
Abstract
Background
Whey is just turning from a waste of milk processing to a renewable raw material in biotechnology for producing single-cell protein, bio-ethanol, or ethyl acetate as an economic alternative. Conversion of whey-borne sugar into ethyl acetate requires yeast biomass as a biocatalyst. A high cell concentration results in a quick ester synthesis, but biomass growth means consumption of sugar at the expense of ester production. Efficient and cost-saving biomass production is thus a practical requirement. Whey is poor in nitrogen and has therefore to be supplemented with a bioavailable N source.
Methods
Several aerobic growth tests were performed with Kluyveromyces marxianus DSM 5422 as a potent producer of ethyl acetate in whey-borne media supplemented with various N sources. Preliminary tests were done in shake flasks while detailed studies were performed in a stirred bioreactor.
Results
Ammonium sulfate resulted in strong acidification due to remaining sulfate, but costly pH control increases the salt load, being inhibitory to yeasts and causing environmental impacts. Ammonium carbonate lessened acidification, but its supplement increased the initial pH to 7.5 and delayed growth. Urea as an alternative N source was easily assimilated by the studied yeast and avoided strong acidification (much less base was required for pH control). Urea was assimilated intracellularly rather than hydrolyzed extracellularly by urease. Conversion of urea to ammonium and usage of formed ammonium for biomass production occurred with a similar rate so that the amount of excreted ammonium was small. Ammonium hydroxide as another N source was successfully added by the pH controller during the growth of K. marxianus DSM 5422, but the medium had to be supplemented with some ammonium sulfate to avoid sulfur limitation and to initiate acidification. Non-limited growth resulted in 82 mg N per g of biomass, but N-limited growth diminished the N content.
Conclusions
K. marxianus could be efficiently produced by supplementing the whey with nitrogen. Urea and ammonia were the favored N sources due to the proton neutrality at assimilation which lessened the salt load and reduced the supply of alkali for pH control or made this even needless.
Similar content being viewed by others

Background
Sustainable and environmentally compatible development requires successive substitution of fossil resources by renewable raw materials. This applies to the energy sector but is also true for production of industrial bulk materials. Such a bulk chemical is ethyl acetate with an annual world production of 1.7 million tons [1]. Ethyl acetate is an organic solvent of moderate polarity with versatile industrial applications. Another prospective application is the biodiesel production from vegetable oil; here, triglycerides are transformed to fatty acid ethyl esters in a lipase-catalyzed transesterification reaction with ethyl acetate as an acyl acceptor instead of methanol [2-5]. Although being an irritant and intoxicant at higher concentrations, ethyl acetate is less toxic to humans compared to many other solvents. Ethyl acetate is an environmentally friendly compound since the ester is easily degraded by bacteria [6-8] and is regarded as a non-persistent pollutant of the atmosphere [9].
Synthesis of ethyl acetate currently proceeds by petrochemical processes, which are based on crude oil constituents or natural gas, run at elevated temperature and pressure and commonly require catalysts [9]. The reactions are often incomplete, and the recovery of ethyl acetate and residual precursors needs a high input of energy.
Microbial production of ethyl acetate from renewables could become an interesting alternative. Various yeast species can synthesize ethyl acetate (reviewed by Löser et al. [9]), but only Pichia anomala, Candida utilis, and Kluyveromyces marxianus produce this ester in larger amounts. K. marxianus is the most promising candidate for large-scale ester production since this dairy yeast with GRAS status grows quickly, converts sugar directly into ethyl acetate without ethanol as an essential intermediate, and produces the ester with a high rate and yield [9-17]. The ester synthesis in K. marxianus is easy to control by the level of iron [11,13,16-18]. K. marxianus exhibits a distinct thermal tolerance which allows cultivation at an elevated temperature [14,19,20] which in turn accelerates the ester stripping and advances its process-integrated recovery. The outstanding capability of K. marxianus for lactose utilization offers the chance for using whey as a resource of ester synthesis. K. marxianus converts whey-borne lactose with a high yield into ethyl acetate [9], but its ability for sucrose and glucose assimilation [19-21] enables production of ethyl acetate with this yeast from renewables like sugarcane, grain, and corn.
Ethanol is another product of microbial sugar conversion, but several factors favor microbial ester over ethanol production [22]: the higher market price of the ester, a reduced number of process stages, a faster process, and a cost-saving product recovery.
Conversion of sugar into ethyl acetate requires yeast biomass as a biocatalyst. A high biomass concentration results in a quick process, but the production of this biomass, on the other hand, is connected with sugar consumption which reduces the portion of sugar available for ester production (as demonstrated in pilot-scale experiments [13]). The right balance between yeast growth and ester synthesis or, in other words, a compromise between a quick process and a high ester yield is of practical importance. This balance can be controlled by the available iron [13,16,23].
An efficient production of the required yeast biomass is an important factor for the economy of the total process of ester and ethanol production. At whey-based bio-ethanol production, the biomass is often considered as a gift and sugar utilization for biomass production is ignored. Such an ‘out-sourcing’ of biomass processing only seemingly improves the economy.
Much research was done in the field of whey-based K. marxianus cultivation for single-cell protein production [24-28]. The results refer to some problems at cultivation of K. marxianus in whey-based media; whey is poor in bioavailable nitrogen so that nitrogen can limit yeast growth [26-29]. Nitrogen-limited growth can even deregulate yeast metabolism and provoke ethanol formation at aerobic conditions [30].
Whey was often supplemented with ammonium as a source of nitrogen to stimulate growth of K. marxianus [10-14,16,27,29,31-37]. Added ammonium increased the yield of biomass [27,32,35] or was without significant effect [29,31,36]. Ammonium is usually supplemented in the form of ammonium sulfate where ammonium is intensively consumed while most of the sulfate remains in the medium and causes an ionic imbalance and acidification [15,38-41]. Such acidification can be inhibitory to K. marxianus since its growth rate is distinctly reduced at pH ≤3.5 [22,28,42,43]. Supplementation of whey with (NH4)2SO4 or NH4Cl requires cost-intensive pH correction with NaOH or KOH which in turn increases the salt load and thus inhibits yeast growth [31,42] and creates waste-water problems.
Urea is an alternative N source since growth with urea exhibits proton neutrality without significant pH changes [38-40]. Assimilation of urea by yeasts is not an exception but the rule; 122 of 123 tested yeasts were able to metabolize urea [44]. There are several potential advantages of urea [25,28,39]: a high amount of nitrogen per unit weight, a low price, and a reduced or even omitted supply of pH correctives. K. marxianus definitely metabolizes urea, and whey was repeatedly supplemented with urea as an N source for this yeast [28,31,32,37,45-47]. However, the effect of added urea on growth was often not described [37,45,46], or the published results were contradictory. Yadav et al. [28] observed an increased biomass yield, Kar and Misra [32] found no positive effect, Mahmoud and Kosikowski [31] described slight inhibition by added urea, and Rech et al. [47] even detected strong inhibition of growth which had been attributed to alkalinization. These inconsistent results require clarification by more detailed studies on this subject.
The first objective of this work was testing the effect of (NH4)2SO4, (NH4)2CO3, or urea as sources of nitrogen at aerobic growth of K. marxianus DSM 5422 in whey-borne medium with special attention on yeast growth and acidification. When using (NH4)2SO4, sulfate remains in the medium and causes proton imbalance and acidification but, when using (NH4)2CO3, carbonate disappears in form of CO2 during cultivation which possibly avoids acidification. The second objective was studying the growth of K. marxianus DSM 5422 in whey-borne medium with urea in detail at defined conditions to get a deeper insight into the urea metabolism. The third objective was testing the pH-controlled feed of ammonia during aerobic growth of K. marxianus DSM 5422 in whey-borne medium. Ammonia as the cheapest source of nitrogen is interesting for large-scale processes.
Methods
Microorganism
K. marxianus DSM 5422 from the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (Braunschweig, Germany) was maintained with the Cryoinstant preservation system (Lomb Scientific Pty Ltd, Vienna, Austria), cultivated on yeast-glucose-chloramphenicol agar (Roth GmbH, Karlsruhe, Germany) for 2 days at 32°C, and then used as an inoculum.
Non-supplemented DW medium
All media originate from concentrated and partially demineralized sweet whey. Sweet whey was ultrafiltrated, concentrated by reverse osmosis, and then demineralized by slight alkalinization and moderate heating to yield the used whey permeate (thus processed in the Sachsenmilch Leppersdorf GmbH, Leppersdorf, Germany). Non-supplemented DW medium was prepared in batches of 1 L by mixing 0.5 L whey permeate with the same volume water. This mixture was autoclaved in a sealed 1-L Schott bottle for 15 min at 121°C. Precipitated minerals were allowed to settle overnight before the upper phase was withdrawn for cultivation experiments. Non-supplemented DW medium should not be confused with DW basic medium (as used in [11-14,16]) which contains 10 g/L (NH4)2SO4.
Shake-flask cultivation
In a first series of shake-flask experiments, various media were prepared like the just-described non-supplemented DW medium with the modification that various sources of nitrogen (10 g/L (NH4)2SO4, 10 g/L (NH4)2CO3, or 5 g/L urea) were added before medium sterilization. The media handling (autoclaving, settlement, withdrawal) occurred as in the above-described manner. These media were supplemented with 2 mL/L autoclaved trace-element solution (preparation as described in [10]) and diluted with water to 1/20 for reducing the content of sugar to 3.9 g/L and, thus, to avoid oxygen-limited growth at a low oxygen-transfer rate in shake flasks. Several 500-mL conical flasks with cotton plugs were filled each with 50 mL medium, inoculated with an agar-plate culture as described in [14], and shaken with 250 rpm at 32°C. After 14 h of pre-cultivation, the process was followed by regular analysis of the optical density (OD) and the pH value over a period of at least 10 h. After a total of 40 h of cultivation, the OD and pH were measured and the residual sugar and formed biomass were analyzed.
A second series of shake-flask experiments was based on autoclaved non-supplemented DW medium. Portions of this medium were supplemented with 2 mL/L sterile trace-element solution and separately autoclaved aqueous stock solutions of (NH4)2SO4, (NH4)2CO3, or urea. The pH was then adjusted to 6.5 with 0.1 M HCl or KOH. These media were diluted with water to reduce the sugar content to 3.9 g/L like in the first series. Discrete sterilization of whey and N sources in this second series avoided unwanted interaction between whey constituents and N compounds such as the Maillard reaction. Inoculation, cultivation, and analyses occurred as in the first series.
Bioreactor cultivation
One reference experiment was done with DW basic medium (DW medium with 10 g/L (NH4)2SO4) and described in detail in [11] while all other presented bioreactor processes were based on non-supplemented DW medium. The sterile 1-L stirred bioreactor with 0.6 mL Antifoam A (Fluka, Sigma Aldrich, St. Louis, USA) was filled with 0.6 L DW medium and 1.2 mL trace-element solution (the latter described in [10]). Further supplements such as (NH4)2SO4, urea, and Na2SO4 were put into 100-mL Schott bottles, autoclaved for 15 min at 121°C, and then transferred to the bioreactor as follows: the bottle was connected with the bioreactor, some culture medium was pumped to the bottle, and, after complete dissolution of the substance, transported back to the reactor. Separate autoclaving avoided unwanted interaction such as the Maillard reaction and sulfate precipitation.
The cultivation occurred as usual [10-12,14]: the medium was inoculated with five loops of biomass, and the reactor was operated at 1,200 rpm and 32°C and gassed with 50 L/h dry and CO2-free air (given for 0°C and 101,325 Pa). The pH was controlled to ≥5 with 2 M KOH or 2 M ammonia solution. All processes lasted at least 36 h. Sampling and sample preparation were performed as usual [10].
Test for urea assimilation
K. marxianus DSM 5422 was cultivated in diluted DW medium with urea (3.9 g/L sugars, 0.25 g/L urea) as described for the second series of shake-flask experiments. After depletion of the sugar, the cell suspension was separated into biomass and supernatant by centrifugation. The biomass was suspended in 50-mM phosphate buffer at pH 6 containing 0.25 g/L urea to yield an initial biomass concentration of 2 g/L, and the supernatant was supplemented with urea to give a concentration of 0.25 g/L. Both mixtures were then shaken with 250 rpm at 32°C. Sampling occurred at the beginning and after 24 h of incubation. The samples were analyzed regarding urea and ammonium.
Analyses
The optical density of cell suspensions was measured photometrically at 600 nm (after pre-dilution to OD <0.4 if required). The biomass dry weight was determined by separating the yeasts via centrifugation, washing the pellet twice, and drying at 103°C. Sugar was quantified by a modified 3,5-dinitrosalicylic-acid method [48] with lactose as a standard. Ammonium, nitrate, nitrite, sulfate, and phosphate were measured by the LCK303, LCK339, LCK342, LCK153, and LCK049 cuvette tests (Hach Lange GmbH, Düsseldorf, Germany). The total dissolved nitrogen was analyzed by the Kjeldahl method (German DIN 38409 H11). Ethanol was quantified by gas chromatography [10].
Urea was measured in the style of Rahmatullah and Boyde [49] with some modifications. This method had been approved for quantifying urea in wine [50] being a matrix similar to culture liquids. Two reagents were prepared: a mixture of 300 mL sulfuric acid (95% to 98%, ρ = 1,840 g/L), 100 mL phosphoric acid (85%, ρ = 1,670 g/L), and 100 mL water; and a solution of 500 mg butane-2,3-dione monoxime (diacetyl monoxime) and 10 mg thiosemicarbazide in 100 mL water. The analysis reagent was prepared from 40 mL acid mixture and 20 mL of the second solution and used immediately. A volume of 0.1 mL sample was mixed with 3 mL analysis reagent. In the case of colored samples, this mixture was measured photometrically at 525 nm as a blind. Then, the mixture was put in a 25-mL test tube with plastic cap and incubated for 20 min in boiling water. After cooling and short mixing, the absorbance was measured at 525 nm. The blind was subtracted, and the obtained ΔA525nm value was interpreted as a urea concentration by a non-linear calibration curve prepared for several solutions of urea in water (from 0 to 500 mg/L): C Urea,L = a · (ΔA525nm)b with a = 520 mg/L and b = 1.37.
Data analyses
The O2 consumption, CO2 formation, and the respiratory quotient (RQ) were calculated as usual [11]. The time-dependent specific growth rate was calculated from masses of formed CO2 assuming a correlation between yeast growth and CO2 formation which only applies to respiratory processes without significant maintenance: μ(t) ≈ Δln(m CO2(t))/Δt. The overall biomass yield (Y X/S) is the ratio between the mass of yeasts grown and the mass of sugar consumed; determination of these masses took losses by sampling and changes of the liquid volume into account. The pH-controlled feeding of 2 M KOH is given as a specific volume, related to the initial liquid volume of the culture. The mass of fed KOH was related to the formed biomass (given as gKOH/gX). The fed 2 M ammonia solution is also expressed as a specific volume and as the mass of supplied ammonia-N. The mass of bioavailable nitrogen was calculated as the sum of initial ammonium-N and urea-N plus the mass of ammonia-N supplied till a given time: \( {m}_{\mathrm{N}}(t)={V}_{\mathrm{L}}\left({t}_0\right)\cdot \left({C}_{{\mathrm{N}\mathrm{H}}_4-\mathrm{N}}\left({t}_0\right)+{\mathrm{C}}_{\mathrm{Urea}\hbox{-} \mathrm{N}}\left({t}_0\right)\right)+{m}_{{\mathrm{N}\mathrm{H}}_4\mathrm{O}\mathrm{H}\hbox{-} \mathrm{N}}(t) \). The consumed nitrogen is this m N(t) value minus the nitrogen not yet used or lost by sampling: \( \Delta {m}_{\mathrm{N}}(t)\kern0.5em =\kern0.5em {m}_{\mathrm{N}}(t)\kern0.5em -\kern0.5em {V}_{\mathrm{L}}(t)\kern0.5em \cdot \left({C}_{{\mathrm{N}\mathrm{H}}_4\hbox{-} \mathrm{N}}(t)+{C}_{\mathrm{Urea}\hbox{-} \mathrm{N}}(t)\right)\kern0.5em -\kern0.5em \varSigma {m}_{\mathrm{N}}\left(\mathrm{sampling}\right) \). These Δm N(t) values were used for calculating biomass-specific consumption rates (r N as mgN/gX/h). The overall biomass yield for nitrogen (Y X/N) is the ratio between the mass of yeasts totally formed and the mass of N altogether consumed. The final N content of the biomass (x N) is the reciprocal of this overall Y X/N value.
Results and discussion
Nitrogen in non-supplemented DW medium
The composition of whey depends on many factors such as origin of milk (cow, goat, or sheep), technology of curd production, and whey processing (e.g., [51,52]). Casein protein is coagulated by acidification (mineral or organic acids directly added or lactic acid produced in the processed milk by bacteria) and/or by using chymosin. Whey processing modifies the composition of whey as well: whey protein is separated by ultrafiltration, solutes are concentrated by reverse osmosis, and/or minerals are partially removed by alkalinization. Preparation of whey-borne culture media also changes the composition by dilution, adding supplements, and heat sterilization. This explains why published compositions of whey-borne media highly fluctuate.
Several batches of non-supplemented DW medium were analyzed regarding potential sources of nitrogen and some other parameters (Table 1). The medium is rich in \sugar and thus exhibits a high potential for biomass growth. An amount of 78 g/L sugar allows formation of 28 g/L K. marxianus biomass assuming Y X/S = 0.36 g/g (as observed at aerobic cultivation in DW basic medium with trace elements [11]). These 28 g/L biomass can only develop when yeast growth is not limited by any other resource than sugar. K. marxianus DSM 5422 grown under such conditions exhibited the following content of minerals (in milligrams of the addressed element per gram dry biomass): x N = 78…79 mg/g [10,11], x P = 10 mg/g, x S = 4 mg/g, and x K = 2 mg/g (unpublished results). These data are similar to published compositions of K. marxianus [45,53-55] and Saccharomyces cerevisiae [40]. Multiplying these x values with the cell concentration of 28 g/L gives the required concentration of the respective element in the culture medium to allow non-limited growth: 2,200 mg/L N, 280 mg/L P, 112 mg/L S, and 56 mg/L K. Nitrogen and sulfur are lacking in non-supplemented DW medium while the phosphorous and potassium content covers the demand (Table 1).
DW medium owns 403 mg/L Kjeldahl-N (Table 1). Detailed analysis of five batches of non-supplemented DW medium gave 89 to 130 mg/L ammonium-N (106 mg/L on an average, σn−1 = 15 mg/L), 38.4 to 45.9 mg/L urea-N (43 mg/L on an average, σn−1 = 2.6 mg/L), 4 mg/L nitrate-N, and <1 mg/L nitrite-N. The Kjeldahl-N minus the urea-N and inorganic N gives a proteinogenic N of 250 mg/L which corresponds to ca. 1.5 g/L proteins (Table 1).
Utilization of whey proteins by K. marxianus is discussed controversially. Raw cheese whey owns 7 g/L protein comprising 50% β-lactoglobulin, 20% α-lactalbumin, 15% glycomacropeptide, and 15% minor protein/peptide components [51]. Their microbial hydrolysis requires excretion of proteases. Decomposition of whey proteins by K. marxianus has been repeatedly studied; the results varied from absent hydrolysis [26], over 20% to 33% [28,29,56], up to 80% hydrolysis [33]. Recent studies confirmed an extracellular serine protease for K. marxianus [57], and Yadav et al. [28] proved modification of whey proteins by K. marxianus via electrophoresis. Indigenous proteases in milk [58] and the proteolytic activity of lactobacilli [56] could also contribute some to protein modification during milk processing. Pre-treatment of whey protein with added proteases resulted in peptides <1 kDa which were efficiently assimilated by K. marxianus [59].
Some yeasts and fungi assimilate nitrate by intracellular reduction to ammonium [60]. K. marxianus seems to be unable for nitrate assimilation; at least several tested strains were negative [61]. This explains why nitrate added to whey did not improve growth of K. marxianus [32]. Some growth of K. marxianus in NaNO3-supplemented medium was possibly caused by the added yeast extract [62]. K. marxianus DSM 5422 proved to be unable to assimilate nitrate.
Due to the uncertainty of whey-protein assimilation by K. marxianus, it is assumed that whey proteins do not contribute to assimilable nitrogen. The utilizable nitrogen in non-supplemented DW medium of ca. 150 mg/L (ammonium-N plus urea-N) is thus much smaller than the calculated demand. Supplementation of DW medium with nitrogen is essentially required.
K. marxianus DSM 5422 grows well with ammonium [10,11], but its growth with urea has not yet been tested. Yeasts generally assimilate urea [44] which should also apply to the studied strain.
A low amount of iron, zinc, and copper in DW medium limits growth of K. marxianus DSM 5422 [11]. Here, the medium was ever supplemented with trace-element solution to avoid such limitation.
Test of several sources of nitrogen without previous pH adjustment
This preliminary test for assimilation of diverse sources of nitrogen by K. marxianus DSM 5422 was performed in shake flasks. Non-supplemented DW medium was spiked with several N sources (Table 2): no supplement as a reference, (NH4)2SO4 as the usual N supplement, (NH4)2CO3 as an alternative ammonium resource, and urea as another N compound. The DW medium contributed ca. 7 mg/L N (5 mg/L NH4-N and 2 mg/L urea-N) to the assimilable N in each culture. The initial pH value was influenced by the added N source: (NH4)2SO4 did not change the pH while urea and (NH4)2CO3 alkalinized the medium (Table 2). All shake flasks were inoculated and pre-cultivated for 14 h before the process was followed by repeated OD and pH measurements (Figure 1).

Aerobic batch cultivation of K. marxianus DSM 5422 without pH adjustment. Aerobic batch cultivation of K. marxianus DSM 5422 in media with various sources of nitrogen without previous pH adjustment. Concentrated and partially demineralized sweet whey was diluted with the same volume of water, supplemented with various sources of nitrogen, and autoclaved; the clear upper phase was supplemented with trace elements and diluted again with water resulting in media with 3.9 g/L sugar; the media were inoculated and cultivated in conical flasks at 250 rpm and 32°C.
The process without a supplement was at first similar to the process with (NH4)2SO4 (Figure 1), but later, nitrogen became a limiting factor so that growth and medium acidification slowed down and nearly stopped after 24 h. The N-limited growth resulted in some residual sugar (Table 2).
With a supplement of (NH4)2SO4, K. marxianus DSM 5422 grew at first with μ = 0.57 h−1 as usual in diluted DW medium [12,14], but then, the growth slowed down due to medium acidification (Figure 1). A pH <3.5 impairs growth of K. marxianus [28,42,43]. This acidification was caused by consumption of ammonium without an equivalent uptake of sulfate. The use of ammonium sulfate as an N source calls for buffered medium (in shake-flask experiments [12,14]) or for pH control (in bioreactor experiments [10,11,13,16,22]).
The supplement of (NH4)2CO3 at first alkalinized the medium. The initial pH of 7.5 impacted growth of K. marxianus DSM 5422 and caused a highly retarded process (Figure 1). Vivier et al. [42] and Antoce et al. [43] found that a pH >7 is adverse for growth of K. marxianus. Continued growth gradually reduced the pH and accelerated yeast growth so that all sugar had been consumed after 40 h. Carbonate as an exchangeable anion can leave the medium in the form of carbon dioxide which counteracted acidification (medium with (NH4)2CO3) while sulfate as a permanent ion remains and resulted in unfavorable acidification (medium with (NH4)2SO4).
The supplement of urea also alkalinized the medium to some degree (initial pH = 7.18) which is, at the first view, surprising since urea reacts neutral in aqueous solution. Autoclaving the medium together with urea maybe caused some hydrolysis of urea to form alkaline ammonium carbonate. This slight alkalinization slowed down yeast growth and retarded the process a little at the beginning, but the yeast metabolism lowered the pH so that the growth rate approached a normal value during pre-cultivation (Table 2). After 21 h, the medium acidification stopped and then the pH increased to a final pH of 7.1 (i.e., only temporary decrease in pH). Absence of enduring pH changes confirms the earlier postulated proton neutrality at growth with urea [38-40].
Test of several sources of nitrogen with previous pH adjustment
The just-described shake-flask experiments were repeated with two modifications: all sources of nitrogen were autoclaved separately to eliminate unwanted interaction between the N sources and medium constituents, and the initial pH value was adjusted to 6.5 for avoiding alkaline conditions.
The convenient initial pH of 6.5 produced a uniformly high specific growth rate of μ = 0.59 h−1 (Table 3) resulting in very similar cell densities at the beginning of the observation period (Figure 2A). This μ value was identical with the growth rate of K. marxianus DSM 5422 in highly diluted and phosphate-buffered DW basic medium [12,14].

Aerobic batch cultivation of K. marxianus DSM 5422 with initial pH adjustment. Aerobic batch cultivation of K. marxianus DSM 5422 in media with various sources of nitrogen with initial pH adjustment. Non-supplemented DW medium was supplemented with various sources of nitrogen and trace elements, the pH was adjusted to 6.5 by HCl or KOH, and these media were diluted with water resulting in 3.9 g/L sugar; the media were inoculated and cultivated in conical flasks at 250 rpm and 32°C.
The culture without a supplement of nitrogen exhibited N-limited growth (Figure 2A), only moderate acidification owing to restricted growth (Figure 2B), a low biomass formation, and some residual sugar after 40 h of cultivation (Table 3). Supplementation with (NH4)2SO4 or (NH4)2CO3 resulted in a nearly identical growth behavior and similar final pH values and biomass yields (Table 2); the pH was suboptimal in both cases which caused impaired growth compared to the process in phosphate-buffered medium [12,14]. A reduced biomass yield for K. marxianus in whey at a low pH has also been described by Yadav et al. [28] and interpreted as diversion of lactose from anabolism (growth) toward catabolism (maintenance). The use of (NH4)2CO3 instead of (NH4)2SO4 was without advantage since the initial pH adjustment of (NH4)2CO3-supplemented medium with hydrochloric acid caused substitution of exchangeable by permanent ions (carbonate replaced by chloride). (NH4)2CO3 is thus not a useful alternative since a high initial pH inhibits yeast growth while preceding pH adjustment with acids eliminates the buffering effect of carbonate.
Separate autoclaving of the N source and initial pH adjustment caused good growth in the urea-supplemented medium from the beginning; then, the growth became somewhat retarded (at 18 to 22 h; Figure 2A), but afterward, the growth accelerated again (at t >22 h) and even exceeded growth with (NH4)2SO4 (Table 3). Quick growth in the first period was possibly based on whey-borne ammonium while the temporal slowdown of growth in the second period perhaps came about through the adaptation of yeast metabolism to urea assimilation after NH4-N depletion. The quite high biomass yield with urea is certainly attributed to an appropriate pH value over the whole growth period (compare [39,62]). Urea is thus a promising source of nitrogen for K. marxianus. The acidification with urea was only of temporal nature; the pH increased again to give a final value being nearly identical with the initial pH (due to proton neutrality at assimilation of urea [38-40]).
Assimilation of urea by K. marxianus DSM 5422
Ammonium is utilized by all common yeasts directly while urea is either hydrolyzed by urease to form ammonium or it is assimilated via the urea amydolyase pathway [25,38]. Urease acts extracellularly while the amydolyase pathway works intracellularly. K. marxianus is regarded as a urease-negative yeast [40] and should metabolize urea only in the latter way being a two-step process [44,60,63]; urea reacts with hydrocarbonate in an energy-consuming process to form allophanate which, in turn, is hydrolyzed to release ammonium:
These two reactions are catalyzed by urea carboxylase and allophanate hydrolase [44,60,63]. The produced ammonium is then metabolized in the same manner as ammonium taken up directly.
An experiment was done for clarifying the way of urea assimilation in K. marxianus DSM 5422. The yeast was cultivated in diluted DW medium with urea, and the obtained culture was then used in an above-described test. K. marxianus DSM 5422 did not excrete urease into the medium since urea added to the cell-free aqueous fraction of this culture was not hydrolyzed at all. This is in accordance with Nahvi and Moeini [61] who found all tested K. marxianus and K. lactis strains being urease negative. These findings argue for assimilation of urea via the amydolyase pathway. The grown yeast biomass was incubated with urea in a phosphate buffer but only a bit urea was reacted to ammonium (average reaction rate 0.25 mg urea/gX/h). Absent sugar obviously suppressed transfer of urea into ammonium which refers to an amydolyase pathway being under transcriptive control.
Bioreactor cultivation with the addition of urea
The shake-flask experiments clearly demonstrated the capability of K. marxianus DSM 5422 for urea assimilation. Urea is a promising source of nitrogen due to the proton neutrality at its consumption during yeast growth [38-40], avoiding strong acidification as happening at growth with (NH4)2SO4. Urea has been repeatedly used at growth of K. marxianus, but the obtained results were inconsistent [28,31,41,47,62]. Hensing et al. [39] referred to the potential risk of an imbalance between release and assimilation of ammonium; the medium could alkalinize when ammonium is quicker released from urea than is incorporated into biomass. Such an alkalinization was observed during cultivation of K. marxianus in urea-supplemented whey [47]. In the above-described experiments, the pH temporally decreased rather than increased. This phenomenon has not yet been understood and requires clarification in a bioreactor experiment.
K. marxianus DSM 5422 was cultivated in a stirred reactor at well-defined conditions (32°C, pO2 ≥30% air saturation, pH ≥5) in DW medium which was supplemented with urea, Na2SO4, and trace-element solution to avoid limitation of growth by nitrogen, sulfur or microelements. Another bioreactor experiment performed with DW medium containing 10 g/L (NH4)2SO4 and trace elements was taken from [11] and used here as a reference. These two processes are depicted in Figure 3, and characteristic parameters are summarized in Table 4.

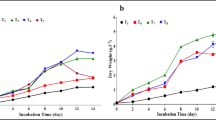
Aerobic batch cultivation of K. marxianus DSM 5422 in stirred bioreactors using ammonium or urea. Aerobic batch cultivation of K. marxianus DSM 5422 in a stirred bioreactor using ammonium (white symbols) or urea (grey symbols) as a source of nitrogen. DW medium with 2 mL/L trace-element solution was supplemented with (NH4)2SO4 or urea plus Na2SO4 (white symbols = 10 g/L (NH4)2SO4; grey symbols = 5 g/L urea and 0.4 g/L Na2SO4); the cultivation occurred in an 1-L stirred reactor at 1,200 rpm, 32°C, and aeration with 50 L/h; the pH was controlled to ≥5 with 2 M KOH; the given growth rate was derived from measured CO2 data.
The courses of yeast growth, sugar consumption, and marginal ethanol formation were very similar for (NH4)2SO4 or urea as the added N sources (Figure 3A). The only marked difference was the amount of formed biomass which was apparently higher with urea (Table 4). This observation is in accordance with Hensing et al. [39] and Rajoka et al. [62]. From the energetic point of view, growth with urea should be less effective compared to growth with ammonium since assimilation of urea via the amydolyase pathway requires ATP [44,60,63]. Cultivation with urea should therefore result in a lower rather than a higher biomass yield. In case of Hensing et al. [39] and Rajoka et al. [62], the observed low growth with ammonium was possibly caused by inhibitory acidification due to absent pH control at shake-flask cultivation. Such an inhibitory acidification was prevented by controlling the pH during bioreactor cultivation (Figure 3B); here, the diverging biomass yields were presumably caused by slightly different amounts of bioavailable nitrogen (1.49 g N in medium with 5 g/L urea, and 1.30 g N in medium with 10 g/L (NH4)2SO4). Nearly all bioavailable nitrogen was assimilated in both processes (only 20 or 30 mg/L residual NH4-N in the culture broth; Table 4) which refers to a slight deficit of nitrogen, and this deficit was more striking with (NH4)2SO4. This argumentation is supported by a higher N content in the biomass grown with urea (Table 4).
In urea-supplemented medium, K. marxianus DSM 5422 grew at first quickly; but later, the growth slowed down more and more (Figure 3D), although sufficient O, N, P, S, K, and trace elements should allow non-limited growth over an extended period. This behavior could be explained with a lack of vitamins in whey [27,29] or with inhibition by whey-borne minerals [31]. Supplementing whey with yeast extract [27,29] or vitamins [29,32,47] stimulated the growth of K. marxianus.
The source of nitrogen distinctly influenced the pH(t) course (Figure 3B). With (NH4)2SO4, the pH decreased due to ammonium consumption until the pH was controlled to pH 5; 82 mL/L 2 M KOH were supplied which corresponds to a specific dosage of 0.34 g KOH per g of produced biomass. The strong acidification with (NH4)2SO4 is explained by a proton imbalance [38-40]: ammonium was consumed while most of the sulfate remained in the medium. With urea, the pH at first a little decreased, then sharply rose to pH 6.8 and reduced again to pH 5 where the pH controller avoided further acidification; the KOH dosage was in fact much smaller (only 10 mL/L 2 M KOH or 0.04 g KOH per g of produced biomass; Table 4). Hensing et al. [39] cultivated K. lactis in galactose medium with (NH4)2SO4 or urea; the acidification was strong and permanent with (NH4)2SO4, while the acidification was moderate and only temporary with urea.
During cultivation of K. marxianus DSM 5422 in urea-supplemented DW medium, the dissolved ammonium-N and urea-N were repeatedly measured and used for calculating the N consumption (Figure 3C). Intracellular conversion of urea to ammonium and usage of this ammonium for biomass growth occurred with nearly the same rate since ammonium excretion was only marginal (some NH4-N originated from the used whey). The N consumption correlated well with the yeast growth (compare Figure 3A and C) and, thus, the courses of the N consumption rate and the specific growth rate were similar (Figure 3D). The quotient of these rates represents a momentary Y X/N value.
The small transient NH4-N accumulation (Figure 3C) partially correlated with the temporary increase in pH. Hensing et al. [39] already referred to the danger of alkalinization when ammonium release exceeds ammonium assimilation. Such an alkalization to pH 8.5 was observed by Rech et al. [47] at cultivation of K. marxianus in urea-supplemented whey causing severe growth inhibition. Here, such an inhibition did not occur (only moderate rise of pH to 6.8).
Bioreactor cultivation at a pH-controlled feed of ammonia
Feeding the required nitrogen in form of ammonia could be a cost-saving alternative. Ammonia was repeatedly used as an N source at cultivation of K. marxianus in whey or other media [34,64-67] but dissolved ammonium or N consumption has not been paid much attention, with exception of Hack and Marchant [65] who depicted the time-dependent supply of ammonia.
In another series of bioreactor experiments, K. marxianus DSM 5422 was cultivated in DW medium as before but 2 M NH4OH was used as the predominating N source which was supplied by the pH controller at pH <5. Three experiments were performed with a varied mass of (NH4)2SO4 which was added as a pure substance to the autoclaved DW medium (0, 0.6, or 1.2 g). These processes were limited neither by oxygen (pO2 always >10%) nor by sulfur (proven by residual sulfate).
In DW medium without an (NH4)2SO4 supplement (Figure 4; white symbols), K. marxianus DSM 5422 grew on whey-borne nitrogen (0.13 g/L NH4-N and 0.04 g/L urea-N), but this nitrogen was quickly depleted (Figure 4C) and the growth became N limited (Figure 4A). The pH temporally rose (Figure 4B) which seemingly correlated with urea consumption (Figure 4C). After depletion of all bioavailable N, the pH decreased only slowly due to a low metabolic activity (look at the sugar concentration in Figure 4A). Later, the pH stagnated above pH 5 and ammonia was thus not dosed (Figure 4B). The low availability of nitrogen (Figure 4D) caused restricted yeast growth (Figure 4A). The total N consumption and the N consumption rate were accordingly low (Figure 4E,F).

Aerobic batch cultivation of K. marxianus DSM 5422 in stirred bioreactors using (NH 4 ) 2 SO 4 and NH 4 OH. Aerobic batch cultivation of K. marxianus DSM 5422 in a stirred bioreactor using various amounts of (NH4)2SO4 and NH4OH as sources of nitrogen. DW medium with 2 mL/L trace-element solution was supplemented with (NH4)2SO4 or Na2SO4 (white symbols = 0.4 g/L Na2SO4; grey symbols = 1 g/L (NH4)2SO4; black symbols = 2 g/L (NH4)2SO4); the cultivation occurred in an 1-L stirred reactor at 1,200 rpm, 32°C, and aeration with 50 L/h; the pH was controlled to ≥5 with 2 M NH4OH; the given growth rates were derived from measured CO2 data.
A supplement of 1 g/L (NH4)2SO4 in the second experiment (Figure 4; grey symbols) increased the bioavailable N (Figure 4D) and allowed better yeast growth due to the higher initial NH4-N which let the pH quickly decrease (Figure 4B). After depletion of this nitrogen, the pH stagnated at 5.05 for a while and then dropped below 5 where the pH controller started dosage of ammonia (Figure 4B). The added ammonia was assimilated immediately, and no ammonium accumulated in the medium (Figure 4C). That is, the yeast growth continued but at N-limited conditions as becoming visible from the low rates of ammonia dosage and N consumption (Figure 4D,F). This deficit of nitrogen slowed down growth and diminished the formed biomass (Figure 4A, Table 4). In the stationary period, some ammonium was released into the medium (Figure 4C) which was also observed by Ghaly and Kamal [26] at the cultivation of K. marxianus in whey and interpreted as decomposition of yeast biomass with release of NH4-N into the medium.
A supplement of 2 g/L (NH4)2SO4 in the third experiment (Figure 4; black symbols) increased the bioavailable N most (Figure 4D) and resulted in fast yeast growth and quick acidification. The feed of ammonia at pH <5 started before the initially added NH4-N had been depleted (Figure 4C). The early start of ammonia dosage prevented limitation of yeast growth by nitrogen (NH4-N always >100 mg/L), and the supply and uptake of nitrogen were well balanced (Figure 4C). The high rate of ammonia dosage corresponded with an accordingly fast growth and intensive N consumption (Figure 4D,E). The rate of N consumption became gradually smaller which is explained by the gently declining growth rate (possible reasons for this fading growth were discussed above). Urea was assimilated co-metabolically with the ammonium (Figure 4C).
The process with a supplement of 2 g/L (NH4)2SO4 and ammonia dosage ran very similar to the process with 10 g/L (NH4)2SO4; the final cell concentrations, formed biomasses, and the overall Y X/S values were nearly identical in both processes (Table 4) which demonstrates effective cultivation of K. marxianus with a pH-controlled feed of ammonia. Supplementing the medium with some (NH4)2SO4 was however required for a quick acidification and for initiation of ammonia dosage. The amount of added (NH4)2SO4 could be reduced by changing the setpoint of the pH controller (e.g., to pH 5.5) so that dosage of ammonia starts earlier and avoids N-limited conditions even at a reduced (NH4)2SO4 supplement, but some (NH4)2SO4 is needed to cover the requirement for sulfur.
Nitrogen in biomass
The content of nitrogen in biomass grown at cultivation in the stirred bioreactor (Figures 3 and 4) is the inverse of the overall biomass yield for nitrogen: x N = 1/Y X/N. The overall Y X/N values were calculated from the produced biomass and the consumed nitrogen, assuming that only ammonium and urea were assimilated (Table 4). This calculation ignores that K. marxianus possibly hydrolyzes some whey-borne proteins and assimilates thus-formed peptides and amino acids. The N content of biomass depended on the extent of N limitation (Table 4): cultivation with enough nitrogen (process with 5 g/L urea and process with 2 g/L (NH4)2SO4 plus dosed ammonia) gave x N values of 81 and 83 mg/g, a slight deficit of nitrogen during the late growth stage (process with 10 g/L (NH4)2SO4) resulted in x N = 76 mg/g, distinct N limitation (process with 1 g/L (NH4)2SO4 plus ammonia) yielded x N = 67 mg/g, while severe N limitation (process without any N supplement) produced an x N value of only 24 mg/g. A diminished N content of K. marxianus was also observed at limitation of growth by trace elements [10,11]. A decreased N content can be explained by a lowered portion of active biomass owing to intracellular storage of polysaccharides (details in [10,11]).
The N content can also be derived from the elemental composition of biomass. Several authors measured the cell composition for K. marxianus by elemental analyzers and transformed these data into biomass formulae: CH1.78O0.75 N0.16 [45], CH1.776O0.575 N0.159 [53], CH1.63O0.54 N0.16 [54], CH1.94O0.76 N0.17 [55]. These formulae represent an N content of 88, 80, 91, or 83 mg/g. The fluctuations originate from measuring errors and from a variable cell composition depending on growth conditions [68].
Stoichiometry of yeast growth
Stoichiometric equations for describing the growth of K. marxianus has been derived here by using the method of Hensing et al. [39] and Mazutti et al. [69]. Such balancing requires a sum formula for biomass. The above-given formulae for K. marxianus biomass are restricted to C, H, O, and N as the predominating elements (derived from elemental analyses [45,53-55]). Here, the elements P, S, and K are included for more precision. The C, H, and O content was taken from the above-given biomass formulae (as averages), the N content of 82 mg/g was taken from own measurements at non-limited yeast growth, and the P, S, and K content was assumed with 10, 4, and 2 mg/g (own assimilation measurements). Combination of these data gives CH1.78O0.66 N0.158P0.009S0.0035K0.0015 (yielding a molar mass of 27.027 g/mol).
Individual stoichiometric balance equations were derived for ammonium, urea, or ammonia as an N source, assuming respiratory growth (without formation of ethanol or ethyl acetate) of K. marxianus DSM 5422 with lactose as a substrate. The included stoichiometry coefficients were determined by balancing each element: seven balance equations were obtained containing nine unknown stoichiometric coefficients. This uncertain algebraic system was dissolved following Hensing et al. [39] by adding a proton balance and introducing the yield coefficient (Y X/S informs about the relation of assimilatory to dissimilatory substrate utilization and allows to establish the mass ratio between formed biomass and consumed lactose). Y X/S = 0.36 g/g was used here uniformly for all balances as found at non-limited growth with ammonium or ammonia (Table 4). Phosphate and sulfate were consumed in form of HPO4 2− and SO4 2− at the prevailing pH. Three equations were obtained for ammonium, ammonium hydroxide, or urea as an N source:
Protons are only formed during growth with ammonium which explains the observed substantial consumption of KOH by the pH controller with ammonium sulfate; the proton release equates to 0.28 g consumed KOH per g grown biomass and hence somewhat deviates from the measured KOH consumption (0.34 gKOH/gX). With NH4OH or urea (Equations 4 and 5), the balances predict a slight alkalinization since OH− ions are formed. The consumption of some KOH with urea as an N source is contradictory to this finding, but it should be kept in mind that the final pH was higher than the initial pH (Figure 3B); synthesis of organic acids (acetate, pyruvate, 2-oxoglutarate, and succinate were by-products of aerobic sugar metabolism of K. marxianus [16,68,70,71]) presumably caused KOH consumption, and consumption of these acidic metabolites after depletion of sugar alkalinized the medium. But such temporary metabolite accumulation was not considered at balancing. Another interfering effect originates from whey-borne lactate (ca. 4 g/L in DW medium [10]) whose microbial utilization also causes some alkalinization.
The balance equations allow to compare calculated with measured masses of consumed oxygen and formed carbon dioxide. The expected masses were calculated from the masses of utilized sugar. The measured masses (Table 4) were 1% to 20% smaller than predicted for unknown reason, but the ratio between formed CO2 and consumed oxygen (the average RQ values) was ca. 1.02 mol/mol (Table 4) and agreed well with the predicted values (1.01 to 1.03 mol/mol).
Conclusions
Whey is poor in nitrogen and requires supplementation with an N source for effective production of yeast biomass. Ammonium sulfate, as usually applied for this reason, causes medium acidification by residual sulfate which requires pH control by alkaline substances to avoid growth inhibition. Application of ammonium carbonate instead of ammonium sulfate is not helpful since added (NH4)2CO3 elevates the pH to inhibitory levels. K. marxianus DSM 5422 assimilates urea as an alternative N source. Consumption of urea means proton neutrality, medium acidification is minor, and only a little pH corrective is required. Moreover, the use of urea reduces the salt load (less inhibition, diminished environmental impact). Dosage of ammonia by the pH controller is a cost-saving alternative, but a suitable supplement of (NH4)2SO4 is needed as a source of sulfur and for initiating dosage of ammonia.
References
Posada JA, Patel AD, Roes A, Blok K, Faaij APC, Patel MK (2013) Potential of bioethanol as a chemical building block for biorefineries: preliminary sustainability assessment of 12 bioethanol-based products. Bioresour Technol 135:490–499
Kim S-J, Jung S-M, Park Y-C, Park K (2007) Lipase catalyzed transesterification of soybean oil using ethyl acetate, an alternative acyl acceptor. Biotechnol Bioprocess Eng 12:441–445
Modi MK, Reddy JRC, Rao BVSK, Prasad RBN (2007) Lipase-mediated conversion of vegetable oils into biodiesel using ethyl acetate as acyl acceptor. Bioresour Technol 98:1260–1264
Uthoff S, Bröker D, Steinbüchel A (2009) Current state and perspectives of producing biodiesel-like compounds by biotechnology. Microb Biotechnol 2:551–565
Röttig A, Wenning L, Bröker D, Steinbüchel A (2010) Fatty acid alkyl esters: perspectives for production of alternative biofuels. Appl Microbiol Biotechnol 85:1713–1733
Hwang S-CJ, Lee C-M, Lee H-C, Pua HF (2003) Biofiltration of waste gases containing both ethyl acetate and toluene using different combinations of bacterial cultures. J Biotechnol 105:83–94
Kam S-K, Kang K-H, Lee M-G (2005) Removal characteristics of ethyl acetate and 2-butanol by a biofilter packed with jeju scoria. J Microbiol Biotechnol 15:977–983
Chan W-C, Su M-Q (2008) Biofiltration of ethyl acetate and amyl acetate using a composite bead biofilter. Bioresour Technol 99:8016–8021
Löser C, Urit T, Bley T (2014) Perspectives for the biotechnological production of ethyl acetate by yeasts. Appl Microbiol Biotechnol 98:5397–5415
Urit T, Löser C, Wunderlich M, Bley T (2011) Formation of ethyl acetate by Kluyveromyces marxianus on whey: studies of the ester stripping. Bioprocess Biosyst Eng 34:547–559
Urit T, Löser C, Stukert A, Bley T (2012) Formation of ethyl acetate by Kluyveromyces marxianus on whey during aerobic batch cultivation at specific trace-element limitation. Appl Microbiol Biotechnol 96:1313–1323
Urit T, Manthey R, Bley T, Löser C (2013) Formation of ethyl acetate by Kluyveromyces marxianus on whey: influence of aeration and inhibition of yeast growth by ethyl acetate. Eng Life Sci 13:247–260
Löser C, Urit T, Stukert A, Bley T (2013) Formation of ethyl acetate from whey by Kluyveromyces marxianus on a pilot scale. J Biotechnol 163:17–23
Urit T, Li M, Bley T, Löser C (2013) Growth of Kluyveromyces marxianus and formation of ethyl acetate depending on temperature. Appl Microbiol Biotechnol 97:10359–10371
Löser C, Urit T, Nehl F, Bley T (2011) Screening of Kluyveromyces strains for the production of ethyl acetate: design and evaluation of a cultivation system. Eng Life Sci 11:369–381
Löser C, Urit T, Förster S, Stukert A, Bley T (2012) Formation of ethyl acetate by Kluyveromyces marxianus on whey during aerobic batch and chemostat cultivation at iron limitation. Appl Microbiol Biotechnol 96:685–696
Kallel-Mhiri H, Engasser J-M, Miclo A (1993) Continuous ethyl acetate production by Kluyveromyces fragilis on whey permeate. Appl Microbiol Biotechnol 40:201–205
Willetts A (1989) Ester formation from ethanol by Candida pseudotropicalis. Antonie Van Leeuwenhoek 56:175–180
Aziz S, Memon HUR, Shah FA, Rajoka MI, Soomro SA (2009) Production of ethanol by indigenous wild and mutant strain of thermotolerant Kluyveromyces marxianus under optimized fermentation conditions. Pak J Anal Environ Chem 10(1+2):25–33
Lertwattanasakul N, Rodrussamee N, Suprayogi LS, Thanonkeo P, Kosaka T, Yamada M (2011) Utilization capability of sucrose, raffinose and inulin and its less-sensitiveness to glucose repression in thermotolerant yeast Kluyveromyces marxianus DMKU 3–1042. AMB Express 1:20
Fonseca GG, de Carvalho NMB, Gompert AK (2013) Growth of the yeast Kluyveromyces marxianus CBS 6556 on different sugar combinations as sole carbon and energy source. Appl Microbiol Biotechnol 97:5055–5067
Löser C, Urit T, Keil P, Bley T (2014) Studies on the mechanism of synthesis of ethyl acetate in Kluyveromyces marxianus DSM 5422. Appl Microbiol Biotechnol. Accepted, doi:10.1007/s00253-014-6098-4
Armstrong DW, Yamazaki H (1984) Effect of iron and EDTA on ethyl acetate accumulation in Candida utilis. Biotechnol Lett 6:819–824
Fonseca GG, Heinzle E, Wittmann C, Gombert AK (2008) The yeast Kluyveromyces marxianus and its biotechnological potential. Appl Microbiol Biotechnol 79:339–354
Ugalde OU, Castrillo IJ (2002) Single cell proteins from fungi and yeasts. In: Dilip KA, George GK (eds) Applied mycology and biotechnology, vol 2, Agriculture and food production. Elsevier, Amsterdam, pp 123–149
Ghaly AE, Kamal MA (2004) Submerged yeast fermentation of acid cheese whey for protein production and pollution potential reduction. Water Res 38:631–644
Schultz N, Chang L, Hauck A, Reuss M, Syldatk C (2006) Microbial production of single-cell protein from deproteinized whey concentrates. Appl Microbiol Biotechnol 69:515–520
Yadav JSS, Bezawada J, Elharche S, Yan S, Tyagi RD, Surampalli RY (2014) Simultaneous single-cell protein production and COD removal with characterization of residual protein and intermediate metabolites during whey fermentation by K. marxianus. Bioprocess Biosyst Eng 37:1017–1029
Parrondo J, García LA, Díaz M (2009) Nutrient balance and metabolic analysis in a Kluyveromyces marxianus fermentation with lactose-added whey. Brazil J Chem Eng 26:445–456
De Nicola R, Hazelwood LA, De Hulster EAF, Walsh MC, Knijnenburg TA, Reinders MJT, Walker GM, Pronk JT, Daran J-M, Daran-Lapujade P (2007) Physiological and transcriptional responses of Saccharomyces cerevisiae to zinc limitation in chemostat cultures. Appl Environ Microbiol 73:7680–7692
Mahmoud MM, Kosikowski FV (1982) Alcohol and single cell protein production by Kluyveromyces in concentrated whey permeates with reduced ash. J Dairy Sci 65:2082–2087
Kar T, Misra AK (1998) Effect of fortification of concentrated whey on growth of Kluyveromyces sp. Rev Argent Microbiol 30(4):163–169
Belem MAF, Lee BH (1999) Fed-batch fermentation to produce oligonucleotides from Kluyveromyces marxianus grown on whey. Process Biochem 34:501–509
Domingues L, Lima N, Teixeira JA (2001) Alcohol production from cheese whey permeate using genetically modified flocculent yeast cells. Biotechnol Bioeng 72:507–514
Moeini H, Nahvi I, Tavassoli M (2004) Improvement of SCP production and BOD removal of whey with mixed yeast culture. Electronic J Biotechnol 7:249–255
Aktaş N, Boyacı İH, Mutlu M, Tanyolaç A (2006) Optimization of lactose utilization in deproteinated whey by Kluyveromyces marxianus using response surface methodology (RSM). Bioresour Technol 97:2252–2259
Gupte AM, Nair AS (2010) β-galactosidase production and ethanol fermentation from whey using Kluyveromyces marxianus NCIM 3551. J Sci Ind Res 69:855–859
Castrillo JI, de Miguel I, Ugalde UO (1995) Proton production and consumption pathways in yeast metabolism. A chemostat culture analysis. Yeast 11:1353–1365
Hensing MCM, Bangma KA, Raamsdonk LM, de Hulster E, van Dijken JP, Pronk JT (1995) Effects of cultivation conditions on the production of heterologous β-galactosidase by Kluyveromyces lactis. Appl Microbiol Biotechnol 43:58–64
Vicente A, Castrillo JI, Teixeira JA, Ugalde U (1998) On-line estimation of biomass through pH control analysis in aerobic yeast fermentation systems. Biotechnol Bioeng 58:445–450
Tovar-Castro L, García-Garibay M, Saucedo-Castañeda G (2008) Lactase production by solid-state cultivation of Kluyveromyces marxianus CDBBL 278 on an inert support: effect of inoculum, buffer, and nitrogen source. Appl Biochem Biotechnol 151:610–617
Vivier D, Ratomahenina R, Moulin G, Galzy P (1993) Study of physicochemical factors limiting the growth of Kluyveromyces marxianus. J Ind Microbiol 11:157–161
Antoce O-A, Antoce V, Takahashi K (1997) Calorimetric study of yeast growth and its inhibition by added ethanol at various pHs and temperatures. Netsu Sokutei 24(4):206–213
Large PJ (1986) Degradation of organic nitrogen compounds by yeasts. Yeast 2:1–34
Castrillo JI, Ugalde UO (1992) Energy metabolism of Kluyveromyces marxianus in deproteinated whey. Chemostat studies. Modelling. J Biotechnol 22:145–152
Castrillo JI, Ugalde UO (1993) Patterns of energy metabolism and growth kinetics of Kluyveromyces marxianus in whey chemostat culture. Appl Microbiol Biotechnol 40:386–393
Rech R, Cassini CF, Secchi A, Ayub MAZ (1999) Utilization of protein-hydrolyzed cheese whey for production of β-galactosidase by Kluyveromyces marxianus. J Ind Microbiol Biotechnol 23:91–96
Hortsch R, Löser C, Bley T (2008) A two-stage CSTR cascade for studying the effect of inhibitory and toxic substances in bioprocesses. Eng Life Sci 8:650–657
Rahmatullah M, Boyde TRC (1980) Improvements in the determination of urea using diacetyl monoxime; methods with and without deproteinisation. Clin Chim Acta 107:3–9
Francis PS (2006) The determination of urea in wine - a review. Aust J Grape Wine Res 12:97–106
Smithers GW (2008) Whey and whey proteins - from ‘gutter-to-gold’. Int Dairy J 18:695–704
Prazeres AR, Carvalho F, Rivas J (2012) Cheese whey management: a review. J Environ Manage 110:48–68
Cordier J-L, Butsch BM, Birou B, von Stockar U (1987) The relationship between elemental composition and heat of combustion of microbial biomass. Appl Microbiol Biotechnol 25:305–312
Krzystek L, Ledakowicz S (2000) Stoichiometric analysis of Kluyveromyces fragilis growth on lactose. J Chem Technol Biotechnol 75:1110–1118
Silva-Santisteban BOY, Converti A, Filho FM (2006) Intrinsic activity of inulinase from Kluyveromyces marxianus ATCC 16045 and carbon and nitrogen balances. Food Technol Biotechnol 44:479–483
Hamme V, Sannier F, Piot J-M, Didelot S, Bordenave-Juchereau S (2009) Crude goat whey fermentation by Kluyveromyces marxianus and Lactobacillus rhamnosus: contribution to proteolysis and ACE inhibitory activity. J Dairy Res 76:152–157
Foukis A, Stergiou P-Y, Theodorou LG, Papagianni M, Papamichael EM (2012) Purification, kinetic characterization and properties of a novel thermo-tolerant extracellular protease from Kluyveromyces marxianus IFO 0288 with potential biotechnological interest. Bioresour Technol 123:214–220
Fox PF (2003) Significance of indigenous enzymes in milk and dairy products. In: Whitaker JR, Voragen AGJ, Wong DWS (eds) Handbook of food enzymology. Marcel Dekker Inc, New York, Basel, pp 255–277
Perea A, Ugalde U, Rodriguez I, Serra JL (1993) Preparation and characterization of whey protein hydrolysates: applications in industrial whey bioconversion processes. Enzyme Microb Technol 15:418–423
Walker GM, White NA (2005) Introduction to fungal physiology. In: Kavanagh K (ed) Fungi: biology and applications. John Wiley & Sons Ltd, Chichester, pp 1–34
Nahvi I, Moeini H (2004) Isolation and identification of yeast strains with high beta-galactosidase activity from dairy products. Biotechnol 3(1):35–40
Rajoka MI, Khan S, Latif F, Shahid R (2004) Influence of carbon and nitrogen sources and temperature on hyperproduction of a thermotolerant β-glucosidase from synthetic medium by Kluyveromyces marxianus. Appl Biochem Biotechnol 117(2):75–92
Whitney PA, Cooper TG (1972) Urea carboxylase and allophanate hydrolase. Two components of adenosine triphosphate: urea amido-lyase in Saccharomyces cerevisiae. J Biol Chem 247:1349–1353
Shay LK, Hunt HR, Wegner GH (1987) High-productivity fermentation process for cultivating industrial microorganisms. J Ind Microbiol 2:79–85
Hack CJ, Marchant R (1998) Characterisation of a novel thermotolerant yeast, Kluyveromyces marxianus var marxianus: development of an ethanol fermentation process. J Ind Microbiol Biotechnol 20:323–327
Nor ZM, Tamer MI, Scharer JM, Moo-Young M, Jervis EJ (2001) Automated fed-batch culture of Kluyveromyces fragilis based on a novel method for on-line estimation of cell specific growth rate. Biochem Eng J 9:221–231
Gélinas P, Barrette J (2007) Protein enrichment of potato processing waste through yeast fermentation. Bioresour Technol 98:1138–1143
Fonseca GG, Gombert AK, Heinzle E, Wittmann C (2007) Physiology of the yeast Kluyveromyces marxianus during batch and chemostat cultures with glucose as the sole carbon source. FEMS Yeast Res 7:422–435
Mazutti MA, Zabot G, Boni G, Skovronski A, de Oliveira D, Di Luccio M, Rodrigues MI, Maugeri F, Treichel H (2010) Mathematical modeling of Kluyveromyces marxianus growth in solid-state fermentation using a packed-bed bioreactor. J Ind Microbiol Biotechnol 37:391–400
Silva-Santisteban BOY, Converti A, Filho FM (2009) Effects of carbon and nitrogen sources and oxygenation on the production of inulinase by Kluyveromyces marxianus. Appl Biochem Biotechnol 152:249–261
Rocha SN, Abrahão-Neto J, Gombert AK (2011) Physiological diversity within the Kluyveromyces marxianus species. Antonie Van Leeuwenhoek 100:619–630
Acknowledgements
Thanet Urit would like to express his thanks to the Nakhon Sawan Rajabhat University (Muang Nakhon Sawan, Thailand) for financial support. We are grateful to Mrs. E. Kneschke for technical assistance, to M. Heller from the Sachsenmilch Leppersdorf GmbH (Germany) for providing whey permeate, and to A. Stukert for performing two bioreactor experiments.
Dedication
This publication is dedicated to Prof. Dr. Andreas Zehnsdorf on the occasion of his 50th birthday.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
CL and TU conceived of the study. EG, CL, and TU explored relevant literature. CL and TU designed the experiments. EG and TU conducted the experiments. CL performed data analysis. CL and TB drafted the manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0), which permits use, duplication, adaptation, distribution, and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Löser, C., Urit, T., Gruner, E. et al. Efficient growth of Kluyveromyces marxianus biomass used as a biocatalyst in the sustainable production of ethyl acetate. Energ Sustain Soc 5, 2 (2015). https://doi.org/10.1186/s13705-014-0028-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13705-014-0028-2