
Abstract
Cancer is a complex disease resulting from abnormal cell growth that is induced by a number of genetic and environmental factors. The tumor microenvironment (TME), which involves extracellular matrix, cancer-associated fibroblasts (CAF), tumor-infiltrating immune cells and angiogenesis, plays a critical role in tumor progression. Cyclic adenosine monophosphate (cAMP) is a second messenger that has pleiotropic effects on the TME. The downstream effectors of cAMP include cAMP-dependent protein kinase (PKA), exchange protein activated by cAMP (EPAC) and ion channels. While cAMP can activate PKA or EPAC and promote cancer cell growth, it can also inhibit cell proliferation and survival in context- and cancer type-dependent manner. Tumor-associated stromal cells, such as CAF and immune cells, can release cytokines and growth factors that either stimulate or inhibit cAMP production within the TME. Recent studies have shown that targeting cAMP signaling in the TME has therapeutic benefits in cancer. Small-molecule agents that inhibit adenylate cyclase and PKA have been shown to inhibit tumor growth. In addition, cAMP-elevating agents, such as forskolin, can not only induce cancer cell death, but also directly inhibit cell proliferation in some cancer types. In this review, we summarize current understanding of cAMP signaling in cancer biology and immunology and discuss the basis for its context-dependent dual role in oncogenesis. Understanding the precise mechanisms by which cAMP and the TME interact in cancer will be critical for the development of effective therapies. Future studies aimed at investigating the cAMP-cancer axis and its regulation in the TME may provide new insights into the underlying mechanisms of tumorigenesis and lead to the development of novel therapeutic strategies.
Similar content being viewed by others


Introduction
Cyclic adenosine monophosphate (cAMP) is a signaling messenger derived from the hydrosis of ATP by the transmembrane or soluble adenylate cyclase (AC). The downstream effectors of cAMP include cAMP-dependent protein kinase (PKA), exchange protein activated by cAMP (EPAC) and ion channels. The first cDNA encoding the catalytic subunits of PKA was isolated in 1986 [1]. Thereafter, more isoforms of the catalytic subunit of PKA were cloned. cAMP-responsive element-binding protein (CREB) is one of the classical effectors of cAMP-PKA/EPAC pathways. As a well-known second messenger, cAMP can regulate cell differentiation, proliferation, survival and migration. Thus, cAMP has pleiotropic effects on both physiological and pathophysiological processes, such as development, reproduction, angiogenesis, cell cycle progression, insulin secretion, energy metabolism, and pluripotent stem cell reprogramming. Besides, nuclear cAMP signaling is important for dopaminergic neurotransmission, learning and memory [2, 3].
Aberrant cAMP signaling may lead to many diseases such as cardiomyopathy, cognitive impairment, oncogenic virus pathogenicity, and autoimmune diseases including multiple sclerosis, rheumatoid arthritis, inflammatory bowel disease and systemic lupus erythematosus [4]. During the latent stage of Epstein-Barr virus (EBV) infection, the cAMP/PKA pathway promotes the stimulation of viral proteins expression by EBNA2, an EBV-encoded protein, and thereby enhances viral persistence and EBV-associated oncogenesis [5]. In addition, cAMP may stimulate HIV-1 transcription in latently infected monocytes/macrophages and reverse HIV latency [6]. However, some isoforms of cAMP responsive element modulator (CREM, also called inducible cAMP early repressor), one of the PKA targets, can inhibit HIV long terminal repeat promoter activity thereby suppressing HIV infection [7]. Moreover, cAMP can suppress the progression of autoimmune diseases by preventing the formation of neutrophil extracellular traps, the activation of NF-κB, the production of inflammatory cytokines such as tumor necrosis factor, IL1β and IL6, T cell activation, and autoantibody production [8,9,10].
Since cAMP signaling regulates cell proliferation and differentiation, abnormal cAMP biogenesis is involved in tumorigenesis, a complex disease resulting from diverse environmental and genetic factors [11]. cAMP has a significant impact on both cancer cells and the tumor microenvironment (TME). Cytokines, hormones and growth factors can modulate cAMP signaling by engaging the generation of cAMP from ATP and the transduction of signals by protein kinases. On the other hand, tumor-associated stromal cells, such as fibroblasts, endothelial cells and immune cells, can produce and release signaling molecules that either stimulate or inhibit cAMP synthesis. cAMP not only directly regulates cancer cell proliferation, survival and migration, but also promotes immune evasion by reprogramming the TME.
Notably, cAMP has paradoxical effects on tumorigenesis, depending on the cancer types, stages and other contexts. Thus, both the antagonists and agonists of cAMP signaling can be exploited for cancer treatment. Although the structure of PKA catalytic subunit has been solved more than 30 years ago [12], there are few PKA inhibitors available in the market. No PKA inhibitors have been approved in the clinic. However, phosphodiesterases (PDEs) inhibitors, which prevent cAMP or cGMP hydrolysis and thereby elevate cAMP/cGMP levels, are developed for the treatment of obstructive respiratory diseases, neuroinflammation, ischemia/reperfusion injury, atopic dermatitis, erectile dysfunction, depression and hypertension [13,14,15]. Preclinical studies demonstrate that PDE inhibitors may have tumor-suppressive effects, especially when combined with chemotherapeutic agents [16, 17]. The present review updates recent advances in cAMP signaling, and discusses the complex roles of cAMP signaling in the TME. Considering the implication of cAMP signaling in cancer biology, the potential to selectively target PKA, EPAC or PDEs, and the possibility of either elevating or reducing cAMP levels in the TME, targeting cAMP signaling pathways holds promise in cancer therapy.
Overview of the cAMP signaling pathways
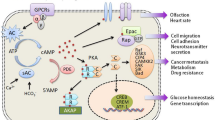
In mammals, there are nine adenylate cyclase genes (AC1-9) encoding transmembrane adenylate cyclases. These transmembrane adenylate cyclases have distinct physiological activities, which are generally regulated by heterotrimeric G proteins upon activation of G protein-coupled receptors (GPCRs) by extracellular hormones (adrenocorticotropic hormone, corticotropin-releasing hormone, follicle-stimulating hormone and thyroid stimulating hormone) [18], neurotransmitters (catecholamines, vasoactive intestinal peptide, glucagon-like peptide 1, gamma-aminobutyric acid, serotonin, etc.), cytokines (prostaglandin E2, netrin-1), and physiological agents such as adenosine and lactate [19, 20] (Fig. 1). Upon stimulation, G proteins dissociate into free Gα and Gβγ subunits. Gαs-coupled GPCRs stimulate adenylate cyclase activity and cAMP synthesis, while Gαi-coupled GPCRs inhibit type I, V and VI adenylate cyclase activation [21]. Except for the plasma membrane-initiated cAMP synthesis, the stimuli-induced internalization of GPCRs to endosomes and/or the trans-Golgi network leads to sustained generation of cAMP. On the other hand, cytosolic or nuclear translocated sAC (soluble adenylate cyclase) can be directly activated by HCO3− or Ca2+, and catalyze cAMP synthesis [22]. The transmembrane adenylate cyclases and sAC co-operatively or independently build cAMP pools in the cytosol, mitochondria and nucleus, allowing compartmentalized cAMP signaling [23]. cAMP signaling is transduced by at least five protein families, including PKA, EPAC, cyclic nucleotide-gated channels and hyperpolarization-activated cyclic nucleotide-gated channels (CNG/HCN), Popeye domain containing protein (POPDC), and cyclic nucleotide receptor involved in sperm function (CRIS). Both PKA and EPAC are widely expressed in many tissues and cancers.

The cAMP signaling pathways. The extracellular cAMP can be converted into AMP by plasma membrane-anchored ecto-PDE, followed by the conversion of AMP into adenosine by CD73. Ligands-engaged Gs- or Gi-coupled receptors may stimulate or inhibit cAMP synthesis, respectively. The ABCC transporter family members ABCC4/5/11 mediate the export of cAMP to extracellular space. cAMP engages PKA, EPAC, POPDC, HCN and CNG channels to regulate diverse signaling pathways and the transcription activities
The cAMP signal transducer PKA
The spatiotemporal propagation of cAMP is critical for appropriate response to various stimuli. PKA is one of the classical cAMP effectors. Both cAMP-synthesizing enzymes and the cAMP effector proteins are confined to intracellular nanodomains that contain distinct isoforms of adenylate cyclase, A kinase anchor proteins (AKAPs) and PDEs. Different AKAPs anchor PKA to distinct sites and thereby enable PKA to phosphorylate specific targets. On the other hand, AKAPs may either positively or negatively regulate adenylate cyclase activity, depending on the types of these enzymes [24]. AKAPs, PDEs and other regulators contribute to local cAMP gradients. PDEs comprise a family of enzymes that catalyze the degradation of cAMP and cGMP to AMP and GMP, respectively. Among the 11 families (PDE1–11) and over 100 estimated isoforms, PDE4/7/8 are specific to cAMP, while PDE5/6/9 selectively hydrolyze cGMP [25]. The other PDEs, including PDE1/2/3/10/11, can degrade both cAMP and cGMP [26,27,28]. The intracellular concentration and distribution of cAMP is determined by the balance between the activities of PDEs and adenylate cyclase.
cAMP not only elicits intracellular signaling transduction, but also releases into extracellular space and the bloodstream. The ABCC transporter family members ABCC4/5/11 mediate the export of cAMP to extracellular space [29]. In turn, the extracellular cAMP is converted into adenosine by ectonucleotide pyrophosphatase/phosphodiesterase 1 (ecto-PDE/ENPP1) and ectonucleotide 5′-nucleotidase (NT5E/CD73) [30, 31]. Finally, adenosine receptors mediate the paracrine action of cAMP in different tissues. While secreted cAMP promotes directional chemotaxis via cAMP receptors in bacteria such as Dictyostelium discoideum [32], it remains to know whether the plasma membrane cAMP receptors are present in mammalian cells.
As a cAMP-dependent protein kinase, PKA assembles as a tetramer consisting of two catalytic (C) subunits and two regulatory (R) subunits [25]. Upon four molecules of cAMP binding to PKA regulatory subunits, the tetramer dissociates into two free C subunits and a R subunit dimer, resulting in PKA activation. In addition, TGFβ-activated SMAD3/4 can interact with PKA regulatory subunits and promote PKA activation [33]. On the other hand, cAMP can enhance TGFβ signaling in breast cancer cells by inducing TGFβ receptor I expression [34]. Therefore, the cAMP and TGFβ signaling pathways are interconnected. Active PKA phosphorylates many proteins including protein kinases, phosphatases, transcription factors, receptor proteins, ion channels and cytoskeleton proteins (Fig. 2). PKA-induced phosphorylation of its targets either enhances or inhibits their activity. CREB is a family of transcription factors that comprises CREB1, CREM and activating transcription factor 1 (ATF-1) [35]. These transcription factors can bind to cAMP response element in the promoter of many genes, including c-Jun, c-Fos, cyclin D1, IL2, and IL10. Phosphorylation of CREB/ATF1 by PKA enhances their transcriptional activity. In contrast, phosphorylation of GSK3 and Raf1 by PKA leads to their inactivation [36, 37]. Furthermore, the mitochondrial AKAP1 and PKA can prevent mitochondrial dysfunction [38]. Since mitochondria are essential for tumor progression [39], the reliance of PKA to relieve mitochondrial stress in cancer warrants further studies.

Representative substrates of PKA. PKA can phosphorylate a number of proteins including kinases, phosphatases, transcription factors, receptors, ion channels, etc. Phosphorylation of these substrates by PKA may either enhance (red font) or inhibit (black font) their activities. The phosphorylation sites in these substrates are shown. *, The phosphorylation site is undefined
The specificity of PKA is largely determined by AKAPs, which deliver PKA to distinct signosomes. AKAPs are a family of functionally related proteins with diverse structure that comprise more than 60 members [40]. The PKA regulatory subunits interact with a 14–18 residue amphipathic helix in AKAPs through the docking and dimerization (D/D) domain. AKAPs serve as the scaffolding proteins to assemble signaling complexes containing other molecules such as phosphatases, kinase, adenylate cyclase and PDEs [41]. Tethering by AKAPs is a pivotal determinant in dictating which PKA substrates become phosphorylated in different cell types and contexts. Most cells express between 10 and 15 different AKAPs [42]. Seven AKAPs have been identified in lipid rafts in T cells and have been shown to contribute to the maintenance of T cell homeostasis [43].
Programmed cell death is an important biological process for physiological homeostasis, tissue renewal, and tumorigenesis. Many targets of PKA are involved in programmed cell death (Fig. 3). PKA phosphorylates and inactivates GSK3 and BAD, thereby inhibiting apoptosis [44]. However, cAMP-PKA also induces the proapoptotic B-cell lymphoma-family protein Bim and promotes the apoptosis of some cell types [45]. In addition, PKA suppresses IL1 maturation and thereby prevents gasdermin D (GSDMD) cleavage and pyroptosis [46]. Phosphorylation of poly(ADP-ribose) polymerase 1 (PARP1) by PKA leads to mitochondrial and nuclear PARP1 activation, thus impairing the function of mitochondria and inducing parthanatos [47]. Also, cAMP may inhibit NETosis, a type of neutrophil cell death associated with the release of neutrophil extracellular traps (NETs) [48]. Ferroptosis is another type of programmed cell death triggered by labile iron and lipid peroxidation. PKA had both pro-ferroptosis and anti-ferroptosis effects. Phosphorylation of CREB by PKA induces GPX4 expression, which antagonizes lipid peroxidation and then inhibits ferroptosis. On the other hand, cAMP-PKA signaling may induce ER stress, leading to autophagy-induced degradation of ferritin heavy chain 1, intracellular accumulation of Fe2+, and ferroptosis [49]. However, recent studies indicate that PKA may inhibit ER stress and ferroptosis as well [50]. Thus, the effects of cAMP and PKA on ferroptosis may be context-dependent.

The effects of cAMP signaling on programmed cell death. PKA can regulate apoptosis, pyroptosis, ferroptosis, parthanatos and NETosis. EPAC also regulates apoptosis, ferroptosis and parthanatos
The cAMP sensor EPAC
EPACs are the cAMP-regulated guanine nucleotide exchange factors that mediate numerous responses to cAMP [51]. Unlike PKA, EPAC proteins are single chain multi-domain polypeptides that contain both catalytic and regulatory elements. Both EPAC1 and EPAC2 consist of a C-terminal catalytic region and an auto-inhibitory N-terminal regulatory region. EPAC2, however, has an additional N-terminal cyclic nucleotide-binding domain [52]. The catalytic region consists of a Ras-exchange motif domain (REM), a Ras-association domain, and a cell division cycle 25 (CDC25) homology domain.
Binding of cAMP to the cAMP-binding domain in the regulatory region of EPAC results in conformational changes and the activation of Ras-related proteins Rap1 and Rap2. G protein-coupled receptor kinase 2 (GRK2) negatively regulates EPAC-mediated Rap1 activation by phosphorylating EPAC1 at Ser-108 [53]. The activation of PLCε by Rap1 in turn activates protein kinase C (PKC). Of note, both PKA and EPAC may mediate the cAMP‐induced activation of PKCε [54]. EPAC-PKC signaling promotes the phosphorylation of mitochondrial enzyme pyruvate dehydrogenase (PDHA1) and thereby enhances mitochondrial respiration [55]. Meanwhile, both PKA and EPAC-mediated activation of PKCε promote phospholamban phosphorylation at Ser-16, which contributes to calcium homeostasis, cardioprotection and germ cell differentiation [56]. In addition, activation of PKC by EPAC1-Rap1 signaling may sensitize TRPV1- and Piezo2-mediated mechanotransduction, which is involved in inflammatory pain and tumor progression [53, 57]. The stimulation of EPAC/Rap pathway by cAMP may enhance PI3K/Akt/mTOR and Raf/Ras/MAPK signaling, which are implicated in various physiological and pathophysiological processes such as cell growth, migration, adhesion and spreading [58, 59]. Except for the guanine nucleotide exchange factor activity of CDC25 homology domain, the REM domain of EPAC is able to induce JNK activation [60].
Similar to PKA, EPAC can regulates apoptosis, parthanatos and ferroptosis (Fig. 3). Activation of Rap1 and inhibition of the pro-apoptotic PUMA by EPAC suppresses apoptosis [61, 62]. In contrast, EPAC promotes ROS-mediated lipid peroxidation and ferroptosis [63]. EPAC also promotes PARP1 expression and inhibits PARP1 cleavage, thereby inducing parthanatos [64]. Thus, EPAC generally inhibits apoptosis, but paradoxically promotes parthanatos and ferroptosis. The effects of EPAC on cell death may be dependent on the stimuli.
Cyclic nucleotide-activated ion channels and POPDC proteins
Cyclic nucleotide-activated ion channels play a fundamental role in a variety of physiological processes. Hyperpolarization-activated cyclic nucleotide-gated channels (HCN) belong to the superfamily of voltage-gated pore loop channels. HCN channels regulate electrical activity in the excitable brain and heart cells [65]. Voltage-dependent opening of these channels is directly regulated by cAMP, contributing to spontaneous rhythmic activity in both brain and heart [65]. The unliganded cyclic nucleotide binding (CNB) fold in the C-terminal region of HCN channels and the transmembrane region co-operatively autoinhibit channel activity, while cAMP binding to the CNB fold relieves the autoinhibition [65, 66]. In addition, cyclic nucleotide-gated (CNG) channels are non-selective cation channels that play a crucial role in visual and olfactory signal transduction [67].
POPDC proteins are three-pass transmembrane proteins with high affinity to bind cAMP [68]. The Popeye domain of POPDC is supposed to undergo a conformational change upon cAMP binding. POPDC are important players in cardiac and skeletal muscle physiology. POPDC1 (also called BVES) serves as an adaptor for the interaction between adenylate cyclase 9 and TREK-1 potassium channels that control heart rate in a cAMP-dependent manner [69]. Similar to that PKA and EPAC exist in complexes containing PDEs, POPDC proteins are found in complexes with PDE4, which ensures an optimal cycle length of Ca2+ transients firing in sinoatrial nodes [70]. In addition, POPDC1 and POPDC3 expression is decreased in breast, brain, colon and gastric cancers [71,72,73]. The frequent silencing of POPDC in cancer may be attributed to promoter hypermethylation [74]. EGFR also downregulates POPDC1 expression in breast cancer [72]. The decrease in POPDC expression correlates with enhanced tumor progression and drug resistance, and poor patient survival, suggesting that POPDC is a tumor suppressor [68]. POPDC1 interacts with the tight junction protein ZO-1 and vesicle-associated membrane protein 3, thereby maintaining epithelial integrity and the recycling of transferrin, transferrin receptor, and integrin [75, 76]. Furthermore, POPDC1 negatively regulates RhoA activity and the expression of several oncogenes including c-myc, β-catenin and MMP2/9, thereby suppressing epithelial-mesenchymal transition (EMT) [77, 78]. Therefore, POPDC may be potential target for cancer therapy.
Contextual roles of cAMP signaling in cancer cells
Accumulating evidence demonstrates that cAMP signaling has paradoxical effects in different types of cancer (Table 1). While the targets of PKA include both oncogene and tumor suppressors, POPDCs are largely tumor suppressive. Therefore, the net effects of cAMP on tumor progression may depend on the balance between tumor-promoting and tumor-suppressing effects. In many types of cancer, cAMP signaling exhibits tumor-promoting properties [35]. The activation of cAMP-mediated PKA and EPAC signaling have been shown to promote tumor cell proliferation, survival, motility, adhesiveness and invasiveness in preclinical models of liver, lung, breast, brain and lymph cancers [35, 61]. PKA is deregulated in several cancers, especially in endocrine tumors such as adrenal tumors (Carney complex, Cushing’s syndrome), thyroid cancer, and growth hormone-secreting pituitary tumors [79,80,81,82,83]. Germline mutations in PRKAR1A cause Carney Complex, a disorder manifested as adrenal cortex hyperplasia/adenomas, cardiac and other myxomas, spotty skin pigmentation, and other abnormalities [84]. The majority of PRKAR1A mutations lead to decreased PRKAR1A mRNA due to nonsense-meditated mRNA decay, thereby reducing the levels of this regulatory subunit and increasing PKA activity [85]. Somatic mutations in PRKACA and PRKACB are also identified as driver mutations in adrenal cortical adenoma [86, 87]. In addition, mutations in PRKACA, PRKACB and PRKAR1A have been detected in liver, stomach, pancreas, lung and ovary cancers. The fusion of PRKACA and DNAJB1 is dominant and oncogenic in fibrolamellar hepatocellular carcinoma (FLC) [88], while the DNAJB1-PRKACA or ATP1B1-PRKACB fusion is also detected in pancreatobiliary neoplasms [89, 90].
As one of the targets of PKA, CREB promotes cancer cell proliferation, survival and migration, and negatively associates with cancer patients survival [136]. cAMP also induces CREB3L1 expression through PKA-p38MAPK pathway [137]. While CREB3L1 promotes the growth and metastasis of anaplastic thyroid carcinoma, it inhibits bladder cancer metastasis and glioblastoma growth [138,139,140]. Contradictory effect of CREB3L1 on triple negative breast cancer has been reported. One study shows that CREB3L1 promotes triple negative breast cancer metastasis [141], whereas another study demonstrates that CREB3L1 inhibits triple negative breast cancer metastasis [142]. The reason for such discrepancy is unclear. Given the versatile effects of diverse cAMP-regulated proteins on cancer, it is not surprising that cAMP may promote the progression of some types of human cancer but inhibit other cancer types. A more fascinating fact is that cAMP can even exert opposing effects on the same type of cancer in a context-specific manner. PKA is one of the dependencies in cancers such as fibrolamellar carcinoma and chondroma [143], while it is a bona-fide tumor suppressor in medulloblastoma.
Roles of cAMP in leukemia and lymphoma
cAMP promotes leukemia progression and inhibits anthracycline-, DNA damage- and ribosome biogenesis inhibitor-induced apoptosis in leukemia cells [91,92,93]. The PKA substrate CREB can promote hematopoietic cell proliferation and myeloproliferative disease [144, 145]. Overexpression of CREB and increased CREB phosphorylation are detected in the majority of acute myeloid leukemia patients, which is associated with poor prognosis in these patients [146]. Microsomal prostaglandin E synthase-1 also induces the expression of metadherin through the PGE2/EP3/cAMP/PKA/CREB pathway and thereby promotes T-cell acute lymphoblastic leukemia progression [147]. In addition, EPAC1/2 overexpression is detected in human acute myeloid leukemia. Inhibition of EPAC1/2 can suppress AML cell survival [93]. The cAMP‐EPAC1/2‐Rap1 survival pathway also contributes to ribosome‐targeting therapy resistance in patients with acute myeloid leukemia [93]. In addition, autophagy, a lysosome-dependent catabolic pathway, is involved in tumor progression and therapy [148]. cAMP signaling may promote PARP1-mediated autophagy and thereby inhibit DNA damage-induced B-cell precursor acute lymphoblastic leukemia cell death [149, 150].
However, cAMP signaling is required for the induction of gene expression by dexamethasone, a popular antileukemia agent [94, 151]. The expression of EP4, a PGE2 receptor, in T-cell acute lymphoblastic leukemia samples leads to an increase in intracellular cAMP levels, which sensitizes T-ALL to dexamethasone. In contrast, downregulation of cAMP synthesis and signaling confers dexamethasone resistance [94]. Moreover, combination of all-trans retinoic acid and arsenic trioxide can successfully treat patients with low-risk acute promyelocytic leukemia [152]. The induction of cAMP/PKA pathway contributes to retinoic acid-induced acute promyelocytic leukemia cell differentiation [153]. Downregulation of adenylate cyclase may reduce intracellular cAMP levels and suppress the induction of acute promyelocytic leukemia cell differentiation by all-trans retinoic acid [154, 155]. Stimulation of cAMP-PKA pathway by forskolin also sensitizes acute myeloid leukemia cells to GSKJ4, a H3K27me2/3 demethylases inhibitor [95].
In diffuse large B-cell lymphoma, cAMP inhibits cell growth in PKA- and EPAC-independent manner. The inhibition of PI3K/Akt pathway contributes to the effect of cAMP on diffuse large B cell lymphoma cell. PDE4B, a main hydrolyzer of cAMP in B cells, promotes diffuse large B-cell lymphoma cell survival and drug resistance by downregulation of cAMP signaling [96]. It is possible that the inhibition of PI3K/Akt by cAMP outweighs other potential tumor-promoting effects of cAMP in diffuse large B-cell lymphoma.
Roles of cAMP in solid tumors
cAMP signaling plays complex roles in various types of solid tumors. Here, we provide an update about the functions of cAMP in lung, liver, stomach, colon, breast, prostate and brain cancer, as well as melanoma. For the findings on cAMP signaling in ovarian cancer, cholangiocarcinoma and thyroid cancer, interested readers can refer to related reviews [82, 156, 157].
Lung cancer
Inactivating mutation of the tumor suppressor STK11/LKB1 is one of the genomic drivers of KRAS-mutated lung adenocarcinoma. LKB1 may inhibit the nuclear translocation of CREB-regulated transcription coactivators and thereby suppress constitutive activation of cAMP/CREB-mediated transcription [158]. Therefore, inactivating LKB1 mutation reinforces the cAMP-PKA-CREB signaling in lung cancer cells [158]. Reciprocally, PKA phosphorylates human and murine LKB1 at Ser-428 and Ser-431, respectively [159]. Phosphorylation of LKB1 at this residue suppresses the activation of AMPK and abrogates the tumor suppressive effects of LKB1 on melanoma [160], while it remains unclear whether the same is true for lung cancer. Of note, PKA can directly phosphorylate and inactivate AMPK, indicating that PKA may inhibit AMPK even in LKB1-mutated cancer [161]. In addition, CREB suppresses lipid peroxidation in lung adenocarcinoma cells by inducing GPX4 transcription and then inhibiting ferroptosis [97]. However, CREB1 phosphorylation contributes to cisplatin sensitivity in lung cancer cells via regulation of the ERK pathway, especially in cancer cells with mutated SET domain containing 2 (SETD2), a histone H3 lysine 36 (H3K36) trimethyltransferase [99]. Except for CREB, the cAMP/PKA-regulated phosphoprotein DARPP-32 enhances ERBB3/EGFR heterodimerization and promotes EGFR inhibitor resistance in EGFR-mutated lung adenocarcinoma [98]. DARPP-32 also promotes small cell lung cancer growth [162]. Moreover, AKAP1 expression correlates with high levels of Myc, phosphorylated mTOR, and reduced lung cancer patients survival [163].
Digestive system cancer
Overexpression of CREB1 and EPAC1 is also associated with gastric cancer progression [100, 101]. In addition, DARPP-32 activates IGF1R and STAT3 signaling in gastric cancer cells [102]. NF-κB activation and hypermethylation-mediated silencing of miR490-3p by H. pylori lead to DARPP-32 overexpression in gastric cancer cells thereby activating PI3K/AKT and STAT3 signaling pathways [103, 104]. Therefore, H. Pylori infection and cAMP signaling may cooperatively promote gastric carcinogenesis. Since STAT3 is a potent oncogene in various types of cancer [164], the PKA-DARPP32-STAT3 axis may contribute to tumorigenesis at sites other than the stomach.
cAMP signaling has complex roles in liver tumorigenesis. While DNAJB1-PRKACA fusion drives oncogenic pathways and induces FLC [105], the mixed-FLC/HCC tumors with inactivating mutations or translocations in the gene encoding BRCA1-associated protein-1 (BAP1) also harbor a chromosome gain of PRKACA and a loss of the inhibitory regulatory subunit PRKAR2A, thus exhibiting addiction to PKA activation [106]. Moreover, GNAS-activating mutation promotes liver tumorigenesis by upregulating cAMP/JAK/STAT3 signaling [107]. Dopamine or PGE2 secretion in the tumor microenvironment also engages dopamine receptor D1 or EP4 receptor, respectively, to activate cAMP/CREB pathway and promote HCC progression [165, 166]. CREB activation promotes HCC metastasis and drug resistance [108,109,110]. However, cAMP/PKA mediates the tumor suppression effects of Exenatide (Ex-4), an antidiabetic drug targeting glucagon-like peptide-1 receptor, on HCC in obese diethylnitrosamine-treated mice [111]. Treatment of hepatoma cells with PDE4 inhibitors increases intracellular cAMP levels but suppresses cell cycle progression and survival [112]. PDE4D depletion also inhibits the expression of cancer-related genes such as IGF2 and the progression of cell cycle in HCC cells [113]. Together, these studies indicate that cAMP has opposing effects on HCC.
The cAMP-PKA-CREB pathway is frequently activated in colorectal cancer. Adenylate cyclase overexpression or PDE4B silencing may increase cAMP levels in colon cancer [167, 168]. In addition, the fermentation ofdietary fiber can generate short-chain fatty acids, which activate free fatty acid receptor 2 (FFAR2/GPR43) and reshape gut microbiota, leading to increased expression of inflammation suppressors and prevention of intestinal carcinogenesis [169]. In contrast, GPR43 deficiency results in enhanced cAMP-PKA-CREB signaling and HDAC expression, thereby promoting colon carcinogenesis by inhibiting the expression of inflammation suppressors and enhancing neutrophil infiltration into colon cancer [114]. Moreover, norepinephrine induces CREB phosphorylation and then promotes human colon cancer cells growth and invasion [115]. CREB crosstalks with KRAS to promote colon carcinogenesis and positively regulates ALCAM (CD166) and PROM1 (CD133) expression, thus promoting colorectal cancer stemness and metastasis [170, 171].
Breast cancer
What roles does cAMP play in breast cancer? About 60% of human breast cancer are estrogen receptor-positive. PKA directly phosphorylates estrogen receptor α, leading to ligand-independent estrogen receptor activation and tamoxifen resistance in estrogen receptor-positive breast cancer [116]. In addition, cAMP/PKA/CREB may act downstream of JAK/STAT3 to promote chemotherapeutic resistance in inflammatory breast carcinoma [117]. However, reduced cAMP levels contribute to the promotion of estrogen receptor inhibitor, HER2 inhibitor and chemotherapeutic resistance by PAQR8 in breast cancer [118]. Inhibition of PDE4A in breast cancer stem cells increases cAMP levels and PKA activity, which upregulates PTEN and induces cell cycle arrest [119]. The PDE4 inhibitor rolipram synergizes with paclitaxel to inhibit breast cancer cell growth [119]. Meanwhile, PKA-mediated ERK1/2 inhibition may enhance the sensitivity of triple negative breast cancer cells to doxorubicin [121]. cAMP/PKA also inhibits NF-κB signaling in breast cancer thereby suppressing the expression of OCT4, a stem cell marker [122]. It appears that cAMP and PKA have tumor-promoting or tumor-suppressing effects in breast cancer. Given that PTEN deficiency is common in breast cancer, PTEN status may shift the balance between the pro-tumor and anticancer effects of cAMP.
One study suggests that PKA may promote the phosphorylation and nuclear translocation of p53 in breast cancer cells during IL24 treatment [120]. While p53 upregulates cAMP levels in breast cancer cells, it prevents cAMP accumulation in osteoblasts by inhibiting parathyroid hormone-related protein [172]. Therefore, the effects of PKA on p53 are inconclusive or context-dependent. In contrast to the stimulatory effect of PKA on p53 phosphorylation and nuclear translocation in breast cancer cells [120], ADRB1-mediated promotion of cAMP synthesis in BRCA1-deficient ovarian cancer cells suppresses p53 accumulation and DNA damage-induced apoptosis [123]. Similar effect of cAMP/PKA on p53 accumulation was detected in acute lymphoblastic leukemia [173, 174]. In addition, the sympathetic nervous system mediator norepinephrine activates ADRB2 and induces cAMP synthesis, leading to CREB-mediated DUSP1 expression, which dephosphorylates JNK and thereby inhibits c-Jun phosphorylation and ovarian cancer cells apoptosis [124]. Of note, DUSPs may promote the resistance of cancer cells to multiple cancer therapeutic approaches including chemotherapy, radiation and molecular targeted therapy [175]. Except for DUSP1, other cAMP-responsive genes may be involved in cancer therapy. In contrast to DUSP, mitochondrial ferritin is a CREB-regulated gene that can enhance cisplatin sensitivity in ovarian cancer cells [176]. By demethylating m6A at the 3'UTR of phosphodiesterases PDE1C and PDE4B, FTO inhibits PDE1C and PDE4B expression and thereby enhances cAMP signaling, which suppresses ovarian cancer cells stemness [177].
Melanoma
The effects of cAMP signaling are even more complex in melanoma. cAMP stimulates primary melanoma growth through EPAC1/2-Rap1/mTORC1 pathways, while it does not promote metastatic melanoma cells growth [125,126,127]. Adenylate cyclase inhibitor impairs cAMP signaling and suppresses melanoma growth, while low levels of CREB phosphorylation correlates with melanoma metastasis and recurrence [126]. miR-23a-3p inhibits adenylate cyclase 1 expression and then reduces cAMP synthesis, leading to suppression of mucosal melanoma growth [178]. It is unclear whether the paradoxical effect of CREB on melanoma aggressiveness involves MITF, a CREB-responsive gene that inhibits melanoma invasiveness [179]. In addition, activation of the cAMP-PKA-CREB/CRTC pathway promotes the resistance of melanoma to BRAF(V600E) inhibitor vemurafenib [128]. PKA activity is negatively regulated by PKARIIβ [180, 181]. Both the autophosphorylation site (Ser-116) and the nuclear location signal KKRK are important for the inhibition of PKA by PKARIIβ [180]. Pharmacological activation of PKARII subunits can inhibit proliferation and increased caspase-3 activity in melanoma cells [182].
Prostate cancer
PKARIIβ overexpression also leads to prostate cancer cell growth inhibition, whereas overexpression of PKARIA stimulates cell growth [181]. PKA phosphorylates the Thr-89 residue in HSP90 and thereby dissociates HSP90-androgen receptor (AR) complex [129]. In turn, HSP27 binds to the released AR and delivers it into the nucleus [129]. AR-V7, a constitutively active AR variant, induces arginine vasopressin receptor 1a expression in prostate cancer cells, which promotes CREB activation and castration resistance [130]. Treatment of prostate cancer cells with abiraterone acetate increases intracellular cAMP levels and PKA activity, leading to CREB1 phosphorylation that promotes abiraterone acetate resistance [131]. Enzalutamide (MDV3100) treatment also enhances CREB activation in AR-positive prostate cancer cells [132]. CREB can enhance the activity of histone methyltransferase EZH2, which represses the anti-angiogenic factor thrombospondin-1 and promotes neuroendocrine differentiation [132]. Sympathetic activation of cAMP-PKA pathway promotes focal adhesion kinase activation and prostate cancer metastasis [133]. Meanwhile, CREB5 promotes enzalutamide resistance in prostate cancer. In AR-positive prostate cancer cells, CREB5 interacts with and enhances FOXA1 and AR activity thereby regulating a subset of targets such as MYC and genes related to cell cycle, Wnt signaling and EMT [183, 184].
Brain tumor
While mutant GNAS drives pancreatic tumorigenesis by promoting PKA-mediated suppression of SIK [185], GNAS acts as a tumor suppressor in neuroblastoma, medulloblastoma and basal cell carcinoma of skin [134, 135, 186]. The development of medulloblastoma and basal cell carcinoma is largely driven by the Sonic Hedgehog (SHH) and Hippo pathways. PKA negatively regulates SHH effectors Smoothened (SMO)-Gli by phosphorylation [187]. Conversely, a PKA-inhibitor motif within SMO physically blocks the active site of PKA catalytic subunit and thereby antagonizes the inhibition of Gli by PKA [188].
In contrast to the tumor suppressive effects of cAMP-PKA signaling in medulloblastoma, the PKA-Dock180 axis mediates the promotion of glioblastoma development and invasion [189]. PKA also phosphorylates glutathione S-transferase P1 (GSTP1), an enzyme for carcinogen and drug metabolism, and c-Jun NH2-terminal kinase signaling [190]. Phosphorylation of GSTP1 at Ser-184 enhances its enzymatic activity and thereby promotes drug resistance in glioblastoma cells [190]. In addition, PGE2-induced cAMP signaling promotes glioblastoma growth, angiogenesis, metastasis and immune evasion [191]. Thus, the cAMP-PKA pathway may have opposing roles in different types of brain tumor.
cAMP signaling in the tumor microenvironment
The microenvironment of solid tumors is characterized by local hypoxia and higher levels of lactate production. In a panel of carcinoma cell lines of various origin, hypoxia induces adenylate cyclase VI and VII expression through HIF1, which leads to the elevation of cAMP levels and stimulation of cell migration and invasion [192, 193]. Moreover, hypoxia and HIF1α activate PKA by repressing PKARIIβ expression in growth hormone-secreting pituitary tumors [83]. Studies have shown that extracellular lactate can decrease cAMP levels and subsequent PKA activation by binding and activating GPR81 [194, 195]. In murine melanoma cells, lactate increases tumor malignancy by facilitating small extracellular vesicles production via GPR81-cAMP-PKA axis [195]. In the preclinical model of colon cancer, lactate-GPR81-cAMP-PKA signaling promotes chemotherapy resistance through regulating the mismatch repair system [196].
Hypoxia, lactate synthesis and secretion, and overexpression of membrane-bound channels/transporters such as Na(+)/H(+) exchanger 1 and bicarbonate symporters in tumor cells may lead to acidic extracellular pH usually ranging from 5.9 to 7.0 [197, 198]. Proton-sensitive GPCRs including GPR4 (GPR19), GPR65, GPR68 and GPR132 are activated by acidic pH via protonation of different histidine residues and other potential acidic residues in these receptors [199, 200]. GPR4 is overexpressed in squamous cell carcinoma of the head and neck, colorectal cancer, breast, ovarian, colon, liver and kidney tumors, Merkel cell carcinoma and melanoma [201,202,203,204]. Acidification of the TME is a well-known promoter of tumor progression and immune evasion [205]. Acidic pH stimulates proton-sensitive GPCRs coupling to the Gs protein and thereby induces adenylate cyclase activation and cAMP accumulation [206]. The migration of GPR4 overexpressing melanoma cells is enhanced in an acidic microenvironment [206]. Overexpression of GPR4 in squamous cell carcinoma of the head and neck also induces angiogenesis via IL6, IL8 and VEGFA secretion at acidic extracellular pH [201]. In addition, TGAG8 helps cancer cells adapt to the acidic microenvironment and enforces tumor progression by promoting PKA and ERK activation [207]. Previous studies have demonstrated that TDAG8 and GPR68 are involved in acidic pH-induced expression of programmed cell death protein 1 (PD-L1), an immune checkpoint molecule, in squamous cell carcinoma and melanoma cells [208, 209], which may contribute to immune escape. However, the induction of PD-L1 expression by acidity may be not mediated by cAMP [208].
Cell stress, cell death, and activation of pannexin/connexin channels on endothelial cells and immune cells may lead to the release of ATP into the extracellular space. Extracellular ATP is rapidly degraded to adenosine through two cell-surface ectonucleotidases, CD39 and CD73. Specifically, ATP is converted to ADP and/or AMP by CD39, followed by CD73-mediated dephosphorylation of AMP to adenosine [210]. There are four subtypes of plasma membrane adenosine receptor, including A1R, A2aR, A2bR, and A3R. Similar to PGE2 receptor, adenosine receptor couples to specific G protein and activates multiple signaling pathways. Both A2aR and A2bR are coupled to Gαs protein and can induce cAMP synthesis [211], while A1R and A3R inhibit transmembrane adenylate cyclase via Gαi protein, leading to a decrease in cAMP levels.
Accumulating evidence indicates that the neuronal circuits are involved in tumor progression. The infiltration and sprouting of nerve fibers into the tumor microenvironment is a driver of tumor growth and metastasis in various cancer types including prostate, gastric, skin, pancreatic and breast cancer [212,213,214,215,216,217,218]. Overexpression of neurotrophic growth factors, such as NGF and brain-derived neurotrophic factor (BDNF), in tumor and stromal cells promotes cancer innervation [213, 215]. Endoplasmic reticulum stress in tumor cells may trigger the expression and release of BDNF into the TME [219]. Given that hypoxia, nutrient shortage and anticancer agents can induce endoplasmic reticulum stress, BDNF expression and release may be induced by diverse stimuli. The sympathetic or parasympathetic nerve fibers in tumor tissue may release neurotransmitters that bind to their receptors in stromal and cancer cells. Catecholamines activates β-adrenergic receptors on tumor or stromal cells and induces cAMP accumulation. In turn, the sympathetic-cAMP-PKA/EPAC signaling pathway promotes tumor growth, invasion and metastasis [133]. β-Adrenergic receptor activation also increases intracellular Ca2+, which feedforwards to promote cAMP synthesis [220]. On the other hand, the cAMP/EPAC/JNK signaling pathway promotes BDNF expression in tumor cells, which feedforwards to enhance tumor innervation [221]. Except for stimulation of neurogenesis, BDNF can induce tumor and endothelial cell migration [222]. Collectively, the interplay among neurotransmitters, cAMP and neurotropic growth factors promotes tumor angiogenesis, growth and metastasis [223]. Except for the hypoxic and acidic microenvironment, and the neural niche, tumor stromal cells such as fibroblasts, macrophages and other immune cells are involved in cAMP signaling.
The roles of cAMP signaling incancer-associated fibroblasts (CAFs)
cAMP/PKA signaling plays an important role in fibroblast cell growth and migration. Inhibition of PKA blocks fibroblast migration in response to serum or PDGF/EGF [224]. Overexpression of RAMP1, a calcitonin gene-related peptide receptor component, can promote mouse skin fibroblast proliferation via Gαi3-PKA-CREB-YAP axis [225]. In human lung fibroblasts, PDE inhibition exerts anti-fibrotic effects via activation of cAMP/PKA signaling and inhibition of TGFβ [226]. Another study reported that PDE4 inhibitors rolipram and roflumilast can antagonize the profibrotic activity of TGFβ1 [227]. Thus, cAMP/PKA signaling is a negative regulator of fibrosis.
Many types of tumor, such as breast, gastric, ovarian, colon, and renal cancer, grow in the anatomical vicinity of adipose tissue. Adipocytes, preadipocytes and adipose tissue support tumorigenesis and metastasis via secreting cytokines and adipokines [228,229,230]. PKA and EPAC both contribute to the differentiation of 3T3-L1 fibroblast to adipocyte [231]. Moreover, cAMP/PKA/CREB signaling stimulates the adipogenesis of 3T3-L1 by suppressing Dlx5 transcription through upregulated C/EBPβ [232]. These data indicate that cAMP promotes fibroblast differentiation and adipogenesis.
CAFs are abundant components of the TME that have important roles in tumor progression [233]. How does cAMP signaling affect CAF? In breast cancer, estrogen induces FOSL2/Wnt5a expression via cAMP/PKA signaling, in turn, the secreted Wnt5a regulates FZD5/NK-kB/ERK signaling in vascular endothelial cells to promote tumor angiogenesis [234]. Moreover, breast tumor cells could activate ER/GPER/cAMP/PKA/CREB axis in CAFs and trigger the aerobic glycolysis, leading to multiple drug resistance [235]. In gastrointestinal cancers, the expression of GPR68, a proton-sensing GPCR, in CAFs is upregulated by cancer cells [236]. The acidic TME promotes GPR68 activation, cAMP synthesis and PKA activation, leading to increased IL6 expression in CAFs [236]. While CAFs-secreted IL6 appears to reciprocally stimulate tumor cell proliferation [236], sustained IL6 secretion also promotes chronic inflammation and immune evasion [237]. Thus, cAMP-mediated crosstalk between tumor cells and CAFs may promote tumor progression through multiple pathways.
The roles of cAMP signaling in immune cells
The immune microenvironment is tightly involved in tumor progression and cancer therapy [238]. Natural killer (NK) cell, macrophage, CD8+ effector T cell, CD4+ T cell, dendritic cell, T regulator cell, myeloid-derived suppressor cell and B lymphocyte are the building blocks of highly heterogenous tumor immune microenvironment [239]. cAMP has pleiotropic effects on immune cells fate, and may affect immune response (Fig. 4).

cAMP signaling in immune cells. The GPCR-mediated cAMP signaling is involved in regulating the differentiation, exhaustion and cytotoxic activities of multiple types of immune cells. Both PKA and EPAC mediate the diverse effects of cAMP on immune cells
Regulatory T cells (Tregs)
Many studies have demonstrated the overexpression of CD39, CD73, A2aR, and A2bR in both tumor cells and immune cells, which contributes to extracellular adenosine accumulation, intracellular cAMP production and metabolic reprogramming [240,241,242,243]. Besides, the contribution of regulatory T cells (Tregs) to adenosine and PGE2 production plays important roles in the TME. Tregs are a small subset of CD4+ T cells which interact with effector T cells and suppress their functions [244]. There are two Tregs subtypes, natural Tregs (nTregs) and inducible Tregs (iTregs). iTregs expands and accumulates in tumor tissues and peripheral blood of cancer patients [244]. iTregs expressing both CD39 and CD73 is able to hydrolyze ATP to adenosine. Tr1, a subset of iTregs, is significantly more induced in COX-2-overexpressed tumors than COX-2-negative tumors, while Tr1 cells themselves are COX-2-positive and able to produce and secrete PGE2 [245, 246].
Tregs play a central role in the maintenance of self-tolerance and homeostasis through suppressing aberrant immune response against self-antigens, and suppress anti-tumor immune response [244]. The infiltration of Tregs into tumor tissues is associated with poor prognosis in cancer patients [247, 248]. cAMP is crucial for Tregs-mediated immune regulation. The levels of cAMP in Tregs are regulated by the expression and activation of ACs and PDEs. IL2 induces AC7 activation and cAMP accumulation in Tregs [249]. FOXP3, a master regulator of the regulatory pathway in the development and function of Tregs, downregulates miR-142-3p to elevate AC9 expression, resulting in increased cAMP production [250]. Moreover, FOXP3 and miR-142-5p, the predominant miR-142 isoform in Tregs, work in concert to repress PDE3B expression and cAMP degradation in Tregs [250, 251]. Furthermore, PDE4B2 and PDE8A expression are significantly reduced in Tregs as compared with effector T cells [252]. On the other hand, TCR-induced FOXP3 expression in Tregs is controlled by CREB/ATF activation [253]. This FOXP3-cAMP-CREB loop enables Tregs to maintain high levels of cAMP.
The ability of Tregs to generate and accumulate high levels of cAMP enables them to transfer it through gap junction into conventional T cells and dendritic cells [254]. Connexin proteins (Cx)-mediated Gap junction is specialized intercellular channels between two adjacent cells. cAMP induces the expression of Cx43, Cx46, Cx31.1, Cx32 and Cx45 in T cells including Tregs [255]. An alternative mechanism to increase cAMP in the target cells involves the conversion of ATP into adenosine by CD39 and CD73 on the surface of Tregs. Tumor-resident FOXP3+ Tregs can decompose ATP to adenosine, which inhibits CD8+ T cells proliferation and survival through the A2aR-cAMP pathway [256]. In addition, cAMP signaling in Tregs regulates the expression of other functionally important molecules. cAMP-elevating agents or β2-adrenergic receptor signaling induces the expression of cytotoxic T lymphocyte antigen-4 (CTLA-4), an immune checkpoint molecule, in a PKA-dependent manner [257, 258].
Conventional T cells (Tcons)
Tcons include naïve T cells, effector T cells (Teffs), and memory T cells. cAMP plays complex roles in T cell activation, proliferation, and production of cytokines. Adenosine engages A2aR on T cells and thereby activates adenylate cyclase to induce cAMP synthesis [259]. cAMP-PKA activation in Tcons results in the translocation of CREB and CREM/ICER into the nucleus. While CREB activation may promote T cell proliferation and function, such as the production of IFN-γ [260], cAMP-induced expression and nuclear translocation of the transcriptional inhibitor ICER inhibits NF-κB-, AP1-, and NFAT-driven transcription, which in turn impedes the transcription of important genes for immune activation, such as CD25 and IL2, but increases the levels of CTLA-4 [261,262,263,264]. In addition, PKA indirectly suppresses Lck and Fyn by phosphorylating Csk at Ser-364, which in turn phosphorylates Lck at Tyr-505 and Fyn at Tyr-528 [265]. The inhibition of Lck/Fyn by Csk prevents zeta-chain-associated protein kinase 70 (ZAP70) phosphorylation, leading to inhibition of TCR signaling [266]. On the other hand, TCR activation by anti-CD3/CD28 stimulation can increase PDE2A and PDE4A/B/D expression in Tcons [267, 268]. Furthermore, evidence suggests that the existence of PKA-independent pathway of cAMP-mediated Tcons suppression. For example, EPAC1 negatively regulates IL2 production and Teffs proliferation in a Rap1-, STAT3- and TGFβ-dependent manner [252, 269]. Selectively activating EPAC1 through 8-pCPT-2’-O-Me-cAMP in mice suppresses CD4+CD25− Teffs proliferation [252].
Dendritic cells (DCs)
DCs are myeloid cells that have either pro-tumor or anticancer effects [270]. PDE4B is a homeostatic regulator of cAMP in DCs. The dynamic expression of PDE4B is dependent on cAMP/PKA [271]. As the main antigen-presenting cells, DCs are the primary targets of Tregs suppression. The DC-Tregs interaction results in reduced T cell stimulatory capacity and secretion of inhibitory cytokines in DCs. Adenosine induces DCs to migrate toward Tregs through cAMP-PKA-EPAC-Rap1 pathway and then attracts DCs away from CD4+ T cells [272, 273]. Furthermore, adenosine-cAMP-PKA/EPAC signaling increases NF-κB expression and immunosuppressive IL10 production, and inhibits IL12p40 production in human monocyte-derived DCs, thereby fully differentiating DCs into a suppressive phenotype [274]. Treatment of DCs with cAMP elevating agent forskolin or co-culture with Tregs leads to a rapid downregulation of co-stimulatory molecules (CD86 and CD80) and upregulation of the inhibitory molecule B7-H3 [275, 276]. Thus, cAMP may reprogram DCs to acquire immune suppressive properties.
The transcriptional activator CREB is a critical regulator of DCs function in immune response. The deletion of CREB in CD11c+ cells results in reduced germinal center responses [277]. Heme oxygenase 1 regulates DCs maturation and antigen presentation by modulating p38MAPK-CREB/ATF1 signaling [278]. Independent of cAMP stimulation, DCs also produce IL10 through innate immune receptors via CREB. C-type lectin MGL enhances IL10 and TNFα secretion by DCs via activating ERK-p90RSK-CREB axis [279]. The calcium-dependent CaMKIV-CREB-Bcl2 axis plays an important role in the activation of DCs by TLR4 stimulation [280].
In semi-mature DCs, cAMP upregulates CTLA-2 expression and enhances TGFβ-dependent FOXP3+ iTreg conversion [281]. Low levels of cAMP in conventional type-2 dendritic cells (cDC2s) promote Th2 differentiation, while increased levels of cAMP reprogram cDC2s from a pro-Th2 to a pro-Th17 phenotype via repression of IRF4 and KLF4 by the PKA-CREB signaling [282,283,284]. Accordingly, PDE4 inhibitor augments the Th17-promoting capability of DCs by enhancing IL23 production [285]. Tumor cell-secreted PGE2 induces IL23 secretion in DCs via EP2/4-cAMP signaling, leading to Th17 cell expansion [286]. Both PKA-induced phosphorylation of CREB and EPAC-induced phosphorylation of C/EBPβ mediate the stimulatory effect of PGE2 on IL23 expression [286]. Th17 cells and Th17-related cytokines have either pro-tumor or anti-tumor roles depending on the cancer type [287, 288]. The exact effects of cAMP-induced Th17-promoting capability of DCs on tumor immune surveillance may be contextual and await further studies.
B cells
Except for T cell-mediated cellular immunity, cAMP is also involved in the B cell-mediated humoral immunity. Previous studies have demonstrated that cAMP inhibits B-cell maturation, the activation of quiescent B cells, and subsequent IgG1 and IgE production [289, 290]. In human CD10+ B-precursor cells, cAMP elevating agents induce a decline in Mcl1 expression and subsequent apoptosis [291]. Adenosine and cAMP block the B cell antigen receptor-mediated NF-κB activation via PKA [292]. Thus, it appears that cAMP/PKA signaling is detrimental for B-cell maturation and survival. However, CREB is critical for B-cell survival and function. To avoid the detrimental effects of cAMP and PKA on B cells, mature B cells take advantage of PKCδ and p90RSK rather than PKA to activate CREB, which contributes to the regulation of numerous CRE-dependent genes involved in B-cell function [293]. Tumor necrosis factor receptor-associated factor 3 (TRAF3) plays a critical role in inhibiting B-cell survival via inhibiting CREB stability [294]. Loss-of-function mutations of TRAF3 are commonly found in multiple myeloma, ovarian cancer and B-cell lymphoma [295, 296].
Macrophages
While phagocytosis of tumor cells by macrophages is a mechanism of immune surveillance, the tumor-associated macrophage (TAM) is a specific subpopulation of macrophages within the TME that are reprogrammed and hijacked by tumor cells to promote tumor progression [297]. cAMP has important roles in macrophage functions such as phagocytosis and microbial killing [298]. In alveolar macrophages, the resident immune effector cells in the lung, EPAC1 activation restrains phagocytosis, while PKA activation suppresses the production of leukotriene B4 (LTB4) and TNFα [299]. It is also reported that cAMP inhibits LPS-induced TNFα secretion via PKA-AKAP95-NFkBp105 in murine macrophage RAW264.7 cells [300]. cAMP and PGE2 stimulate the production of IL10 and G-CSF through PKA in RAW264.7 cells, which induces a pro-tumorigenic macrophage phenotype [300,301,302,303]. PGE2 secreted from breast cancer cells suppresses CCL5 secretion in LPS-activated macrophages through cAMP/PKA signaling pathway [304]. Moreover, cAMP signaling stimulates the M2 polarization of macrophages through a PKA-C/EBPβ-CREB dependent pathway in murine macrophages [305]. PGE2 enhances M2 polarization via the CREB-mediated induction of KLF4 [306]. Db-cAMP, a cAMP mimetic, promotes reprogramming of bone-marrow-derived macrophages to a M2 phenotype through increasing Arg-1/CD206/Ym-1 expression and IL10 levels in a PKA-dependent manner [307]. β2-Adrenoceptors in tumor-associated macrophages contribute to HCC progression through activating cAMP/PKA/CREB and cAMP/IL6 signaling pathways [308]. Thus, PKA is tightly involved in the promotion of pro-tumor macrophage polarization.
NK cells
NK cells are immune effectors that can directly recognize and kill tumor cells [309]. The presence of immunosuppressive factors in the TME, including PGE2, adenosine and cAMP, limits tumor-infiltrating NK cells persistence. cAMP signaling modulates the cytotoxicity of NK cells. When NK cells are exposed to lysis-sensitive tumor target cells, there is an increase in intracellular cAMP in NK cells, but no increase in response to lysis-resistant tumor target cells [310]. Several studies have identified that cAMP can inhibit the cytotoxicity and secretion of granzyme B, perforin and IFN-γ from NK cells through PKA/CREB signaling [311,312,313,314,315].
Recently, a study has shown that PDE4A confers resistance to PGE2-mediated suppression in NK cells by reducing intracellular levels of cAMP [316]. Moreover, adenosine suppresses various cytokines/chemokines production and inhibits the cytotoxic activity of human and murine NK cells via stimulation of A2aR/AC/cAMP signaling and subsequently activation of PKA type I [313, 317]. Blocking regulatory, but not catalytic, subunits of PKA type I abrogates the inhibitory effects of adenosine [313]. These data suggest that the regulatory subunits of PKA type I are the prominent contributor to the cAMP-mediated inhibitory effect on NK cells.
Myeloid-derived suppressor cells (MDSCs)
MDSCs are natural immunosuppressive cells, which block adaptive immunity by inhibiting the activation of CD4+ and CD8+ T cells and suppress innate immunity by inhibiting NK cells [318, 319]. PGE2 can induces COX2 expression in cultured peripheral blood-isolated monocytes, blocking their differentiation into CD1a+ DCs and promoting their development into MDSCs [320]. In a 4T1 mammary carcinoma model, PGE2 promoted tumor progression by inducing MDSCs partially through the EP2 receptor [321]. In addition to PGE2, selective EP2 and EP4 agonists, but not EP1/3 agonists, also induce MDSCs development through the AC/cAMP/PKA/CREB signaling pathway [320, 321]. MDSCs play a prominent role in tumor progression [322]. In a B16F10 mouse melanoma model, mice treated with a selective A2bR agonist Bay60-6583 showed increased VEGF production from MDSCs and tumor vessel density [323]. Pharmacological blockade of A2bR with PSB1115 reduced tumor-infiltrating MDSCs and restored an efficient antitumor T cell response, leading to a significant suppression of melanoma growth [323, 324]. Given that MDSCs has emerged as important contributor to tumor progression, more powerful evidence is needed to determine whether cAMP signaling can directly induce the development and activation of MDSCs.
Targeting cAMP signaling for cancer therapy
PKA inhibitor
Since the cAMP-PKA pathway is pro-tumorigenic and immunosuppressive in many types of cancer, PKA inhibitors may have anticancer effects in some types of cancer. There are three types of PKA inhibitor, including cAMP analog, small-molecule PKA inhibitor, and peptide inhibitor (Table 2). cAMP analogs have been shown to inhibit PKA activity by binding to the ATP-binding pocket of the PKA regulatory subunit [325]. Rp-8-Br-cAMP is a commonly used cAMP analog that competes with endogenous cAMP to bind to the PKA regulatory subunit. H89 is a widely used PKA inhibitor in preclinical studies, while it is non-specific. Treatment with H89 suppressed small cell lung cancer and Ewing sarcoma progression in animal models [326, 327]. Given that the PKA catalytic subunit also promotes the immunosuppressive macrophage phenotype in tumors, inhibition of PKA may have immunotherapeutic effects, especially when combined with αCTLA-4 antibody [328]. Liposomal H89 complexes with a diameter of about 1000 nm can be taken up by cancer-associated macrophages and thereby inhibit tumor growth and metastasis by promoting T cell activation [328]. KT-5720 is another PKA inhibitor with tumor suppressive effects. The MDR1-mediated drug resistance in hematological malignancies could be reversed by KT-5720 [329]. Daphnetin, a dehydroxylated derivative of coumarin isolated from plants Daphne species, has PKA-inhibitory activity with an IC50 value of 9.33 µM; however, it also non-specifically inhibits EGFR and PKC [330, 331]. Preclinical studies demonstrate that daphnetin has anticancer potential against leukemia, osteosarcoma, breast, ovary, kidney, colon, and liver cancers [332,333,334,335,336,337]. Recently, a novel diarylcyclohexanone derivative MHY4571 was developed as an orally active PKA inhibitor with anticancer effects [11].
Protein kinase inhibitor peptide (PKI) is another category of specific PKA inhibitor (Table 2). The endogenous PKI isoforms, PKIα/β/γ, are negative regulators of PKA activity. Several synthetic PKA inhibitor peptides have been developed. Among these peptides, PKI-(6-22)-amide is the most potent PKA inhibitor, followed by PKI-(5-24)-amide and PKI-(14-24)-amide. For detailed information on the development of PKA inhibitor peptides, we refer readers to the review by Liu et al. [325].
EPAC inhibitor
PKA can negatively feedback to inhibit cAMP synthesis. Thus, PKA inhibition may result in an increase in cAMP levels and divert cAMP signaling toward EPAC [338]. While EPAC activation may be detrimental to tumor suppression, H89 synergizes with the oncolytic virus M1 to inhibit tumor growth through EPAC activation [339]. Hence, this seemingly adverse effect may be exploited for combined therapy. It warrants further studies to determine the effects of 8-CPT-cAMP and 8-pCPT-2'-O-Me-cAMP-AM, two EPAC agonists, in tumor progression. In addition, I942 and SY009 are two EPAC1-specific non-cyclic nucleotide agonists which effects in cancer are poorly studied [340, 341].
3-(5-tert-butyl-isoxazol-3-yl)-2-[(3-chloro-phenyl)-hydrazono]-3-oxo-propionitrile (ESI-09) is a non-cyclic nucleotide EPAC-selective inhibitor that suppresses Rap1 activation, Akt phosphorylation and pancreatic cancer cell invasion [342, 343]. Other pan-EPAC inhibitors include ESI-08 and HJC0197. Further studies develop EPAC1- or EPAC2-specific inhibitors. While AAK-399, AAD-026 and MAY0132 are EPAC2-specific inhibitors [344], AM-001 and the (R)-enantiomer of CE3F4 (R)-CE3F4 are potent EPAC1 antagonists [345, 346]. So far, there are little reports on the effects of these EPAC inhibitors on tumor therapy. The inhibition of insulin secretion by AAK-399 and AAD-026 may be an adverse effect [344].
Adenylate cyclase and sAC inhibitors
Instead of inhibiting PKA and EPAC, another strategy to target cAMP signaling is direct inhibition of cAMP synthesis. SQ22536 (9-(tetrahydro-2-furanyl)-9H-purin-6-amine) is an adenine-like adenylate cyclase inhibitor. Another adenylate cyclase 1 inhibitor, NB-001, has analgesic effect on cancer pain [347]. The safety of NB001 is being tested in human clinical trials [348]. While the chromone-based ST034307 selectively inhibits adenylate cyclase 1 [349], the oxadiazole-based AC10065 can suppress both adenylate cyclase 1 and adenylate cyclase 8 [350]. The adenylate cyclase inhibitor MDL-12,330A suppresses cAMP synthesis in tumor tissue, leading to immune system-dependent inhibition of tumor progression [351]. In addition, the GPCR-adenylate cyclase-cAMP-PKA-CREB pathway is involved in the resistance of melanoma to MAPK inhibitors [352]. Except for the transmembrane adenylate cyclase, sAC is another target for cancer therapy. Bithionol and hexachlorophene are potent sAC-specific inhibitors that bind to bicarbonate-binding site [353]. Bithionol has been shown to enhance the sensitivity of ovarian cancer cells to cisplatin and paclitaxel [354]. These sAC inhibitors can suppress mitochondrial respiration and thereby limit the energy source in tumor cells [355].
Chimeric antigen receptor (CAR) T-cell therapy, an adoptive T-cell therapy involving the ex vivo transduction of a patient’s T cells with an engineered CAR that targets a defined tumor antigen, is a promising immunotherapy for hematological malignancies [356]. Since the adenosine-A2aR-adenylate cyclase-cAMP axis suppresses the function of CD8+ T-cell as well as CAR T-cell [357], blockade of A2aR in CAR-T cells can improve the efficacy of adoptively transferred T cells [358, 359]. It remains to know whether blockade of the predominant adenylate cyclase isoforms in CAR T-cells can achieve similar effect. Currently, the efficacy of CAR T-cell therapy is relatively poor in solid tumors [360], targeting the A2aR-adenylate cyclase-cAMP axis in CAR T-cells holds promise in improving CAR T-cell therapy for solid tumors.
cAMP-elevating agents
While inhibition of cAMP synthesis or PKA/EPAC can inhibit some types of cancer and stimulate anticancer immunity, cAMP-elevating agents such as the adenylate cyclase agonist forskolin and the cAMP analog 8-Br-cAMP also emerge as potential cancer therapeutics, especially when combined with other anticancer agents. Forskolin enhances paclitaxel- and H3K27me2/3 demethylases inhibitor GSKJ4-induced cytotoxicity in non-small-cell lung cancer and acute myeloid leukemia cells [95, 361]. Co-delivery of paclitaxel and forskolin by liquid crystal nanoparticles can inhibit the stemness of breast cancer stem cells and reverse chemoresistance, thereby eliciting potent antitumor activity [362]. Given the favorable safety profile, forskolin has been used to treat patients with glaucoma, asthma and heart failure [363]. However, the poor pharmacokinetic profile may make forskolin less effective for cancer therapy. The clearance rate and half-life for forskolin are 0.53 L/h and 3.9 h in mice, respectively [362]. It may be necessary to develop new delivery system to improve the efficacy of forskolin. cAMP analogs also inhibit medullary thyroid cancer cell growth [364]. PKA-mediated inhibitory phosphorylation of Raf1 contributes to the anticancer effects of cAMP, while cAMP activators also induce p21 expression independent of PKA. 5-Demethylnobiletin, a natural polymethoxyflavone in the extract of citrus fruits peels, is able to stimulate cAMP signaling and inhibit cancer [365, 366]. Nevertheless, these natural agents usually have multiple targets.
PDE inhibitors
An alternative choice to elevate intracellular cAMP levels is the suppression of PDE. Isobutylmethylxanthine is an anticancer agent that inhibits PDE3/4/5 with the IC50 values ranging from 6.5 to 31.7 µM [367]. Cilostamide and cilostazol are PDE3 inhibitors that have tumor suppressive activity in both solid tumors and hematological malignancies. Cilostamide synergizes with imatinib to inhibit cancer [368]. In humans receiving single oral administration of cilostazol at dose of 100 mg, the mean maximum concentration in serum (Cmax), concentration–time curve (AUC) and half-life (T1/2) are 701 ng/ml, 13,724 ng × h/mL and 13.5 h, respectively (https://drugs.ncats.io/drug/N7Z035406B). Combined treatment with statin and cilostazol can inhibit AML and MM cells survival [369]. In addition, cAMP can induce diffuse large B-cell lymphoma cell apoptosis by inhibiting spleen tyrosine kinase (SYK)/PI3K/AKT pathway, which is independent of PKA and EPAC [96]. Many receptor or non-receptor tyrosine kinases are the targets of cancer therapy. FDA-approved SYK inhibitors have been used to treat cancer [370]. The inhibition of SYK by cAMP may underlie the anticancer effects of certain agents that elevate intracellular cAMP levels. While PKA reportedly inhibits the activity of SYK in neutrophils [371], it promotes SYK activation in platelet [372]. It remains unclear how cAMP inhibits SYK in diffuse large B-cell lymphoma cell. Further studies are needed to determine the role of PKA in the regulation of SYK by cAMP in diffuse large B-cell lymphoma cell.
Rolipram is an inhibitor of PDE4A, PDE4B and PDE4D with its IC50 values of 4, 20 and 33 nM, respectively. Rolipram and another PDE4 inhibitor, eggmanone, suppress SHH and Hedgehog signaling thereby inhibiting medulloblastoma growth in GNAS-mutant mice [134, 373, 374]. PDE4A and PDE4D promote HIF signaling in lung cancer through cAMP-PKA/EPAC pathways. Treatment of lung cancer with PDE4 inhibitor suppresses cancer cell growth and angiogenesis [375]. PDE4D is overexpressed in LKB1-mutated lung cancer; therefore, PDE4 inhibitors may be more effective in treating LKB1-mutated lung cancer [376]. In addition, the PDE4D inhibitor roflumilast suppresses the growth of medulloblastoma that is resistant to SHH antagonist vismodegib [377]. Given that PDE4D interacts with mTORC1 and promotes its activation by inhibiting PKA-mediated raptor phosphorylation at Ser-791, pharmacological inhibition of PDE4D by roflumilast or GEBR-7b suppresses pancreatic cancer cell growth in vitro and in vivo [16]. Pharmacologic inhibition of PDE4D also suppresses prostate cancer growth by blocking sonic hedgehog, androgen receptor and MAPK pathways, and reverse tamoxifen resistance in estrogen receptor-positive breast cancer [378, 379]. Moreover, adenylate cyclase activator in combination with PDE inhibitor more significantly elevates cAMP levels and overcomes chemoresistance. Forskolin together with rolipram potently inhibits chemoresistant colon cancer cell growth [380]. PDE4B is overexpressed in diffuse large B-cell lymphoma and prevents cAMP-induced apoptosis [96, 381]. Hence, PDE4B inhibitors may have tumor suppressive effects in diffuse large B-cell lymphoma. PDE4B also restricts cAMP levels in colon cancer and abrogates cAMP-induced anticancer effect [382]. These studies indicate that PDE4 inhibitors may have anticancer effects on various types of cancer with specific genetic background.
PDE5 usually controls the degradation of the second messenger cGMP, an activator of protein kinase G (PKG); however, cGMP has been shown to inhibit the degradation of cAMP by other PDE family members [383]. Thus, the PDE5 inhibitors may indirectly elevate cAMP levels in tumor cells. Many PDE5 inhibitors such as sildenafil, vardenafil, tadalafil, and avanafil have been approved to treat pulmonary arterial hypertension. PDE5 inhibition also eliminates cancer stem cells, possibly through cAMP-PKA signaling [384]. Sildenafil, tadalafil and vardenafil impact cAMP-specific PDE8 isoforms-linked second messengers and steroid production in a mouse Leydig tumor cell line [383]. Accumulating evidence demonstrates that PDE5 inhibitors have anticancer effects by synergizing with chemotherapeutic agents [385,386,387]. Sildenafil and vardenafil have been shown to inhibit B-cell chronic lymphocytic leukemia cells growth [384]. Treatment of cancer cells with sildenafil increases the sensitivity of other chemotherapeutic drugs such as vincristine, etoposide, doxorubicin and cisplatin [388].
Some natural agents can also inhibit PDEs. Luteolin and 6-gingerol are PDE inhibitors from artichoke (Cynara scolymus) and ginger (Zingiber officinale), respectively [389]. In addition, the antimalarial agent artemisinin can inhibit calmodulin-mediated activation of PDEs and elevate cAMP levels [390]. These phytochemicals have anticancer effects especially when combining with other cytotoxic drugs [391,392,393,394]. Since these agents have multiple targets, it remains to know how cAMP may be involved in the anticancer effects of these agents.
Clinical development of anticancer agents targeting cAMP signaling pathways
Although PKA inhibitors can suppress tumor progression in some animal models, there are no PKA inhibitors undergoing clinical trials for cancer therapy. Global inhibition of cAMP, PKA and EPAC is challenging, due to the ubiquitous expression and activity of PKA and EPAC in normal physiology such as heart rhythm, optimum cardiac performance, synaptic plasticity, insulin secretion, sleep, learning and memory [395,396,397,398]. In addition, the pharmacological PKA inhibitors H89 and KT5720 have widespread effects independent of PKA [399]. Therefore, these inhibitors have on-target and off-target side effects. Some of the adverse effects, such as anxiety and depression disorders, may be intolerable [400]. To enable safe administration of PKA inhibitors for cancer therapy, future efforts may be made to precisely deliver PKA inhibitors to tumor site.
While the progression of PKA inhibitors into the clinic has been held back by severe adverse effects, many PDE inhibitors have been used in clinical setting. The PDE4 inhibitors roflumilast (Daliresp, Daxas), apremilast (Otezla), and crisaborole (Eucrisa) are in the market for treating chronic obstructive pulmonary disease, psoriasis, and moderate atopic dermatitis, respectively [401]. There are some clinical trials aiming to evaluate the safety and efficacy of PDE inhibitors in cancer patients (Table 3). In humans receiving single oral administration of roflumilast at a dose of 0.5 mg, the Cmax, AUC and T1/2 are 12.5 ng/ml, 65.1 ng × h/mL and 19.9 h, respectively (https://drugs.ncats.io/drug/0P6C6ZOP5U). Roflumilast in combination with prednisone was safe in patients with advanced B-cell malignancies; in this small-scale clinical trial, combination of roflumilast and prednisone resulted in partial response or stable disease in 66% of patients [402]. It remains to know the outcome of this regimen in larger clinical trials. The plasma concentration of roflumilast levels can reach 8.2 ng/ml, which is markedly above the IC50 value for inhibition of PDE4 (0.8 nM) [402]. In another pilot clinical study on relapsed/refractory diffuse large B-cell lymphoma (DLBCL), roflumilast in combination with etoposide, cisplatin, methylprednisolone and cytarabine resulted in better CR (46.2% vs. 34.6%), ORR (76.9% vs. 53.8%), and 1-year PFS (50.0% vs. 25.9%) compared with the chemotherapy-alone group [403]. There was no difference in median overall survival and 1-year overall survival between the roflumilast and chemotherapy-only arms. Importantly, PDE4B was a key downstream effector of DPY30, and the PDE4 inhibitor rolipram preferentially targeted DPY30-expressing cells [404]. It remains to know whether DPY30 can serve as a biomarker for precision treatment with PDE4 inhibitor such as roflumilast.
While the association between PDE5 inhibitors and colorectal cancer risk is controversial [408, 409], one study indicates that the use of PDE5 inhibitor after surgical resection may be related to a reduced risk of colorectal cancer-specific mortality [410]. Exisulind is a sulindac metabolite without anti-inflammatory activity, while it inhibits PDE5 [411]. One clinical trial has evaluated the effectiveness and safety of PDE5 inhibitors in treating metastatic breast cancer. Treatment of breast cancer patients with exisulind and capecitabine was well tolerated, while the effectiveness may be limited at the tested dosage [412]. Another phase I clinic trial explored the pharmacokinetic profile of exisulind in patients with advanced solid tumors [413]. There was a significant correlation between the plasma concentrations of exisulind and gastrointestinal toxicities [413]. In addition, clinical trials demonstrated that tadalafil increased T-cell expansion, reduced peripheral MDSC and Tregs numbers, and enhanced tumor-specific immunity in response to head and neck squamous cell carcinoma (HNSCC) lysate [414, 415]. Another clinical trial of combining tadalafil with the MUC1/polyICLC vaccine also demonstrates the safety and immunologic potential of this regimen in HNSCC patients [405]. However, the expression of PDL1 is upregulated after the treatment, indicating additional immune evasion [405]. A neoadjuvant randomized trial in patients with resectable HNSCC demonstrates that combination of the PDE5 inhibitor tadalafil with the PD-1 inhibitor nivolumab is safe and enhances immune-mediated anticancer effects, leading to pathologic treatment response in more than 20% patients [406]. Sildenafil, another PDE5 inhibitor that has been used in the clinic. A phase I trial has been conducted to evaluate the combination of sildenafil and regorafenib, a multi-kinase inhibitor, in advanced solid tumors [407]. Sildenafil can increase intratumoral T cell infiltration and activation, thereby improving the anticancer effectiveness of adoptive T cell treatment. More clinical studies are needed to evaluate the anticancer effects of PDE inhibition in combination with immunotherapy.
Concluding remarks
In conclusion, cAMP and the tumor microenvironment play a complex and interdependent role in cancer progression. cAMP may promote or inhibit cancer cell growth and metastasis in cancer types- and context-dependent manners. Activation of PKA by cAMP may either enhance or inhibit the activity of different oncogenes or tumor suppressors. Therefore, the type of oncogenic alteration is a determinant of the complex roles for cAMP signaling in cancer. Tumors with the DNAJB1–PRKACA gene fusion or BAP1-mutation share a molecular hallmark, namely aberrant activation of PKA [416]. Certainly, there are other driving events that lead to hyperactivation of cAMP signaling pathways during the development of some types of cancer. While cAMP promotes tumors such as FLC and basal cell carcinoma, it functions as a tumor suppressor in medulloblastoma by inactivating Gli. In addition, BRAF mutation may override the inhibition of Raf1 by PKA and thereby shift the balance between the tumor-promoting and tumor-suppressing effects of cAMP.
Although selective inhibitor of PKA can suppress the growth of PKA-addictive tumors, clinical development of such an inhibitor remains to be highly problematic because of the toxicity resulting from inhibition of the critical physiological functions of normal PKA. The discovery of selective small-molecule inhibitors of the DNAJB1–PRKACA chimera would be wonderful for precise treatment. Future efforts aimed at tumor-targeted delivery of PKA inhibitor may help break these bottlenecks. Alternatively, understanding the precise mechanisms by which cAMP, PKA and other effectors interact with the tumor microenvironment to promote tumor progression and immune resistance will be helpful for the development of new and effective therapies that target the key nodes in cAMP signaling. Evidence indicates that inhibition of cAMP synthesis can restore antitumor immunity and suppress tumorigenesis [351]. Except for the immune checkpoints, the RIG-I-like receptors/MAVS signaling pathway is emerging as another target for cancer immunotherapy [417]. cAMP/PKA promotes MAVS degradation by phosphorylating it at T54 [418]. It remains to know whether PKA inhibitors can improve the RIG-I-like receptors-targeted immunotherapy.
cAMP has both proapoptotic and antiapoptotic effects [44]. Currently, more efforts are paid for exploring the efficacy of cAMP-elevating compounds on cancer therapy. Indeed, cAMP and its signaling transducers such as PKA not only upregulate many oncogenes but also inactivate some oncogenes. Some types of cancer, such as basal cell carcinoma and medulloblastoma, are driven by oncogenes that are suppressed by cAMP and PKA. Therefore, elevating cAMP levels is supposed to be effective in treating these types of cancer. PDE inhibitors hold promise for cancer therapy. Many PDE inhibitors have been safely used for treating different diseases, and can be repurposed for cancer therapy. While PKA can inhibit cell apoptosis, it paradoxically promotes caspase-independent parthanatos, a type of programmable cell death characteristic of chromatinolysis and large-size DNA fragmentation, by phosphorylating PARP1 [47]. Also, the activation of EPAC may contribute to parthanatos by inhibiting PARP cleavage and upregulating PARP expression [419]. Thus, PDE inhibitors may synergize with agents that induce parthanatos. In addition, activation of PKA and EPAC by cAMP may sensitize cancer cells to lysosome-dependent cell death. PDE inhibitors in combination with lysosome-destabilizing agent may synergistically induce cancer cell death. Together, both tumor types and the exact nature of combined agents should be taken into consideration when cAMP-elevating compounds are supplemented with other anticancer agents. Another critical concern for the administration of cAMP-elevating agents for cancer therapy is the immune-suppressive effects of cAMP. Nonetheless, recent study indicates that the PDE4 inhibitor roflumilast does not reduce the clinical activity of immune checkpoint inhibitors, the mainstay in cancer immunotherapy [420, 421]. It warrants further studies to confirm this issue. Given that ferroptosis is inhibited by PKA but promoted by EPAC, PKA inhibition may enhance the efficacy of cAMP-elevating compound in combination with ferroptosis-inducing agents.
Abbreviations
- AC:
-
Adenylate cyclase
- AKAP:
-
A kinase anchor protein
- CAF:
-
Cancer-associated fibroblast
- cAMP:
-
Cyclic adenosine monophosphate
- CREB:
-
cAMP-responsive element-binding protein
- DCs:
-
Dendritic cells
- EBV:
-
Epstein–Barr virus
- EPAC:
-
Exchange protein activated by cAMP
- FLC:
-
Fibrolamellar hepatocellular carcinoma
- GPCR:
-
G protein-coupled receptor
- MDSC:
-
Myeloid-derived suppressor cell
- NETs:
-
Neutrophil extracellular traps
- PDE:
-
Phosphodiesterase
- PKA:
-
cAMP-dependent protein kinase
- PKI:
-
Protein kinase inhibitor peptide
- TAM:
-
Tumor-associated macrophage
- TME:
-
Tumor microenvironment
References
Uhler MD, Carmichael DF, Lee DC, Chrivia JC, Krebs EG, McKnight GS. Isolation of cDNA clones coding for the catalytic subunit of mouse cAMP-dependent protein kinase. Proc Natl Acad Sci USA. 1986;83:1300–4.
Caldwell JL, Lee IJ, Ngo L, Wang L, Bahriz S, Xu B, et al. Whole-heart multiparametric optical imaging reveals sex-dependent heterogeneity in cAMP signaling and repolarization kinetics. Sci Adv. 2023;9:eadd5799.
Martinez JM, Shen A, Xu B, Jovanovic A, de Chabot J, Zhang J, et al. Arrestin-dependent nuclear export of phosphodiesterase 4D promotes GPCR-induced nuclear cAMP signaling required for learning and memory. Sci Signal. 2023;16:eade3380.
Roy D, Balasubramanian S, Krishnamurthy PT, Sola P, Rymbai E. Phosphodiesterase-4 inhibition in Parkinson’s disease: molecular insights and therapeutic potential. Cell Mol Neurobiol. 2023;43:2713–41.
Fuentes-Pananá EM, Peng R, Brewer G, Tan J, Ling PD. Regulation of the Epstein–Barr virus C promoter by AUF1 and the cyclic AMP/protein kinase A signaling pathway. J Virol. 2000;74:8166–75.
Moar P, Sushmita K, Kateriya S, Tandon R. Transcriptional profiling indicates cAMP-driven reversal of HIV latency in monocytes occurs via transcription factor SP-1. Virology. 2020;542:40–53.
Luo Z, Li M, Li TW, Lv Z, Ye Z, Cisneros WJ, et al. Differential expression of CREM/ICER isoforms is associated with the spontaneous control of HIV infection. mBio. 2022;13:e0197921.
Xu K, Cooney KA, Shin EY, Wang L, Deppen JN, Ginn SC, et al. Adenosine from a biologic source regulates neutrophil extracellular traps (NETs). J Leukoc Biol. 2019;105:1225–34.
Ravani A, Vincenzi F, Bortoluzzi A, Padovan M, Pasquini S, Gessi S, et al. Role and function of A2A and A3 adenosine receptors in patients with ankylosing spondylitis, psoriatic arthritis and rheumatoid arthritis. Int J Mol Sci. 2017;18:697.
Zhang L, Yang N, Wang S, Huang B, Li F, Tan H, et al. Adenosine 2A receptor is protective against renal injury in MRL/lpr mice. Lupus. 2011;20:667–77.
Chung JH, Choi HJ, Kang YJ, Kim YS, Lee SY, Kwon RJ, et al. MHY4571, a novel diarylcyclohexanone derivative, exerts anti-cancer activity by regulating the PKA-cAMP-response element-binding protein pathway in squamous cell lung cancer. Exp Hematol Oncol. 2022;11:68.
Knighton DR, Zheng JH, Ten Eyck LF, Ashford VA, Xuong NH, Taylor SS, et al. Crystal structure of the catalytic subunit of cyclic adenosine monophosphate-dependent protein kinase. Science. 1991;253:407–14.
Stolfa I, Page C. Phosphodiesterase inhibitors and lung diseases. Adv Pharmacol. 2023;98:55–81.
Tsuji G, Yamamura K, Kawamura K, Kido-Nakahara M, Ito T, Nakahara T. Novel therapeutic targets for the treatment of atopic dermatitis. Biomedicines. 2023;11:1303.
Sadeghi MA, Nassireslami E, Yousefi Zoshk M, Hosseini Y, Abbasian K, Chamanara M. Phosphodiesterase inhibitors in psychiatric disorders. Psychopharmacology. 2023;240:1201–19.
Jeong MH, Urquhart G, Lewis C, Chi Z, Jewell JL. Inhibition of phosphodiesterase 4D suppresses mTORC1 signaling and pancreatic cancer growth. JCI Insight. 2023;8:e158098.
Lee A, Lebedyeva I, Zhi W, Senthil V, Cheema H, Brands MW, et al. A non-systemic phosphodiesterase-5 inhibitor suppresses colon proliferation in mice. Int J Mol Sci. 2023;24:9397.
Godbole A, Lyga S, Lohse MJ, Calebiro D. Internalized TSH receptors en route to the TGN induce local G(s)-protein signaling and gene transcription. Nat Commun. 2017;8:443.
Arkhipov A, Khuzakhmetova V, Petrov AM, Bukharaeva EA. Catecholamine-dependent hyperpolarization of the junctional membrane via β2-adrenoreceptor/G(i)-protein/α2-Na-K-ATPase pathway. Brain Res. 2022;1795:148072.
Tang G, Guo Y, Zhang L, Wang T, Li R, Yang J, et al. 5-HT(1B) receptors in the basolateral amygdaloid nucleus regulate anxiety-like behaviors through AC-PKA signal pathway in a rat model of Parkinson’s disease. Behav Brain Res. 2023;449:114488.
Ostrom KF, LaVigne JE, Brust TF, Seifert R, Dessauer CW, Watts VJ, et al. Physiological roles of mammalian transmembrane adenylyl cyclase isoforms. Physiol Rev. 2022;102:815–57.
Pizzoni A, Zhang X, Naim N, Altschuler DL. Soluble cyclase-mediated nuclear cAMP synthesis is sufficient for cell proliferation. Proc Natl Acad Sci USA. 2023;120:e2208749120.
Tenner B, Getz M, Ross B, Ohadi D, Bohrer CH, Greenwald E, et al. Spatially compartmentalized phase regulation of a Ca(2+)-cAMP-PKA oscillatory circuit. Elife. 2020;9:e55013.
Piggott LA, Bauman AL, Scott JD, Dessauer CW. The A-kinase anchoring protein Yotiao binds and regulates adenylyl cyclase in brain. Proc Natl Acad Sci USA. 2008;105:13835–40.
Maurice DH, Ke H, Ahmad F, Wang Y, Chung J, Manganiello VC. Advances in targeting cyclic nucleotide phosphodiesterases. Nat Rev Drug Discov. 2014;13:290–314.
Beavo JA. Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiol Rev. 1995;75:725–48.
Andrew TB, Joseph AB. Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use. Pharmacol Rev. 2006;58:488.
Paes D, Schepers M, Rombaut B, van den Hove D, Vanmierlo T, Prickaerts J. The molecular biology of phosphodiesterase 4 enzymes as pharmacological targets: an interplay of isoforms, conformational states, and inhibitors. Pharmacol Rev. 2021;73:1016–49.
Sassi Y, Ahles A, Truong DJ, Baqi Y, Lee SY, Husse B, et al. Cardiac myocyte-secreted cAMP exerts paracrine action via adenosine receptor activation. J Clin Invest. 2014;124:5385–97.
Jackson EK, Raghvendra DK. The extracellular cyclic AMP-adenosine pathway in renal physiology. Annu Rev Physiol. 2004;66:571–99.
Giron MC, Bin A, Brun P, Etteri S, Bolego C, Florio C, et al. Cyclic AMP in rat ileum: evidence for the presence of an extracellular cyclic AMP-adenosine pathway. Gastroenterology. 2008;134:1116–26.
Brzostowski JA, Sawai S, Rozov O, Liao XH, Imoto D, Parent CA, et al. Phosphorylation of chemoattractant receptors regulates chemotaxis, actin reorganization and signal relay. J Cell Sci. 2013;126:4614–26.
Yang H, Li G, Wu JJ, Wang L, Uhler M, Simeone DM. Protein kinase A modulates transforming growth factor-β signaling through a direct interaction with Smad4 protein. J Biol Chem. 2013;288:8737–49.
Oerlecke I, Bauer E, Dittmer A, Leyh B, Dittmer J. Cyclic AMP enhances TGFβ responses of breast cancer cells by upregulating TGFβ receptor I expression. PLOS ONE. 2013;8:e54261.
Zhang H, Kong Q, Wang J, Jiang Y, Hua H. Complex roles of cAMP-PKA-CREB signaling in cancer. Exp Hematol Oncol. 2020;9:32.
Lin D, Chen Y, Koksal AR, Dash S, Aydin Y. Targeting ER stress/PKA/GSK-3β/β-catenin pathway as a potential novel strategy for hepatitis C virus-infected patients. Cell Commun Signal. 2023;21:102.
Maurice DH. PDE8A runs interference to limit PKA inhibition of Raf-1. Proc Natl Acad Sci USA. 2013;110:6248–9.
Soundararajan R, Hernández-Cuervo H, Stearns TM, Griswold AJ, Patil SS, Fukumoto J, et al. A-Kinase Anchor Protein 1 deficiency causes mitochondrial dysfunction in mouse model of hyperoxia induced acute lung injury. Front Pharmacol. 2022;13:980723.
Inigo JR, Chandra D. The mitochondrial unfolded protein response (UPR(mt)): shielding against toxicity to mitochondria in cancer. J Hematol Oncol. 2022;15:98.
Omar MH, Scott JD. AKAP signaling Islands: venues for precision pharmacology. Trends Pharmacol Sci. 2020;41:933–46.
Bucko PJ, Scott JD. Drugs that regulate local cell signaling: AKAP targeting as a therapeutic option. Annu Rev Pharmacol Toxicol. 2021;61:361–79.
Pidoux G, Taskén K. Specificity and spatial dynamics of protein kinase A signaling organized by A-kinase-anchoring proteins. J Mol Endocrinol. 2010;44:271–84.
Ruppelt A, Mosenden R, Grönholm M, Aandahl EM, Tobin D, Carlson CR, et al. Inhibition of T cell activation by cyclic adenosine 5’-monophosphate requires lipid raft targeting of protein kinase A type I by the A-kinase anchoring protein ezrin. J Immunol. 2007;179:5159–68.
Insel PA, Zhang L, Murray F, Yokouchi H, Zambon AC. Cyclic AMP is both a pro-apoptotic and anti-apoptotic second messenger. Acta Physiol (Oxf). 2012;204:277–87.
Keshwani MM, Kanter JR, Ma Y, Wilderman A, Darshi M, Insel PA, et al. Mechanisms of cyclic AMP/protein kinase A- and glucocorticoid-mediated apoptosis using S49 lymphoma cells as a model system. Proc Natl Acad Sci USA. 2015;112:12681–6.
Ye J, Zeng B, Zhong M, Li H, Xu L, Shu J, et al. Scutellarin inhibits caspase-11 activation and pyroptosis in macrophages via regulating PKA signaling. Acta Pharm Sin B. 2021;11:112–26.
Brunyanszki A, Olah G, Coletta C, Szczesny B, Szabo C. Regulation of mitochondrial poly(ADP-Ribose) polymerase activation by the β-adrenoceptor/cAMP/protein kinase A axis during oxidative stress. Mol Pharmacol. 2014;86:450–62.
Shishikura K, Horiuchi T, Sakata N, Trinh DA, Shirakawa R, Kimura T, et al. Prostaglandin E2 inhibits neutrophil extracellular trap formation through production of cyclic AMP. Br J Pharmacol. 2016;173:319–31.
Shi L, Chen H, Chen K, Zhong C, Song C, Huang Y, et al. The DRD2 antagonist haloperidol mediates autophagy-induced ferroptosis to increase temozolomide sensitivity by promoting endoplasmic reticulum stress in glioblastoma. Clin Cancer Res. 2023;29:3172–88.
Guan Q, Wang Z, Hu K, Cao J, Dong Y, Chen Y. Melatonin ameliorates hepatic ferroptosis in NAFLD by inhibiting ER stress via the MT2/cAMP/PKA/IRE1 signaling pathway. Int J Biol Sci. 2023;19:3937–50.
Robichaux WG 3rd, Cheng X. Intracellular cAMP sensor EPAC: physiology, pathophysiology, and therapeutics development. Physiol Rev. 2018;98:919–1053.
Kumar N, Prasad P, Jash E, Saini M, Husain A, Goldman A, et al. Insights into exchange factor directly activated by cAMP (EPAC) as potential target for cancer treatment. Mol Cell Biochem. 2018;447:77–92.
Singhmar P, Huo X, Eijkelkamp N, Berciano SR, Baameur F, Mei FC, et al. Critical role for Epac1 in inflammatory pain controlled by GRK2-mediated phosphorylation of Epac1. Proc Natl Acad Sci USA. 2016;113:3036–41.
Khaliulin I, Bond M, James AF, Dyar Z, Amini R, Johnson JL, et al. Functional and cardioprotective effects of simultaneous and individual activation of protein kinase A and Epac. Br J Pharmacol. 2017;174:438–53.
Goode DJ, Molliver DC. Regulation of mitochondrial function by Epac2 contributes to acute inflammatory hyperalgesia. J Neurosci. 2021;41:2883–98.
Okumura S, Fujita T, Cai W, Jin M, Namekata I, Mototani Y, et al. Epac1-dependent phospholamban phosphorylation mediates the cardiac response to stresses. J Clin Invest. 2014;124:2785–801.
Gu Y, Li G, Huang LM. Inflammation induces Epac-protein kinase C alpha and epsilon signaling in TRPV1-mediated hyperalgesia. Pain. 2018;159:2383–93.
Goode DJ, Molliver DC. Phospho-substrate profiling of Epac-dependent protein kinase C activity. Mol Cell Biochem. 2019;456:167–78.
Hua H, Zhang H, Chen J, Wang J, Liu J, Jiang Y. Targeting Akt in cancer for precision therapy. J Hematol Oncol. 2021;14:128.
Hochbaum D, Tanos T, Ribeiro-Neto F, Altschuler D, Coso OA. Activation of JNK by Epac is independent of its activity as a Rap guanine nucleotide exchanger. J Biol Chem. 2003;278:33738–46.
Wehbe N, Slika H, Mesmar J, Nasser SA, Pintus G, Baydoun S, et al. The role of Epac in cancer progression. Int J Mol Sci. 2020;21:6489.
Lakhter AJ, Naidu SR. Cyclic AMP-Epac signaling pathway contributes to repression of PUMA transcription in melanoma cells. Melanoma Res. 2017;27:411–6.
Musheshe N, Oun A, Sabogal-Guáqueta AM, Trombetta-Lima M, Mitchel SC, Adzemovic A, et al. Pharmacological inhibition of Epac1 averts ferroptosis cell death by preserving mitochondrial integrity. Antioxidants (Basel). 2022;11:314.
Zhang T, Luu MDA, Dolga AM, Eisel ULM, Schmidt M. The old second messenger cAMP teams up with novel cell death mechanisms: potential translational therapeutical benefit for Alzheimer’s disease and Parkinson’s disease. Front Physiol. 2023;14:1207280.
Page DA, Magee KEA, Li J, Jung M, Young EC. Cytoplasmic autoinhibition in HCN channels is regulated by the transmembrane region. J Membr Biol. 2020;253:153–66.
Magee KE, Madden Z, Young EC. HCN channel C-terminal region speeds activation rates independently of autoinhibition. J Membr Biol. 2015;248:1043–60.
Evans EGB, Morgan JLW, DiMaio F, Zagotta WN, Stoll S. Allosteric conformational change of a cyclic nucleotide-gated ion channel revealed by DEER spectroscopy. Proc Natl Acad Sci USA. 2020;117:10839–47.
Amunjela JN, Tucker SJ. POPDC proteins as potential novel therapeutic targets in cancer. Drug Discov Today. 2016;21:1920–7.
Baldwin TA, Li Y, Marsden AN, Rinné S, Garza-Carbajal A, Schindler RFR, et al. POPDC1 scaffolds a complex of adenylyl cyclase 9 and the potassium channel TREK-1 in heart. EMBO Rep. 2022;23:e55208.
Tibbo AJ, Mika D, Dobi S, Ling J, McFall A, Tejeda GS, et al. Phosphodiesterase type 4 anchoring regulates cAMP signaling to Popeye domain-containing proteins. J Mol Cell Cardiol. 2022;165:86–102.
Gingold-Belfer R, Kessler-Icekson G, Morgenstern S, Rath-Wolfson L, Zemel R, Boltin D, et al. The transition from gastric intestinal metaplasia to gastric cancer involves POPDC1 and POPDC3 downregulation. Int J Mol Sci. 2021;22:5359.
Amunjela JN, Tucker SJ. POPDC1 is suppressed in human breast cancer tissues and is negatively regulated by EGFR in breast cancer cell lines. Cancer Lett. 2017;406:81–92.
Wu Y, Zhang J. Study on differentially expressed genes between stage M and stage MS neuroblastoma. Front Oncol. 2022;12:1083570.
Kim M, Jang HR, Haam K, Kang TW, Kim JH, Kim SY, et al. Frequent silencing of popeye domain-containing genes, BVES and POPDC3, is associated with promoter hypermethylation in gastric cancer. Carcinogenesis. 2010;31:1685–93.
Osler ME, Chang MS, Bader DM. Bves modulates epithelial integrity through an interaction at the tight junction. J Cell Sci. 2005;118:4667–78.
Hager HA, Roberts RJ, Cross EE, Proux-Gillardeaux V, Bader DM. Identification of a novel Bves function: regulation of vesicular transport. Embo J. 2010;29:532–45.
Parang B, Kaz AM, Barrett CW, Short SP, Ning W, Keating CE, et al. BVES regulates c-Myc stability via PP2A and suppresses colitis-induced tumourigenesis. Gut. 2017;66:852–62.
Han P, Fu Y, Luo M, He J, Liu J, Liao J, et al. BVES inhibition triggers epithelial-mesenchymal transition in human hepatocellular carcinoma. Dig Dis Sci. 2014;59:992–1000.
Ramms DJ, Raimondi F, Arang N, Herberg FW, Taylor SS, Gutkind JS. Gαs-protein kinase A (PKA) pathway signalopathies: the emerging genetic landscape and therapeutic potential of human diseases driven by aberrant Gαs-PKA signaling. Pharmacol Rev. 2021;73:155–97.
Djari C, Sahut-Barnola I, Septier A, Plotton I, Montanier N, Dufour D, et al. Protein kinase A drives paracrine crisis and WNT4-dependent testis tumor in Carney complex. J Clin Invest. 2021;131:e146910.
Beuschlein F, Fassnacht M, Assié G, Calebiro D, Stratakis CA, Osswald A, et al. Constitutive activation of PKA catalytic subunit in adrenal Cushing’s syndrome. N Engl J Med. 2014;370:1019–28.
Pitsava G, Stratakis CA, Faucz FR. PRKAR1A and thyroid tumors. Cancers (Basel). 2021;13:3834.
Lucia K, Wu Y, Garcia JM, Barlier A, Buchfelder M, Saeger W, et al. Hypoxia and the hypoxia inducible factor 1α activate protein kinase A by repressing RII beta subunit transcription. Oncogene. 2020;39:3367–80.
Bolger GB. The cAMP-signaling cancers: Clinically-divergent disorders with a common central pathway. Front Endocrinol (Lausanne). 2022;13:1024423.
Horvath A, Bertherat J, Groussin L, Guillaud-Bataille M, Tsang K, Cazabat L, et al. Mutations and polymorphisms in the gene encoding regulatory subunit type 1-alpha of protein kinase A (PRKAR1A): an update. Hum Mutat. 2010;31:369–79.
Sato Y, Maekawa S, Ishii R, Sanada M, Morikawa T, Shiraishi Y, et al. Recurrent somatic mutations underlie corticotropin-independent Cushing’s syndrome. Science. 2014;344:917–20.
Espiard S, Knape MJ, Bathon K, Assié G, Rizk-Rabin M, Faillot S, et al. Activating PRKACB somatic mutation in cortisol-producing adenomas. JCI Insight. 2018;3:e98296.
Neumayer C, Ng D, Jiang CS, Qureshi A, Lalazar G, Vaughan R, et al. Oncogenic addiction of fibrolamellar hepatocellular carcinoma to the fusion kinase DNAJB1-PRKACA. Clin Cancer Res. 2023;29:271–8.
Vyas M, Hechtman JF, Zhang Y, Benayed R, Yavas A, Askan G, et al. DNAJB1-PRKACA fusions occur in oncocytic pancreatic and biliary neoplasms and are not specific for fibrolamellar hepatocellular carcinoma. Mod Pathol. 2020;33:648–56.
Singhi AD, Wood LD, Parks E, Torbenson MS, Felsenstein M, Hruban RH, et al. Recurrent rearrangements in PRKACA and PRKACB in intraductal oncocytic papillary neoplasms of the pancreas and bile duct. Gastroenterology. 2020;158:573–82.
Gausdal G, Wergeland A, Skavland J, Nguyen E, Pendino F, Rouhee N, et al. Cyclic AMP can promote APL progression and protect myeloid leukemia cells against anthracycline-induced apoptosis. Cell Death Dis. 2013;4:e516.
Naderi EH, Findley HW, Ruud E, Blomhoff HK, Naderi S. Activation of cAMP signaling inhibits DNA damage-induced apoptosis in BCP-ALL cells through abrogation of p53 accumulation. Blood. 2009;114:608–18.
Kusnadi EP, Trigos AS, Cullinane C, Goode DL, Larsson O, Devlin JR, et al. Reprogrammed mRNA translation drives resistance to therapeutic targeting of ribosome biogenesis. Embo J. 2020;39:e105111.
Roderick JE, Gallagher KM, Murphy LC, O’Connor KW, Tang K, Zhang B, et al. Prostaglandin E2 stimulates cAMP signaling and resensitizes human leukemia cells to glucocorticoid-induced cell death. Blood. 2021;137:500–12.
Illiano M, Conte M, Sapio L, Nebbioso A, Spina A, Altucci L, et al. Forskolin sensitizes human acute myeloid leukemia cells to H3K27me2/3 demethylases GSKJ4 inhibitor via protein kinase A. Front Pharmacol. 2018;9:792.
Smith PG, Wang F, Wilkinson KN, Savage KJ, Klein U, Neuberg DS, et al. The phosphodiesterase PDE4B limits cAMP-associated PI3K/AKT-dependent apoptosis in diffuse large B-cell lymphoma. Blood. 2005;105:308–16.
Wang Z, Zhang X, Tian X, Yang Y, Ma L, Wang J, et al. CREB stimulates GPX4 transcription to inhibit ferroptosis in lung adenocarcinoma. Oncol Rep. 2021;45:88.
Alam SK, Zhang Y, Wang L, Zhu Z, Hernandez CE, Zhou Y, et al. DARPP-32 promotes ERBB3-mediated resistance to molecular targeted therapy in EGFR-mutated lung adenocarcinoma. Oncogene. 2022;41:83–98.
Kim IK, McCutcheon JN, Rao G, Liu SV, Pommier Y, Skrzypski M, et al. Acquired SETD2 mutation and impaired CREB1 activation confer cisplatin resistance in metastatic non-small cell lung cancer. Oncogene. 2019;38:180–93.
Wang YW, Chen X, Gao JW, Zhang H, Ma RR, Gao ZH, et al. High expression of cAMP-responsive element-binding protein 1 (CREB1) is associated with metastasis, tumor stage and poor outcome in gastric cancer. Oncotarget. 2015;6:10646–57.
Sun DP, Fang CL, Chen HK, Wen KS, Hseu YC, Hung ST, et al. EPAC1 overexpression is a prognostic marker and its inhibition shows promising therapeutic potential for gastric cancer. Oncol Rep. 2017;37:1953–60.
Zhu S, Soutto M, Chen Z, Blanca Piazuelo M, Kay Washington M, Belkhiri A, et al. Activation of IGF1R by DARPP-32 promotes STAT3 signaling in gastric cancer cells. Oncogene. 2019;38:5805–16.
Zhu S, Soutto M, Chen Z, Peng D, Romero-Gallo J, Krishna US, et al. Helicobacter pylori-induced cell death is counteracted by NF-κB-mediated transcription of DARPP-32. Gut. 2017;66:761–2.
Zhu S, Khalafi S, Chen Z, Poveda J, Peng D, Lu H, et al. Silencing of miR490-3p by H. pylori activates DARPP-32 and induces resistance to gefitinib. Cancer Lett. 2020;491:87–96.
Gummadi J, Wang X, Xie C. Current advances in the treatment of fibrolamellar carcinoma of liver. J Hepatocell Carcinoma. 2023;10:745–52.
Hirsch TZ, Negulescu A, Gupta B, Caruso S, Noblet B, Couchy G, et al. BAP1 mutations define a homogeneous subgroup of hepatocellular carcinoma with fibrolamellar-like features and activated PKA. J Hepatol. 2020;72:924–36.
Nault JC, Fabre M, Couchy G, Pilati C, Jeannot E, van Nhieu JT, et al. GNAS-activating mutations define a rare subgroup of inflammatory liver tumors characterized by STAT3 activation. J Hepatol. 2012;56:184–91.
Li Y, Fu Y, Hu X, Sun L, Tang D, Li N, et al. The HBx-CTTN interaction promotes cell proliferation and migration of hepatocellular carcinoma via CREB1. Cell Death Dis. 2019;10:405.
Zhang H, Yang S, Wang J, Jiang Y. Blockade of AMPK-mediated cAMP-PKA-CREB/ATF1 signaling synergizes with aspirin to inhibit hepatocellular carcinoma. Cancers (Basel). 2021;13:1738.
Wang J, Ma L, Weng W, Qiao Y, Zhang Y, He J, et al. Mutual interaction between YAP and CREB promotes tumorigenesis in liver cancer. Hepatology. 2013;58:1011–20.
Zhou M, Mok MT, Sun H, Chan AW, Huang Y, Cheng AS, et al. The anti-diabetic drug exenatide, a glucagon-like peptide-1 receptor agonist, counteracts hepatocarcinogenesis through cAMP-PKA-EGFR-STAT3 axis. Oncogene. 2017;36:4135–49.
Massimi M, Cardarelli S, Galli F, Giardi MF, Ragusa F, Panera N, et al. Increase of intracellular cyclic AMP by PDE4 inhibitors affects HepG2 cell cycle progression and survival. J Cell Biochem. 2017;118:1401–11.
Ragusa F, Panera N, Cardarelli S, Scarsella M, Bianchi M, Biagioni S, et al. Phosphodiesterase 4D depletion/inhibition exerts anti-oncogenic properties in hepatocellular carcinoma. Cancers (Basel). 2021;13:2182.
Pan P, Oshima K, Huang YW, Agle KA, Drobyski WR, Chen X, et al. Loss of FFAR2 promotes colon cancer by epigenetic dysregulation of inflammation suppressors. Int J Cancer. 2018;143:886–96.
Han J, Jiang Q, Ma R, Zhang H, Tong D, Tang K, et al. Norepinephrine-CREB1-miR-373 axis promotes progression of colon cancer. Mol Oncol. 2020;14:1059–73.
Hany D, Vafeiadou V, Picard D. CRISPR-Cas9 screen reveals a role of purine synthesis for estrogen receptor α activity and tamoxifen resistance of breast cancer cells. Sci Adv. 2023;9:eadd3685.
Stevens LE, Peluffo G, Qiu X, Temko D, Fassl A, Li Z, et al. JAK-STAT signaling in inflammatory breast cancer enables chemotherapy-resistant cell states. Cancer Res. 2023;83:264–84.
Chen S, Paul MR, Sterner CJ, Belka GK, Wang D, Xu P, et al. PAQR8 promotes breast cancer recurrence and confers resistance to multiple therapies. Breast Cancer Res. 2023;25:1.
Mukherjee P, Bagchi A, Banerjee A, Roy H, Bhattacharya A, Biswas A, et al. PDE4 inhibitor eliminates breast cancer stem cells via noncanonical activation of mTOR. J Cell Biochem. 2022;123:1980–96.
Persaud L, Mighty J, Zhong X, Francis A, Mendez M, Muharam H, et al. IL-24 promotes apoptosis through cAMP-dependent PKA pathways in human breast cancer cells. Int J Mol Sci. 2018;19:3561.
Illiano M, Sapio L, Salzillo A, Capasso L, Caiafa I, Chiosi E, et al. Forskolin improves sensitivity to doxorubicin of triple negative breast cancer cells via Protein Kinase A-mediated ERK1/2 inhibition. Biochem Pharmacol. 2018;152:104–13.
Hao N, Shen W, Du R, Jiang S, Zhu J, Chen Y, et al. Phosphodiesterase 3A represents a therapeutic target that drives stem cell-like property and metastasis in breast cancer. Mol Cancer Ther. 2020;19:868–81.
Yue W, Ma J, Xiao Y, Wang P, Gu X, Xie B, et al. The apoptotic resistance of BRCA1-deficient ovarian cancer cells is mediated by cAMP. Front Cell Dev Biol. 2022;10:889656.
Kang Y, Nagaraja AS, Armaiz-Pena GN, Dorniak PL, Hu W, Rupaimoole R, et al. Adrenergic stimulation of DUSP1 impairs chemotherapy response in ovarian cancer. Clin Cancer Res. 2016;22:1713–24.
Rodríguez CI, Castro-Pérez E, Prabhakar K, Block L, Longley BJ, Wisinski JA, et al. EPAC-RAP1 axis-mediated switch in the response of primary and metastatic melanoma to cyclic AMP. Mol Cancer Res. 2017;15:1792–802.
Rodríguez CI, Castro-Pérez E, Longley BJ, Setaluri V. Elevated cyclic AMP levels promote BRAF(CA)/Pten(-/-) mouse melanoma growth but pCREB is negatively correlated with human melanoma progression. Cancer Lett. 2018;414:268–77.
Krishnan A, Bhasker AI, Singh MK, Rodriguez CI, Pérez EC, Altameemi S, et al. EPAC regulates melanoma growth by stimulating mTORC1 signaling and loss of EPAC signaling dependence correlates with melanoma progression. Mol Cancer Res. 2022;20:1548–60.
Ostojić J, Yoon YS, Sonntag T, Nguyen B, Vaughan JM, Shokhirev M, et al. Transcriptional co-activator regulates melanocyte differentiation and oncogenesis by integrating cAMP and MAPK/ERK pathways. Cell Rep. 2021;35:109136.
Dagar M, Singh JP, Dagar G, Tyagi RK, Bagchi G. Phosphorylation of HSP90 by protein kinase A is essential for the nuclear translocation of androgen receptor. J Biol Chem. 2019;294:8699–710.
Zhao N, Peacock SO, Lo CH, Heidman LM, Rice MA, Fahrenholtz CD, et al. Arginine vasopressin receptor 1a is a therapeutic target for castration-resistant prostate cancer. Sci Transl Med. 2019;11:eaaw4636.
Pan W, Zhang Z, Kimball H, Qu F, Berlind K, Stopsack KH, et al. Abiraterone acetate induces CREB1 phosphorylation and enhances the function of the CBP-p300 complex, leading to resistance in prostate cancer cells. Clin Cancer Res. 2021;27:2087–99.
Zhang Y, Zheng D, Zhou T, Song H, Hulsurkar M, Su N, et al. Androgen deprivation promotes neuroendocrine differentiation and angiogenesis through CREB-EZH2-TSP1 pathway in prostate cancers. Nat Commun. 2018;9:4080.
Cheng Y, Gao XH, Li XJ, Cao QH, Zhao DD, Zhou JR, et al. Depression promotes prostate cancer invasion and metastasis via a sympathetic-cAMP-FAK signaling pathway. Oncogene. 2018;37:2953–66.
He X, Zhang L, Chen Y, Remke M, Shih D, Lu F, et al. The G protein α subunit Gαs is a tumor suppressor in Sonic Hedgehog-driven medulloblastoma. Nat Med. 2014;20:1035–42.
Iglesias-Bartolome R, Torres D, Marone R, Feng X, Martin D, Simaan M, et al. Inactivation of a Gα(s)-PKA tumour suppressor pathway in skin stem cells initiates basal-cell carcinogenesis. Nat Cell Biol. 2015;17:793–803.
Chowdhury MAR, An J, Jeong S. The pleiotropic face of CREB family transcription factors. Mol Cells. 2023;46:399–413.
Delidaki M, Gu M, Hein A, Vatish M, Grammatopoulos DK. Interplay of cAMP and MAPK pathways in hCG secretion and fusogenic gene expression in a trophoblast cell line. Mol Cell Endocrinol. 2011;332:213–20.
Pan Z, Xu T, Bao L, Hu X, Jin T, Chen J, et al. CREB3L1 promotes tumor growth and metastasis of anaplastic thyroid carcinoma by remodeling the tumor microenvironment. Mol Cancer. 2022;21:190.
Rose M, Schubert C, Dierichs L, Gaisa NT, Heer M, Heidenreich A, et al. OASIS/CREB3L1 is epigenetically silenced in human bladder cancer facilitating tumor cell spreading and migration in vitro. Epigenetics. 2014;9:1626–40.
Saito A, Kamikawa Y, Ito T, Matsuhisa K, Kaneko M, Okamoto T, et al. p53-independent tumor suppression by cell-cycle arrest via CREB/ATF transcription factor OASIS. Cell Rep. 2023;42:112479.
Feng YX, Jin DX, Sokol ES, Reinhardt F, Miller DH, Gupta PB. Cancer-specific PERK signaling drives invasion and metastasis through CREB3L1. Nat Commun. 2017;8:1079.
Mellor P, Kendall S, Smith S, Saxena A, Anderson DH. Reduced CREB3L1 expression in triple negative and luminal a breast cancer cells contributes to enhanced cell migration, anchorage-independent growth and metastasis. PLoS ONE. 2022;17:e0271090.
Sharifnia T, Wawer MJ, Goodale A, Lee Y, Kazachkova M, Dempster JM, et al. Mapping the landscape of genetic dependencies in chordoma. Nat Commun. 2023;14:1933.
Cheng JC, Kinjo K, Judelson DR, Chang J, Wu WS, Schmid I, et al. CREB is a critical regulator of normal hematopoiesis and leukemogenesis. Blood. 2008;111:1182–92.
Cho EC, Mitton B, Sakamoto KM. CREB and leukemogenesis. Crit Rev Oncog. 2011;16:37–46.
Sandoval S, Kraus C, Cho EC, Cho M, Bies J, Manara E, et al. Sox4 cooperates with CREB in myeloid transformation. Blood. 2012;120:155–65.
Li Y, Chen J, Yang W, Liu H, Wang J, Xiao J, et al. mPGES-1/PGE2 promotes the growth of T-ALL cells in vitro and in vivo by regulating the expression of MTDH via the EP3/cAMP/PKA/CREB pathway. Cell Death Dis. 2020;11:221.
Seo W, Silwal P, Song IC, Jo EK. The dual role of autophagy in acute myeloid leukemia. J Hematol Oncol. 2022;15:51.
Skah S, Richartz N, Duthil E, Gilljam KM, Bindesbøll C, Naderi EH, et al. cAMP-mediated autophagy inhibits DNA damage-induced death of leukemia cells independent of p53. Oncotarget. 2018;9:30434–49.
Richartz N, Pietka W, Gilljam KM, Skah S, Skålhegg BS, Bhagwat S, et al. cAMP-mediated autophagy promotes cell survival via ROS-Induced activation of PARP1: implications for treatment of acute lymphoblastic leukemia. Mol Cancer Res. 2022;20:400–11.
Byun JM, Min CK, Kim K, Bang SM, Lee JJ, Kim JS, et al. Phase II trial of daratumumab with DCEP in relapsed/refractory multiple myeloma patients with extramedullary disease. J Hematol Oncol. 2022;15:150.
Korsos V, Miller WH Jr. How retinoic acid and arsenic transformed acute promyelocytic leukemia therapy. J Mol Endocrinol. 2022;69:T69-t83.
Zhao Q, Tao J, Zhu Q, Jia PM, Dou AX, Li X, et al. Rapid induction of cAMP/PKA pathway during retinoic acid-induced acute promyelocytic leukemia cell differentiation. Leukemia. 2004;18:285–92.
He B, Chang Y, Yang C, Zhang Z, Xu G, Feng X, et al. Adenylate cyclase 7 regulated by miR-192 promotes ATRA-induced differentiation of acute promyelocytic leukemia cells. Biochem Biophys Res Commun. 2018;506:543–7.
Zhuang LK, Xu GP, Pan XR, Lou YJ, Zou QP, Xia D, et al. MicroRNA-181a-mediated downregulation of AC9 protein decreases intracellular cAMP level and inhibits ATRA-induced APL cell differentiation. Cell Death Dis. 2014;5:e1161.
Kilanowska A, Ziółkowska A, Stasiak P, Gibas-Dorna M. cAMP-dependent signaling and ovarian cancer. Cells. 2022;11:3835.
Baiocchi L, Lenci I, Milana M, Kennedy L, Sato K, Zhang W, et al. Cyclic AMP signaling in biliary proliferation: A possible target for cholangiocarcinoma treatment? Cells. 2021;10:1692.
Komiya T, Coxon A, Park Y, Chen WD, Zajac-Kaye M, Meltzer P, et al. Enhanced activity of the CREB co-activator Crtc1 in LKB1 null lung cancer. Oncogene. 2010;29:1672–80.
Sapkota GP, Kieloch A, Lizcano JM, Lain S, Arthur JS, Williams MR, et al. Phosphorylation of the protein kinase mutated in Peutz-Jeghers cancer syndrome, LKB1/STK11, at Ser431 by p90(RSK) and cAMP-dependent protein kinase, but not its farnesylation at Cys(433), is essential for LKB1 to suppress cell vrowth. J Biol Chem. 2001;276:19469–82.
Zheng B, Jeong JH, Asara JM, Yuan YY, Granter SR, Chin L, et al. Oncogenic B-RAF negatively regulates the tumor suppressor LKB1 to promote melanoma cell proliferation. Mol Cell. 2009;33:237–47.
Pearah A, Ramatchandirin B, Liu T, Wolf RM, Ikeda A, Radovick S, et al. Blocking AMPKαS496 phosphorylation improves mitochondrial dynamics and hyperglycemia in aging and obesity. Cell Chem Biol. 2023;30:1585-600.e6.
Alam SK, Wang L, Ren Y, Hernandez CE, Kosari F, Roden AC, et al. ASCL1-regulated DARPP-32 and t-DARPP stimulate small cell lung cancer growth and neuroendocrine tumour cell proliferation. Br J Cancer. 2020;123:819–32.
Rinaldi L, Sepe M, Delle Donne R, Conte K, Arcella A, Borzacchiello D, et al. Mitochondrial AKAP1 supports mTOR pathway and tumor growth. Cell Death Dis. 2017;8:e2842.
Mo J, Deng L, Peng K, Ouyang S, Ding W, Lou L, et al. Targeting STAT3-VISTA axis to suppress tumor aggression and burden in acute myeloid leukemia. J Hematol Oncol. 2023;16:15.
Yan Y, Pan J, Chen Y, Xing W, Li Q, Wang D, et al. Increased dopamine and its receptor dopamine receptor D1 promote tumor growth in human hepatocellular carcinoma. Cancer Commun (Lond). 2020;40:694–710.
Xia S, Ma J, Bai X, Zhang H, Cheng S, Zhang M, et al. Prostaglandin E2 promotes the cell growth and invasive ability of hepatocellular carcinoma cells by upregulating c-Myc expression via EP4 receptor and the PKA signaling pathway. Oncol Rep. 2014;32:1521–30.
Yi H, Wang K, Jin JF, Jin H, Yang L, Zou Y, et al. Elevated adenylyl cyclase 9 expression is a potential prognostic biomarker for patients with colon cancer. Med Sci Monit. 2018;24:19–25.
Pleiman JK, Irving AA, Wang Z, Toraason E, Clipson L, Dove WF, et al. The conserved protective cyclic AMP-phosphodiesterase function PDE4B is expressed in the adenoma and adjacent normal colonic epithelium of mammals and silenced in colorectal cancer. PLOS Genet. 2018;14:e1007611.
Sivaprakasam S, Gurav A, Paschall AV, Coe GL, Chaudhary K, Cai Y, et al. An essential role of Ffar2 (Gpr43) in dietary fibre-mediated promotion of healthy composition of gut microbiota and suppression of intestinal carcinogenesis. Oncogenesis. 2016;5:e238.
Fujishita T, Kojima Y, Kajino-Sakamoto R, Mishiro-Sato E, Shimizu Y, Hosoda W, et al. The cAMP/PKA/CREB and TGFβ/SMAD4 pathways regulate stemness and metastatic potential in colorectal cancer cells. Cancer Res. 2022;82:4179–90.
Steven A, Heiduk M, Recktenwald CV, Hiebl B, Wickenhauser C, Massa C, et al. Colorectal carcinogenesis: connecting K-RAS-induced transformation and CREB activity in vitro and In vivo. Mol Cancer Res. 2015;13:1248–62.
Walia MK, Taylor S, Ho PWM, Martin TJ, Walkley CR. Tolerance to sustained activation of the cAMP/Creb pathway activity in osteoblastic cells is enabled by loss of p53. Cell Death Dis. 2018;9:844.
Safa M, Kazemi A, Zaker F, Razmkhah F. Cyclic AMP-induced p53 destabilization is independent of EPAC in pre-B acute lymphoblastic leukemia cells in vitro. J Recept Signal Transduct Res. 2011;31:256–63.
Naderi EH, Skah S, Ugland H, Myklebost O, Sandnes DL, Torgersen ML, et al. Bone marrow stroma-derived PGE2 protects BCP-ALL cells from DNA damage-induced p53 accumulation and cell death. Mol Cancer. 2015;14:14.
Zandi Z, Kashani B, Alishahi Z, Pourbagheri-Sigaroodi A, Esmaeili F, Ghaffari SH, et al. Dual-specificity phosphatases: therapeutic targets in cancer therapy resistance. J Cancer Res Clin Oncol. 2022;148:57–70.
Gong S, Chen Y, Meng F, Zhang Y, Li C, Zhang G, et al. Roflumilast enhances cisplatin-sensitivity and reverses cisplatin-resistance of ovarian cancer cells via cAMP/PKA/CREB-FtMt signalling axis. Cell Prolif. 2018;51:e12474.
Huang H, Wang Y, Kandpal M, Zhao G, Cardenas H, Ji Y, et al. FTO-dependent N (6)-methyladenosine modifications inhibit ovarian cancer stem cell self-renewal by blocking cAMP signaling. Cancer Res. 2020;80:3200–14.
Ma M, Dai J, Tang H, Xu T, Yu S, Si L, et al. MicroRNA-23a-3p inhibits mucosal melanoma growth and progression through targeting adenylate cyclase 1 and attenuating cAMP and MAPK pathways. Theranostics. 2019;9:945–60.
Carreira S, Goodall J, Denat L, Rodriguez M, Nuciforo P, Hoek KS, et al. Mitf regulation of Dia1 controls melanoma proliferation and invasiveness. Genes Dev. 2006;20:3426–39.
Budillon A, Cereseto A, Kondrashin A, Nesterova M, Merlo G, Clair T, et al. Point mutation of the autophosphorylation site or in the nuclear location signal causes protein kinase A RII beta regulatory subunit to lose its ability to revert transformed fibroblasts. Proc Natl Acad Sci USA. 1995;92:10634–8.
Neary CL, Nesterova M, Cho YS, Cheadle C, Becker KG, Cho-Chung YS. Protein kinase A isozyme switching: eliciting differential cAMP signaling and tumor reversion. Oncogene. 2004;23:8847–56.
Mantovani G, Bondioni S, Lania AG, Rodolfo M, Peverelli E, Polentarutti N, et al. High expression of PKA regulatory subunit 1A protein is related to proliferation of human melanoma cells. Oncogene. 2008;27:1834–43.
Hwang JH, Seo JH, Beshiri ML, Wankowicz S, Liu D, Cheung A, et al. CREB5 promotes resistance to androgen-receptor antagonists and androgen deprivation in prostate cancer. Cell Rep. 2019;29:2355-70.e6.
Hwang JH, Arafeh R, Seo JH, Baca SC, Ludwig M, Arnoff TE, et al. CREB5 reprograms FOXA1 nuclear interactions to promote resistance to androgen receptor-targeting therapies. Elife. 2022;11:e73223.
Patra KC, Kato Y, Mizukami Y, Widholz S, Boukhali M, Revenco I, et al. Mutant GNAS drives pancreatic tumourigenesis by inducing PKA-mediated SIK suppression and reprogramming lipid metabolism. Nat Cell Biol. 2018;20:811–22.
Rao R, Salloum R, Xin M, Lu QR. The G protein Gαs acts as a tumor suppressor in sonic hedgehog signaling-driven tumorigenesis. Cell Cycle. 2016;15:1325–30.
Pan Y, Wang C, Wang B. Phosphorylation of Gli2 by protein kinase A is required for Gli2 processing and degradation and the Sonic Hedgehog-regulated mouse development. Dev Biol. 2009;326:177–89.
Happ JT, Arveseth CD, Bruystens J, Bertinetti D, Nelson IB, Olivieri C, et al. A PKA inhibitor motif within SMOOTHENED controls Hedgehog signal transduction. Nat Struct Mol Biol. 2022;29:990–9.
Feng H, Hu B, Vuori K, Sarkaria JN, Furnari FB, Cavenee WK, et al. EGFRvIII stimulates glioma growth and invasion through PKA-dependent serine phosphorylation of Dock180. Oncogene. 2014;33:2504–12.
Lo HW, Antoun GR, Ali-Osman F. The human glutathione S-transferase P1 protein is phosphorylated and its metabolic function enhanced by the Ser/Thr protein kinases, cAMP-dependent protein kinase and protein kinase C, in glioblastoma cells. Cancer Res. 2004;64:9131–8.
Qiu J, Li Q, Li J, Zhou F, Sang P, Xia Z, et al. Complementary roles of EP2 and EP4 receptors in malignant glioma. Br J Pharmacol. 2023;180:2623–40.
Simko V, Iuliano F, Sevcikova A, Labudova M, Barathova M, Radvak P, et al. Hypoxia induces cancer-associated cAMP/PKA signalling through HIF-mediated transcriptional control of adenylyl cyclases VI and VII. Sci Rep. 2017;7:10121.
Nakayama K. cAMP-response element-binding protein (CREB) and NF-κB transcription factors are activated during prolonged hypoxia and cooperatively regulate the induction of matrix metalloproteinase MMP1. J Biol Chem. 2013;288:22584–95.
Feng J, Yang H, Zhang Y, Wei H, Zhu Z, Zhu B, et al. Tumor cell-derived lactate induces TAZ-dependent upregulation of PD-L1 through GPR81 in human lung cancer cells. Oncogene. 2017;36:5829–39.
Luo M, Zhu J, Ren J, Tong Y, Wang L, Ma S, et al. Lactate increases tumor malignancy by promoting tumor small extracellular vesicles production via the GPR81-cAMP-PKA-HIF-1α axis. Front Oncol. 2022;12:1036543.
Qu J, Sun Z, Peng C, Li D, Yan W, Xu Z, et al. C. tropicalis promotes chemotherapy resistance in colon cancer through increasing lactate production to regulate the mismatch repair system. Int J Biol Sci. 2021;17:2756–69.
Boedtkjer E, Pedersen SF. The acidic tumor microenvironment as a driver of cancer. Annu Rev Physiol. 2020;82:103–26.
Zhong X, He X, Wang Y, Hu Z, Huang H, Zhao S, et al. Warburg effect in colorectal cancer: the emerging roles in tumor microenvironment and therapeutic implications. J Hematol Oncol. 2022;15:160.
Sisignano M, Fischer MJM, Geisslinger G. Proton-sensing GPCRs in health and disease. Cells. 2021;10:2050.
Rowe JB, Kapolka NJ, Taghon GJ, Morgan WM, Isom DG. The evolution and mechanism of GPCR proton sensing. J Biol Chem. 2021;296:100167.
Jing Z, Xu H, Chen X, Zhong Q, Huang J, Zhang Y, et al. The proton-sensing G-protein coupled receptor GPR4 promotes angiogenesis in head and neck cancer. PLOS ONE. 2016;11:e0152789.
Sin WC, Zhang Y, Zhong W, Adhikarakunnathu S, Powers S, Hoey T, et al. G protein-coupled receptors GPR4 and TDAG8 are oncogenic and overexpressed in human cancers. Oncogene. 2004;23:6299–303.
Yu M, Cui R, Huang Y, Luo Y, Qin S, Zhong M. Increased proton-sensing receptor GPR4 signalling promotes colorectal cancer progression by activating the hippo pathway. EBioMedicine. 2019;48:264–76.
Klatt W, Wallner S, Brochhausen C, Stolwijk JA, Schreml S. Expression profiles of proton-sensing G-protein coupled receptors in common skin tumors. Sci Rep. 2020;10:15327.
Harmon C, Robinson MW, Hand F, Almuaili D, Mentor K, Houlihan DD, et al. Lactate-mediated acidification of tumor microenvironment induces apoptosis of liver-resident NK cells in colorectal liver metastasis. Cancer Immunol Res. 2019;7:335–46.
Stolwijk JA, Wallner S, Heider J, Kurz B, Pütz L, Michaelis S, et al. GPR4 in the pH-dependent migration of melanoma cells in the tumor microenvironment. Exp Dermatol. 2023;32:479–90.
Ihara Y, Kihara Y, Hamano F, Yanagida K, Morishita Y, Kunita A, et al. The G protein-coupled receptor T-cell death-associated gene 8 (TDAG8) facilitates tumor development by serving as an extracellular pH sensor. Proc Natl Acad Sci USA. 2010;107:17309–14.
Mori D, Tsujikawa T, Sugiyama Y, Kotani SI, Fuse S, Ohmura G, et al. Extracellular acidity in tumor tissue upregulates programmed cell death protein 1 expression on tumor cells via proton-sensing G protein-coupled receptors. Int J Cancer. 2021;149:2116–24.
Yi M, Niu M, Xu L, Luo S, Wu K. Regulation of PD-L1 expression in the tumor microenvironment. J Hematol Oncol. 2021;14:10.
Vijayan D, Young A, Teng MWL, Smyth MJ. Targeting immunosuppressive adenosine in cancer. Nat Rev Cancer. 2017;17:709–24.
Guieu R, Degioanni C, Fromonot J, De Maria L, Ruf J, Deharo JC, et al. Adenosine, adenosine receptors and neurohumoral syncope: from molecular basis to personalized treatment. Biomedicines. 2022;10:1127.
Magnon C, Hall SJ, Lin J, Xue X, Gerber L, Freedland SJ, et al. Autonomic nerve development contributes to prostate cancer progression. Science. 2013;341:1236361.
Hayakawa Y, Sakitani K, Konishi M, Asfaha S, Niikura R, Tomita H, et al. Nerve growth factor promotes gastric tumorigenesis through aberrant cholinergic signaling. Cancer Cell. 2017;31:21–34.
Peterson SC, Eberl M, Vagnozzi AN, Belkadi A, Veniaminova NA, Verhaegen ME, et al. Basal cell carcinoma preferentially arises from stem cells within hair follicle and mechanosensory niches. Cell Stem Cell. 2015;16:400–12.
Renz BW, Takahashi R, Tanaka T, Macchini M, Hayakawa Y, Dantes Z, et al. β2 adrenergic-neurotrophin feedforward loop promotes pancreatic cancer. Cancer Cell. 2018;34:863–7.
Kamiya A, Hayama Y, Kato S, Shimomura A, Shimomura T, Irie K, et al. Genetic manipulation of autonomic nerve fiber innervation and activity and its effect on breast cancer progression. Nat Neurosci. 2019;22:1289–305.
Vaes N, Idris M, Boesmans W, Alves MM, Melotte V. Nerves in gastrointestinal cancer: from mechanism to modulations. Nat Rev Gastroenterol Hepatol. 2022;19:768–84.
Takahashi R, Ijichi H, Fujishiro M. The role of neural signaling in the pancreatic cancer microenvironment. Cancers (Basel). 2022;14:4269.
Jiang CC, Marsland M, Wang Y, Dowdell A, Eden E, Gao F, et al. Tumor innervation is triggered by endoplasmic reticulum stress. Oncogene. 2022;41:586–99.
Pon CK, Lane JR, Sloan EK, Halls ML. The β2-adrenoceptor activates a positive cAMP-calcium feedforward loop to drive breast cancer cell invasion. Faseb J. 2016;30:1144–54.
Allen JK, Armaiz-Pena GN, Nagaraja AS, Sadaoui NC, Ortiz T, Dood R, et al. Sustained adrenergic signaling promotes intratumoral innervation through BDNF induction. Cancer Res. 2018;78:3233–42.
Tsai YF, Tseng LM, Hsu CY, Yang MH, Chiu JH, Shyr YM. Brain-derived neurotrophic factor (BDNF) -TrKB signaling modulates cancer-endothelial cells interaction and affects the outcomes of triple negative breast cancer. PLoS ONE. 2017;12:e0178173.
Contreras-Zárate MJ, Day NL, Ormond DR, Borges VF, Tobet S, Gril B, et al. Estradiol induces BDNF/TrkB signaling in triple-negative breast cancer to promote brain metastases. Oncogene. 2019;38:4685–99.
Edin ML, Howe AK, Juliano RL. Inhibition of PKA blocks fibroblast migration in response to growth factors. Exp Cell Res. 2001;270:214–22.
Yin S, Song R, Ma J, Liu C, Wu Z, Cao G, et al. Receptor activity-modifying protein 1 regulates mouse skin fibroblast proliferation via the Gαi3-PKA-CREB-YAP axis. Cell Commun Signal. 2022;20:52.
Wójcik-Pszczoła K, Chłoń-Rzepa G, Jankowska A, Ślusarczyk M, Ferdek PE, Kusiak AA, et al. A novel, pan-PDE inhibitor exerts anti-fibrotic effects in human lung fibroblasts via inhibition of TGF-β signaling and activation of cAMP/PKA signaling. Int J Mol Sci. 2020;21:4008.
Togo S, Liu X, Wang X, Sugiura H, Kamio K, Kawasaki S, et al. PDE4 inhibitors roflumilast and rolipram augment PGE2 inhibition of TGF-{beta}1-stimulated fibroblasts. Am J Physiol Lung Cell Mol Physiol. 2009;296:L959–69.
Nieman KM, Romero IL, Van Houten B, Lengyel E. Adipose tissue and adipocytes support tumorigenesis and metastasis. Biochim Biophys Acta. 2013;1831:1533–41.
Kim HS, Jung M, Choi SK, Woo J, Piao YJ, Hwang EH, et al. IL-6-mediated cross-talk between human preadipocytes and ductal carcinoma in situ in breast cancer progression. J Exp Clin Cancer Res. 2018;37:200.
Amemori S, Ootani A, Aoki S, Fujise T, Shimoda R, Kakimoto T, et al. Adipocytes and preadipocytes promote the proliferation of colon cancer cells in vitro. Am J Physiol Gastrointest Liver Physiol. 2007;292:G923–9.
Martini CN, Plaza MV, Vila MC. PKA-dependent and independent cAMP signaling in 3T3-L1 fibroblasts differentiation. Mol Cell Endocrinol. 2009;298:42–7.
Lee HL, Qadir AS, Park HJ, Chung E, Lee YS, Woo KM, et al. cAMP/protein kinase A signaling inhibits Dlx5 expression via activation of CREB and subsequent C/EBPβ induction in 3T3-L1 preadipocytes. Int J Mol Sci. 2018;19:3161.
Liu T, Han C, Fang P, Ma Z, Wang X, Chen H, et al. Cancer-associated fibroblast-specific lncRNA LINC01614 enhances glutamine uptake in lung adenocarcinoma. J Hematol Oncol. 2022;15:141.
Wan X, Guan S, Hou Y, Qin Y, Zeng H, Yang L, et al. FOSL2 promotes VEGF-independent angiogenesis by transcriptionnally activating Wnt5a in breast cancer-associated fibroblasts. Theranostics. 2021;11:4975–91.
Yu T, Yang G, Hou Y, Tang X, Wu C, Wu XA, et al. Cytoplasmic GPER translocation in cancer-associated fibroblasts mediates cAMP/PKA/CREB/glycolytic axis to confer tumor cells with multidrug resistance. Oncogene. 2017;36:2131–45.
Wiley SZ, Sriram K, Liang W, Chang SE, French R, McCann T, et al. GPR68, a proton-sensing GPCR, mediates interaction of cancer-associated fibroblasts and cancer cells. Faseb J. 2018;32:1170–83.
Orange ST, Leslie J, Ross M, Mann DA, Wackerhage H. The exercise IL-6 enigma in cancer. Trends Endocrinol Metab. 2023;34:749–63.
Xu L, Zou C, Zhang S, Chu TSM, Zhang Y, Chen W, et al. Reshaping the systemic tumor immune environment (STIE) and tumor immune microenvironment (TIME) to enhance immunotherapy efficacy in solid tumors. J Hematol Oncol. 2022;15:87.
Cheng X, Wang H, Wang Z, Zhu B, Long H. Tumor-associated myeloid cells in cancer immunotherapy. J Hematol Oncol. 2023;16:71.
Jin H, Lee JS, Kim DC, Ko YS, Lee GW, Kim HJ. Increased extracellular adenosine in radiotherapy-resistant breast cancer cells enhances tumor progression through A2AR-Akt-β-catenin signaling. Cancers (Basel). 2021;13:2105.
Hajizadeh F, Masjedi A, Heydarzedeh Asl S, Karoon Kiani F, Peydaveisi M, Ghalamfarsa G, et al. Adenosine and adenosine receptors in colorectal cancer. Int Immunopharmacol. 2020;87:106853.
Guan S, Suman S, Amann JM, Wu R, Carbone DP, Wang J, et al. Metabolic reprogramming by adenosine antagonism and implications in non-small cell lung cancer therapy. Neoplasia. 2022;32:100824.
Li N, Tang N, Cheng C, Hu T, Wei X, Han W, et al. Improving the anti-solid tumor efficacy of CAR-T cells by inhibiting adenosine signaling pathway. Oncoimmunology. 2020;9:1824643.
Paluskievicz CM, Cao X, Abdi R, Zheng P, Liu Y, Bromberg JS. T regulatory cells and priming the suppressive tumor microenvironment. Front Immunol. 2019;10:2453.
Whiteside TL, Jackson EK. Adenosine and prostaglandin e2 production by human inducible regulatory T cells in health and disease. Front Immunol. 2013;4:212.
Mandapathil M, Szczepanski MJ, Szajnik M, Ren J, Jackson EK, Johnson JT, et al. Adenosine and prostaglandin E2 cooperate in the suppression of immune responses mediated by adaptive regulatory T cells. J Biol Chem. 2010;285:27571–80.
Tanaka A, Sakaguchi S. Targeting Treg cells in cancer immunotherapy. Eur J Immunol. 2019;49:1140–6.
Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017;27:109–18.
Bazhin AV, Kahnert S, Kimpfler S, Schadendorf D, Umansky V. Distinct metabolism of cyclic adenosine monophosphate in regulatory and helper CD4+ T cells. Mol Immunol. 2010;47:678–84.
Kuchen S, Resch W, Yamane A, Kuo N, Li Z, Chakraborty T, et al. Regulation of microRNA expression and abundance during lymphopoiesis. Immunity. 2010;32:828–39.
Anandagoda N, Willis JC, Hertweck A, Roberts LB, Jackson I, Gökmen MR, et al. microRNA-142-mediated repression of phosphodiesterase 3B critically regulates peripheral immune tolerance. J Clin Invest. 2019;129:1257–71.
Vang AG, Housley W, Dong H, Basole C, Ben-Sasson SZ, Kream BE, et al. Regulatory T-cells and cAMP suppress effector T-cells independently of PKA-CREM/ICER: a potential role for Epac. Biochem J. 2013;456:463–73.
Kim HP, Leonard WJ. CREB/ATF-dependent T cell receptor-induced FoxP3 gene expression: a role for DNA methylation. J Exp Med. 2007;204:1543–51.
Klein M, Bopp T. Cyclic AMP represents a crucial component of Treg cell-mediated immune regulation. Front Immunol. 2016;7:315.
Valdebenito S, Barreto A, Eugenin EA. The role of connexin and pannexin containing channels in the innate and acquired immune response. Biochim Biophys Acta Biomembr. 2018;1860:154–65.
Shi L, Feng M, Du S, Wei X, Song H, Yixin X, et al. Adenosine generated by regulatory T cells induces CD8(+) T cell exhaustion in gastric cancer through A2aR pathway. Biomed Res Int. 2019;2019:4093214.
Riccomi A, Gesa V, Sacchi A, De Magistris MT, Vendetti S. Modulation of phenotype and function of human CD4(+)CD25(+) T regulatory lymphocytes mediated by cAMP-elevating agents. Front Immunol. 2016;7:358.
Guereschi MG, Araujo LP, Maricato JT, Takenaka MC, Nascimento VM, Vivanco BC, et al. Beta2-adrenergic receptor signaling in CD4+ Foxp3+ regulatory T cells enhances their suppressive function in a PKA-dependent manner. Eur J Immunol. 2013;43:1001–12.
Ohta A, Gorelik E, Prasad SJ, Ronchese F, Lukashev D, Wong MK, et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc Natl Acad Sci USA. 2006;103:13132–7.
Yano S, Ghosh P, Kusaba H, Buchholz M, Longo DL. Effect of promoter methylation on the regulation of IFN-gamma gene during in vitro differentiation of human peripheral blood T cells into a Th2 population. J Immunol. 2003;171:2510–6.
Vaeth M, Gogishvili T, Bopp T, Klein M, Berberich-Siebelt F, Gattenloehner S, et al. Regulatory T cells facilitate the nuclear accumulation of inducible cAMP early repressor (ICER) and suppress nuclear factor of activated T cell c1 (NFATc1). Proc Natl Acad Sci USA. 2011;108:2480–5.
Jenabian MA, Seddiki N, Yatim A, Carriere M, Hulin A, Younas M, et al. Regulatory T cells negatively affect IL-2 production of effector T cells through CD39/adenosine pathway in HIV infection. PLOS Pathog. 2013;9:e1003319.
Wehbi VL, Taskén K. Molecular mechanisms for cAMP-mediated immunoregulation in T cells-role of anchored protein kinase A signaling units. Front Immunol. 2016;7:222.
Bodor J, Bopp T, Vaeth M, Klein M, Serfling E, Hünig T, et al. Cyclic AMP underpins suppression by regulatory T cells. Eur J Immunol. 2012;42:1375–84.
Tasken K, Ruppelt A. Negative regulation of T-cell receptor activation by the cAMP-PKA-Csk signalling pathway in T-cell lipid rafts. Front Biosci. 2006;11:2929–39.
Kurelic R, Krieg PF, Sonner JK, Bhaiyan G, Ramos GC, Frantz S, et al. Upregulation of phosphodiesterase 2A augments T cell activation by changing cGMP/cAMP cross-talk. Front Pharmacol. 2021;12:748798.
Mosenden R, Taskén K. Cyclic AMP-mediated immune regulation–overview of mechanisms of action in T cells. Cell Signal. 2011;23:1009–16.
Peter D, Jin SL, Conti M, Hatzelmann A, Zitt C. Differential expression and function of phosphodiesterase 4 (PDE4) subtypes in human primary CD4+ T cells: predominant role of PDE4D. J Immunol. 2007;178:4820–31.
Almahariq M, Mei FC, Wang H, Cao AT, Yao S, Soong L, et al. Exchange protein directly activated by cAMP modulates regulatory T-cell-mediated immunosuppression. Biochem J. 2015;465:295–303.
Wang Y, Johnson KCC, Gatti-Mays ME, Li Z. Emerging strategies in targeting tumor-resident myeloid cells for cancer immunotherapy. J Hematol Oncol. 2022;15:118.
Chinn AM, Salmerón C, Lee J, Sriram K, Raz E, Insel PA. PDE4B Is a homeostatic regulator of cyclic AMP in dendritic cells. Front Pharmacol. 2022;13:833832.
Rueda CM, Jackson CM, Chougnet CA. Regulatory T-cell-mediated suppression of conventional T-cells and dendritic cells by different cAMP intracellular pathways. Front Immunol. 2016;7:216.
Ring S, Pushkarevskaya A, Schild H, Probst HC, Jendrossek V, Wirsdörfer F, et al. Regulatory T cell-derived adenosine induces dendritic cell migration through the Epac-Rap1 pathway. J Immunol. 2015;194:3735–44.
Kayhan M, Koyas A, Akdemir I, Savas AC, Cekic C. Adenosine receptor signaling targets both PKA and Epac pathways to polarize dendritic cells to a suppressive phenotype. J Immunol. 2019;203:3247–55.
Fassbender M, Gerlitzki B, Ullrich N, Lupp C, Klein M, Radsak MP, et al. Cyclic adenosine monophosphate and IL-10 coordinately contribute to nTreg cell-mediated suppression of dendritic cell activation. Cell Immunol. 2010;265:91–6.
Ring S, Karakhanova S, Johnson T, Enk AH, Mahnke K. Gap junctions between regulatory T cells and dendritic cells prevent sensitization of CD8(+) T cells. J Allergy Clin Immunol. 2010;125:237–46.
Ohl K, Schippers A, Tenbrock K. CD11c-specific deletion reveals CREB as a critical regulator of DC function during the germinal center response. J Immunol Res. 2018;2018:8947230.
Al-Huseini LM, Aw Yeang HX, Hamdam JM, Sethu S, Alhumeed N, Wong W, et al. Heme oxygenase-1 regulates dendritic cell function through modulation of p38 MAPK-CREB/ATF1 signaling. J Biol Chem. 2014;289:16442–51.
van Vliet SJ, Bay S, Vuist IM, Kalay H, García-Vallejo JJ, Leclerc C, et al. MGL signaling augments TLR2-mediated responses for enhanced IL-10 and TNF-α secretion. J Leukoc Biol. 2013;94:315–23.
Illario M, Giardino-Torchia ML, Sankar U, Ribar TJ, Galgani M, Vitiello L, et al. Calmodulin-dependent kinase IV links Toll-like receptor 4 signaling with survival pathway of activated dendritic cells. Blood. 2008;111:723–31.
Silva-Vilches C, Pletinckx K, Lohnert M, Pavlovic V, Ashour D, John V, et al. Low doses of cholera toxin and its mediator cAMP induce CTLA-2 secretion by dendritic cells to enhance regulatory T cell conversion. PLOS ONE. 2017;12:e0178114.
Lee J, Kim TH, Murray F, Li X, Choi SS, Broide DH, et al. Cyclic AMP concentrations in dendritic cells induce and regulate Th2 immunity and allergic asthma. Proc Natl Acad Sci USA. 2015;112:1529–34.
Datta SK, Sabet M, Nguyen KP, Valdez PA, Gonzalez-Navajas JM, Islam S, et al. Mucosal adjuvant activity of cholera toxin requires Th17 cells and protects against inhalation anthrax. Proc Natl Acad Sci USA. 2010;107:10638–43.
Lee J, Zhang J, Chung YJ, Kim JH, Kook CM, González-Navajas JM, et al. Inhibition of IRF4 in dendritic cells by PRR-independent and -dependent signals inhibit Th2 and promote Th17 responses. Elife. 2020;9:e49416.
Bros M, Montermann E, Cholaszczyńska A, Reske-Kunz AB. The phosphodiesterase 4 inhibitor roflumilast augments the Th17-promoting capability of dendritic cells by enhancing IL-23 production, and impairs their T cell stimulatory activity due to elevated IL-10. Int Immunopharmacol. 2016;35:174–84.
Qian X, Gu L, Ning H, Zhang Y, Hsueh EC, Fu M, et al. Increased Th17 cells in the tumor microenvironment is mediated by IL-23 via tumor-secreted prostaglandin E2. J Immunol. 2013;190:5894–902.
Ji Y, Zhang W. Th17 cells: positive or negative role in tumor? Cancer Immunol Immunother. 2010;59:979–87.
Asadzadeh Z, Mohammadi H, Safarzadeh E, Hemmatzadeh M, Mahdian-Shakib A, Jadidi-Niaragh F, et al. The paradox of Th17 cell functions in tumor immunity. Cell Immunol. 2017;322:15–25.
Huang R, Cioffi J, Berg K, London R, Cidon M, Maayani S, et al. B cell differentiation factor-induced B cell maturation: regulation via reduction in cAMP. Cell Immunol. 1995;162:49–55.
Roper RL, Phipps RP. Prostaglandin E2 and cAMP inhibit B lymphocyte activation and simultaneously promote IgE and IgG1 synthesis. J Immunol. 1992;149:2984–91.
Myklebust JH, Josefsen D, Blomhoff HK, Levy FO, Naderi S, Reed JC, et al. Activation of the cAMP signaling pathway increases apoptosis in human B-precursor cells and is associated with downregulation of Mcl-1 expression. J Cell Physiol. 1999;180:71–80.
Minguet S, Huber M, Rosenkranz L, Schamel WW, Reth M, Brummer T. Adenosine and cAMP are potent inhibitors of the NF-kappa B pathway downstream of immunoreceptors. Eur J Immunol. 2005;35:31–41.
Blois JT, Mataraza JM, Mecklenbraüker I, Tarakhovsky A, Chiles TC. B cell receptor-induced cAMP-response element-binding protein activation in B lymphocytes requires novel protein kinase Cdelta. J Biol Chem. 2004;279:30123–32.
Mambetsariev N, Lin WW, Stunz LL, Hanson BM, Hildebrand JM, Bishop GA. Nuclear TRAF3 is a negative regulator of CREB in B cells. Proc Natl Acad Sci USA. 2016;113:1032–7.
Zorea J, Motro Y, Mazor RD, Carmi YK, Shulman Z, Mahajna J, et al. TRAF3 suppression encourages B cell recruitment and prolongs survival of microbiome-intact mice with ovarian cancer. J Exp Clin Cancer Res. 2023;42:107.
Bangalore-Prakash P, Stunz LL, Mambetsariev N, Whillock AL, Hostager BS, Bishop GA. The oncogenic membrane protein LMP1 sequesters TRAF3 in B-cell lymphoma cells to produce functional TRAF3 deficiency. Blood Adv. 2017;1:2712–23.
Li W, Wang F, Guo R, Bian Z, Song Y. Targeting macrophages in hematological malignancies: recent advances and future directions. J Hematol Oncol. 2022;15:110.
Peters-Golden M. Putting on the brakes: cyclic AMP as a multipronged controller of macrophage function. Sci Signal. 2009;2:pe37.
Aronoff DM, Canetti C, Serezani CH, Luo M, Peters-Golden M. Cutting edge: macrophage inhibition by cyclic AMP (cAMP): differential roles of protein kinase A and exchange protein directly activated by cAMP-1. J Immunol. 2005;174:595–9.
Wall EA, Zavzavadjian JR, Chang MS, Randhawa B, Zhu X, Hsueh RC, et al. Suppression of LPS-induced TNF-alpha production in macrophages by cAMP is mediated by PKA-AKAP95-p105. Sci Signal. 2009;2:28.
Goldsmith M, Avni D, Ernst O, Glucksam Y, Levy-Rimler G, Meijler MM, et al. Synergistic IL-10 induction by LPS and the ceramide-1-phosphate analog PCERA-1 is mediated by the cAMP and p38 MAP kinase pathways. Mol Immunol. 2009;46:1979–87.
Ernst O, Glucksam-Galnoy Y, Bhatta B, Athamna M, Ben-Dror I, Glick Y, et al. Exclusive temporal stimulation of IL-10 expression in LPS-stimulated mouse macrophages by cAMP inducers and type I interferons. Front Immunol. 2019;10:1788.
Ernst O, Glucksam-Galnoy Y, Athamna M, Ben-Dror I, Ben-Arosh H, Levy-Rimler G, et al. The cAMP pathway amplifies early MyD88-dependent and type I interferon-independent LPS-induced interleukin-10 expression in mouse macrophages. Mediators Inflamm. 2019;2019:3451461.
Qian X, Zhang J, Liu J. Tumor-secreted PGE2 inhibits CCL5 production in activated macrophages through cAMP/PKA signaling pathway. J Biol Chem. 2011;286:2111–20.
Polumuri S, Perkins DJ, Vogel SN. cAMP levels regulate macrophage alternative activation marker expression. Innate Immun. 2021;27:133–42.
Luan B, Yoon YS, Le Lay J, Kaestner KH, Hedrick S, Montminy M. CREB pathway links PGE2 signaling with macrophage polarization. Proc Natl Acad Sci USA. 2015;112:15642–7.
Negreiros-Lima GL, Lima KM, Moreira IZ, Jardim BLO, Vago JP, Galvão I, et al. Cyclic AMP regulates key features of macrophages via PKA: recruitment, reprogramming and efferocytosis. Cells. 2020;9:128.
Wu JJ, Yang Y, Peng WT, Sun JC, Sun WY, Wei W. G protein-coupled receptor kinase 2 regulating β2-adrenergic receptor signaling in M2-polarized macrophages contributes to hepatocellular carcinoma progression. Onco Targets Ther. 2019;12:5499–513.
LamersKok N, Panella D, Georgoudaki AM, Liu H, Özkazanc D, Kučerová L, et al. Natural killer cells in clinical development as non-engineered, engineered, and combination therapies. J Hematol Oncol. 2022;15:164.
Whalen MM, Crews JD. Inhibition of a phosphodiesterase III in the lysis-sensitive target-induced elevation of cyclic AMP (cAMP) in human natural killer cells. Biochem Pharmacol. 2000;60:499–506.
Sun Z, Hou D, Liu S, Fu W, Wang J, Liang Z. Norepinephrine inhibits the cytotoxicity of NK92-MI cells via the β2-adrenoceptor/cAMP/PKA/p-CREB signaling pathway. Mol Med Rep. 2018;17:8530–5.
Salinthone S, Schillace RV, Marracci GH, Bourdette DN, Carr DW. Lipoic acid stimulates cAMP production via the EP2 and EP4 prostanoid receptors and inhibits IFN gamma synthesis and cellular cytotoxicity in NK cells. J Neuroimmunol. 2008;199:46–55.
Raskovalova T, Lokshin A, Huang X, Jackson EK, Gorelik E. Adenosine-mediated inhibition of cytotoxic activity and cytokine production by IL-2/NKp46-activated NK cells: involvement of protein kinase A isozyme I (PKA I). Immunol Res. 2006;36:91–9.
Holt D, Ma X, Kundu N, Fulton A. Prostaglandin E(2) (PGE (2)) suppresses natural killer cell function primarily through the PGE(2) receptor EP4. Cancer Immunol Immunother. 2011;60:1577–86.
Martinet L, Jean C, Dietrich G, Fournié JJ, Poupot R. PGE2 inhibits natural killer and gamma delta T cell cytotoxicity triggered by NKR and TCR through a cAMP-mediated PKA type I-dependent signaling. Biochem Pharmacol. 2010;80:838–45.
Chen Z, Yang Y, Neo SY, Shi H, Chen Y, Wagner AK, et al. Phosphodiesterase 4A confers resistance to PGE2-mediated suppression in CD25(+)/CD54(+) NK cells. EMBO Rep. 2021;22:e51329.
Hoskin DW, Mader JS, Furlong SJ, Conrad DM, Blay J. Inhibition of T cell and natural killer cell function by adenosine and its contribution to immune evasion by tumor cells. Int J Oncol. 2008;32:527–35.
Hegde S, Leader AM, Merad M. MDSC: markers, development, states, and unaddressed complexity. Immunity. 2021;54:875–84.
Yu S, Ren X, Li L. Myeloid-derived suppressor cells in hematologic malignancies: two sides of the same coin. Exp Hematol Oncol. 2022;11:43.
Obermajer N, Kalinski P. Generation of myeloid-derived suppressor cells using prostaglandin E2. Transplant Res. 2012;1:15.
Sinha P, Clements VK, Fulton AM, Ostrand-Rosenberg S. Prostaglandin E2 promotes tumor progression by inducing myeloid-derived suppressor cells. Cancer Res. 2007;67:4507–13.
Gabrilovich DI. Myeloid-derived suppressor cells. Cancer Immunol Res. 2017;5:3–8.
Sorrentino C, Miele L, Porta A, Pinto A, Morello S. Myeloid-derived suppressor cells contribute to A2B adenosine receptor-induced VEGF production and angiogenesis in a mouse melanoma model. Oncotarget. 2015;6:27478–89.
Iannone R, Miele L, Maiolino P, Pinto A, Morello S. Blockade of A2b adenosine receptor reduces tumor growth and immune suppression mediated by myeloid-derived suppressor cells in a mouse model of melanoma. Neoplasia. 2013;15:1400–9.
Liu C, Ke P, Zhang J, Zhang X, Chen X. Protein kinase inhibitor peptide as a tool to specifically inhibit protein kinase A. Front Physiol. 2020;11:574030.
Xia Y, Zhan C, Feng M, Leblanc M, Ke E, Yeddula N, et al. Targeting CREB pathway suppresses small cell lung cancer. Mol Cancer Res. 2018;16:825–32.
Luo W, Xu C, Ayello J, Dela Cruz F, Rosenblum JM, Lessnick SL, et al. Protein phosphatase 1 regulatory subunit 1A in ewing sarcoma tumorigenesis and metastasis. Oncogene. 2018;37:798–809.
Na YR, Kwon JW, Kim DY, Chung H, Song J, Jung D, et al. Protein kinase A catalytic subunit is a molecular switch that promotes the pro-tumoral function of macrophages. Cell Rep. 2020;31:107643.
Galski H, Sivan H, Lazarovici P, Nagler A. In vitro and in vivo reversal of MDR1-mediated multidrug resistance by KT-5720: implications on hematological malignancies. Leuk Res. 2006;30:1151–8.
Yang EB, Zhao YN, Zhang K, Mack P. Daphnetin, one of coumarin derivatives, is a protein kinase inhibitor. Biochem Biophys Res Commun. 1999;260:682–5.
Nam G, An SK, Park IC, Bae S, Lee JH. Daphnetin inhibits α-MSH-induced melanogenesis via PKA and ERK signaling pathways in B16F10 melanoma cells. Biosci Biotechnol Biochem. 2022;86:596–609.
Javed M, Saleem A, Xaveria A, Akhtar MF. Daphnetin: A bioactive natural coumarin with diverse therapeutic potentials. Front Pharmacol. 2022;13:993562.
Fan X, Xie M, Zhao F, Li J, Fan C, Zheng H, et al. Daphnetin triggers ROS-induced cell death and induces cytoprotective autophagy by modulating the AMPK/Akt/mTOR pathway in ovarian cancer. Phytomedicine. 2021;82:153465.
Liu C, Pan J, Liu H, Lin R, Chen Y, Zhang C. Daphnetin inhibits the survival of hepatocellular carcinoma cells through regulating Wnt/β-catenin signaling pathway. Drug Dev Res. 2022;83:952–60.
Fukuda H, Nakamura S, Chisaki Y, Takada T, Toda Y, Murata H, et al. Daphnetin inhibits invasion and migration of LM8 murine osteosarcoma cells by decreasing RhoA and Cdc42 expression. Biochem Biophys Res Commun. 2016;471:63–7.
Finn GJ, Creaven BS, Egan DA. Daphnetin induced differentiation of human renal carcinoma cells and its mediation by p38 mitogen-activated protein kinase. Biochem Pharmacol. 2004;67:1779–88.
Li T, Yang G, Hao Q, Zhang X, Zhang X. Daphnetin ameliorates the expansion of chemically induced hepatocellular carcinoma via reduction of inflammation and oxidative stress. J Oleo Sci. 2022;71:575–85.
Hoy JJ, Salinas Parra N, Park J, Kuhn S, Iglesias-Bartolome R. Protein kinase A inhibitor proteins (PKIs) divert GPCR-Gαs-cAMP signaling toward EPAC and ERK activation and are involved in tumor growth. Faseb J. 2020;34:13900–17.
Li K, Liang J, Lin Y, Zhang H, Xiao X, Tan Y, et al. A classical PKA inhibitor increases the oncolytic effect of M1 virus via activation of exchange protein directly activated by cAMP 1. Oncotarget. 2016;7:48443–55.
Parnell E, McElroy SP, Wiejak J, Baillie GL, Porter A, Adams DR, et al. Identification of a novel, small molecule partial agonist for the cyclic AMP sensor, EPAC1. Sci Rep. 2017;7:294.
Beck EM, Parnell E, Cowley A, Porter A, Gillespie J, Robinson J, et al. Identification of a novel class of benzofuran oxoacetic acid-derived ligands that selectively activate cellular EPAC1. Cells. 2019;8:1425.
Almahariq M, Tsalkova T, Mei FC, Chen H, Zhou J, Sastry SK, et al. A novel EPAC-specific inhibitor suppresses pancreatic cancer cell migration and invasion. Mol Pharmacol. 2013;83:122–8.
Wang X, Luo C, Cheng X, Lu M. Lithium and an EPAC-specific inhibitor ESI-09 synergistically suppress pancreatic cancer cell proliferation and survival. Acta Biochim Biophys Sin (Shanghai). 2017;49:573–80.
Qureshi U, Khan MI, Ashraf S, Hameed A, Hafizur RM, Rafique R, et al. Identification of novel Epac2 antagonists through in silico and in vitro analyses. Eur J Pharm Sci. 2020;153:105492.
Courilleau D, Bouyssou P, Fischmeister R, Lezoualc’h F, Blondeau JP. The (R)-enantiomer of CE3F4 is a preferential inhibitor of human exchange protein directly activated by cyclic AMP isoform 1 (Epac1). Biochem Biophys Res Commun. 2013;440:443–8.
Foret-Lucas C, Figueroa T, Bertin A, Bessière P, Lucas A, Bergonnier D, et al. EPAC1 pharmacological inhibition with AM-001 prevents SARS-CoV-2 and influenza a virus replication in cells. Viruses. 2023;15:319.
Kang WB, Yang Q, Guo YY, Wang L, Wang DS, Cheng Q, et al. Analgesic effects of adenylyl cyclase inhibitor NB001 on bone cancer pain in a mouse model. Mol Pain. 2016;12:1744806916652409.
Wang W, Chen QY, Zhao P, Zhong J, Wang Y, Li X, et al. Human safety study of a selective neuronal adenylate cyclase 1 inhibitor NB001 which relieves the neuropathic pain and blocks ACC in adult mice. Mol Pain. 2022;18:17448069221089596.
Brust TF, Alongkronrusmee D, Soto-Velasquez M, Baldwin TA, Ye Z, Dai M, et al. Identification of a selective small-molecule inhibitor of type 1 adenylyl cyclase activity with analgesic properties. Sci Signal. 2017;10:eaah5381.
Kaur J, Soto-Velasquez M, Ding Z, Ghanbarpour A, Lill MA, van Rijn RM, et al. Optimization of a 1,3,4-oxadiazole series for inhibition of Ca(2+)/calmodulin-stimulated activity of adenylyl cyclases 1 and 8 for the treatment of chronic pain. Eur J Med Chem. 2019;162:568–85.
Johann K, Bohn T, Shahneh F, Luther N, Birke A, Jaurich H, et al. Therapeutic melanoma inhibition by local micelle-mediated cyclic nucleotide repression. Nat Commun. 2021;12:5981.
Johannessen CM, Johnson LA, Piccioni F, Townes A, Frederick DT, Donahue MK, et al. A melanocyte lineage program confers resistance to MAP kinase pathway inhibition. Nature. 2013;504:138–42.
Kleinboelting S, Ramos-Espiritu L, Buck H, Colis L, van den Heuvel J, Glickman JF, et al. Bithionol potently inhibits human soluble adenylyl cyclase through binding to the allosteric activator site. J Biol Chem. 2016;291:9776–84.
Ayyagari VN, Diaz-Sylvester PL, Hsieh TJ, Brard L. Evaluation of the cytotoxicity of the Bithionol-paclitaxel combination in a panel of human ovarian cancer cell lines. PLOS ONE. 2017;12:e0185111.
Jakobsen E, Lange SC, Andersen JV, Desler C, Kihl HF, Hohnholt MC, et al. The inhibitors of soluble adenylate cyclase 2-OHE, KH7, and bithionol compromise mitochondrial ATP production by distinct mechanisms. Biochem Pharmacol. 2018;155:92–101.
Wu JF, Dhakal B. BCMA-targeted CAR-T cell therapies in relapsed and/or refractory multiple myeloma: latest updates from 2023 ASCO Annual Meeting. J Hematol Oncol. 2023;16:86.
Kjaergaard J, Hatfield S, Jones G, Ohta A, Sitkovsky M. A(2A) adenosine receptor gene deletion or synthetic A(2A) sntagonist liberate tumor-reactive CD8(+) T cells from tumor-induced immunosuppression. J Immunol. 2018;201:782–91.
Masoumi E, Jafarzadeh L, Mirzaei HR, Alishah K, Fallah-Mehrjardi K, Rostamian H, et al. Genetic and pharmacological targeting of A2a receptor improves function of anti-mesothelin CAR T cells. J Exp Clin Cancer Res. 2020;39:49.
Giuffrida L, Sek K, Henderson MA, Lai J, Chen AXY, Meyran D, et al. CRISPR/Cas9 mediated deletion of the adenosine A2A receptor enhances CAR T cell efficacy. Nat Commun. 2021;12:3236.
Yan T, Zhu L, Chen J. Current advances and challenges in CAR T-Cell therapy for solid tumors: tumor-associated antigens and the tumor microenvironment. Exp Hematol Oncol. 2023;12:14.
Salzillo A, Ragone A, Spina A, Naviglio S, Sapio L. Forskolin affects proliferation, migration and paclitaxel-mediated cytotoxicity in non-small-cell lung cancer cell lines via adenylyl cyclase/cAMP axis. Eur J Cell Biol. 2023;102:151292.
Singh D, Singh P, Pradhan A, Srivastava R, Sahoo SK. Reprogramming cancer stem-like cells with nanoforskolin enhances the efficacy of paclitaxel in targeting breast cancer. ACS Appl Bio Mater. 2021;4:3670–85.
Sapio L, Gallo M, Illiano M, Chiosi E, Naviglio D, Spina A, et al. The natural cAMP elevating compound forskolin in cancer therapy: Is it time? J Cell Physiol. 2017;232:922–7.
Dicitore A, Grassi ES, Caraglia M, Borghi MO, Gaudenzi G, Hofland LJ, et al. The cAMP analogs have potent anti-proliferative effects on medullary thyroid cancer cell lines. Endocrine. 2016;51:101–12.
Wang HM, Qu LQ, Ng JPL, Zeng W, Yu L, Song LL, et al. Natural Citrus flavanone 5-demethylnobiletin stimulates melanogenesis through the activation of cAMP/CREB pathway in B16F10 cells. Phytomedicine. 2022;98:153941.
Ding H, You Q, Li D, Liu Y. 5-Demethylnobiletin: Insights into its pharmacological activity, mechanisms, pharmacokinetics and toxicity. Phytomedicine. 2022;104:154285.
Wu BN, Lin RJ, Lo YC, Shen KP, Wang CC, Lin YT, Chen IJ. KMUP-1, a xanthine derivative, induces relaxation of guinea-pig isolated trachea: the role of the epithelium cyclic nucleotides and K+ channels. Br J Pharmacol. 2004;142:1105–14.
Vandenberghe P, Hagué P, Hockman SC, Manganiello VC, Demetter P, Erneux C, et al. Phosphodiesterase 3A: a new player in development of interstitial cells of Cajal and a prospective target in gastrointestinal stromal tumors (GIST). Oncotarget. 2017;8:41026–43.
Longo J, Pandyra AA, Stachura P, Minden MD, Schimmer AD, Penn LZ. Cyclic AMP-hydrolyzing phosphodiesterase inhibitors potentiate statin-induced cancer cell death. Mol Oncol. 2020;14:2533–45.
Liu D, Mamorska-Dyga A. Syk inhibitors in clinical development for hematological malignancies. J Hematol Oncol. 2017;10:145.
Venkataraman C, Chelvarajan RL, Cambier JC, Bondada S. Interleukin-4 overcomes the negative influence of cyclic AMP accumulation on antigen receptor stimulated B lymphocytes. Mol Immunol. 1998;35:997–1014.
Makhoul S, Trabold K, Gambaryan S, Tenzer S, Pillitteri D, Walter U, et al. cAMP- and cGMP-elevating agents inhibit GPIbα-mediated aggregation but not GPIbα-stimulated Syk activation in human platelets. Cell Commun Signal. 2019;17:122.
Williams CH, Hempel JE, Hao J, Frist AY, Williams MM, Fleming JT, et al. An in vivo chemical genetic screen identifies phosphodiesterase 4 as a pharmacological target for hedgehog signaling inhibition. Cell Rep. 2015;11:43–50.
Schmidt AL, de Farias CB, Abujamra AL, Kapczinski F, Schwartsmann G, Brunetto AL, et al. BDNF and PDE4, but not the GRPR, regulate viability of human medulloblastoma cells. J Mol Neurosci. 2010;40:303–10.
Pullamsetti SS, Banat GA, Schmall A, Szibor M, Pomagruk D, Hänze J, et al. Phosphodiesterase-4 promotes proliferation and angiogenesis of lung cancer by crosstalk with HIF. Oncogene. 2013;32:1121–34.
He N, Kim N, Song M, Park C, Kim S, Park EY, et al. Integrated analysis of transcriptomes of cancer cell lines and patient samples reveals STK11/LKB1-driven regulation of cAMP phosphodiesterase-4D. Mol Cancer Ther. 2014;13:2463–73.
Ge X, Milenkovic L, Suyama K, Hartl T, Purzner T, Winans A, et al. Phosphodiesterase 4D acts downstream of Neuropilin to control Hedgehog signal transduction and the growth of medulloblastoma. Elife. 2015;4:e07068.
Powers GL, Hammer KD, Domenech M, Frantskevich K, Malinowski RL, Bushman W, et al. Phosphodiesterase 4D inhibitors limit prostate cancer growth potential. Mol Cancer Res. 2015;13:149–60.
Mishra RR, Belder N, Ansari SA, Kayhan M, Bal H, Raza U, et al. Reactivation of cAMP pathway by PDE4D inhibition represents a novel druggable axis for overcoming tamoxifen resistance in ER-positive breast cancer. Clin Cancer Res. 2018;24:1987–2001.
McEwan DG, Brunton VG, Baillie GS, Leslie NR, Houslay MD, Frame MC. Chemoresistant KM12C colon cancer cells are addicted to low cyclic AMP levels in a phosphodiesterase 4-regulated compartment via effects on phosphoinositide 3-kinase. Cancer Res. 2007;67:5248–57.
Kim SW, Rai D, McKeller MR, Aguiar RC. Rational combined targeting of phosphodiesterase 4B and SYK in DLBCL. Blood. 2009;113:6153–60.
Kim DU, Kwak B, Kim SW. Phosphodiesterase 4B is an effective therapeutic target in colorectal cancer. Biochem Biophys Res Commun. 2019;508:825–31.
Limoncella S, Lazzaretti C, Paradiso E, D’Alessandro S, Barbagallo F, Pacifico S, et al. Phosphodiesterase (PDE) 5 inhibitors sildenafil, tadalafil and vardenafil impact cAMP-specific PDE8 isoforms-linked second messengers and steroid production in a mouse Leydig tumor cell line. Mol Cell Endocrinol. 2022;542:111527.
Klutzny S, Anurin A, Nicke B, Regan JL, Lange M, Schulze L, et al. PDE5 inhibition eliminates cancer stem cells via induction of PKA signaling. Cell Death Dis. 2018;9:192.
Pantziarka P, Verbaanderd C, Sukhatme V, Rica Capistrano I, Crispino S, Gyawali B, et al. ReDO_DB: the repurposing drugs in oncology database. Ecancermedicalscience. 2018;12:886.
Cruz-Burgos M, Losada-Garcia A, Cruz-Hernández CD, Cortés-Ramírez SA, Camacho-Arroyo I, Gonzalez-Covarrubias V, et al. New approaches in oncology for repositioning drugs: the case of PDE5 inhibitor sildenafil. Front Oncol. 2021;11:627229.
Li Q, Shu Y. Pharmacological modulation of cytotoxicity and cellular uptake of anti-cancer drugs by PDE5 inhibitors in lung cancer cells. Pharm Res. 2014;31:86–96.
Roberts JL, Booth L, Conley A, Cruickshanks N, Malkin M, Kukreja RC, et al. PDE5 inhibitors enhance the lethality of standard of care chemotherapy in pediatric CNS tumor cells. Cancer Biol Ther. 2014;15:758–67.
Röhrig T, Pacjuk O, Hernández-Huguet S, Körner J, Scherer K, Richling E. Inhibition of cyclic adenosine monophosphate-specific phosphodiesterase by various food plant-derived phytotherapeutic agents. Medicines (Basel). 2017;4:80.
Noori S, Hassan ZM, Rezaei B, Rustaiyan A, Habibi Z, Fallahian F. Artemisinin can inhibit the calmodulin-mediated activation of phosphodiesterase in comparison with Cyclosporin A. Int Immunopharmacol. 2008;8:1744–7.
Zhang J, Xu HX, Zhu JQ, Dou YX, Xian YF, Lin ZX. Natural Nrf2 inhibitors: A review of their potential for cancer treatment. Int J Biol Sci. 2023;19:3029–41.
Chen YH, Wu JX, Yang SF, Hsiao YH. Synergistic combination of luteolin and asiatic acid on cervical cancer in vitro and in vivo. Cancers (Basel). 2023;15:548.
Furlan V, Bren U. Protective effects of [6]-gingerol against chemical carcinogens: mechanistic insights. Int J Mol Sci. 2020;21:695.
Wang J, Zhang J, Guo Z, Hua H, Zhang H, Liu Y, et al. Targeting HSP70 chaperones by rhein sensitizes liver cancer to artemisinin derivatives. Phytomedicine. 2024;122:155156.
Zou X, Shanmugam SK, Kanner SA, Sampson KJ, Kass RS, Colecraft HM. Divergent regulation of KCNQ1/E1 by targeted recruitment of protein kinase A to distinct sites on the channel complex. Elife. 2023;12:e83466.
Ireton KE, Xing X, Kim K, Weiner JC, Jacobi AA, Grover A, et al. Regulation of the Ca(2+) channel Ca(V)1.2 supports spatial memory and its flexibility and LTD. J Neurosci. 2023;43:5559–73.
Honda T, Fujiyama T, Miyoshi C, Ikkyu A, Hotta-Hirashima N, Kanno S, et al. A single phosphorylation site of SIK3 regulates daily sleep amounts and sleep need in mice. Proc Natl Acad Sci USA. 2018;115:10458–63.
Wang XT, Zhou L, Dong BB, Xu FX, Wang DJ, Shen EW, et al. cAMP-EPAC-PKCε-RIM1α signaling regulates presynaptic long-term potentiation and motor learning. Elife. 2023;12:e80875.
Murray AJ. Pharmacological PKA inhibition: all may not be what it seems. Sci Signal. 2008;1:re4.
Gao F, Yang S, Wang J, Zhu G. cAMP-PKA cascade: An outdated topic for depression? Biomed Pharmacother. 2022;150:113030.
Baillie GS, Tejeda GS, Kelly MP. Therapeutic targeting of 3’,5’-cyclic nucleotide phosphodiesterases: inhibition and beyond. Nat Rev Drug Discov. 2019;18:770–96.
Kelly K, Mejia A, Suhasini AN, Lin AP, Kuhn J, Karnad AB, et al. Safety and pharmacodynamics of the PDE4 inhibitor roflumilast in advanced B-cell malignancies. Clin Cancer Res. 2017;23:1186–92.
Kim DY, Nam J, Chung JS, Kim SW, Shin HJ. Role of roflumilast combined with ESHAP chemotherapy in relapsed/refractory patients with diffuse large B-cell lymphoma. Cancer Res Treat. 2022;54:301–13.
Dixit D, Prager BC, Gimple RC, Miller TE, Wu Q, Yomtoubian S, et al. Glioblastoma stem cells reprogram chromatin in vivo to generate selective therapeutic dependencies on DPY30 and phosphodiesterases. Sci Transl Med. 2022;14:eabf3917.
Weed DT, Zilio S, Reis IM, Sargi Z, Abouyared M, Gomez-Fernandez CR, et al. The reversal of immune exclusion mediated by tadalafil and an anti-tumor vaccine also induces PDL1 upregulation in recurrent head and neck squamous cell carcinoma: interim analysis of a phase I clinical trial. Front Immunol. 2019;10:1206.
Luginbuhl AJ, Johnson JM, Harshyne LA, Linnenbach AJ, Shukla SK, Alnemri A, et al. Tadalafil enhances immune signatures in response to neoadjuvant nivolumab in resectable head and neck squamous cell carcinoma. Clin Cancer Res. 2022;28:915–27.
Booth L, Roberts JL, Rais R, Cutler RE Jr, Diala I, Lalani AS, et al. Neratinib augments the lethality of [regorafenib + sildenafil]. J Cell Physiol. 2019;234:4874–87.
Sutton SS, Magagnoli J, Cummings TH, Hardin JW. The association between phosphodiesterase-5 inhibitors and colorectal cancer in a national cohort of patients. Clin Transl Gastroenterol. 2020;11:e00173.
Zhang Y, Lo CH, Giovannucci EL. Phosphodiesterase 5 inhibitor use and risk of conventional and serrated precursors of colorectal cancer. Cancer Epidemiol Biomarkers Prev. 2021;30:419–21.
Huang W, Sundquist J, Sundquist K, Ji J. Phosphodiesterase-5 inhibitors use and risk for mortality and metastases among male patients with colorectal cancer. Nat Commun. 2020;11:3191.
Ren Y, Zheng J, Yao X, Weng G, Wu L. Essential role of the cGMP/PKG signaling pathway in regulating the proliferation and survival of human renal carcinoma cells. Int J Mol Med. 2014;34:1430–8.
Pusztai L, Zhen JH, Arun B, Rivera E, Whitehead C, Thompson WJ, et al. Phase I and II study of exisulind in combination with capecitabine in patients with metastatic breast cancer. J Clin Oncol. 2003;21:3454–61.
Witta SE, Gustafson DL, Pierson AS, Menter A, Holden SN, Basche M, et al. A phase I and pharmacokinetic study of exisulind and docetaxel in patients with advanced solid tumors. Clin Cancer Res. 2004;10:7229–37.
Califano JA, Khan Z, Noonan KA, Rudraraju L, Zhang Z, Wang H, et al. Tadalafil augments tumor specific immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res. 2015;21:30–8.
Weed DT, Vella JL, Reis IM, De la Fuente AC, Gomez C, Sargi Z, et al. Tadalafil reduces myeloid-derived suppressor cells and regulatory T cells and promotes tumor immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res. 2015;21:39–48.
Rüland L, Andreatta F, Massalini S, de Sousa Lopes SC, Clevers H, Hendriks D, et al. Organoid models of fibrolamellar carcinoma mutations reveal hepatocyte transdifferentiation through cooperative BAP1 and PRKAR2A loss. Nat Commun. 2023;14:2377.
Jiang Y, Zhang H, Wang J, Chen J, Guo Z, Liu Y, et al. Exploiting RIG-I-like receptor pathway for cancer immunotherapy. J Hematol Oncol. 2023;16:8.
Yan BR, Zhou L, Hu MM, Li M, Lin H, Yang Y, et al. PKACs attenuate innate antiviral response by phosphorylating VISA and priming it for MARCH5-mediated degradation. PLOS Pathog. 2017;13:e1006648.
Grandoch M, de Jesús ML, Oude Weernink PA, Weber AA, Jakobs KH, Schmidt M. B cell receptor-induced growth arrest and apoptosis in WEHI-231 immature B lymphoma cells involve cyclic AMP and Epac proteins. Cell Signal. 2009;21:609–21.
Sasi B, Ethiraj P, Myers J, Lin AP, Jiang S, Qiu Z, et al. Regulation of PD-L1 expression is a novel facet of cyclic-AMP-mediated immunosuppression. Leukemia. 2021;35:1990–2001.
Wu M, Huang Q, Xie Y, Wu X, Ma H, Zhang Y, et al. Improvement of the anticancer efficacy of PD-1/PD-L1 blockade via combination therapy and PD-L1 regulation. J Hematol Oncol. 2022;15:24.
Acknowledgements
Not applicable.
Funding
The work in the authors’ lab was supported by Sichuan Natural Science Foundation (Grant No. 2023NSFSC0737).
Author information
Authors and Affiliations
Contributions
YJ, HH and HZ conceived the review and wrote the main manuscript text. HZ and JW prepared the figures in this manuscript. JL, JC and YL prepared the tables and revised the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
YJ is an editorial board member of J Hematol Oncol. Other authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, H., Liu, Y., Liu, J. et al. cAMP-PKA/EPAC signaling and cancer: the interplay in tumor microenvironment. J Hematol Oncol 17, 5 (2024). https://doi.org/10.1186/s13045-024-01524-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13045-024-01524-x