V. A. Luzadis, M. Ramsey, K. L. Schulz & C. M. Mayer, 2010. The good, the bad, and the algae: Perceiving ecosystem services and disservices generated by zebra and quagga mussels. Journal of Great Lakes Research 36: 86–92.
Lindeman, P. V., 2006. Zebra and Quagga Mussels (Dreissena spp.) and other prey of a Lake Erie population of common map turtles (Emydidae: Graptemys geographica). Copeia 2: 268–273.
Logvinenko, B. M. & Y. I. Starobogatov, 1968. Mollusca. In Birshstein, Y. A. et al. (ed), Atlas of invertebrates of the Caspian Sea. Pishchevaya Promyshlennost Press, Moscow: 308–385, plate 5 (in Russian).
Lohner, R., V. Sigler, C. Mayer & C. Balogh, 2007. A comparison of the benthic bacterial communities within and surrounding Dreissena clusters in lakes. Microbial Ecology 54: 469–477.
Lovell, S. J., S. F. Stone & L. Fernandez, 2006. The economic impacts of aquatic invasive species: a review of the literature. Agricultural and Resource Economics Review 35: 195–208.
Lowe, R. L. & R. W. Pillsbury, 1995. Shifts in benthic algal community structure and function following the appearance of zebra mussel (Dreissena polymorpha) in Saginaw Bay, Lake Huron. Journal of Great Lakes Research 21: 558–566.
Lozano, S. J., J. V. Scharold & T. F. Nalepa, 2001. Recent declines in benthic macroinvertebrate densities in Lake Ontario. Canadian Journal of Fisheries and Aquatic Sciences 58: 518–529.
Lucy, F. E., 2009. Zebra mussels: Review of ecology and impacts since invasion in Ireland. In Mackie, G. & R. Claudi (eds), Monitoring and Control of Macrofouling Molluscs in Freshwater Systems CRC Press, Boca Raton, Florida, USA: 386–389.
Lucy, F. E., T. K. Graczyk, L. Tamang, A. Miraflor & D. Minchin, 2008. Biomonitoring of surface and coastal water for Cryptosporidium, Giardia, and human-virulent microsporidia using molluscan shellfish. Parasitology Research 103: 1369.
Lucy, F. E., M. Connolly, T. K. Graczyk, L. Tamang, M. Sullivan & S. E. Mastitsky, 2010. Zebra mussels (Dreissena polymorpha) are effective sentinels of water quality irrespective of their size. Aquatic Invasions 5: 49–57.
Lucy, F. E., L. E. Burlakova, A. Y. Karatayev, S. E. Mastitsky & D. T. Zanatta, 2014. Zebra mussel impacts on unionids: A synthesis of trends in North America and Europe. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, Florida, USA: 623–646.
Luferov, V. P., 1965. Organisms living anabiotically frozen in ice of the littoral zone of Rybinskoe Reservoir. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 8: 151–154 (in Russian).
Lund, K., K. B. Cattoor, E. Fieldseth, J. Sweet & M. A. McCartney, 2018. Zebra mussel (Dreissena polymorpha) eradication efforts in Christmas Lake, Minnesota. Lake and Reservoir Management 34: 7–20.
Luukkonen, D. R., E. N. Kafcas, B. T. Shirkey & S. R. Winterstein, 2014. Impacts of dreissenid mussels on the distribution and abundance of diving ducks on Lake St. Clair. Chapter 41 In Nalepa, T. & D. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control. 2nd Edition. CRC Press, Boca Raton, Florida, USA: 647–659.
Lvova, A. A., 1977. The ecology of Dreissena polymorpha (Pall.) in Uchinskoe Reservoir. Candidate Dissertation, Moscow State University, Moscow, USSR: 116 pp. (in Russian).
Lvova, A. A., 1980. Ecology of Dreissena (Dreissena polymorpha polymorpha (Pall.)). Trudy Vsesouznogo Gidrobiologicheskogo Obschestva 23: 101–119 (in Russian).
Lvova, A. A., 2004. On invasion of Dreissena bugensis (Bivalvia, Dreissenidae) in the Ucha Reservoir (Moscow obla.t) and the Moscow River. Zoologicheskii Zhurnal 83: 766–768 (in Russian with English summary).
Lvova, A. A. & G. E. Makarova, 1994. Gametogenesis and reproductive cycle. In Starobogatov, J. I. (ed), Freshwater zebra mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, ecology, practical meaning. Nauka Press, Moscow, USSR: 138–148 pp. (in Russian).
Lvova, A. A., G. E. Makarova, A. F. Alimov, A. Y. Karatayev, M. P. Miroshnichenko, V. P. Zakutsky & M. Ya. Nekrasova, 1994a. Growth and production. In Starobogatov, J. I. (ed), Freshwater zebra mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, ecology, practical meaning. Nauka Press, Moscow, USSR: 156–179 (in Russian).
Lvova, A. A., A. Y. Karatayev & G. E. Makarova, 1994b. Planktonic larva. In Starobogatov, J. I. (ed), Freshwater zebra mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, ecology, practical meaning. Nauka Press, Moscow, USSR: 149–155 (in Russian).
Lyagina, T. N. & V. D. Spanowskaya, 1963. Morphological distinctions of some fish in Uchinskoe reservoir. In Sokolova, N. Y. (ed), Uchinskoe and Mozhaiskoe Reservoirs Moskovskii Universitet Press, Moscow: 269–310. (in Russian).
Lyakhnovich, V. P., A. Y. Karatayev & P. A. Mitrakhovich, 1983. Effect of Dreissena polymorpha Pallas on ecosystem of a eutrophic lake. Biologiya Vnutrennikh Vod. Informacionnyi Bulletin 60: 25–28 (in Russian).
Lyakhnovich, V. P., A. Y. Karatayev, P. A. Mitrakhovich, L. V. Guryanova & G. G. Vezhnovets, 1988. Productivity and prospects for utilizing the ecosystem of Lake Lukoml, thermoelectric station cooling reservoir. Soviet Journal of Ecology 18: 255–259.
Lyakhnovich, V. P., A. Y. Karatayev, N. I. Andreev, S. I. Andreeva, S. A. Afanasiev, A. K. Dyga, V. P. Zakutskiy, V. I. Zolotareva, A. A. Lvova, M. Y. Nekrasova, V. F. Osadchikh, Y. V. Pligin, A. A. Protasov & G. M. Tischikov, 1994. Living conditions. In Starobogatov, J. I. (ed), Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow, USSR: 109–119 (in Russian).
Lyakhov, S. M. & V. P. Mikheev, 1964. Distribution and density of Dreissena in the Kuibyshevskoe Reservoir seven years after its construction. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 7(10): 3–18 (in Russian).
MacIsaac, H. J., 1994. Comparative growth and survival of Dreissena polymorpha and Dreissena bugensis, exotic molluscs introduced to the Great Lakes. Journal of Great Lakes Research 20: 783–790.
MacIsaac, H. J., 1996. Potential abiotic and biotic impacts of zebra mussels on the inland waters of North America. American Zoologist 36: 287–299.
Mackie, G. L., 1991. Biology of the exotic zebra mussel, Dreissena polymorpha, in relation to native bivalves and its potential impact in Lake St. Clair. Hydrobiologia 219: 251–268.
Mackie, G. L. & C. A. Wright, 1994. Ability of the zebra mussel, Dreissena polymorpha to biodeposit and remove phosphorus and BOD from diluted activated sewage sludge. Water Research 28: 1123–1130.
Mackie, G. L. & D. W. Schloesser, 1996. Comparative Biology of Zebra Mussels in Europe and North America: An Overview. American Zoologist 36: 244–258.
Mackie, G. & R. Claudi, 2010. Monitoring and Control of Macrofouling Mollusks in Fresh Water Systems, 2nd ed. CRC Press. Boca Raton, FL: 508.
Madenjian, C. P., S. A. Pothoven, P. J. Schneeberger, M. P. Ebener, L. C. Mohr, T. F. Nalepa & J. R. Bence, 2010. Dreissenid mussels are not a “dead end” in Great Lakes food webs. Journal of Great Lakes Research 36: 73–77.
Madenjian, C. P., M. A. Stapanian, L. D. Witzel, S. A. Pothoven, D. W. Einhouse & H. L. Whitford, 2011. Evidence for predatory control of the invasive Round Goby. Biological Invasions 13: 987–1002.
Madenjian, C. P., D. B. Bunnell, D. M. Warner, S. A. Pothoven, G. L. Fahnenstiel, T. F. Nalepa, H. A. Vanderploeg, I. Tsehaye, R. M. Claramunt & R. D. Clark Jr., 2015. Changes in the Lake Michigan food web following dreissenid mussel invasions: a synthesis. Journal of Great Lakes Research 41(Suppl. 3): 217–231.
Madon, S. P., D. W. Schneider, J. A. Stoeckel & R. E. Sparks, 1998. Effects of inorganic sediment and food concentrations on energetic processes of the zebra mussel, Dreissena polymorpha: implication for growth in turbid rivers. Canadian Journal of Fisheries and Aquatic Sciences 55: 401–413.
Magoulick, D. D. & L. C. Lewis, 2002. Predation on exotic zebra mussels by native fishes: Effects on predator and prey. Freshwater Biology 47: 1908–1918.
Makarevich, T. A., S. E. Mastitsky & Y. V. Savich, 2008. Phytoperiphyton on the shells of Dreissena polymorpha (Pallas) in Lake Naroch. Aquatic Invasions 3: 283–295.
Malinovskaja, L. V., A. A. Filippov, V. F. Osadchikh & N. V. Aladin, 1998. Benthic macroinvertebrates of the northern Caspian Sea during recent rises in water-level. International Journal of Salt Lake Research 7: 211–233.
Malkin, S. Y., G. M. Silsbe, R. E. H. Smith & E. T. Howell, 2012. A deep chlorophyll maximum nourishes benthic filter feeders in the coastal zone of a large clear lake. Limnology and Oceanography 57: 735–748.
Mainnín, T. O., 2021. Invasive species flexing its “mussels” on the Shannon. Raidió Teilifís Éireann, Ireland. https://www.rte.ie/news/regional/2021/0726/1237346-invasive-mussels-ireland/.
Marchowski, D., G. Neubauer, Ł Ławicki, A. Woźniczka, D. Wysocki, S. Guentzel & M. Jarzemski, 2015. The importance of non-native prey, the zebra mussel Dreissena polymorpha, for the declining greater scaup Aythya marila: A case study at a key European staging and wintering site. PLoS ONE 10: e0145496.
Marescaux, J., A. Vaate & K. Van Doninck, 2012. First records of Dreissena rostriformis bugensis (Andrusov, 1897) in the Meuse River. BioInvasions Records 1(2): 109–114.
Marescaux, J., P. Boets, J. Lorquet, R. Sablon, K. Van Doninck & J.-N. Beise, 2015. Sympatric Dreissena species in the Meuse River: towards a dominance shift from zebra to quagga mussels. Aquatic Invasions 10(3): 287–298.
Marin Jarrin, J. R., K. L. Pangle, J. M. Reichert, T. B. Johnson, J. Tyson & S. A. Ludsin, 2015. Influence of habitat heterogeneity on the foraging ecology of first feeding yellow perch larvae, Perca flavescens, in western Lake Erie. Journal of Great Lakes Research 41: 208–214.
Marsden, J. E., 1991. Development of a standardized sampling protocol to monitor the spread and density of zebra mussel (Dreissena polymorpha) populations. Dreissena Polymorpha Information Review. New York Zebra Mussel Information Clearinghouse 1(2): 4–5.
Marsden, J. E., N. Trudeau & T. Keniry, 1993. Zebra mussel study of Lake Michigan. Illinois Natural History Survey, Aquatic Ecology Technical Report 93/14: 35 pp.
Marsden, J. E. & M. A. Chotkowski, 2001. Lake trout spawning on artificial reefs and the effect of zebra mussels: fatal attraction? Journal of Great Lakes Research 27: 33–43.
Martel, A., 1993. Dispersal and recruitment of zebra mussel (Dreissena polymorpha) in a nearshore area in west-central Lake Erie: the significance of postmetamorphic drifting. Canadian Journal of Fisheries and Aquatic Sciences 50: 3–12.
Martel, A., 1995. Demography and growth of the exotic zebra mussel (Dreissena polymorpha) in the Rideau River (Ontario). Canadian Journal of Zoology 73: 2244–2250.
Martel, A. L., B. S. Baldwin, R. M. Dermott & R. A. Lutz, 2001. Species and epilimnion/hypolimnion-related differences in size at larval settlement and metamorphosis in Dreissena (Bivalvia). Limnology and Oceanography 46: 707–713.
Mastitsky, S. E., 2004. Endosymbionts of bivalve mollusc Dreissena polymorpha in waterbodies of Belarus. PhD Thesis. Institute of Zoology of the National Academy of Sciences of the Republic of Belarus, Minsk, Belarus.
Mastitsky, S. E. & V. G. Gagarin, 2004. Nematodes which infect the mollusc Dreissena polymorpha (Bivalvia: Dreissenidae) in the Narochanskie Lakes. Vestnik Belorusskogo Universiteta. Ser. 2(3): 22–25 (in Russian with English summary).
Mastitsky, S. E. & V. M. Samoilenko, 2005. Larvae of chironomids (Insecta, Diptera) encountered in the mantle cavity of zebra mussel, Dreissena polymorpha (Bivalvia, Dreissenidae). International Review of Hydrobiology 90: 42–50.
Matthews, J., G. Van der Velde, A. bij de Vaate, F. P. L. Collas, K. R. Koopman & R. S. E. W. Leuven, 2014. Rapid range expansion of the invasive quagga mussel in relation to zebra mussel presence in The Netherlands and Western Europe. Biological Invasions 16: 23–42.
Matthews, J., A. M. Schipper, A. J. Hendriks, T. T. Yen Le, A. bij de Vaate, G. van der Velde & R. S. E. W. Leuven, 2015. A dominance shift from the zebra mussel to the invasive quagga mussel may alter the trophic transfer of metals. Environmental Pollution 203: 183–190.
Mayer, C. M., L. G. Rudstam, E. L. Mills, S. G. Cardiff & C. A. Bloom, 2001. Zebra mussels (Dreissena polymorpha), habitat alteration, and yellow perch (Perca flavescens) foraging: system-wide effects and behavioural mechanisms. Canadian Journal of Fisheries and Aquatic Sciences 58: 2459–2467.
Mayer, C. M., R. A. Keats, L. G. Rudstam & E. L. Mills, 2002. Scale-dependent effects of zebra mussels on benthic invertebrates in a large eutrophic lake. Journal of the North American Benthological Society 21: 616–633.
Mayer, C. M., L. E. Burlakova, P. Eklöv, D. Fitzgerald, A. Y. Karatayev, S. A. Ludsin, S. Millard, E. L. Mills, A. P. Ostapenya, L. G. Rudstam, B. Zhu & T. V. Zhukova, 2014. The benthification of freshwater lakes: exotic mussels turning ecosystems upside down. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 575–586.
McCorquodale-Bauer, K. & N. Cicek, 2020. Zebra mussel shells as an alternative mineral resource for lime production as a phosphorus precipitant. Environmental Technology 22: 1–12.
McDonnell, E., 1996. Composting Zebra Mussels. Cornell Composting. Cornell Waste Management Institute, Department of Crop and Soil Sciences, Cornell University. http://compost.css.cornell.edu/zebra.html.
McLaughlan, C. & D. C. Aldridge, 2013. Cultivation of zebra mussels (Dreissena polymorpha) within their invaded range to improve water quality in reservoirs. Water Research 47: 4357–4369.
McLaughlan, C., P. Rose & D. C. Aldridge, 2014. Making the best of a pest: the potential for using invasive Zebra mussel (Dreissena polymorpha) biomass as a supplement to commercial chicken feed. Environmental Management 54: 1102–1109.
McMahon, R. F., 1991. Mollusca: Bivalvia. In Thorp, J. H. & A. P. Covich (eds), Ecology and classification of North American freshwater invertebrates Academic Press, New York: 315–399.
McMahon, R. F., 1996. The physiological ecology of the Zebra Mussel, Dreissena polymorpha, in North America and Europe. American Zoologist 36: 339–363.
McMahon, R. F. & A. F. Bogan, 2001. Mollusca: bivalvia. In Thorp, J. H. & A. P. Covich (eds), Ecology and classification of North American freshwater invertebrates Academic Press, New York: 331–430.
Meehan, S., B. Gruber & F. Lucy, 2014. Zebra mussel control using Zequanox® in an Irish waterway. Management of Biological Invasion 5: 279–286.
Mellina, E. & J. B. Rasmussen, 1994. Patterns in the distribution and abundance of zebra mussel (Dreissena polymorpha) in rivers and lakes in relation to substrate and other physicochemical factors. Canadian Journal of Fisheries and Aquatic Sciences 51: 1024–1036.
Mellina, E., J. B. Rasmussen & E. L. Mills, 1995. Impact of zebra mussel (Dreissena polymorpha) on phosphorus cycling and chlorophyll in lakes. Canadian Journal of Fisheries and Aquatic Sciences 52: 2553–2573.
Mercer, J. L., M. G. Fox & C. D. Metcalfe, 1999. Changes in benthos and three littoral zone fishes in a shallow, eutrophic Ontario Lake following the invasion of the zebra mussel (Dreissena polymorpha). Lake and Reservoir Management 15: 310–323.
Merritt, R. W., K. W. Cummins & T. M. Burton, 1984. The role of aquatic insects in the processing and cycling of nutrients. In Resh, V. H. & D. M. Rosenberg (eds), The Ecology of Aquatic Insects Praeger Scientific, New York: 134–163.
Metz, O., A. Temmen, K. C. M. von Oheimb, C. Albrecht, P. Schubert & T. Wilke, 2018. Invader vs. invader: intra- and interspecific competition mechanisms in zebra and quagga mussels. Aquatic Invasions 13(4): 473–480.
Mezek, T., E. Sverko, M. D. Rudy, D. Zaruk, A. Capretta, C. E. Hebert, A. T. Fisk, D. J. McGoldrick, T. J. Newton, T. M. Sutton, M. A. Koops, A. M. Muir, T. B. Johnson, M. P. Ebener & M. T. Arts, 2011. Polymethylene-interrupted fatty acids: Biomarkers for native and exotic mussels in the Laurentian Great Lakes. Journal of Great Lakes Research 37: 289–297.
Mezzanotte, V., F. Marazzi, M. Bissa, S. Pacchioni, A. Binelli, M. Parolini, S. Magni, F. M. Ruggeri, C. D. C. Morghen, C. Zanotto & A. Radaelli, 2016. Removal of enteric viruses and Escherichia coli from municipal treated effluent by zebra mussels. Science of the Total Environment 539: 395–400.
Micklin, P., N. V. Aladin, T. Chida, N. Boroffka, I. S. Plotnikov, S. Krivonogov & K. White, 2020. The Aral Sea: A story of devastation and partial recovery of a large lake. In: S. Mischke (ed) Large Asian Lakes in a Changing World Natural State and Human Impact. Springer Nature, Switzerland: 109–141.
Micu, D. & A. Telembici, 2004. First record of Dreissena bugensis (Andrusov 1897) from the Romanian stretch of River Danube. In Abstracts of the International Symposium of Malacology, Sibiu, Romania.
Mida, J. L., D. Scavia, G. L. Fahnenstiel, S. A. Pothoven, H. A. Vanderploeg & D. M. Dolan, 2010. Long–term and recent changes in southern Lake Michigan water quality with implications for present trophic status. Journal of Great Lakes Research 36(Suppl. 3): 42–49.
Mihuc, T. B., J. M. Battle, J. R. Mihuc & C. F. Bryan, 1999. Zebra mussel (Dreissena polymorpha) seasonal colonization patterns in a sub-tropical floodplain river. Hydrobiologia 392: 121–128.
Mikheev, V. P., 1964. Linear growth of Dreissena polymorpha Pallas in some reservoirs of the European part of the USSR. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 7(10): 55–65 (in Russian).
Mikheev, V. P., 1994. Composition and quantity of Dreissena food in natural conditions. In J. I. Starobogatov (ed). Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow: 129–132 (in Russian).
Mikulski, J., B. Adamczak, L. Bittel, R. Bohr, D. Bronisz, W. Donderski, A. Gizinski, M. Luscinska, M. Rejewski, E. Strzelczyk, N. Wolnomiejski, W. Zawislak & R. Zytkowicz, 1975. Basic regularities of productive processes in the Ilawa Lakes and in the Goplo Lake from the point of view of utility values of the water. Polish Archives of Hydrobiology 22: 101–122.
Millane, M., M. O’Grady, K. Delanty & M. Kelly-Quinn, 2012. An assessment of fish predation on the zebra mussel, Dreissena polymorpha (Pallas 1771) after recent colonisation of two managed brown trout lake fisheries in Ireland. Biology and Environment: Proceedings of the Royal Irish Academy 112B: 1–9.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-being. Island Press, Washington, DC. https://www.millenniumassessment.org/en/index.html
Mills, E. L., R. M. Dermott, E. F. Roseman, D. Dustin, E. Mellina, D. B. Corm & A. P. Spidle, 1993. Colonization, ecology, and population structure of the “quagga” mussel (Bivalvia: Dreissenidae) in the lower Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 50: 2305–2314.
Mills, E. L., R. O’Gorman, E. F. Roseman, C. Adams & R. W. Owens, 1995. Planktivory by alewife (Alosa pseudoharengus) and rainbow smelt (Osmerus mordax) on microcrustacean zooplankton and dreissenid (Bivalvia: Dreissenidae) veligers in southern Lake Ontario. Canadian Journal of Fisheries and Aquatic Sciences 52: 925–935.
Mills, E. L., G. Rosenberg, A. P. Spidle, M. Ludyansky & Y. Pligin, 1996. A review of the biology and ecology of the quagga mussel (Dreissena bugensis), a second species of freshwater dreissenid introduced to North America. American Zoologist 36: 271–286.
Minchin, D., 2000. Dispersal of zebra mussel in Ireland. Verhandlungen Des Internationalen Verein Limnologie 27: 1576–1579.
Minchin, D., F. Lucy & M. Sullivan, 2002. Zebra mussel: impacts and spread. In Leppakoski, E., S. Gollach & S. Olenin (eds), Invasive Aquatic Species of Europe: Distribution, Impacts and Management Kluwer Academic Publishers, Dordreicht: 135–146.
Miroshnichenko, M. P., 1987. Molluscs of the Tsimlyanskoe Reservoir and their role in increasing fish productivity. Sbornik Nauchnykh Trudov. Gosudarstvennyii Nauchno-Issledovatel’skii Institut Ozernogo i Rechnogo Rybnogo Khozyaistva 270: 61–70 (in Russian).
Miroshnichenko, M. P., 1990. Growth, production and practical use of Dreissena polymorpha (Pallas) in food web of Tsimlyanskoe Reservoir. In Khmeleva, N. N., A. P. Golubev, N. Y. Sokolova & V. E. Roschin (eds), Species Within their Range: Biology, Ecology, and Productivity of Aquatic Invertebrates Navuka i Tekhnika Press, Minsk, Belarus: 141–146. (in Russian).
Miroshnichenko, M. P., L. I. Volvich & B. O. Skabichevskii, 1984. Production and energetic balance of the most abundant molluscs in Tsimlyanskoe Reservoir. Sbornik Nauchnykh Trudov. Gosudarstvennyii Nauchno-Issledovatel’skii Institut Ozernogo i Rechnogo Rybnogo Khozyaistva 218: 19–30 (in Russian).
Mitchell, C. A. & J. Carlson, 1993. Lesser scaup forage on zebra mussels at Cook Nuclear Plant, Michigan. Journal of Field Ornithology 64: 219–222.
Mitrakhovich, P. A. & A. Y. Karatayev, 1986. The role of Dreissena polymorpha Pallas larvae in zooplankton of the cooling reservoir of the Lukoml thermal power plant. Vestnik Belorusskogo Universiteta, Ser. 2(1): 38–41 (in Russian).
Mitrakhovich, P. A., V. M. Samoilenko, Z. K. Kartashevich, A. A. Svirid, E. A. Kozlov, G. N. Korolev & N. A. Papko, 2008. Ecosystem of Lukoml Thermoelectric Station Cooling Reservoir. Pravo and Economica Press, Minsk, Belarus: 144 pp. (in Russian).
Molloy, D. P., 2002. Biological control of zebra mussels. In Proceedings of the Third California Conference on Biological Control. University of California, Davis: 86–94.
Molloy, D. P., J. Powell & P. Ambrose, 1994. Short-term reduction of adult zebra mussels (Dreissena polymorpha) in the Hudson River near Catskill, New York: An effect of juvenile blue crab (Callinectes sapidus) predation? Journal of Shellfish Research 13: 367–371.
Molloy, D. P., A. Y. Karatayev, L. E. Burlakova, D. P. Kurandina & F. Laruelle, 1997. Natural enemies of zebra mussels: predators, parasites, and ecological competitors. Reviews in Fisheries Science 5: 27–97.
Molloy, D. P., D. H. Lynn & L. Giamberini, 2005. Ophryoglena hemophaga n. sp. (Ciliophora: Ophryoglenidae): A parasite of the digestive gland of zebra mussels Dreissena polymorpha. Diseases of Aquatic Organisms 65: 237–243.
Molloy, D. P., A. bij de Vaate, T. Wilke & L. Giamberini, 2007. Discovery of Dreissena rostriformis bugensis (Andrusov 1897) in Western Europe. Biological Invasions 9: 871–874.
Molloy, D. P., L. Giamberini, L. E. Burlakova, A. Y. Karatayev, J. R. Cryan, S. L. Trajanovski & S. P. Trajanovska, 2010. Investigation of the endosymbionts of Dreissena stankovici with morphological and molecular confirmation of host species. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 227–237.
Molloy, D. P., L. Giamberini, N. A. Stokes, E. M. Burreson & M. A. Ovcharenko, 2012. Haplosporidium raabei n. sp. (Haplosporidia): a parasite of zebra mussels, Dreissena polymorpha (Pallas, 1771). Parasitology 139(4): 463–477.
Molloy, D. P., D. A. Mayer, L. Giamberini & M. J. Gaylo, 2013a. Mode of action of Pseudomonas fluorescens strain CL145A, a lethal control agent of dreissenid mussels (Bivalvia: Dreissenidae). Journal of Invertebrate Pathology 113: 115–121.
Molloy, D., D. Mayer, M. Gaylo, L. Burlakova, A. Karatayev, K. Presti, P. Sawyko, J. Morse & E. Paul, 2013b. Non-target trials with Pseudomonas fluorescens strain CL145A, a lethal control agent of dreissenid mussels (Bivalvia: Dreissenidae). Management of Biological Invasions 4: 71–79.
Mordukhai-Boltovskoi, F. D., 1960. Caspian Fauna in the Azov and Black Sea Basins. Moscow: Academia Nauk Press: 286 pp. (in Russian).
Mordukhai-Boltovskoi, F. D., 1972. The present state of the Aral Sea fauna. Hydrobiological Journal 8(3): 14–20 (in Russian).
Morrison, T. W., W. E. Lynch Jr. & K. Dabrowski, 1997. Predation on zebra mussels by freshwater drum and yellow perch in western Lake Erie. Journal of Great Lakes Research 23: 177–189.
Morse, J. T., 2009. Assessing the effects of application time and temperature on the efficacy of hot-water sprays to mitigate fouling by Dreissena polymorpha (zebra mussels Pallas). Biofouling 25: 605–610.
Mörtl, M., S. Werner & K. O. Rothhaupt, 2010. Effects of predation by wintering water birds on zebra mussels and on associated macroinvertebrates. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 239–249.
Morton, B., 1969a. Studies on the biology of Dreissena polymorpha Pall. III. Population dynamics. Proceedings of the Malacological Society of London 38: 471–481.
Morton, B., 1969b. Studies on the biology of Dreissena polymorpha Pall. II. Correlation of the rhythms of adductor activity, feeding, digestion and excretion. Proceedings of the Malacological Society of London 38: 401–414.
Morton, B. S., 1970. The evolution of the heteromyarian condition in the Dreissenacea (Bivalvia). Palaeontology 13: 563–572.
Morton, B. S., 1993. The anatomy of Dreissena polymorpha and the evolution and success of the heteromyarian form in the Dreissenoidea. In Nalepa, T. F. & D. W. Schloesser (eds), Zebra Mussels: Biology, Impacts, and Control Lewis Publishers, Ann Arbor, MI: 185–212.
Morton, B. S., 1996. The evolutionary history of the Bivalvia. In Taylor, J. (ed), Origin and Evolutionary Radiation in the Mollusca Oxford University Press, New York: 337–359.
Mosley, C. & H. Bootsma, 2015. Phosphorus recycling by profunda quagga mussels (Dreissena rostriformis bugensis) in Lake Michigan. Journal of Great Lakes Research 41: 38–48.
Müller, J. C., D. Hidde & A. Seitz, 2002. Canal construction destroys the barrier between major European invasion lineages of the zebra mussel. Proceeding of the Royal Society B. Biological Sciences 269: 1139–1142.
Nack, C. C., K. E. Limburg & R. E. Schmidt, 2015. Diet composition and feeding behaviour of larval American Shad, Alosa sapidissima (Wilson), after the introduction of the invasive zebra mussel, Dreissena polymorpha (Pallas), in the Hudson River Estuary, NY. Northeastern Naturalist 22: 437–450.
Naddafi, R. & L. G. Rudstam, 2013. Predator induced behavioural defences in two competitive invasive species. Animal Behavior 86: 1275–1284.
Naddafi, R. & L. G. Rudstam, 2014. Predator-induced morphological defences in two invasive dreissenid mussels: implications for species replacement. Freshwater Biology 59: 703–713.
Naddafi, R., P. Eklöv & K. Pettersson, 2009. Stoichiometric constraints do not limit successful invaders: zebra mussels in Swedish lakes. PLoS ONE 4(4): e5345.
Nalepa, T. F., 1994. Decline of native unionid bivalves in Lake St. Clair after infestation by the zebra mussel, Dreissena polymorpha. Canadian Journal of Fisheries and Aquatic Sciences 51: 2227–2233.
Nalepa, T. F. & D. W. Schloesser, 1993. Zebra Mussels Biology, Impacts, and Control, Lewis Publishers, Ann Arbor, Michigan, USA:, 810.
Nalepa, T. F. & D. W. Schloesser, 2014. Quagga and Zebra Mussels: Biology, Impacts, and Control, 2nd ed. CRC Press, Boca Raton, Florida, USA: 775.
Nalepa, T. E., J. A. Wojcik, D. L. Fanslow & G. A. Lang, 1995. Initial colonization of the zebra mussel (Dreissena polymorpha) in Saginaw Bay, Lake Huron: population recruitment, density and size structure. Journal of Great Lakes Research 21: 417–434.
Nalepa, T. E., D. J. Hartson, G. W. Gostenik, D. L. Fanslow & G. A. Lang, 1996. Changes in the freshwater mussel community of Lake St. Clair: from Unionidae to Dreissena polymorpha in eight years. Journal of Great Lakes Research 22: 354–369.
Nalepa, T. F., D. L. Fanslow, M. B. Lansing & G. A. Lang, 2003. Trends in the benthic macroinvertebrate community of Saginaw Bay, Lake Huron, 1987 to 1996: responses to phosphorus abatement and the zebra mussel, Dreissena polymorpha. Journal of Great Lakes Research 29: 14–33.
Nalepa, T. F., D. L. Fanslow, A. J. Foley III., G. A. Lang, B. J. Eadie & M. A. Quigley, 2006. Continued disappearance of the benthic amphipod Diporeia spp. in Lake Michigan: is there evidence for food limitation? Canadian Journal of Fisheries and Aquatic Sciences 63: 872–890.
Nalepa, T. F., D. L. Fanslow, S. A. Pothoven, A. J. Foiley III. & G. A. Lang, 2007. Long-term trends in benthic macroinvertebrate populations in Lake Huron over the past four decades. Journal of Great Lakes Research 33: 42–436.
Nalepa, T. F., D. L. Fanslow & G. A. Lang, 2009a. Transformation of the offshore benthic community in Lake Michigan: recent shift from the native amphipod Diporeia spp. to invasive mussel Dreissena rostriformis bugensis. Freshwater Biology 54: 466–479.
Nalepa, T. F., S. A. Pothoven & D. L. Fanslow, 2009b. Recent changes in benthic macroinvertebrate populations in Lake Huron and impact on the diet of lake whitefish (Coregonus clupeaformus). Aquatic Ecosystem Health and Management 12: 2–10.
Nalepa, T. F., D. L. Fanslow & S. A. Pothoven, 2010. Recent changes in density, biomass, recruitment, size structure, and nutritional state of Dreissena populations in southern Lake Michigan. Journal of Great Lakes Research 36: 5–19.
Nalepa, T. F., D. L. Fanslow, G. A. Lang, K. Mabrey & M. Rowe, 2014. Lake-wide benthic surveys in Lake Michigan in 1994–95, 2000, 2005, and 2010: Abundances of the amphipod Diporea spp. and abundances and biomass of the mussels Dreissena polymorpha and Dreissena rostriformis bugensis. NOAA Technical Memorandum GLERL-164: 20 pp.
Nalepa, T. F., L. E. Burlakova, A. K. Elgin, A. Y. Karatayev, G. A. Lang & K. Mehler, 2020. NOAA Tech Memo GLERL-175. Abundance and Biomass of Benthic Macroinvertebrates in Lake Michigan in 2015, With a Summary of Temporal Trends. Technical Memorandum, NOAA Great Lakes Environmental Research Laboratory, Ann Arbor Michigan, USA: 41 pp.
Neumann, D. & H. A. Jenner, 1992. Studies on the ecology and ecotoxicology of the zebra mussel Dreissena polymorpha. In Neumann, D. & H. A. Jenner (eds), The Zebra Mussel Dreissena polymorpha Ecology, Biology, Monitoring and First Applications in the Water Quality Management Gustav Fischer, New York: 1–4.
Neumann, D., J. Borcherding & B. Jantz, 1993. Growth and seasonal reproduction of Dreissena polymorpha in the Rhine River and adjacent waters. In Nalepa, T. F. & D. W. Schloesser (eds), Zebra Mussels: Biology, Impacts, and Control Lewis Publishers, Ann Arbor, MI: 95–109.
Nichols, S. J., 1993. Spawning of zebra mussels (Dreissena polymorpha) and rearing of the veligers under laboratory conditions. In Nalepa, T. F. & D. W. Schloesser (eds), Zebra Mussels: Biology, Impacts, and Control Lewis Publishers, Ann Arbor, MI: 315–329.
Nichols, S. J. & M. G. Black, 1994. Identification of larvae: the zebra mussel (Dreissena polymorpha), quagga mussel (Dreissena rostriformis bugensis), and Asian clam (Corbicula fluminea). Canadian Journal of Zoology 72: 406–417.
Nichols, S. J. & D. Wilcox, 1997a. Coexistence of zebra mussels and native clams in a lake coastal wetland. Journal of Shellfish Research 16: 346.
Nichols, S. J. & D. Wilcox, 1997b. Burrowing saves Lake Erie clams. Nature 389: 921–921.
Noordhuis, R. H., H. Reeders & A. bij de Vaate, 1992. Filtration rate and pseudofeces production in zebra mussels and their application in water quality management. In Neumann, D. & H. A. Jenner (ed), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management. Gustav Fisher, Stuttgart, Germany: 101–114.
Noordhuis, R., B. G. van Zuidam, E. T. H. M. Peeters & G. J. van Geestet, 2016. Further improvements in water quality of the Dutch Border lakes: two types of clear states at different nutrient levels. Aquatic Ecology 50: 521–539.
Nuttall, C. P., 1990. Review of the Caenozoic heterodont bivalve superfamily Dreissenacea. Palaeontology (London) 33: 707–737.
Office of Technology Assessment, 1993. Harmful non-indigenous species in the United States (Publication OTA-F-565). Washington, DC: United States Congress. http://www.princeton.edu/~ota/disk1/1993/9325/9325.pdf.
Olney, P., 1963. The food and feeding habits of tufted duck Aythya fuligula. Ibis 105: 55–62.
O’Neill, C. R., Jr., 1996. The zebra mussel: impacts and control. Cornell Cooperative Extension Information Bulletin 238: 62.
O’Neill, C. R., Jr., 1997. Economic impact of zebra mussels – Results of the 1995 National Zebra Mussel Information Clearinghouse study. Great Lakes Research Review 3: 35–44.
O’Neill, C.R. Jr., 2008. The Silent Invasion: Finding Solutions to Minimize the Impacts of Invasive Quagga Mussels on Water Rates, Water Infrastructure and the Environment. Hearing of the U.S. House of Representatives Committee on Natural Resources—Subcommittee on Water and Power, Washington, D.C. Available at: http://naturalresources.house.gov/uploadedfiles/oneilltestimony06.24.08.pdf.
Orlova, M. I., 2014. Origin and spread of quagga mussels (Dreissena rostriformis bugensis) in eastern Europe with notes on size structure of populations. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 93–102.
Orlova, M. I. & G. K. Shcherbina, 2001. Dreissena bugensis (Andr.) (Dreissenidae, Bivalvia): range expansion in Europe, invasion history, patterns and future invasion perspectives. In William, P. G. et al. (eds), Abstracts of the U.S.–Russia invasive species workshop. Yaroslavl’ State Technical University Press, Yaroslavl, Russia: 150–152.
Orlova, M. I. & V. E. Panov, 2004. Establishment of the Zebra Mussel, Dreissena polymorpha (Pallas), in the Neva Estuary (Gulf of Finland, Baltic Sea): distribution, population structure and possible impact on local unionid bivalves. Hydrobiologia 514(1–3): 207–217.
Orlova, M. I., J. R. Muirhead, P. I. Antonov, G. K. Shcherbina, Y. I. Starobogatov, G. I. Biochino, T. W. Therriault & H. J. MacIsaac, 2004. Range expansion of quagga mussels Dreissena rostriformis bugensis in the Volga River and Caspian Sea basin. Aquatic Ecology 38: 561–573.
Orlova, M. I., T. W. Therriault, P. I. Antonov & G. K. Shcherbina, 2005. Invasion ecology of quagga mussels (Dreissena rostriformis bugensis): a review of evolutionary and phylogenetic impacts. Aquatic Ecology 39: 401–418.
Ozersky, T., D. R. Barton & D. O. Evans, 2011. Fourteen years of dreissenid presence in the rocky littoral zone of a large lake: effects on macroinvertebrate abundance and diversity. Journal of the North American Benthological Society 30: 913–922.
Ozersky, T., D. O. Evans & B. K. Ginn, 2015. Invasive mussels modify the cycling, storage and distribution of nutrients and carbon in a large lake. Freshwater Biology 60: 827–843.
Padilla, D. K., M. A. Chotkowski & L. A. J. Buchan, 1996. Predicting the spread of zebra mussels (Dreissena polymorpha) to inland waters using boater movement patterns. Global Ecology and Biogeography Letters 5: 353–359.
Paldavičiene, A., A. Zaiko, H. Mazur-Marzec & A. Razinkovas-Baziukas, 2015. Bioaccumulation of microcystins in invasive bivalves: a case study from the boreal lagoon ecosystem. Oceanologia 57: 93–101.
Pallas, P. S., 1771. Reise durch verschiedene Provinzen des Russischen Reichs. Theil 1. Physicalische Reise durch verschiedene Provinzen des Russischen Reichs im 1768- und 1769 sten Jahren. Kayserliche Akademie der Wissenschaften. St. Petersburg: 505 pp. (in German).
Pallas, P. S., 1773. Travels Through Various Provinces of the Russian Empire. St. Petersburg. Part 1. Supplement.
Park, J.-K. & D. Ó. Foighil, 2000. Sphaeriid and corbiculid clams represent separate heterodont bivalve radiations into freshwater environments. Molecular Phylogenetics and Evolution 14: 75–88.
Passamaneck, Y. J., 2018. Literature review and synthesis of invasive mussel control techniques. U.S. Department of the Interior Bureau of Reclamation Research and Development Office Science and Technology Program Final Report ST-2018-1868-01: 43 pp.
Patterson, J. C. & P. V. Lindeman, 2009. Effects of Zebra and Quagga Mussel (Dreissena spp.) invasion on the feeding habits of Sternotherus odoratus (Stinkpot) on Presque Isle. Northwestern Pennsylvania. Northeastern Naturalist 16: 365–374.
Patterson, M. W. R., J. J. H. Ciborowski & D. R. Barton, 2005. The distribution and abundance of Dreissena species (Dreissenidae) in Lake Erie, 2002. Journal of Great Lakes Research 31(Suppl. 2): 223–237.
Paukstis, G. L., F. J. Janzen & J. K. Tucker, 1996. Response of aerially-exposed zebra mussels (Dreissena polymorpha) to subfreezing temperatures. Journal of Freshwater Ecology 11: 513–519.
Pedroli, J.-C., 1981. La phénologie des Fuligules hivernants sur le lac de Neuchâtel. Nos Oiseaux 36: 157–163.
Pejchar, L. & H. A. Mooney, 2009. Invasive species, ecosystem services and human well-being. Trends in Ecology & Evolution 24: 497–504.
Perez-Fuentetaja, A., S. A. Mackintosh, L. R. Zimmerman, M. D. Clapsadl, M. Alaee & D. S. Aga, 2015. Trophic transfer of flame retardants (PBDEs) in the food web of Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 72: 1886–1896.
Pergl, J., P. Pyšek, F. Essl, J. M. Jeschke, F. Courchamp, J. Geist, M. Hejda, I. Kowarik, A. Mill, C. Musseau, P. Pipek, W.-C. Saul, M. von Schmalensee & D. Strayer, 2020. Need for routine tracking of biological invasions. Conservation Biology 34: 1311–1314.
Perrings, C., M. Williamson & S. Dalmazzone (eds), 2001. The Economics of Biological Invasions. Edward Elgar, Northampton (USA).
Petrie, S. & R. Knapton, 1999. Rapid increase and subsequent decline of zebra and quagga mussels in Long Point Bay, Lake Erie: possible influence of waterfowl predation. Journal of Great Lakes Research 25: 772–782.
Peyer, S. M., A. J. McCarthy & C. E. Lee, 2009. Zebra mussels anchor byssal threads faster and tighter than quagga mussels in flow. Journal of Experimental Biology 212: 2026–2035.
Peyer, S. M., J. C. Hermanson & C. E. Lee, 2010. Developmental plasticity of shell morphology of quagga mussels from shallow and deep-water habitats of the Great Lakes. Journal of Experimental Biology 213: 2602–2609.
Piesik, Z., 1974. The role of the crayfish Orconectes limosus (Raf.) in extinction of Dreissena polymorpha (Pall.) subsisting on steelon net. Polish Archives of Hydrobiology 21: 401–410.
Piesik, Z., 1983. Biology of Dreissena polymorpha (Pall.) settling on stylon nets and the role of this mollusc in eliminating the seston and the nutrients from the watercourse. Polish Archives of Hydrobiology 30: 353–361.
Pimentel, D., 2005. Aquatic nuisance species in the New York State Canal and Hudson River systems and the Great Lakes Basin: an economic and environmental assessment. Environmental Management 35: 692–701.
Pimentel, D. (ed), 2011. Biological invasions. Economic and environmental costs of alien plant, animal, and microbe species (2nd Edition). CRC Press, Boca Raton, Florida, USA: 1–449.
Pligin, Y. V., 1979. Habitat extension of Dreissena bugensis Andr. In Likharev, I. M. (ed), Molluscs: Main Results of Their Study Nauka Press, Leningrad, Abstracts of the Sixth Meeting on the Investigation of Molluscs: 222–224 (in Russian).
Poddubnyi, A. G., 1966. A note on an adaptive response of a roach population to a change in environmental conditions. Proceedings of Institute of Biology of Inland Waters of Academy of Science of the USSR 10: 131–138 (in Russian).
Pollux, B., D. Minchin, G. Van Der Velde, T. Van Alen, S. Moon-Van Der Staay & J. Hackstein, 2003. Zebra mussels (Dreissena polymorpha) in Ireland, AFLP-fingerprinting and boat traffic both indicate an origin from Britain. Freshwater Biology 48: 1127–1139.
Pollux, B. J. A., G. van der Velde & A. biji de Vaate, 2010. A perspective on global spread of Dreissena polymorpha: a review on possibilities and limitations. In van der Velde, G., S. Rajagopal & A. biji de Vaate (eds), The Zebra Mussel in Europe Backhuys Publishers, Leiden: 45–58.
Ponyi, J. E., 1992. The distribution and biomass of Unionidae (Mollusca, Bivalvia), and the production of Unio tumidus Retzius in Lake Balaton (Hungary). Archiv Für Hydrobiologie 125: 245–251.
Pothoven, S. A. & C. P. Madenjian, 2008. Changes in consumption by alewives and lake whitefish after dreissenid mussel invasions in Lake Michigan and Huron. North American Journal of Fisheries Management 28: 308–320.
Pothoven, S. A. & G. L. Fahnenstiel, 2014. Lake Michigan after dreissenid mussel invasion. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton, FL: 545–553.
Pothoven, S. A. & A. K. Elgin, 2019. Dreissenid veliger dynamics along a nearshore to offshore transect in Lake Michigan. Journal of Great Lakes Research 45: 300–306.
Pothoven, S. A. & H. A. Vanderploeg, 2020. Seasonal patterns for Secchi depth, chlorophyll a, total phosphorus, and nutrient limitation differ between nearshore and offshore in Lake Michigan. Journal of Great Lakes Research 46: 519–527.
Pothoven, S. A., T. Nalepa, P. Schneeberger & S. Brandt, 2001. Changes in diet and body condition of lake whitefish in southern Lake Michigan associated with changes in benthos. North American Journal of Fisheries Management 21: 876–883.
Pothoven, S. A., C. P. Madenjian & T. O. Hook, 2017. Feeding ecology of the walleye (Percidae, Sander vitreus), a resurgent piscivore in Lake Huron (Laurentian Great Lakes) after shifts in the prey community. Ecology of Freshwater Fish 26: 676–685.
Prejs, A., 1976. Fishes and their feeding habits. In Pieczynska, E. (ed), Selected Problems of Lake Littoral Ecology. University of Warsaw Press, Warsaw, Poland: 155–171.
Props, R., M. L. Schmidt, J. Heyse, H. A. Vanderploeg, N. Boon & V. J. Denef, 2018. Flow cytometric monitoring of bacterioplankton phenotypic diversity predicts high population-specific feeding rates by invasive dreissenid mussels. Environmental Microbiology 20: 521–534.
Protasov, A. A., K. D. Starodub & S. A. Afanasiev, 1983. Species composition of zooperiphyton in the cooling reservoir of the Chernobyl Atomic Power Station. Hydrobiological Journal 19(2): 51–55 (in Russian).
Protasov, A. A., S. A. Afanasjev, O. O. Sinicyna & B. Zdanowski, 1994. Composition and functioning of benthic communities. Archives of Polish Fisheries 2: 257–284.
Rajagopal, S., G. van der Velde & H. A. Jenner, 2010. Turning the heat on Dreissena polymorpha: temperature as a control option. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds), The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 363–370.
Rakovic, M., N. Popovic, V. Kalafatic & V. Martinovic-Vitanovic, 2013. Spreading of Dreissena rostriformis bugensis (Andrusov, 1897) in the Danube River (Serbia). Acta Zoologica Bulgarica 65(3): 349–357.
Ram, J. L., P. F. Fong & D. W. Garton, 1996. Physiological aspects of zebra mussel reproduction: maturation, spawning, and fertilization. American Zoologist 36: 326–338.
Ramcharan, C. W., D. K. Padilla & S. I. Dodson, 1992. Models to predict potential occurrence and density of the zebra mussel, Dreissena polymorpha. Canadian Journal of Fisheries and Aquatic Sciences 49: 2611–2620.
Rebelo, M. F., L. F. Afonso, J. A. Americo, L. da Silva, J. L. B. Neto, F. Dondero, A. Crisanti & Q. Zhang, 2018. A sustainable synthetic biology approach for the control of the invasive golden mussel (Limnoperna fortunei). PeerJ. https://doi.org/10.7287/peerj.preprints.27164v1.
Reed-Andersen, T., S. R. Carpenter, D. K. Padilla & R. C. Lathrop, 2000. Predicted impact of zebra mussel (Dreissena polymorpha) invasion on water clarity in Lake Mendota. Canadian Journal of Fisheries and Aquatic Sciences 57: 1617–1626.
Reeders, H. H. & A. bij de Vaate, 1990. Zebra mussels (Dreissena polymorpha): a new perspective for water quality management. Hydrobiologia 200/201: 437–450.
Reeders, H. H., A. bij de Vaate & F. J. Slim, 1989. The filtration rate of Dreissena polymorpha (Bivalvia) in three Dutch lakes with reference to biological water quality management. Freshwater Biology 22: 133–141.
Reeders, H. H., A. Vaate & R. Noordhuis, 1993. Potential of the zebra mussel (Dreissena polymorpha) for water quality management. In Nalepa, T. F. & D. W. Schloesser (eds), Zebra Mussels. Biology, Impacts, and Control Lewis Publishers, Ann Arbor, Michigan, USA: 439–451.
Reid, N. J., M. A. Anderson & W. D. Taylor, 2010. Distribution of quagga mussel veligers, Dreissena bugensis, in the reservoirs of the Colorado River Aqueduct. Lake and Reservoir Management 26: 328–335.
Rennie, M. D. & D. O. Evans, 2012. Decadal changes in benthic invertebrate biomass and community structure in Lake Simcoe. Freshwater Science 31: 733–749.
Rennie, M. D., W. G. Sprules & T. B. Johnson, 2009. Resource switching in fish following a major food web disruption. Oecologia 159: 789–802.
Reynolds, S. A. & D. C. Aldridge, 2021. Impacts of invasive quagga mussels (Dreissena rostriformis bugensis) on reservoir water quality, as revealed by progressive-change BACIPS analysis. Water Research 197: 117105.
Ricciardi, A. & H. J. MacIsaac, 2000. Recent mass invasion of the North American Great Lakes by Ponto-Caspian species. Trends in Ecology and Evolution 15: 62–65.
Ricciardi, A. & F. G. Whoriskey, 2004. Exotic species replacement: shifting dominance of dreissenid mussels in the Soulanges Canal, upper St. Lawrence River, Canada. Journal of the North American Benthological Society 23: 507–514.
Ricciardi, A., F. G. Whoriskey & J. B. Rasmussen, 1995. Predicting the intensity and impact of Dreissena infestation on native unionid bivalves from Dreissena field density. Canadian Journal of Fisheries and Aquatic Sciences 52: 1449–1461.
Ricciardi, A., F. G. Whoriskey & J. B. Rasmussen, 1996. Impact of the Dreissena invasion on native unionid bivalves in the upper St. Lawrence River. Canadian Journal of Fisheries and Aquatic Sciences 53: 1434–1444.
Roberts, L., 1990. Zebra mussel invasion threatens U.S. waters. Science 249: 1370–1372.
Robertson, A. & G. G. Lauenstein, 1998. Distribution of chlorinated organic contaminants in dreissenid mussels along the southern shores of the Great Lakes. Journal of Great Lakes Research 24: 608–619.
Roe, S. L. & H. J. MacIsaac, 1997. Deepwater population structure and reproductive state of quagga mussels (Dreissena bugensis) in Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 54: 2428–2433.
Roper, J. M., D. S. Cherry, J. W. Simmers & H. E. Tatem, 1996. Bioaccumulation of toxicants in the zebra mussel, Dreissena polymorpha, at the Times Beach Confined Disposal Facility, Buffalo, New York. Environmental Pollution 94: 117–129.
Rosenberg, G. & M. L. Ludyanskiy, 1994. A nomenclatural review of Dreissena (Bivalvia: Dreissenidae) with identification of the quagga mussel as Dreissena bugensis. Canadian Journal of Fisheries and Aquatic Sciences 51: 1474–1484.
Rowe, M. D., D. R. Obenour, T. F. Nalepa, H. A. Vanderploeg, F. Yousef & W. C. Kerfoot, 2015. Mapping the spatial distribution of the biomass and filter-feeding effect of invasive dreissenid mussels on the winter-spring phytoplankton bloom in Lake Michigan. Freshwater Biology 60: 2270–2285.
Rudstam, L. G. & C. J. Gandino, 2020. Zebra or quagga mussel dominance depends on trade-offs between growth and defense-Field support from Onondaga Lake, NY. PLoS ONE 15: e0235387.
Ruiz, G. & J. Carlton, 2003. Invasive Species: Vectors and Management Strategies, Island Press, Washington, DC:, 536.
Sablon, R., T. Vercauteren & P. Jacobs, 2010. De quaggamossel (Dreissena rostriformis bugensis (Andrusov, 1897)), een recent gevonden invasieve zoetwatermossel in Vlaanderen. Antenne 4: 32–36. (in Dutch).
Schaller, J. & B. Planer-Friedrich, 2017. The filter feeder Dreissena polymorpha affects nutrient, silicon, and metal(loid) mobilization from freshwater sediments. Chemosphere 174: 531–537.
Schernewski, G., N. Stybel & T. Neumann, 2012. Zebra mussel farming in the Szczecin (Oder) Lagoon: Water-quality objectives and cost-effectiveness. Ecology and Society 17: 4.
Schernewski, G., R. Friedland, A.-L. Buer, S. Dahlke, B. Drews, S. Höft, T. Klumpe, M. Schadach, J. Schumacher & A. Zaiko, 2019. Ecological-social-economic assessment of zebra-mussel cultivation scenarios for the Oder (Szczecin) Lagoon. Journal of Coastal Conservation 23: 913–929.
Schloesser, D. W. & A. bij de Vaate & A. Zimmerman, 1994. A bibliography of Dreissena polymorpha in European and Russian waters: 1964–1993. Journal of Shellfish Research 13: 243–267.
Schloesser, D. W. & C. Schmuckal, 2012. Bibliography of Dreissena polymorpha (zebra mussels) and Dreissena rostriformis bugensis (quagga mussels): 1989 to 2011. Journal of Shellfish Research 31: 1205–1263.
Schneider, D. W., S. P. Madon, J. A. Stoeckel & R. E. Sparks, 1998. Seston quality controls zebra mussel (Dreissena polymorpha) energetics in turbid rivers. Oecologia 117: 331–341.
Schneider, D. W., J. A. Stoeckel, C. R. Rehmann, K. D. Blodgett, R. E. Sparks & D. K. Padilla, 2003. A developmental bottleneck in dispersing larvae: implications for spatial population dynamics. Ecology Letters 6: 352–360.
Schummer, M. L., S. S. Badzinski, S. A. Petrie, Y. W. Chen & N. Belzile, 2010. Selenium accumulation in sea ducks wintering at Lake Ontario. Archives of Environmental Contamination and Toxicology 58: 854–862.
Schwab, A., U. Bornhauser-Sieber & V. Keller, 2001. Wintering waterbirds in the Lucerne part of Vierwaldstättersee (Switzerland) 1954/55-2000/01. Ornithologische Beobachter 98: 179–208.
Sebestyen, O., 1937. Colonization of two new fauna-elements of Pontus-origin (Dreissena polymorpha Pall. and Corophium curvispinum G. O. Sars forma devium Wundsch) in Lake Balaton. Verhandlungen Der Internationalen Vereinigung Für Theoretische Und Angewandte Limnologie 8: 169–182.
Selegean, J. P. & T. M. Heidtke, 1994. Helpful zebra mussels? Water Environment and Technology 6: 31–32.
Selegean, J. P. W., R. Kusserow, R. Patel, T. M. Heidtke & J. L. Ram, 2001. Using zebra mussels to monitor Escherichia coli in environmental waters. Journal of Environmental Quality 30: 171–179.
Shevtsova, L. V., 1968a. Distinctive features of the reproduction and development of Dreissena in the canal of the Dnieper-Krivoy Rog. Hydrobiological Journal 4(5): 70–72 (in Russian).
Shevtsova, L. V., 1968b. Animals in the periphyton and benthos of the canals of the south Ukraine. Summary of the Candidate Dissertation, Hydrobiological Institute of the Academy of Science of the Ukrainian SSR, Kiev: 202 pp. (in Russian).
Shevtsova, L. V., 2000. Establishment of populations of bivalve molluscs of the genus Dreissena in the Dniester River. In Starobogatov, Y. I. & E. N. Egorova (eds), Molluscs: Problems of Taxonomy, Ecology and Phylogeny Nauka Press, St. Petersburg: 86–88 (in Russian).
Shields, R. C. & D. W. Beckman, 2015. Assessment of variation in age, growth, and prey of freshwater drum (Aplodinotus grunniens) in the Lower Missouri River. Southwestern Naturalist 60: 360–365.
Shkorbatov, G. L., 1986a. Intraspecific differentiation and wholeness of a species as a system. In Kontrimavichus, V. L., et al., (eds), System of Species Integration Lithuanian SSR Press, Vilnius, Institute of Zoology and Parasitology Acad. Sci: 118–137 (in Russian).
Shkorbatov, G. L. & P. I. Antonov, 1986b. Study of the eco-physiological and eco-morphological variability of Dreissena polymorpha from waterbodies of the Volga cascade. In Braginskiy, L. P., et al., (eds), Abstracts of the Fifth Congress of the All-Union Hydrobiological Society, Vol. 1. Kuibyshev District Committee of the Communist Party Press, Kuibyshev: 178–180 (in Russian).
Shkorbatov, G. L., A. F. Karpevich & P. I. Antonov, 1994. Ecological physiology. In Starobogatov, J. I. (ed), Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow, USSR: 67–108 (in Russian).
Simberloff, D. & L. Gibbons, 2004. Now you see them, now you don’t! – population crashes of established introduced species. Biological Invasions 6: 161–172.
Sinitsina, O. O. & A. A. Protasov, 1993. Structural-functional peculiarities of periphyton as a criterion standardization of thermal action on cooling ponds. Hydrobiological Journal 29(5): 17–30 (in Russian).
Skalskaya, I. A., 1976a. The impact of heated water from the Kostromskaya Power Plant on zooperyphiton of artificial substrates. Summary of the Candidate Dissertation, Moscow State University, Moscow: 14 pp. (in Russian).
Skalskaya, I. A., 1976b. Colonization of new substrates in Gorkovskoe Reservoir by Dreissena polymorpha Pallas. Biologiya Vnutrennikh Vod. Informatsionnyi Bulletin. 31: 30–34 (in Russian).
Skubinna, J. P., T. G. Coon & T. R. Batters, 1995. Increased abundance and depth of submersed macrophytes in response to decreased turbidity in Saginaw Bay, Lake Huron. Journal of Great Lakes Research 21: 476–478.
Smit, H., A. biji de Vaate & A. Fioole, 1992. Shell growth of the zebra mussel Dreissena polymorpha Pallas in relation to selected physiochemical parameters in the Lower Rhine and some associated lakes. Archiv für Hydrobiologie 124(3): 257–280.
Smit, H., A. bij de Vaate, E. H. Van Nes & R. Noordhuis, 1993. Colonization, ecology, and positive aspects of zebra mussels (Dreissena polymorpha) in the Netherlands. In Nalepa, T. F. & D. W. Schloesser (ed), Zebra Mussels. Biology, Impacts, and Control. Lewis Publishers, Ann Arbor, Michigan, USA: 55–77.
Sokolova, N. Y., 1959. On periphyton fauna of reservoirs and lakes. Proceedings of VI Meeting on Problems on Biology of Inland Waters. Moscow, Academy of Sciences Press: 287–291.
Son, M. O., 2007. Native range of the zebra mussel and quagga mussel and new data on their invasions within the Ponto-Caspian region. Aquatic Invasions 2: 169–179.
Sousa, R., J. L. Gutierrez & D. C. Aldridge, 2009. Nonindigenous invasive bivalves as ecosystem engineers. Biological Invasions 11: 2367–2385.
Sousa, R., F. Pilotto & D. C. Aldridge, 2011. Fouling of European freshwater bivalves (Unionidae) by the invasive zebra mussel (Dreissena polymorpha). Freshwater Biology 56: 867–876.
Spiridonov, Y. I., 1972. Polarographic investigation of the respiration of Dreissena. In Topics in Physiological and Population Ecology. 2. Saratov University Press, Saratov: 15–21 (in Russian).
Sprecher, S. L. & K. D. Getsinger, 2000. Zebra mussel chemical control guide. ERDC/El Tr-00-1, U.S. Army Engineer Research and Development Center, Vicksburg, Mississippi, USA: 1–89.
Sprung, M., 1987. Ecological requirements of developing Dreissena polymorpha eggs. Archiv für Hydrobiologi—Supplements 79: 69–86.
Sprung, M., 1989. Field and laboratory observations of Dreissena polymorpha larvae: abundance, growth, mortality, and food demands. Archiv Für Hydrobiologie 115: 537–561.
Sprung, M., 1991. Costs of reproduction: a study on metabolic requirements of the gonads and fecundity of the bivalve Dreissena polymorpha. Malacologia 33: 63–70.
Sprung, M., 1992. Observation on shell growth and mortality of Dreissena polymorpha in Lakes. In Neumann, D. & H. A. Jenner (ed), The Zebra Mussel Dreissena polymorpha: Ecology, Biological Monitoring and First Applications in the Water Quality Management. Gustav Fisher, Stuttgart, Germany: 19–28.
Sprung, M., 1995a. Physiological energetics of the zebra mussel Dreissena polymorpha in lakes I. Growth and Reproductive Effort. Hydrobiologia 304: 117–132.
Sprung, M., 1995b. Physiological energetics of the zebra mussel Dreissena polymorpha in lakes. II. Food uptake and gross growth efficiency. Hydrobiologia 304(133–1): 46.
Stanczykowska, A., 1977. Ecology of Dreissena polymorpha (Pall.) (Bivalvia) in lakes. Polish Archives of Hydrobiology 24: 461–530.
Stanczykowska, A. & K. Lewandowski, 1993. Thirty years of studies of Dreissena polymorpha ecology in Mazurian Lakes of Northeastern Poland. In Nalepa, T. F. & D. W. Schloesser (ed), Zebra Mussels. Biology, Impacts, and Control. Lewis Publishers, Ann Arbor, Michigan, USA: 3–33.
Stanczykowska, A. & K. Lewandowski, 1995. Individual growth of the freshwater mussel Dreissena polymorpha (Pall.) in Mikolajskie Lake; estimates in situ. Ekologia Polska 43(3–4): 267–276.
Starobogatov, J. I., 1994a. (ed) Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow, USSR: 240 pp. (in Russian).
Starobogatov, J. I., 1994b. Systematics and paleontology. In Starobogatov, J. I. (ed), Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow, USSR: 18–46.
Starobogatov, Y. I., 1994c. Fauna and Zoogeography of Molluscs of Turkmenistan. In Fet, V. & K. I. Atamuradov (eds) Biogeography and Ecology of Turkmenistan. Monographiae Biologicae, Springer, Dordrecht 72: 535–543.
Starobogatov, J. I. & S. I. Andreeva, 1994. Distribution and history. In Starobogatov, J. I. (ed), Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Systematics, Ecology, Practical Meaning. Nauka Press, Moscow, USSR: 47–55.
Stempniewicz, L., 1974. The effect of feeding of the coot (Fulica atra L.) on the character of the shoals of Dreissena polymorpha Pall. in the Lake Goplo. Acta Universitatis Nicolai Copernici, Nauki Matematyczno-Przyrodnicze 34: 83–103.
Stepien, C. A., A. N. Hubers & J. L. Skidmore, 1999. Diagnostic genetic markers and evolutionary relationships among invasive dreissenoid and corbiculoid bivalves in North America: phylogenetic signal from mitochondrial 16S rDNA. Molecular Phylogenetics and Evolution 13: 31–49.
Stepien, C., B. Morton, K. Dabrowska, R. Guarnera, T. Radja & B. Radja, 2001. Genetic diversity and evolutionary relationships of the troglodytic “living fossil” Congeria kusceri (Bivalvia: Dreissenidae). Molecular Ecology 10: 1873–1879.
Stepien, C. A., C. D. Taylor, I. A. Grigorovich, S. V. Shirman, R. Wei, A. V. Korniushin & K. A. Dabrowska, 2003. DNA and systematic analysis of invasive and native dreissenid mussels: Is Dreissena bugensis really D. rostriformis?. Aquatic Invaders 14: 8–18.
Stepien, C. A., I. Grigorovich, M. Gray, T. Sullivan, S. Yerga-Woolwine & G. Kalayci, 2014. Evolutionary, biogeographic, and population genetic relationships of dreissenid mussels, with revision of component taxa. In Nalepa, T. F. & D. W. Schloesser (eds), Quagga and Zebra Mussels: Biology, Impacts, and Control 2nd ed. CRC Press, Boca Raton: 403–444.
Stewart, T. W., J. G. Miner & R. L. Lowe, 1998. Quantifying mechanisms for zebra mussel effects on benthic macroinvertebrates: Organic matter production and shell-generated habitat. Journal of the North American Benthological Society 17: 81–94.
Stoeckel, A., C. R. Rehmann, D. W. Schnieder & D. K. Padilla, 2004. Retention and supply of zebra mussel larvae in a large river system: importance of an upstream lake. Freshwater Biology 49: 919–930.
Stoeckmann, A., 2003. Physiological energetics of Lake Erie dreissenid mussels: a basis for the displacement of Dreissena polymorpha by Dreissena bugensis. Canadian Journal of Fisheries and Aquatic Sciences 60: 126–134.
Stoeckman, A. & D. Garton, 1997. A seasonal energy budget for zebra mussels (Dreissena polymorpha) in western Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 54: 2743–2751.
Strayer, D. L., 1991. Projected distribution of zebra mussel, Dreissena polymorpha in North America. Canadian Journal of Fisheries and Aquatic Sciences 48: 1389–1395.
Strayer, D. L., 1999. Effects of alien species on freshwater mollusks in North America. Journal of the North American Benthological Society 18: 74–98.
Strayer, D. L., 2012. Eight questions about invasions and ecosystem functioning. Ecology Letters 15: 1199–1210.
Strayer, D. L. & L. C. Smith, 1993. Distribution of zebra mussel (Dreissena polymorpha) in estuaries and brackish waters. In Nalepa, T. F. & D. W. Schloesser (ed), Zebra Mussels. Biology, Impacts, and Control. Lewis Publishers, Ann Arbor, Michigan, USA: 715–727.
Strayer, D. L. & H. M. Malcom, 2006. Long-term demography of a zebra mussel (Dreissena polymorpha) population. Freshwater Biology 51: 117–130.
Strayer, D. L. & H. M. Malcom, 2007. Effects of zebra mussels (Dreissena polymorpha) on native bivalves: the beginning of the end or the end of the beginning? Journal of the North American Benthological Society 26(1): 111–122.
Strayer, D. L., J. Powell, P. Ambrose, L. C. Smith, M. L. Pace & D. T. Fischer, 1996. Arrival, spread, and early dynamics of a zebra mussel (Dreissena polymorpha) population in the Hudson River estuary. Canadian Journal of Fisheries and Aquatic Science 53: 1143–1149.
Strayer, D. L., N. F. Caraco, J. J. Cole, S. Findlay & M. L. Pace, 1999. Transformation of freshwater ecosystems by bivalves—a case study of zebra mussels in the Hudson River. BioScience 49: 19–28.
Strayer, D. L., K. A. Hattala & A. W. Kahnle, 2004. Effects of an invasive bivalve (Dreissena polymorpha) on fish in the Hudson River estuary. Canadian Journal of Fisheries and Aquatic Science 61: 924–941.
Strayer, D. L., V. T. Eviner, J. M. Jeschke & M. L. Pace, 2006. Understanding the long-term effects of species invasions. Trends Ecology and Evolution. https://doi.org/10.1016/j.tree.2006.07.00.
Strayer, D. L., B. V. Adamovich, R. Adrian, D. C. Aldridge, C. Balogh, L. E. Burlakova, H. Fried-Petersen, L. G.-Tóth, A. L. Hetherington, T. S. Jones, A. Y. Karatayev, J. B. Madill, O. A. Makarevich, J. E. Marsden, A. L. Martel, D. Minchin, T. F. Nalepa, R. Noordhuis, T. J. Robinson, L. G. Rudstam, A. N. Schwalb, D. R. Smith, A. D. Steinman & J. M. Jeschke, 2019. Long-term population dynamics of dreissenid mussels (Dreissena polymorpha and D. rostriformis): a cross-system analysis. Ecosphere 10: e02701
Stumpf, P., K. Failing, T. Papp, J. Nazir, R. Böhm & R. E. Marschang, 2010. Accumulation of a low pathogenic avian influenza virus in zebra mussels (Dreissena polymorpha). Avian Diseases 54: 1183–1190.
Sturtevant, R. A., D. M. Mason, E. S. Rutherford, A. Elgin, E. Lower & F. Martinez, 2019. Recent history of nonindigenous species in the Laurentian Great Lakes; an update to Mills et al., 1993 (25 years later). Journal of Great Lakes Research 45: 1011–1035.
Stybel, N., C. Fenske & G. Schernewski, 2009. Mussel cultivation to improve water quality in the Szczecin Lagoon. Journal of Coastal Research, Special Issue 56: 1459–1463.
Summers, R. B., J. H. Thorp, J. E. Alexander & R. D. Fell, 1996. Respiratory adjustment of dreissenid mussels (Dreissena polymorpha and Dreissena bugensis) in response to chronic turbidity. Canadian Journal of Fisheries and Aquatic Science 53: 1626–1631.
Suter, W., 1982a. Die Bedeutung von Untersee-Ende/Hochrhein (Bodensee) als wichtiges Überwinterungsgewässer für Tauchenten Aythya, Bucephala und Bläßhuhn Fulica atra. Der Ornithologische Beobachter 79: 73–96.
Suter, W., 1982b. Vergleichende Nahrungsökologie von überwinternden Tauchenten Bucephala, Aythya und Bläßhuhn Fulica atra am Untersee-Ende/Hochrhein (Bodensee). Der Ornithologische Beobachter 79: 225–254.
Suter, W., 1994. Overwintering waterfowl on Swiss lakes: How are abundance and species richness influenced by trophic status and lake morphology? Hydrobiologia 279–280: 1–14.
Tang, H., H. A. Vanderploeg, T. H. Johengen & J. R. Liebig, 2014. Quagga mussel (Dreissena rostriformis bugensis) selective feeding of phytoplankton in Saginaw Bay. Journal of Great Lakes Research Supplement 40: 83–94.
Tellier, J. M., N. I. Kalejs, B. S. Leonhardt, D. Cannon, T. O. Hӧӧk & P. D. Collingsworth, 2022. Widespread prevalence of hypoxia and the classification of hypoxic conditions in the Laurentian Great Lakes. Journal of Great Lakes Research 48: 13–23.
Therriault, T. W., M. F. Docker, M. I. Orlova, D. D. Heath & H. J. MacIsaac, 2004. Molecular resolution of Dreissenidae (Mollusca: Bivalvia) including the first report of Mytilopsis leucophaeta in the Black Sea basin. Molecular Phylogenetics and Evolution 30: 479–489.
Therriault, T. W., M. I. Orlova, M. F. Docker, H. J. MacIsaac & D. D. Heath, 2005. Invasion genetics of a freshwater mussel (Dreissena rostriformis bugensis) in eastern Europe: high gene flow and multiple introductions. Heredity 95: 16–23.
Thomaz, S. M., 2021. Ecosystem services provided by freshwater macrophytes. Hydrobiologia. https://doi.org/10.1007/s10750-021-04739-y.
Thompson, K., 2014. Where do camels belong?: Why invasive species aren’t all bad. Profile Books, London, UK.
Thorp, J. H. & A. F. Casper, 2002. Potential effects on zooplankton from species shifts in planktivorous mussels: a field experiment in the St Lawrence River. Freshwater Biology 47(1): 107–119.
Thorp, J. H., M. D. Delong & A. F. Casper, 1998. In situ experiments on predatory regulation of a bivalve mollusc (Dreissena polymorpha) in the Mississippi and Ohio Rivers. Freshwater Biology 39: 649–661.
Turgeon, D. D., J. F. Quinn, Jr., A. E. Bogan, E. V. Coan, F. G. Hochberg, W. G. Lyons, P. M. Mikkelsen, R. J. Neves, C. F. E. Roper, G. Rosenberg, B. Roth, A. Scheltema, F. G. Thompson, M. Vecchione & J. D. Williams, 1998. Common and scientific names of aquatic invertebrates from the United States and Canada: Mollusks, 2nd Edition. American Fisheries Society Special Publication 26: 526 pp.
Turschak, B. A. & H. A. Bootsma, 2015. Lake Michigan trophic structure as revealed by stable C and N isotopes. Journal of Great Lakes Research 41: 185–196.
Van Beneden, P. J., 1835. Histoire naturelle et anatomique du Dreissena polymorpha, genre nouveau dans la familie de mytilacées [sic]. Bulletin De L’académie Royale Des Sciences, Des Lettres Et Des Beaux Arts De Belgique 2(25–26): 44–47.
van der Velde, G. & D. Platvoet, 2007. Quagga mussels (Dreissena rostriformis bugensis) (Andrusov, 1897) in Main River (Germany). Aquatic Invasions 2: 265–267.
van der Velde, G., S. Rajagopal & A. Bij de Vaate (eds.), 2010. The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands.
van der Velde, G., S. Rajagopal & A. bij de Vaate, 2010b. From zebra mussels to quagga mussels: an introduction to the Dreissenidae. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds). The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 1–10.
Vanderploeg, H. A., J. R. Liebig, W. W. Carmichael, M. A. Agy, T. H. Johengen, G. L. Fahnenstiel & T. F. Nalepa, 2001. Zebra mussel (Dreissena polymorpha) selective filtration promoted toxic Microcystis blooms in Saginaw Bay (Lake Huron) and Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 58: 1208–1221.
Vanderploeg, H. A., T. F. Nalepa, D. J. Jude, E. L. Mills, K. T. Holeck, J. R. Liebig, I. A. Grigorovich & H. Ojaveer, 2002. Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 59: 1209–1228.
Vanderploeg, H. A., T. H. Johengen & J. R. Liebig, 2009. Feedback between zebra mussel selective feeding and algal composition affects mussel condition: did the regime changer pay a price for its success? Freshwater Biology 54: 47–63.
Vanderploeg, H. A., J. R. Liebig, T. F. Nalepa, G. L. Fahnenstiel & A. Pothoven, 2010. Dreissena and the disappearance of the spring phytoplankton bloom in Lake Michigan. Journal of Great Lakes Research 36: 50–59.
Van Weelden, T. B. & B. C. Anderson, 2003. Phosphorus adsorption capacity of zebra mussel shells as a potential filter medium in constructed wetlands. Proceedings, Annual Conference—Canadian Society for Civil Engineering: 797–805.
Vaughn, C. C., 2017. Ecosystem services provided by freshwater mussels. Hydrobiologia 810: 15–27.
Vaughn, C. C. & T. J. Hoellein, 2018. Bivalve impacts in freshwater and marine ecosystems. Annual Review of Ecology, Evolution, and Systematics 49: 183–208.
Verween, A., M. Vincz & S. Degraer, 2010. Mytilopsis leucophaeta: The brackish water equivalent of Dreissena polymorpha? A review. In van der Velde, G., S. Rajagopal & A. bij de Vaate (eds). The Zebra Mussel in Europe. Backhuys Publishers, Leiden, The Netherlands: 29–43.
Vimercati, G., S. Kumschick, A. F. Probert, L. Volery & S. Bacher, 2020. The importance of assessing positive and beneficial impacts of alien species. NeoBiota 62: 525–545.
Vladimirov, M. Z., 1983. The influence of heated water from the Moldavian thermal power plant on composition, biotopical distribution and density of molluscs in Kuchurgankiy cooling-liman. In Likharev, I. M. (ed), Molluscs: Systematics, Ecology and Patterns of Occurrence. Abstracts of the Seventh Meeting on the Investigation of Molluscs. Nauka Press, Leningrad: 240–241 (in Russian).
Vorobiev, V. P., 1949. The Benthos of the Azov Sea, Krimizdat Press, Simpheropol (in Russian).
Waajen, G., N. C. B. Van Bruggen, L. M. D. Pires, W. Lengkeek & M. Lurling, 2016. Biomanipulation with quagga mussels (Dreissena rostriformis bugensis) to control harmful algal blooms in eutrophic urban ponds. Ecological Engineering 90: 141–150.
Wacker, A. & E. Von Elert, 2003. Settlement pattern of the zebra mussel, Dreissena polymorpha, as a function of depth in Lake Constance. Archiv Für Hydrobiologie 158: 289–301.
Walker, G. K., M. G. Black & C. A. Edwards, 1996. Comparative morphology of zebra (Dreissena polymorpha) and quagga (Dreissena bugensis) mussel sperm: light and electron microscopy. Canadian Journal of Zoology 74: 809–815.
Wallace, J. S. & D. M. Blersch, 2015. Dynamic modeling predicts continued bioaccumulation of polybrominated diphenyl ethers (PBDEs) in smallmouth bass (Micropterus dolomiu) post phase-out due to invasive prey and shifts in predation. Environmental Pollution 206: 289–297.
Waller, D. L. & M. R. Bartsch, 2018. Use of carbon dioxide in zebra mussel (Dreissena polymorpha) control and safety to a native freshwater mussel (Fatmucket, Lampsilis siliquoidea) Management of Biological Invasions 9(4): 439–450.
Walton, W. C., 1996. Occurrence of zebra mussel (Dreissena polymorpha) in the oligohaline Hudson River, New York. Estuaries 19(3): 612–618.
Walz, N., 1973. Untersuchungen zur Biologie von Dreissena polymorpha Pallas im Bodensee. Archiv für Hydrobiologie—Supplements 42: 452–482.
Walz, N., 1978. The energy balance of the freshwater mussel Dreissena polymorpha Pallas in laboratory experiments and in Lake Constance. II. Reproduction. Archiv für Hydrobiologie—Supplements 55: 106–119.
Walz, N., 1978. The energy balance of the freshwater mussel Dreissena polymopha Pallas in laboratory experiments and in Lake Constance. IV. Growth in Lake Constance. Archiv für Hydrobiologie—Supplements 55: 142–156.
Walz, N., 1978. The energy balance of the freshwater mussel Dreissena polymorpha Pallas in laboratory experiments and in Lake Constance. III. Growth under standard conditions. Archiv für Hydrobiologie—Supplements 55: 121–141.
Walz, N., 1978d. The production and significance of the Dreissena population in the nutrient cycle in Lake Constance. Archiv Für Hydrobiologie 82: 482–499.
Wang, J., K. R. Koopman, F. P. L. Collas, L. Posthuma, T. de Nijs, R. S. E. W. Leuven & A. J. Hendriks, 2021. Towards an ecosystem service-based method to quantify the filtration services of mussels under chemical exposure. Science of the Total Environment 763: 144196.
Ward, J. M. & A. Ricciardi, 2007. Impact of Dreissena invasions on benthic macroinvertebrate communities: a meta-analysis. Diversity and Distributions 13: 155–165.
Watkins, J. M., R. Dermott, S. J. Lozano, E. L. Mills, L. G. Rudstam & J. V. Scharold, 2007. Evidence for remote effects of dreissenids mussels on the amphipod Diporea: analysis of Lake Ontario benthic surveys, 1997–2003. Journal of Great Lakes Research 33: 642–657.
Watzin, M. C., K. Joppe-Mercure, J. Rowder, B. Lancaster & L. Bronson, 2008. Significant fish predation on zebra mussels Dreissena polymorpha in Lake Champlain, USA. Journal of Fish Biology 73: 1585–1599.
Wawrzyniak-Wydrowska, B., T. Radziejewska, A. Skrzypacz & A. Woz’niczka, 2019. Two non-indigenous dreissenids (Dreissena polymorpha and D. rostriformis bugensis) in a southern Baltic coastal lagoon: variability in populations of the “Old” and a “New” immigrant. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2019.00076.
Weber, M. J., J. M. Dettmers, D. H. Wahl & S. J. Czesny, 2011. Size preferences and behaviors of native yellow perch foraging on invasive round gobies. Journal of Great Lakes Research 37: 584–587.
Wegner, B., A. L. Kronsbein, M. Gillefalk, K. van de Weyer, J. Köhler, E. Funke, M. T. Monaghan & S. Hilt, 2019. Mutual facilitation among invading nuttall’s waterweed and quagga mussels. Frontiers in Plant Science. https://doi.org/10.3389/fpls.2019.00789.
Werner, S., M. Mortl, H. G. Bauer & K. O. Rothhaupt, 2005. Strong impact of wintering waterbirds on zebra mussel (Dreissena polymorpha) populations at Lake Constance, Germany. Freshwater Biology 50: 1412–1426.
Whitten, A. L., J. R. M. Jarrin & A. S. McNaught, 2018. A mesocosm investigation of the effects of quagga mussels (Dreissena rostriformis bugensis) on Lake Michigan zooplankton assemblages. Journal of Great Lakes Research 44: 105–113.
Wiktor, K., 1958. Larvae of Dreissena polymorpha Pall. as a food for fish spawn. Przegląd Zoologiczny 2: 182–184.
Wiktor, K., 1963. Research on the ecology of Dreissena polymorpha Pall. in the Szczecin Lagoon (Zatew Szczecinski). Ekologiya Polska, Seria A 11: 275–280.
Williams, K., 2020. Invasive mussels clear the water and coat the wrecks at Thunder Bay National Marine Sanctuary. Great Lakes Echo, February 27, 2020. https://greatlakesecho.org/2020/02/27/invasive-mussels-clear-the-water-and-coat-the-wrecks-at-thunder-bay-national-marine-sanctuary/.
Wimbush, J., M. Frischer, J. Zarzynski & S. Nierzwicki-Bauer, 2009. Eradication of colonizing populations of zebra mussels (Dreissena polymorpha) by early detection and SCUBA removal: Lake George, NY. Aquatic Conservation: Marine and Freshwater Ecosystems 19: 703–713.
Winkler, G., C. Martineau, J. J. Dodson, W. F. Vincent & L. E. Johnson, 2007. Trophic dynamics of two sympatric mysid species in an estuarine transition zone. Marine Ecology Progress Series 332: 171–187.
Withers, J. L., T. M. Sesterhenn, C. J. Foley, C. D. Troy & T. O. Höök, 2015. Diets and growth potential of early stage larval yellow perch and alewife in a nearshore region of southeastern Lake Michigan. Journal of Great Lakes Research 41(Suppl. 3): 197–209.
Wojtal-Frankiewicz, A. & P. Frankiewicz, 2010. The Influence of the Zebra Mussel (Dreissena polymorpha) on magnesium and calcium concentration in water. Acta Universitatis Lodziensis. Folia Biologica Et Oecologica 6: 81–101.
Wong, W. H., J. S. Levinton, B. S. Twining & N. Fisher, 2003. Assimilation of micro- and mesozooplankton by zebra mussels; a demonstration of the food web link between zooplankton and benthic suspension feeders. Limnology and Oceanography 48: 308–312.
Wong, W. H., S. Gerstenberger, W. Baldwin & B. Moore, 2012. Settlement and growth of quagga mussels (Dreissena rostriformis bugensis Andrusov, 1897) in Lake Mead, Nevada-Arizona, USA. Aquatic Invasions 7(1): 7–19.
Wright, D. A., E. M. Setzler-Hamilton, J. A. Magee & R. Harvey, 1996. Laboratory culture of zebra (Dreissena polymorpha) and quagga (D. bugensis) mussel larvae using estuarian algae. Journal of Great Lakes Research 22: 46–54.
Yablonskaya, E. A., 1985. The Caspian Sea: Fauna and Biological Productivity, Nauka, Moscow, USSR: 290 pp. (in Russian).
Yablonskaya, E. A., T. A. Kortunova & G. B. Gavrilov, 1973. Long-term changes in the Aral Sea benthos. Trudy Vsesoyuznogo Nauchno-Issledovatel’skogo Instituta Morskogo Rybnogo Khozyaistva i Okeanografii 80: 147–158 (in Russian).
Yaroshenko, M. F. & A. I. Naberezhny, 1971. On biology of Dreissena polymorpha Pallas in Kuchurganskiy cooling-liman of the Moldavian thermal power plant. Biologicheskie Resursy Vodoemov Moldavii 8: 31–41 (in Russian).
Yonge, C. M. & J. I. Campbell, 1968. On the heteromyarian condition in the Bivalvia with special reference to Dreissena polymorpha and certain Mytilacea. Transactions of the Royal Society of Edinburgh 68: 4–42.
Yu, N. & D. A. Culver, 1999. In situ survival and growth of zebra mussels (Dreissena polymorpha) under chronic hypoxia in a stratified lake. Hydrobiologia 392: 205–215.
Zenkevich, L. A., 1963. Biology of the USSR Seas. Academy of Science Press, Moscow: 739 pp. (in Russian).
Zhadin, V. I., 1946. The traveling shellfish Dreissena. Priroda 5: 29–37 (in Russian).
Zhadin, V.I., 1952. Mollusks of Fresh and Brackish Waters of the USSR. Academy of Sciences of the USSR Press Moscow Russia (in Russian).
Zhu, B., D. G. Fitzgerald, C. M. Mayer, L. G. Rudstam & E. L. Mills, 2006. Alteration of ecosystem function by zebra mussels in Oneida Lake: Impacts on submerged macrophytes. Ecosystems 9: 1017–1028.
Zhulidov, A. V., D. F. Pavlov, T. F. Nalepa, G. H. Scherbina, D. A. Zhulidov & TYu. Gurtovaya, 2004. Relative distributions of Dreissena bugensis and Dreissena polymorpha in the lower Don River system, Russia. International Review of Hydrobiology 89: 326–333.
Zhulidov, A. V., D. A. Zhulidov, D. F. Pavlov, T. F. Nalepa & T. Y. Gurtovaya, 2005. Expansion of the invasive bivalve mollusk Dreissena bugensis (quagga mussel) in the Don and Volga River Basins: revisions based on archived specimens. Ecohydrology and Hydrobiology 5(2): 127–133.
Zhulidov, A. V., T. F. Nalepa, A. V. Kozhara, D. A. Zhulidov & TYu. Gurtovaya, 2006. Recent trends in relative abundance of two dreissenid species, Dreissena polymorpha and Dreissena bugensis in the Lower Don River system, Russia. Archiv Fur Hydrobiologie 165: 209–220.
Zhulidov, A. V., A. V. Kozhara, G. H. Scherbina, T. F. Nalepa, A. Protasov, S. A. Afanasiev, E. G. Pryanichnikova, D. A. Zhulidov, T. Yu & Gurtovaya & D. F. Pavlov, 2010. Invasion history, distribution, and relative abundances of Dreissena bugensis in the Old World: a synthesis of data. Biological Invasions 12: 1923–1940.
Zhuravel, P. A., 1957. New invasive species from liman invertebrates in the Dnieprovskoe Reservoir. In Roll, Y. V. (ed), Problems of Ecology, Vol. 1. Kiev University Press, Kiev: 97–102 (in Russian).
Zhuravel, P. A., 1965. Wide spreading and mass growth of Dreissena bugensis in the canals and reservoirs of the Ukraine. In Likharev, I. M. (ed), Questions of Theoretical and Applied Malacology. Nauka Press, Leningrad: 63–64 (in Russian).
Zhuravel, P. A., 1967. Settling of Dreissena bugensis in artificial reservoirs. Hydrobiological Journal 3: 87–90 (in Russian).
Acknowledgements
This research was supported by the Great Lakes Restoration Initiative via a cooperative agreement with Cornell University, Department of Natural Resources under Prime Agreement Award GL 00E02259-2 from the U.S. EPA (PI L. Rudstam, Co-PIs Burlakova & Karatayev). The authors would like to thank Demetrio Boltovskoy for edits and suggestions to the earlier versions of this manuscript and for map graphics, Administrative Assistant S. Dickinson (GLC) for manuscript proofreading, and two anonymous reviewers for helpful suggestions. The views expressed in this publication are those of the authors and do not necessarily represent the views or policies of the U.S. EPA. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. EPA.
Funding
LEB supported by a cooperative agreement with Cornell University, Department of Natural Resources 1187 under Prime Agreement Award GL 00E02259-2 from the U.S. EPA “Great Lakes Long-Term Biological 1188 Monitoring Program 2017–2022” (PI L. Rudstam, Co-PIs Burlakova & Karatayev).
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study and performed the literature search and data analysis. The first draft of the manuscript was written by AYK and both authors reviewed and commented on previous versions of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest and no competing interests to declare regarding the publication of this article. The authors have no financial or proprietary interests in any material discussed in this article. All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript.
Ethical approval
Not applicable—this study is a review of published data and did not involve animals.
Consent to participate
Not applicable—this study did not involve human participants.
Additional information
Handling editor: Sidinei Magela Thomaz
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Manuel P. M. Lopes-Lima, Lyubov E. Burlakova, Ting Hui Ng, Alexandra Zieritz & Ronaldo G. Sousa / Biology and Impact of Invasive Freshwater Molluscs
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Karatayev, A.Y., Burlakova, L.E. What we know and don’t know about the invasive zebra (Dreissena polymorpha) and quagga (Dreissena rostriformis bugensis) mussels. Hydrobiologia (2022). https://doi.org/10.1007/s10750-022-04950-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10750-022-04950-5