
Abstract
Cold stress adversely affects rice (Oryza sativa L.) growth and productivity, and has so far determined its geographical distribution. Dissecting cold stress-mediated physiological changes and understanding their genetic causes will facilitate the breeding of rice for cold tolerance. Here, we review recent progress in research on cold stress-mediated physiological traits and metabolites, and indicate their roles in the cold-response network and cold-tolerance evaluation. We also discuss criteria for evaluating cold tolerance and evaluate the scope and shortcomings of each application. Moreover, we summarize research on quantitative trait loci (QTL) related to cold stress at the germination, seedling, and reproductive stages that should provide useful information to accelerate progress in breeding cold-tolerant rice.
Similar content being viewed by others


Introduction
Crops are exposed to varied environmental conditions during their life cycle. Cold stress, which can be classified as chilling (0-15°C) and freezing (<0°C) stress, is a major environmental factor limiting the growth, productivity, and geographical distribution of crops (Zhu et al. [2007]). Rice (Oryza sativa L.), one of the most world's most important staple crops, feeds more than 2.7 billion people worldwide and is extensively grown by more than half of the world's farmers (Fairhurst and Dobermann [2002]; Shelton et al. [2002]). Due to its origin in tropical and subtropical regions, rice is more sensitive to cold stress than other cereal crops such as wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.). Therefore, in temperate areas, the production of rice is severely limited by cold stress (Xie et al. [2012]). Low temperatures that occur at critical reproductive stages can adversely affect grain quality or cause yield reductions in high-latitude or high-altitude regions of China, Japan, Korea, and other parts of the world (Jena et al. [2012]).
Over the past 20 years, extensive efforts have been made to improve cold tolerance in rice, which is a very complex trait (Maruyama et al. [2014]). Cold stress affects chlorophyll content and fluorescence, and thus interferes with photosynthesis in rice (Kanneganti and Gupta [2008]; Kim et al. [2009]). Moreover, increased contents of reactive oxygen species (ROS) and malondialdehyde (MDA) that accumulate during cold stress in rice can impair metabolism via cellular oxidative damage (Xie et al. [2009]; Nakashima et al. [2007]). On the other hand, rice also possesses strategies to cope with or adapt to cold stress. For example, cold-treated rice plants accumulate proline, an amino acid that stabilizes protein synthesis, and thereby maintains the optimal function of rice cells (Kandpal and Rao [1985]). Under cold stress, contents of antioxidant species also increase to scavenge ROS and protect rice plants against oxidative damage (Sato et al. [2011]). Such physiological changes that occur upon cold treatment of rice, whether mediators or symptoms of cold damage, can also be used as indicators to evaluate the cold tolerance of rice.
Due to diverse growing locations and climatic factors, rice cultivars face cold stress at specific growth stages (Saito et al. [2001]). Researchers have established many growth-stage specific criteria to evaluate and select cold-tolerant rice. Evaluation of rice cultivars typically takes place during seedling and reproductive stages that are critical to production of rice. However, in high-latitude or high-altitude regions, low temperatures during long, cold springs can severely inhibit germination and constrain early seedling growth. So evaluation of cold tolerance at the germination stage is especially significant for these regions.
As the development of molecular markers and linkage maps progresses, marker-assisted selection becomes an effective way to breed cold-tolerant cultivars. Many cold-tolerance related QTL have been identified in the past 20 years. The QTL Ctb1, qCTB2a, qPSST-3, qLTB3 are related to cold tolerance at the reproductive stage; qCTP11 is related to cold tolerance at the germination stage; and qCtss11 and qCTS4a are related to cold tolerance at the seedling stage. Because breeding for abiotic-stress resistance is urgently needed, progress in the identification of cold tolerance-related QTL has been a significant development for facilitating molecular marker-assisted selection (MAS).
In this review, we focus on clarifying the role of various metabolites during the response to cold stress in rice, and summarize the diverse criteria that are useful for evaluating the cold tolerance of rice at different growth stages. In addition, we discuss QTL and markers related to cold tolerance that can be used to facilitate marker-assisted breeding through recurrent selection in rice.
Review
Changes in physiological parameters of rice under cold stress
Low temperature not only inflicts obvious physical damage to rice plants, including low germination rate, stunted seedling growth, high death rate, and low spikelet fertility, but also initiates physiological fluctuations, such as increased electrolyte leakage (EL), changes in chlorophyll fluorescence, and increases in amounts of ROS, MDA, sucrose, lipid peroxidation, proline, and other metabolites. Analysis of metabolites in cold-stressed Arabidopsis by gas chromatography-mass spectrometry (GC-MS) has detected a total of 434 low-molecular-weight carbohydrates, amines, organic acids, and other polar molecules. Of these metabolites, levels of 325 (75%) had increased, and levels of 114 (35%) metabolites had increased at least 5-fold upon cold stress (Cook et al. [2004]). These metabolites could be organized into six categories related to 1) photosynthesis, 2) electrolyte leakage, 3) ROS and MDA, 4) stress-related soluble sugars, 5) cold-response related amino acids, and 6) antioxidants (Table 1). Thus, changes in the concentrations of these metabolites can be useful indicators for analysis of cold-stress responses in rice.
Chlorophyll content and fluorescence indicate effects of cold stress on photosynthesis
Chlorophyll content and fluorescence, measured as the ratio of variable fluorescence to maximum fluorescence (Fv/Fm), are two photosynthetic properties whose changes are relevant to cold responses in plants. Chlorophyll content can signify nutrient stress in general, and nitrogen or sulfur stress in particular (Haboudane et al. [2002]). Cold stress can inhibit chlorophyll synthesis and chloroplast formation in rice leaves. Thus, reduced chlorophyll content can indicate the effect of low temperature on rice plants (Sharma et al. [2005]).
Fv/Fm is a measure of chlorophyll fluorescence that is commonly used to determine the maximum quantum efficiency of Photosystem II (PSII), which indicates whether cold stress has compromised PSII in its dark-adapted state (McFarlane et al. [1980]). During cold stress, Fv/Fm values decrease slightly in plants that tolerate cold, but decrease significantly in plants that are sensitive to cold (Bonnecarrère et al. [2011]; Zahedi and Alahrnadi [2007]). This parameter is thus useful for assaying the cold tolerance and sensitivity of plants that differ for this trait due either to genetics or acclimation.
The expression of several genes known to be involved in stress signal transduction can influence chlorophyll content or fluorescence. For example, chlorophyll content is much higher compared to that of control plants in both rice and tobacco transgenic lines overexpressing OsiSAP8, a member of the SAP gene family in rice. OsiSAP8 encodes a cytoplasmic zinc finger protein that acts early in the signal transduction of responses to various stresses including cold (Kanneganti and Gupta [2008]).
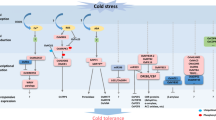
Additionally, ectopic expression of another cold-responsive gene, OsAsr1, which functions in abscisic acid (ABA)-response and fruit ripening, results in twofold higher values for Fv/Fm in transgenic rice seedlings, a finding consistent with enhanced cold tolerance (Kim et al. [2009]). The phytohormone ABA is involved in various aspects of plant growth and development, and one of its major roles is to mediate adaptive responses to various environmental stresses (Hossain et al. [2010b]). In cold-stressed rice, ABA accumulates and initiates the ABA signaling cascade, which affects the expression of ABA-responsive genes via cis-acting ABA-response elements (ABRE) and the ABRE-binding bZIP transcription factor (ABF) (Hossain et al. [2010a]). The OsNAC gene transduces the ABA signal through an ABRE in its promoter and regulates the expression of NACRS-containing genes to control cold tolerance in rice (Figure 1A) (Nakashima et al. [2012]; Nakashima et al. [2007]; Song et al. [2011]).

Cold sensing and responsive pathway in rice. (A) ABA signaling is initiated by ABA accumulation, and is transduced by ABF and the ABRE-containing OsNAC gene, which regulates the expression of a NACRS-containing target gene to increase cold tolerance. (B) The DREB-CRT/DRE pathway is initiated by a Ca2+-influx signal that is transduced by calcium-dependent protein kinases (CDPKs), OsMYB3R-2, OsMYB2, and OsICE1/2. The expression of OsDREB1B and OsDREB2A are down-regulated by MYBS3. (C) The MAPK cascade (OsMKK6-OsMPK3 and OsMPK6) is activated by ROS and is negatively regulated by OsTRX23. Solid arrows indicate direct activation; broken arrows indicate indirect activation; lines ending with a bar indicate negative regulation.
Yamburenko reported that exogenous ABA reduced chlorophyll content and differentially affected the expression of chloroplast genes, suggesting a role of ABA in the regulation of proteins with functions in photosynthesis in barley leaves (Yamburenko et al. [2013]). However, in rice, the role of ABA in regulation of photosynthesis-related genes has not yet been fully explained. Therefore, progress in this field will help to bridge the gaps in our knowledge of the crosstalk between cold stress, ABA signaling, and photosynthesis.
Changes in membrane fluidity initiate cellular cold responsive
Changes in ambient temperature can affect cell membranes quickly, although this is a reversible process (Murata and Los [1997]; Los and Murata [2004]; Sangwan et al. [2002]). Changes in membrane fluidity are measured by the membrane polarization index, p, an inverse indicator of membrane fluidity (Sangwan et al. [2002]).
Rice cells can sense cold stress based on changes in membrane rigidity, the physical state of membrane proteins, and osmotic pressure (Los and Murata [2004]). Low temperatures initiate increased membrane rigidity and can lead to increased EL, which can indicate the activity of cold tolerance-relevant genes, including TERF2, OVP1, and OsNAC5 (Yun et al. [2010]; Tian et al. [2011]; Zhang et al. [2011a]; Song et al. [2011]). Functional characterization of the cold-activated Stress-activated MAPK (SAMK) reveals that a MAPK signaling cascade is triggered by increased membrane rigidity and altered ion conductance within cells and tissues (Sangwan et al. [2002]). Also, the influx of Ca2+ into the cytoplasm, an early event in cold stress, may be mediated by Ca2+ channels that are activated by membrane rigidification, ligands, or mechanical stimuli (Chinnusamy et al. [2006]). This cold-sensing signal can be interpreted and amplified by a calcium signaling cascade that subsequently activates the DREB-CRT/DRE (dehydration-responsive element-binding proteins-C-repeat/dehydration-responsive elements) pathway in rice (Zhang et al. [2013]), which is an important cascade for cold sensing and response in rice (Figure 1B) (Zhu et al. [2007]; Chinnusamy et al. [2007]).
ROS and MDA mediate cold damage and cold sensing in rice
ROS are chemically reactive molecules that contain oxygen, including superoxide (O2-), hydrogen peroxide (H2O2), or the hydroxyl radical (HO-) that are produced at low levels as normal byproducts of plant cellular metabolism, mainly in organelles such as chloroplasts, mitochondria, and peroxisomes. However, both biotic and abiotic stresses can lead to excessive production of ROS that can then react rapidly with proteins, DNA, and lipids to cause cellular oxidative damage (Apel and Hirt [2004]; Skopelitis et al. [2006]; Mittal et al. [2012]).
In chloroplasts, ROS may cause over-reduction of the electron transport chain, limit CO2 fixation, and interfere with the photosynthetic process. ROS can also cause damage during stress through their effects upon the electron transport chain in mitochondria (Suzuki and Mittler [2006]). ROS degrade polyunsaturated lipids to form MDA, a reactive aldehyde that initiates toxic stress in cells and subsequently causes cellular dysfunction and tissue damage (Pamplona [2011]). In a study evaluating QTL associated with cold tolerance, elevated MDA content (0.00–0.57 nmol–g-1 FW) was found in 92.16% (47/51) of rice cultivars (Kim and Tai [2011]). Other studies have revealed that the accumulation of ROS in cells of cold-treated rice triggers expression of cold-responsive genes and regulation of the cold-responsive signaling network via the OsMKK6-OsMPK3 (MAPKK-MAPK) pathway (Figure 1C) (Xie et al. [2009]).
Soluble sugars, proline, and antioxidants protect rice from further damage due to cold stress
Soluble sugars that accumulate in plants under stress include sucrose, hexose, raffinose, glucose, fructose, and trehalose. These sugars act as compatible solutes in freezing stress, serving as osmoprotectants against freezing-dehydration damage (Nagao et al. [2005]; Shao et al. [2007]; Yuanyuan et al. [2010]). Because sucrose, trehalose, raffinose, and stachyose contents can increase under low temperature, these metabolites can be used as indicators to evaluate the potential cold tolerance of rice varieties (Morsy et al. [2007]).
Moreover, proline accumulation is also enhanced by cold stress. In addition to acting as a reservoir of carbon and nitrogen, proline also protects cellular enzymes from denaturation (Shah and Dubey [1997]). Proline stabilizes the polyribosome and thereby maintains the operation of protein synthesis (Kandpal and Rao [1985]). Proline is also involved in removal of stress-related excess H+ and maintains oxidative respiration at optimal cytosolic pH (Venekamp [1989]). Moreover, proline increases protein water-binding ability through its hydrophobic interactions with the surface residues of proteins (Schobert and Tschesche [1978]). Increased proline content has been widely observed in rice varieties under low temperatures. Finally, the significant correlations between proline contents and cold tolerance have help to confirm the function of proline during the cold response in rice (Kim and Tai [2011]).
In another metabolic adaptation, antioxidant species scavenge ROS to protect rice plants against oxidative damage induced by cold stress. These antioxidant species include ascorbic acid (AsA) and glutathione (GSH) (Kim and Tai [2011]; Xie et al. [2009]). Such antioxidants occur at high concentrations in chloroplasts and other cellular compartments, and are crucial for defense of plants against oxidative stress (Noctor and Foyer [1998]; Mittler [2002]). Sato reported that overexpression of OsAPXa, an ascorbate peroxidase gene, improves cold tolerance by increasing ascorbate peroxidase activity to reduce the levels of H2O2 and lipid peroxidation under cold stress (Sato et al. [2011]).
Current criteria for the evaluation of cold tolerance in various rice cultivars
Evaluation of rice cultivars for improved cold tolerance is mainly performed during the seedling and reproductive stages. Cold tolerance at the germination stage is a significant component of rapid seedling establishment and development of a uniform crop stand, especially in direct-seeding production systems (Krishnasamy and Seshu [1989]). Because rice is cultivated in regions that range from tropical to temperate, considerable variation in the extent of cold tolerance is exhibited by seedlings of various O. sativa germplasm accessions (Kim and Tai [2011]; Sharifi [2010]). So evaluation of cold tolerance at the seedling stage is important for adaptation to high-altitude or high-latitude locations that can experience cold temperatures at night or early or late in the growing season. Finally, cold tolerance during reproductive stages is critical for pollen survival, seed set, and grain filling to ensure maximal yield (Suh et al. [2010]). Criteria for the evaluation of cold tolerance in rice at different growth stages are summarized in Table 2 and described in detail below.
Evaluation of cold tolerance in rice at the germination stage
Germination vigor and seedling survival rate are the two main criteria used for the evaluation of cold tolerance in rice at the germination stage. The vigor of seed germination is recorded at 7 d, 11 d, 14 d, and 17 d following germination at 14°C in the dark.
Germination vigor (%) = (Number of germinated grain/Number of total grain) × 100.
The standard assessment of whether a rice grain has germinated is determined as the point at which the bud length equals half the length of the seed, and the root length equals the seed length (Han et al. [2006]).
The seedling survival rate for cold tolerance is evaluated as follows. When shoots are about 5 mm long, the germinated seedlings are planted in soil and are subjected to cold treatment at 2°C for 3 d, and are then moved to a sunny indoor environment where the temperature is above 20°C to ensure normal growth. Seedling survival rates are assessed after 7 d recovery growth and cold tolerance evaluation indices are calculated as:
Seedling survival rate (%) = surviving seedlings/budding seeds × 100 (Zhou et al. [2012]).
During cultivation, rice seeds are germinated in early spring at the appropriate temperature (usually 32°C), and the germinated seeds are then sown in the field when the shoots are approximately 5 mm long. Thus, rice seedlings are more likely to encounter cold stress after sprouting than during germination. Therefore, seedling survival rate (%) is a more practical criterion for the evaluation of cold tolerance than are traits that are assessed at the germination stage.
Evaluation of cold tolerance in rice at the seedling stage
Both visual and physiological indicators are used to evaluate cold tolerance at the seedling stage in rice. Five criteria are typically used for visual assessment of cold tolerance, including fresh weight, survival rate, new leaf emergence, seedling growth, and leaf growth.
As water loss often occurs concomitantly with plant damage, changes in fresh weight can be used to indicate water loss and growth retardation of rice plants under cold stress (Bonnecarrère et al. [2011]). However, water loss is not always an accurate indicator of cold stress because it can also be affected by traits including plant variety and leaf size, and also by other stressors.
The survival rate (%) (the number of surviving plants divided by the total number of plants treated × 100) is determined after 4°C treatment, which is a severe condition for rice growth that explicitly distinguishes the cold tolerance of cultivars in short time periods ranging from 6 to 7 d (Zhang et al. [2011b]). As this method clearly and efficiently distinguishes degrees of cold tolerance among cultivars and individuals, it is recommended for gauging cold tolerance in the laboratory. Moreover, new leaf emergence can also be used to assess cold tolerance in transgenic rice in terms of maintained vigor and increased growth (Xie et al. [2012]). For some cold-tolerant wild type plants and their transgenic lines, cold stress (4°C treatment) only retards growth rather than causing lethal damage. So, new leaf emergence would be a better choice for distinguishing the cold tolerance of these lines.
The seedling growth scale is derived from the standard evaluation system for rice that was developed by the International Rice Research Institute (IRRI [2002]). After 14 d of cold treatment at 9°C, this assay is performed by scoring seedlings as 1 = dark green seedlings, 3 = light green seedlings, 5 = yellow seedlings, 7 = brown seedlings, 9 = dead seedlings (Kim and Tai [2011]; Andaya and Tai [2006]). Using a similar visual scaling approach, the leaf growth scale is based on the degree of leaf wilting and is scored on a scale of 1-3 (tolerant, all leaves normal, no apparent visual injury) to 4-9 (susceptible, all leaves wilted, seedlings apparently dead) (Suh et al. [2012]). However, unlike the seedling growth scale, in which seedlings are examined immediately after low temperature treatment, the leaf growth assay is based on symptoms that are apparent on the 7th day of the recovery period (Suh et al. [2012]). Because of the moderate treatment (9-10°C), the seedling growth scale and leaf growth assay are often used to show minor differences in cold tolerance between cultivars. Moreover, because the recovery period at 25°C lasts 7 d, the leaf growth assay can better distinguish cultivars with similar cold tolerance than can the seedling growth scale.
As visual ratings are limited by their tendency to be subjective, physiological parameters thus complement the evaluation of cold tolerance in rice. These parameters include measurements of EL, proline, MDA, AsA, and GSH (Tian et al. [2011]; Zhang et al. [2011a]; Yang et al. [2012]), changes in which can be attributed to cold-responsive metabolism. For example, temperate O. japonica varieties exhibit less EL, while O. indica varieties tend to accumulate higher levels of proline, MDA, AsA, and GSH (Kim and Tai [2011]). The differences in these physiological parameters also indicate the relative degrees of cold tolerance among rice cultivars. Moreover, because MDA is related to the accumulation of ROS, the examination of MDA can indicate whether the cold tolerance in transgenic lines is related to ROS signaling.
In addition, activities of antioxidant enzymes such as peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) can also be used to evaluate cold tolerance in rice. The higher activities of antioxidant enzymes often expressed in transgenic rice lines can indicate their relatively improved cold tolerance compared to wild-type plants (Bonnecarrère et al. [2011]; Huang et al. [2009]). Increased activities of antioxidant enzyme, including OsPOX1, APXa, and related kinase OsTrx23 (Kim et al. [2011]; Sato et al. [2011]; Xie et al. [2012]) may serve to maintain the functions of many other genes during cold stress. Therefore, rather than only physical evaluation of whole-plant cold tolerance, evaluation of antioxidant enzyme activities could both reveal cold-tolerant plants and reveal the mechanisms of cold tolerance.
Evaluation of cold tolerance in rice at the reproductive stage
Exposure to low temperatures during the reproductive stage in rice can cause male sterility and thereby severe yield loss. Cold tolerance at this stage can be evaluated by spikelet fertility based on cold greenhouse cultivation (CGC) or cold deep-water irrigation (CDWI).
To obtain synchronously developing panicles for calculating spikelet fertility using CGC, extra tillers are removed from each greenhouse-grown (26°C/19°C day/night temperature regime) plant at the tillering stage, leaving the main culm. Pollen development is estimated using the auricle distance method (Satake and Hayase [1970]). When the auricle of the flag leaf is approximately 5 cm below the auricle of the penultimate leaf on each plant, the pollen should have undergone meiosis, and each pot is then transferred to a greenhouse maintained at 12°C. After 5-6 d, the cold-treated pots are returned to the warmer greenhouse, where they remain until the plants reach maturity (Andaya and Mackill [2003b]; Suh et al. [2010]). At heading, each plant is tagged with the heading date to verify the developmental stage that pollen had reached after cold treatment, then mean spikelet fertility is calculated for the evaluation of cold tolerance (Sato et al. [2011]).
Spikelet fertility can also be assessed after cold treatment by CDWI. When young panicles begin to differentiate, rice plants are transferred to tanks filled to 20-25 cm depth with water maintained at 18–19°C. Plants are maintained in the cold deep-water irrigation tanks during the entire booting stage. Spikelet fertility is calculated based on the percentage of the number of fertile grains relative to the number of florets (Shirasawa et al. [2012]). Spikelet fertility using CDWI was developed to examine the cold tolerance of rice accessions about 30 years ago, and it is still widely used for selecting cold-tolerant lines because it is highly reliable (Matsunaga [2005]; Shimono et al. [2011]; Shirasawa et al. [2012]).
Both of these methods can be used to select more cold-tolerant rice varieties. Compared to CGC, CDWI exposes plants to a more moderate treatment temperature and a longer treatment period, and is conducted directly in field. Therefore, CDWI is more suitable for distinguishing the cold tolerance of relatively cold-sensitive cultivars and for evaluating the cold tolerance of QTL mapping populations.
QTL identified in various cultivars facilitate the breeding of cold-tolerant rice
Cold tolerance in rice is a quantitative trait controlled by multiple genes. Because it is often difficult to directly associate plant phenotypes with the genes responsible for cold tolerance, marker-assisted selection is an effective means of developing cold-tolerant cultivars (Shirasawa et al. [2012]; Foolad et al. [1999]). The development of molecular markers and linkage maps has made it possible to identify QTL that control cold tolerance in rice (Table 3). QTL analyses have been carried out using rice populations with large levels of genetic variation for cold tolerance (Futsuhara and Toriyama [1969]).
QTL related to cold tolerance at the germination stage
A set of F2:3 populations including 200 individuals and lines derived from a cross between the indica and japonica varieties `Milyang 23' x `Jileng 1' was used to locate QTL for low-temperature vigor of germination (LVG) and to develop a cold-response index for vigor of germination (CIVG). In that study, the QTL qLVG2 was detected in the region of RM29-RM262 on chromosome 2, and qLVG7-2 and qCIVG7-2 were mapped to the region near RM336-RM118 on chromosome 7 (Han et al. [2006]). A major QTL for low temperature germination ability, qLTG3-1, was identified by Fujino on chromosome 3 using backcross inbred lines (BIL) derived from a cross between `Italica Livorno' and `Hayamasari' (varieties with vigorous or weak low-temperature germination ability, respectively) (Fujino et al. [2008]). This QTL explains 30% of the total phenotypic variation for low-temperature germination in their mapping population and was thought to be involved in tissue weakening, a key process during seed germination.
Two major QTL (qCTP11 and qCTP12) for cold tolerance at the plumule stage were identified in genetic stocks derived from 34 cultivated (Oryza sativa) and 23 wild (Oryza rufipogon) rice strains (Baruah et al. [2009]). Both tropical and temperate japonica subpopulations, and also annual, intermediate, and perennial O. rufipogon types were used for this study (Vaughan et al. [2003]). Cold tolerance was scored based on vigor of germination. In another study, cold tolerance at the bud burst stage (CTB) was evaluated at 5°C in a set of 95 chromosome-segment substitution lines (CSSL), derived from indica rice accession 9311 and japonica rice cultivar `Nipponbare', which has the genetic background of 9311. In this study, QTL qCTB-5-1, qCTB-5-2, and qCTB-5-3 were mapped in the regions of RM267-RM1237, RM2422-RM6054, and RM3321-RM1054, respectively, at positions 21.3 cM, 27.4 cM, and 12.7 cM on rice chromosome 5. Additionally, the QTL qCTB-7 mapped to a 6.8-cM region near RM11-RM2752 on rice chromosome 7 (Lin et al. [2010]).
QTL related to cold tolerance at the seedling stage
A set of recombinant inbred lines (RIL) that were derived from a cross between M202 and IR50 (indica, highly sensitive to cold stress) was used to identify QTL conferring tolerance to cold stress at the seedling stage. Using these RIL, Andaya and Mackill mapped a major QTL, qCTS12a, to chromosome 12 that accounted for 41% of the phenotypic variation in seedling growth after cold stress. A number of other QTL with smaller effects have also been detected on eight rice chromosomes (Andaya and Mackill [2003a]). Using RM5746 and RM3103, Andaya and Tai screened 1954 5-F10 lines to find recombinants in the qCTS12 region. Additional microsatellite markers were identified from publicly available genomic sequences and used to fine map qCTS12 within a region of approximately 87 kb on the BAC clone OSJNBb0071I17. Subsequently, open reading frame analyses delimited the QTL to a region of about 55 kb. The most likely candidates for the gene(s) underlying qCTS12 are OsGSTZ1 and OsGSTZ2 (Andaya and Tai [2006]). OsGSTZ1 appears to function in improving cold tolerance at the germination stage, as overexpression of this gene in transgenic rice triggered enhanced germination and growth of seedlings at low temperature (Takesawa et al. [2002]). Moreover, the qCTS4 locus, which is associated with tolerance to yellowing and stunting of rice seedlings, was mapped to chromosome 4 using the same RIL mapping population (Andaya and Tai [2007]).
A set of 282 F13 RIL (recombinant inbred lines) derived by single-seed descent from a cross between `Lemont' (japonica) and `Teqing' (indica) were used to map QTL controlling seedling vigor. A total of 34 QTL for seedling vigor were identified among these RIL. Of the QTL identified, the majority (82%) clustered within five genomic regions, and these were designated qSV-3-1, qSV-3-2, qSV-5, qSV-8-1, and qSV-8-2, respectively (Zhang et al. [2005]). A major QTL (LOD = 15.09), qCTS-2, was detected on chromosome 2 flanked by RM561 and RM341 in doubled haploid (DH) lines that were derived from a cross between a cold-tolerant japonica variety (`AAV002863') and a cold-sensitive indica cultivar (`Zhenshan97B') (Lou et al. [2007]). An F2 population derived from a cold-tolerant wild relative of rice, W1943 (Oryza rufipogon), and a sensitive indica cultivar, `Guang-lu-ai 4' (GLA4), were screened by seedling growth to identify QTL that control cold tolerance in rice. A major QTL for cold tolerance at the seedling stage, qCtss11, was fine mapped to a 60 kb candidate region defined by markers AK24 and GP0030 on chromosome 11, in which six genes have been annotated. Expression analyses and resequencing of these six candidate genes indicated that the Os11g0615600 gene is expressed only from the GLA4 allele, and that the Os11g0615600 gene has a premature stop codon in the GLA4 haplotype. This suggests that either Os11g0615600 or Os11g0615900, or both, might control seedling cold tolerance in this population derived from the cross between W1943 and GLA4 (Koseki et al. [2010]). Moreover, the deduced protein from the Os11g0615900 gene contains the NB-ARC (nucleotide-binding adaptor shared by APAF-1, R protein, and CED-4) domain, a conserved motif found in disease resistance proteins and involved in hypersensitive response (HR) (DeYoung and Innes [2006]). The further functional studies of the Os11g0615900 gene under both cold stress and biotic stress would help to define the QTL qCtss11, and help discern whether the Os11g0615900 gene functions in crosstalk between biotic and abiotic stress. Therefore, these genes would be good candidate genes for future comparative functional analyses in cold-sensitive and cold-tolerant populations.
QTL related to cold tolerance at the reproductive stage
Many QTL related to cold tolerance at the reproductive stage have been identified in recent years. Saito and Miura detected two QTL, Ctb1 and Ctb2, on chromosome 4 using a set of near-isogenic lines (NIL) derived from the backcross `Kirara397'/'Norin-PL8'/'Kirara397' (Saito et al. [2001]). The QTL Ctb1 for cold tolerance as assessed by spikelet fertility was fine mapped to a 56 kb region (Saito et al. [2004]). Moreover, Saito located Ctb1 within a 17 kb region and finally identified the first gene found to confer cold tolerance at the booting stage of rice. Based on a two-hybrid screen, Saito suggested that the F-box protein encoded by the Ctb1 gene functions as part of the E3 ubiquitin ligase complex (Saito et al. [2010]). Other QTL related to cold tolerance at the booting stage as assessed by spikelet fertility, qCTB2a and qCTB3, were mapped using 191 RIL derived from a cross between a temperate japonica rice variety, M-202, and a tropical indica variety, IR50 (Andaya and Mackill [2003b]). QTL for Dth (days to heading), cl (culm length), fer (spikelet fertility), pe (panicle neck exsertion), and dc (discoloration) were identified among RIL developed from a cross between the indica cultivar, `Milyang 23' and the japonica weedy rice, `Hapcheonaengmi 3' (Oh et al. [2004]). A set of NIL with cold tolerance at the booting stage have been developed by backcrossing the strongly cold-tolerant japonica landrace, `Kunmingxiaobaigu' (KMXBG) to the cold-sensitive japonica cultivar, `Towada', as the pollen recipient parent. Eight QTL based on variation in spikelet fertility, qCTB-1-1, qCTB-4-1, qCTB-4-2, qCTB-5-1, qCTB-5-2, qCTB-10-1, qCTB-10-2 and qCTB-11-1, were mapped in this population on chromosomes 1, 4, 5, 10, and 11, respectively. All of the alleles for cold tolerance were contributed by KMXBG, and the marker intervals containing these QTL were narrowed to 0.6-5.6 cM (Xu et al. [2008]). QTL analysis with simple sequence repeat (SSR) markers and composite interval mapping has identified three main-effect QTL assessed by spikelet fertility and growth during the reproductive stage, including qPSST-3, qPSST-7, and qPSST-9, respectively, on chromosomes 3, 7, and 9. In addition, a new source of cold-tolerance measured by spikelet fertility, line IR66160-121-4-4-2, was used as the pollen donor parent in a cross with a cold-sensitive cultivar, `Geumobyeo', to produce 153 F8 RIL for QTL analysis (Suh et al. [2010]). Another QTL for cold tolerance, qLTB3, was identified on the long arm of chromosome 3 from the cold-tolerant breeding line `Ukei 840' in F2 and BC1F2 populations from crosses between `Ukei 840' and `Hitomebore'. The cold tolerance of `Ukei 840' is derived from the Chinese cultivar `Lijiangxintuanheigu' (Shirasawa et al. [2012]). Cold tolerance related to qLTB3 in these lines was assessed by seed fertility.
Taken together, QTL for cold tolerance have been detected in rice at the seedling stage (Andaya and Mackill [2003a]; Zhang et al. [2005]), booting stage (Xu et al. [2008]), bud burst stage (Lin et al. [2010]), and plumule stage (Baruah et al. [2009]), suggesting that cold tolerance could be both developmentally regulated and growth-stage specific (Suh et al. [2012]; Foolad and Lin [2001]).
Conclusions
Systematic studies have been carried out to improve our understanding of the physiological and genetic basis of cold tolerance in rice, which will promote the development of rice cultivars with improved cold tolerance. Cold stress interferes with metabolism and initiates changes in various physiological properties of plants. In rice, these changes are part of the cold-sensing system that initiates the cold-responsive signaling network. Reduced chlorophyll content and fluorescence (Fv/Fm) triggered by cold stress indicate injury to photosynthesis, as increased ROS and MDA subsequently mediate cold damage and cold sensing. Soluble sugars, proline, and antioxidants then accumulate and protect rice plants from further damage. Moreover, measurements of these physiological properties can also be used to evaluate the extent of cold tolerance exhibited by various rice cultivars to identify genes controlling cold tolerance for breeding purposes.
Currently, there are many criteria for the evaluation of cold tolerance; but is there a "gold standard" among techniques for assessing cold tolerance of a specific rice cultivar? The appropriate technique may depend on a number of factors, so a strong correlation between the measured trait and the ultimately desired trait, whether survival or yield after cold stress, is very important. Although a decrease in fresh weight can indicate cold stress, water loss is not always an accurate indicator of cold stress, because plant variety, leaf area, and other stressors can also affect water content measurements. Visual evaluations of cold stress using the seedling growth scale are performed immediately after a 14 d cold treatment at 9°C, while those using leaf growth are performed after 7 d of recovery from 9°C cold stress. These techniques, particularly the latter, can help make fine distinctions in cold tolerance between varieties or individuals. Seedling survival rate (%) can be a more practical criterion for cold tolerance than germination-stage cold tolerance because seedlings are more likely to actually experience cold stress in the field. Seedling survival would also be useful for addressing the discrepancies between greenhouse- and field-based results. For sublethal cold stress, such as 4°C cold stress that merely slows growth but does not cause severe damage, new leaf emergence is a better choice for distinguishing the cold tolerance of lines stressed at 4°C. But an important disadvantage of all germination- and seedling-stage assays is that they do not necessarily predict reproductive-stage cold tolerance, or may do so in a variety-specific manner.
For selection of appropriate parents for cold tolerance breeding, evaluation of cold tolerance at the reproductive stage is important. And because it results in strong selection, the examination of spikelet fertility after CGC seems to be an ideal approach for improving cold tolerance relatively quickly. To identify cold-tolerance related QTL that operate at the reproductive stage, the examination of spikelet fertility after CDWI is particularly useful because it is conducted directly in the field. Mechanistic studies to evaluate metabolic parameters and enzyme activities can be valuable for researchers who are interested in the functional characterization of genes related to cold response, because changes in metabolism and enzyme activities are useful for revealing the role of genes that function in the cold-sensing and cold-response networks.
The development of molecular markers and linkage maps has allowed detection of many QTL related to cold tolerance at various growth stages. Most QTL have been identified in mapping populations derived from crosses between varieties or accessions derived fromO. japonica and O. indica, and between cultivated rice and its wild relatives. The future application of these QTL to MAS will significantly accelerate the breeding of cold-tolerant rice for temperate environments and high-altitude areas. However, the gap between greenhouse research and field application is a major concern in cold tolerance research. To resolve this problem, more researchers are evaluating cold tolerance using criteria that are conduct directly in the field, such as spikelet fertility after CDWI, to identify cold-tolerance related QTL. Moreover, Suh and Jena used both greenhouse and direct field methods to cross-validate their results (Suh et al. [2010]; Jena et al. [2012]), a positive trend for cold tolerance research in rice. Furthermore, in order to characterize and confirm the function of QTL identified in the greenhouse, Saito and Koseki conduct expression analysis or transgenic studies of candidate genes (Saito et al. [2010]; Koseki et al. [2010]). These approaches help to validate the function of these QTL in cold-tolerance and are necessary before initiating selection. In addition to these QTL and molecular markers, the mapping populations that have been established and used in mapping projects are valuable tools that are directly useful in cold tolerance breeding. In the future, the integration physiological mechanistic studies of cold tolerance and QTL identification will accelerate the improvement of rice for the traits related to cold tolerance.
Authors' contributions
QZ, QC, and SW performed the analysis, and prepared the figure and tables. QZ, YH, and ZW designed and wrote the paper. All authors read and approved the final manuscript.
Abbreviations
- QTL:
-
Quantitative trait loci
- ROS:
-
Reactive oxygen species
- MDA:
-
Malondialdehyde
- MAS:
-
Molecular marker-assisted selection
- EL:
-
Electrolyte leakage
- GC-MS:
-
Gas chromatography-mass spectrometry
- ABA:
-
Abscisic acid
- ABRE:
-
ABA-responsive elements
- ABF:
-
ABRE-binding bZIP transcription factor
- AsA:
-
Ascorbic acid
- GSH:
-
Glutathione
- POD:
-
Peroxidase
- SOD:
-
Superoxide dismutase
- CAT:
-
Catalase
- APX:
-
Ascorbate peroxidase
- CGC:
-
Cold greenhouse cultivation
- CDWI:
-
Cold deep-water irrigation
References
Andaya V, Mackill D: Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J Exp Bot 2003, 54(392):2579–2585. 10.1093/jxb/erg243
Andaya V, Mackill D: QTLs conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica × indica cross. Theor Appl Genet 2003, 106(6):1084–1090.
Andaya V, Tai T: Fine mapping of the qCTS12 locus, a major QTL for seedling cold tolerance in rice. Theor Appl Genet 2006, 113(3):467–475. 10.1007/s00122-006-0311-5
Andaya VC, Tai TH: Fine mapping of the qCTS4 locus associated with seedling cold tolerance in rice ( Oryza sativa L.). Mol Breed 2007, 20(4):349–358. 10.1007/s11032-007-9096-8
Apel K, Hirt H: Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 2004, 55: 373–399. 10.1146/annurev.arplant.55.031903.141701
Baruah AR, Ishigo-Oka N, Adachi M, Oguma Y, Tokizono Y, Onishi K, Sano Y: Cold tolerance at the early growth stage in wild and cultivated rice. Euphytica 2009, 165(3):459–470. 10.1007/s10681-008-9753-y
Bonnecarrère V, Borsani O, Díaz P, Capdevielle F, Blanco P, Monza J: Response to photoxidative stress induced by cold in japonica rice is genotype dependent. Plant Sci 2011, 180(5):726–732. 10.1016/j.plantsci.2011.01.023
Chinnusamy V, Zhu J, Zhu JK: Gene regulation during cold acclimation in plants. Physiol Plant 2006, 126(1):52–61. 10.1111/j.1399-3054.2006.00596.x
Chinnusamy V, Zhu J, Zhu JK: Cold stress regulation of gene expression in plants. Trends Plant Sci 2007, 12(10):444–451. 10.1016/j.tplants.2007.07.002
Cook D, Fowler S, Fiehn O, Thomashow MF: A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis . Proc Natl Acad Sci U S A 2004, 101(42):15243–15248. 10.1073/pnas.0406069101
DeYoung BJ, Innes RW: Plant NBS-LRR proteins in pathogen sensing and host defense. Nat Immunol 2006, 7(12):1243–1249. 10.1038/ni1410
Fairhurst T, Dobermann A: Rice in the global food supply. World 2002, 5(7,502):454. 349–511,675 349-511,675
Foolad M, Lin G: Genetic analysis of cold tolerance during vegetative growth in tomato, Lycopersicon esculentum Mill. Euphytica 2001, 122(1):105–111. 10.1023/A:1012616231637
Foolad M, Lin G, Chen F: Comparison of QTLs for seed germination under non-stress, cold stress and salt stress in tomato. Plant Breed 1999, 118(2):167–173. 10.1046/j.1439-0523.1999.118002167.x
Fujino K, Sekiguchi H, Matsuda Y, Sugimoto K, Ono K, Yano M: Molecular identification of a major quantitative trait locus, qLTG3–1 , controlling low-temperature germinability in rice. Proc Natl Acad Sci U S A 2008, 105(34):12623–12628. 10.1073/pnas.0805303105
Futsuhara Y, Toriyama K: Genetic studies on cool tolerance in rice. IV. Direct and indirect effects of selection and cold tolerance. Jpn J Breed 1969, 19: 286–292. 10.1270/jsbbs1951.19.286
Gothandam KM, Nalini E, Karthikeyan S, Shin JS: OsPRP3 , a flower specific proline-rich protein of rice, determines extracellular matrix structure of floral organs and its overexpression confers cold-tolerance. Plant Mol Biol 2010, 72(1):125–135. 10.1007/s11103-009-9557-z
Haboudane D, Miller JR, Tremblay N, Zarco-Tejada PJ, Dextraze L: Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens Environ 2002, 81(2):416–426. 10.1016/S0034-4257(02)00018-4
Han LZ, Zhang YY, Qiao YL, Cao GL, Zhang SY, Kim JH, Koh HJ: Genetic and QTL analysis for low-temperature vigor of germination in rice. Acta Genet Sin 2006, 33(11):998–1006. 10.1016/S0379-4172(06)60135-2
Hossain MA, Cho JI, Han M, Ahn CH, Jeon JS, An G, Park PB: The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J Plant Physiol 2010, 167(17):1512–1520. 10.1016/j.jplph.2010.05.008
Hossain MA, Lee Y, Cho J-I, Ahn C-H, Lee S-K, Jeon J-S, Kang H, Lee C-H, An G, Park PB: The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol Biol 2010, 72(4-5):557–566. 10.1007/s11103-009-9592-9
Huang J, Sun SJ, Xu DQ, Yang X, Bao YM, Wang ZF, Tang HJ, Zhang H: Increased tolerance of rice to cold, drought and oxidative stresses mediated by the overexpression of a gene that encodes the zinc finger protein ZFP245. Biochem Biophys Res Commun 2009, 389(3):556–561. 10.1016/j.bbrc.2009.09.032
Irri I: Standard evaluation system for rice. International Rice Research Institute, Philippine; 2002.
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M, Shinozaki K, Yamaguchi-Shinozaki K: Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol 2006, 47(1):141–153. 10.1093/pcp/pci230
Jena KK, Kim SM, Suh JP, Yang CI, Kim YJ: Identification of Cold-Tolerant Breeding Lines by Quantitative Trait Loci Associated with Cold Tolerance in Rice. Crop Sci 2012, 51(2):517–523. 10.2135/cropsci2010.12.0733
Kandpal RP, Rao NA: Alterations in the biosynthesis of proteins and nucleic acids in finger millet ( Eleucine coracana ) seedlings during water stress and the effect of proline on protein biosynthesis. Plant Sci 1985, 40(2):73–79. 10.1016/0168-9452(85)90044-5
Kanneganti V, Gupta AK: Overexpression of OsiSAP8 , a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol Biol 2008, 66(5):445–462. 10.1007/s11103-007-9284-2
Kim S-H, Choi H-S, Cho Y-C, Kim S-R: Cold-Responsive Regulation of a Flower-Preferential Class III Peroxidase Gene, OsPOX1 , in Rice ( Oryza sativa L.). J Plant Biol 2011, 55(2):123–131. 10.1007/s12374-011-9194-3
Kim S-I, Tai TH: Evaluation of seedling cold tolerance in rice cultivars: a comparison of visual ratings and quantitative indicators of physiological changes. Euphytica 2011, 178(3):437–447. 10.1007/s10681-010-0343-4
Kim SJ, Lee SC, Hong SK, An K, An G, Kim SR: Ectopic expression of a cold-responsive OsAsr1 cDNA gives enhanced cold tolerance in transgenic rice plants. Mol Cells 2009, 27(4):449–458. 10.1007/s10059-009-0055-6
Koseki M, Kitazawa N, Yonebayashi S, Maehara Y, Wang ZX, Minobe Y: Identification and fine mapping of a major quantitative trait locus originating from wild rice, controlling cold tolerance at the seedling stage. Mol Genet Genomics 2010, 284(1):45–54. 10.1007/s00438-010-0548-1
Krishnasamy V, Seshu D: Seed germination rate and associated characters in rice. Crop Sci 1989, 29(4):904–908. 10.2135/cropsci1989.0011183X002900040012x
Lee SC, Huh KW, An K, An G, Kim SR: Ectopic expression of a cold-inducible transcription factor, CBF1/DREB1b, in transgenic rice ( Oryza sativa L.). Mol Cells 2004, 18(1):107–114.
Lee SC, Kwon SY, Kim SR: Ectopic expression of a cold-responsive CuZn superoxide dismutase gene, SodCc1 , in transgenic rice ( Oryza sativa L.). J Plant Biol 2009, 52(2):154–160. 10.1007/s12374-009-9017-y
Lin J, Zhu W, Zhang Y, Zhu Z, Zhao L, Chen T, Zhao Q, Zhou L, Fang X, Wang Y: Detection of quantitative trait loci for cold tolerance at the bud bursting stage by using chromosome segment substitution lines in rice ( Oryza sativa ). Chin J Rice Sci 2010, 24: 233–236.
Liu K, Wang L, Xu Y, Chen N, Ma Q, Li F, Chong K: Overexpression of OsCOIN , a putative cold inducible zinc finger protein, increased tolerance to chilling, salt and drought, and enhanced proline level in rice. Planta 2007, 226(4):1007–1016. 10.1007/s00425-007-0548-5
Los DA, Murata N: Membrane fluidity and its roles in the perception of environmental signals. Biochimica et Biophysica Acta (BBA)-Biomembranes 2004, 1666(1):142–157. 10.1016/j.bbamem.2004.08.002
Lou Q, Chen L, Sun Z, Xing Y, Li J, Xu X, Mei H, Luo L: A major QTL associated with cold tolerance at seedling stage in rice ( Oryza sativa L.). Euphytica 2007, 158(1):87–94. 10.1007/s10681-007-9431-5
Ma Q, Dai X, Xu Y, Guo J, Liu Y, Chen N, Xiao J, Zhang D, Xu Z, Zhang X, Chong K: Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol 2009, 150(1):244–256. 10.1104/pp.108.133454
Maruyama K, Urano K, Yoshiwara K, Morishita Y, Sakurai N, Suzuki H, Kojima M, Sakakibara H, Shibata D, Saito K: Integrated analysis of the effects of cold and dehydration on rice metabolites, phytohormones, and gene transcripts. Plant Physiol 2014, 164(4):1759–1771. 10.1104/pp.113.231720
Matsunaga K: Establishment of an evaluation method for cold tolerance at the booting stage of rice using deep water irrigation system and development of highly cold-tolerant rice varieties by combining cold tolerance genes. Bull Miyagi Furukawa Agric Exp Stn 2005, 4: 1–78.
McFarlane J, Watson R, Theisen A, Jackson RD, Ehrler W, Pinter P Jr, Idso SB, Reginato R: Plant stress detection by remote measurement of fluorescence. Appl Opt 1980, 19(19):3287–3289. 10.1364/AO.19.003287
Mittal D, Madhyastha DA, Grover A: Genome-Wide Transcriptional Profiles during temperature and oxidative stress reveal coordinated expression Patterns and overlapping regulons in rice. PLoS One 2012, 7(7):e40899. 10.1371/journal.pone.0040899
Mittler R: Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 2002, 7(9):405–410. 10.1016/S1360-1385(02)02312-9
Morsy MR, Almutairi AM, Gibbons J, Yun SJ, De Los Reyes BG: The OsLti6 genes encoding low-molecular-weight membrane proteins are differentially expressed in rice cultivars with contrasting sensitivity to low temperature. Gene 2005, 344: 171–180. 10.1016/j.gene.2004.09.033
Morsy MR, Jouve L, Hausman JF, Hoffmann L, Stewart JMD: Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice ( Oryza sativa L.) genotypes contrasting in chilling tolerance. J Plant Physiol 2007, 164(2):157–167. 10.1016/j.jplph.2005.12.004
Murata N, Los DA: Membrane fluidity and temperature perception. Plant Physiol 1997, 115(3):875.
Nagao M, Minami A, Arakawa K, Fujikawa S, Takezawa D: Rapid degradation of starch in chloroplasts and concomitant accumulation of soluble sugars associated with ABA-induced freezing tolerance in the moss Physcomitrella patens. J Plant Physiol 2005, 162(2):169–180. 10.1016/j.jplph.2004.06.012
Nakashima K, Takasaki H, Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K: NAC transcription factors in plant abiotic stress responses. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2012, 1819(2):97–103. 10.1016/j.bbagrm.2011.10.005
Nakashima K, Tran LS, Van Nguyen D, Fujita M, Maruyama K, Todaka D, Ito Y, Hayashi N, Shinozaki K, Yamaguchi-Shinozaki K: Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J 2007, 51(4):617–630. 10.1111/j.1365-313X.2007.03168.x
Noctor G, Foyer CH: Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Biol 1998, 49(1):249–279. 10.1146/annurev.arplant.49.1.249
Oh CS, Choi YH, Lee SJ, Yoon DB, Moon HP, Ahn SN: Mapping of quantitative trait loci for cold tolerance in weedy rice. Breed Sci 2004, 54(4):373–380. 10.1270/jsbbs.54.373
Oliver SN, Van Dongen JT, Alfred SC, Mamun EA, Zhao X, Saini HS, Fernandes SF, Blanchard CL, Sutton BG, Geigenberger P: Cold-induced repression of the rice anther-specific cell wall invertase gene OSINV4 is correlated with sucrose accumulation and pollen sterility. Plant Cell Environ 2005, 28(12):1534–1551. 10.1111/j.1365-3040.2005.01390.x
Pamplona R: Advanced lipoxidation end-products. Chem-Biol Interact 2011, 192(1):14–20. 10.1016/j.cbi.2011.01.007
Saijo Y, Hata S, Kyozuka J, Shimamoto K, Izui K: Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J 2001, 23(3):319–327. 10.1046/j.1365-313x.2000.00787.x
Saito K, Hayano-Saito Y, Kuroki M, Sato Y: Map-based cloning of the rice cold tolerance gene Ctb1 . Plant Sci 2010, 179(1):97–102. 10.1016/j.plantsci.2010.04.004
Saito K, Hayano-Saito Y, Maruyama-Funatsuki W, Sato Y, Kato A: Physical mapping and putative candidate gene identification of a quantitative trait locus Ctb1 for cold tolerance at the booting stage of rice. Theor Appl Genet 2004, 109(3):515–522. 10.1007/s00122-004-1667-z
Saito K, Miura K, Nagano K, Hayano-Saito Y, Araki H, Kato A: Identification of two closely linked quantitative trait loci for cold tolerance on chromosome 4 of rice and their association with anther length. Theor Appl Genet 2001, 103(6):862–868. 10.1007/s001220100661
Sangwan V, Örvar BL, Beyerly J, Hirt H, Dhindsa RS: Opposite changes in membrane fluidity mimic cold and heat stress activation of distinct plant MAP kinase pathways. Plant J 2002, 31(5):629–638. 10.1046/j.1365-313X.2002.01384.x
Satake T, Hayase H: Male sterility caused by cooling treatment at the young micro-spore stage in rice plants. V. Estimations of pollen developmental stage and the most sensitive stage to coolness. Proc Crop Sci Soc Jpn 1970, 39(4):468–473. 10.1626/jcs.39.468
Sato Y, Masuta Y, Saito K, Murayama S, Ozawa K: Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene, OsAPXa . Plant Cell Rep 2011, 30(3):399–406. 10.1007/s00299-010-0985-7
Sato Y, Murakami T, Funatsuki H, Matsuba S, Saruyama H, Tanida M: Heat shock-mediated APX gene expression and protection against chilling injury in rice seedlings. J Exp Bot 2001, 52(354):145–151. 10.1093/jexbot/52.354.145
Schobert B, Tschesche H: Unusual solution properties of proline and its interaction with proteins. Biochimica et Biophysica Acta (BBA)-General Subjects 1978, 541(2):270–277. 10.1016/0304-4165(78)90400-2
Shah K, Dubey R: Effect of cadmium on proline accumulation and ribonuclease activity in rice seedlings: role of proline as a possible enzyme protectant. Biol Plant 1997, 40(1):121–130. 10.1023/A:1000956803911
Shao HB, Guo QJ, Chu LY, Zhao XN, Su ZL, Hu YC, Cheng JF: Understanding molecular mechanism of higher plant plasticity under abiotic stress. Colloids Surf B: Biointerfaces 2007, 54(1):37–45. 10.1016/j.colsurfb.2006.07.002
Sharifi P: Evaluation on sixty-eight rice germplasms in cold tolerance at germination stage. Rice Sci 2010, 17(1):77–81. 10.1016/S1672-6308(08)60107-9
Sharma P, Sharma N, Deswal R: The molecular biology of the low-temperature response in plants. BioEssays 2005, 27(10):1048–1059. 10.1002/bies.20307
Shelton AM, Zhao J-Z, Roush RT: Economic, ecological, food safety, and social consequences of the deployment of Bt transgenic plants. Annu Rev Entomol 2002, 47(1):845–881. 10.1146/annurev.ento.47.091201.145309
Shimono H, Ishii A, Kanda E, Suto M, Nagano K: Genotypic Variation in Rice Cold Tolerance Responses during Reproductive Growth as a Function of Water Temperature during Vegetative Growth All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher. Crop Sci 2011, 51(1):290–297. 10.2135/cropsci2010.05.0300
Shirasawa S, Endo T, Nakagomi K, Yamaguchi M, Nishio T: Delimitation of a QTL region controlling cold tolerance at booting stage of a cultivar, `Lijiangxintuanheigu', in rice, Oryza sativa L. Theor Appl Genet 2012, 124: 937–946. 10.1007/s00122-011-1758-6
Skopelitis DS, Paranychianakis NV, Paschalidis KA, Pliakonis ED, Delis ID, Yakoumakis DI, Kouvarakis A, Papadakis AK, Stephanou EG, Roubelakis-Angelakis KA: Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 2006, 18(10):2767–2781. 10.1105/tpc.105.038323
Song SY, Chen Y, Chen J, Dai XY, Zhang WH: Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234(2):331–345. 10.1007/s00425-011-1403-2
Suh J, Jeung J, Lee J, Choi Y, Yea J, Virk P, Mackill D, Jena K: Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerant genotypes of rice ( Oryza sativa L.). Theor Appl Genet 2010, 120(5):985–995. 10.1007/s00122-009-1226-8
Suh J, Lee C, Lee J, Kim J, Kim S, Cho Y, Park S, Shin J, Kim Y, Jena K: Identification of quantitative trait loci for seedling cold tolerance using RILs derived from a cross between japonica and tropical japonica rice cultivars. Euphytica 2012, 184(1):101–108. 10.1007/s10681-011-0575-y
Suzuki N, Mittler R: Reactive oxygen species and temperature stresses: a delicate balance between signaling and destruction. Physiol Plant 2006, 126(1):45–51. 10.1111/j.0031-9317.2005.00582.x
Takesawa T, Ito M, Kanzaki H, Kameya N, Nakamura I: Over-expression of ζ glutathione S-transferase in transgenic rice enhances germination and growth at low temperature. Mol Breed 2002, 9(2):93–101. 10.1023/A:1026718308155
Tian Y, Zhang H, Pan X, Chen X, Zhang Z, Lu X, Huang R: Overexpression of ethylene response factor TERF2 confers cold tolerance in rice seedlings. Transgenic Res 2011, 20(4):857–866. 10.1007/s11248-010-9463-9
Vannini C, Locatelli F, Bracale M, Magnani E, Marsoni M, Osnato M, Mattana M, Baldoni E, Coraggio I: Overexpression of the rice Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants. Plant J 2004, 37(1):115–127. 10.1046/j.1365-313X.2003.01938.x
Vaughan DA, Morishima H, Kadowaki K: Diversity in the Oryza genus. Curr Opin Plant Biol 2003, 6(2):139–146. 10.1016/S1369-5266(03)00009-8
Venekamp JH: Regulation of cytosol acidity in plants under conditions of drought. Physiol Plant 1989, 76(1):112–117. 10.1111/j.1399-3054.1989.tb05461.x
Xie G, Kato H, Imai R: Biochemical identification of the OsMKK6-OsMPK3 signalling pathway for chilling stress tolerance in rice. Biochem J 2012, 443(1):95–102. 10.1042/BJ20111792
Xie G, Kato H, Sasaki K, Imai R: A cold-induced thioredoxin h of rice, OsTrx23, negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro. FEBS Lett 2009, 583(17):2734–2738. 10.1016/j.febslet.2009.07.057
Xu LM, Zhou L, Zeng YW, Wang FM, Zhang HL, Shen SQ, Li ZC: Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice near-isogenic line. Plant Sci 2008, 174(3):340–347. 10.1016/j.plantsci.2007.12.003
Xu M, Li L, Fan Y, Wan J, Wang L: ZmCBF3 overexpression improves tolerance to abiotic stress in transgenic rice ( Oryza sativa ) without yield penalty. Plant Cell Rep 2011, 30(10):1949–1957. 10.1007/s00299-011-1103-1
Yamburenko MV, Zubo YO, Vanková R, Kusnetsov VV, Kulaeva ON, Börner T: Abscisic acid represses the transcription of chloroplast genes. J Exp Bot 2013, 64(14):4491–4502. 10.1093/jxb/ert258
Yang A, Dai X, Zhang WH: A R2R3-type MYB gene, OsMYB2 , is involved in salt, cold, and dehydration tolerance in rice. J Exp Bot 2012, 63(7):2541–2556. 10.1093/jxb/err431
Yuanyuan M, Yali Z, Jiang L, Hongbo S: Roles of plant soluble sugars and their responses to plant cold stress. Afr J Biotechnol 2010, 8(10):2004–2010.
Yun KY, Park MR, Mohanty B, Herath V, Xu F, Mauleon R, Wijaya E, Bajic VB, Bruskiewich R, de Los Reyes BG: Transcriptional regulatory network triggered by oxidative signals configures the early response mechanisms of japonica rice to chilling stress. BMC Plant Biol 2010, 10: 16. 10.1186/1471-2229-10-16
Zahedi H, Alahrnadi SMJ: Effects of drought stress on chlorophyll fluorescence parameters, chlorophyll content and grain yield of wheat cultivars. J Biol Sci 2007, 7(6):841–847. 10.3923/jbs.2007.841.847
Zhang J, Li J, Wang X, Chen J: OVP1 , a vacuolar H+-translocating inorganic pyrophosphatase (V-PPase), overexpression improved rice cold tolerance. Plant Physiol Biochem 2011, 49(1):33–38. 10.1016/j.plaphy.2010.09.014
Zhang Q, Jiang N, Wang G-L, Hong Y, Wang Z: Advances in understanding cold sensing and the cold-responsive network in rice. Adv Crop Sci Tech 2013, 1(1):104.
Zhang X, Guo X, Lei C, Cheng Z, Lin Q, Wang J, Wu F, Wang J, Wan J: Overexpression of SlCZFP1 , a novel TFIIIA-type zinc finger protein from tomato, confers enhanced cold tolerance in transgenic Arabidopsis and rice. Plant Mol Biol Report 2011, 29(1):185–196. 10.1007/s11105-010-0223-z
Zhang ZH, Qu XS, Wan S, Chen LH, Zhu YG: Comparison of QTL controlling seedling vigour under different temperature conditions using recombinant inbred lines in rice ( Oryza sativa ). Ann Bot 2005, 95(3):423–429. 10.1093/aob/mci039
Zhou L, Zeng Y, Hu G, Pan Y, Yang S, You A, Zhang H, Li J, Li Z: Characterization and identification of cold tolerant near-isogenic lines in rice. Breed Sci 2012, 62(2):196. 10.1270/jsbbs.62.196
Zhu J, Dong CH, Zhu JK: Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr Opin Plant Biol 2007, 10(3):290–295. 10.1016/j.pbi.2007.04.010
Acknowledgements
This work was supported, in part, by grants from the National Transgenic Project (2012ZX08009001), the National 973 Project (2012CB723000), the National 863 Project (2014AA10A604), the National Natural Science Foundation of China (31071674, 31371927), the State Key Laboratory Breeding Base for Zhejiang Sustainable Pest and Disease Control (2010DS700124-KF1210), and the Hunan Province University Innovation Platform Open Fund Project (10 K031). ZW was also supported by the Program for Innovative Research Teams in University (IRT1239), the Aid Program for Science and Technology Innovative Research Team in Higher Educational Institutions of Hunan Province, the Hunan Provincial Key Laboratory for Crop Germplasm Innovation and Utilization (11KFXM01), and Hunan Agricultural University (11YJ13).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Zhang, Q., Chen, Q., Wang, S. et al. Rice and cold stress: methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 7, 24 (2014). https://doi.org/10.1186/s12284-014-0024-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12284-014-0024-3