
Abstract
Background
Animal models are widely used to study pathological processes and drug (side) effects in a controlled environment. There is a wide variety of methods available for establishing animal models depending on the research question. Commonly used methods in tumor research include xenografting cells (established/commercially available or primary patient-derived) or whole tumor pieces either orthotopically or heterotopically and the more recent genetically engineered models—each type with their own advantages and disadvantages. The current systematic review aimed to investigate the meningioma model types used, perform a meta-analysis on tumor take rate (TTR), and perform critical appraisal of the included studies. The study also aimed to assess reproducibility, reliability, means of validation and verification of models, alongside pros and cons and uses of the model types.
Methods
We searched Medline, Embase, and Web of Science for all in vivo meningioma models. The primary outcome was tumor take rate. Meta-analysis was performed on tumor take rate followed by subgroup analyses on the number of cells and duration of incubation. The validity of the tumor models was assessed qualitatively. We performed critical appraisal of the methodological quality and quality of reporting for all included studies.
Results
We included 114 unique records (78 using established cell line models (ECLM), 21 using primary patient-derived tumor models (PTM), 10 using genetically engineered models (GEM), and 11 using uncategorized models). TTRs for ECLM were 94% (95% CI 92–96) for orthotopic and 95% (93–96) for heterotopic. PTM showed lower TTRs [orthotopic 53% (33–72) and heterotopic 82% (73–89)] and finally GEM revealed a TTR of 34% (26–43).
Conclusion
This systematic review shows high consistent TTRs in established cell line models and varying TTRs in primary patient-derived models and genetically engineered models. However, we identified several issues regarding the quality of reporting and the methodological approach that reduce the validity, transparency, and reproducibility of studies and suggest a high risk of publication bias. Finally, each tumor model type has specific roles in research based on their advantages (and disadvantages).
Systematic review registration: PROSPERO-ID CRD42022308833.
Similar content being viewed by others

Introduction
Meningiomas account for 40% of all primary intracranial tumors [1] and are primarily benign (90–95%). They display as a group a heterogenous epigenetic/genetic profiles [2,3,4,5,6,7]. The preferred treatment for a symptomatic and/or growing meningioma is surgery [8, 9]. Pharmaceutical therapies are primarily used pre-clinically or in protocolled trials and have so far had limited success in humans [2]. In vivo research is thus paramount.
While a perfectly designed tumor animal model is a utopian thought, we can strive towards a model that resembles the actual tumor as closely as possible. Scientific advances over the last 70 years have led to huge progress in the field of animal models, including models for meningioma. The first successes in establishing a meningioma model via xenotransplantation occurred in the 1940s and 50 s [10, 11], and studies over the next 30 years have used a variety of approaches [12,13,14,15,16,17,18,19].
The availability of immunodeficient animals and established immortalized cell-lines (i.e. IOMM-Lee, CH-157, and BEN-MEN-1) has revolutionized the field in terms of tumor take rate (TTR) (which is close to 100% in many cases) and has led to stable models for testing new treatment options [20,21,22]. The use of immortalized cells provides a needed weapon against senescence, but at the same time it limits conclusions as to treatment effects, and many pharmaceutical successes from animal studies have proven ineffective in human clinical trials [23]. The use of primary cell models without the use of immortalization provides a patient-specific model for more targeted therapies. However, these approaches lack immune system interaction and show varying TTRs, especially for benign primary tumors [24,25,26]. The first genetically engineered (meningioma) model (GEM) emerged twenty years ago using a conditional knockout Nf2Flox2/Flox2 via recombinant high-titer adenovirus expressing Cre recombinase (AdCre) to ensure growth [27], with a few papers (primarily from the same group) replicating the results [28,29,30,31]. Although GEM provides a solution to the significant problem of adaptive immune system involvement, its low TTRs and long tumor induction time limit the model’s use in treatment studies.
So far, a few narrative reviews [32,33,34] and a systematic review have been conducted on the topic [35]. However, the systematic review did not entail a search through multiple literature platforms and did not assess methodological quality/risk of bias. In the current paper, we present the first systematic review and meta-analysis of meningioma animal models that includes an assessment of the quality of reporting and the methodological quality of the included studies.
The overall aim of the review was to investigate which method of tumor development/growth—established patient-derived cell lines (orthotopic and heterotopic), primary patient-derived tumor cells/material (orthotopic and heterotopic), or genetically modified animals—provides the best tumor take rate and at what duration of incubation. The specific objectives were firstly, to search the literature for all available research regarding the different models and to assess their reproducibility and reliability, advantages, and disadvantages. Secondly, to determine how the models should be verified and which modalities are necessary to compare xenograft or genetically modified tumors to the parent tumor. Thirdly, to present possible future aspects of meningioma animal models in relation to optimal tests of future therapeutics in human meningiomas. Fourthly, to identify and analyze knowledge gaps that could help identify future research initiatives.
Methods
Protocol and registration
This systematic review was performed in accordance with the updated 2020 Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [36]. The PRISMA checklist and PRISMA abstract checklists are provided in the Additional files 1 and 2). The review was prospectively registered with PROSPERO at https://www.crd.york.ac.uk/PROSPERO/ as an animal study (CRD42022308833) prior to first full-text screening on February 14, 2022. The full original protocol that was uploaded to PROSPERO is available in the Additional file 3. The minor changes to the original uploaded protocol are reported in the Additional file 4.
Eligibility criteria
We sought to include all original records published in peer-reviewed journals with full texts that had the Population and Outcome (PO) of in vivo experiments with the intention of meningioma growth (population) and the corresponding meningioma growth rate and induction duration (outcome).
We excluded review records, systematic reviews, human studies, and conference abstracts as well as records describing spontaneous meningiomas in animals without the use of genetic modification specifically aimed at meningiomas. Models that were not meningioma models were excluded. No restrictions were applied to study design as long as the record described an in vivo experiment aiming for meningioma growth. We included records in any language if an English title or abstract was available.
Information sources and search strategy
We searched the following electronic databases on June 18, 2021, and again on August 8, 2022, (the latter with the limitations set to 2021–2022): Medline, Embase, and Web of Science. The search strategy was reviewed by a research librarian at the University of Southern Denmark. Search terms were sourced from already published papers that we were familiar with, e.g. meningioma animal model, xenograft, and genetically engineered model. The full original search strategy is available in the Additional file 5.
Study selection
All papers extracted via the search string were screened by title and abstract by two reviewers (MSA and MSK) in a blinded fashion. During the first round of screening (title and abstract), all papers that were deemed eligible by either of the authors were included for the second round (full-text screening). Cohen’s Kappa index [37] was performed to assess initial screening agreement. The first 200 papers were rated un-blinded to adjust the screening method. Full-text screening was performed in a blinded fashion by two reviewers (MSA and MSK), with disagreements being settled by discussion with a senior author (FRP).
Papers were divided into four categories: established cell line models (ECLM), primary patient-derived tumor models (PTM), genetically engineered models (GEM), and other models not fitting into the previous three categories (uncategorized). Papers eligible for inclusion but older than 40 years were ultimately excluded due lack of relevance. Records containing more than one animal model type were included in all relevant categories.
Data extraction fields
A pre-defined data extraction sheet was developed for each model type. Data were extracted independently by two reviewers (MSA and MSK) in a blinded fashion. Data were extracted on record meta-data, animal data, and tumor model characteristics. The data extraction sheet (including a description of the fields) is available in the Additional file 6.
Synthesis of results and summary measures
All analyses were conducted using the freely available software R (https://www.r-project.org/). We performed random-effect meta-analyses on the proportion of animals that developed tumors (TTR) for five model types: ECLM (orthotopic and heterotopic separately), PTM (orthotopic and heterotopic separately), and GEM. Meta-analyses used the metaprop function from the meta package.
Heterogeneity was assessed by visual inspection of forest plots and by using the I2 measure as recommended in the Cochrane Handbook [38]. If heterogeneity was identified, it was examined through subgroup analyses. The following characteristics were examined as potential sources of heterogeneity in xenografted models: number of cells, cell concentration, established cell line used, parent tumor grade (WHO grade 1–3), and duration of incubation. For GEMs, the following characteristics were examined in subgroup analyses: genomic lesion type, method of gaining lesion, incubation time.
Due to the risk of confounding, all subgroup analyses were considered exploratory and were interpreted cautiously. Survival studies on duration of incubation were excluded from the meta-analyses (apart from GEMs, which were mainly survival studies) and commented on narratively. The validity of different tumor models and their relation to human tumors were discussed narratively, including a discussion of the methodological quality of relevant trials. The results of the mixed category ‘uncategorized’, which contains various tumor models, were presented narratively due to significant heterogeneity.
Critical appraisal of quality of reporting, methodological quality, and risk of bias
As no validated tool exists for the critical assessment of the types of records included in this review, we developed a critical appraisal tool (Critical appraisal of methodological quality and quality of reporting—CRIME-Q) [39] for use in this review and other animal reviews in the future. CRIME-Q is inspired primarily by Macleod et al. [40] and the ARRIVE 2.0 guidelines for Animal Research: Reporting of In Vivo experiments [41], but it also includes items from the recommended and validated SYRCLE’s Risk of Bias tool [42].
Two reviewers (MSA and MSK) individually used CRIME-Q to assess all included studies. Any discrepancies were solved via a third reviewer (FRP). The domains included in CRIME-Q are described in Table 1. We summarized the overall quality of included studies narratively and included this in the interpretation of our results. A full table of all assessed studies are available in the Additional file 7. A predetermined list of information needed to obtain the various grades of Yes/No/Partly/Unclear/Not applicable (NA) is available in the Additional file 8.
Results
Study selection
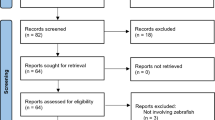
The search strategy yielded 2175 unique studies, of which 151 were potentially eligible. The kappa index for the two reviewers at the screening stage of the first round was 0.9. One record was initially screened negative but was added through other literature alongside two other studies. Of the 117 studies that met the inclusion criteria, three were discarded due to retraction from the journal. The top-up search conducted in August 2022 led to the inclusion of 11 additional studies. We identified 78 studies on ECLM [6, 20,21,22, 24, 43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115], 21 studies on PTM [20, 24,25,26, 73, 106, 116,117,118,119,120,121,122,123,124,125,126,127,128,129], 10 studies on GEM [27,28,29,30,31, 130,131,132,133,134], and 11 studies categorized as “uncategorized” [30, 132, 135,136,137,138,139,140,141,142,143]. We decided to exclude studies older than 40 years due to lack of relevance. The PRISMA flow diagram for the study selection process is presented in Fig. 1. The original data extraction sheets for ECLM, PTM, GEM, and uncategorized studies are available in the Additional files 9, 10, 11, 12).

Flow chart. Established cell line models (ECLM), primary tumor models (PTM), genetically engineered models (GEM). *One record was initially screened negative on the abstract, and the other two studies were found through other literature (one as a historical perspective and the other as an ECLM)
Primary analysis
We conducted meta-analyses on the tumor take rate (TTR) for the five model groups. The overall TTR was 95% (95% CI 93–96%) for ECLM heterotopic models and 94% (92–96%) for ECLM orthotopic models, 82% (73–89%) for PTM heterotopic models and 53% (33–72%) for PTM orthotopic models, and 34% (26–43%) for GEM. While no statistical heterogeneity was found for the ECLM (I2 = 0%), the other meta-analyses showed substantial heterogeneity with I2 over 50%. Forest plots for these meta-analyses are available in the Additional file 13.
Subgroup analyses
Table 2 shows the results of subgroup analyses of TTR against duration of incubation, number of cells, injection volume, cell line and WHO grade, where significant subgroup interactions were found for at least one of the model types. The full results of subgroup analyses, including forest plots, can be found in the Additional file 13.
Meningioma animal models
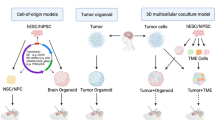
There is a plethora of methods available for creating a meningioma animal model depending on the research question. Commonly used methods include xenografting cells (established/commercially available or primary patient-derived) or whole tumor pieces either orthotopically or heterotopically and the more recent genetically engineered models (Fig. 2).

Overview of the most common model types. Both ECLM (established cell line models) and PTM (primary patient-derived tumor models) require immunocompromised hosts. PTM include cell injection models and whole tumor ‘PDX’ models. Syngeneic modelling is achievable in immunocompetent hosts. RCAS/TVA: Replication-competent avian leukosis virus splice-acceptor/tumor virus A. Illustrator: Mikkel Schou Andersen
Patient-derived xenotransplantation and xenografts
Patient-derived xenotransplantation is the transplantation of tissue foreign to the host e.g., where meningioma tissue or cells derived from patients are transplanted into small animals such as immunocompromised mice. As a minimum, the animals need to have absent mature T-cells such as the BALB/c nude, Athymic nude, CD-1 nude, NMRI nude, NU/NU, and Swiss nude mice or the severe combined immune-deficient animals such as NSG, NRG and NOD SCID, and SCID mice that lack T cells and B-cells and an innate immune system [144]. Most studies use animals up to 10 weeks old, with some exceptions (12–15 [62], 10–22 [53], 7–11 [62], and 12–16 [25] weeks old).
The two main types of models used are the orthotopic models and the heterotopic models. An overview of these models is provided in Table 4 (for ECLM) and Table 5 (for PTM). Further details are available in the Additional files 9, 10, 11).
The orthotopic model requires implantation of material intracranially, most commonly through a burr hole in the frontal region of the skull (typically 1–3 mm anterior and 1–3 mm right of bregma) either superficially or at the skull base. All the reviewed orthotopic models inject between 0.5–10 μl of volume, with 2–10 μl being most common without noteworthy issues. Heterotopic models have a higher injection volume (commonly 100–500 μl), and a larger number of cells are needed to obtain a large tumor. Thus, while orthotopic models typically use 105–106 cells, most of the heterotopic models inject > 106–107 depending on the type of cells used. Since there are potential big differences in TTR between the immortalized cell line models and the primary patient-derived cell/tissue models, we have chosen to distinguish between the two in the sections below.
Established/commercially available cell line models (ECLM)
Meningioma established patient-derived cell lines have been used for decades for in vitro and in vivo research. Most commonly used is the Nf2 + , malignant, IOMM-Lee, which was established from an intraosseous malignant meningioma from 1990 [22]. Other noteworthy cell lines include the Nf2−, benign BEN-MEN-1, which was hTERT- immortalized [21], the malignant KT21 [100] with heterozygous loss of Nf2, and the malignant CH-157, whose origin remains a mystery [20, 145, 146]. Other cell lines have been produced for various purposes. Some of these have complex karyotypes such as KCI-MENG1 [105] and SF3061 [109], which is also Nf2+, or the MeTSC, which is Nf2− [69]. Others have a simple karyotype such as SF4433 [147], and BEN-MEN-1 [20]. Table 3 shows all established/commercially available meningioma cell lines used in vivo including both the origin paper and papers in which the cell lines have been used.
Over the last 40 years of research, about 70% of meningioma studies have used ECLM (Table 4). The most commonly used, IOMM-Lee, shows a high tumor take rate in subgroup analysis for both orthotopic models 87% (95% CI 95–98%) and heterotopic models 94% (90–96%), and it shows median survival of 10–27 days in orthotopic models, depending on the number of cells. In general, 104–2.5 × 105 cells are needed for orthotopic and heterotopic ECLM (Table 4). CH-157 has a TTR of 89% (81–94%) in orthotopic models and 97% (90–96%) in heterotopic models and a similar median survival of 12–24 days in orthotopic models with cell concentrations of 104–106 cells. The most used benign cell type is the BEN-MEN-1, which has only been studied in orthotopic models (TTR of 97% (81–100%); using 0.5–1.0 × 106 cells, researchers have created a long-term model (Ki67 < 1%) up to 180 days [21]. (For further details, see Additional file 9).
Primary patient-derived tumor models (PTM)
The non-immortalized patient-derived models are comprised of all studies describing implantation either straight from surgery as cells or whole tumor pieces [25, 106, 122, 125, 127] or after fewer than 10 passages (usually after 3–6 passages) [24, 26, 67, 72, 118,119,120,121, 123, 124]. For orthotopic models, subgroup analyses (Table 2) showed varying TTRs ≤ 50% with a benign model TTR of 47% (95% CI 17–79%), atypical tumor model TTR of 36% (7–81%), and malignant tumor model TTR of 50% (18–82%) (see Table 5 for individual TTR). McCutcheon et al. [24] tested various durations for different WHO tumor grades and showed succesful tumor take time of 21–56 days for benign, 14–28 days for atypical, and 4–12 for malignant. Zhang et al. [25] performed survival studies on atypical tumors up to 240 days and malignant tumors up to 160 days. In general, successful benign models required 106 cells, atypical models 105–106 cells, and malignant down to 103 cells.
For heterotopic models, subgroup analyses showed TTRs for benign tumor models of 88% (95% CI 78–94%), atypical tumor models 59% (9–95%), and malignant tumor models 75% (51–89%) (Table 2). Benign models have been more often used and show more consistent results with 105–5 × 107 cells and duration of incubation ranging from one month to almost one year. Duration of incubation for atypical tumors is up to 180 days [122] while that for malignant tumors is significantly shorter ranging from 30 to 56 days [106, 118, 124, 129].
Some studies in both the orthotopic and heterotopic groups do not describe WHO grade or tumor subtype, and thus valuable information is difficult to obtain [116, 117, 125, 127] (for further details, see Additional file 10).
Strengths and weaknesses of xenografting to immune-incompetent/compromised animals
The advantages of xenografting material or cells into immunocompromised animals are the lower cost and higher availability compared to for instance GEM. The strength of the orthotopic models is that tumors grow in the appropriate microenvironment (without taking alterations of the immune system into account), making them suitable for drug testing. The heterotopic models are performed outside of the central nervous tissue most commonly via a flank/subcutaneous injection model and are the easiest model to set up and perform. Subcutaneous injections of meningioma cells have been successful both with and without the use of Matrigel (which forms a solid gel at 37 °C to keep cells close together during tumor development [170]) or fibrin clots (for further details, see Additional file 9, 10, 11, 12). There are concerns, however, that Matrigel enhances tumorigenicity or even modulates characteristics of the original tumor. It may even increase drug resistance in vivo [171] and might transform pre-malignant to malignant cells [172]. The translatability of heterotopic models is debatable [32].
Meningioma established cell lines have been used for decades for in vitro and in vivo research. Some cell lines (such as IOMM-Lee, BEN-MEN-1, and CH-157) have the advantage of being thoroughly characterized on every level from histology to genetic profiles [20, 149]. The use of established cell lines produces consistent and homogeneous results across studies. Using established (immortalized) cell lines also negates the great issue of senescence that is often experienced in patient-derived primary cells (non-immortalized). A drawback of immortalized cell lines is that they are very far away from human meningioma conditions, e.g. IOMM-Lee’s complex karyotype probably due to long-term culturing [114]. ECLMs do not display normal meningioma pathology, disease nature, or heterogeneity, which in general makes them unsuitable for pharmaceutical studies.
In contrast to established cell lines, the primary patient-derived non-immortalized models show varying degree of tumor take within and between studies. Zhang et al. [25] reported TTRs ranging from 0% for most of the tumors to 90% (for a malignant tumor), and none of their xenografted benign cell lines could be detected even after a full year. Other studies also show inconsistency both inter-and intratumorally [24, 67]. Our own group has experienced similar issues with verified benign meningiomas (unpublished data). Despite these obstacles, primary patient-derived non-immortalized models display inter-patient tumor variability more accurately for possible targeted personalized treatment.
The major limitation to xenotransplant models is that they must be performed in immunocompromised animals, thereby circumventing natural response by the adaptive immune system (whether this is anti- or pro-tumorigenic) [173, 174]. In addition, the heterotopic animal models do not provide the correct microenvironment for the meningioma cells and can alter the way they grow and express cell markers. Finally, the tumor development is not de novo, meaning that xenograft studies are not useful for studying tumor origins.
Genetically engineered models (GEM)
GEM are based on mice that have undergone genome alterations through various genetic engineering techniques. There are multiple ways of achieving the desired genetic lesions. The following approaches have been used in meningioma research: The Cre-loxP system, which utilizes Cre-Lox recombination that can produce deletions and insertions at specific sites in the DNA. The DNA modification can be triggered by an external stimulus (e.g. recombinant adenovirus—AdCre) or be cell-type specific (i.e. is ‘conditional’). The alteration is performed by the splicing of previously inserted LoxP DNA sites using the enzyme Cre recombinase [151]. And the RCAS/TVA system, which utilizes retroviral infection via vectors that can only infect cells expressing the corresponding receptor TVA. The possibility of cloning the TVA receptor gene in mammalian cells has led to the creation of TVA-expressing transgenic mice [152]. RCAS is the vector and derives from the Rous sarcoma virus A [153]. The technology utilizes transfection of embryonic chicken fibroblast cell line DF-1 with the RCAS vectors, which then target ectopic TVA on pre-specified cells. The RCAS/TVA system is another example of a ‘conditional knockout’ system. In contrast to the Cre-loxP system, the RCAS/TVA system allows for simultaneous introduction of several genes of interest in the same cell [152].
As a group, meningiomas contain a plethora of DNA mutations depending on WHO grade and tumor location. Mutations in Nf2 [154], TRAF7, KLF4, AKT1, and SMO are present in approximately 80% of sporadic meningiomas [3, 155]. Especially the rare genetic disorder neurofibromatosis type 2 (Nf2) at q22 predisposes to meningiomas of which approximately 50% display alteration of the tumor suppressor [155,156,157]. Table 6 displays all lesions and outcomes of studies involving GEM (for further details, see Additional file 11). Only a few genes have been studied, mostly the Nf2 gene [27, 28, 30, 31, 130].
The TTR for Nf2(flox2/flox2) was 29% (95% CI 19–41) via AdCre injection orthotopically at PN1-3 to only target the Nf2 gene on both alleles over a duration of 117–450 days. The tumors were benign histologically, with transitional, meningothelial, and fibroblastic subtypes. However, a variety of other pathologies arose such as osteomas at the burr hole (51%), liver tumors (17%), and hydrocephalus (34%). Nf2Flox transgenic mice have been crossed with various other genes to assess the interactions. Waldt et al. [130] tested loss of the potential meningioma tumor suppressor receptor-like density-enhanced phosphatase-1 (DEP-1) [99], encoded by PTPRJ. They showed no TTR in PTPRJ−/− transgenic mice alone but raised TTRs ranging from 6% (0–27%) in their Nf2Flox2/Flox2 to 25% (13–40%) in Nf2(flox2/flox2);Ptprj(−/−), all over the same time period of one year for typical meningiomas with whorls and psammoma bodies, thus suggesting an interaction between the two genes in meningioma development.
It is well known that loss of the tumor suppressor p53 can cause tumor development through various pathways[158], as tested in congruency with Nf2 by Kalamarides et al.’s first GEM paper from 2002 [27]. They showed a 30% TTR in the Nf2 lesion alone and only 13% (6–28%) in Nf2flox/flox;p53± (heterozygous p53), however with a 91% rate of sarcomas/osteosarcomas over the course of a mere 165 days (median survival). A conditional homozygous lesion of p53flox/− with Nf2flox/− was also tested by the same group using the cell-specific prostaglandin D2 synthase (PDGS;Cre), which affects the fetus during intrauterine development. The authors found a higher TTR for Nf2(flox2/flox2);p53(flox2/−) of 45% (23–68%), but it was still lower than the TTR of 50% (29–71%) for the corresponding Nf2flox2/− alone. There was again a high number of malignant tumors, 79% osteosarcomas, and an even shorter survival of 135 days. The authors identify PGDS + arachnoid cells as a cell of origin for meningiomas [29].
Also of great interest are the tumor suppressor genes CDKN2A/B (located at 9p21 in humans). In meningiomas, alterations of CDKN2A/B are more common in higher grade tumors and are associated with high clinical recurrence [159, 160]. CDKN2A encodes the p16INK4a and p14arf (p19arf at chromosome 4 in mice [161]). p16INK4a regulates G1/S-phase via inhibition of cyclin-dependent kinases Cdk4 and Cdk6 [162], and p14arf regulates activity of p53 [163]. Adjacent to CDKN2A lies CDKN2B, which encodes the p15INK4b that also inhibits Cdk4 and 6 [164]. A TTR of 37% (95% CI 23–55%) was obtained by exploring only CDKN2A alteration (INK4a) using the AdCre method [28] and a TTR of 82% (95% CI 60–95%) from exploring the PDGS;Cre method [29]; these were primarily in benign tumors with few tumors showing atypical features and with other pathologies such as osteomas (78% and 88%, respectively) and hydrocephalus (56% and 31%, respectively). Further exploration of full CDKN2A/B hetero- and homozygous deletion led to creation of a Nf2(flox2/flox2);ink4ab(−/(−/+)) AdCre model. This showed a higher TTR of 76% (95% CI 61–86%) in homozygous [30, 132] compared to heterozygous 50% (36–64%) [30], with a higher take in sarcomas (34% vs 9%) and liver tumors (79% vs 59%). The homozygous deletion found 66% grade 1, 32% grade 2, and 3% grade 3, whereas the heterozygous deletion found 75% grade 1, 14% grade 2, and 11% grade 3 [30]. However, all homozygous tumors would be classified as malignant in accordance with the newly implemented WHO classification 2021 [165] due to CDKN2A/B homozygous deletion as an independent criterion of WHO grade 3 meningiomas.
Lastly, Morrison et al. [131] induced tumors using transgenic mice models of p16 and p19 wildtype, hetero- and homozygous and the carcinogenic compound N-ethyl-N-nitrosourea (ENU) as intraperitoneal injection at E14. They showed a TTR of 6% (95% CI 2–19%) and survival of 98–133 days for homozygous vs TTR of 31% (95% CI 7–75%) and survival of 210–273 days for heterozygous. Concomitant alveola-bronchiolar adenomas were present in almost all tumor-bearing mice. Wild type showed no meningioma tumors.
It has long been suggested that platelet-derived growth factor (PDGF) exhibits tumorigenic properties in meningiomas [166, 167]. Using the RCAS/TVA system, a PDGF-B model was created that showed a TTR of 27% (95% CI 12–48), all benign. However, the model also yielded 88% gliomas and 65% with hydrocephalus with a survival median of 240 days [31]. Furthermore, PDGF in combination with AdCre;Nf2 gave a higher TTR of 52% (33–71%) with 66% being grade 1, 40% grade 2, and 20% grade 3; however, there was still a high number of gliomas (48%) and a shorter median survival of 189 days. Lastly, they combined PDGF-B;Nf2;CDKN2AB lesions and found an even higher tumor take rate of 79% (54–94%) with 33% grade 1, 47% grade 2, and 20% grade 3 (but the same applies here as with the above CDKN2A/B−/− in relation to the malignancy grade). Median survival was greatly decreased to 54 days, and glioma incident remained high (79%).
SMO is a member of the Hedgehog (Hh) signaling pathway and is present in a small percentage of meningiomas (5%), specifically the meningothelial subtype [168]. It is a suggested oncogenic driver and is frequently associated with PI3K/AKT/mTOR pathway in driving tumor formation in meningiomas [168]. Boetto et al. explored this utilizing both PDGSCre; SMO and AdCre; SMO GEMs [133]. They found a TTR of 21% (95% CI 10–37%) in PDGSCre vs 2% (0–10%) in AdCre; all were meningothelial subtype with median survival of 426 days vs 84 days. The AdCre model besides having a shorter survival also produced medulloblastomas in 8%. The results suggest that SMO activation is restricted to a prenatal window E12.5 as is the case with PDGSCre.
Finally, Szulzewsky et al. [134] recently explored Yes-associated protein 1 (YAP1), which is involved in functional inactivation of Nf2 in heterozygous cases. YAP1 is a transcriptional coactivator of cell growth that is regulated by the Hippo signaling pathway and is especially associated with pediatric Nf2 wild-type meningiomas [169]. YAP1-MAML2 exerts oncogenic YAP activity that is resistant to inhibitory Hippo pathway signaling and relies on the interaction with TEAD transcription factors. Utilizing RCAS/TVA system and a Nestin/TVA CDKN2AB null mouse strain, it was found that the TTR for YAP1-MALM2(v1 and v2)’s was 42% (95% CI 15–72%) to 60% (95% CI 36–80%) of cases over the course of 67–164 days. A nuclear localization sequence (NLS)-2SA-YAP1 lesion—which constitutively activated YAP1 to inactivate Nf2—was explored to determine whether it would suffice to produce meningioma-like tumors. The authors showed a very high TTR in 97% (95% CI 83–100%) of the animals and verified their results with RNA sequencing. The study did not describe other pathologies present in the animals.
Strengths and weaknesses/limitations of GEM in meningiomas
In contrast to xenotransplantation in immunodeficient animals, GEM develop de novo tumors in immunocompetent animals [175]. GEM can thus be used to investigate candidate cancer genes (e.g. driver mutations), determine cancer cells-of-origin by altering specific targeted cells, and study the contribution of tumor microenvironment due to the intact immune system. GEM could thus be helpful in validating drug targets [175].
Although GEM have been of great value in cancer research, they have several disadvantages. The major weakness of Cre-loxP is that it does not allow for sequential and time-specific stepwise activation or inactivation of specific genes in vivo. This means that although Cre-loxP can assess single gene lesions, the model does not accurately reflect all aspects of sporadic multistep carcinogenesis [176]. The major weakness of RCAS/TVA is the limited insert capacity of the virus (2.8 kb), which limits the genes that can be evaluated [177]. Furthermore, producing a germline GEM is labor-intensive, time-consuming, and expensive [175]. Despite the clear advantages of a de novo tumor in an immunocompetent environment, the tumor remains a mouse tumor and not a human tumor—and these may act/react differently, e.g. CDKN2A/B is present at chromosome 9 in humans but chromosome 4 in mice [161, 162].
Uncategorized models
These studies were deemed too heterogenous and too few to be included in the meta-analysis and subgroup analyses. A narrative approach was chosen to describe the studies of most interest, in addition to a table (Table 7) (for further details Additional file 12).
Syngeneic models were first tried in 1994 by Yamate et al. [142], who excised a highly malignant cerebellar tumor from an F344 rat; this was consecutively passaged and transplanted with great success (100% TTR) into new animals. Furthermore, the models showed similar histological traits to the parent tumor. Peyre and Kalamarides in collaboration with Yeung et al. [30, 132, 135] have performed 100% TTR fast-growing syngeneic malignant models (both orthotopic and heterotopic) using cell lines derived from Peyre et al. GEM (MGS1-3) [30]. This model type provides a stable method of assessing WHO grade 3 meningiomas and their interaction with the immune system as the animals are immunocompetent. Thus, it is a genetically modifiable alternative to the spontaneous animal-derived tumor studies from the 1990s [142, 143].
Developing meningiomas in an animal model has also been successful from patient-derived tumor stem-like cells [140, 141]. These studies show tumors reflecting the histological features and immunohistochemistry of the parent tumors. This model type could be used for assessing tumorigenesis of various progenitor cells. Baia et al. [139] transfected non-neoplastic arachnoidal cells with Yes-Associated Protein-1 and found 100% tumor in comparison to no tumor in controls. Tumorigenic studies on specific proteins or other factors could be performed in a similar fashion.
Designing a model: type and validation of growth and verification of tumor
The preferred model depends on multiple factors such as the type of research and availability of resources (specialized equipment, scans, laboratory analyses). This section gives an overview via Table 8 of the advantages and disadvantages of the different model types, and we present a separate section on validation of growth and verification of tumor.
Validation of growth and verification of tumor
During any experiment, it is important to follow growth at various time-points, e.g. before, during, and after treatment to assess the efficacy of the intervention. Due to the associated skin and fatty tissue, the size of a tumor xenografted subcutaneously is more difficult to estimate via caliper. A subrenal capsule approach can give more precise measurements but requires surgery for each measurement at the cost of animal strain [106, 127, 129]. A single study reports peritoneal injection [58], but this approach has the same disadvantages as orthotopic models without any benefits. It is difficult to assess tumor growth in orthotopic models without sacrificing the animals and without specialized non-invasive methods such as bioluminescence, MRI, and PET (Tables 4, 5 and 6). Bioluminescence is widely used for real-time imaging in vivo. Once cells have been transduced with luciferase (Fluc) prior to implantation, it is relatively easy and inexpensive [178]. MRI scans are also widely used but usually require special high-field MRI scanners for optimal resolution on a small scale (3.0T [67], 4.7T [28, 30, 53, 61, 62], 9.4T [25, 83]), but can be achieved to a certain degree with a clinical 1.5Tesla MRI [81].
Meningioma verification in the clinic depends primarily on HE histology and perhaps the immunohistochemical markers vimentin, epithelial membrane antigen (EMA), somatostatin receptor 2 (SSTR2), and Ki67/MIB-1. However, newer guidelines include genome sequencing on special cases [165]. Particularly when implanting primary patient-derived cells or tissue, researchers should perform a panel of HE and immunohistochemical markers and compare to the parent tumor at the very least, however it is recommended to compare at a genomic [179] or epigenomic level. During inoculation of cancer cells in mice, the stroma is replaced by mouse cells, resulting in a mix of DNA [180]. It was shown in pancreas cancer that xenografts in immunocompromised animals were contaminated with 47% (17–73%) mouse DNA [181]. To our knowledge, no such experiments have been conducted in meningioma research, but it is important to take this into account when discussing results of a treatment. In regard to verifying and classifying GEM-derived tumors, Kalamarides and colleagues have suggested a specific classification based on histological analysis, which reflects tumor composition more accurately than the human classification [35], which should be used in case of GEM.
Critical appraisal of methodological quality and quality of reporting (CRIME-Q)
All studies included in this meta-analysis were peer-reviewed, assuring a certain quality. We considered 67% of all studies to sufficiently describe the bench-top part of the experiments, but only 46% described the animals properly. Almost all studies did not calculate sample size, and no studies provided a full description of the sample size calculation. A full description of the in vivo experiment elements was present in 59% of the studies, and 27% had such deficient descriptions that the experiment could not be replicated. Around two-thirds of the studies described compliance with animal welfare, and almost no papers (6%) were blinded in any way. It is not standard practice in animal research to discuss model limitations, which was reflected in three-quarters of the studies not mentioning this. In general, we found a higher percentage of studies with good methodological approach (2Y, 3Y, 5Y) than with good quality of reporting (2X, 3X, 5X), see Fig. 3A.

CRIME-Q—Quality of Reporting, Methodological Quality and SYRCLE’s Risk of Bias. Graphical display of all categories 1–10 including all included studies A Quality of Reporting items (X) and Methodological Quality (Y) Green = Yes, Yellow = Partly, Red = No. B Results of Items from SYRCLE’s Risk of Bias tool. Green = Yes, Yellow = Unclear, Red = No, Blue = Not applicable. 1X Peer-review, 2X Bench-top/laboratory work related to establishing model, 2Y Methodological quality of 2X, 3X Animals, 3Y Methodological quality of 3X, 3Z: Selection bias (baseline characteristics), 4Y Sample size, 5X in vivo design and performance, 5Y: Methodological quality of 5X, 5Z(1): Selection bias (Sequence generation), 5Z(2): Performance bias (Random housing), 5Z(3): Detection bias (Random outcome assessment), 6X: Compliance with animal welfare regulations, 7X Blinding, 7Z(1) Performance bias (Blinding), 7Z(2) Allocation bias (allocation concealment), 7Z(3) Detection bias (blinding), 8X Congruency between methods and results, 8Z(1) Attrition bias (incomplete outcome data), 8Z(2) Reporting bias (Selective outcome reporting), 9X Presentation of limitations, 10X Statement of potential conflict of interest, 10Z Other bias (Publication bias). See Table 1 for further descriptions
SYRCLE’s risk of bias items were included in the CRIME-Q tool. Items 3Z, 5Z [1,2,3], and 7 [1,2,3] were mostly unclear in 67–89% of the included studies. A few studies were non-interventional, so some items were not applicable. We found that 79% of studies had a low risk of excluding outcome data (Attrition bias, 8Z [1]). Although no study protocols were apparently available, it was clear in 79% of studies that the published reports included all expected outcomes (i.e., comparing the Methods and Result sections) and reported key elements, which would have been expected from such studies (i.e. tumor-take rate) (Selection bias 8Z [2]). No risk of influence from third party was found in 56% of studies (Publication bias 10Z) Fig. 3B.
Discussion
This meta-analysis showed that the tumor take rate (TTR) for established cell line models (ECLMs) was near 100% for both orthotopic models [94% (95% CI 92–96)] and heterotopic models [95% (95% CI 93–96)]. These results proved consistent regardless of subgroup category, time frame (duration of incubation), number of cells, injection volume, cell concentration, and cell line (Additional file 13). TTR was more variable for primary patient-derived tumor models (PTMs), with 53% (95% CI 33–72) for orthotopic models and 82% (95% CI 73–89) for heterotopic models. Subgroup analyses for PTM showed varying TTRs, e.g. high (98%; 89–100%) for orthotopic duration of incubation between 14 and 30 days, but low [8% (95% CI 3–21%)] for 200–499 days of incubation. We could not identify any pattern in take rate according to the number of cells, e.g. 100–1000 cells had high TTR [86% (95% CI 42–100%)], 10.000–100.000 had very low TTR [15% (95% CI 5–37%)], and 500.000–1.000.000 cells had higher again [91% (95% CI 77–97%)].
Two aspects should be considered before applying these results in models. First, we found few published papers on PTM section and secondly, only a few studies have published negative tumor takes (0%) in some samples, namely Ragel et al. [67], Malham et al. [122], Hu et al. [121], and most importantly, Zhang et al. [25] who published several individual tumors unable to obtain growth in vivo. To our knowledge there has been no investigation into why some tumors simply fail to take in animals, and no papers have focused on extensive genomic/epigenomic alterations in meningioma xenografts. One explanation could be lack of certain driver mutations in tumors that help develop tumors in the animals. Our own experiences regarding inter-tumor heterogeneity of take-rate and knowledge within the field lead us to believe, there is a high risk of publication bias in the current analysis as not all eligible studies may have been published. If unpublished studies are more likely to show negative results, this may have skewed our results. There is a larger focus on publishing negative results today [183], but it remains an issue overall as suggested in this paper. Although submission of animal study protocols is recommended and feasible through open access journals without peer-review, it is still not common practice.
Most (70%) of the studies presented here were based on immortalized cell lines, which by far mimic proper tumor qualities. The ability of these cell lines to provide a steady, fast, and homogeneous growth makes them especially suitable to quickly test experimental treatment strategies as an ‘add on’ in often times very well performed in vitro studies with (often) very few animals i.e. [57, 64, 74, 75, 78, 80, 88, 93, 102, 103]. This may explain the low quality of reporting, which is problematic for the research community. These studies sometimes use a heterotopic model and a very early treatment start day (1–2 weeks) [52], thus not mimicking normal tumor pathology or treatment in any way. Despite this, ECLMs have a role to play in exploring, for example, scan modalities, cognitive function studies, and tumor load studies, where the specific tumor characteristics are less important.
The TTR of 34% across all GEM studies indicate a problem of finding a strong oncogenic driver that can be used to model meningioma with its typical benign nature and slow growth, which presents challenges in preclinical research [134]. The only study that reported solid growth in almost 100% of the cases was based on constitutively activated YAP1 [134], but meningiomas initiated by YAP1 fusion are a rare subset of childhood and young adulthood meningiomas [184]. Given sufficient time, sample size, and appropriate design to avoid an underpowered study [185], the lower take rates of GEM would not be problematic in pre-clinical pharmaceutical tests. Otherwise, the syngeneic model could be used for faster growth at higher take. Translatability from GEM (mouse tumor) to human in preclinical treatment studies in meningioma still remains to be seen.
The average rate of successful translation from animal models to clinical cancer trials is generally around 8% [23], and meningiomas are no exception. As there are currently no known pharmaceuticals with sufficient clinical benefits [186,187,188], in vitro and in vivo research in this field is paramount. Possible issues in relation to this loss in translation are meningioma heterogeneity as a group in terms of genetics/epigenetics and histology, microenvironmental challenges and design/use of model. Gene expression, epigenetic profiling, molecular markers, and DNA-technologies have over the past decade especially helped uncover the heterogeneity. And in line with this, the continuing search for the perfect model, which has not been discovered and might not exist. However, we can strive towards model improvement and development. Furthermore, we ought to use the ‘right’ type of model, which mimics the condition researched/reflects research question as close as possible to decrease the gap in translation.
Poor study reporting leads to irreproducible and uncertain findings [189, 190]. In this study, we present a new tool (CRIME-Q) to assess all kinds of animal studies quickly and thoroughly on multiple parameters. CRIME-Q identified issues in reporting, which influences methodological quality and risk of bias greatly, further emphasizing the need for research to report their methods and findings more thoroughly to ensure transparency, replicability and ultimately usability of models. CRIME-Q unifies multiple aspects of quality of reporting, methodological quality, and risk of bias from bench-top to in vivo design and performance and related items in between including risk of bias and presents a clearer overall assessment of included studies in systematic reviews on animal research. Used alongside the ARRIVE 2.0 guidelines for animal studies [41], CRIME-Q can help improve study transparency and replicability. We found it necessary to develop our own broader method, which also includes bench-top assessment, since we found no suitable options available [191]. Our tool is not yet externally validated, but we present all the results here in a transparent manner for further inspection.
It is important to note that this review intended to assess animal models only. Many of the included papers used animal studies to underpin the in vitro findings or as proof of concept, which will have influenced the quality of reporting, methodological quality, and risk of bias. The focus of these papers simply is not animal studies and thus our judgements should not be regarded as overall judgements of quality of the studies, but rather as judgements of the reporting and methodological quality in relation to our objectives. Individual study scores can be assessed in the Additional file 13, where a short description and grade is given for each category. Furthermore, many of the studies were published over 15 years ago, when there was less focus on full reporting.
Study strengths and limitations
A strength of this study is the systematic approach in accordance with the PRISMA guidelines, where we made individual assessments of study eligibility and data extraction. The structured critical appraisal of methodological quality and quality of reporting of all included studies allowed us to judge the overall reliability of the studied body of evidence. A further strength is the meta-analysis on TTRs for the various model types, allowing us to assess the effectiveness of different models.
The study also has several limitations. First, there is a high risk of publication bias, as noted above. If studies with high success rates are more likely to be published, meta-analyses might overestimate the overall success rate. Likewise, if studies using specific methods are more likely to be published, this might limit the ability to compare success rates for different methods. Secondly, while we critically appraised all included studies, we used a non-validated tool. We believe we have included the most important methodological aspects, but some could have been overlooked, and we welcome feedback and criticism of the tool from the scientific community. Lastly, the methodological quality and the quality of reporting of included studies varied. Our results (including from the meta-analyses and subgroup analyses) should thus be interpreted with caution.
Future perspectives for meningioma models and knowledge gaps
We describe here some selected points of interest in our search to identify and analyze knowledge gaps in the use of meningioma animal models for optimal preclinical tests.
First, previous successful in vivo pharmaceutical trials in xenograft models in meningiomas have failed to translate to human conditions. This is probably due to microenvironmental challenges from the lack of a proper immune response in immune-incompetent animals. Use of humanized animals [192] could help this, i.e., by engrafting CD34 + human hematopoietic stem and precursor cells to encourage development of a normal immune system [193]. The use of humanized animals with co-engrafted stromal and immune components is not yet a perfect science [192], but the technology has great potential for both tumor growth and drug response studies. Its uses remain to be seen in meningiomas.
Second, 68 Ga-DOTA(TOC/NOC/TATE) PET-CT provides high-contrast images of meningiomas due to the abundance of somatostatin receptor 2 in meningiomas but not in brain (except the pituitary gland) and bone. PET-CT has shown to be more sensitive than MRI to detect meningiomas [194, 195] and is widely used in the clinic [196]. Only two preclinical studies have assessed PET in heterotopic models, both of which showed easy distinction between healthy tissue and tumor [95, 96]. These findings need to be verified in an orthotopic model that could be used in both GEM and xenograft models.
Third, previous comparisons between primary patient-derived tumor and corresponding xenograft in meningioma research have primarily used histology and immunohistochemistry. Only Zhang et al. [25] assessed two tumors (WHO grade II and III) and corresponding xenograft gene expression profiles using RNA sequence to assess DNA copy number variations and genes. The xenograft tumors retained all of the copy number variations seen in the original tumors. When comparing original tumor to xenograft and normal brain tissue, the authors found high correlation between original tumor and xenograft genes. Moreover, a large study involving several hundred xenograft cancer models of many types showed no enrichment of cancer-related genes in xenografts and concluded a lack of systematic copy number evolution driven by the PDX mouse host [197]. Despite these findings, we lack epigenetic knowledge of meningioma xenograft using DNA-methylation (especially in the benign tumors, which have not yet been tested). We should explore this approach further to identify the model type that best mimics human conditions.
Finally, we have limited knowledge of GEMs and oncogenic drivers, as well as the impact of different genes e.g., TRAF7, AKT1. The observed TTR of 34% for GEMs indicates challenges in finding a strong oncogenic driver that can be used to model this disease with the Cre-recombinase and RCAS/TVA systems. Conditional or ‘time- and site-specific’ DNA alteration is important in various diseases as some gene alterations are not compatible with intrauterine development [151], e.g. homozygous Nf2-deleted mouse embryos fail in development [198]. The CRISPR/Cas9 technology can target any genomic point through single-guide RNAs [199] and can be used to introduce defined mutations or loxP-sites [175]. It is especially useful in non-germline models based on direct gene editing in vivo. This could be used to quickly identify oncogenic genes without the need for extensive breeding to obtain the proper transgenic model [200].
Conclusion
This systematic review shows high consistent tumor take rates in established cells lines and varying tumor take rates in primary-patient derived material and genetically engineered models. However, we identified various issues across the studies regarding the quality of reporting, the methodological approach, and a high risk of publication bias. Each tumor model type has specific roles. ECLMs are useful for modality testing and other tumor burden studies, while PTMs (orthotopic) mimic tumor heterogeneity and have low cost and technical skill requirements, making them useful in initial pharmaceutical testing if appropriate examinations are performed (e.g. DNA sequencing, DNA methylation). Finally, GEMs are useful in assessing and validating driver mutations and determining cells-of-origin, making them relevant in pre-clinical testing due to an intact immune system; they may also be beneficial in validating drug targets.
Availability of data and materials
All data generated or analyzed during this study are included in this published record [and its Additional files].
References
Ostrom QT, Price M, Neff C, Cioffi G, Waite KA, Kruchko C, et al. CBTRUS statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2015–2019. Neuro Oncol. 2022;24(Suppl 5):v1–95.
Nigim F, Wakimoto H, Kasper EM, Ackermans L, Temel Y. Emerging medical treatments for meningioma in the molecular era. Biomedicines. 2018;6(3):86.
Bi WL, Mei Y, Agarwalla PK, Beroukhim R, Dunn IF. Genomic and epigenomic landscape in meningioma. Neurosurg Clin N Am. 2016;27(2):167–79.
Suppiah S, Nassiri F, Bi WL, Dunn IF, Hanemann CO, Horbinski CM, et al. Molecular and translational advances in meningiomas. Neuro Oncol. 2019;21(Suppl 1):i4–17.
Sahm F, Schrimpf D, Stichel D, Jones DTW, Hielscher T, Schefzyk S, et al. DNA methylation-based classification and grading system for meningioma: a multicentre, retrospective analysis. Lancet Oncol. 2017;18(5):682–94.
Choudhury A, Magill ST, Eaton CD, Prager BC, Chen WC, Cady MA, et al. Meningioma DNA methylation groups identify biological drivers and therapeutic vulnerabilities. Nat Genet. 2022;54(5):649–59.
Driver J, Hoffman SE, Tavakol S, Woodward E, Maury EA, Bhave V, et al. A molecularly integrated grade for meningioma. Neuro Oncol. 2022;24(5):796–808.
Simpson D. The recurrence of intracranial meningiomas after surgical treatment. J Neurol Neurosurg Psychiatry. 1957;20(1):22–39.
Goldbrunner R, Stavrinou P, Jenkinson MD, Sahm F, Mawrin C, Weber DC, et al. EANO guideline on the diagnosis and management of meningiomas. Neuro Oncol. 2021;23(11):1821–34.
Krementz ET, Greene HS. Heterologous transplantation of human neural tumors. Cancer. 1953;6(1):100–10.
Greene HS, Arnold H. The homologous and heterologous transplantation of brain and brain tumors. J Neurosurg. 1945;2:315–29.
Ishida YS, Sato K, Niibe H. Experimental induction of gliogenous tumors in rats. A histopathologic and electron microscope study. Gunma J Med Sci. 1963;12(1):36–72.
Greene HS. The transplantation of human brain tumors to the brains of laboratory animals. Cancer Res. 1953;13(6):422–6.
Miyawaki H, Ishii S. The heterologous intracerebral transplantation of human brain tumors. Arch Pathol. 1960;70:508–19.
Maekawa A, Odashima S, Nakadate M. Induction of tumors in the stomach and nervous system of the ACI/N rat by continuous oral administration of 1-methyl-3-acetyl-1-nitrosourea. Z Krebsforsch Klin Onkol Cancer Res Clin Oncol. 1976;86(2):195–207.
Ponomarkov V, Tomatis L. Long-term testing of vinylidene chloride and chloroprene for carcinogenicity in rats. Oncology. 1980;37(3):136–41.
Anigstein L, Anigstein DM, Unterharnscheidt FJ. Mouse transplantable tumor induced by human spinal meningioma implant in mice inoculated with human thymus antiserum. Tex Rep Biol Med. 1969;27(2):341–66.
Ueyama Y, Ohsawa N, Tamaoki N, Nomura T. Heterotransplantation of human neoplasms in nude mice. Keio J Med. 1975;24(4):415–21.
Ueyama Y, Morita K, Ochiai C, Ohsawa N, Hata J, Tamaoki N. Xenotransplantation of a human meningioma and its lung metastasis in nude mice. Br J Cancer. 1978;37(4):644–7.
Ragel BT, Couldwell WT, Gillespie DL, Wendland MM, Whang K, Jensen RL. A comparison of the cell lines used in meningioma research. Surg Neurol. 2008;70(3):295–307.
Puttmann S, Senner V, Braune S, Hillmann B, Exeler R, Rickert CH, et al. Establishment of a benign meningioma cell line by hTERT-mediated immortalization. Lab Invest. 2005;85(9):1163–71.
Lee WH. Characterization of a newly established malignant meningioma cell line of the human brain: IOMM-Lee. Neurosurgery. 1990;27(3):389–95.
Mak IW, Evaniew N, Ghert M. Lost in translation: animal models and clinical trials in cancer treatment. Am J Transl Res. 2014;6(2):114–8.
McCutcheon IE, Friend KE, Gerdes TM, Zhang BM, Wildrick DM, Fuller GN. Intracranial injection of human meningioma cells in athymic mice: an orthotopic model for meningioma growth. J Neurosurg. 2000;92(2):306–14.
Zhang H, Qi L, Du Y, Huang LF, Braun FK, Kogiso M, et al. Patient-derived orthotopic xenograft (PDOX) mouse models of primary and recurrent meningioma. Cancers (Basel). 2020;12(6):147.
Friedrich S, Schwabe K, Klein R, Krusche CA, Krauss JK, Nakamura M. Comparative morphological and immunohistochemical study of human meningioma after intracranial transplantation into nude mice. J Neurosci Methods. 2012;205(1):1–9.
Kalamarides M, Niwa-Kawakita M, Leblois H, Abramowski V, Perricaudet M, Janin A, et al. Nf2 gene inactivation in arachnoidal cells is rate-limiting for meningioma development in the mouse. Genes Dev. 2002;16(9):1060–5.
Kalamarides M, Stemmer-Rachamimov AO, Takahashi M, Han ZY, Chareyre F, Niwa-Kawakita M, et al. Natural history of meningioma development in mice reveals: a synergy of Nf2 and p16(Ink4a) mutations. Brain Pathol. 2008;18(1):62–70.
Kalamarides M, Stemmer-Rachamimov AO, Niwa-Kawakita M, Chareyre F, Taranchon E, Han ZY, et al. Identification of a progenitor cell of origin capable of generating diverse meningioma histological subtypes. Oncogene. 2011;30(20):2333–44.
Peyre M, Stemmer-Rachamimov A, Clermont-Taranchon E, Quentin S, El-Taraya N, Walczak C, et al. Meningioma progression in mice triggered by Nf2 and Cdkn2ab inactivation. Oncogene. 2012;32:4264.
Peyre M, Salaud C, Clermont-Taranchon E, Niwa-Kawakita M, Goutagny S, Mawrin C, et al. PDGF activation in PGDS-positive arachnoid cells induces meningioma formation in mice promoting tumor progression in combination with Nf2 and Cdkn2ab loss. Oncotarget. 2015;6(32):32713–22.
Mawrin C. Animal models of meningiomas. Chin Clin Oncol. 2017. https://doi.org/10.21037/cco.2017.05.03.
Kalamarides M, Peyre M, Giovannini M. Meningioma mouse models. J Neurooncol. 2010;99(3):325–31.
Cimino PJ. Malignant progression to anaplastic meningioma: neuropathology, molecular pathology, and experimental models. Exp Mol Pathol. 2015;99(2):354–9.
Boetto J, Peyre M, Kalamarides M. Mouse models in meningioma research: a systematic review. Cancers (Basel). 2021;13(15):3712.
Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021;372: n71.
Cohen J. A coefficient of agreement for nominal scales. Educ Psychol Measur. 1960;20:37–46.
Higgins J, Thomas J, Chandler J, Cumpston M, Li T, Page M, et al. Cochrane handbook for systematic reviews of interventions. 2nd ed. Chichester: Wiley; 2019.
Andersen MS, Kofoed MS, Paludan-Müller AS, Pedersen CB, Mathiesen T, Mawrin C, et al. A unifying tool for critical appraisal of methodological quality, quality of reporting and risk of bias in animal research. Submitted to BMC Medical Research Methodology. 2023
Macleod MR, O’Collins T, Howells DW, Donnan GA. Pooling of animal experimental data reveals influence of study design and publication bias. Stroke. 2004;35(5):1203–8.
Percie du Sert N, Ahluwalia A, Alam S, Avey MT, Baker M, Browne WJ, et al. Reporting animal research: explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020;18(7): e3000411.
Hooijmans CR, Rovers MM, de Vries RB, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE’s risk of bias tool for animal studies. BMC Med Res Methodol. 2014;14:43.
Gogineni VR, Nalla AK, Gupta R, Dinh DH, Klopfenstein JD, Rao JS. Chk2-mediated G2/M cell cycle arrest maintains radiation resistance in malignant meningioma cells. Cancer Lett. 2011;313(1):64–75.
Das A, Alshareef M, Henderson F Jr, Martinez Santos JL, Vandergrift WA 3rd, Lindhorst SM, et al. Ganoderic acid A/DM-induced NDRG2 over-expression suppresses high-grade meningioma growth. Clin Transl Oncol. 2020;22(7):1138–45.
Bao Z, Hua L, Ye Y, Wang D, Li C, Xie Q, et al. MEF2C silencing downregulates NF2 and E-cadherin and enhances erastin-induced ferroptosis in meningioma. Neuro Oncol. 2021;23(12):2014–27.
Kim H, Park KJ, Ryu BK, Park DH, Kong DS, Chong K, et al. Forkhead box M1 (FOXM1) transcription factor is a key oncogenic driver of aggressive human meningioma progression. Neuropathol Appl Neurobiol. 2020;46(2):125–41.
von Spreckelsen N, Waldt N, Poetschke R, Kesseler C, Dohmen H, Jiao HK, et al. KLF4(K409Q)-mutated meningiomas show enhanced hypoxia signaling and respond to mTORC1 inhibitor treatment. Acta Neuropathol Commun. 2020;8(1):41.
Guo L, Cui J, Wang H, Medina R, Zhang S, Zhang X, et al. Metformin enhances anti-cancer effects of cisplatin in meningioma through AMPK-mTOR signaling pathways. Mol Ther Oncolytics. 2021;20:119–31.
Giles AJ, Hao S, Padget M, Song H, Zhang W, Lynes J, et al. Efficient ADCC killing of meningioma by avelumab and a high-affinity natural killer cell line, haNK. JCI Insight. 2019;4(20): e130688.
Ho WS, Sizdahkhani S, Hao S, Song H, Seldomridge A, Tandle A, et al. LB-100, a novel protein phosphatase 2A (PP2A) inhibitor, sensitizes malignant meningioma cells to the therapeutic effects of radiation. Cancer Lett. 2018;415:217–26.
Wang S, Liu X, Wang W, Tu Y, Wang C, Mei J, et al. The effects of silencing the Her2 gene on proliferation and angiogenesis of meningioma cells in vivo and in vitro. Ann Clin Lab Sci. 2018;48(5):580–6.
Zhang Q, Song LR, Huo XL, Wang L, Zhang GB, Hao SY, et al. MicroRNA-221/222 inhibits the radiation-induced invasiveness and promotes the radiosensitivity of malignant meningioma cells. Front Oncol. 2020;10:1441.
Tuchen M, Wilisch-Neumann A, Daniel EA, Baldauf L, Pachow D, Scholz J, et al. Receptor tyrosine kinase inhibition by regorafenib/sorafenib inhibits growth and invasion of meningioma cells. Eur J Cancer. 2017;73:9–21.
Bahr O, Gross S, Harter PN, Kirches E, Mawrin C, Steinbach JP, et al. ASA404, a vascular disrupting agent, as an experimental treatment approach for brain tumors. Oncol Lett. 2017;14(5):5443–51.
Das A, Alshareef M, Martinez Santos JL, Porto GBF, McDonald DG, Infinger LK, et al. Evaluating anti-tumor activity of palbociclib plus radiation in anaplastic and radiation-induced meningiomas: pre-clinical investigations. Clin Transl Oncol. 2020;22(11):2017–25.
Wang Z, Wang W, Xu S, Wang S, Tu Y, Xiong Y, et al. The role of MAPK signaling pathway in the Her-2-positive meningiomas. Oncol Rep. 2016;36(2):685–95.
Park KJ, Yu MO, Song NH, Kong DS, Park DH, Chae YS, et al. Expression of astrocyte elevated gene-1 (AEG-1) in human meningiomas and its roles in cell proliferation and survival. J Neurooncol. 2015;121(1):31–9.
Iwami K, Momota H, Fujii M, Natsume A, Yagi S, Toriyama K, et al. Anaplastic meningioma with rapid growth after omental flap transposition: a case report and experimental study. Brain Tumor Pathol. 2015;32(2):137–44.
Wilisch-Neumann A, Kliese N, Pachow D, Schneider T, Warnke JP, Braunsdorf WE, et al. The integrin inhibitor cilengitide affects meningioma cell motility and invasion. Clin Cancer Res. 2013;19(19):5402–12.
Iwami K, Natsume A, Ohno M, Ikeda H, Mineno J, Nukaya I, et al. Adoptive transfer of genetically modified Wilms’ tumor 1-specific T cells in a novel malignant skull base meningioma model. Neuro Oncol. 2013;15(6):747–58.
Pachow D, Andrae N, Kliese N, Angenstein F, Stork O, Wilisch-Neumann A, et al. mTORC1 inhibitors suppress meningioma growth in mouse models. Clin Cancer Res. 2013;19(5):1180–9.
Kliese N, Gobrecht P, Pachow D, Andrae N, Wilisch-Neumann A, Kirches E, et al. miRNA-145 is downregulated in atypical and anaplastic meningiomas and negatively regulates motility and proliferation of meningioma cells. Oncogene. 2013;32(39):4712–20.
Haase D, Schmidl S, Ewald C, Kalff R, Huebner C, Firsching R, et al. Fatty acid synthase as a novel target for meningioma therapy. Neuro Oncol. 2010;12(8):844–54.
Ge Y, Xu K. Alpha-synuclein contributes to malignant progression of human meningioma via the Akt/mTOR pathway. Cancer Cell Int. 2016;16:86.
Kargiotis O, Chetty C, Gogineni V, Gondi CS, Pulukuri SM, Kyritsis AP, et al. uPA/uPAR downregulation inhibits radiation-induced migration, invasion and angiogenesis in IOMM-Lee meningioma cells and decreases tumor growth in vivo. Int J Oncol. 2008;33(5):937–47.
Baia GS, Dinca EB, Ozawa T, Kimura ET, McDermott MW, James CD, et al. An orthotopic skull base model of malignant meningioma. Brain Pathol. 2008;18(2):172–9.
Ragel BT, Elam IL, Gillespie DL, Flynn JR, Kelly DA, Mabey D, et al. A novel model of intracranial meningioma in mice using luciferase-expressing meningioma cells. Lab Investig J Neurosurg. 2008;108(2):304–10.
Tummalapalli P, Gondi CS, Dinh DH, Gujrati M, Rao JS. RNA interference-mediated targeting of urokinase plasminogen activator receptor and matrix metalloproteinase-9 gene expression in the IOMM-lee malignant meningioma cell line inhibits tumor growth, tumor cell invasion and angiogenesis. Int J Oncol. 2007;31(1):5–17.
Cargioli TG, Ugur HC, Ramakrishna N, Chan J, Black PM, Carroll RS. Establishment of an in vivo meningioma model with human telomerase reverse transcriptase. Neurosurgery. 2007;60(4):750–9 (discussion 9–60).
Tummalapalli P, Spomar D, Gondi CS, Olivero WC, Gujrati M, Dinh DH, et al. RNAi-mediated abrogation of cathepsin B and MMP-9 gene expression in a malignant meningioma cell line leads to decreased tumor growth, invasion and angiogenesis. Int J Oncol. 2007;31(5):1039–50.
Gupta V, Su YS, Samuelson CG, Liebes LF, Chamberlain MC, Hofman FM, et al. Irinotecan: a potential new chemotherapeutic agent for atypical or malignant meningiomas. J Neurosurg. 2007;106(3):455–62.
Ragel BT, Jensen RL, Gillespie DL, Prescott SM, Couldwell WT. Celecoxib inhibits meningioma tumor growth in a mouse xenograft model. Cancer. 2007;109(3):588–97.
Ragel BT, Gillespie DL, Kushnir V, Polevaya N, Kelly D, Jensen RL. Calcium channel antagonists augment hydroxyurea- and ru486-induced inhibition of meningioma growth in vivo and in vitro. Neurosurgery. 2006;59(5):1109–20 (discussion 20-1).
Kondraganti S, Gondi CS, Gujrati M, McCutcheon I, Dinh DH, Rao JS, et al. Restoration of tissue factor pathway inhibitor inhibits invasion and tumor growth in vitro and in vivo in a malignant meningioma cell line. Int J Oncol. 2006;29(1):25–32.
Kondraganti S, Gondi CS, McCutcheon I, Dinh DH, Gujrati M, Rao JS, et al. RNAi-mediated downregulation of urokinase plasminogen activator and its receptor in human meningioma cells inhibits tumor invasion and growth. Int J Oncol. 2006;28(6):1353–60.
Surace EI, Lusis E, Haipek CA, Gutmann DH. Functional significance of S6K overexpression in meningioma progression. Ann Neurol. 2004;56(2):295–8.
Salhia B, Rutka JT, Lingwood C, Nutikka A, Van Furth WR. The treatment of malignant meningioma with verotoxin. Neoplasia. 2002;4(4):304–11.
Rao Gogineni V, Kumar Nalla A, Gupta R, Gorantla B, Gujrati M, Dinh DH, et al. Radiation-inducible silencing of uPA and uPAR in vitro and in vivo in meningioma. Int J Oncol. 2010;36(4):809–16.
Waldt N, Kesseler C, Fala P, John P, Kirches E, Angenstein F, et al. Crispr/Cas-based modeling of NF2 loss in meningioma cells. J Neurosci Methods. 2021;356: 109141.
Gogineni VR, Gupta R, Nalla AK, Velpula KK, Rao JS. uPAR and cathepsin B shRNA impedes TGF-beta1-driven proliferation and invasion of meningioma cells in a XIAP-dependent pathway. Cell Death Dis. 2012;3(12): e439.
van Furth WR, Laughlin S, Taylor MD, Salhia B, Mainprize T, Henkelman M, et al. Imaging of murine brain tumors using a 1.5 Tesla clinical MRI system. Can J Neurol Sci. 2003;30(4):326–32.
Nalla AK, Gogineni VR, Gupta R, Dinh DH, Rao JS. Suppression of uPA and uPAR blocks radiation-induced MCP-1 mediated recruitment of endothelial cells in meningioma. Cell Signal. 2011;23(8):1299–310.
John P, Waldt N, Liebich J, Kesseler C, Schnabel S, Angenstein F, et al. AKT1(E17K) -mutated meningioma cell lines respond to treatment with the AKT inhibitor AZD5363. Neuropathol Appl Neurobiol. 2022;48(2): e12780.
Nakano T, Fujimoto K, Tomiyama A, Takahashi M, Achiha T, Arita H, et al. Eribulin prolongs survival in an orthotopic xenograft mouse model of malignant meningioma. Cancer Sci. 2022;113(2):697–708.
Horbinski C, Xi G, Wang Y, Hashizume R, Gopalakrishnan M, Phillips JJ, et al. The effects of palbociclib in combination with radiation in preclinical models of aggressive meningioma. Neurooncol Adv. 2021;3(1):vdab085.
Deng Y, Hu B, Miao Y, Wang J, Zhang S, Wan H, et al. A nicotinamide phosphoribosyltransferase inhibitor, FK866, suppresses the growth of anaplastic meningiomas and inhibits immune checkpoint expression by regulating STAT1. Front Oncol. 2022;12: 836257.
Chen X, Tian F, Lun P, Feng Y. Curcumin inhibits HGF-induced EMT by regulating c-MET-dependent PI3K/Akt/mTOR signaling pathways in meningioma. Evid Based Complement Alternat Med. 2021;2021:5574555.
Yang A, Yang X, Wang J, Wang X, Wu H, Fan L, et al. Effects of the tight junction protein CLDN6 on cell migration and invasion in high-grade meningioma. World Neurosurg. 2021;151:e208–16.
Angus SP, Oblinger JL, Stuhlmiller TJ, DeSouza PA, Beauchamp RL, Witt L, et al. EPH receptor signaling as a novel therapeutic target in NF2-deficient meningioma. Neuro Oncol. 2018;20(9):1185–96.
Chow HY, Dong B, Duron SG, Campbell DA, Ong CC, Hoeflich KP, et al. Group I Paks as therapeutic targets in NF2-deficient meningioma. Oncotarget. 2015;6(4):1981–94.
Burns SS, Akhmametyeva EM, Oblinger JL, Bush ML, Huang J, Senner V, et al. Histone deacetylase inhibitor AR-42 differentially affects cell-cycle transit in meningeal and meningioma cells, potently inhibiting NF2-deficient meningioma growth. Cancer Res. 2013;73(2):792–803.
Chang LS, Oblinger JL, Smith AE, Ferrer M, Angus SP, Hawley E, et al. Brigatinib causes tumor shrinkage in both NF2-deficient meningioma and schwannoma through inhibition of multiple tyrosine kinases but not ALK. PLoS ONE. 2021;16(7): e0252048.
Chen K, Si Y, Ou J, Guan JS, Kim S, Ernst P, et al. Antibody-drug conjugate to treat meningiomas. Pharmaceuticals (Basel). 2021;14(5):427.
Karsy M, Hoang N, Barth T, Burt L, Dunson W, Gillespie DL, et al. Combined hydroxyurea and verapamil in the clinical treatment of refractory meningioma: human and orthotopic xenograft studies. World Neurosurg. 2016;86:210–9.
Soto-Montenegro ML, Pena-Zalbidea S, Mateos-Perez JM, Oteo M, Romero E, Morcillo MA, et al. Meningiomas: a comparative study of 68Ga-DOTATOC, 68Ga-DOTANOC and 68Ga-DOTATATE for molecular imaging in mice. PLoS ONE. 2014;9(11): e111624.
Cal-Gonzalez J, Vaquero JJ, Herraiz JL, Perez-Liva M, Soto-Montenegro ML, Pena-Zalbidea S, et al. Improving PET quantification of small animal [(68)Ga]DOTA-labeled PET/CT studies by using a CT-based positron range correction. Mol Imaging Biol. 2018;20(4):584–93.
Dijkstra BM, de Jong M, Stroet MCM, Andreae F, Dulfer SE, Everts M, et al. Evaluation of Ac-Lys(0)(IRDye800CW)Tyr(3)-octreotate as a novel tracer for SSTR(2)-targeted molecular fluorescence guided surgery in meningioma. J Neurooncol. 2021;153(2):211–22.
La Cava F, Fringuello Mingo A, Irrera P, Di Vito A, Cordaro A, Brioschi C, et al. Orthotopic induction of CH157MN convexity and skull base meningiomas into nude mice using stereotactic surgery and MRI characterization. Anim Model Exp Med. 2019;2(1):58–63.
Petermann A, Haase D, Wetzel A, Balavenkatraman KK, Tenev T, Guhrs KH, et al. Loss of the protein-tyrosine phosphatase DEP-1/PTPRJ drives meningioma cell motility. Brain Pathol. 2011;21(4):405–18.
Tanaka K, Sato C, Maeda Y, Koike M, Matsutani M, Yamada K, et al. Establishment of a human malignant meningioma cell line with amplified c-myc oncogene. Cancer. 1989;64(11):2243–9.
Skibinski CG, Williamson T, Riggins GJ. Mebendazole and radiation in combination increase survival through anticancer mechanisms in an intracranial rodent model of malignant meningioma. J Neurooncol. 2018;140(3):529–38.
Takeda H, Okada M, Kuramoto K, Suzuki S, Sakaki H, Sanomachi T, et al. Antitumor activity of gemcitabine against high-grade meningioma in vitro and in vivo. Oncotarget. 2017;8(53):90996–1008.
Kanno H, Nishihara H, Wang L, Yuzawa S, Kobayashi H, Tsuda M, et al. Expression of CD163 prevents apoptosis through the production of granulocyte colony-stimulating factor in meningioma. Neuro Oncol. 2013;15(7):853–64.
Ishiwata I, Ishiwata C, Ishiwata E, Sato Y, Kiguchi K, Tachibana T, et al. In vitro culture of various typed meningiomas and characterization of a human malignant meningioma cell line (HKBMM). Hum Cell. 2004;17(4):211–7.
Michelhaugh SK, Guastella AR, Varadarajan K, Klinger NV, Parajuli P, Ahmad A, et al. Development of patient-derived xenograft models from a spontaneously immortal low-grade meningioma cell line, KCI-MENG1. J Transl Med. 2015;13:227.
Yazaki T, Takamiya Y, Costello PC, Mineta T, Menon AG, Rabkin SD, et al. Inhibition of angiogenesis and growth of human non-malignant and malignant meningiomas by TNP-470. J Neurooncol. 1995;23(1):23–9.
Jalali S, Singh S, Agnihotri S, Wataya T, Salehi F, Alkins R, et al. A role for matrix remodelling proteins in invasive and malignant meningiomas. Neuropathol Appl Neurobiol. 2015;41(2):e16-28.
Saydam O, Senol O, Schaaij-Visser TB, Pham TV, Piersma SR, Stemmer-Rachamimov AO, et al. Comparative protein profiling reveals minichromosome maintenance (MCM) proteins as novel potential tumor markers for meningiomas. J Proteome Res. 2010;9(1):485–94.
Senol O, Schaaij-Visser TB, Erkan EP, Dorfer C, Lewandrowski G, Pham TV, et al. miR-200a-mediated suppression of non-muscle heavy chain IIb inhibits meningioma cell migration and tumor growth in vivo. Oncogene. 2015;34(14):1790–8.
Jungwirth G, Yu T, Moustafa M, Rapp C, Warta R, Jungk C, et al. Identification of KIF11 as a novel target in meningioma. Cancers (Basel). 2019;11(4):545.
Jungwirth G, Yu T, Cao J, Eddine MA, Moustafa M, Warta R, et al. KIF11 inhibitors filanesib and ispinesib inhibit meningioma growth in vitro and in vivo. Cancer Lett. 2021;506:1–10.
Ding Y, Ge Y, Wang D, Liu Q, Sun S, Hua L, et al. LncRNA-IMAT1 promotes invasion of meningiomas by suppressing KLF4/hsa-miR22-3p/Snai1 pathway. Mol Cells. 2022;45(6):388–402.
Nigim F, Esaki S, Hood M, Lelic N, James MF, Ramesh V, et al. A new patient-derived orthotopic malignant meningioma model treated with oncolytic herpes simplex virus. Neuro Oncol. 2016;18(9):1278–87.
Nigim F, Kiyokawa J, Gurtner A, Kawamura Y, Hua L, Kasper EM, et al. A monoclonal antibody against beta1 integrin inhibits proliferation and increases survival in an orthotopic model of high-grade meningioma. Target Oncol. 2019;14(4):479–89.
Yu T, Cao J, Alaa Eddine M, Moustafa M, Mock A, Erkut C, et al. Receptor-tyrosine kinase inhibitor ponatinib inhibits meningioma growth in vitro and in vivo. Cancers (Basel). 2021;13(23):5898.
Linsler S, Muller SJ, Muller A, Senger S, Oertel JM. Fluorescence image-guided resection of intracranial meningioma: an experimental in vivo study on nude mice. Ann Anat. 2021;237: 151752.
Jiang C, Song T, Li J, Ao F, Gong X, Lu Y, et al. RAS promotes proliferation and resistances to apoptosis in meningioma. Mol Neurobiol. 2017;54(1):779–87.
Ding MH, Wang Z, Jiang L, Fu HL, Gao J, Lin XB, et al. The transducible TAT-RIZ1-PR protein exerts histone methyltransferase activity and tumor-suppressive functions in human malignant meningiomas. Biomaterials. 2015;56:165–78.
Friedrich S, Schwabe K, Grote M, Krauss JK, Nakamura M. Effect of systemic celecoxib on human meningioma after intracranial transplantation into nude mice. Acta Neurochir (Wien). 2013;155(1):173–82.
Chen J, Zhang H, Wang H. Experimental study on the inhibitory effects of verapamil on the proliferation of meningiomas cells. J Huazhong Univ Sci Technol Med Sci. 2007;27(1):88–90.
Hu D, Wang X, Mao Y, Zhou L. Identification of CD105 (endoglin)-positive stem-like cells in rhabdoid meningioma. J Neurooncol. 2012;106(3):505–17.
Malham GM, Thomsen RJ, Synek BJ, Baguley BC. Establishment of primary human meningiomas as subcutaneous xenografts in mice. Br J Neurosurg. 2001;15(4):328–34.
Jensen RL, Wurster RD. Calcium channel antagonists inhibit growth of subcutaneous xenograft meningiomas in nude mice. Surg Neurol. 2001;55(5):275–83.
McCutcheon IE, Flyvbjerg A, Hill H, Li J, Bennett WF, Scarlett JA, et al. Antitumor activity of the growth hormone receptor antagonist pegvisomant against human meningiomas in nude mice. J Neurosurg. 2001;94(3):487–92.
Schrell UM, Rittig MG, Anders M, Kiesewetter F, Marschalek R, Koch UH, et al. Hydroxyurea for treatment of unresectable and recurrent meningiomas. I. Inhibition of primary human meningioma cells in culture and in meningioma transplants by induction of the apoptotic pathway. J Neurosurg. 1997;86(5):845–52.
Matsuda Y, Kawamoto K, Kiya K, Kurisu K, Sugiyama K, Uozumi T. Antitumor effects of antiprogesterones on human meningioma cells in vitro and in vivo. J Neurosurg. 1994;80(3):527–34.
Medhkour A, Van Roey M, Sobel RA, Fingert HJ, Lee J, Martuza RL. Implantation of human meningiomas into the subrenal capsule of the nude mouse. A model for studies of tumor growth. J Neurosurg. 1989;71(4):545–50.
Olson JJ, Beck DW, Schlechte JA, Loh PM. Effect of the antiprogesterone RU-38486 on meningioma implanted into nude mice. J Neurosurg. 1987;66(4):584–7.
Markert JM, Coen DM, Malick A, Mineta T, Martuza RL. Expanded spectrum of viral therapy in the treatment of nervous system tumors. J Neurosurg. 1992;77(4):590–4.
Waldt N, Scharnetzki D, Kesseler C, Kirches E, Stroscher N, Bohmer FD, et al. Loss of PTPRJ/DEP-1 enhances NF2/Merlin-dependent meningioma development. J Neurol Sci. 2020;408: 116553.
Morrison JP, Satoh H, Foley J, Horton JL, Dunnick JK, Kissling GE, et al. N-ethyl-N-nitrosourea (ENU)-induced meningiomatosis and meningioma in p16(INK4a)/p19(ARF) tumor suppressor gene-deficient mice. Toxicol Pathol. 2007;35(6):780–7.
Peyre M, Clermont-Taranchon E, Stemmer-Rachamimov A, Kalamarides M. Miniaturized handheld confocal microscopy identifies focal brain invasion in a mouse model of aggressive meningioma. Brain Pathol. 2013;23(4):371–7.
Boetto J, Apra C, Bielle F, Peyre M, Kalamarides M. Selective vulnerability of the primitive meningeal layer to prenatal Smo activation for skull base meningothelial meningioma formation. Oncogene. 2018;37(36):4955–63.
Szulzewsky F, Arora S, Arakaki AKS, Sievers P, Almiron Bonnin DA, Paddison PJ, et al. Both YAP1-MAML2 and constitutively active YAP1 drive the formation of tumors that resemble NF2 mutant meningiomas in mice. Genes Dev. 2022;36(13–14):857–70.
Yeung J, Yaghoobi V, Miyagishima D, Vesely MD, Zhang T, Badri T, et al. Targeting the CSF1/CSF1R axis is a potential treatment strategy for malignant meningiomas. Neuro Oncol. 2021;23(11):1922–35.
Toktas ZO, Akgun E, Ozkan A, Bozkurt SU, Bekiroglu N, Seker A, et al. Relationship of angiogenic potential with clinical features in cranial meningiomas: a corneal angiogenesis study. Neurosurgery. 2010;67(6):1724–32 (discussion 32).
Kilic K, Avsar T, Akgun E, Ozkan A, Toktas ZO, Seker A, et al. Gamma knife radiosurgery inhibits angiogenesis of meningiomas: in vivo rat corneal assay. World Neurosurg. 2013;80(5):598–604.
Brooks SE, Adachi M, Hoffman LM, Stein MR, Brooks J, Schneck L. Induction of lymphomas and fibrosarcomas in nude mice after implantation of simian virus 40-transformed human meningioma. Lab Invest. 1988;58(5):518–23.
Baia GS, Caballero OL, Orr BA, Lal A, Ho JS, Cowdrey C, et al. Yes-associated protein 1 is activated and functions as an oncogene in meningiomas. Mol Cancer Res. 2012;10(7):904–13.
Hueng DY, Sytwu HK, Huang SM, Chang C, Ma HI. Isolation and characterization of tumor stem-like cells from human meningiomas. J Neurooncol. 2011;104(1):45–53.
Rath P, Miller DC, Litofsky NS, Anthony DC, Feng Q, Franklin C, et al. Isolation and characterization of a population of stem-like progenitor cells from an atypical meningioma. Exp Mol Pathol. 2011;90(2):179–88.
Yamate J, Tajima M, Saitoh T, Shibuya K. Biological behaviour and morphological characteristics of a transplantable tumour (MM-KMY) derived from a malignant meningioma in an F344 rat. J Comp Pathol. 1994;111(3):243–57.
Tsujino K, Yamate J, Tsukamoto Y, Kumagai D, Kannan Y, Jippo T, et al. Establishment and characterization of cell lines derived from a transplantable rat malignant meningioma: morphological heterogeneity and production of nerve growth factor. Acta Neuropathol. 1997;93(5):461–70.
Yang QE. Human cancer xenografts in immunocompromised mice provide an advanced genuine tumor model for research and drug development: a revisit of murine models for human cancers. Biochim Biophys Acta Gen Subj. 2021;1865(8): 129929.
Tsai JC, Goldman CK, Gillespie GY. Vascular endothelial growth factor in human glioma cell lines: induced secretion by EGF, PDGF-BB, and bFGF. J Neurosurg. 1995;82(5):864–73.
Tsai JC, Hsiao YY, Teng LJ, Shun CT, Chen CT, Goldman CK, et al. Regulation of vascular endothelial growth factor secretion in human meningioma cells. J Formos Med Assoc. 1999;98(2):111–7.
Baia GS, Slocum AL, Hyer JD, Misra A, Sehati N, VandenBerg SR, et al. A genetic strategy to overcome the senescence of primary meningioma cell cultures. J Neurooncol. 2006;78(2):113–21.
Yamazaki S, Ohka F, Hirano M, Shiraki Y, Motomura K, Tanahashi K, et al. Newly established patient-derived organoid model of intracranial meningioma. Neuro Oncol. 2021. https://doi.org/10.1093/neuonc/noab155.
Mei Y, Bi WL, Greenwald NF, Agar NY, Beroukhim R, Dunn GP, et al. Genomic profile of human meningioma cell lines. PLoS ONE. 2017;12(5): e0178322.
Akat K, Mennel HD, Kremer P, Gassler N, Bleck CK, Kartenbeck J. Molecular characterization of desmosomes in meningiomas and arachnoidal tissue. Acta Neuropathol. 2003;106(4):337–47.
Metzger D, Chambon P. Site- and time-specific gene targeting in the mouse. Methods. 2001;24(1):71–80.
von Werder A, Seidler B, Schmid RM, Schneider G, Saur D. Production of avian retroviruses and tissue-specific somatic retroviral gene transfer in vivo using the RCAS/TVA system. Nat Protoc. 2012;7(6):1167–83.
Hughes SH, Greenhouse JJ, Petropoulos CJ, Sutrave P. Adaptor plasmids simplify the insertion of foreign DNA into helper-independent retroviral vectors. J Virol. 1987;61(10):3004–12.
Perry A, Giannini C, Raghavan R, Scheithauer BW, Banerjee R, Margraf L, et al. Aggressive phenotypic and genotypic features in pediatric and NF2-associated meningiomas: a clinicopathologic study of 53 cases. J Neuropathol Exp Neurol. 2001;60(10):994–1003.
Yuzawa S, Nishihara H, Tanaka S. Genetic landscape of meningioma. Brain Tumor Pathol. 2016;33(4):237–47.
Goutagny S, Kalamarides M. Meningiomas and neurofibromatosis. J Neurooncol. 2010;99(3):341–7.
Bi WL, Greenwald NF, Abedalthagafi M, Wala J, Gibson WJ, Agarwalla PK, et al. Genomic landscape of high-grade meningiomas. NPJ Genom Med. 2017. https://doi.org/10.1038/s41525-017-0014-7.
Mello SS, Attardi LD. Deciphering p53 signaling in tumor suppression. Curr Opin Cell Biol. 2018;51:65–72.
Barresi V, Simbolo M, Fioravanzo A, Piredda ML, Caffo M, Ghimenton C, et al. Molecular profiling of 22 primary atypical meningiomas shows the prognostic significance of 18q heterozygous loss and CDKN2A/B homozygous deletion on recurrence-free survival. Cancers (Basel). 2021;13(4):903.
Sievers P, Hielscher T, Schrimpf D, Stichel D, Reuss DE, Berghoff AS, et al. CDKN2A/B homozygous deletion is associated with early recurrence in meningiomas. Acta Neuropathol. 2020;140(3):409–13.
Matsuoka M, Kurita M, Sudo H, Mizumoto K, Nishimoto I, Ogata E. Multiple domains of the mouse p19ARF tumor suppressor are involved in p53-independent apoptosis. Biochem Biophys Res Commun. 2003;301(4):1000–10.
Ruas M, Peters G. The p16INK4a/CDKN2A tumor suppressor and its relatives. Biochim Biophys Acta. 1998;1378(2):F115–77.
Prives C, Hall PA. The p53 pathway. J Pathol. 1999;187(1):112–26.
Xia Y, Liu Y, Yang C, Simeone DM, Sun TT, DeGraff DJ, et al. Dominant role of CDKN2B/p15INK4B of 9p21.3 tumor suppressor hub in inhibition of cell-cycle and glycolysis. Nat Commun. 2021;12(1):2047.
Louis DN, Perry A, Wesseling P, Brat DJ, Cree IA, Figarella-Branger D, et al. The 2021 WHO classification of tumors of the central nervous system: a summary. Neuro Oncol. 2021;23(8):1231–51.
Maxwell M, Galanopoulos T, Hedley-Whyte ET, Black PM, Antoniades HN. Human meningiomas co-express platelet-derived growth factor (PDGF) and PDGF-receptor genes and their protein products. Int J Cancer. 1990;46(1):16–21.
Black PM, Carroll R, Glowacka D, Riley K, Dashner K. Platelet-derived growth factor expression and stimulation in human meningiomas. J Neurosurg. 1994;81(3):388–93.
Brastianos PK, Horowitz PM, Santagata S, Jones RT, McKenna A, Getz G, et al. Genomic sequencing of meningiomas identifies oncogenic SMO and AKT1 mutations. Nat Genet. 2013;45(3):285–9.
Sievers P, Chiang J, Schrimpf D, Stichel D, Paramasivam N, Sill M, et al. YAP1-fusions in pediatric NF2-wildtype meningioma. Acta Neuropathol. 2020;139(1):215–8.
Passaniti A, Kleinman HK, Martin GR. Matrigel: history/background, uses, and future applications. J Cell Commun Signal. 2022;16(4):621–6.
Fridman R, Giaccone G, Kanemoto T, Martin GR, Gazdar AF, Mulshine JL. Reconstituted basement membrane (matrigel) and laminin can enhance the tumorigenicity and the drug resistance of small cell lung cancer cell lines. Proc Natl Acad Sci USA. 1990;87(17):6698–702.
Fridman R, Sweeney TM, Zain M, Martin GR, Kleinman HK. Malignant transformation of NIH-3T3 cells after subcutaneous co-injection with a reconstituted basement membrane (matrigel). Int J Cancer. 1992;51(5):740–4.
Pompili L, Porru M, Caruso C, Biroccio A, Leonetti C. Patient-derived xenografts: a relevant preclinical model for drug development. J Exp Clin Cancer Res. 2016;35(1):189.
Day CP, Merlino G, Van Dyke T. Preclinical mouse cancer models: a maze of opportunities and challenges. Cell. 2015;163(1):39–53.
Kersten K, de Visser KE, van Miltenburg MH, Jonkers J. Genetically engineered mouse models in oncology research and cancer medicine. EMBO Mol Med. 2017;9(2):137–53.
Jonkers J, Berns A. Conditional mouse models of sporadic cancer. Nat Rev Cancer. 2002;2(4):251–65.
Ahronian LG, Lewis BC. Using the RCAS-TVA system to model human cancer in mice. Cold Spring Harb Protoc. 2014;2014(11):1128–35.
Williams SJ, Prescher JA. Building biological flashlights: orthogonal luciferases and luciferins for in vivo imaging. Acc Chem Res. 2019;52(11):3039–50.
Risbridger GP, Lawrence MG. Towards best practice in establishing patient-derived xenografts. In: Wang Y, Lin D, Gout PW, editors. Patient-derived xenograft models of human cancer. Cham: Springer International Publishing; 2017. p. 11–28.
Hahn SA, Seymour AB, Hoque AT, Schutte M, da Costa LT, Redston MS, et al. Allelotype of pancreatic adenocarcinoma using xenograft enrichment. Cancer Res. 1995;55(20):4670–5.
Lin MT, Tseng LH, Kamiyama H, Kamiyama M, Lim P, Hidalgo M, et al. Quantifying the relative amount of mouse and human DNA in cancer xenografts using species-specific variation in gene length. Biotechniques. 2010;48(3):211–8.
Iwami K, Momota H, Natsume A, Kinjo S, Nagatani T, Wakabayashi T. A novel method of intracranial injection via the postglenoid foramen for brain tumor mouse models. J Neurosurg. 2012;116(3):630–5.
ter Riet G, Korevaar DA, Leenaars M, Sterk PJ, Van Noorden CJ, Bouter LM, et al. Publication bias in laboratory animal research: a survey on magnitude, drivers, consequences and potential solutions. PLoS ONE. 2012;7(9): e43404.
Schieffer KM, Agarwal V, LaHaye S, Miller KE, Koboldt DC, Lichtenberg T, et al. YAP1-FAM118B fusion defines a rare subset of childhood and young adulthood meningiomas. Am J Surg Pathol. 2021;45(3):329–40.
Charan J, Kantharia ND. How to calculate sample size in animal studies? J Pharmacol Pharmacother. 2013;4(4):303–6.
Shahbandi A, Shah DS, Hadley CC, Patel AJ. The role of pharmacotherapy in treatment of meningioma: a systematic review. Cancers (Basel). 2023;15(2):483.
Jensen LR, Maier AD, Lomstein A, Graillon T, Hrachova M, Bota D, et al. Somatostatin analogues in treatment-refractory meningioma: a systematic review with meta-analysis of individual patient data. Neurosurg Rev. 2022;45(5):3067–81.
Kaley T, Barani I, Chamberlain M, McDermott M, Panageas K, Raizer J, et al. Historical benchmarks for medical therapy trials in surgery- and radiation-refractory meningioma: a RANO review. Neuro Oncol. 2014;16(6):829–40.
Begley CG, Ioannidis JP. Reproducibility in science: improving the standard for basic and preclinical research. Circ Res. 2015;116(1):116–26.
Begley CG, Ellis LM. Drug development: raise standards for preclinical cancer research. Nature. 2012;483(7391):531–3.
Krauth D, Woodruff TJ, Bero L. Instruments for assessing risk of bias and other methodological criteria of published animal studies: a systematic review. Environ Health Perspect. 2013;121(9):985–92.
Morton JJ, Bird G, Refaeli Y, Jimeno A. Humanized mouse xenograft models: narrowing the tumor-microenvironment gap. Cancer Res. 2016;76(21):6153–8.
Shultz LD, Lyons BL, Burzenski LM, Gott B, Chen X, Chaleff S, et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J Immunol. 2005;174(10):6477–89.
Rachinger W, Stoecklein VM, Terpolilli NA, Haug AR, Ertl L, Poschl J, et al. Increased 68Ga-DOTATATE uptake in PET imaging discriminates meningioma and tumor-free tissue. J Nucl Med. 2015;56(3):347–53.
Afshar-Oromieh A, Giesel FL, Linhart HG, Haberkorn U, Haufe S, Combs SE, et al. Detection of cranial meningiomas: comparison of (6)(8)Ga-DOTATOC PET/CT and contrast-enhanced MRI. Eur J Nucl Med Mol Imaging. 2012;39(9):1409–15.
Kowalski ES, Khairnar R, Gryaznov AA, Kesari V, Koroulakis A, Raghavan P, et al. (68)Ga-DOTATATE PET-CT as a tool for radiation planning and evaluating treatment responses in the clinical management of meningiomas. Radiat Oncol. 2021;16(1):151.
Woo XY, Giordano J, Srivastava A, Zhao ZM, Lloyd MW, de Bruijn R, et al. Conservation of copy number profiles during engraftment and passaging of patient-derived cancer xenografts. Nat Genet. 2021;53(1):86–99.
McClatchey AI, Saotome I, Ramesh V, Gusella JF, Jacks T. The Nf2 tumor suppressor gene product is essential for extraembryonic development immediately prior to gastrulation. Genes Dev. 1997;11(10):1253–65.
Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 2013;339(6121):819–23.
Maresch R, Mueller S, Veltkamp C, Ollinger R, Friedrich M, Heid I, et al. Multiplexed pancreatic genome engineering and cancer induction by transfection-based CRISPR/Cas9 delivery in mice. Nat Commun. 2016;7:10770.
Acknowledgements
We would like to thank Claire Gudex, University of Southern Denmark, for proofreading the paper and Morten Winkler Møller, Department of Neurosurgery, Odense University Hospital, for manuscript discussions throughout the process.
Funding
Open access funding provided by University Library of Southern Denmark.
Author information
Authors and Affiliations
Contributions
MSA: conceptualization, methodology, screening, full-text data extraction, analyses, figures, and tables; MSK: screening, full-text data extraction; ASPM: conceptualization, methodology; FRP: conceptualization, methodology, screening; BH: animal protocol assessment; BBO; cell-protocol assessment. All authors performed review, writing, editing, and approved final draft.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
The PRISMA checklist.
Additional file 2:
The PRISMA abstract checklist.
Additional file 3:
Uploaded PROSPERO protocol.
Additional file 4:
Protocol breaches.
Additional file 5:
Full original search strategy.
Additional file 6:
Data extraction fields.
Additional file 7:
CRIME-Q: Full spread sheet.
Additional file 8:
CRIME-Q: Predetermined list of information.
Additional file 9:
ECLM full data extraction sheet.
Additional file 10:
PTM full data extraction sheet.
Additional file 11:
GEM full data extraction sheet.
Additional file 12:
Uncategorized full data extraction sheet.
Additional file 13:
Meta- and subgroup analyses.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Andersen, M.S., Kofoed, M.S., Paludan-Müller, A.S. et al. Meningioma animal models: a systematic review and meta-analysis. J Transl Med 21, 764 (2023). https://doi.org/10.1186/s12967-023-04620-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-023-04620-7