
Abstract
Background
The salinity threat represents an environmental challenge that drastically affects plant growth and yield. Besides salinity stress, the escalating world population will greatly influence the world’s food security in the future. Therefore, searching for effective strategies to improve crop salinity resilience and sustain agricultural productivity under high salinity is a must. Seed priming is a reliable, simple, low-risk, and low-cost technique. Therefore, this work aimed to evaluate the impact of seed priming with 0.5 mM NaHS, as a donor of H2S, in mitigating salinity effects on sunflower seedlings. Primed and nonprime seeds were established in nonsaline soil irrigated with tape water for 14 d, and then exposed to 150 mM NaCl for 7 d.
Results
Salinity stress significantly reduced the seedling growth, biomass accumulation, K+, Ca2+, and salinity tolerance index while elevating Na+ uptake and translocation. Salinity-induced adverse effects were significantly alleviated by H2S priming. Upregulation in gene expression (HaSOS2, HaGST) under NaCl stress was further enhanced by H2S priming. Also, H2S reduced lipid peroxidation, electrolyte leakage, and H2O2 content, but elevated the antioxidant defense system. NaCl-induced levels of ascorbate, glutathione, and α tocopherol, as well as the activities of AsA-GSH cycle enzymes: ascorbate peroxidase, monodehydroascorbate reductase, dehydroascorbate reductase, glutathione reductase, and glutathione S-transferase, were further enhanced by H2S priming. Increased level of H2S and total thiol by NaCl was also further stimulated by H2S priming.
Conclusion
H2S priming has proved to be an efficient strategy to improve sunflower seedlings’ salinity tolerance by retaining ion homeostasis, detoxifying oxidative damage, modulating gene expression involved in ion homeostasis and ROS scavenging, and boosting endogenous H2S. These findings suggested that H2S acts as a regulatory molecule activating the functional processes responsible for sunflower adaptive mechanisms and could be adopted as a crucial crop management strategy to combat saline conditions. However, it would be of great interest to conduct further studies in the natural saline field to broaden our understanding of crop adaptive mechanisms and to support our claims.
Similar content being viewed by others
Background
Salinity is one of the most critical environmental constraints, affecting plant growth, development, and productivity, especially in the arid and semi-arid regions. High soil salinity has arisen as an important global concern, which hampers sustainable crop production in many agrarian countries like Egypt. In addition, water shortage, hot and dry climate, and rising sea levels due to global warming aggravate the existing salinity problems, thus worsening crop production in inland and coastal areas [1]. Excessive salinity can induce various negative effects in plant cells, including ionic, osmotic, and oxidative stress [2]. Salinity stress builds up toxic Na+ and Cl− while decreasing beneficial K+ and Ca2+ levels, which causes the ionic impact of salinity. High salt concentrations in the soil lower its water potential, which imposes water deficit or osmotic stress of salinity. Plants therefore have difficulty absorbing water from the soil. It is worth mentioning that osmotic stress shows its effects at short-term salinity exposure whereas ionic component effects prevail at salinity long exposure [1]. Further, high salinity induces the overproduction of reactive oxygen species (ROS) that oxidize cellular proteins, nucleic acids, and lipids. Plant cells minimize the ROS level because ROS may act as protective agents, not always damaging, as they play a role in signaling pathways and thus stress tolerance under saline conditions, which depend upon ROS concentration and time [3].
Creating salinity-tolerant crop germplasm is becoming urgent to counterbalance and avoid amplifying these hazardous issues of salinity stress. Salinity tolerance in crops is a physiologically multifaceted trait and is controlled by multiple mechanisms. One crucial mechanism to minimize the deleterious effects of toxic ions is to regulate ion concentrations in response to saline conditions, i.e., ion homeostasis [4]. That is, plants need to reduce Na+ content to avoid cytoplasmic injury and toxicity and to retain continued uptake of K+ under saline environments, which takes place through minimizing Na+ uptake by roots and/or increasing Na+ efflux back to the soil, intracellular Na+ sequestration, K+ retention in the cytosol, control of xylem ion loading and excluding Na+ from the shoot [5,6,7]. The Salt Overly Sensitive (SOS) pathway is essential for ion homeostasis and plant adaptation to salinity stress [8]. Among the three proteins (SOS1, SOS2, SOS3) of the SOS pathway, the protein kinase SOS2 functions as a network hub in the SOS pathway, and its kinase activity is rapidly activated by salinity stress, interacting with SOS3 and forms a complex in the cell membrane, which in turn activates the SOS1 antiporter [8, 9]. This antiporter plays a key role in Na+ exclusion to outer spaces and in controlling long-distance Na+ transport from the root to the shoot [5, 10]. Also, SOS2 has been illustrated to increase both the transcription of the SOS1 gene and the activity of the SOS1 protein through direct protein–protein interaction [9], indicating a central role in SOS-mediated Na+ extrusion. Additionally, Verslues et al. [11] demonstrated that SOS2 interacts with nucleoside diphosphate kinase 2 and with catalases 2 and 3, suggesting that SOS2 is part of a signaling node connecting salinity stress response with ROS signaling. Other plasma membrane channels and transporters that have key functions in K+ maintenance and ion homeostasis under high salinity include AKT1 (K+ in channels) and KUP1/HAK/KT (high-affinity K+-H+ symporters) [12].
Owing to the lowering of the soil water potential induced by high concentrations of salts in the soil solution, plants must reduce their water potential to maintain water gradient and subsequently continue water absorption. Plants therefore cope with this osmotic action of salinity by the accumulation of organic/inorganic solutes; that is an osmotic adjustment [13, 14]. These compatible solutes include soluble sugars, polyols, proline, glycine betaine, polyamines, and phenolic compounds, which are innocent to cellular metabolism even at high concentrations [14, 15]. Inorganic ions contribute to osmotic adjustment including Na+, K+, and Cl−, which constitute 80–95% of the osmotic pressure of the cell sap in halophytes and in glycophytes contribute between 50 and 70% [4].
Salinity-triggered oxidative stress through the production of ROS in chloroplasts, mitochondria, peroxisomes, plasma membrane, and apoplast, is a secondary stress [16]. Therefore, another component of salinity tolerance is ROS detoxification and scavenging which is closely related to the maintenance of cellular redox balance by activating and upregulating the antioxidant system including non-enzymatic antioxidants and enzymatic antioxidants [2, 14, 15]. Superoxide dismutase, catalase, glutathione reductase, and peroxidase are examples of enzymatic antioxidants, while non-enzymatic antioxidants include ascorbic acid, glutathione, glycine betaine, proline, carotenoids, total phenolics, flavonoids, and tocopherols [15, 16]. Enzymatic and nonenzymatic defense systems thus scavenge ROS resulting in maintained cell ultrastructures and hence participate in salinity tolerance. In particular, the activation of the ascorbate–glutathione (AsA-GSH) cycle and modulation of the content of ascorbic acid and glutathione have been shown to play central roles in the stability of redox homeostasis and thus crop salinity resilience [17].
Seed priming has been reported as one of the most promising strategies for improving crop growth, development, and productivity, and balancing ionic homeostasis under salinity stress [18,19,20]. Seed priming with different agents has been successfully adopted to alleviate the adverse effects and induce crop tolerance and productivity to various stresses [21,22,23]. Hydrogen sulfide (H2S) has recently emerged as an important gaseous multifunctional signaling molecule regulating a myriad of physiological processes in plants and is a powerful tool in modifying plants’ adaptability against multiple abiotic stresses [22, 24,25,26]. Further, H2S promotes stress tolerance to abiotic stress by reinstating redox equilibrium, increasing osmolyte buildup, preserving ion balance, modulating gene expression, and regulating ROS-processing systems by transcriptional or posttranslational modifications [27,28,29,30,31,32].
Sunflower (Helianthus annuus L.) is an economically important oilseed crop, which ranks the fourth largest source of edible oil after soybean, rapeseed, and safflower [33]. Sunflower is a high-yielding oilseed crop and has the potential to bridge the gap existing between consumption and domestic production of edible oil in Egypt. Egypt's self-sufficiency of palatable vegetable oils during the 1960s was 95%, which declined to 31.6% in 2007 [34]. However, it is reported that sunflower growing in the central Egyptian province of Fayoum brings economic hope to farmers and contributes to reducing an exorbitant vegetable oil import bill [33, 34]. This necessitates researchers in Egypt and the Mediterranean region to search for approaches to improve sunflower crop performance under saline conditions and hence exploit the saline soils and elevate oil production. This for sure will help to reduce the oil gap in Egypt. In addition, the sunflower crop is not only used in the feeding of humans but also for industrial and energy uses as well as a high-quality forage by livestock producers [35]. Despite priming and foliar application of H2S have been illustrated to have an ameliorative impact on crop species in response to various abiotic stresses [22, 26, 30, 31], to the best of our knowledge, the alleviating impact of seed priming with H2S on overcoming the adverse effects of saline conditions on sunflower has not been studied yet in detail. Therefore, works aiming to analyze the ameliorative mechanisms triggered by H2S could be a novel approach to enhance crop resilience and productivity under salinity stress, which might contribute to global food security. Herein, we thus provide a comprehensive assessment of the impact of seed priming with H2S on sunflower growth, the AsA-GSH system, ion homeostasis, and enzymatic and nonenzymatic antioxidants. In addition, the H2S priming impact on HaSOS2 and HaGST gene expression modulation of sunflower seedlings exposed to NaCl stress has also been investigated. Our study thus characterizes the underlying processes contributing to seed H2S priming-induced salinity tolerance in sunflower seedlings, with particular emphasis on ion homeostasis, regulating H2S metabolism, modulating gene expression, and detoxifying oxidative stress. The results indicated that H2S priming upregulated expression levels of genes related to ion homeostasis and antioxidant system, SOS pathway, and GSH pathway, which largely contributed to enhanced sunflower tolerance to high salinity.
Results
Growth parameters, Na+uptake, Na+ translocation, and salinity tolerance index (STI)
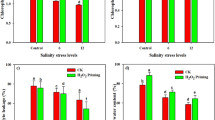
The shoot length, root length, shoot, and root FW, and shoot and root DW were significantly reduced by 21.9%, 29.3%, 16.4%, 19.7%, 12.7, and 53.6% under salinity stress, respectively, compared with their controls (Fig. 1a-c). However, applying NaHS significantly ameliorated these parameters relative to their corresponding stressed seedlings (Fig. 1a-c). The NaCl-stressed sunflower seedlings recorded the greatest Na+ uptake and translocation reaching about 277.9% and 80%, respectively, compared with the control ones (Fig. 1d, e). NaHS pretreatment significantly diminished Na+ uptake and translocation by 22.9% and 9%, respectively, compared with the NaCl-stressed alone plants (Fig. 1d, e). In response to NaCl stress, a significant reduction in STI percentage to 24.1 of the value of control seedlings was observed, while NaHS priming significantly enhanced the STI percentage of NaCl-stressed seedlings up to 46.5 (Fig. 1f).

Effect of NaHS priming in the presence and absence of 150 mM NaCl on the shoot and root (a) length, (b) FW, (c) DW, (d) sodium uptake, (e) sodium translocation, and (f) salinity tolerance index (STI) of sunflower shoot and root. Each value is the mean ± SD of three replicates. Bars with different letters indicate significant differences at P ≤ 0.05
Mineral contents and HaSOS2 gene expression
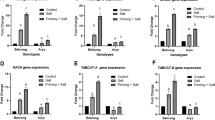
NaCl stress significantly increased the Na+ accumulation in the shoots (500%) and roots (233%) of sunflower seedlings relative to their controls, while decreasing the K+ content in the shoots (36.8%) and roots (58%) resulting in higher Na+/K+ ratios (3.12% shoot, 19.1% root) (Fig. 2a-c). NaHS priming significantly reduced the Na+ content (31.8% shoot, 25% root), and increased K+ content (24% shoot, 117% root), thus lowering Na+/K+ ratios (53.9% shoot, 74.6% root) compared with NaCl-stressed alone (Fig. 2a-c). The contents of Ca2+, Mg2+, and P were decreased by 36.5%, 33%, and 33.3%, respectively, in the shoots of salinity-stressed plants relative to their controls. As for the roots, salinity stress caused a decrease in the contents of Ca2+, Mg2+, and P by 48.7%, 38.3%, and 63.6%, respectively, relative to their controls (Fig. 2d-f). In comparison with salinity-stressed plants, NaHS-pretreated salinity-stressed plants showed increased Ca2+, Mg2+, and P contents by 40.7%, 20.7%, and 24.1%, respectively, in the shoots and by 39.5%, 67.9%, and 83.3%, respectively, in the roots (Fig. 2d-f). Real-time RT-PCR analysis showed that NaCl treatment significantly increased the transcript level of the HaSOS2 gene by 209.5% in sunflower roots compared with their control (Fig. 2g). Further elevation in the relative expression of the HaSOS2 gene by 71.4% was obtained in salinity-stressed roots treated with NaHS compared with seedlings treated with NaCl alone (Fig. 2g).

Effect of NaHS priming in the presence and absence of 150 mM NaCl on (a) Na+ content, (b) K+ content, (c) Na+/K+ ratio, (d) Ca2+ content, (e) Mg.2+ content, (f) P content, and (g) SOS2 gene expression of sunflower shoot and root. Each value is the mean ± SD of three replicates. Bars with different letters indicate significant differences at P ≤ 0.05
H2O2, malondialdehyde (MDA), electrolyte leakage (EL), and 2,2‐diphenyl‐1‐picrylhydrazyl (DPPH)
Salinity stress significantly enhanced the contents of H2O2 by 57.3% and 49.8% and that of MDA by 96.1% and 60% in the shoots and roots, respectively, compared with their controls (Fig. 3a, b). On the other hand, NaHS seed priming before salinity treatment reduced H2O2 (38.3% and 18.4%) and MDA (40% and 33.3%) contents in shoots and roots, respectively, compared with the seedlings that received only NaCl stress (Fig. 3a, b). Although exposure of the sunflower seedlings to NaCl stress significantly increased EL by 77.1% in the shoots and by 28.4% in the roots relative to their controls, NaHS priming significantly reduced the EL to 18.1% and 25.5% in the shoots and roots, respectively, compared with the plants received NaCl only (Fig. 3c). Salinity stress significantly reduced DPPH radical scavenging activity by 21.9% in the shoots and by 20.8% in the roots relative to their controls (Fig. 3d) while NaHS pretreatment significantly restored DPPH radical scavenging activity to the control level in the shoots and roots of salinity-stressed sunflower seedlings (Fig. 3d).

Effect of NaHS priming in the presence and absence of 150 mM NaCl on (a) H2O2 content, (b) malondialdehyde (MDA), (c) electrolyte leakage (EL), and (d) DPPH scavenging percentage of sunflower shoot and root. Each value is the mean ± SD of three replicates. Bars with different letters indicate significant differences at P ≤ 0.05
Antioxidant enzyme activities and HaGST gene expression
The activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) were enhanced by 102.2%, 577.3%, and 309.6% in the shoots and by 42%, 130.6%, and 72% in the roots of NaCl-treated plants, respectively, concerning their respective controls (Fig. 4a-c). On the other hand, NaHS seed priming significantly decreased SOD, CAT, and PX activities in the shoots by 26.9%, 48.3%, and 34.5%, and in the roots by 41.2%, 35.4%, and 42.9%, respectively, as compared with NaCl-treated seedlings (Fig. 4a-c). As for the enzymes of the ascorbate–glutathione (AsA-GSH) cycle, salinity stress significantly enhanced the activities of ascorbate peroxidase (APX), Monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and glutathione reductase (GR) in the shoots by 48.5%, 50%, 66.7%, and 91.7% and in the roots by 21.3%, 100%, 166.7%, and 75%, respectively, relative to those of the controls (Fig. 4d-g). NaHS pretreatment displayed further enhancement in the APX, MDHAR, DHAR, and GR activities for the shoots by 30.6%, 66.7%, 35%, and 43.5% as well as for the roots by 15.8%, 100% 12.5%, and 42.9%, respectively, in comparison with NaCl-treated plants (Fig. 4d-g). Similarly, under NaCl-free conditions, NaHS priming significantly increased the enzyme activities of the AsA-GSH cycle relative to their untreated controls (Fig. 4d-g).

Effect of NaHS priming in presence and absence of 150 mM NaCl on (a) superoxide dismutase (SOD), (b) catalase (CAT), (c) peroxidase (POD), (d) ascorbate peroxidase (APX), (e) monodehydroascorbate reductase (MDHAR), (f) dehydroascorbate reductase (DHAR), (g) glutathione reductase (GR), (h) glutathione S-transferase (GST), and (i) GST gene expression of sunflower shoot and root. Each value is the mean ± SD of three replicates. Bars with different letters indicate significant differences at P ≤ 0.05
For GST as one of the GSH-metabolizing enzymes, salinity stress significantly enhanced the GST activity in sunflower shoots and roots by 40% and 23.6%, respectively, relative to their controls, and NaHS priming further increased GST activity reaching 25% and 56.2% in the shoots and roots, respectively, compared with NaCl-treated plants (Fig. 4h). Real-time RT-PCR analysis showed that NaCl treatment resulted in significant increases in HaGST transcripts (102.4% and 124.3% in the shoots and roots, respectively), whereas NaHS priming exhibited further enhancement in HaGST transcription level by 22.1% and 18.2% in both stressed shoots and roots, respectively, relative to plants received only NaCl treatment (Fig. 4i).
Nonenzymatic antioxidants, endogenous H2S, and total thiol (TT)
Compared with untreated plants, a significant increase in the concentration of ascorbic acid (AsA) (69.9% shoot, 35.4% root) and reduced glutathione (GSH) (87.5% shoot, 22.9% root) were observed in the shoots and roots of NaCl-stressed plants (Fig. 5a, b). NaHS priming further enhanced the levels of AsA and GSH by 72.1% and 20.8% in the shoots and by 77.2% and 28.3% in the roots, respectively, relative to the seedlings treated with NaCl only (Fig. 5a, b). Additionally, salinity stress significantly promoted α-tocopherol level in the shoots by 155.1% over the untreated control, and NaHS priming further elevated α-tocopherol level by 65.6% in the stressed seedlings compared with those received only NaCl stress (Fig. 5c). The roots of sunflower seedlings showed no α-tocopherol content in response to the different treatments (Fig. 5c). On the other hand, NaHS priming significantly reduced the NaCl-induced accumulation of total phenolic content (TPC) in the roots (67.5%) (Fig. 5d). Compared with the controls, NaCl treatment stimulated the endogenous H2S level in the shoots (134.8%) and roots (188.6%) which was further boosted by NaHS priming to 252.8% and 693.5% in the shoots and roots, respectively, relative to the NaCl-stressed seedlings (Fig. 5e). NaHS pretreatment alone also increased the endogenous H2S level in the shoot (111.6%) and in the roots (180.3%) compared with the plants that received no NaCl stress (Fig. 5e). NaCl-induced elevation in the TT content of the sunflower shoots and roots was further increased by NaHS priming relative to seedlings grown under saline and non-saline conditions (Fig. 5f).

Effect of NaHS priming in the presence and absence of 150 mM NaCl on (a) ascorbic acid content (AsA), (b) reduced glutathione (GSH), (c) α-tocopherol content, (d) total phenolic content (TPC), (e) H2S content, and (f) total thiols (TT) of sunflower shoot and root. Each value is the mean ± SD of three replicates. Bars with different letters indicate significant differences at P ≤ 0.05
Discussion
One of the most negative impacts of salinity stress is the reduction in plant growth and productivity despite the escalating demand for food all over the world [13, 36]. NaCl-induced growth reduction in sunflower seedlings observed in this study was most probably resulted from Na+ toxicity [36, 37], declined essential nutrient elements [38,39,40], reduced water absorption resulting from osmotic stress [41], and/or high ROS production [40, 42, 43]. These salinity-hazardous impacts adversely affect crop species by impairing various physiological, biochemical, and molecular processes which were reflected in growth and yield reduction [16, 40, 43, 44]. Our study showed that seed priming with NaHS significantly alleviated the deleterious effects of salinity on sunflower seedling growth, which is consistent with earlier studies by Sun and Luo [45] in cucumber, Chen et al. [46] in barley, Chen et al. [43] in Cyclocarya paliurus, Dawood et al. [39] in common bean, and Ding et al. [47] in wheat, who reported improved seed germination, biomass production, growth, and salinity tolerance of crop plants pretreated with NaHS under saline conditions. The ameliorative effect of H2S observed in the current work is most likely attributed to H2S impact on the retention of essential minerals (e.g., K+, Ca2+, Mg2+), decreased Na+ uptake and translocation that may effectively activate various events associated with stress adaptation and enhanced growth. Similarly, a strong correlation between salinity stress tolerance and crop ability to prevent NaCl-induced K+ leak from roots and retain low Na+ concentration by H2S application has been reported by other researchers in different crops [39, 48,49,50,51]. Additionally, H2S priming-stimulated ion homeostasis was possibly a major factor that participated in the improved salinity tolerance index of sunflower seedlings reported in this study, which agrees with previous works that demonstrate NaHS-enhanced salinity tolerance is associated with ion homeostasis in various crop species [46, 49,50,51,52,53].
Maintenance of ion homeostasis and enough nutrients are critically important for plants to preserve their structure and vital physiological activities under salinity stress [10]. Exposure of sunflower seedlings to 150 mM NaCl induced excessive Na+ accumulation and K+ insufficiency resulting in an increased Na+/K+ ratio, and reduced levels of Ca2+, Mg2+, and P. This disturbed ion homeostasis in response to NaCl exposure has been previously reported in other crops [38, 47, 51, 54]. The result could be attributed to the antagonistic impact of Na+ on K+ binding sites of the root plasma membranes [55], salinity-induced plasma membrane depolarization-initiated K+ leak [47, 56], the competitive uptake of Na+ and Cl− with Ca2+, Mg2+, and P nutrients in the root plasma membranes [57], resulting in essential nutrient deficiency and elevated toxic ions. However, H2S priming enhanced the mineral content of sunflower seedlings and restored the nutrient pool, which is crucial for triggering several physiological and biochemical events related to stress adaptation. Ion homeostasis restoration induced by H2S treatment under salinity stress most probably is explained by H2S-improved maintenance of the plasma membrane integrity, thus preventing the K+ efflux and maintaining Ca2+ and other beneficial nutrients [52, 53, 58]. Another possible explanation might be due to H2S-enhanced K+ and Ca2+ levels that induce the plasma membrane ATPase activity providing the chemical potential gradient required for Na+/H+ antiport work [5, 46, 50, 59], thus accelerating K+ influx and Na+ efflux (reflected in lower Na+/K+ ratio obtained in this study) on one hand. On the other hand, ATPase activity possibly hyperpolarized the plasma membrane which also promoted the entry of K+ [5, 10, 60].
As SOS pathway is reported to be involved in regulating the ion levels in the cytosol [7, 61, 62], and HaSOS2 has been shown as a candidate gene to enhance salinity tolerance and as a central regulator of SOS pathway [14, 63], HaSOS2 gene transcription in H2S primed and salinity stressed sunflower seedlings was determined. Supplementation of H2S significantly further induced upregulation of HaSOS2 transcripts in salinity-stressed sunflower seedlings, which agrees with previously published works that indicate H2S upregulation of the expression of HaSOS genes triggered SOS-mediated Na+ efflux as an important strategy for maintenance of ionic homeostasis and crop salinity tolerance [49, 64,65,66]. Therefore, H2S-enhanced HaSOS2 gene transcription might be an important strategy for the maintenance of ionic homeostasis and salinity tolerance in sunflower seedlings, and we suggest it as a biotechnological target for promoting crop tolerance to saline soil. NaCl-induced HaSOS2 gene transcription most likely was not sufficient to reduce Na+ level and contribute to ion homeostasis under NaCl stress alone.
A growing body of evidence indicates that salinity imposition is associated with oxidative stress resulting from excessive ROS production [13, 16, 58, 67,68,69,70]. Research by Jiang et al. [58] demonstrated NaCl treatment triggered the accumulation of H2O2 that was associated with significant increments in EL and MDA levels in the leaves and roots of cucumber plants. Ahanger et al. [68] also illustrated reduced membrane stability in response to salinity-mediated accumulation of H2O2 in potato seedlings. Similarly, elevated oxidative stress indicators (MDA, H2O2) are reported in maize under a saline environment [67, 70, 71]. In the present study, NaCl stress increased EL, H2O2, and MDA overproduction in both sunflower shoots and roots, which was also confirmed by the impaired ROS scavenging capacity as evidenced by the dropped level of DPPH. Seed priming with H2S, however, successfully alleviated salinity-induced oxidative damage as proved by reduced levels of H2O2, MDA, and EL along with enhanced DPPH radical scavenging activity in both sunflower shoots and roots. Consistent with our results, NaHS pretreatment has mitigated salinity stress by lowering H2O2, MDA, and EL in rice [53], cucumber [58], Malus hupehensis [66, 72], and common bean [39]. Three different mechanisms are suggested to mediate H2S amelioration of NaCl-induced oxidative damage: first, H2S can interact directly with oxidants like H2O2, superoxide radical, the hydroxyl radical, and peroxynitrite, which effectively detoxified their deleterious impact [73]; second, H2S may induce higher levels of NO that acts as a scavenger of ROS [74] as well as H2S crosstalk with NO might modulate oxidative stress by inducing the antioxidant system [42]; third, H2S priming decreased the buildup of Na+, retained K+ and Ca2+ mediating ionic and redox homeostasis, which regulates ROS production [16, 62, 75]. In agreement with our assumption, Amooaghaie and Enteshari [76] reported that H2S application directly reacts with lipid radicals to prevent lipid oxidation, and/or H2S can indirectly amplify the activation of the antioxidant system to decrease salinity-induced oxidative damage.
One crucial mechanism evolved by plants to adapt to salinity stress includes the induction of enzymatic and non-enzymatic antioxidants, which are important for eliminating ROS and maintaining cellular redox potential [14, 77]. Under abiotic-induced oxidative stress, SOD scavenges O⋅ − into O2 and H2O2 as the first stage of the defense system, and the peroxidases act on the generated H2O2 reducing it to H2O. Further, APX, MDHAR, DHAR, and GR of the ascorbate–glutathione cycle act coordinately in eliminating H2O2 and regenerating AsA and GSH, which are crucial for promoting plant tolerance to abiotic stresses [78]. Although NaCl induced the activities of SOD, CAT, POD, APX, MDHAR, DHAR, and GR enzymes, elevated level of H2O2 was still obtained in the sunflower shoots and roots, suggesting H2O2 production exceeded ROS-scavenging capacity and might be considered a harmful impact of NaCl-stressed sunflower seedlings in absence of H2S; a similar response has been previously reported in rice by Mishra et al. [79]. NaHS-pretreated stressed sunflower seedlings, on the other hand, exhibited a significant reduction in oxidative stress-induced cellular injuries through an evident declined level of H2O2 concomitant with enhanced activities of the AsA-GSH cycle enzymes and increased content of GSH and AsA, indicative of H2S function was directly associated with the activation of AsA-GSH pathway [22]. In agreement, Lai et al. [52], Mostofa et al. [53], Dawood et al. [39], and Kumari et al. [22] have reported that H2S induction of the AsA-GSH cycle contributed to maintained redox status in various crop species. Moreover, to scavenge ROS directly, GSH detoxifies endobiotic substrates with the assistance of the GST enzyme [80]. The results of this research clearly showed that H2S boosted GST activity probably by promoting its biosynthesis through upregulating the HaGST gene expression, which might also maintain the cellular integrity and GST-mediated electrophile scavenging potential under salinity stress. Our proposal is supported by the data of Mostofa et al. [53], Ding et al. [48], and Dawood et al. [39] who reported NaHS-pretreatment significantly triggered the enhancement of GSH metabolizing enzymes by regulating their gene expression and activities, which results in scavenging of the oxidative damage induced by NaCl stress.
Besides enzymatic antioxidants, plants possess antioxidant compounds such as AsA, GSH, proline, glycine betaine, phenolics, trehalose, and α-tocopherols which work coordinately to scavenge ROS efficiently under stress [13, 78, 81]. H2S priming enhanced the overproduction of α-tocopherol content in salinity-stressed sunflower shoots, which may contribute to the enhancement of the antioxidant defense system and quick elimination of ROS. In support, α-tocopherol is involved in the direct scavenging of singlet oxygen (1O2) and hydrogen peroxide (H2O2) in tomato and wheat under abiotic stresses [82, 83]. Notably, α-tocopherol was not detected in sunflower roots in response to NaCl and/or H2S treatment. Presumably, α-tocopherol is synthesized and accumulated in the shoots because one of its main roles is to preserve the redox state in the chloroplasts and maintain the thylakoid membrane structure and function during plant development and response to stresses [84], while the main form of tocopherols prevailing in roots and seeds has been indicated to be γ-tocopherol, which can functionally substitute α-tocopherol [85, 86]. As for phenolic compounds, H2S pretreatment reduced the salinity-induced accumulation of phenols particularly in the sunflower roots; most likely H2S executed its antioxidant role via reestablishing redox homeostasis mainly by increasing the levels of AsA and GSH rather than phenols.
Although different kinds of environmental stresses caused differential dynamic changes in endogenous H2S metabolism, H2S application has been shown to greatly induce endogenous H2S production [39], which was also observed in this work. NaHS treatment significantly boosts NaCl-induced endogenous H2S and was associated with enhanced total thiol contents in both shoots and roots of sunflower seedlings. The elevation in the endogenous H2S level after stress treatment is indicative of H2S ability to be an important secondary messenger of stress detection, which in turn modulates plant physiological changes and downstream gene expressions [64]. As such, other studies reported salinity induction of increments in the endogenous H2S levels in strawberry [64], alfalfa [52], and common bean [39]. Enhancing TT content along with increasing endogenous H2S obtained in this study may be attributed to H2S-induced regulation of dynamic sulfur metabolism in plants, thus promoting the production of sulfur derivatives particularly GSH, as well as sulfur-containing proteins [66]. Also, H2S-mediated NO production and the interaction between them can generate new nitrosothiols, increasing TT content [87]. In accordance, the results of Chen et al. [88] showed increased internal contents of H2S, glutathione, cysteine, and TT in response to NaHS treatment in Spinacia oleracea seedlings.
Conclusions
The present study provided a novel approach for unraveling the potential role of H2S in the alleviation of salinity-induced adversities in sunflower seedlings. H2S-mediated responses that confer salinity tolerance through two mechanisms: 1) maintained ion homeostasis by restricting the uptake of Na+, increasing K+ and Ca2+ contents as well as regulating the expression of HaSOS2 gene, and 2) reduced oxidative damage, principally by regulating the AsA-GSH cycle and scavenging of peroxides through upregulation of HaGST gene along with GST activity stimulation, leading to declining ROS accumulation and consequently their cellular injuries. Therefore, H2S acts as a regulatory molecule activating these functional processes responsible for sunflower adaptive mechanisms and could be adopted as a crucial crop management strategy to combat saline conditions in future research. However, using environmentally friendly substances for exogenous applications in natural field conditions will be of great interest to a full understanding of salinity-resistant mechanisms and improving crop yield, and may support our claims of the results observed in the controlled environments.
Materials and methods
Sunflower (Helianthus annuus L.) seeds (cv. Sakha 53) were obtained from the Agricultural Research Center, Giza, Egypt, and kept in the dark at 4 °C. Sunflower generally has been considered a sensitive plant to abiotic stresses (Tyagi et al., 2018). For the experiment, the seeds were surface sterilized by immersion in 1% (w/v) sodium hypochlorite solution for 10 min and then rinsed thoroughly with distilled water. The sterilized sunflower seeds were divided into two groups at room temperature (25 ± 2 °C): the first group was primed with distilled water (control). The second group was primed for 2 h with NaHS (Sigma, USA) as the exogenous H2S donor at a concentration of 0.5 mM. The H2S concentration was used based on a trial experiment that exhibited the best results on sunflower germination and growth. The seeds were then dried on filter paper for approximately 24 h before germination. Ten seeds were directly sown in plastic pots (diameter 15 cm, height 30 cm) containing 1.5 kg of sieved air-dried clay soil and peat moss (peat moss: clay, 1:1 v/v). The seedlings were grouped into three replications of ten seedlings per replicate. The experiment was conducted using a completely randomized design using a controlled growth chamber (model V3-DM, Vision Scientific Company, Daejeon-Si, Korea) which was maintained at 27/18 °C day/night temperatures, a 50% relative humidity, a photosynthetic photon flux density (PPFD) of 400 μmol m−2 s−1, and a 14-h photoperiod. NaHS-pretreated and non-pretreated seedlings were irrigated every other day with tape water until seedling establishment for 14 d, and then they were exposed to two levels of NaCl (0 and 150 mM) for 7 d. Therefore, the following treatments of sunflower seedlings were established: (1) control, untreated and irrigated with tape water; (2) NaCl, untreated and irrigated with 150 mM NaCl solution; (3) NaHS, NaHS-priming and irrigated with tape water; (4) NaHS + NaCl, NaHS-priming and irrigated with 150 mM NaCl solution. The seedlings were watered every other day with 75 mL of the respective treatment. At the end of the experiment, 21-day-old seedlings were harvested and growth criteria, STI, and EL were measured. The remaining seedlings were immediately frozen in liquid nitrogen and stored at —80 °C for other physiological and biochemical analyses.
Determination of growth parameters and STI
The plants were cut at the shoot base to give the shoots and roots, which were immediately weighed to determine their fresh weight (FW, g per shoot or root). The shoots and roots were then dried at 60 °C for 96 h to obtain the dry weight (DW, g per shoot or root). The STI was calculated as the total plant dry weight of salinity stress compared with the total plant dry weight of the control (0 salinity) based on the equation of Bağci et al. [89]:
Where STI = salinity tolerance index, S1 = control, Sx = salinity treatment.
Elemental analysis, Na+ uptake, and translocation
To determine the shoot and root mineral nutrient contents; Na+, K+, Ca2+, Mg2+, and P as well as the Na+/K+ ratio, dried samples (0.1 g) were ground and digested with an HNO3:HClO4 (5:1 v/v) mixture at 80°C until the yellow color disappeared. The contents of Na+, K+, Ca2+, and Mg2+ were measured using flame atomic absorption spectrophotometry (Savant AA, GBC, Australia). P was determined calorimetrically (HI 835200 Multiparameter Bench photometer, UK) according to the method of Jackson [90]. The results were expressed as µg of the metal per g DW of the sample. Sodium uptake at the sunflower root surface and ionic Na+ translocation from root to shoot were calculated according to Malik et al. [91] using the following equations:
Determination of EL and the content of MDA
As salinity imposition induces excessive ROS generation resulting in oxidative stress, membrane integrity in response to salinity and H2S priming was evaluated in the leaves and roots by measuring the EL as described by Valentovic et al. [92] using the HANNA conductivity meter (HI8733, UK). The EL was determined according to the following equation:
Where L1 refers to the electric conductivity (EC) of the outer de-ionized water where the leaves were soaked for 24 h at 25°C, and L2 is the EC of the outer de-ionized water after autoclaving at 120 °C for 20 min.
MDA was determined by the thiobarbituric acid (TBA)–based colorimetric method as described by Heath and Packer [93]. Half a gram of leaf sample was extracted in 3 mL of trichloroacetic acid (TCA) and centrifuged at 11,500 × g for 12 min. The supernatant (1 mL) was integrated with 4 mL of thiobarbituric acid (TBA) reagent (0.5% TBA + 20% TCA) in a 95 °C water bath for 30 min. After cooling, the density of the colored chromophore was observed using a colorimeter. The results were expressed as µmol MDA g−1 FW using the molar extinction coefficient of 155 mM−1 cm−1.
Determination of H2O2 content
H2O2 was determined by the methods of Velikova et al. [94]. Leaf tissues (0.5 g) were homogenized in an ice bath with 5 mL 0.1% (w/v) trichloroacetic acid. The homogenate was centrifuged at 12,000 g for 15 min and 0.5 mL of the supernatant was added to 0.5 mL of 10 mM potassium phosphate buffer (pH 7.0) and 1 mL of 1 M KI. The absorbance of the supernatant was measured at 390 nm. The content of H2O2 was determined using a standard curve and expressed as µmol g−1 FW.
DPPH radical scavenging assay
DPPH radical scavenging is a popular spectrophotometric method that has a wide application area and is used for determining the antioxidant capacity of various compounds. The activity of 2,2‐diphenyl‐1‐picrylhydrazyl (DPPH) radical scavenging was determined using the method of Hatano et al. [95]. The radical-scavenging activity was calculated as a percentage according to the equation:
Determination of antioxidant enzyme activities and nonenzymatic antioxidant contents
Fresh samples (0.5 g) were ground in liquid nitrogen and homogenized in Tris–HCl (100 mM, pH 8.0) extraction buffer containing EDTA (1 mM), DTT (5 mM), Triton X-100 (0.02%, v/v), and glycerol (10%, v/v). The resulting homogenates were centrifuged at 17,000 g for 20 min at 4 °C. The supernatants were used for the determination of enzyme activities. SOD (EC 1.15.1.1) activity was measured according to Marklund and Marklund [96]. CAT (EC 1.11.1.6) activity was assayed following the method of Aebi [97]. POD (EC 1.11.1.7) activity was determined based on the method of Shannon et al. [98]. DHAR (EC 1.8.5.1) activity was measured using the method of De Tullio et al. [99]. MDHAR (EC 1.6.5.4) activity was assayed according to the method of Hossain and Asada [100]. GR (EC 1.6.4.2) was assayed by the method of Goldberg and Spooner [101] using kits (Biodiagnostics, Egypt). GST (EC 2.5.1.18) was carried out according to Habig and Jakoby [102]. APX (EC 1.11.1.11) activity was measured by the method of Chen and Asada [103].
Non-enzymatic antioxidants including TPC and AsA were determined according to Makkar et al. [104] and Mukherjee and Choudhuri [105], respectively. GSH was determined according to the modified method of Vlachaki and Meyn [106]. The content of α-tocopherol was quantified by using high-performance liquid chromatography (HPLC; instrument E-Chrom Tech, LC 1620, USA) with electrochemical detection according to Desai [107]. Extraction from the samples was performed with 1 mL of methanol and 4 mL of hexane. After the samples were centrifuged at 1500 g for 10 min, the hexane phase was removed and evaporated to dryness under N2. Samples were dissolved in methanol: ethanol (1:1, v/v) and injected for HPLC analysis. d,l-α T from synthetic phytol (Sigma, USA) was used as standard.
Determination of endogenous H2S and TT contents
The endogenous H2S content of sunflower leaves and roots was determined using 2 g FW each, which was extracted and homogenized in 20 mM Tris–HCl buffer (pH 8.0) containing 10 mM EDTA and 20 mM zinc acetate to stabilize H2S. After centrifugation at 15,000 g for 15 min at 4 °C, 1 mL of 30 mM FeCl3 dissolved in 1.2 M HCl and 1 mL of 20 mM DMPD dissolved in 7.2 M HCl mix were added. The test mixture was incubated at room temperature for 15 min, and the absorbance was read at 670 nm. The content of H2S was determined and expressed as μg g−1 FW from a standard curve of an appropriate donor of NaHS based on the method of Li et al. [108].
Total thiols were estimated as described by Nagalakshmi and Prasad [109]. Fresh shoots (0.5 g) were homogenized in 20 mM ascorbate buffer containing 20 mM EDTA, and the homogenates were centrifuged at 12,000 g for 20 min at 4°C. Aliquots (0.5 mL) of the supernatants were mixed with 2.4 mL of 200 mM Tris–HCl buffer (pH 8.2) and 0.1 mL of 10 mM DTNB. The color was allowed to develop for 15 min, and then the absorbance was measured at 412 nm. The TT content was calculated and expressed as μg g−1 FW via a standard curve prepared with known concentrations of GSH.
RNA extraction and Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from 30 mg of leaf FW for each treatment using a Gene JET™ RNA purification Kit (Thermo Fisher Scientific, MA, USA). One μg of total RNA was reverse transcribed into cDNA using the Revert Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, MA, USA). Primer sequences for the HaSOS2 gene reactions were 5′-AATTTGGATGATATTCGTGCAGTTTTTG-3′ and 5′-TTAACATTTAAATGGAATTGACC-3′/ synthesized from Gene link, USA, as described by Halfter et al. [110].
The primer pairs used for the HaGST gene were 5′-TTGTGGAGAGGATCAGAGG-3′ and 5′-TTTAGCCGAAAAGGGTATT-3′ as described by Ma et al. [111]. Actin sequence from sunflower 5′-AGGGCGGTCTTTCCAAGTAT -3′ and 5′-ACATACATGGCGGGAACATT -3′ was used as a reference gene to normalize the relative transcription and to minimize different copy numbers of cDNA templates. PCR amplification specificity was verified using melting curve analysis and data were analyzed using the 2−ΔΔCt method according to Livak and Schmittgen [112] after normalizing to the expression of each actin gene.
Statistical analysis
The results were subjected to a one-way analysis of variance (ANOVA) using the software package SPSS v20.0 (SPSS Inc., Chicago, USA). The means of different treatments were compared using Duncan’s multiple range test at a significance level of 5% (P ≤ 0.05).
Availability of data and materials
The datasets analyzed during the study can be obtained from the corresponding author on request.
Abbreviations
- APX:
-
Ascorbate peroxidase
- AsA-GSH cycle:
-
Ascorbate-glutathione cycle
- CAT:
-
Catalase
- DHAR:
-
Dehydroascorbate reductase
- DPPH:
-
2,2‐Diphenyl‐1‐picrylhydrazyl
- EC:
-
Electric conductivity
- EL:
-
Electrolyte leakage
- GR:
-
Glutathione reductase
- GST:
-
Glutathione S-transferase
- HPLC:
-
High-performance liquid chromatography
- MDA:
-
Malondialdehyde
- MDHAR:
-
Monodehydroascorbate reductase
- POD:
-
Peroxidase
- qRT-PCR:
-
Quantitative Real-Time PCR
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- SOS:
-
Salt Overly Sensitive
- STI:
-
Salt tolerance index
- TPC:
-
Total phenolic content
- TT:
-
Total thiol
References
Tarolli P, Luo J, Straffelini E, Liou Y-A, Nguyen K-A, Laurenti R, et al. Saltwater intrusion and climate change impact on coastal agriculture. PLOS Water. 2023;2:e0000121.
Ashraf M, Munns R. Evolution of approaches to increase the salt tolerance of crops. CRC Crit Rev Plant Sci. 2022;41:128–60.
Sah SK, Reddy KR, Li J. Abscisic acid and abiotic stress tolerance in crop plants. Front Plant Sci. 2016;7:571.
Mansour MMM, Salama KHA. Cellular mechanisms of plant salt tolerance. In: Giri B, Varma A, editors. Microorganisms in Saline Environment: Strategies and Functions. Switzerland: Springer; 2019. p. 169–210.
Mansour MMF. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against NaCl stress. Plant Physiol Biochem. 1998;36:767–72.
Ketehouli T, Idrice Carther KF, Noman M, Wang F-W, Li X-W, Li H-Y. Adaptation of plants to salt stress: characterization of Na+ and K+ transporters and role of CBL gene family in regulating salt stress response. Agronomy. 2019;9:687.
Mansour MMF. Role of vacuolar membrane transport systems in plant salinity tolerance. J Plant Growth Regul. 2023;42:1364–401.
Brindha C, Vasantha S, Raja AK, Tayade AS. Characterization of the salt overly sensitive pathway genes in sugarcane under salinity stress. Physiol Plant. 2021;171:677–87.
Chen C, He G, Li J, Perez-Hormaeche J, Becker T, Luo M, et al. A salt stress-activated GSO1-SOS2-SOS1 module protects the Arabidopsis root stem cell niche by enhancing sodium ion extrusion. EMBO J. 2023;42:e113004.
Mansour MMF, Hassan FAS. How salt stress-responsive proteins regulate plant adaptation to saline conditions? Plant Mol Biol. 2022;108:175–224.
Verslues PE, Batelli G, Grillo S, Agius F, Kim Y-S, Zhu J, et al. Interaction of SOS2 with nucleoside diphosphate kinase 2 and catalases reveals a point of connection between salt stress and H2O2 signaling in Arabidopsis thaliana. Mol Cell Biol. 2007;27:7771–80.
Mansour MMF. The plasma membrane transport systems and adaptation to salinity. J Plant Physiol. 2014;171:1787–800.
Mansour MMF, Salama KHA. Proline and abiotic stresses: responses and adaptation. In: Hasanuzzaman M, editor. Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II, Mechanisms of Adaptation and Stress Amelioration. Singapore: Springer; 2020. p. 357–97.
Mansour MMF, Emam MM, Salama KHA, Morsy AA. Sorghum under saline conditions: Responses, tolerance mechanisms, and management strategies. Planta. 2021;254:1–38.
Surówka E, Hura T. Osmoprotectants and nonenzymatic antioxidants in halophytes. Handb Halophytes From Mol to Ecosyst Towar Biosaline Agric. 2020;5:1670553.
Mansour MMF, Ali EF, Salama KHA. Does seed priming play a role in regulating reactive oxygen species under saline conditions? In: Hasanuzzaman M, Fotopoulos V, Nahar K, Fujita M, editors. Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms. New Jersey: Wiley; 2019. p. 437–88.
Hasanuzzaman M, Bhuyan MHMB, Anee TI, Parvin K, Nahar K, Mahmud J Al, et al. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants. 2019;8:384.
Farooq M, Rehman A, Al-Alawi AKM, Al-Busaidi WM, Lee D-J. Integrated use of seed priming and biochar improves salt tolerance in cowpea. Sci Hortic. 2020;272:109507.
Huang P, He L, Abbas A, Hussain S, Hussain S, Du D, et al. Seed priming with sorghum water extract improves the performance of camelina (Camelina sativa (L.) crantz.) under salt stress. Plants. 2021;10:749.
Biswas S, Seal P, Majumder B, Biswas AK. Efficacy of seed priming strategies for enhancing salinity tolerance in plants: an overview of the progress and achievements. Plant Stress. 2023;9:100186.
Hussain M, Farooq M, Lee D. Evaluating the role of seed priming in improving drought tolerance of pigmented and non-pigmented rice. J Agron Crop Sci. 2017;203:269–76.
Kumari S, Kaur H, Jain A, Hussain SJ, Siddiqui MH, Khan MIR. Hydrogen sulfide modulates ascorbate-glutathione system, osmolytes production, nutrient content and yield responses under salt stress in wheat. South Afri J Bot. 2023;160:295–308.
Zhang H, Zhang X, Gao G, Ali I, Wu X, Tang M, et al. Effects of various seed priming on morphological, physiological, and biochemical traits of rice under chilling stress. Front Plant Sci. 2023;14:1146285.
Corpas FJ, Palma JM. H2S signaling in plants and applications in agriculture. J Adv Res. 2020;24:131–7.
Zhang J, Zhou M, Zhou H, Zhao D, Gotor C, Romero LC, et al. Hydrogen sulfide, a signaling molecule in plant stress responses. J Integr Plant Biol. 2021;63:146–60.
Ocvirk D, Špoljarević M, Kristić M, Hancock JT, Teklić T, Lisjak M. The effects of seed priming with sodium hydrosulphide on drought tolerance of sunflower (Helianthus annuus L.) in germination and early growth. Ann Appl Bio. 2021;178:400–13.
Luo S, Calderón-Urrea A, Yu J, Liao W, Xie J, Lv J, et al. The role of hydrogen sulfide in plant alleviates heavy metal stress. Plant Soil. 2020;449:1–10.
Chen T, Tian M, Han Y. Hydrogen sulfide: A multi-tasking signal molecule in the regulation of oxidative stress responses. J Exp Bot. 2020;71:2862–9.
Goyal V, Jhanghel D, Mehrotra S. Emerging warriors against salinity in plants: Nitric oxide and hydrogen sulphide. Physiol Plant. 2021;171:896–908.
Khan MN, Siddiqui MH, Mukherjee S, Alamri S, Al-Amri AA, Alsubaie QD, et al. Calcium-hydrogen sulfide crosstalk during K+-deficient NaCl stress operates through regulation of Na+/H+ antiport and antioxidative defense system in mung bean roots. Plant Physiol Biochem. 2021;159:211–25.
Srivastava V, Chowdhary AA, Verma PK, Mehrotra S, Mishra S. Hydrogen sulfide-mediated mitigation and its integrated signaling crosstalk during salinity stress. Physiol Plant. 2022;174:e13633.
Corpas FJ, Palma JM. Functions of NO and H2S Signal Molecules Against Plant Abiotic Stress. In: Plant Abiotic Stress Signaling. Springer; 2023. 97–109.
Adeleke BS, Babalola OO. Oilseed crop sunflower (Helianthus annuus) as a source of food: Nutritional and health benefits. Food Sci Nutr. 2020;8:4666–84.
El-Hamidi M, Zaher FA. Production of vegetable oils in the world and in Egypt: an overview. Bullet Nat Res Cent. 2018;42:19.
Bonciu E, Pandia O, Olaru AL, Saracin I, Rosculete E. Some aspects regarding the genetic and biotechnological progress of the Helianthus annuus L. Manag Econ Eng Agric Rural Dev. 2020;20:105–10.
Naeem M, Basit A, Ahmad I, Mohamed HI, Wasila H. Effect of salicylic acid and salinity stress on the performance of tomato plants. Gesunde Pflanz. 2020;72:393–402.
Mohamed NA, Saad MF, Shukry M, El-Keredy AMS, Nasif O, Van Doan H, et al. Physiological and ion changes of Nile tilapia (Oreochromis niloticus) under the effect of salinity stress. Aquac Reports. 2021;19:100567.
Calzada K, Calero Hurtado A, Olivera Viciedo D, Habermann E, de Mello PR, de Oliveira R, et al. Regulatory role of silicon on growth, potassium uptake, ionic homeostasis, proline accumulation, and antioxidant capacity of soybean plants under salt stress. J Plant Growth Regul. 2023;42:4528–40.
Dawood MFA, Sofy MR, Mohamed HI, Sofy AR, Abdel-kader HAA. Hydrogen Sulfide Modulates Salinity Stress in Common Bean Plants by Maintaining Osmolytes and Regulating Nitric Oxide Levels and Antioxidant Enzyme Expression. J Soil Sci Plant Nutr. 2022;22:3708–26.
Shalaby OAE-S, Farag R, Ibrahim MFM. Effect of hydrogen sulfide and hydrogen peroxide on growth, yield and nutrient content of broccoli plants grown under saline conditions. Sci Hortic. 2023;316:112035.
Arif Y, Singh P, Siddiqui H, Bajguz A, Hayat S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol Biochem. 2020;156:64–77.
Chen T, Shabala S, Niu Y, Chen ZH, Shabala L, Meinke H, et al. Molecular mechanisms of salinity tolerance in rice. Crop J. 2021;9:506–20.
Joshi S, Nath J, Singh AK, Pareek A, Joshi R. Ion transporters and their regulatory signal transduction mechanisms for salinity tolerance in plants. Physiol Plant. 2022;174:e13702.
Parihar P, Singh S, Singh R, Singh VP, Prasad SM. Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res. 2015;22:4056–75.
Sun YD, Luo WR. Effects of exogenous hydrogen sulphide on seed germination and seedling growth of cucumber (Cucumis sativus) under sodium bicarbonate stress. Seed Sci Technol. 2014;42:126–31.
Chen J, Wang WH, Wu FH, He EM, Liu X, Shangguan ZP, et al. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci Rep. 2015;5:12516.
Chen P, Yang W, Jin S, Liu Y. Hydrogen sulfide alleviates salinity stress in Cyclocarya paliurus by maintaining chlorophyll fluorescence and regulating nitric oxide level and antioxidant capacity. Plant Physiol Biochem. 2021;167:738–47.
Ding H, Ma D, Huang X, Hou J, Wang C, Xie Y, et al. Exogenous hydrogen sulfide alleviates salt stress by improving antioxidant defenses and the salt overly sensitive pathway in wheat seedlings. Acta Physiol Plant. 2019;41:123.
Deng Y-Q, Bao J, Yuan F, Liang X, Feng Z-T, Wang B-S. Exogenous hydrogen sulfide alleviates salt stress in wheat seedlings by decreasing Na+ content. Plant Growth Regul. 2016;79:391–9.
Zhao N, Zhu H, Zhang H, Sun J, Zhou J, Deng C, et al. Hydrogen sulfide mediates K+ and Na+ homeostasis in the roots of salt-resistant and salt-sensitive poplar species subjected to NaCl stress. Front Plant Sci. 2018;9:1366.
Ali M, Afzal S, Parveen A, Kamran M, Javed MR, Abbasi GH, et al. Silicon mediated improvement in the growth and ion homeostasis by decreasing Na+ uptake in maize (Zea mays L.) cultivars exposed to salinity stress. Plant Physiol Biochem. 2021;158:208–18.
Lai D, Mao Y, Zhou H, Li F, Wu M, Zhang J, et al. Endogenous hydrogen sulfide enhances salt tolerance by coupling the reestablishment of redox homeostasis and preventing salt-induced K+ loss in seedlings of Medicago sativa. Plant Sci. 2014;225:117–29.
Mostofa MG, Saegusa D, Fujita M, Phan Tran LS. Hydrogen sulfide regulates salt tolerance in rice by maintaining Na+/K+ balance, mineral homeostasis and oxidative metabolism under excessive salt stress. Front Plant Sci. 2015;6:1055.
Chen Z, Zhou M, Newman IA, Mendham NJ, Zhang G, Shabala S. Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance. Funct Plant Biol. 2007;34:150–62.
Li Z, Zhang Z, Feng Z, Chen J, Zhao L, Gao Y, et al. Energy transfer analysis of the SH626 sheet rotary dryer on the production system perspective. Energy Rep. 2022;8:13–20.
Shabala S, Demidchik V, Shabala L, Cuin TA, Smith SJ, Miller AJ, et al. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 2006;141:1653–65.
Zeng Y, Li Q, Wang H, Zhang J, Du J, Feng H, et al. Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol J. 2018;16:310–21.
Jiang JL, Tian Y, Li L, Yu M, Hou RP, Ren XM. H2S alleviates salinity stress in cucumber by maintaining the Na+/K+ balance and regulating H2S metabolism and oxidative stress response. Front Plant Sci. 2019;10:678.
Li J, Jia H, Wang J, Cao Q, Wen Z. Hydrogen sulfide is involved in maintaining ion homeostasis via regulating plasma membrane Na+/H+ antiporter system in the hydrogen peroxide-dependent manner in salt-stress Arabidopsis thaliana root. Protoplasma. 2014;251:899–912.
Sun J, Dai S, Wang R, Chen S, Li N, Zhou X, et al. Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol. 2009;29:1175–86.
Abid MA, Liang C, Malik W, Meng Z, Tao Z, Meng Z, et al. Cascades of ionic and molecular networks involved in expression of genes underpin salinity tolerance in cotton. J Plant Growth Regul. 2018;37:668–79.
Manishankar P, Wang N, Köster P, Alatar AA, Kudla J. Calcium signaling during salt stress and in the regulation of ion homeostasis. J Exp Bot. 2018;69:4215–26.
Yang Y, Tang R, Jiang C, Li B, Kang T, Liu H, et al. Overexpression of the PtSOS2 gene improves tolerance to salt stress in transgenic poplar plants. Plant Biotechnol J. 2015;13:962–73.
Christou A, Manganaris GA, Papadopoulos I, Fotopoulos V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways. J Exp Bot. 2013;64:1953–66.
Yang Y, Guo Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018;217:523–39.
Li H, Shi J, Wang Z, Zhang W, Yang H. H2S pretreatment mitigates the alkaline salt stress on Malus hupehensis roots by regulating Na+/K+ homeostasis and oxidative stress. Plant Physiol Biochem. 2020;156:233–41.
Sacała E. The influence of increasing doses of silicon on maize seedlings grown under salt stress. J Plant Nutr. 2017;40:819–27.
Ahanger MA, Alyemeni MN, Wijaya L, Alamri SA, Alam P, Ashraf M, et al. Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS One. 2018;13:e0202175.
Abdelrhim AS, Mazrou YSA, Nehela Y, Atallah OO, El-Ashmony RM, Dawood MFA. Silicon dioxide nanoparticles induce innate immune responses and activate antioxidant machinery in wheat against Rhizoctonia solani. Plants. 2021;10:2758.
Kamran M, Parveen A, Ahmar S, Malik Z, Hussain S, Chattha MS, et al. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int J Mol Sci. 2019;21:148.
AbdElgawad H, Zinta G, Hegab MM, Pandey R, Asard H, Abuelsoud W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front Plant Sci. 2016;7:276.
Wei GQ, Zhang WW, Cao H, Yue SS, Li P, Yang HQ. Effects hydrogen sulfide on the antioxidant system and membrane stability in mitochondria of malus hupehensis under NaCl stress. Biol Plant. 2019;63:228–36.
Filipovic MR, Zivanovic J, Alvarez B, Banerjee R. Chemical biology of H2S signaling through persulfidation. Chem Rev. 2018;118:1253–337.
Del Castello F, Nejamkin A, Cassia R, Correa-Aragunde N, Fernández B, Foresi N, et al. The era of nitric oxide in plant biology: Twenty years tying up loose ends. Nitric Oxide. 2019;85:17–27.
Xu D, Wang W, Gao T, Fang X, Gao X, Li J, et al. Calcium alleviates decreases in photosynthesis under salt stress by enhancing antioxidant metabolism and adjusting solute accumulation in Calligonum mongolicum. Conserv Physiol. 2017;5:cox060.
Amooaghaie R, Enteshari S. Role of two-sided crosstalk between NO and H2S on improvement of mineral homeostasis and antioxidative defense in Sesamum indicum under lead stress. Ecotoxicol Environ Saf. 2017;139:210–8.
Younes NA, Rahman MM, Wardany AA, Dawood MFA, Mostofa MG, Keya SS, et al. Antioxidants and bioactive compounds in licorice root extract potentially contribute to improving growth, bulb quality and yield of onion (Allium cepa). Molecules. 2021;26:2633.
Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010;48:909–30.
Mishra P, Bhoomika K, Dubey RS. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings. Protoplasma. 2013;250:3–19.
Ahmad P, Tripathi DK, Deshmukh R, Pratap Singh V, Corpas FJ. Revisiting the role of ROS and RNS in plants under changing environment. Environ Exp Bot. 2019;161:1–3.
Khan AA, Lu LX, Yao F-J, Fang M, Wang P, Zhang YM, Meng JJ, Ma XX, He Q, Shao KS, Wei YH, Xu B. Characterization, antioxidant activity, and mineral profiling of Auricularia cornea mushroom strains. Front Nutr. 2023;10:1167805.
Spicher L, Glauser G, Kessler F. Lipid antioxidant and galactolipid remodeling under temperature stress in tomato plants. Front Plant Sci. 2016;7:167.
Saeed S, Ullah A, Ullah S, Elshikh MS, Noor J, Eldin SM, et al. Salicylic acid and α-tocopherol ameliorate salinity impact on wheat. ACS Omega. 2023;29:26122–35.
Mesa T, Munné-Bosch S. α-Tocopherol in chloroplasts: nothing more than an antioxidant? Curr Opin Plant Biol. 2023;74:102400.
Munné-Bosch S, Alegre L. The function of tocopherols and tocotrienols in plants. CRC Crit Rev Plant Sci. 2002;21:31–57.
Munoz P, Munne-Bosch S. Vitamin E in plants: biosynthesis, transport, and function. Trends Plant Sci. 2019;24:1040–51.
Kharbech O, Ben Massoud M, Sakouhi L, Djebali W, Jose Mur LA, Chaoui A. Exogenous application of hydrogen sulfide reduces chromium toxicity in maize seedlings by suppressing NADPH oxidase activities and methylglyoxal accumulation. Plant Physiol Biochem. 2020;154:646–56.
Chen J, Wu F-H, Wang W-H, Zheng C-J, Lin G-H, Dong X-J, et al. Hydrogen sulphide enhances photosynthesis through promoting chloroplast biogenesis, photosynthetic enzyme expression, and thiol redox modification in Spinacia oleracea seedlings. J Exp Bot. 2011;62:4481–93.
Baǧci SA, Ekiz H, Yilmaz A. Determination of the salt tolerance of some barley genotypes and the characteristics affecting tolerance. Turkish J Agric For. 2003;27:253–60.
Jackson KW, West TS, Balchin L. Determination of arsenic (III) at the parts-per-billion level by differential pulse polarography. Anal Chem. 1973;45:267–71.
Malik AA, Li WG, Lou LN, Weng JH, Chen JF. Biochemical/physiological characterization and evaluation of in vitro salt tolerance in cucumber. African J Biotechnol. 2010;9:3284–92.
Valentovič P, Luxová M, Kolarovič L, Gašparíková O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant, Soil Environ. 2006;52:186–91.
Heath RL, Packer L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys. 1968;125:189–98.
Velikova V, Yordanov I, Edreva A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci. 2000;151:59–66.
Hatano T, Kagawa H, Yasuhara T, Okuda T. Two new flavonoids and other constituents in licorice root. Their relative astringency and radical scavenging effects. Chem Pharm Bull. 1988;36:2090–7.
Marklund S, Marklund G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem. 1974;47:469–74.
Aebi H. Catalase in vitro. Methods Enzymol. 1984;105:121–6.
Shannon LM, Kay E, Lew JY. Peroxidase isozymes from horseradish roots: I. Isolation and physical properties. J Biol Chem. 1966;241:2166–72.
De Tullio MC, De Gara L, Paciolla C, Arrigoni O. Dehydroascorbate-reducing proteins in maize are induced by the ascorbate biosynthesis inhibitor lycorine. Plant Physiol Biochem. 1998;36:433–40.
Hossain MA, Nakano Y, Asada K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984;25:385–95.
Goldberg DM, Spooner RJ. UV method for determination of glutathione reductase activity. Methods Enzym Anal. 1983;3:258–65.
Habig WH, Jakoby WB. Glutathione S-transferases (rat and human). Methods Enzymol. 1981;77:218–31.
Chen G-X, Asada K. Ascorbate peroxidase in tea leaves: occurrence of two isozymes and the differences in their enzymatic and molecular properties. Plant Cell Physiol. 1989;30:987–98.
Makkar HPS, Singh B, Vats SK, Sood RP. Total phenols, tannins and condensed tannins in different parts of Rumex hastatus. Bioresour Technol. 1993;45:69–71.
Mukherjee SP, Choudhuri MA. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol Plant. 1983;58:166–70.
Vlachaki MT, Meyn RE. Astro research fellowship: the role of bcl-2 and glutathione in an antioxidant pathway to prevent radiation-induced apoptosis. Int J Radiat Oncol Biol Phys. 1998;42:185–90.
Desai ID. Vitamin E analysis methods for animal tissues. Methods Enzymol. 1984;105:138–47.
Li Z-G. Quantification of hydrogen sulfide concentration using methylene blue and 5, 5′-dithiobis (2-nitrobenzoic acid) methods in plants. Methods Enzymol. 2015;554:101–10.
Nagalakshmi N, Prasad MNV. Responses of glutathione cycle enzymes and glutathione metabolism to copper stress in Scenedesmus bijugatus. Plant Sci. 2001;160:291–9.
Halfter U, Ishitani M, Zhu J-K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc Natl Acad Sci. 2000;97:3735–40.
Ma L, Zhang Y, Meng Q, Shi F, Liu J, Li Y. Molecular cloning, identification of GSTs family in sunflower and their regulatory roles in biotic and abiotic stress. World J Microbiol Biotechnol. 2018;34:1–12.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods. 2001;25:402–8.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research did not receive any specific grant from funding agencies in the public, commercial, or nonprofit sectors.
Author information
Authors and Affiliations
Contributions
AAY, MMFM: Conceptualization, design of the experiment, and validation. AAY: Methodology, formal analyses, and wrote the original draft. MMFM: Data curation, review, and editing of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Younis, A.A., Mansour, M.M.F. Hydrogen sulfide priming enhanced salinity tolerance in sunflower by modulating ion hemostasis, cellular redox balance, and gene expression. BMC Plant Biol 23, 525 (2023). https://doi.org/10.1186/s12870-023-04552-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04552-w