
Abstract
Background
Tsetse flies are vectors of human and animal African trypanosomiasis. In spite of many decades of chemotherapy and vector control, the disease has not been eradicated. Other methods like the transformation of tsetse fly symbionts to render the fly refractory to trypanosome infection are being evaluated. The aim of the present study was to evaluate the association between trypanosome infections and the presence of symbionts in these tsetse species. Tsetse flies were trapped in two villages of the “Faro and Déo” Division of the Adamawa region of Cameroon. In the field, tsetse fly species were identified and their infection by trypanosomes was checked by microscopy. In the laboratory, DNA was extracted from their midguts and the presence of symbionts (Sodalis glossinidius and Wolbachia sp.) and trypanosomes was checked by PCR. Symbionts/trypanosomes association tests were performed.
Results
Three tsetse fly species including Glossina tachinoides (90.1%), Glossina morsitans submorsitans (9.4%) and Glossina fuscipes fuscipes (0.5%) were caught. In all the population we obtained an occurrence rate of 37.2% for Sodalis glossinidius and 67.6% for Wolbachia irrespective to tsetse flies species. S. glossinidius and Wolbachia sp. occurrence rates were respectively 37 and 68% for G. tachinoides and 28.6 and 59.5% for G. m. submorsitans. Between Golde Bourle and Mayo Dagoum significant differences were observed in the prevalence of symbionts. Prevalence of trypanosomes were 34.8% for Glossina tachinoides and 40.5% for Glossina morsitans submorsitans. In G. tachinoides, the trypanosome infection rates were 11, 2.6 and 13.7%, respectively, for T. brucei s.l., T. congolense forest type and T. congolense savannah type. In G. m. submorsitans, these infection rates were 16.7, 9.5 and, 2.4% respectively, for T. brucei s.l., T. congolense forest type and T. congolense savannah type.
Conclusions
The rate of tsetse fly infection by trypanosomes was low compared to those obtained in HAT foci of south Cameroon, and this rate was not statistically linked to the rate of symbiont occurrence. This study allowed to show for the first time the presence of Wolbachia sp. in the tsetse fly sub-species Glossina morsitans submorsitans and Glossina tachinoides.
Similar content being viewed by others

Background
Tsetse flies are vectors of parasitic diseases, namely, sleeping sickness or Human African Trypanosomiasis (HAT) in humans and nagana or Animal African Trypanosomiasis (AAT) in cattle. Nagana remains one of major obstacles constraining livestock development in several sub-Saharan African regions. It is responsible for the death of about 3 million cattle every year, the loss of 26% of dairy yield and the reduction of 50% of the number of cattle herds used in high potential agricultural areas [1]. In addition, the shortfall in agriculture due to nagana is estimated at 4.5 billion US$ per year [2]. AAT constitutes therefore one of the major causes of famine and poverty in several sub-Saharan countries [3, 4]. Both vector control and the drug administration for preventive or curative therapy are currently used for the control of AAT. The drugs currently used for the treatment of trypanosome infections in animals are not very efficient due to the increasing number of drug resistant strains in several African countries [5] including Cameroon [6]. For vector control, several tools have been developed (the trapping, the use of insecticides and the sterile insect technique) (see [7] for a review). However, the implementation and the field sustainability of these tools remain a challenge. To improve vector control, investigations on endosymbionts which could be implicated in the vector competence of tsetse flies have been undertaken in different tsetse species.
Indeed, besides the parasite, tsetse flies harbor three species of symbiotic bacteria which show different levels of relation with their host [8]. Wiggleworthia glossinidia, which is the obligatory symbiont, provides food supplements to maintain the fecundity of the tsetse fly and is therefore important for their larval development and contributes later to the maturation of the immune system [9]. Sodalis glossinidius, a maternally transmitted endosymbiont was suspected to be involved in tsetse fly vector competence by favoring parasite installation in the insect midgut [10] through a complex biochemical mechanism involving the production of N-acetyl glucosamine [11,12,13].
Bacteria belonging to the genus Wolbachia are a group of intracellular alpha proteobacteria (order Rickettsiales) that are maternally inherited and occur in numerous arthropod (65% of insects) and filarial nematode species [14]. This bacteria is associated with the reproductive tissues and causes reproductive abnormalities such as cytoplasmic incompatibility (CI), parthenogenesis, male death and feminization [15]. The ability to induce reproductive phenotypes allows them to spread effectively and quickly in host populations [16]. Experimental studies conducted by Geiger et al. [17, 18] showed an association between the presence of symbionts and the ability of tsetse flies to harbor trypanosomes. Moreover, a positive association between the presence of S. glossinidius in G. p. palpalis and trypanosome infections has been observed in the sleeping sickness foci of Campo and Bipindi in Cameroon [10, 19]. Despite these interesting data, there is little information in the relationship between the presence of endosymbionts and trypanosome infections in other tsetse fly species.
The aim of this study was to determine the prevalence of different trypanosome species and two endosymbionts (S. glossinidius and Wolbachia sp.) in two tsetse species of two AAT endemic villages of the Adamawa region of Cameroon, in order to evaluate the association between the trypanosome infections and the presence of symbionts in these tsetse species. The study is a preliminary stage of the IAEA-CRP D42015 program that aims to enhance tsetse fly refractoriness to trypanosome infection.
Methods
Study site
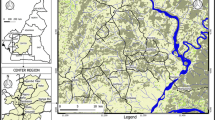
This study was carried out in the villages of Mayo Dagoum (longitude 07°33.06′ and latitude 12°07.11′) and Golde Bourle (longitude 07°33.16′ and latitude 12°07.25′) of the “Faro and Déo” Division in Adamawa region (Fig. 1). The Adamawa plateau covers more than 72, 000 km2 with very suitable environments for intensive cattle rearing. Trypanosomiasis is the principal reason of bovine veterinary consultation [20]. The “Faro and Déo” Division is located in northern part of the Adamawa plateau and covers about 11, 000 km2 with an altitude of 1000 to 1100 m. The climate is of Sudano-Sahelian type with an average rainfall of 1800 mm [21]. This area has a dense hydrographic network and during the year offers pastures for livestock. However, the pastoral resources are not well exploited because the presence of tsetse flies and the high rate (61.1%) of trypanosome infections in cattle [22] are major constraints to agro pastoral activities.

Sampling sites in the Animal African Trypanosomiasis focus of Dodeo
Sampling and dissection of tsetse flies
For this study, entomological surveys were conducted in January 2013 at Mayo Dagoum and Golde Bourle villages. During these surveys, 86 traps were put in suitable tsetse fly biotopes for five consecutive days. The geographical coordinates of each trap were recorded using a global positioning system (GPS) device (Garmin Etrex). Tsetse flies were collected twice a day at 11 am and 4 pm to obtain fresh samples for dissection. Species and sex were identified according to morphological criteria [23]. Young flies that had never taken a blood meal, called teneral flies, characterized by the slack aspect of their thorax were excluded from the sample [24]. The non-teneral flies were dissected under a binocular magnifying glass in a drop of sterile 0.9% saline solution. Dissection tools were carefully cleaned successively in 0.5 N NaOH and distilled water after the dissection of each fly to prevent contamination. Tsetse fly midguts were examined under the light microscope at magnification 10X for trypanosome detection and 40X for the confirmation. Midguts and other body parts were separately transferred into Eppendorf tubes containing ethanol (95°). During field manipulations, the tubes were maintained at room temperature; and in the laboratory, they were stored at − 20 °C until use.
DNA extraction
Frozen samples were thawed and air-dried for 90 min. DNA of tsetse flies’ mid-guts was extracted using cethyl trimethyl ammonium bromide (CTAB) according to the protocol described by Maniatis et al. [25] and Navajas et al. [26]. The mid-guts of the tsetse flies were homogenized with a pestle in a CTAB buffer (CTAB 5%; 1 M Tris, pH 8; 0.5 M EDTA pH 8; 5 M NaCl, 20 μL of β-mercaptoethanol and sterile water) and incubated at 60 °C for 30 min. The DNA was extracted from the lysis mixture with chloroform/isoamylic alcohol (24/1; V/V) and precipitated by adding isopropanol (V/V). After centrifugation (10,000 g × 10 min), the pellet was rinsed with 70% ethanol, air-dried, and re-suspended in distilled sterile water (20 μL). A quantity of 2 μL of this extracted DNA were launched on 1.5% agarose gel electrophoresis to check the quality of the extracts. The DNA samples were stored at − 20 °C until PCR amplification.
Confirmation of tsetse fly species
A diagnostic PCR method based on the length polymorphism of ITS1 (internal transcribed spacer 1) fragments between tsetse species was performed to confirm species identification [27]. PCR was used to amplify sequences of ITS1 using the primer pairs given in Table 1. The PCR mix contained 6.45 μL double distilled water, 1.5 μL 10X PCR buffer (Bioline), 0.5 μL of 10 mM dNTP, 1 μL of each 10 pmol primer, 1.5 μL of 50 mM MgCl2 (final concentration 2.5 mM), 0.5 U (0.05 μL) of BIOTaq DNA polymerase. A quantity of 3 μL of 1/10e dilution of template extracted DNA was added to PCR mix. Temperature cycles: 5 min 94 °C, 30 cycles of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 1 min, then 72 °C for 10 min.
Molecular identification of different trypanosomes
PCR amplifications of animal trypanosomes contained in midguts were performed as described by Moser et al. [28], Masiga et al. [29], Majiwa et al. [30] and Herder et al. [31]. In general, a denaturing step at 94 °C for 5 min was followed by 40 amplification cycles. Each cycle included a denaturing step at 94 °C for 30 s, an annealing step at 58 °C (T. brucei sensu lato) or 60 °C (T. congolense “forest” and “savannah” types, T. vivax) for 30 s and an extension step at 72 °C for 1 min. A final extension was performed at 72 °C for 10 min.
Molecular identification of S. glossinidius and Wolbachia sp.
S. glossinidius identification was done according to the protocol described by Darby et al. [32] and Wolbachia sp. was identified following the method of Baldo et al. [33] and Werren and Windsor [34] methods (Table 1). For PCR amplification of S. glossinidius, a denaturing step at 94 °C for 5 min was followed by 40 amplification cycles. Each cycle included a denaturing step at 94 °C for 30 s, an annealing step at 50 °C for 30 s and an extension step at 72 °C for 1 min. A final extension was performed at 72 °C for 10 min. For PCR amplification of Wolbachia sp. only the number of cycles (37) and annealing step (59 °C for 45 s) differed from the Sodalis PCR protocol.
The amplified products were separated on a 2% agarose gel and visualized under UV illumination. Positive controls corresponding to PCR reagents with reference DNA of trypanosome or symbiont species and negative controls (PCR reagent without any DNA) were included in each set of PCR amplification experiments.
Expression of results and statistical analysis
Entomological data were expressed in terms of abundance of tsetse flies, estimated by fly apparent density per trap per day (ADT) according to the following expression: ADT = Nc/TD (where Nc is the number of captured tsetse flies, T, the number of traps and D the number of trapping days.
Trypanosomes and symbionts hosted by tsetse flies were expressed in terms of parasite infection rate and symbiont occurrence rate. The Pearson’s chi-square test (χ2) was used to compare infection rates between villages [35]. A logistic regression model [36] was used to analyse the association between the presence of symbionts (S. glossinidius and Wolbachia sp.) and trypanosome infections in each tsetse fly species. All the statistical tests were performed with the software PASW Statistics 18 (SPSS Inc., Chicago, IL, USA), and the level of significance was fixed at 0.05.
Results
Entomological surveys
A total of 446 tsetse flies belonging to three species or subspecies were caught: 402 (90.1%) Glossina tachinoides, 42 (9.4%) Glossina morsitans submorsitans and only 2 (0.5%) Glossina fuscipes fuscipes. This morphological identification was confirmed by the amplification of ITS1 sequence that revealed a DNA fragment of 221 bp for G. tachinoides and a fragment of less than 221 bp for G. m. submorsitans.
The mean apparent density (ADT) was 1.037. Its values were 0.785 at Mayo-Dagoum and 1.501 at Golde Bourle (Table 2). Of the 446 flies captured, 30 (6.72%) were teneral and 102 were dead/dessicated. For the remaining 314 flies, 270 G. tachinoides and 42 G. m. submorsitans were dissected while the two G. f. fuscipes samples were not, and were excluded from further analyses, because of the small size of this species. Microscopy enabled the identification of midgut infections of trypanosomes in two G. m. submorsitans and one G. tachinoides, giving an infection rate of 0.95% (Table 2).
Molecular identification of different trypanosomes
Of the 270 midguts of G. tachinoides analyzed, 94 (34.81%) were positive with at least one trypanosome species; 72 (44.7%) trypanosome infections in flies of Golde Bourle and 22 (20.2%) in flies of Mayo-Dagoum. A significant difference (χ2 = 17.243; p < 0.0001) in the global infection rate was found between the two villages. The numbers of tsetse flies infected by T. brucei s.l., T. congolense “forest type” and T. congolense “savannah type” were respectively 30 (11.1%), 7 (2.6%) and 37 (13.7%). Trypanosome infection rates were similar between Golde Bourle and Mayo Dagoum for T. brucei and T. congolense “forest type” but different for T. congolense “savannah type” (p = 0.003). No T. vivax infection was identified. When looking at the type of infection (single or mixed), 74 (27.40%) flies carried single infections and 20 (7.40%) carried mixed infections including 18 double infections (13 TcS/TcF, TcS/Tb sl and TcF/Tb sl) and 3 triple infections (TcS/TcF /Tb sl) (Table 3). The rates of single infections (18.3% versus 33.5%) and mixed infections (1.8% versus 11.2%) significantly differed between Mayo Dagoum and Golde Bourle (p = 0.006 and p = 0.004) respectively (Table 3).
Of the 42 midguts of G. m. submorsitans, 17 (40.5%) were infected with at least one trypanosome species: 3 (18.8%) in flies of Mayo Dagoum and 14 (53.8%) in tsetse of Golde Bourle. The global infection rates differed significantly between the two villages (p < 0.05). The rates of single infections differed significantly between Mayo Dagoum (6.3%) and Golde Bourle (46.2%) (p < 0.01). The number of flies infected with each trypanosome species or subspecies were 7 (16.7%), 4 (9.5%), 1 (2.4%) and 1 (2.4%), respectively, for T. brucei s.l., T. congolense “savannah type”, T. congolense forest type and T. vivax. The differences in these infection rates were not tested between the villages because of the low numbers of the infected flies (Table 4).
Molecular identification of symbionts
S. glossinidius was identified in 100 (37%) of the 270 G. tachinoides: 50 (45.9%) in flies of Mayo Dagoum and 50 (31.1%) in flies of Golde Bourle. Comparing the percentages of flies harboring S. glossinidius, a significant difference (χ2 = 6.118; p = 0.013) was obtained between Mayo Dagoum and Golde Bourle (Table 3). For the 42 G. m. submorsitans, S. glossinidius was identified in 12 (28.6%) of them: 7 (43.8%) in Mayo Dagoum and 5 (19.2%) in Golde Bourle (Table 4). These values did not differ between villages (p > 0.05).
Of the 270 G. tachinoides, Wolbachia sp. was identified in 184 (68.1%) of them. Between Mayo Dagoum and Golde Bourle, no significant difference (χ2 = 0.037; p = 0.848) was found in the percentage of flies harboring Wolbachia sp. (Table 3). Of the 42 G. m. submorsitans, Wolbachia sp. was identified in 59.5% of them. Comparing the percentage of flies harboring Wolbachia sp., no significant difference (P > 0.1) was observed between Mayo Dagoum and Golde Bourle (Table 4).
Relationship between trypanosome infections and the presence of symbionts
Of the 100 G. tachinoides harboring S. glossinidius, 37 (37%) were infected by at least one trypanosome species (these flies were noticed (S + trp+)) and 63 (63%) were not (S + trp-). Of the 170 S. glossinidius negative flies, 58 (34.12%) were infected by trypanosomes (S-trp+) while 112 (65.88%) were not (S-trp-). Of 184 G. tachinoides harboring Wolbachia sp., 60 (32.6%) were infected by trypanosomes (W + trp+) and 124 (67.4%) were not (W + trp-). For the remaining 86 G. tachinoides for which Wolbachia sp. was not identified, 32 (37.2%) were infected by trypanosomes (W-trp+). No significant difference was observed between tsetse flies harboring together trypanosome infections and Wolbachia sp., or/and S. glossinidius and flies with only trypanosome infections (Table 5).
Discussion
Tsetse flies trapping
The entomological survey has revealed the presence of three tsetse fly species whereas previous studies detected only two taxa (G. tachinoides and G. m. submorsitans) in forest galleries and pasture bordering the watercourse in the Adamawa savannah [21]. Moreover, the high frequency of G. tachinoides compared to G. m. submorsitans suggests a modification of the distribution compared to previous results [37, 38] that reported high densities of G. m. submorsitans in the Adamawa plateau. This discrepancy may be explained by both environmental and climatic changes that have induced high or low harmful effects according to tsetse fly species, and the development of agriculture zones. The high density of tsetse flies at Golde Bourle (1.506) compared to Mayo-Dagoum (0.785) can be explained by the presence of a forest gallery with diverse wild animals along the great river of Golde Bourle which has created favorable microenvironment for tsetse flies.
Trypanosome infection rate
Trypanosome infection rates were relatively low, varying from 2.4 to 16.7% in G. m. submorsitans and from 2.6 to 13.7% in G. tachinoides, as compared to values reported in G. m. submorsitans (1.5 to 36.4%) and G. tachinoides (2.3 to 20.7%) in Yankari National Park and Wuya areas (Nigeria) by Isaac et al. [39]. T. congolense was the predominant species, confirming the high transmission of that parasite in “Faro and Déo” as previously observed by Mamoudou et al. [6, 40] and Tanembe et al. [22] when studying cattle trypanosomes and the main parasites responsible for AAT in livestock. However, the use of species-specific primers could not reveal all the trypanosome species and might have lowered the infection risk. The association of DNA sequencing of Glyceraldehyde 3-Phosphate Dehydrogenase and the Fluorescent Fragment Length Barcoding methods would have been useful. The low prevalence of T. vivax could be due to the fact that DNA was extracted only from the midguts.
Symbiont occurrence rates
In G. tachinoides, the mean occurrence rate of S. glossinidius (37%) is low compared to 54.9% revealed in G. p. palpalis of Bipindi and Campo in the Southern Cameroon. However, this value is higher than 3.7 and 16% reported, respectively, for G. austeni and G. pallidipes in Shimba Hills National Reserve in Kenya [41]. The differences observed between these studies may be due to the diversity of tsetse species and the occurrence of different genotypes of symbionts [18, 19].
The high prevalence of Wolbachia sp. in G. tachinoides (68.1%) and in G. m. submorsitans (58.5%) does not corroborate results of Doudoumis et al. [42] who reported the absence of Wolbachia in tsetse flies of the G. palpalis group. However, the species G. m. submorsitans was not included in their study. These observations suggest that the prevalence of Wolbachia sp. may depend on the ecological conditions of tsetse fly populations [43, 44].
Sodalis and Trypanosoma sp. relationships
In the present study, no significant association was found between the presence of symbionts and trypanosome infections in G. tachinoides. The lack of association may be due to several reasons: firstly, the low trypanosome infection rate observed in tsetse flies (2.5 to 13.7%) compared to 9.1 to 15.6% in South Cameroon was not a favorable condition for the association tests; secondly, tsetse fly species might differ in whether Sodalis affects them (G. p. palpalis versus G. tachinoides); the association may also vary according to specific genotypes of S. glossinidius rather than the presence or absence of the symbiont as shown by Geiger et al. [18].
S. glossinidius is suspected to be involved in the vector competence of Glossina by favoring parasite installation in the insect midgut through a complex biochemical mechanism involving the production of N-acetyl glucosamine [11, 12]. This sugar was reported to inhibit tsetse-midgut lectin lethal for the procyclic forms of the trypanosome [12, 13]. The presence of S. glossinidius would thus allow the trypanosome to establish in the midgut [12, 45, 46]. The studies conducted by Geiger et al. [17, 18] supported this hypothesis but did not consider it as an absolute condition. Recently, Farikou et al. [10, 19] observed a positive association between the presence of S. glossinidius and the rate of some trypanosome infections in Campo and Bipindi HAT foci in South Cameroon. These results suggested that S. glossinidius could favor establishment of trypanosome infections in G. p. palpalis. The identification of S. glossinidius genotypes occurring in these flies would allow to better understand these relations.
Wolbachia sp. and Trypanosoma sp. relationships
More than 50% of tsetse flies harbored Wolbachia sp. among which 26% of G. tachinoides and 23% of G. m. submorsitans were infected by trypanosomes, but no association test was significant. These tests should be performed with different haplotypes of Wolbachia sp. revealed by genotyping and sequencing of the alleles identified, to allow a better exploration of the possible existence of relationships between the presence of Wolbachia sp. and the level of trypanosome infection.
Conclusions
This study revealed the presence of three species of tsetse flies dominated by G. tachinoides. Some tsetse flies harbored symbionts (Sodalis glossinidius and/or Wolbachia sp.) and animal trypanosomes, but no association between the presence of symbionts and the level of trypanosome infection was significant. Additional studies searching for associations between the presence of some symbiont haplotypes and trypanosome infections are required to better elucidate tsetse fly/symbiont/trypanosome relations.
Abbreviations
- AAT:
-
Animal African trypanosomosis
- ADT:
-
Apparent density per trap per day
- CTAB:
-
Cethyl trimethyl ammonium bromide
- DNA:
-
Deoxyribonucleic acid
- GPS:
-
Global positioning system
- HAT:
-
Human African trypanosomosis
- ITS:
-
Internal transcribed spacer
- MSEG:
-
Special Mission for Tsetse Eradication
- NaOH:
-
Sodium hydroxide
- PCR:
-
Polymerase chain reaction
- Tb:
-
Trypanosoma brucei
- Tcf:
-
Trypanosoma congolense forest type
- Tcs:
-
Trypanosoma congolense savannah type
- Trp:
-
Trypanosoma
References
Talaki E. Etude de la résistance des trypanosomes à l’isométamidium et au diminazène dans la zone cotonnière de l’Afrique de l’Ouest (Mali-Guinée-Burkina Faso). Thèse de doctorat, Université de Bobo Dioulasso. 2008.
Reinhardt E. Travailler ensemble: la mouche tsé-tsé et la pauvreté rurale. Available from: Chronique ONU, ONU Editor. 2002. http://www.un.org/french/pubs/chronique/2002/numero2/0202p17_la_mouche_tsetse.html. Accessed 02 Sept 2002.
Feldmann U, Dyck VA, Mattioli RC, Jannin J. Potential impact of tsetse fly control involving the sterile insect technique. In: Dyck VA, Hendrichs J, Robinson AS, editors. Sterile Insect Technique. Dordrecht: Springer; 2005. p. 701–23.
Kioy D, Jannin J, Mattock N. Human African trypanosomiasis. Nat Rev Microbiol. 2004;2(3):186–7.
Talaki E, Sidibe I, Akoda K, Belem AMG, Pangui LJ. Chimiorésistance aux trypanocides dans les élevages en Afrique subsaharienne. RASPA. 2013;11(S):77–83.
Mamoudou A, Zoli A, Tchoua P. Parasitological prevalence of bovine trypanosomosis in the Faro and Deo division valley of the Adamaoua plateau, Cameroon. Int J Biol Chem Sci. 2009;3(5):1192–7.
Kuzoe FAS, Schofield CJ. Strategic review of traps and targets for tsetse and African trypanosomiasis control. World Health Organization TDR/IDE/TRY/05.1; 2005. p. 58.
Aksoy S. Tsetse-A haven for microorganisms. Parasitol Today. 2000;16:114–8.
Weiss B, Wang J, Aksoy S. Tsetse immune system maturation requires the presence of obligate symbionts in larvae. PLoS Biol. 2011. https://doi.org/10.1371/journal.pbio.1000619.
Farikou O, Njiokou F, Mbida Mbida JA, Njitchouang GR, Djeunga HN, Asonganyi T, Simarro PP, Cuny G, Geiger A. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes an epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect Genet Evol. 2010;10:115–21.
Maudlin I, Ellis DS. Association between intracellular rickettsia-like infections of midgut cells and susceptibility to trypanosome infection in Glossina spp. Z Parasitenkd. 1985;71:683–7.
Welburn SC, Maudlin I. Tsetse–trypanosome interactions: rites of passage. Parasitol Today. 1999;15:399–403.
Dale C, Welburn SC. The endosymbionts of tsetse flies: manipulating host-parasite interactions. Int J Parasitol. 2001;31:628–31.
Bandi C, Anderson TJ, Genchi C, Blaxter ML. Phylogeny of Wolbachia in filarial nematodes. Proc R Soc Lond B. 1998;265:2407–13.
Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008;6:741–51.
Hoffmann AA, Hercus M, Dagher H. Population dynamics of the Wolbachia infection causing cytoplasmic incompatibility in Drosophila melanogaster. Genetics. 1998;148:221–31.
Geiger A, Ravel S, Frutos R, Cuny G. Sodalis glossinidius (Enterobacteriaceae) and vectorial competence of Glossina palpalis gambiensis and Glossina morsitans morsitans for Trypanosoma congolense savannah type. Curr Microbiol. 2005;51:35–40.
Geiger A, Ravel S, Mateille T, Janelle J, Patrel D, Cuny G, Frutos R. Vector competence of Glossina palpalis gambiensis for Trypanosoma brucei s.l. and genetic diversity of the symbiont Sodalis glossinidius. Mol Biol Evol. 2007;24:102–9.
Farikou O, Njiokou F, Cuny G, Geiger A. Microsatellite genotyping reveals diversity within populations of Sodalis glossinidius, the secondary symbiont of tsetse flies. Vet Microbiol. 2011;150:207–10.
Sarwissi S. Rapport annuel (1996). Ngaoundéré: Mission spéciale d’éradication des glossines; 1997. p. 52.
Cuisance D, Boutrais J. Evaluation de la situation et de la stratégie de lutte contre les glossines et les trypanosomoses dans l’Adamaoua (Cameroun). Rapport de mission. Maisons-Alfort, France, Cirad-emvt; 1995. p. 63.
Tanenbe C, Gambo H, Musongong AG, Boris O, Achukwi MD. Prévalence de la trypanosomose bovine dans les départements du Faro et Déo, et de la Vina au Cameroun: bilan de vingt années de lutte contre les glossines. Revue Elev Méd Vét Pays Trop. 2010;63(3–4):63–9.
Pollock JN. Manuel de lutte contre la mouche Tsé-tsé: Biologie, systématique et répartition des tsé-tsé, vol. 1. Rome: FAO; 1992. p. 310.
Grébaut P, Mbida JA, Kondjio CA, Njiokou F, Penchenier L, Laveissiere C. Spatial and temporal patterns of human African trypanosomiasis (HAT) transmission risk in the Bipindi focus, in the forest zone of southern Cameroon. Vector Borne Zoo Dis. 2004;4:230–8.
Maniatis T, Fritsch EF, Sambrook J. Molecular cloning: a laboratory manual. New York: Cold Spring Harbor Publications; 1982.
Navajas M, Lagnel J, Gutierrez J, Boursot P. Species-wide homogeneity of nuclear ribosomal ITS2 sequences in the spider mite Tetranychusurticae contrasts with extensive mitochondrial COI polymorphism. Heredity. 1998;80:742–52.
Dyer NA, Lawton SP, Ravel S, Choi KS, Lehane MJ, Robinson AS, Okedi LM, Hall MJR, Solano P, Donnelly MJ. Molecular phylogenetics of tsetse flies (Diptera: Glossinidae) based on mitochondrial (COI, 16S, ND2) and nuclear ribosomal DNA sequences, with an emphasis on the palpalis group. Mol Phylogenet Evol. 2008;49:227–39.
Moser DR, Cook GA, Ochs DE, Bailey CP, McKane MR, Donelson JE. Detection of Trypanosoma congolense and Trypanosoma brucei subspecies by DNA amplification using the polymerase chain reaction. Parasitology. 1989;99:57–66.
Masiga DK, Smyth AJ, Ayes P, Bromidge TJ, Gibson WC. Sensitive detection of trypanosomes in tsetse flies by DNA amplification. Int J Parasitol. 1992;22:909–18.
Majiwa PAO, Thatthi R, Moloo SK, Nyeko JHP, Otieno LH, Maloo S. Detection of trypanosome infections in the saliva of tsetse flies and buffy-coat samples from antigenaemic but aparasitaemic cattle. Parasitology. 1994;108:313–22.
Herder S, Simo G, Nkinin S, Njiokou F. Identification of trypanosomes in wild animals from southern Cameroon using the polymerase chain reaction (PCR). Parasite. 2002;9:345–9.
Darby AC, Lagnel J, Matthew CZ, Bourtzis K, Maudlin I, Welburn SC. Extrachromosomal DNA of the symbiont Sodalis glossinidius. J Bacteriol. 2005;187:5003–7.
Baldo L, Dunning HJC, Jolley KA, Bordenstein SR, Biber SA, Choudhury RR, Hayashi C, Maiden MC, Tettelin H, Werren JH. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol. 2006;72:7098–110.
Werren JH, Windsor DM. Wolbachia infection frequencies in insects: evidence of a global equilibrium? Proc Biol Sci. 2000;267(1450):1277–85.
Schwartz D. Méthodes statistiques à l’usage des médecins et des biologistes. 4th ed. Paris: Médecine Sciences Flammarion; 1996. p. 314.
Bouyer J, Hemon D, Cordier S, Derriennic F, Stucker I, Stengel B, Clavel J. Epidémiologie-Principes et Méthodes quantitatives. Les Editions INSERM. 1993.
Banser JT. The evolution of tsetse and trypanosomiasis control in Cameroon. International Scientific Council for Trypanosomiasis Research and Control (ISCTRC) 16th Meeting, Yaoundé; 1979. p. 459–61.
Hurault J. Comment a été vécue à Banyo et à Tibati la crise résultante de l’invasion des glossines (1990–1991). Cameroun: Rapport de mission; 1993.
Isaac C, Ciosi M, Hamilton A, Scullion KM, Dede P, Igbinosa IB, Nmorsi OPG, Masiga D, Turner CMR. Molecular identification of different trypanosome species and subspecies in tsetse flies of northern Nigeria. Parasit Vectors. 2016;9:301.
Mamoudou A, Zoli A, Mbahin N, Tanenbe C, Bourdanne, Clausen P-H, Marcotty T, et al. Prevalence and incidence of bovine trypanosomosis on the Adamaoua plateau in Cameroon ten years after the tsetse eradication campaign. Vet Parasitol. 2006;142:16–22.
Wamwiri FN, Alam U, Thande P, Aksoy E, Ngure RM, Aksoy S, et al. Wolbachia, Sodalis and trypanosome co-infections in natural populations of Glossina austeni and Glossina pallidipes. Parasit Vectors. 2013;6:232.
Doudoumis V, Tsiamis G, Wamwiri F, Brelsfoard C, Alam U, Aksoy E, et al. Detection and characterization of Wolbachia infections in laboratory and natural populations of different species of tsetse flies (genus Glossina). BMC Microbiol. 2012;1:S3.
Mouton L, Henri H, Charif D, Boulétreau M, Vavre F. Interaction between host genotype and environmental conditions affects bacterial density in Wolbachia symbiosis. Biol Lett. 2007;3(2):210–3.
Yun Y, Lei C, Peng Y, Liu F, Chen J, Chen L. Wolbachia strains typing in different geographic population spider, Hylyphantes graminicola (Linyphiidae). Curr Microbiol. 2010;62(1):139–45.
Welburn SC, Arnold K, Maudlin I, Gooday GW. Rickettsia-like organisms and chitinase production in relation to transmission of trypanosomes by tsetse flies. Parasitology. 1993;107:141–5.
Dale C, Maudlin I. Sodalis gen. nov. and Sodalis glossinidius sp. nov., a microaerophilic secondary endosymbiont of the tsetse fly Glossina morsitans morsitans. Int J Syst Bacteriol. 1999;49(1):267–75.
Acknowledgements
The authors thank M. Nana Djeunga Hugues Clotaire for his help in statistical analysis.
Funding
This study received IAEA funds through Coordinated Research Project D42015 (Enhancing Vector Refractoriness to Trypanosome Infection). The Laboratory phase was hosted by the University of Yaoundé 1.
Availability of data and materials
All the datasets are presented in the main manuscript. The raw datasets used and or analysed during the current study are available from the corresponding author on reasonable request.
About this supplement
This article has been published as part of BMC Microbiology Volume 18 Supplement 1, 2018: Enhancing Vector Refractoriness to Trypanosome Infection. The full contents of the supplement are available online at https://bmcmicrobiol.biomedcentral.com/articles/supplements/volume-18-supplement-1.
Author information
Authors and Affiliations
Contributions
GIKN collected field data, carried out molecular biology experiments and drafted the manuscript. FN participated in the conception and the coordination of the study, designed the experiments, improve the quality of the manuscript. TTMT collected field data. OF collected field data and helped to draft the manuscript. GS participated in the conception of the study and in the design of experiments, collected field data and helped to improve the quality of the manuscript. AG participated in the conception of the study and pre-reviewed the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
This is an open access article distributed under the terms of the Creative Commons Attribution IGO License (https://creativecommons.org/licenses/by/3.0/igo/) which permits unrestricted use, distribution, and reproduction in any medium, provided appropriate credit to the original author(s) and the source is given.
About this article

Cite this article
Kame-Ngasse, G.I., Njiokou, F., Melachio-Tanekou, T.T. et al. Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa region of Cameroon. BMC Microbiol 18 (Suppl 1), 159 (2018). https://doi.org/10.1186/s12866-018-1286-5
Published:
DOI: https://doi.org/10.1186/s12866-018-1286-5