
Abstract
Key message
Recent developments in understanding the distribution and distinctive features of recombination hotspots are reviewed and approaches are proposed to increase recombination frequency in coldspot regions.
Abstract
Recombination events during meiosis provide the foundation and premise for creating new varieties of crops. The frequency of recombination in different genomic regions differs across eukaryote species, with recombination generally occurring more frequently at the ends of chromosomes. In most crop species, recombination is rare in centromeric regions. If a desired gene variant is linked in repulsion with an undesired variant of a second gene in a region with a low recombination rate, obtaining a recombinant plant combining two favorable alleles will be challenging. Traditional crop breeding involves combining desirable genes from parental plants into offspring. Therefore, understanding the mechanisms of recombination and factors affecting the occurrence of meiotic recombination is important for crop breeding. Here, we review chromosome recombination types, recombination mechanisms, genes and proteins involved in the meiotic recombination process, recombination hotspots and their regulation systems and discuss how to increase recombination frequency in recombination coldspot regions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Meiosis is an essential part of sexual reproduction in most organisms. This process halves chromosome numbers by coupling a single round of DNA replication with two consecutive rounds of nuclear division (meiosis I and meiosis II) to produce haploid gametes (Hillers et al. 2017; Kleckner 1996). During meiosis I, replicated homologous chromosomes align and undergo recombination between non-sister chromatids before separating. Then, during meiosis II, sister chromatids segregate, ultimately producing four gametes (Kleckner 1996) with each gamete containing one set of chromosomes. When one gamete from one sex combines with a gamete of the opposite sex, the chromosome number of the subsequent generation returns to the parental level, maintaining a steady state (Hillers et al. 2017). This process leads to new combinations of alleles present in the progenies. As a result, chromosome recombination during meiosis is regarded as the foundation for genetic diversity and the evolution of species.
Recombination rates vary across different regions of chromosomes (Blair et al. 2018; Henderson 2012; Kauppi et al. 2004). In most crop species, recombination rates are positively correlated with the distance from the centromere and gene densities but negatively correlated with transposable elements (Barakate et al. 2014; Blair et al. 2018; Henderson 2012; Kauppi et al. 2004; Phillips et al. 2010; Shen et al. 2017). The genomic region that has a relatively higher recombination frequency is referred to as recombination hotspot, while the region with a lower recombinant frequency is called a coldspot. Traditional crop breeding relies heavily on incorporating beneficial gene alleles from parental chromosomes into their offspring. Consequently, low recombination frequency hampers the selection of lines that pyramid favorable close-linked traits within coldspots in crop breeding programs. Due to increased food demand and rapidly deteriorating climate change, it is an urgent requirement to create new varieties that would possess high yields and better quality while being climate resilient. However, previous studies indicated that many genes, such as around one-third in barley, are located in recombination coldspot regions (Higgins et al. 2014). This poses a challenge for crop breeders, as achieving their objectives leads to a significant increase in breeding costs and cycle times.
Although recombination mechanisms are not yet fully understood, substantial progress has been made in studying recombination models. This review provides an overview of gene recombination types and genetic mechanisms, the genes participating in the recombination process, the classification of recombination pathways, the distribution of recombination events and their regulatory systems in plants. Additionally, we explore the potential for enhancing recombination rates in coldspot regions.
Gene recombination and genetic mechanisms
Recombination types
In genetics, recombination refers to the process of rearranging genetic material from different chromosomes or regions to create new DNA combinations (Rice 2002; Stapley et al. 2017). It can occur naturally in both eukaryotes and prokaryotes and can also be induced in a laboratory (Baker et al. 1976; Camerini-Otero and Hsieh 1995; Covo et al. 2012; Gratia 2017; Paques and Haber 1999; Schnable et al. 1998). Recombination types are primarily classified into four groups: homologous recombination, non-homologous recombination, site-specific recombination and transposition (Fig. 1).

Four recombination types. a The process of homologous recombination involves the DNA double-strand break and rejoining the strands. It results in an exchange of genetic information between the homologous chromosomes. b The yellow and orange blocks represent the segments at the sites of nonhomologous recombination. It does not require sequence homology between the DNA molecules and can introduce mutations at the site of the break. c Site-specific recombination catalyzed by site-specific recombinase enzymes, is usually observed between two different DNA molecules among bacteriophages, bacteria, and unicellular eukaryotes. d The red block represents the transposable element of DNA which could be integrated into the genome
Homologous recombination, known as general recombination, typically occurs during meiosis in eukaryotic cells (Camerini-Otero and Hsieh 1995). It involves the exchange of genetic information between the alleles of homologous chromosomes and generates genetic diversity in offspring. The occurrence of homologous recombination requires homologous segments that have large-scale significant similar sequences to line up in proximity. Non-homologous recombination process ligates the broken ends of DNA together directly with no requirement of a homologous sequence to serve as a template to repair DNA double-stranded breaks (DSBs) (Pannunzio et al. 2018). It is more prone to mistakes and can give rise to the deletion or insertion of genetic material at the break site and even chromosomal abnormalities. These outcomes can have significant consequences for gene function and regulation (Pannunzio et al. 2018). Site-specific recombination process is catalyzed by site-specific recombinases and reintegrates the DNA segments at specific sites (Grindley et al. 2006). The transposition process involves the movement of transposable elements within the genome. Replicative transpositions may result in the creation of a new copy (Fedoroff 2000). A homologous recombination during meiosis is the primary type of recombination employed in crop breeding programs. Therefore, our focus will be on this specific type of recombination.
Homologous recombination process
The genetic recombination process has a variety of forms and presents great complexities, varying among species. In the meiosis of eukaryotes, the widely accepted model of homologous recombination is primarily based on studies of DSB repair in Saccharomyces cerevisiae (Aylon and Kupiec 2004; Osman et al. 2011). This process is initiated by programmed DSB and involves rejoining of DNA sequences (Lake and Hawley 2016; Murakami and Keeney 2008). DSB repair could generate either crossover (CO) recombination or non-crossover (NCO) recombination through different pathways, including double Holliday junction (dHJ) model and synthesis-dependent strand annealing (SDSA) model. The CO recombination modifies two chromatids by exchanging large DNA fragments, while NCO only involves copying and replacing a short stretch of DNA without exchange (Fig. 2). Homologous chromosome pairing and recombination occur in the prophase phase of meiosis I (prophase I) (Azumi et al. 2002; Zickler and Kleckner 2015). The chromosomes start to condense and become thin filaments that could be visible under the light microscope during the Prophase I leptotene stage (Hartl and Ruvolo 2012). At the zygotene stage, homologous chromosomes align closely through the formation of a synaptonemal complex (SC), a unique proteinaceous structure (Fraune et al. 2012; Hartl and Ruvolo 2012; Heyting 1996; Hillers et al. 2017; Page and Hawley 2004). The SC is completely assembled during pachytene stage, and is considered to promote the initiation of recombination events (Fraune et al. 2012; Hartl and Ruvolo 2012; Hayashi et al. 2010; Hernandez-Hernandez et al. 2016; Hillers et al. 2017; Kouznetsova et al. 2011; Schramm et al. 2011). The CO takes place between two non-sister chromatids of the homologous chromosomes during the pachytene stage (Gilbert and Barresi 2016; Hartl and Ruvolo 2012). In the diplotene stage, homologous chromosomes start to separate from each other with the dissolution of SC and are only attached at chiasmata (Armstrong and Jones 2003; Hartl and Ruvolo 2012; Heyting 1996). Finally, the chromosomes become fully condensed during the diakinesis stage (Hartl and Ruvolo 2012; Taiz et al. 2015). Some studies suggest that homology along the chromosome arms is the main determinant of the recognition and pairing of homologous chromosomes, with centromeres playing a negligible role in this process during meiosis (Corredor et al. 2007; Lefrancois et al. 2016).

CO and NCO produced from the homologous recombination. The homologous recombination begins with the DNA double-strand breaks of one of the homologous DNA duplexes, shown as blue strands. The 5′ end of the DSB is resected by specific nucleases to generate 3′ single-stranded DNA. One of the 3′ ends invades another homologous DNA duplexes which are shown as red strands, forming a displacement loop (D-loop) structure. DNA polymerase extends the invading 3′ end strand to generate the new DNA. For SDSA, the newly synthesized strand is displaced from the D-loop and then anneals, and typically produces NCO products. The formation of dHJ is derived from capturing the second end of the break after DNA synthesis extends the invading strand. It is resolved by cutting the noncrossed strands and producing NCO or by cutting the crossed strands and creating COs
Genes involved in DSB formation
DSB could be caused by exogenous or endogenous factors in a variety of circumstances. Genes that function in DSB formation are exceptionally diverse, which makes the study of the mechanism extremely complex (Table 1). During meiosis, the conserved SPO11 protein is one of the primary participants involved directly in the DSB process (Keeney and Neale 2006; Lam and Keeney 2015). It shares homology with the subunit A of archaeal topoisomerase VI (TopVIA), a type II DNA topoisomerase (Bergerat et al. 1997; Gadelle et al. 2003; Keeney 2008). Two hybrid active sites of the Spo11 contain tyrosine which reacts with the phosphodiester linkage of DNA to cleave DNA strands (Diaz et al. 2002; Nichols et al. 1999; Shingu et al. 2010). In addition, the homolog of archaeal topoisomerase VI subunit B (TopVIB), known as the meiotic topoisomerase VI B subunit (MTOPVIB), forms a complex with SPO11 and is also required for DSB formation in the meiotic recombination process (An et al. 2011; Fu et al. 2016; Robert et al. 2016; Tang et al. 2017; Vrielynck et al. 2016; Xue et al. 2016). Studies in S. cerevisiae indicate that SPO11 alone is insufficient to generate DSB. There are at least nine other proteins that promote DSB formation, namely Ski8, Mei4, Mer2, Mre11, Rad50, Rec102, Rec104, Rec114, and Xrs2 (Cole et al. 2010; Lam and Keeney 2015; Neale et al. 2005). They can form several different interacting subcomplexes, Spo11-Ski8, Rec102-Rec104, Rec114-Mei4-Mer2 and Mre11-Rad50-Xrs2 (MRX) (Lam and Keeney 2015; Li et al. 2006; Maleki et al. 2007). It is worth noting that the MRX complex is not only recruited during DSB formation but also plays a role in the subsequent DSB repair processes (Borde et al. 2004; Williams et al. 2007).
There are functional divergences and significant sequence evolutionary divergences among some DSB proteins across different species. For example, AtMRE11 and AtRAD50 only participate in the DNA repair process rather than generating DSB, and Ski8 is involved in DSB formation in S. cerevisiae but not in Arabidopsis (Jolivet et al. 2006; Osman et al. 2011). In rice, OsSPO11-1 and OsSPO11-4 participate in DSB formation, while OsSPO11-2 and OsSPO11-3 are not involved in DSB formation (An et al. 2011; Yu et al. 2010). Arabidopsis is widely accepted as a model system in plant scientific research, containing three SPO11 homologs, AtSPO11-1, AtSPO11-2, and AtSPO11-3 (Hartung and Puchta 2000; Stacey et al. 2006). However, only AtSPO11-1 and AtSPO11-2 are essential for DSB formation, likely acting as heterodimers in meiotic recombination (Grelon et al. 2001; Hartung et al. 2007; Stacey et al. 2006). Numerous proteins, such as AtPRD1, AtPRD2, AtPRD3 and AtDFO, participate in DSB formation in Arabidopsis (Muyt et al. 2009, 2007; Zhang et al. 2012). The protein AtPRD1 shows interactions with AtSPO11-1, AtSPO11-2, MTOPVIB, AtPRD3 and AtDFO, although there is currently no evidence of its interaction with AtPRD2 (Muyt et al. 2007; Tang et al. 2017).
DSB end processing and a single strand DNA invasion
After DSB, SPO11 remains covalently linked to the 5′ terminal of each broken DNA strand (Keeney et al. 1997; Lam and Keeney 2015; Neale et al. 2005). The MRX complex (composed of MRE11, RAD50, Xrs2) works with Com1/Sae2 (Table 1) to resect the 5′ end on each side of the DSB and remove SPO11 (Aylon and Kupiec 2004; Cannavo and Cejka 2014). Further resections of the 5′ termini are conducted by exonuclease 1 (EXO1), resulting in the generation of 3′ ssDNA tails (Garcia et al. 2011). With the assistance of recombinases, these ssDNA tails invade the homologous duplex DNA to form a recombination intermediate known as D-loop (Hunter 2015; Hunter and Kleckner 2001; Lichten 2001; Martinez-Perez and Colaiacovo 2009; Wang and Copenhaver 2018). A replication protein A (RPA) binds the 3′ terminus to prevent degradation and remove secondary structures, facilitating the recruitment of recombinases (Soustelle et al. 2002; Wold 1997). RPA is a heterotrimeric complex that consists of three subunits: RPA1, RPA2 and RPA3 (Ribeiro et al. 2016) (Table 1). RPA also act on DNA annealing which is promoted by Rad52 protein during second-end capture (Nimonkar et al. 2009; Sugiyama et al. 2006; Wang and Haber 2004). Once the 3′ ssDNA tails are protected, recombinase A (RecA)-like related recombinases are loaded to form presynaptic nucleoprotein filaments. These filaments facilitate the invasion of free 3′ end ssDNA into the duplex DNA of the paired homologous chromosome, forming the D-Loop in yeast (Brown and Bishop 2015; Shinohara et al. 1992). Two homologs of the bacterial RecA, Rad51 and Dmc1, have been discovered in most eukaryotic organisms (Table 1) (Bishop et al. 1992; Brown and Bishop 2015; Shinohara et al. 1992). Both recombinases play crucial roles in efficient meiotic recombination. Rad51 not only directly facilitates recombination during mitosis but also participates in meiotic recombination, whereas Dmc1 is merely required for meiotic recombination (Bishop 2012; Bishop et al. 1992; Shinohara et al. 1992).
DSB repair
The current meiotic DSB repair model is broadly divided into two categories: the dHJ model and the SDSA model. Both of these models have been observed in yeast system (Mitchel et al. 2010). In the dHJ model, the invading 3′ end function as a primer to initiate the DNA synthesis using invaded DNA as the template (Szostak et al. 1983). Consequently, the newly synthesized DNA contains a specified sequence which is the same as the invaded DNA. The strand invasion and second end capture lead to the formation of dHJ, which are resolved to form CO or NCO products (Szostak et al. 1983; Wyatt and West 2014). The characteristic of the SDSA model is strand displacement, where the invading strand can anneal with the other 3′ single-stranded end (Szostak et al. 1983). In the SDSA model, only one DNA terminal participates in the invasion process, while another one utilizes newly synthesized DNA as a template for synthesis. This process results in the formation of NCO products (Szostak et al. 1983).
In general, the majority of CO products are formed through the dHJ intermediate, while most NCO is primarily produced via the SDSA (Allers and Lichten 2001; McMahill et al. 2007). These intermediates can either undergo a repair, resulting in gene conversions, or they can segregate during the next round of replication. In addition, the dissolution of dHJs could also give rise to some NCO products (Wyatt and West 2014). Usually, only a small proportion of meiotic DSBs are repaired into COs in plants. Meiotic DSBs are generated in excess with more than 90% of plant DSBs being repaired using the sister chromatid as a template or being resolved as NCO (Mercier et al. 2015).
DSB and CO events are associated with chromosome number and size
The identification of recombination distribution provides valuable insights into genome evolution and plant breeding. Distribution patterns of DSB and CO, along with their hotspots, have been reported in various species (Table 2).
DSB varies considerably across different species. About 200 DSB events in Arabidopsis (Sanchez-Moran et al. 2007) and ~ 500 DSBs in maize (Pawlowski et al. 2003) have been detected. More DSB events (2100) have been identified in bread wheat as wheat has a larger genome size and a greater number of chromosomes (Gardiner et al. 2019). This suggests a correlation between the number of chromosomes and the genome size with the frequency of DSB events.
Only a small amount of DSBs are repaired into CO, for example, about 4% in maize (Sidhu et al. 2015). In Arabidopsis (2n = 10), the number of COs ranged from 7 to 13 across different studies, with six NCOs reported. In soybean (2n = 40), the average number of COs per recombinant line is 49–59, while NCOs are about 25, about half of the CO events. In cucumber (2n = 14), a dicot species, the average number of COs per individual ranges from 12.9 to 13.8, approximately one for each chromosome. In maize (2n = 20), a monocot species, approximately 500 DSBs were identified per cell, while only 20 COs were formed. In bread wheat, 55 COs were identified, while in barley (2n = 14), 19–24 COs were estimated across 45 genetic mapping populations (Table 2). The total number of COs is associated with the number of chromosomes. Soybean and bread wheat contain similar pairs of chromosome numbers, 20 and 21, respectively, and their COs are both around 55 (Table 2). For the species with a smaller number of chromosomes (5—10 pairs), e.g. Arabidopsis, cucumber, maize and barley, their corresponding COs are less than 25, confirming that the number of COs is positively correlated with the number of chromosomes. Besides, chromosome size also shows influence on CO events. For example, both cucumber and barley have seven pairs of chromosomes, but the genome size in barley is 13.5 times greater than that of cucumber. The considerably longer chromosomes in barley compared to cucumber lead to nearly twice as many.
In plants, typically one to two COs are distributed across most individual chromosomes (Jones and Franklin 2006; Sidhu et al. 2015), with the majority of DSBs being repaired as NCOs through DNA synthesis, utilizing the homologous chromosome as a template or the sister chromatid (Allers and Lichten 2001). Nevertheless, regardless of the mechanisms, as indicated by the studies listed in Table 2, the majority of genetic variation generated by meiotic recombination in plants originates from COs.
The factors determining whether COs or NCOs form from DSB are poorly understood. Studies conducted in mouse, C. elegans, and budding yeast have indicated that CO numbers are not impacted by DSB numbers, even when the number of DSB varies significantly (Cole et al. 2012; Martini et al. 2006; Rosu et al. 2011; Yokoo et al. 2012). In contrast, a strong correlation between the number of meiotic DSBs and COs has been found in maize (Sidhu et al. 2015) with 25.8% of bivalents having single chiasma, 72.6% forming two chiasmata and only 1.7% displaying three chiasma (Sidhu et al. 2015).
Furthermore, variations in recombination frequency exist between genders within the same species (Lenormand and Dutheil 2005; Martinez-Perez and Colaiacovo 2009). For example, in Arabidopsis, the recombination rate is higher males than in females, particularly in the sub-telomeric region (Giraut et al. 2011). A similar difference has been found in barley, with male gametes showing more COs than female gametes (Phillips et al. 2015).
CO hotspot distribution across chromosomes
A prerequisite for the formation of recombination is the occurrence of DSB. In the search of DSB hotspots, researchers have found a close relationship between the number of hotspots and chromosome length, with the average hotspot length being 1 ~ 2 kb (He et al. 2017; Paul et al. 2016). In maize, there is a low frequency of CO hotspots occur in the centromeric and pericentromeric chromosome regions, while these regions exhibit a high frequency of the DSB hotspots (He et al. 2017). In S. cerevisiae, hypomorphic mutants of spo11 show a decrease in DSB number but not CO numbers (Martini et al. 2006). As the repair of DSB can proceed via either the CO pathway or the NCO pathway, an increase in CO may occur at the cost of NCOs, maintaining homeostasis (Martini et al. 2006). Consequently, there isn’t an absolute correlation between the spo11 alleles and COs. Therefore, it is impossible to solely identify the recombination spots based on DSB spots.
Meiotic recombination events are unevenly distributed and are restricted to certain regions, particularly at the distal ends of chromosome arms (He et al. 2017; Lukaszewski 1992; Paigen and Petkov 2010; Petes 2001). The preferential distribution of CO hotspots is in gene-rich regions where the chromatin is easily accessible by DSB complex. Furthermore, structure variations, such as large inversions, have been reported to influence the recombination rate in barley. For example, no recombination event can be identified within a large 141 Mb inversion region on chromosome 7H from the DH population of RGT Planet and Hindmarsh (Jayakodi et al. 2020).
Some studies show that recombination hotspot tends to present near gene promoters (Choi et al. 2013; Mancera et al. 2008; Pan et al. 2011; Petes 2001; Wu and Lichten 1994). Research on the hexaploid wheat genome has shown that recombination hotspots typically occur near the coding regions of the chromosomes (Darrier et al. 2017) with about 95% of the recombination being distributed in 18 major and 30 minor gene-rich regions (Erayman et al. 2004). Similarly, studies in maize have suggested that approximately 90% hotspots are distributed in gene-rich regions (Fu et al. 2001; Gore et al. 2009; Kianian et al. 2018; Li et al. 2015; Rodgers-Melnick et al. 2015; Sidhu et al. 2015). A recent comprehensive study in cucumber revealed that over 93% of the COs are either in genes or their 10 kb regions. Among these, about 45% of COs occurred in distal intergenic regions, 25% in the promoter regions (2 kb upstream), 13% in introns, 10% in coding sequences, and 7% in untranslated regions (Wang et al. 2023).
McConaughy et al. (2023) identified 451 CO hotspots from two soybean mapping populations. These hotspots are distributed across the entire soybean genome with around 27% of them located in the pericentromeric regions. In barley, recombination is severely suppressed in some regions (Kunzel et al. 2000). Repetitive sequences are associated with distinct chromatin modifications, and their expansion suppresses the recombination rate (Henderson 2012). Hotspots account for less than 5% of the genome region. Distal CO occurrence is 25 times greater than interstitial chiasmata (Higgins et al. 2012).
Hotspots DNA motifs
Recombination events are correlated with the presence of specific DNA sequences (Zelkowski et al. 2019). DNA and chromatin features are associated with DSB hotspots. The popular motifs with the hotspots include CCN repeat motif, poly-A motif, and min-inverted-repeat transposable elements.
In Arabidopsis, DSB hotspots are correlated to CO hotspots. Three DNA motifs (A‐rich, CCN and CTT) have been found to be enriched in CO regions (Shilo et al. 2015). In maize, an associated 20‐bp‐long, GC‐rich sequence motif is similar to the CCN motif identified in Arabidopsis (Shilo et al. 2015). The recombination hotspot in maize is located in the bronze and Stc1 locus (Fu et al. 2001; He and Dooner 2009) and the recombination event is significantly enriched in GC-rich regions, which is similar to the CCN motif identified in Arabidopsis and yeast (Gerton et al. 2000; Liu et al. 2009; Sidhu et al. 2015). In cucumber, numerous hotpot motifs are identified for DSB, including the TATA repeat (Wang et al. 2023).
The regulation of recombination
The precise control of the frequency and distribution of meiotic recombination events remains challenging. Previous studies indicated that homologous chromosome pairing typically results in generated at least one CO event per chromosome (Bishop and Zickler 2004; Hillers 2004; Kleckner et al. 2004; Martini et al. 2006; Shinohara et al. 2008). However, a recent report showed that the absence of COs in some chromosomes in a few F2 lines in cucumber (Wang et al. 2023). Meiotic recombination distribution is uneven along chromosomes, and its regulation can be classified into chromosome-level regulation, genome-level regulation and other mechanisms (Kauppi et al. 2004; Lichten and Goldman 1995; Petes 2001; Sidhu et al. 2015).
Chromosome level regulation
The frequency of COs increases from centromeres to telomeres, with notably low frequency observed in the telomeric region (Chen et al. 2008; Higgins et al. 2012; Liu et al. 2009; Saintenac et al. 2009, 2011; Salome et al. 2012; Sidhu et al. 2015). In most organisms, each chromosome pair usually every undergoes one or two COs (Martini et al. 2006). When more than two COs present on the homologous recombination, one CO tends to suppress the occurrence of others in nearby regions, a phenomenon known as CO interference (Hillers 2004; Jones 1984; Kleckner et al. 2004; Muller 1916). This phenomenon also appears during the DSB period. For example, the occurrence of a DSB on one chromosome of S. cerevisiae suppresses the frequency of DSB generation on its homolog at the same and nearby positions (Fukuda et al. 2008; Xu and Kleckner 1995).
In cucumber, however, Wang et al. (2023) observed the absence of COs on chromosomes 3,4 and 5 in the individual line Y-154, no CO on chromosome 3 and 4 in the line X-69, no CO on chromosome 7 in line Y-231, and no CO on 1,5,6 in Y-284 (Wang et al. 2023), suggesting that this regulation system may not work in cucumber.
Genome level regulation
Recombination events tend to cluster in short specific genome regions of the genome (Marand et al. 2017). Studies in the mammalian species revealed the correlation between hotspot location and certain sequence motifs (Buard and de Massy 2007; Myers et al. 2005; Parvanov et al. 2010; Shifman et al. 2006; Smagulova et al. 2011). In humans and mice, the meiotic recombination hotspots are closely linked to the specific recognition DNA sequence of PRDM9 zinc finger protein (Borde and de Massy 2013; De Massy 2013). However, PRDM subfamilies are absent in plants (Zhang and Ma 2012). In plants, meiotic recombination hotspots tend to occur in regions close to gene promoters and terminators associated with active chromatin modifications (Choi et al. 2013; Drouaud et al. 2013; Fu et al. 2002; He et al. 2017; Wang and Copenhaver 2018). Previous studies in Arabidopsis indicated a consistency between recombination hotspots and DSB hotspot regions (Choi et al. 2013; Horton et al. 2012). However, recombination events in maize are only associated with the DSBs close to the genes (He and Dooner 2009; Yao et al. 2002).
Other regulation factors
Recombination events can be influenced by extrinsic conditions, such as biotic stress, extreme temperature, chemical substances, nutrients, and UV radiation (Boyko et al. 2007; Higgins et al. 2012; Lucht et al. 2002; Mickelbart et al. 2015; Molinier et al. 2006; Phillips et al. 2015). The formation of COs relies on the ZMM protein or the endonuclease Mus81 (Berchowitz et al. 2007; Bishop and Zickler 2004; Borner et al. 2004; Santos et al. 2003; Mercier et al. 2005; Shinohara et al. 2008). Posttranslational modification could influence the activity and stability of proteins related to meiotic recombination, thereby regulating recombination events (Wang and Copenhaver 2018). DNA methylation has also implicated in regulating the formation of meiotic recombination (Buard and de Massy 2007).
The effect of temperature on miotic recombination has been reported in several species. In Allium ursinum, exposure to 35 °C for 30 h resulted in a detrimental effect on chromosome synapsis (Loidl 1989), whereas in barley, synapsis failed to occur at 35 °C (Higgins et al. 2012). In addition, the distribution and frequency of Chiasmata were altered when exposed to temperatures of 30 °C and 22 °C. At 30 °C, there was an increase in interstitial/proximal chiasmata, but the average number of chiasmata and COs per cell were significantly decreased (Higgins et al. 2012).
Phillips et al. (2015) found that the recombination rate during male meiosis consistently suppressed that of females. Moreover, in barley, as the temperature increased from 15 °C to 25 °C and 30 °C, the recombination rate increased during male meiosis but decreased during female meiosis. Similarly, in Arabidopsis, Giraut et al. (2011) demonstrated higher CO frequencies during male meiosis.
How to increase CO frequency
CRISPR-Cas9
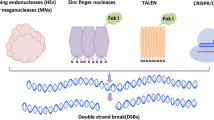
The main limitation of targeted homologous recombination is DSB formation. The homologous recombination frequency can be enhanced dramatically when the DSB occurs at the target locus (Hayut et al. 2017; Puchta and Fauser 2013). Therefore, it is essential to find effective methods for inducing greater DSB formation. Sequence-specific nucleases (SSNs) are recognized for their capability to generate DSB at a specific site (Belhaj et al. 2015). The development of sequence-specific nuclease, including zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs), has already proven successful in targeted gene editing in plants (Mao et al. 2019; Podevin et al. 2013; Voytas and Gao 2014; Wang et al. 2019). However, challenges associated with the design and construction of large modular proteins have hindered their widespread adoption (Doudna and Charpentier 2014). In addition, ZFNs have shown a high fault rate during DNA sequence recognition and cleavage (Voytas 2013). In contrast, the CRISPR-Cas method has emerged as a versatile solution. In recent years, the CRISPR-Cas system, a relatively easy and powerful gene-editing tool, has achieved rapid development. Most studies have used CRISPR-Cas9 technology to edit genes in homozygous tissues, potentially increasing the occurrence of DSB. Recently, the CRISPR-Cas9 system has been used in targeted recombination in tomatoes (Hayut et al. 2017). In this study, the F1 hybrid seed was used for targeted DNA editing, resulting in homologous CO events. Applying the allele specific recombination analysis suggests that the homologous recombination rate can be increased by generating DSB (Hayut et al. 2017).
Mutation
Mutating genes involved in COs is a powerful tool to increase CO frequency. FANCM and RECQ4 are key players in the CO pathway, and the impact of mutations in these genes on CO rate has been extensively studied in Arabidopsis (Crismani et al. 2012; Seguela-Arnaud et al. 2015). For instance, fancm (Crismani et al. 2012) and recq4a/b (Seguela-Arnaud et al. 2015) mutants exhibit a nearly 3–5.9 folds increase in recombination rate (Fernandes et al. 2018b).
The AAA‐ATPase FIDGETIN‐like 1 (FIGL1) negatively regulates CO formation at the early stages (Girard et al. 2015). The figl1 mutation enhances the CO rate by 1.5 times (Girard et al. 2015) and when combined with arecq4a/b double mutation, the CO rate is significantly increased (7.8‐fold) (Fernandes et al. 2018b). Remarkably, this mutant exhibits 60.7 COs per meiosis, compared to only 7.8 COs in the wild type (Fernandes et al. 2018b).
Other approaches
Higher temperatures increase recombination rates in male meiosis in barley (Phillips et al. 2015) with CO events increasing by 40% when the temperature rose from 15 °C to 30 °C (Phillips et al. 2015). This approach can be tried in other crops to increase the recombination rate. Furthermore, DNA methylation occurs across the plant genome, regulating gene expression (Jeddeloh et al. 1998) and silencing transposable element activity (Slotkin and Martienssen 2007). Changing DNA methylation patterns has been shown to alter CO distribution in mutant plants. For instance, loss of CG methylation in Arabidopsis leads to changes in CO distribution (Melamed-Bessudo and Levy 2012; Mirouze et al. 2012). These studies suggest that modifying methylation patterns can remodel CO distribution in plants.
Conclusion
While there are slight variations in CO distribution patterns, most recombination events occur toward the ends of chromosomes. In plant breeding programs, changing temperatures, creating mutations, reducing methylation patterns, and targeting CRISPR-Cas9 system can be used to regulate the recombination frequency within coldspots. With the development of whole genome sequencing, the identification of recombination spots becomes more precise. High-density genetic maps and resequencing data can be used to identify the precise location of the CO events and their associated motifs. A better understanding of CO and targeting hotspots will facilitate CO regulation in crop breeding programs.
Availability of data and materials
No datasets were generated or analysed during the current study.
References
Aklilu BB, Soderquist RS, Culligan KM (2014) Genetic analysis of the replication protein A large subunit family in Arabidopsis reveals unique and overlapping roles in DNA repair, meiosis and DNA replication. Nucleic Acids Res 42:3104–3118
Allers T, Lichten M (2001) Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106:47–57
An XJ, Deng ZY, Wang T (2011) OsSpo11-4, a rice homologue of the archaeal TopVIA protein, mediates double-strand DNA cleavage and interacts with OsTopVIB. PLoS ONE 6:e20327
Anderson LK, Doyle GG, Brigham B, Carter J, Hooker KD, Lai A, Rice M, Stack SM (2003) High-resolution crossover maps for each bivalent of Zea mays using recombination nodules. Genetics 165:849–865
Armstrong SJ, Jones GH (2003) Meiotic cytology and chromosome behaviour in wild-type Arabidopsis thaliana. J Exp Bot 54:1–10
Aylon Y, Kupiec M (2004) DSB repair: the yeast paradigm. DNA Repair 3:797–815
Azumi Y, Liu DH, Zhao DZ, Li WX, Wang GF, Hu Y, Ma H (2002) Homolog interaction during meiotic prophase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. EMBO J 21:3081–3095
Baker BS, Boyd JB, Carpenter ATC, Green MM, Nguyen TD, Ripoll P, Smith PD (1976) Genetic controls of meiotic recombination and somatic DNA metabolism in Drosophila melanogaster. P Natl Acad Sci USA 73:4140–4144
Barakate A, Higgins JD, Vivera S, Stephens J, Perry RM, Ramsay L, Colas I, Oakey H, Waugh R, Franklin FCH, Armstrong SJ, Halpin C (2014) The synaptonemal complex protein ZYP1 is required for imposition of meiotic crossovers in barley. Plant Cell 26:729–740
Barakate A, Arrieta M, Macaulay M, Vivera S, Davidson D, Stephens J, Orr J, Schreiber M, Ramsay L, Halpin C, Waugh R (2021) Downregulation of barley regulator of telomere elongation helicase 1 alters the distribution of meiotic crossovers. Front Plant Sci 12:745070
Belhaj K, Chaparro-Garcia A, Kamoun S, Patron NJ, Nekrasov V (2015) Editing plant genomes with CRISPR/Cas9. Curr Opin Biotechnol 32:76–84
Berchowitz LE, Francis KE, Bey AL, Copenhaver GP (2007) The role of AtMUS81 in interference-insensitive crossovers in A. thaliana. Plos Genet 3:e132
Bergerat A, deMassy B, Gadelle D, Varoutas PC, Nicolas A, Forterre P (1997) An atypical topoisomerase II from archaea with implications for meiotic recombination. Nature 386:414–417
Bishop DK (2012) Rad51, the lead in mitotic recombinational DNA repair, plays a supporting role in budding yeast meiosis. Cell Cycle 11:4105–4106
Bishop DK, Zickler D (2004) Early decision: meiotic crossover interference prior to stable strand exchange and synapsis. Cell 117:9–15
Bishop DK, Park D, Xu LZ, Kleckner N (1992) DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell 69:439–456
Blair MW, Cortes AJ, Farmer AD, Huang W, Ambachew D, Penmetsa RV, Carrasquilla-Garcia N, Assefa T, Cannon SB (2018) Uneven recombination rate and linkage disequilibrium across a reference SNP map for common bean (Phaseolus vulgaris L.). PLoS ONE 13:e0189597
Blary A, Gonzalo A, Eber F, Berard A, Berges H, Bessoltane N, Charif D, Charpentier C, Cromer L, Fourment J, Genevriez C, Le Paslier MC, Lode M, Lucas MO, Nesi N, Lloyd A, Chevre AM, Jenczewski E (2020) FANCM limits meiotic crossovers in Brassica Crops. Front Plant Sci 11:337590
Bleuyard JY, White CI (2004) The Arabidopsis homologue of Xrcc3 plays an essential role in meiosis. Embo J 23:439–449
Bleuyard JY, Gallego ME, White CI (2004) Meiotic defects in the Arabidopsis rad50 mutant point to conservation of the MRX complex function in early stages of meiotic recombination. Chromosoma 113:197–203
Borde V, de Massy B (2013) Programmed induction of DNA double strand breaks during meiosis: setting up communication between DNA and the chromosome structure. Curr Opin Genet Dev 23:147–155
Borde V, Lin W, Novikov E, Petrini JH, Lichten M, Nicolas A (2004) Association of Mre11p with double-strand break sites during yeast meiosis. Mol Cell 13:389–401
Borner GV, Kleckner N, Hunter N (2004) Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell 117:29–45
Boyko A, Kathiria P, Zemp FJ, Yao YL, Pogribny I, Kovalchuk I (2007) Transgenerational changes in the genome stability and methylation in pathogen-infected plants (Virus-induced plant genome instability). Nucleic Acids Res 35:1714–1725
Brown MS, Bishop DK (2015) DNA strand exchange and RecA homologs in meiosis. Csh Perspect Biol 7:a016659
Buard J, de Massy B (2007) Playing hide and seek with mammalian meiotic crossover hotspots. Trends Genet 23:301–309
Byun MY, Kim WT (2014) Suppression of OsRAD51D results in defects in reproductive development in rice (Oryza sativa L.). Plant J 79:256–269
Camerini-Otero RD, Hsieh P (1995) Homologous recombination proteins in prokaryotes and eukaryotes. Annu Rev Genet 29:509–552
Cannavo E, Cejka P (2014) Sae2 promotes dsDNA endonuclease activity within Mre11-Rad50-Xrs2 to resect DNA breaks. Nature 514:122–125
Chang L, Ma H, Xue HW (2009a) Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice. Cell Res 19:768–782
Chang YX, Gong L, Yuan WY, Li XW, Chen GX, Li XH, Zhang QF, Wu CY (2009b) Replication protein A (RPA1a) is required for meiotic and somatic DNA repair but is dispensable for DNA replication and homologous recombination in rice. Plant Physiol 151:2162–2173
Chelysheva L, Vezon D, Chambon A, Gendrot G, Pereira L, Lemhemdi A, Vrielynck N, Le Guin S, Novatchkova M, Grelon M (2012) The Arabidopsis HEI10 Is a new ZMM protein related to Zip3. Plos Genet 8:e1002799
Chen CB, Zhang W, Timofejeva L, Gerardin Y, Ma H (2005) The Arabidopsis ROCK-N-ROLLERS gene encodes a homolog of the yeast ATP-dependent DNA helicase MER3 and is required for normal meiotic crossover formation. Plant J 43:321–334
Chen SY, Tsubouchi T, Rockmill B, Sandler JS, Richards DR, Vader G, Hochwagen A, Roeder GS, Fung JC (2008) Global analysis of the meiotic crossover landscape. Dev Cell 15:401–415
Choi KH, Zhao XH, Kelly KA, Venn O, Higgins JD, Yelina NE, Hardcastle TJ, Ziolkowski PA, Copenhaver GP, Franklin FCH, McVean G, Henderson IR (2013) Arabidopsis meiotic crossover hot spots overlap with H2A. Z nucleosomes at gene promoters. Nat Genet 45:1327–1336
Cole F, Keeney S, Jasin M (2010) Evolutionary conservation of meiotic DSB proteins: more than just Spo11. Gene Dev 24:1201–1207
Cole F, Kauppi L, Lange J, Roig I, Wang R, Keeney S, Jasin M (2012) Homeostatic control of recombination is implemented progressively in mouse meiosis. Nat Cell Biol 14:424–430
Corredor E, Lukaszewski AJ, Pachon P, Allen DC, Naranjo T (2007) Terminal regions of wheat chromosomes select their pairing partners in meiosis. Genetics 177:699–706
Couteau F, Belzile F, Horlow C, Grandjean O, Vezon D, Doutriaux MP (1999) Random chromosome segregation without meiotic arrest in both male and female meiocytes of a dmc1 mutant of Arabidopsis. Plant Cell 11:1623–1634
Covo S, Ma WJ, Westmoreland JW, Gordenin DA, Resnick MA (2012) Understanding the origins of UV-induced recombination through manipulation of sister chromatid cohesion. Cell Cycle 11:3937–3944
Crismani W, Girard C, Froger N, Pradillo M, Santos JL, Chelysheva L, Copenhaver GP, Horlow C, Mercier R (2012) FANCM limits meiotic crossovers. Science 336:1588–1590
Da Ines O, Abe K, Goubely C, Gallego ME, White CI (2012) Differing requirements for RAD51 and DMC1 in meiotic pairing of centromeres and chromosome arms in Arabidopsis thaliana. Plos Genet 8:245–256
Darrier B, Rimbert H, Balfourier F, Pingault L, Josselin AA, Servin B, Navarro J, Choulet F, Paux E, Sourdille P (2017) High-resolution mapping of crossover events in the hexaploid wheat genome suggests a universal recombination mechanism. Genetics 206:1373–1388
De Massy B (2013) Initiation of meiotic recombination: how and where? Conservation and specificities among eukaryotes. Annu Rev Genet 47(47):563–599
De Maagd RA, Loonen A, Chouaref J, Pele A, Meijer-Dekens F, Fransz P, Bai YL (2020) CRISPR/Cas inactivation of RECQ4 increases homeologous crossovers in an interspecific tomato hybrid. Plant Biotechnol J 18:805–813
De Muyt A, Vezon D, Gendrot G, Gallois JL, Stevens R, Grelon M (2007) AtPRD1 is required for meiotic double strand break formation in Arabidopsis thaliana. Embo J 26:4126–4137
De Muyt A, Pereira L, Vezon D, Chelysheva L, Gendrot G, Chambon A, Laine-Choinard S, Pelletier G, Mercier R, Nogue F, Grelon M (2009) A high throughput genetic screen identifies new early meiotic recombination functions in Arabidopsis thaliana. Plos Genet 5:e1000654
Deng ZY, Wang T (2007) OsDMC1 is required for homologous pairing in Oryza sativa. Plant Mol Biol 65:31–42
Diaz RL, Alcid AD, Berger JM, Keeney S (2002) Identification of residues in yeast Spo11p critical for meiotic DNA double-strand break formation. Mol Cell Biol 22:1106–1115
Dion E, Li LL, Jean M, Beizile F (2007) An Arabidopsis MLH1 mutant exhibits reproductive defects and reveals a dual role for this gene in mitotic recombination. Plant J 51:431–440
Doudna JA, Charpentier E (2014) The new frontier of genome engineering with CRISPR-Cas9. Science 346:1077–1086
Drouaud J, Khademian H, Giraut L, Zanni V, Bellalou S, Henderson IR, Falque M, Mezard C (2013) Contrasted patterns of crossover and non-crossover at Arabidopsis thaliana meiotic recombination hotspots. Plos Genet 9:e1003922
Duroc Y, Lemhemdi A, Larcheveque C, Hurel A, Cuacos M, Cromer L, Horlow C, Armstrong SJ, Chelysheva L, Mercier R (2014) The kinesin AtPSS1 promotes synapsis and is required for proper crossover distribution in meiosis. Plos Genet 10:e1004674
Erayman M, Sandhu D, Sidhu D, Dilbirligi M, Baenziger PS, Gill KS (2004) Demarcating the gene-rich regions of the wheat genome. Nucleic Acids Res 32:3546–3565
Fedoroff N (2000) Transposons and genome evolution in plants. P Natl Acad Sci USA 97:7002–7007
Fernandes JB, Duhamel M, Seguela-Arnaud M, Froger N, Girard C, Choinard S, Solier V, De Winne N, De Jaeger G, Gevaert K, Andrey P, Grelon M, Guerois R, Kumar R, Mercier R (2018a) FIGL1 and its novel partner FLIP form a conserved complex that regulates homologous recombination. Plos Genet 14:e1007317
Fernandes JB, Seguela-Arnaud M, Larcheveque C, Lloyd AH, Mercier R (2018b) Unleashing meiotic crossovers in hybrid plants. P Natl Acad Sci USA 115:2431–2436
Fraune J, Schramm S, Alsheimer M, Benavente R (2012) The mammalian synaptonemal complex: protein components, assembly and role in meiotic recombination. Exp Cell Res 318:1340–1346
Fu HH, Park WK, Yan XH, Zheng ZW, Shen BZ, Dooner HK (2001) The highly recombinogenic bz locus lies in an unusually gene-rich region of the maize genome. P Natl Acad Sci USA 98:8903–8908
Fu HH, Zheng ZW, Dooner HK (2002) Recombination rates between adjacent genic and retrotransposon regions in maize vary by 2 orders of magnitude. P Natl Acad Sci USA 99:1082–1087
Fu M, Wang C, Xue FY, Higgins J, Chen MJ, Zhang DB, Liang WQ (2016) The DNA topoisomerase VI-B subunit OsMTOPVIB is essential for meiotic recombination initiation in rice. Mol Plant 9:1539–1541
Fukuda T, Kugou K, Sasanuma H, Shibata T, Ohta K (2008) Targeted induction of meiotic double-strand breaks reveals chromosomal domain-dependent regulation of Spo11 and interactions among potential sites of meiotic recombination. Nucleic Acids Res 36:984–997
Gadelle D, Filee J, Buhler C, Forterre P (2003) Phylogenomics of type II DNA topoisomerases. BioEssays 25:232–242
Garcia V, Phelps SEL, Gray S, Neale MJ (2011) Bidirectional resection of DNA double-strand breaks by Mre11 and Exo1. Nature 479:241–244
Gardiner LJ, Wingen LU, Bailey P, Joynson R, Brabbs T, Wright J, Higgins JD, Hall N, Griffiths S, Clavijo BJ, Hall A (2019) Analysis of the recombination landscape of hexaploid bread wheat reveals genes controlling recombination and gene conversion frequency. Genome Biol 20:1–16
Gerton JL, DeRisi J, Shroff R, Lichten M, Brown PO, Petes TD (2000) Global mapping of meiotic recombination hotspots and coldspots in the yeast Saccharomyces cerevisiae. P Natl Acad Sci USA 97:11383–11390
Gilbert SF, Barresi MJF (2016) Developmental biology, 11th edition. Sinauer Associates Inc., Sunderland
Girard C, Chelysheva L, Choinard S, Froger N, Macaisne N, Lehmemdi A, Mazel J, Crismani W, Mercier R (2015) AAA-ATPase FIDGETIN-LIKE 1 and helicase FANCM antagonize meiotic crossovers by distinct mechanisms. Plos Genet 11:e1005369
Giraut L, Falque M, Drouaud J, Pereira L, Martin OC, Mezard C (2011) Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes. Plos Genet 7:e1002354
Gore MA, Chia JM, Elshire RJ, Sun Q, Ersoz ES, Hurwitz BL, Peiffer JA, McMullen MD, Grills GS, Ross-Ibarra J, Ware DH, Buckler ES (2009) A first-generation haplotype map of maize. Science 326:1115–1117
Gratia JP (2017) Genetic recombinational events in prokaryotes and their viruses: insight into the study of evolution and biodiversity. Anton Leeuw Int J G 110:1493–1514
Grelon M, Vezon D, Gendrot G, Pelletier G (2001) AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J 20:589–600
Griffiths S, Sharp R, Foote TN, Bertin I, Wanous M, Reader S, Colas I, Moore G (2006) Molecular characterization of Ph1 as a major chromosome pairing locus in polyploid wheat. Nature 439:749–752
Grindley NDF, Whiteson KL, Rice PA (2006) Mechanisms of site-specific recombination. Annu Rev Biochem 75:567–605
Hartl DL, Ruvolo M (2012) Genetics : analysis of genes and genomes, 8th edn. Jones & Bartlett Learning, Burlington
Hartung F, Puchta H (2000) Molecular characterisation of two paralogous SPO11 homologues in Arabidopsis thaliana. Nucleic Acids Res 28:1548–1554
Hartung F, Suer S, Bergmann T, Puchta H (2006) The role of AtMUS81 in DNA repair and its genetic interaction with the helicase AtRecQ4A. Nucleic Acids Res 34:4438–4448
Hartung F, Wurz-Wildersinn R, Fuchs J, Schubert I, Suer S, Puchta H (2007) The catalytically active tyrosine residues of both SPO11-1 and SPO11-2 are required for meiotic double-strand break induction in Arabidopsis. Plant Cell 19:3090–3099
Hayashi M, Mlynarczyk-Evans S, Villeneuve AM (2010) The synaptonemal complex shapes the crossover landscape through cooperative assembly, crossover promotion and crossover inhibition during Caenorhabditis elegans meiosis. Genetics 186:45–58
Hayut SF, Bessudo CM, Levy AA (2017) Targeted recombination between homologous chromosomes for precise breeding in tomato. Nat Commun 8:1–9
He LM, Dooner HK (2009) Haplotype structure strongly affects recombination in a maize genetic interval polymorphic for Helitron and retrotransposon insertions. P Natl Acad Sci USA 106:8410–8416
He Y, Wang MH, Dukowic-Schulze S, Zhou A, Tiang CL, Shilo S, Sidhu GK, Eichten S, Bradbury P, Springer NM, Buckler ES, Levy AA, Sun Q, Pillardy J, Kianian PMA, Kianian SF, Chen CB, Pawlowski WP (2017) Genomic features shaping the landscape of meiotic double-strand-break hotspots in maize. P Natl Acad Sci USA 114:12231–12236
Henderson IR (2012) Control of meiotic recombination frequency in plant genomes. Curr Opin Plant Biol 15:556–561
Hernandez-Hernandez A, Masich S, Fukuda T, Kouznetsova A, Sandin S, Daneholt B, Hoog C (2016) The central element of the synaptonemal complex in mice is organized as a bilayered junction structure. J Cell Sci 129:2239–2249
Heyting C (1996) Synaptonemal complexes: structure and function. Curr Opin Cell Biol 8:389–396
Higgins JD, Armstrong SJ, Franklin FCH, Jones GH (2004) The Arabidopsis MutS homolog AtMSH4 functions at an early step in recombination: evidence for two classes of recombination in Arabidopsis. Gene Dev 18:2557–2570
Higgins JD, Sanchez-Moran E, Armstrong SJ, Jones GH, Franklin FCH (2005) The Arabidopsis synaptonemal complex protein ZYP1 is required for chromosome synapsis and normal fidelity of crossing over. Gene Dev 19:2488–2500
Higgins JD, Buckling EF, Franklin FCH, Jones GH (2008a) Expression and functional analysis of AtMUS81 in Arabidopsis meiosis reveals a role in the second pathway of crossing-over. Plant J 54:152–162
Higgins JD, Vignard J, Mercier R, Pugh AG, Franklin FCH, Jones GH (2008b) AtMSH5 partners AtMSH4 in the class I meiotic crossover pathway in Arabidopsis thaliana, but is not required for synapsis. Plant J 55:28–39
Higgins JD, Perry RM, Barakate A, Ramsay L, Waugh R, Halpin C, Armstrong SJ, Franklin FCH (2012) Spatiotemporal asymmetry of the meiotic program underlies the predominantly distal distribution of meiotic crossovers in barley. Plant Cell 24:4096–4109
Higgins JD, Osman K, Jones GH, Franklin FC (2014) Factors underlying restricted crossover localization in barley meiosis. Annu Rev Genet 48:29–47
Hillers KJ (2004) Crossover interference. Curr Biol 14:1036–1037
Hillers KJ, Jantsch V, Martinez-Perez E, Yanowitz JL (2017) Meiosis. Wormbook 2017:1–43
Horton MW, Hancock AM, Huang YS, Toomajian C, Atwell S, Auton A, Muliyati NW, Platt A, Sperone FG, Vilhjalmsson BJ, Nordborg M, Borevitz JO, Bergelson J (2012) Genome-wide patterns of genetic variation in worldwide Arabidopsis thaliana accessions from the RegMap panel. Nat Genet 44:212–216
Hunter N (2015) Meiotic recombination: the essence of heredity. Cold Spring Harb Perspect Biol 7:a016618
Hunter N, Kleckner N (2001) The single-end invasion: an asymmetric intermediate at the double-strand break to double-holliday junction transition of meiotic recombination. Cell 106:59–70
Jackson N, Sanchez-Moran E, Buckling E, Armstrong SJ, Jones GH, Franklin FCH (2006) Reduced meiotic crossovers and delayed prophase I progression in AtMLH3-deficient Arabidopsis. Embo J 25:1315–1323
Jayakodi M, Padmarasu S, Haberer G, Bonthala VS, Gundlach H, Monat C, Lux T, Kamal N, Lang DI, Himmelbach A, Ens J, Zhang XQ, Angessa TT, Zhou GF, Tan C, Hill C, Wang PH, Schreiber M, Boston LB, Plott C, Jenkins J, Guo Y, Fiebig A, Budak H, Xu DD, Zhang J, Wang CC, Grimwood J, Schmutz J, Guo GG, Zhang GP, Mochida K, Hirayama T, Sato K, Chalmers KJ, Langridge P, Waugh R, Pozniak CJ, Scholz U, Mayer KFX, Spannagl M, Li CD, Mascher M, Stein N (2020) The barley pan-genome reveals the hidden legacy of mutation breeding. Nature 588:284–289
Jeddeloh JA, Bender J, Richards EJ (1998) The DNA methylation locus DDM1 is required for maintenance of gene silencing in Arabidopsis. Gene Dev 12:1714–1725
Ji JH, Tang D, Wang KJ, Wang M, Che LX, Li M, Cheng ZK (2012) The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis. Plant J 72:18–30
Ji JH, Tang D, Wang M, Li YF, Zhang L, Wang KJ, Li M, Cheng ZK (2013) MRE11 is required for homologous synapsis and DSB processing in rice meiosis. Chromosoma 122:363–376
Jolivet S, Vezon D, Froger N, Mercier R (2006) Non conservation of the meiotic function of the Ski8/Rec103 homolog in Arabidopsis. Genes Cells 11:615–622
Jones GH (1984) The control of chiasma distribution. Symp Soc Exp Biol 38:293–320
Jones GH, Franklin FCH (2006) Meiotic crossing-over: obligation and interference. Cell 126:246–248
Kauppi L, Jeffreys AJ, Keeney S (2004) Where the crossovers are: recombination distributions in mammals. Nat Rev Genet 5:413–424
Keeney S (2008) Spo11 and the formation of DNA double-strand breaks in meiosis. Genome Dyn Stab 2:81–123
Keeney S, Neale MJ (2006) Initiation of meiotic recombination by formation of DNA double-strand breaks: mechanism and regulation. Biochem Soc T 34:523–525
Keeney S, Giroux CN, Kleckner N (1997) Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 88:375–384
Kianian PMA, Wang MH, Simons K, Ghavami F, He Y, Dukowic-Schulze S, Sundararajan A, Sun Q, Pillardy J, Mudge J, Chen CB, Kianian SF, Pawlowski WP (2018) High-resolution crossover mapping reveals similarities and differences of male and female recombination in maize. Nat Commun 9:2370
Kleckner N (1996) Meiosis: how could it work? P Natl Acad Sci USA 93:8167–8174
Kleckner N, Zickler D, Jones GH, Dekker J, Padmore R, Henle J, Hutchinson J (2004) A mechanical basis for chromosome function. P Natl Acad Sci USA 101:12592–12597
Knoll A, Higgins JD, Seeliger K, Reha SJ, Dangel NJ, Bauknecht M, Schropfer S, Franklin FCH, Puchta H (2012) The Fanconi anemia ortholog FANCM ensures ordered homologous recombination in both somatic and meiotic cells in Arabidopsis. Plant Cell 24:1448–1464
Kou YJ, Chang YX, Li XH, Xiao JH, Wang SP (2012) The rice RAD51C gene is required for the meiosis of both female and male gametocytes and the DNA repair of somatic cells. J Exp Bot 63:5323–5335
Kouznetsova A, Benavente R, Pastink A, Hoog C (2011) Meiosis in mice without a synaptonemal complex. PLoS ONE 6:e28255
Kunzel G, Korzun L, Meister A (2000) Cytologically integrated physical restriction fragment length polymorphism maps for the barley genome based on translocation breakpoints. Genetics 154:397–412
Kuromori T, Azumi Y, Hayakawa S, Kamiya A, Imura Y, Wada T, Shinozaki K (2008) Homologous chromosome pairing is completed in crossover defective atzip4 mutant. Biochem Bioph Res Co 370:98–103
Lake CM, Hawley RS (2016) Becoming a crossover-competent DSB. Semin Cell Dev Biol 54:117–125
Lam I, Keeney S (2015) Mechanism and regulation of meiotic recombination initiation. Csh Perspect Biol 7:a016634
Lefrancois P, Rockmill B, Xie P, Roeder GS, Snyder M (2016) Multiple pairwise analysis of non-homologous centromere coupling reveals preferential chromosome size-dependent interactions and a role for bouquet formation in establishing the interaction pattern. Plos Genet 12:e1006347
Lenormand T, Dutheil J (2005) Recombination difference between sexes: a role for haploid selection. Plos Biol 3:396–403
Li J, Hooker GW, Roeder GS (2006) Saccharomyces cerevisiae Mer2, Mei4 and Rec114 form a complex required for meiotic double-strand break formation. Genetics 173:1969–1981
Li J, Harper LC, Golubovskaya I, Wang CR, Weber D, Meeley RB, McElver J, Bowen B, Cande WZ, Schnable PS (2007) Functional analysis of maize RAD51 in meiosis and double-strand break repair. Genetics 176:1469–1482
Li XW, Chang YX, Xin XD, Zhu CM, Li XH, Higgins JD, Wu CY (2013) Replication protein A2c coupled with replication protein A1c regulates crossover formation during meiosis in rice. Plant Cell 25:3885–3899
Li X, Li L, Yan JB (2015) Dissecting meiotic recombination based on tetrad analysis by single-microspore sequencing in maize. Nat Commun 6:6648
Lian QC, Solier V, Walkemeier B, Durand S, Huettel B, Schneeberger K, Mercier R (2022) The megabase-scale crossover landscape is largely independent of sequence divergence. Nat Commun 13:3828
Lichten M (2001) Meiotic recombination: breaking the genome to save it. Curr Biol 11:R253–R256
Lichten M, Goldman ASH (1995) Meiotic recombination hotspots. Annu Rev Genet 29:423–444
Liu SZ, Yeh CT, Ji TM, Ying K, Wu HY, Tang HM, Fu Y, Nettleton D, Schnable PS (2009) Mu transposon insertion sites and meiotic recombination events co-localize with epigenetic marks for open chromatin across the maize genome. PLoS Genet 5:e1000733
Liu Y, Deng YT, Li G, Zhao J (2013) Replication factor C1 (RFC1) is required for double-strand break repair during meiotic homologous recombination in Arabidopsis. Plant J 73:154–165
Loidl J (1989) Effects of elevated-temperature on meiotic chromosome synapsis in Allium ursinum. Chromosoma 97:449–458
Lu PL, Han XW, Qi J, Yang JG, Wijeratne AJ, Li T, Ma H (2012) Analysis of Arabidopsis genome-wide variations before and after meiosis and meiotic recombination by resequencing Landsberg erecta and all four products of a single meiosis. Genome Res 22:508–518
Lucht JM, Mauch-Mani B, Steiner HY, Metraux JP, Ryals J, Hohn B (2002) Pathogen stress increases somatic recombination frequency in Arabidopsis. Nat Genet 30:311–314
Lukaszewski AJ (1992) A comparison of physical distribution of recombination in chromosome 1R in diploid rye and in hexaploid triticale. Theor Appl Genet 83:1048–1053
Luo Q, Tang D, Wang M, Luo WX, Zhang L, Qin BX, Shen Y, Wang KJ, Li YF, Cheng ZK (2013) The role of OsMSH5 in crossover formation during rice meiosis. Mol Plant 6:729–742
Ma X, Fan L, Zhang ZF, Yang X, Liu YC, Ma YM, Pan Y, Zhou GA, Zhang M, Ning HL, Kong FJ, Ma JK, Liu SL, Tian ZX (2023) Global dissection of the recombination landscape in soybean using a high-density 600K SoySNP array. Plant Biotechnol J 21:606–620
Maleki S, Neale MJ, Arora C, Henderson KA, Keeney S (2007) Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae. Chromosoma 116:471–486
Mancera E, Bourgon R, Brozzi A, Huber W, Steinmetz LM (2008) High-resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature 454:479-U471
Mao YF, Botella JR, Liu YG, Zhu JK (2019) Gene editing in plants: progress and challenges. Natl Sci Rev 6:421–437
Marand AP, Jansky SH, Zhao HN, Leisner CP, Zhu XB, Zeng ZX, Crisovan E, Newton L, Hamernik AJ, Veilleux RE, Buell CR, Jiang JM (2017) Meiotic crossovers are associated with open chromatin and enriched with Stowaway transposons in potato. Genome Biol 18:1–6
Martinez-Perez E, Colaiacovo MP (2009) Distribution of meiotic recombination events: talking to your neighbors. Curr Opin Genet Dev 19:105–112
Martini E, Diaz RL, Hunter N, Keeney S (2006) Crossover homeostasis in yeast meiosis. Cell 126:285–295
McConaughy S, Amundsen K, Song QJ, Pantalone V, Hyten D (2023) Recombination hotspots in soybean [Glycine max (L.) Merr.]. G3 Genes Genomes Genet 13:075
McMahill MS, Sham CW, Bishop DK (2007) Synthesis-dependent strand annealing in meiosis. PLoS Biol 5:2589–2601
Melamed-Bessudo C, Levy AA (2012) Deficiency in DNA methylation increases meiotic crossover rates in euchromatic but not in heterochromatic regions in Arabidopsis. P Natl Acad Sci USA 109:E981–E988
Mercier R, Jolivet S, Vezon D, Huppe E, Chelysheva L, Giovanni M, Nogue F, Doutriaux MP, Horlow C, Grelon M, Mezard C (2005) Two meiotic crossover classes cohabit in Arabidopsis: one is dependent on MER3, whereas the other one is not. Curr Biol 15:692–701
Mercier R, Mezard C, Jenczewski E, Macaisne N, Grelon M (2015) The molecular biology of meiosis in plants. Annu Rev Plant Biol 66(66):297–327
Miao CB, Tang D, Zhang HG, Wang M, Li YF, Tang SZ, Yu HX, Gu MH, Cheng ZK (2013) Central region component1, a novel synaptonemal complex component, is essential for meiotic recombination initiation in rice. Plant Cell 25:2998–3009
Mickelbart MV, Hasegawa PM, Bailey-Serres J (2015) Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat Rev Genet 16:237–251
Mirouze M, Lieberman-Lazarovich M, Aversano R, Bucher E, Nicolet J, Reinders J, Paszkowski J (2012) Loss of DNA methylation affects the recombination landscape in Arabidopsis. P Natl Acad Sci USA 109:5880–5885
Mitchel K, Zhang HS, Welz-Voegele C, Jinks-Robertson S (2010) Molecular structures of crossover and noncrossover intermediates during gap repair in yeast: implications for recombination. Mol Cell 38:211–222
Molinier J, Ries G, Zipfel C, Hohn B (2006) Transgeneration memory of stress in plants. Nature 442:1046–1049
Muller HJ (1916) The mechanism of crossing-over. Am Nat 50:193–221
Murakami H, Keeney S (2008) Regulating the formation of DNA double-strand breaks in meiosis. Gene Dev 22:286–292
Myers S, Bottolo L, Freeman C, McVean G, Donnelly P (2005) A fine-scale map of recombination rates and hotspots across the human genome. Science 310:321–324
Neale MJ, Pan J, Keeney S (2005) Endonucleolytic processing of covalent protein-linked DNA double-strand breaks. Nature 436:1053–1057
Nichols MD, DeAngelis K, Keck JL, Berger JM (1999) Structure and function of an archaeal topoisomerase VI subunit with homology to the meiotic recombination factor Spo11. EMBO J 18:6177–6188
Nimonkar AV, Sica RA, Kowalczykowski SC (2009) Rad52 promotes second-end DNA capture in double-stranded break repair to form complement-stabilized joint molecules. P Natl Acad Sci USA 106:3077–3082
Nonomura KL, Nakano M, Fukuda T, Eiguchi M, Miyao A, Hirochika H, Kurata N (2004) The novel gene HOMOLOGOUS PAIRING ABERRATION IN RICE MEIOSIS1 of rice encodes a putative coiled-coil protein required for homologous chromosome pairing in meiosis. Plant Cell 16:1008–1020
Osman K, Higgins JD, Sanchez-Moran E, Armstrong SJ, Franklin FC (2011) Pathways to meiotic recombination in Arabidopsis thaliana. New Phytol 190:523–544
Page SL, Hawley RS (2004) The genetics and molecular biology of the synaptonemal complex. Annu Rev Cell Dev Biol 20:525–558
Paigen K, Petkov P (2010) Mammalian recombination hot spots: properties, control and evolution. Nat Rev Genet 11:221–233
Pan J, Sasaki M, Kniewel R, Murakami H, Blitzblau HG, Tischfield SE, Zhu X, Neale MJ, Jasin M, Socci ND, Hochwagen A, Keeney S (2011) A hierarchical combination of factors shapes the genome-wide topography of yeast meiotic recombination initiation. Cell 144:719–731
Pannunzio NR, Watanabe G, Lieber MR (2018) Nonhomologous DNA end-joining for repair of DNA double-strand breaks. J Biol Chem 293:10512–10523
Panoli AP, Ravi M, Sebastian J, Nishal B, Reddy TV, Marimuthu MPA, Subbiah V, Vijaybhaskar V, Siddiqi I (2006) AtMND1 is required for homologous pairing during meiosis in Arabidopsis. Bmc Mol Biol 7:1–12
Paques F, Haber JE (1999) Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol Mol Biol R 63:349–404
Parvanov ED, Petkov PM, Paigen K (2010) Prdm9 controls activation of mammalian recombination hotspots. Science 327:835–835
Paul P, Nag D, Chakraborty S (2016) Recombination hotspots: models and tools for detection. DNA Repair (amst) 40:47–56
Pawlowski WP, Golubovskaya IN, Cande WZ (2003) Altered nuclear distribution of recombination protein RAD51 in maize mutants suggests the involvement of RAD51 in meiotic homology recognition. Plant Cell 15:1807–1816
Pawlowski WP, Golubovskaya IN, Timofejeva L, Meeley RB, Sheridan WF, Cande WZ (2004) Coordination of meiotic recombination, pairing, and synapsis by PHS1. Science 303:89–92
Petes TD (2001) Meiotic recombination hot spots and cold spots. Nat Rev Genet 2:360–369
Phillips D, Nibau C, Ramsay L, Waugh R, Jenkins G (2010) Development of a molecular cytogenetic recombination assay for barley. Cytogenet Genome Res 129:154–161
Phillips D, Jenkins G, Macaulay M, Nibau C, Wnetrzak J, Fallding D, Colas I, Oakey H, Waugh R, Ramsay L (2015) The effect of temperature on the male and female recombination landscape of barley. New Phytol 208:421–429
Podevin N, Davies HV, Hartung F, Nogue F, Casacuberta JM (2013) Site-directed nucleases: a paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol 31:375–383
Puchta H, Fauser F (2013) Gene targeting in plants: 25 years later. Int J Dev Biol 57:629–637
Puizina J, Siroky J, Mokros P, Schweizer D, Riha K (2004) Mre11 deficiency in Arabidopsis is associated with chromosomal instability in somatic cells and Spo11-dependent genome fragmentation during meiosis. Plant Cell 16:1968–1978
Ribeiro J, Abby E, Livera G, Martini E (2016) RPA homologs and ssDNA processing during meiotic recombination. Chromosoma 125:265–276
Rice WR (2002) Experimental tests of the adaptive significance of sexual recombination. Nat Rev Genet 3:241–251
Robert T, Nore A, Brun C, Maffre C, Crimi B, Bourbon HM, de Massy B (2016) The TopoVIB-Like protein family is required for meiotic DNA double-strand break formation. Science 351:943–949
Rodgers-Melnick E, Bradbury PJ, Elshire RJ, Glaubitz JC, Acharya CB, Mitchell SE, Li CH, Li YX, Buckler ES (2015) Recombination in diverse maize is stable, predictable, and associated with genetic load. P Natl Acad Sci USA 112:3823–3828
Ronceret A, Doutriaux MP, Golubovskaya IN, Pawlowski WP (2009) PHS1 regulates meiotic recombination and homologous chromosome pairing by controlling the transport of RAD50 to the nucleus. P Natl Acad Sci USA 106:20121–20126
Rosu S, Libuda DE, Villeneuve AM (2011) Robust crossover assurance and regulated interhomolog access maintain meiotic crossover number. Science 334:1286–1289
Saintenac C, Falque M, Martin OC, Paux E, Feuillet C, Sourdille P (2009) Detailed recombination studies along chromosome 3B provide new insights on crossover distribution in wheat (Triticum aestivum L.). Genetics 181:393–403
Saintenac C, Faure S, Remay A, Choulet F, Ravel C, Paux E, Balfourier F, Feuillet C, Sourdille P (2011) Variation in crossover rates across a 3-Mb contig of bread wheat (Triticum aestivum) reveals the presence of a meiotic recombination hotspot. Chromosoma 120:185–198
Salome PA, Bomblies K, Fitz J, Laitinen RAE, Warthmann N, Yant L, Weigel D (2012) The recombination landscape in Arabidopsis thaliana F-2 populations. Heredity 108:447–455
Sanchez-Moran E, Armstrong SJ, Santos JL, Franklin FCH, Jones GH (2002) Variation in chiasma frequency among eight accessions of Arabidopsis thaliana. Genetics 162:1415–1422
Sanchez-Moran E, Santos JL, Jones GH, Franklin FCH (2007) ASY1 mediates AtDMC1-dependent interhomolog recombination during meiosis in Arabidopsis. Gene Dev 21:2220–2233
Santos TDL, Hunter N, Lee C, Larkin B, Loidl J, Hollingsworth NM (2003) The Mus81/Mms4 endonuclease acts independently of double-Holliday junction resolution to promote a distinct subset of crossovers during meiosis in budding yeast. Genetics 164:81–94
Schnable PS, Hsia AP, Nikolau BJ (1998) Genetic recombination in plants. Curr Opin Plant Biol 1:123–129
Schramm S, Fraune J, Naumann R, Hernandez-Hernandez A, Hoog C, Cooke HJ, Alsheimer M, Benavente R (2011) A novel mouse synaptonemal complex protein is essential for loading of central element proteins, recombination, and fertility. PLoS Genet 7:e1002088
Seguela-Arnaud M, Crismani W, Larcheveque C, Mazel J, Froger N, Choinard S, Lemhemdi A, Macaisne N, Van Leene J, Gevaert K, De Jaeger G, Chelysheva L, Mercier R (2015) Multiple mechanisms limit meiotic crossovers: TOP3 alpha and two BLM homologs antagonize crossovers in parallel to FANCM. P Natl Acad Sci USA 112:4713–4718
Seguela-Arnaud M, Choinard S, Larcheveque C, Girard C, Froger N, Crismani W, Mercier R (2017) RMI1 and TOP3 alpha limit meiotic CO formation through their C-terminal domains. Nucleic Acids Res 45:1860–1871
Shen Y, Tang D, Wang KJ, Wang M, Huang J, Luo WX, Luo Q, Hong LL, Li M, Cheng ZK (2012) ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis. J Cell Sci 125:2581–2591
Shen C, Li XM, Zhang RT, Lin ZX (2017) Genome-wide recombination rate variation in a recombination map of cotton. PLoS ONE 12:e0188682
Shifman S, Bell JT, Copley RR, Taylor MS, Williams RW, Mott R, Flint J (2006) A high-resolution single nucleotide polymorphism genetic map of the mouse genome. PLoS Biol 4:2227–2237
Shilo S, Melamed-Bessudo C, Dorone Y, Barkai N, Levy AA (2015) DNA crossover motifs associated with epigenetic modifications delineate open chromatin regions in Arabidopsis. Plant Cell 27:2427–2436
Shingu Y, Mikawa T, Onuma M, Hirayama T, Shibata T (2010) A DNA-binding surface of SPO11-1, an Arabidopsis SPO11 orthologue required for normal meiosis. FEBS J 277:2360–2374
Shinohara A, Ogawa H, Ogawa T (1992) Rad51 protein involved in repair and recombination in S. cerevisiae is a RecA-like protein. Cell 69:457–470
Shinohara M, Oh SD, Hunter N, Shinohara A (2008) Crossover assurance and crossover interference are distinctly regulated by the ZMM proteins during yeast meiosis. Nat Genet 40:299–309
Shultz RW, Tatineni VM, Hanley-Bowdoin L, Thompson WF (2007) Genome-wide analysis of the core DNA replication machinery in the higher plants Arabidopsis and rice. Plant Physiol 144:1697–1714
Siaud N, Dray E, Gy I, Gerard E, Takvorian N, Doutriaux MP (2004) Brca2 is involved in meiosis in Arabidopsis thaliana as suggested by its interaction with Dmc1. Embo J 23:1392–1401
Sidhu GK, Fang C, Olson MA, Falque M, Martin OC, Pawlowski WP (2015) Recombination patterns in maize reveal limits to crossover homeostasis. P Natl Acad Sci USA 112:15982–15987
Slotkin RK, Martienssen R (2007) Transposable elements and the epigenetic regulation of the genome. Nat Rev Genet 8:272–285
Smagulova F, Gregoretti IV, Brick K, Khil P, Camerini-Otero RD, Petukhova GV (2011) Genome-wide analysis reveals novel molecular features of mouse recombination hotspots. Nature 472:375–378
Soustelle C, Vedel M, Kolodner R, Nicolas A (2002) Replication protein A is required for meiotic recombination in Saccharomyces cerevisiae. Genetics 161:535–547
Stacey NJ, Kuromori T, Azumi Y, Roberts G, Breuer C, Wada T, Maxwell A, Roberts K, Sugimoto-Shirasu K (2006) Arabidopsis SPO11-2 functions with SPO11-1 in meiotic recombination. Plant J 48:206–216
Stapley J, Feulner PGD, Johnston SE, Santure AW, Smadja CM (2017) Recombination: the good, the bad and the variable. Philos Trans R Soc B Biol Sci 372:20170279
Su H, Cheng ZH, Huang JY, Lin J, Copenhaver GP, Ma H, Wang YX (2017) Arabidopsis RAD51, RAD51C and XRCC3 proteins form a complex and facilitate RAD51 localization on chromosomes for meiotic recombination. PLoS Genet 13:e1006827
Sugiyama T, Kantake N, Wu Y, Kowalczykowski SC (2006) Rad52-mediated DNA annealing after Rad51-mediated DNA strand exchange promotes second ssDNA capture. EMBO J 25:5539–5548
Szostak JW, Orrweaver TL, Rothstein RJ, Stahl FW (1983) The double-strand-break repair model for recombination. Cell 33:25–35
Taiz L, Zeiger E, Møller IM, Murphy AS (2015) Plant physiology and development, 6th edition. Sinauer Associates Inc., Sunderland
Tang D, Miao CB, Li YF, Wang HJ, Liu XF, Yu HX, Cheng ZK (2014) OsRAD51C is essential for double-strand break repair in rice meiosis. Front Plant Sci 5:86630
Tang Y, Yin ZN, Zeng YJ, Zhang QX, Chen LQ, He Y, Lu PL, Ye D, Zhang XQ (2017) MTOPVIB interacts with AtPRD1 and plays important roles in formation of meiotic DNA double-strand breaks in Arabidopsis. Sci Rep 7:10007
Uanschou C, Siwiec T, Pedrosa-Harand A, Kerzendorfer C, Sanchez-Moran E, Novatchkova M, Akimcheva S, Woglar A, Klein F, Schlogelhofer P (2007) A novel plant gene essential for meiosis is related to the human CtIP and the yeast COM1/SAE2 gene. EMBO J 26:5061–5070
Uanschou C, Ronceret A, Von Harder M, De Muyt A, Vezon D, Pereira L, Chelysheva L, Kobayashi W, Kurumizaka H, Schlogelhofer P, Grelon M (2013) Sufficient amounts of functional HOP2/MND1 complex promote interhomolog DNA repair but are dispensable for intersister DNA repair during meiosis in Arabidopsis. Plant Cell 25:4924–4940
Vignard J, Siwiec T, Chelysheva L, Vrielynck N, Gonord F, Armstrong SJ, Schlogelhofer P, Mercier R (2007) The interplay of RecA-related proteins and the MND1-HOP2 complex during meiosis in Arabidopsis thaliana. PLoS Genet 3:1894–1906
Voytas DF (2013) Plant genome engineering with sequence-specific nucleases. Annu Rev Plant Biol 64:327–350
Voytas DF, Gao C (2014) Precision genome engineering and agriculture: opportunities and regulatory challenges. PLoS Biol 12:e1001877
Vrielynck N, Chambon A, Vezon D, Pereira L, Chelysheva L, De Muyt A, Mezard C, Mayer C, Grelon M (2016) A DNA topoisomerase VI-like complex initiates meiotic recombination. Science 351:939–943
Wang YX, Copenhaver GP (2018) Meiotic recombination: mixing it up in plants. Annu Rev Plant Biol 69:577–609
Wang X, Haber JE (2004) Role of Saccharomyces single-stranded DNA-binding protein RPA in the strand invasion step of double-strand break repair. PLoS Biol 2:104–112
Wang KJ, Wang M, Tang D, Shen Y, Miao CB, Hu Q, Lu TG, Cheng ZK (2012) The role of rice HEI10 in the formation of meiotic crossovers. PLoS Genet 8:e1002809
Wang YX, Xiao R, Wang HF, Cheng ZH, Li WX, Zhu GF, Wang Y, Ma H (2014) The Arabidopsis RAD51 paralogs RAD51B, RAD51D and XRCC2 play partially redundant roles in somatic DNA repair and gene regulation. New Phytol 201:292–304
Wang X, Lu J, Lao K, Wang S, Mo X, Xu X, Chen X, Mo B (2019) Increasing the efficiency of CRISPR/Cas9-based gene editing by suppressing RNAi in plants. Sci China Life Sci 62:982–984
Wang Y, Dong Z, Ma Y, Zheng Y, Huang S, Yang X (2023) Comprehensive dissection of meiotic DNA double-strand breaks and crossovers in cucumber. Plant Physiol 193:1913–1932
Waterworth WM, Altun C, Armstrong SJ, Roberts N, Dean PJ, Young K, Weil CF, Bray CM, West CE (2007) NBS1 is involved in DNA repair and plays a synergistic role with ATM in mediating meiotic homologous recombination in plants. Plant J 52:41–52
Williams RS, Williams JS, Tainer JA (2007) Mre11-Rad50-Nbs1 is a keystone complex connecting DNA repair machinery, double-strand break signaling, and the chromatin template. Biochem Cell Biol 85:509–520
Wold MS (1997) Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem 66:61–92
Wu TC, Lichten M (1994) Meiosis-induced double-strand break sites determined by yeast chromatin structure. Science 263:515–518
Wyatt HDM, West SC (2014) Holliday junction resolvases. Cold Spring Harb Perspect Biol 6:a023192
Xu LH, Kleckner N (1995) Sequence non-specific double-strand breaks and interhomolog interactions prior to double-strand break formation at a meiotic recombination hot spot in yeast. EMBO J 14:5115–5128
Xue ZH, Li YF, Zhang L, Shi WQ, Zhang C, Feng MS, Zhang FF, Tang D, Yu HX, Gu MH, Cheng ZK (2016) OsMTOPVIB promotes meiotic DNA double-strand break formation in rice. Mol Plant 9:1535–1538
Yao H, Zhou Q, Li J, Smith H, Yandeau M, Nikolau BJ, Schnable PS (2002) Molecular characterization of meiotic recombination across the 140-kb multigenic a1-sh2 interval of maize. P Natl Acad Sci USA 99:6157–6162
Yokoo R, Zawadzki KA, Nabeshima K, Drake M, Arur S, Villeneuve AM (2012) COSA-1 reveals robust homeostasis and separable licensing and reinforcement steps governing meiotic crossovers. Cell 149:75–87
Yu HX, Wang M, Tang D, Wang KJ, Chen FL, Gong ZY, Gu MH, Cheng ZK (2010) OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice. Chromosoma 119:625–636
Zelkowski M, Olson MA, Wang MH, Pawlowski W (2019) Diversity and determinants of meiotic recombination landscapes. Trends Genet 35:359–370
Zhang LS, Ma H (2012) Complex evolutionary history and diverse domain organization of SET proteins suggest divergent regulatory interactions. New Phytol 195:248–263
Zhang C, Song Y, Cheng ZH, Wang YX, Zhu J, Ma H, Xu L, Yang ZN (2012) The Arabidopsis thaliana DSB formation (AtDFO) gene is required for meiotic double-strand break formation. Plant J 72:271–281
Zhou SR, Wang Y, Li WC, Zhao ZG, Ren YL, Wang Y, Gu SH, Lin QB, Wang D, Jiang L, Su N, Zhang X, Liu LL, Cheng ZJ, Lei CL, Wang JL, Guo XP, Wu FQ, Ikehashi H, Wang HY, Wan JM (2011) Pollen Semi-Sterility1 encodes a Kinesin-1-Like protein important for male meiosis, anther dehiscence, and fertility in rice. Plant Cell 23:111–129
Zickler D, Kleckner N (2015) Recombination, pairing, and synapsis of homologs during meiosis. Cold Spring Harb Perspect Biol 7:a016626
Acknowledgements
This work was supported by the postgraduate scholarship from University of Tasmania and Grain Research & Development Corporation (GRDC).
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Grains Research and Development Corporation
Author information
Authors and Affiliations
Contributions
MLZ conducted the data collection and analysis and wrote the manuscript. CZ, SS, and MXZ revised the manuscript. MXZ designed and supervised the research. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Communicated by Rajeev K. Varshney.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zou, M., Shabala, S., Zhao, C. et al. Molecular mechanisms and regulation of recombination frequency and distribution in plants. Theor Appl Genet 137, 86 (2024). https://doi.org/10.1007/s00122-024-04590-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00122-024-04590-4