
Abstract
The Ets1 proto-oncoprotein is a member of the Ets family of transcription factors that share a unique DNA binding domain, the Ets domain. The DNA binding activity of Ets1 is controlled by kinases and transcription factors. Some transcription factors, such as AML-1, regulate Ets1 by targeting its autoinhibitory module. Others, such as Pax-5, alter Ets1 DNA binding properties. Ets1 harbors two phosphorylation sites, threonine-38 and an array of serines within the exon VII domain. Phosphorylation of threonine-38 by ERK1/2 activates Ets1, whereas phosphorylation of the exon VII domain by CaMKII or MLCK inhibits Ets1 DNA binding activity. Ets1 is expressed by numerous cell types. In haemotopoietic cells, it contributes to the regulation of cellular differentiation. In a variety of other cells, including endothelial cells, vascular smooth muscle cells and epithelial cancer cells, Ets1 promotes invasive behavior. Regulation of MMP1, MMP3, MMP9 and uPA as well as of VEGF and VEGF receptor gene expression has been ascribed to Ets1. In tumors, Ets1 expression is indicative of poorer prognosis.
Similar content being viewed by others


Introduction
Ets proteins comprise a family of transcription factors that share a unique DNA binding domain, the Ets domain [1–4]. The name "Ets" stems from a sequence that was detected in an avian erythroblastosis virus, E26, where it formed a transforming gene together with Δgag and c-myb [5, 6]. The newly discovered sequence was called E 26 t ransformation specific s equence or Ets. Later, a cellular homologue to the viral ets (v-ets) gene, c-ets1, was found suggesting that v-ets derived from c-ets1 [7, 8].
Structure of c-Ets1
The c-Ets1 protein is closely related to c-Ets2. It is believed that these two proteins derived from the same ancestor gene by duplication [9, 10]. The genomes of Drosophila and sea urchin contain only one Ets1-related gene, D-ets2 (Pointed) and SU-ets2, respectively, whereas birds, reptiles, amphibians and mammals harbor both the ets1 and ets2 genes [11, 12]. In humans, the ets1 and ets2 genes are located on two distinct chromosomes, ets1 on chromosome #11, ets2 on chromosome #21 [7]. No ets1-related gene has been found in the genome of the Caenorhabditis elegans, although this nematode worm does express a variety of other Ets proteins [13].
In birds and reptiles, the c-ets1 locus contains two different start sites leading to the expression of p68c-ets1 and p54c-ets1. These two proteins differ in their N-terminal sequence [14]. Exons α (I) and β (II) code for the N-terminus of p68c-ets1, whereas exon I54 (A) encodes the N-terminus of p54c-ets1 (Fig. 1). The exon β domain of p68c-ets1 shows a high degree of homology to the N-terminus of c-Ets2 and was thus called the Ets1-beta/Ets2-conserved sequence (BEC). In mammals, only exon I54 is present, therefore, p68c-ets1 is not expressed.

The domains of the Ets1 protein. BEC = Ets1-beta/Ets2-conserved sequence, TAD = transactivation domain, NLS = nuclear localization sequence, HI-1/2 = inhibitory α-helices 1/2, H4 = α-helix 4, S = serines, T = threonine-38.
The human TATA-less ets1 gene contains eight exons (A, III-IX) [15]. Transcripts either harbor all exons or lack exon IV- or exon VII- or exon IV/VII-specific sequence [16]. Only two proteins are generated from these RNAs, p54c-ets1 (full length Ets1) and p42c-ets1 (ΔVII-Ets1) (Fig. 1). The Ets1 protein sequence is highly conserved among species. E.g., the DNA and protein sequence of chicken Ets1 is 85% or 95%, respectively, identical to the corresponding sequence of human Ets1 [17]. The Ets1 protein can be divided into six domains, A-F (Fig. 1). The E-domain is the DNA binding Ets-domain. The adjoining D- (or exon VII-) domain and the F-domain are regulatory domains that control the activity of the E-domain. The A- und B- domains also function as regulatory units, whereas the C-domain is the activation domain of p54c-ets1 and p42c-ets1.
The Ets domain
Ets domain proteins belong to the superfamily of winged helix-turn-helix (wHTH) DNA-binding proteins which includes also hepatocyte nuclear factor HFN-3γ, heat shock factor HSF and catabolite activator protein CAP [18]. The Ets domain, composed of 85 amino acids, comprises three α-helices and four β-strands that are arranged in the order H1-S1-S2-H2-H3-S3-S4 (Fig. 2). The Ets domain specifically recognizes DNA sequences that contain a GGAA/T core element [19]. In the Ets1 protein, the Ets domain stretches from residue 331 to residue 415. As revealed by NMR spectroscopy, the helix H3 of Ets1 interacts with the GGAA motif in the major groove, whereas the "wing", formed by the loop between strands 3 and 4 of the β-sheet, contacts the 5' minor groove [20]. Helices 1–3, the "turn" between helix H2 and helix H3 contribute to the interaction of the Ets1 domain with the 3' minor groove. Strikingly, the distantly related Ets factor PU.1 binds in a similar way to DNA [21] demonstrating that the Ets domain/DNA interaction is highly preserved. However, Ets proteins differ significantly in their preference for the sequence flanking the GGAA/T core motif. The consensus sequence for Ets1 is PuCC/a-GGAA/T-GCPy as determined by several rounds of selection and amplification of Ets1-binding sites [19]. Yet, many natural Ets1-responsive GGAA/T elements differ from this consensus sequence [22]. It is possible that other transcription factors may facilitate Ets1 binding to unfavorable DNA sequences (see below).

The inhibitory module of Ets1. Details are described in the text. For simplicity, only the major contact of the H3-helix with DNA is shown. ID = inhibitory domain, H = α-helix, S = β-sheet, HI= inhibitory α-helix, I-1 = HI-1 after having unfolded to a random coil, AML1 = acute-myeloid-leukemia-1
Ets1 is predominantly found in the nucleus, though its presence in the cytoplasm of quiescent endothelial cells as well as of endometrial and ovarian cancer cells has also been demonstrated [23–25]. Deletion of the C-terminal part of the Ets domain excludes Ets1 from the nucleus [26] showing that the nuclear localization sequence of Ets1 resides on its Ets domain (Fig. 1).
The Pointed domain and the Ras-responsive phosphorylation site
The Pointed (PNT) domain, named after the Ets1-related protein Pointed-P2 in Drosophila, is shared by many Ets proteins. The Ets1 PNT domain, located between amino acid 54 and 135, consists of five α-helices. Though originally thought to adopt a helix-loop-helix (HLH) conformation, NMR analysis revealed that the PNT domain forms a globular structure that does not resemble any other known protein- or DNA-binding interaction interface [27]. The N-terminal sequence of Ets1 contains also a Ras-responsive phosphorylation site at threonine-38 [28, 29] (Fig. 1). Phosphorylation of this residue strongly increases the transcriptional activity of Ets1.
The exon VII domain, the autoinhibitory module and calcium-dependent phosphorylation
The D- or exon VII domain resides in the Ets1 protein between amino acids 243 and 331 (Fig. 1). It comprises two regulatory units. The C-terminal unit is part of an autoinhibitory module [30], the N-terminal sequence contains a calcium-responsive phosphorylation site [31]. The two units are functionally connected as phosphorylation within the N-terminal sequence increases the inhibitory effect of the C-terminal unit [32]. An inhibitory sequence within the exon VII domain was first proposed when studies showed that deletion of the exon VII domain results in an enhanced DNA binding activity [33]. Since a similar increase in DNA binding was also observed when the C-terminus (F-domain) was deleted or mutated [34–36] an interplay between the exon VII domain and the C-terminus of the Ets1 protein was suggested. Later, it became clear that the exon VII-specific sequences between 301 and 331 and the C-terminal sequence between 415 and 440 form a metastable autoinhibitory module [30]. The key structural unit was found to be formed by three inhibitory helices (HI-1 and HI-2 within exon VII domain and H4 at the C-terminus). These helices cooperatively block Ets DNA binding activity by interacting with the H1-helix of the Ets domain and, thereby, freeze the Ets domain in a closed confirmation [37, 38]. The blockage is transient and can be relieved when the HI-1 helix spontaneously unfolds to form a random coil (Fig. 2). This structural switch allows other proteins, such as the transcription factor AML-1 (acute myeloid leukemia-1), to modulate Ets1 DNA binding activity [37].
Calcium-dependent phosphorylation of the exon VII domain also interferes with the activity of the Ets1 autoinhibitory module and decreases the stability of the Ets1 protein in T-lymphocytes [39]. It involves phosphorylation of a cluster of serines, including Ser251, Ser257, Ser282 and Ser285, within the N-terminal part of exon VII [31] (Fig. 1). It results in stabilization of the inhibitory structure and in a strongly reduced DNA binding activity [32]. The natural splicing variant ΔVII-Ets1 lacks an essential part of the autoinhibitory module and the calcium-sensitive serines. This confers the ability to ΔVII-Ets1 to form stable complexes with DNA [40] and renders this protein resistant to the negative action of calcium. The viral v-Ets1 protein contains a modified C-terminus which inactivates the autoinhibitory module as well and allows v-Ets1 to bind stably to DNA [36].
The activation domains
The activation domain or C-domain is located between amino acids 130–242 and contains a high content of acidic residues. It is essential for p54c-ets1 and p42c-ets1 to activate transcription [41]. Chicken p68c-ets1 harbors a second N-terminal transcriptional domain, BEC (as mentioned above), that it shares with Ets2 [10]. The C-domain is also necessary for the interaction of Ets1 with the CREB binding factor (CBP)/p300 [42]. CBP and p300 have dual functions as histone acetyl transferases and co-activators that facilitate cooperativity between transcription factors [43, 44].
Interaction with other proteins
Ets1 functionally and physically interacts with a variety of transcription factors and other proteins (Table 1). Most proteins that directly contact Ets1 bind to the Ets domain (Fig. 3). Others associate with the exon VII domain, the N-terminal part of Ets1 or the activation domain. Some proteins, such as Sp100 or TFE3, interact with several domains of the Ets1 protein [45, 46].

Proteins that physically interact with Ets1. Proteins that cooperate with Ets1 to activate genes are shown in light yellow, proteins that either repress Ets1 activity or cooperate with Ets1 to repress gene activity are shown in pink. Proteins whose specific sites of interaction on the Ets1 protein are not yet defined (Sp1, HTLV-I Tax, mut p53) are indicated below. AML1 = acute-myeloid-leukemia-1, AP-1 = activator protein 1, ATF2 = activating transcription factor 2, CBP = CREB binding protein, ERK1/2 = extracellular-signal-regulated kinase 1/2, HIF-2α = hypoxia-inducible factor 2α, HTLV-I Tax = human T-cell lymphotropic virus-I transactivating protein of region X, MafB = musculoaponeurotic fibrosarcoma B, NFAT = nuclear factor of activated T-cells, Pax5 = paired box containing gene 5, Pit-1 = pituitary-specific trans-acting factor 1, Sp100 = speckled, 100 kD, Stat5 = signal transducer and activators of transcription 5, EAP = Ets1-associated protein, UBC9 = ubiquitin-conjugating enzyme 9, USF-1 = upstream stimulatory factor 1, VDR = vitamin D receptor
Transcription factors
A number of transcription factors have been shown to regulate the transcriptional activity of Ets1 by modulating Ets1 DNA binding affinity (Fig. 4). AML-1, Pit-1 (pituitary-specific transacting factor 1) and HIF-2α (hypoxia-inducible factor 2α) bind to the exon VII domain and block the inhibitory module of Ets1 [49, 50, 64, 73]. In turn, Ets1 may also increase the DNA binding activity of its partner. E.g., Ets1 stimulates DNA binding activity of AML-1 by associating with its NRDB domain (negative regulatory domain for DNA binding) [50, 89] (Fig. 4). In the presence of a palindromic Ets binding site, the exon VII domain also mediates homodimerization of Ets1 proteins [61]. The dimerization blocks the autoinhibitory mechanism allowing these proteins to mutually increase their DNA binding activities and to bind cooperatively to DNA. Pax5 (paired box containing gene 5), a transcription factor that contacts the Ets domain of Ets1, uses a different way to increase Ets1 affinity to DNA. It reorientates Tyr-395 of the DNA-contacting helix 3 such that Ets1 can even bind to an unfavorable GGAG containing element [70]. Often, synergism between Ets1 and another transcription factor require certain stimuli to induce activation and/or recruitment of the Ets1 interaction partner. Both phorbol ester and ionomycin are needed for the cooperative effect of Ets1 with GATA3 on the human interleukin-5 (IL-5) promoter, for the synergistic action of Ets1 with AP1 (activator protein 1)and NFκB (nuclear factor of κB) on the granulocyte-macrophage colony stimulating factor (GM-CSF) promoter and for the ability of Ets1 and Stat5 to cooperatively activate promoters through a GAS (interferon γ-activated sequence)/Ets combinatorial element [56, 63, 84]. Whereas, TGFβ (transforming growth factor β) is essential for the synergistic action of Ets1 and Smad3 (Sma/Mother against Decapentaplegic) on the PTHrP (parathyroid-hormone-related protein) P3 promoter [77]. However, the importance of vitamin D for the functional interaction of Ets1 with the vitamin D receptor (VDR) seems to depend on the target gene. While vitamin D was required for the Ets1/VDR synergistic effect on the rat cytochrome P450C24 gene promoter [90], it was dispensable for the Ets1/VDR combined effect on rat prolactin promoter [62]. The cooperative action of Ets1 with a transcription factor is sometimes mediated by a co-factor. This is the case for the Ets1/wildtype p53 synergism, where CBP is required for the interaction between these two proteins [76]. The combinatorial effect of Ets1 with another transcription factor can also lead to gene repression. This has been shown for the interaction of Ets1 with Daxx/EAPI (Ets1 associated protein I) which induces the suppression of the bcl2 and mmp1 genes [87].

Ternary complexes of Ets1 with AML1, Pit-1 or Pax5 and DNA. TAD = transactivation domain, NRDB = negative regulatory domain for DNA binding (autoinhibitory domain of AML1), runt = DNA binding domain of AML1, POU-HD = POU homeo domain (C-terminal DNA binding domain of Pit-1), POU-S = POU-specific (N-terminal DNA binding domain of Pit-1), CTD = C-terminal DNA binding domain of Pax5, NTD = N-terminal DNA binding domain.
Some nuclear proteins, EAPI/II, MafB and ZEB, are able to inhibit the transcriptional activity of Ets1 [55, 67, 88]. ZEB-induced Ets1 repression can be relieved by c-Myb, a protein that can synergize with Ets1 [67].
Kinases
Several kinases have been demonstrated to phosphorylate Ets1 and modulate its activity. Among them is calmodulin-dependent kinase II (CaMKII) which can mimick calcium in its ability to induce phosphorylation of the exon VII domain [32, 47]. This blocks Ets1 DNA binding activity and, hence, inhibits Ets1 transcriptional activity. Recent data suggest that this inactivation may even convert Ets1 from an activator to a repressor protein. This has been shown for the GM-CSF (granulocyte/macrophage-colony stimulating factor) promoter, whose activity was found to be repressed when CaMKII-phosphorylated Ets1 was present [47]. It is thought that CaMKII-inactivated Ets1 may act as a dominant negative protein by interfering with the activity of related, CaMKII-resistant Ets proteins, such as ΔVII-Ets1. Besides CaMKII, also myosin light-chain kinase (MLCK) and a constitutively active form of protein kinase Cα (PKCα) are able to phosphorylate the exon VII domain [74, 91]. Like CaMKII, MLCK was shown to inactivate Ets1 [91], whereas PKCα increased the transcriptional activity of Ets1 [74]. Whether exon VII phosphorylation and Ets1 activation by PKCα are independent events or whether the PKCα-mediated exon VII phosphorylation is different to that induced by CaMKII and MLCK remains to be seen. In support of the latter, PKCα-dependent exon VII phosphorylation was found to be independent of calcium [74], whereas, like CaMKII, MLCK mediates calcium-dependent exon VII phosphorylation [91]. Phosphorylation of the exon VII domain was also observed, when leukaemic cells go into mitosis [92].
The other known phosphorylation site on the Ets1 protein is threonine-38. Activation of the GTPase Ras leads to phosphorylation of threonine-38 and increased Ets1 activity [28, 29]. The Ras effect is mediated by the mitogen activated kinases ERK1 and ERK2, the two effector kinases of the Ras/Raf/MEK(MAPK/ERK kinase 1)/ERK 1/2 (extracellular-signal-regulated kinase) pathway [28, 29, 59, 93]. Phosphorylation of threonine-38 requires docking of ERK onto the PNT domain [60]. Typical Ras responsive genes harbor combinatorial Ets1/AP1 recognition motives through which Ets1 and AP1 synergistically activate transcription when stimulated by Ras [94]. Cooperative effects with Ras have also been reported for other Ets proteins, namely Ets2 and Drosophila Pointed. In both cases, a sequence similar to (114)LXLXXXF(120), which is crucial for ERK binding, is present within the PNT domain. Also a kinase-sensitive threonine is located exactly 16 amino acids upstream from the PNT domain. This suggests that the mechanism by which Ras induces phosphorylation of Ets1-related proteins is conserved.
Acetyl transferases
Ets1 interacts with the acetyl transferases CBP/p300 and ATF-2 (activating transcription factor 2) [42, 48, 58]. ATF-2, originally defined as a CRE (cAMP responsive element)-binding transcription factor, has recently also found to display acetyl transferase activity [95]. Interestingly, CBP and ATF-2 are the only Ets1-interacting proteins that so far have been demonstrated to associate with the transactivation domain of the Ets1 protein [42, 48]. This may suggest that they serve a common function, such as acetylating histones in response to Ets1 recruitment to an Ets1-regulated promoter. ATF-2 has been demonstrated to support Ets1-dependent activation of the T-cell receptor α (TCRα) gene in cooperation with the transcription factors AML-1 and LEF-1 (lymphocyte enhancer-binding factor 1)/TCF (T-cell factor) [48]. CPB and p300 are required for the synergistic action of Ets1 and wildtype p53 on pro-apoptotic genes in embryonal stem cells [76] and increased induction of the MMP-3 (matrix metalloprotease-3) promoter by Ets1 [58]. Recently, Ets1 has been reported to be itself a target for acetyl transferases. When treated with TGFβ, human dermal fibroblasts were found to acetylate Ets1 [96]. This was accompanied by inhibition of TGFβ-dependent gene expression. Since Ets1 can also synergize with TGFβ to activate genes [77], it is possible that acetylation triggers Ets1 to display an anti-TGFβ activity.
Ets1 expression
The Ets1 protein is produced by a variety of tissues. In most cases, its truncated form, ΔVII-Ets1, is expressed along with full-length Ets1. However, ΔVII-Ets1 is barely detectable in astrocytes, astrocytoma and invasive mammary carcinoma cells [91, 97].
The lymphoid and haematopoietic tissue
In adults, expression of Ets1 was originally thought to be restricted to lymphoid tissues [8]. Here, Ets1 is present in T- and B-cells at all stages of their development [98]. Ets1 is also expressed in natural killer cells [99]. In chicken, it seems that resting peripheral blood mononuclear cells produce less Ets1 than cells of thymus, spleen and bursa [100]. In the same species, Ets1 expression increased when T-cells were activated and correlated well with the presence of CD-28. In contrast, the Ets1 level was reported to be downregulated in human T-cells upon activation [101, 102]. Ets1 is also expressed in erythroid cells, where it is upregulated during differentiation [103].
The vascular system
Ets1 is also detected in other tissues. The two main blood vessel forming cell types, endothelial cells (EC) and vascular smooth muscle cells (VSMC), transiently produce Ets1 upon activation by angiogenic factors, such as vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF), angiotensin II, endothelin-1, tumor necrosis factor α (TNFα) or hydrogen peroxide [104–108]. Activation of ECs and VSMCs results in proliferation, migration and invasion of these cells [109]. Inhibition of Ets1 expression by trans-dominant negative Ets1 mutant proteins or by anti-sense oligonucleotides directed against Ets1 abrogated the ability of ECs to migrate, to adopt an invasive behavior or to form tubes in response to angiogenic growth factors [107, 110–112]. Conversely, constitutive Ets1 expression mimicked angiogenic stimulation and induced an invasive phenotype [113]. In addition, inhibition of Ets1 activity was shown to suppress hepatocyte growth factor (HGF) and FGF-2 induced angiogenesis in animals [114, 115]. During the menstrual cycle, EC-dependent Ets1 expression increases in the proliferative phase and drops in the secretory phase, following changes in the VEGF level [116]. During pregnancy, Ets1 is found in endothelial cells of the villous trophoblast, but not in ECs of maternal endothelial cells [117]. In the developing hypothalamo-hypophyseal system of the rat, Ets1 is detected during angiogenesis [118]. Under pathological conditions, Ets1 is produced by ECs that form new blood vessels in the synovial membrane of the joint in patients with active rheumatoid arthritis [119, 120], by ECs of the ulcerous gastric mocusa during the early phase of the healing process [121], in ECs within arteriovenous malformations [122] and, in rats, by ECs in the kidney during glomerulonephritis [123]. Tumor-induced neo-angiogenesis also involves Ets1. Ets1 expression is found in the vascular stroma of cancerous lesions [124] and often correlates with tumor microvessel density [23, 125–127]. In colon cancer, vascular Ets1 expression was associated with poor prognosis and higher incidence of lymph node metastasis [126].
Tumors
Ets1 is produced by a variety of solid tumors, including epithelial tumors, sarcomas and astrocytomas (Tab. 2). Dependent on the tumor type, Ets1 expression is either increased or exclusively found in invasive higher grade tumors. High Ets1 levels in breast, ovary and cervix carcinoma correlates with poorer prognosis [25, 127–129]. Ets1 was found to be an independent prognostic marker of breast cancer that was not linked to other tumor markers, such as nodal status, tumor size, histological grade or estrogen receptor status [128]. In lung, colorectal and squamous cell carcinoma, Ets1 expression was associated with a higher incidence of lymph node metastasis [126, 130, 131]. In endometrial and ovarian cancer, the presence of Ets1 correlated with a higher histological grade [24, 25]. In addition to advanced solid tumors, high Ets1 expression has also found in leukemic T-cells [132].
Other tissues
Ets1 is also expressed in astrocytes, in certain cells of the ovary, in hepatic stellate cells as well as in glandular epithelial cells and stromal cells of the endometrium during the menstrual cycle [91, 152–154]. When murine breast epithelial cells were stimulated to form tubules, Ets1 mRNA was detected at their growing tips [155]. Fibroblasts in tumor-associated stroma, but not in normal tissue has also been found to be Ets1 positive [124]. High non-vascular stromal Ets1 level in colon cancers was found to be associated with a higher risk for developing metastastic lesions in the lung [51]. In rats, progression of glomerulonephritis was accompanied with Ets1 expression in glomerular epithelial cells and interstitial cells [123].
Ets1 is also expressed in a variety of tissues throughout the embryonal development [156]. During the first trimester of pregnancy, Ets1 was found in extravillous trophoblastic cells invading the uterine vessels [117]. At embryonic day 15 of murine embryo development, Ets1 could be detected in all organs [157], whereas, later, Ets1 was predominantly present in lymphoid tissues, brain and other organs during branching morphogenesis. In the developing nervous system, Ets1 could be detected in the neural pituitary and adenohypophysis at early stages and in the hypothalamic magnocellular nuclei at later stages [118] as well as in the hindbrain regions, neural tube, neural crest and in the first and second branchial arches [158]. Ets1 was also found in developing vascular structures, such as heart, arteries, capillaries and meninges. In the developing bone, Ets1 is found in mesenchymal cells, whereas Ets1 is not detected in the cartilage [159].
Regulation of Ets1 expression
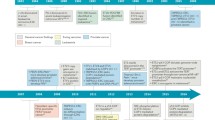
Ets1 expression can be modulated by a variety of factors (Fig. 5). The human Ets1 promoter is a TATA-less promoter containing both positive and negative regulatory elements [160]. Among the positive elements are binding sites for Ets1, AP1, AP2 and Sp1 in the proximal part of the promoter. More upstream, recognition sequence for Oct and another Ets site and the two negative regulatory elements are found. The Ets1/AP1 combinatorial element allows positive autoregulation of the Ets1 gene by a cooperative Ets1/AP1 interaction [161, 162]. Recently, recognition elements for retinoic acid receptor and for hypoxia-inducible factor HIF-1 have also been identified, through which retinoic acid or hypoxia, respectively, activates Ets1 transcription [163–165]. Ets1 synthesis is also induced by HGF (hepatocyte growth factor), PDGF (platelet-derived growth factor) or TNFα (tumor necrosis factor α) via activation of the Ras/Raf/MEK1/ERK1/2 pathway [59, 108, 166]. In breast carcinoma, melanoma and osteosarcoma cells, Ets1 expression is controlled by protein kinase C (PKC) ([77] and J. Dittmer, unpublished results). In activated endothelial cells, the MAP kinase p38 is involved in Ets1 regulation [104]. In invasive breast cancer cells, Ets1 expression can be induced by TGFβ [167]. Some factors have been shown to repress Ets1 expression. Wildtype p53, but not mutant p53, was found to repress the Ets1 promoter [15] and, in endothelial cells, retinoic acid negatively modulates Ets1 expression [168]. Different regions of the Ets1 promoter drive Ets1 expression during murine embryonal development [169]. A 2.4 kbp 5'-flanking sequence is sufficient to allow Ets1 transcription in the neural tube at gestational day 8.5, whereas also the first exon and 9 kpb of the first intron are required for Ets1 expression in the developing vessels, meninges and choriod plexus. This suggest a rather complex regulation of Ets1 gene activity.

Factors that modulate Ets1 expression. Transcription factors that directly bind to the Ets1 promoter are shown in light yellow, kinases are in orange, growth factors and other factors are in green. AP-1 = activator protein 1, ERK1/2 = extracellular-signal-regulated kinase 1/2, HGF = hepatocyte growth factor, HIF-1 = hypoxia-inducible factor 1, PDGF = platelet derived growth factor, PKC = protein kinase C, RA = retinoic acid, RAR = retinoic acid receptor, TNFα = tumor necrosis factor α, wt p53 = wildtype p53.
Ets1 function
Numerous genes have been shown to respond to the transcription factor Ets1 [22]. However, whether a particular gene is indeed regulated by Ets1 or, alternatively, by a related Ets factor can often not be answered with certainty. To address this problem, Ets1-specific anti-sense technology and trans-dominant negative protein treatments are frequently applied. However, neither method is as specific as desirable. Trans-dominant Ets1 proteins may potentially also interfere with the function of closely related Ets proteins, such as Ets2. Likewise, non-specific interactions of anti-sense oligonucleotides may also affect the expression of unrelated genes [170]. Nevertheless, based on data obtained by such approaches and by the knock-out technology in mouse a number of major functions has been ascribed to Ets1 (Fig. 6). Some of those are of more general importance, others are cell-type specific.

Main proposed functions of Ets1 in mammals. Only some of the potential Ets1 target genes are shown, ES = embryonic stem cells, EC = endothelial cells, VSMC = vascular smooth muscle cells, TCR = T-cell receptor, Ig = immunoglobulin, Tfr = transferrin receptor, VEGF = vascular endothelial growth factor, VEGFR = VEGF receptor
Haematopoietic development
Ets1 plays an important role in the development of lymphoid tissues [156]. By applying the knock-out technology in mouse, a link between the presence of Ets1 and T-lymphocyte activation was established, showing that Ets1-deficient T-cells are defective in responding to activation signals and are more prone to undergo apoptosis [171, 172]. Ets1 is also required for the development of natural killer cells [99]. In cooperation with AML-1, which is primarily expressed in T-cells [173], Ets1 activates a variety of T-cell specific genes, such as TCRα and β [48, 50]. The transcription factors LEF-1/TCF and ATF-2 support the AML-1/Ets1 cooperative effect on TCRα expression [48]. The Ets1-Stat5 synergism plays a role in the activation of genes in response to T-cell activation [84]. In the Th2 subtype of T-helper cells, Th2-specific transcription factor GATA3 is able to functionally interact with Ets1 to transactivate the interleukin-5 promoter [63]. A function of Ets1 is also proposed for B-cell development. The mb-1 (immunoglobulin alpha chain) gene which is involved in B-cell maturation has been demonstrated to be activated by a concerted action of Ets1 with the B-cell specific activator protein Pax5 [71, 72]. A role for Ets1 in regulating the immunoglobulin heavy chain gene enhancer in B-cells by cooperating with TFE3 and the distantly related Ets factor PU.1 has also been suggested [46]. In addition, it seems that Ets1 can inhibit the differentiation of B-cells to plasma cells [171, 172]. In erythroid differentiation, Ets1 expression is linked to hemoglobinization [103]. Ets1 is suggested to regulate genes that are involved in heme synthesis [174]. Accordingly, Ets1-interacting protein MafB downregulates transferrin receptor gene and porphobilinogen deaminase expression by repressing Ets1 activity [88]. The haematopoietic transcription factor c-Myb cooperates with Ets1 to transactivate the promoters of important haematopoietic cell markers, such as mim-1 and α4 integrin [67, 68]. Also janus kinase 3 (Jak3), a non-receptor tyrosine kinase that is predominantly expressed in haematopoietic cells and that is important for cytokine-mediated activation, is under the control of Ets transcription factors [175]. Chromatin immunoprecipitation assays revealed that Ets1/2 binds to the Jak3 promoter.
Invasiveness and tumor progression
The Ets1 protein has oncogenic potential. It is able to transform murine NIH3T3 cells [162] and it allows rat embryo fibroblasts to grow in serum-free medium [176]. There is also a growing body of evidence that Ets1 plays a key role in the acquisition of an invasive behavior. How could Ets1 regulate invasiveness? Among the genes that respond to Ets1 are those that code for certain proteases, such as matrix metalloproteases MMP-1, MMP-3, MMP-9, and urokinase type plasminogen activator (uPA) [22]. These proteases are known to be involved in ECM (extracellular matrix)-degradion, a key event in invasion. In angiosarcoma of the skin, Ets1 is co-expressed with MMP-1 [151]. Ovarial carcinoma cells and stromal fibroblasts in breast and ovarian cancer produce MMP-1 and MMP-9 along with Ets1 [135, 145]. In lung and brain tumors, Ets1 expression correlates with that of uPA [134, 144, 177]. Similarily, some maternal cell populations during mouse implantation and placentation were found to co-express uPA and Ets1 [178]. However, not always does Ets1 and uPA production overlap, as shown for trophoblasts that heavily synthesize uPA although they lack Ets1 [178]. By modulating Ets1 activity further evidence for a link between Ets1 and certain proteases could be accumulated. Inhibition of Ets1 expression in activated endothelial cells by anti-sense DNA reduced the synthesis of MMP-1 and uPA [110–112, 115]. Introduction of a trans-dominant negative form of Ets1 into Ets1-expressing epithelial cell lines blunted the activity of uPA [155]. In invasive MDA-MB-231 breast cancer cells, RNA interference-mediated downregulation of Ets1 down-modulated the expression of MMP-1 and MMP-9, but not that of MMP-3 and uPA (J. Dittmer, unpublished results). When overexpressed in endothelial cells or hepatoma cells, Ets1 induced the production of MMP-1, MMP-3 plus MMP-9 or MMP-1, MMP-9 plus uPA, respectively [113, 179, 180]. It seems that the cellular context dictates whether a certain protease is regulated by Ets1 or not.
The integrin αv subunit also promotes migration and invasion of cells and is involved in MMP activation [181]. In glioma cells, expression of the integrin αv gene, which contains Ets binding sites [182], was found to be reduced in the presence of a transdominant-negative form of Ets1 [183]. There is also growing evidence demonstrating that Ets1 may be involved in the regulation of c-Met, the receptor of hepatocyte growth factor/scatter factor (HGF/SF). c-Met induces migration ("scattering"), proliferation and epithelial tube formation [184]. In esophageal cancer, Met expression was found to correlate with that of Ets1 [141]. In murine liver progenitor cells and hepatoma cell lines, Ets1 was shown to induce c-Met expression and to render hepatoma cells more susceptible to HGF/SF [180, 185]. Also Epstein-Barr virus specific LMP-1 (latent membrane protein-1)-mediated induction of Ets1 expression in MDCK epithelial cells resulted in an increased production of c-Met, which could be prevented by addition of a trans-dominant negative form of Ets1 [186]. In endothelial cells, c-Met and HGF/SF expression could be reduced by anti-sense DNA directed against Ets1 [115] and high glucose treatment decreased both HGF/SF and Ets1 [187]. Conversely, c-Met also activates Ets1, as HGF/SF is able to stimulate Ets1 activity in MDCK cells through the Ras/Raf/MEK1/ERK1/2 pathway [59] and to induce Ets1 expression in endothelial cells [115]. Ets1 may even be an effector of c-Met. Not only was Ets1 found to mediate the effect of HGF/SF on MMP1 in human hepatic stellate cells [188], but it was also demonstrated to be capable of mimicking c-Met in its ability to induce "scattering" of liver progenitor cells [185].
A number of other Ets1-responsive genes are known to be involved in tumor progression. Among them is PTHrP [80, 189], expressed by numerous tumors and inducer of hypercalcaemia of malignancy [190]. It is not only a potent angiogenic factor [191], but also promotes expansion of metastatic breast cancer cells in the bone by inducing bone degradation [192]. This leads to activation of TGFβ which further stimulates PTHrP gene expression. The TGFβ effect on PTHrP involves a synergistic action between Ets1 and Smad3 and is attentuated by inhibitors that abrogate Ets1 expression [77].
Overexpression of the multidrug resistance (MDR) gene increases aggressive behavior of MCF-7 cells [193]. Mutant, but not wildtype, p53 was found to cooperate with Ets1 to increase the transcription from the MDR gene [75]. The N-acetylglucosaminyltransferase V (GlcNAc-TV) has also been reported to be a target of Ets1 [194]. Expression of GlcNAc-TV, a Golgi enzyme that catalyzes β 1,6-GlcNAc-branching of N-glycans, is associated with metastatic activities of tumor cells [195].
An additional new function of Ets1 emerged when Ets1 was found to interact with Sp100 [45], a component of PML (promyelocytic leukemia protein) oncogenic domains (PODs), nuclear bodies, which harbor also a number of other proteins, such as p53, Rb and Ets1-interacting Daxx/EAPI [196]. It could be shown that Ets1 decreased the number of PODs in HeLa cells [45]. Since PML can act as a tumor suppressor [197], Ets1 may, by interfering with PODs, negatively influence PML function and promote tumorigenesis. Several important cellular processes, such as apoptosis, cell proliferation and senescence, are ascribed to PODs [196]. Ets1 may participate in controlling some of these events. In support of this notion, Ets1 has been shown to be involved in the regulation of apoptosis (see below). Ets1 has also been found to be overexpressed in senescent, but not in young normal human fibroblasts [198]. The effect of Ets1 on PODs may involve sumoylation. By sumoylation an approximately 100 kD protein, called SUMO (small ubiquitin-like modifier), is ligated to another protein [199]. This process requires ubiquitin conjugating enzyme Ubc9 which has been shown to interact with Ets1 and to enhance its transcriptional activity [85]. Sumoylation of TEL, an Ets repressor protein, leads to the formation of TEL nuclear bodies [200]. This process requires Ubc9 to bind to the POINTED domain of TEL. Sumoylation seems to be important for packaging proteins in nuclear bodies and may effect the activity of transcription factors.
Angiogenesis
It has been well documented that Ets1 is required for endothelial cells to adopt an angiogenic, blood vessel forming phenotype [110–113, 201]. Since acquisition of invasive behavior is part of the endothelial activation program, Ets1 may be responsible for stimulating the necessary proteases [202]. However, it seems that Ets1 fulfills other functions in angiogenesis as well [203]. Anti-sense DNA directed against Ets1 downregulated VEGF expression in endothelial cells [115] suggesting that the VEGF production by endothelial cells is regulated by Ets1. Furthermore, VEGF was found to be co-expressed together with Ets1 in the vascular stroma of endometrial and colorectal carcinoma [125, 126]. Ets1 was also found to activate the promoter of the endothelial VEGF receptor 1 gene, flt-1, [204]. Accordingly, production of Ets1 and of the VEGF receptor 1 (VEGFR-1) was co-induced in endothelial cells that was co-cultured with estrogen-treated breast cancer cells [201]. Also, endothelial expression of VEGFR-1 correlated with that of Ets1 in astrocytic cancers [23]. In addition, by cooperating with the hypoxia-inducible factor HIF-2α, Ets1 is able to induce transcription from the VEGFR-2 (VEGF receptor 2) gene (flk-1) promoter [64]. However, in contrast to VEGFR-1 production, VEGFR-2 expression was not associated with Ets1 expression in astrocytic cancers [23]. The RNA level of neuropilin, an enhancer of the VEGF-A effect, was also found to be linked to Ets1 activity. It was increased in the presence of Ets1 and decreased when Ets1 activity was suppressed [205]. Thus, Ets1 seems be involved in the regulation of activities of VEGF and its receptors. As was proposed for the c-Met/HGF/SF-Ets1 cooperativity, a positive feedback loop may exist for the Ets1/VEGF interaction. By inducing Ets1 expression, VEGF may trigger its own production leading to further Ets1 synthesis, finally resulting in endothelial cell activation. However, the function of Ets1 in endothelial cells may be more complex. Overexpression of Ets1 can also lead to reduced growth activity of endothelial cells at higher cellular density [206]. This is accompanied by an increased expression of VE-cadherin, which plays an important role in vascular morphogenesis and growth control.
Apoptosis
There are conflicting data on the role of Ets1 in apoptosis. On the one hand, Ets1-deficiency increases T-cells apoptosis [171, 172] and overexpression of Ets1 protects VSMCs from undergoing apoptosis by activating p21WAF1/Cip1 [207]. On the other hand, deletion of the Ets1 gene cells render embryonic stem cells resistant to UV-induced apoptosis [76]. In this case, Ets1 acts synergistically with wildtype p53 to activate p53-responsive pro-apoptotic genes, such as bax. Similarly, Ets1 induces apoptosis in endothelial cells by stimulating the expression of pro-apoptotic genes, such as bid [202, 208]. In colon cancer cells, Ets1 cooperates with Daxx/EAPI to suppress expression of the anti-apoptotic Bcl2 [87]. This effect may be linked to the ability of Ets1 to regulate the activity of PODs, since PODs contain Daxx/EAPI (see above). Also in colon cancer cells, the natural Ets1 variant ΔVII-Ets1 was found to induce the expression of caspase 1 and to render these cells more susceptible to Fas-mediated apoptosis [209]. When overexpressed in invasive breast cancer cells, ΔVII-Ets1 reduced the survival of these cells [97]. Invasive cancer cells that depend on Ets1 may, therefore, be forced to protect themselves against the pro-apoptotic effect of Ets1, in particular of ΔVII-Ets1. One way to do this is by adopting a mechanism that selectively suppress expression of ΔVII-Ets1 [97]. T-cells in the contrary, where Ets1 acts anti-apoptotically, may tolerate high protein levels of ΔVII-Ets1.
Other functions
The presence of Ets1 in a variety of murine embryonic tissues, particular in the developing vascular system, in angiogenically active tissues and in those organs that undergo morphogenesis, suggest a fundamental role of Ets1 in embryonic development. Yet, Ets1 k.o. mice develop normally and are viable, although they show an increased perinatal mortality [171]. On the other hand, when Ets1 was downregulated together with Ets2, chicken embryos displayed defects in the epicardium, the coronary circulation and in the myocardium [210] that resembled those found in mice deficient in VCAM-1 (vascular cell adhesion molecule 1) and α4 integrin [211, 212], two Ets-responsive genes [67, 213]. Since no obvious effect on the vascular system was reported when the Ets2 gene alone was disrupted in mouse [214] it is likely that a double-knock down of Ets1 plus Ets2 is required to induce defects in the developing vascular system. Since Ets1 and Ets2 expression overlap in the developing embryo at some areas, such as in the developing heart [156], Ets2 may partially substitute for Ets1 in embryonic development. Several genes involved in embryonic development have been found to be Ets1-responsive. Among these are the gene that codes for transcription factors AP-2α (activating protein 2α) which plays a pivotal role in differentiation of the Ets1-expressing trophoblast [215].
The interaction of Ets1 with transcription factors, such as AML-1, Pit-1 or Pax5 whose expression is limited to certain tissues, predicts that Ets1 serves functions in specific tissues. As discussed above, AML-1/Ets1 and Pax-5/Ets1 cooperations may play important roles in T- and B-cell differentiation, respectively. Pit-1 is specifically produced by the anterior pituitary gland, where it is required for the expression of various genes and for the development of this gland [216, 217]. Pit-1 cooperates with pituitary Ets1 to stimulate prolactin synthesis [218, 219] suggesting that Ets1 plays also a role in pituitary hormone secretion. Interaction of Ets1 with USF (upstream stimulatory factor) [86] seems to mediate Ets1-dependent transactivation of the DOR (δ-opioid receptor) gene through an EBS (Ets binding site)/E-box combinatorial element in neuronal cells [220].
Ets1 is also involved in viral transformation. Ets1 is important for mediating transcriptional activation of specific genes by the transforming viral protein Tax, a transactivator that is encoded by the region X of the human T-cell lymphotropic virus I (HTLV-I). Being unable to bind to DNA by itself, Tax interacts with transcription factors, to get in close contact with the transcriptional machinery [221]. Tax can bind directly to Ets1 and can form a ternary complex with Ets1 and Sp1 [65]. Tax/Ets1 synergism is involved in Tax-mediated activation of the PTHrP and interleukin-5 promoter as well as of the HTLV-I LTR (long terminal repeat) [63, 189, 222]. Ets1 was also shown to be required for efficient transcription from the LTR of the human immunodeficiency virus-1 (HIV-1) [69, 86]. Accordingly, transdominant-negative Ets1 mutant protein was found to suppress HIV replication in T-cells [223].
Conclusions
The currently available data demonstrate distinct roles for Ets1 in haematopoietic cell differentiation. Ets1 seems also to participate in the regulation of invasive behavior of many normal and tumor cells alike. Adoption of invasive behavior is an important step for endothelial cells to convert to an angiogenic phenotype. Accordingly, Ets1 is abundant in regenerating adult tissues or in areas of the developing embryo that require new blood vessel to be formed. Likewise, Ets1 is overexpressed in tissues under pathological conditions that involve angiogenic activities. The link between Ets1 and invasiveness is not limited to endothelial cells, also vascular smooth muscle, epithelial and fibroblastic cells seem to need Ets1 to become invasive. For epithelial cancer, Ets1 may fulfill a dual function; it may provide the cancer cells with nutrients and oxygen by inducing tumor vascularization and it may promote tumor invasion by activating ECM-degrading proteases in the cancer and/or in stromal cells. Consequently, high levels of Ets1 in tumors often correlates with poorer prognosis. Therefore, it would be desirable to develop therapies that target the Ets1 gene. It seems that PKC is a major regulator of Ets1 expression in cancer cells. Thus, PKC inhibitors, which already entered clinical trials [224], may be a useful tool to control Ets1 activity in cancer.
As mentioned above, the current understanding of the Ets1 function is based on data partially obtained by methods which may not clearly allow distinction between the involvement of Ets1 or that of another Ets protein. In future, more reliable results may be obtained by using the recently developed method of post-transcriptional silencing by RNA interference [225]. In addition, new DNA binding assays, such as chromatin immunoprecipitation assays, may help to demonstrate direct binding of Ets1 to the gene of interest in vivo.
References
Sharrocks AD, Brown AL, Ling Y, Yates PR: The ETS-domain transcription factor family. Int J Biochem Cell Biol. 1997, 29: 1371-1387. 10.1016/S1357-2725(97)00086-1
Graves BJ, Petersen JM: Specificity within the ets family of transcription factors. Adv Cancer Res. 1998, 75: 1-55.
Ghysdael J, Boureux A: The ETS family of transcriptional regulators. In Oncogenes as transcriptional regulators. Edited by: Yaniv M, Ghysdael J. 1997, 1: 29-88. Basel: Birkhäuser
Dittmer J, Nordheim A: Ets transcription factors and human disease. Biochim Biophys Acta. 1998, 1377: F1-11. 10.1016/S0304-419X(97)00039-5
Blair DG, Athanasiou M: Ets and retroviruses - transduction and activation of members of the Ets oncogene family in viral oncogenesis. Oncogene. 2000, 19: 6472-6481. 10.1038/sj.onc.1204046
Nunn MF, Seeburg PH, Moscovici C, Duesberg PH: Tripartite structure of the avian erythroblastosis virus E26 transforming gene. Nature. 1983, 306: 391-395.
Watson DK, McWilliams-Smith MJ, Nunn MF, Duesberg PH, O'Brien SJ, Papas TS: The ets sequence from the transforming gene of avian erythroblastosis virus, E26, has unique domains on human chromosomes 11 and 21: both loci are transcriptionally active. Proc Natl Acad Sci U S A. 1985, 82: 7294-7298.
Ghysdael J, Gegonne A, Pognonec P, Dernis D, Leprince D, Stehelin D: Identification and preferential expression in thymic and bursal lymphocytes of a c-ets oncogene-encoded Mr 54, 000 cytoplasmic protein. Proc Natl Acad Sci U S A. 1986, 83: 1714-1718.
Lautenberger JA, Burdett LA, Gunnell MA, Qi S, Watson DK, O'Brien SJ, Papas TS: Genomic dispersal of the ets gene family during metazoan evolution. Oncogene. 1992, 7: 1713-1719.
Albagli O, Soudant N, Ferreira E, Dhordain P, Dewitte F, Begue A, Flourens A, Stehelin D, Leprince D: A model for gene evolution of the ets-1/ets-2 transcription factors based on structural and functional homologies. Oncogene. 1994, 9: 3259-3271.
Laudet V, Niel C, Duterque-Coquillaud M, Leprince D, Stehelin D: Evolution of the ets gene family. Biochem Biophys Res Commun. 1993, 190: 8-14. 10.1006/bbrc.1993.1002
Hsu T, Schulz RA: Sequence and functional properties of Ets genes in the model organism Drosophila. Oncogene. 2000, 19: 6409-6416. 10.1038/sj.onc.1204033
Hart AH, Reventar R, Bernstein A: Genetic analysis of ETS genes in C. elegans. Oncogene. 2000, 19: 6400-6408. 10.1038/sj.onc.1204040
Macleod K, Leprince D, Stehelin D: The ets gene family. Trends Biochem Sci. 1992, 17: 251-256. 10.1016/0968-0004(92)90404-W
Iotsova V, Crepieux P, Montpellier C, Laudet V, Stehelin D: TATA-less promoters of some Ets-family genes are efficiently repressed by wild-type p53. Oncogene. 1996, 13: 2331-2337.
Jorcyk CL, Watson DK, Mavrothalassitis GJ, Papas TS: The human ETS1 gene: genomic structure, promoter characterization and alternative splicing. Oncogene. 1991, 6: 523-532.
Watson DK, McWilliams MJ, Lapis P, Lautenberger JA, Schweinfest CW, Papas TS: Mammalian ets-1 and ets-2 genes encode highly conserved proteins. Proc Natl Acad Sci U S A. 1988, 85: 7862-7866.
Donaldson LW, Petersen JM, Graves BJ, McIntosh LP: Secondary structure of the ETS domain places murine Ets-1 in the superfamily of winged helix-turn-helix DNA-binding proteins. Biochemistry. 1994, 33: 13509-13516.
Nye JA, Petersen JM, Gunther CV, Jonsen MD, Graves BJ: Interaction of murine ets-1 with GGA-binding sites establishes the ETS domain as a new DNA-binding motif. Genes Dev. 1992, 6: 975-990.
Werner MH, Clore GM, Fisher CL, Fisher RJ, Trinh L, Shiloach J, Gronenborn AM: Correction of the NMR structure of the ETS1/DNA complex. J Biomol NMR. 1997, 10: 317-328. 10.1023/A:1018399711996
Kodandapani R, Pio F, Ni CZ, Piccialli G, Klemsz M, McKercher S, Maki RA, Ely KR: A new pattern for helix-turn-helix recognition revealed by the PU.1 ETS-domain-DNA complex. Nature. 1996, 380: 456-460. 10.1038/380456a0
Sementchenko VI, Watson DK: Ets target genes: past, present and future. Oncogene. 2000, 19: 6533-6548. 10.1038/sj.onc.1204034
Valter MM, Hugel A, Huang HJ, Cavenee WK, Wiestler OD, Pietsch T, Wernert N: Expression of the Ets-1 transcription factor in human astrocytomas is associated with Fms-like tyrosine kinase-1 (Flt-1)/vascular endothelial growth factor receptor-1 synthesis and neoangiogenesis. Cancer Res. 1999, 59: 5608-5614.
Takai N, Miyazaki T, Fujisawa K, Nasu K, Miyakawa I: Expression of c-Ets1 is associated with malignant potential in endometrial carcinoma. Cancer. 2000, 89: 2059-2067. 10.1002/1097-0142(20001115)89:10<2059::AID-CNCR5>3.0.CO;2-3
Takai N, Miyazaki T, Nishida M, Nasu K, Miyakawa I: c-Ets1 is a promising marker in epithelial ovarian cancer. Int J Mol Med. 2002, 9: 287-292.
Boulukos KE, Pognonec P, Rabault B, Begue A, Ghysdael J: Definition of an Ets1 protein domain required for nuclear localization in cells and DNA-binding activity in vitro. Mol Cell Biol. 1989, 9: 5718-5721.
Slupsky CM, Gentile LN, Donaldson LW, Mackereth CD, Seidel JJ, Graves BJ, McIntosh LP: Structure of the Ets-1 pointed domain and mitogen-activated protein kinase phosphorylation site. Proc Natl Acad Sci U S A. 1998, 95: 12129-12134. 10.1073/pnas.95.21.12129
Wasylyk C, Bradford AP, Gutierrez-Hartmann A, Wasylyk B: Conserved mechanisms of Ras regulation of evolutionary related transcription factors, Ets1 and Pointed P2. Oncogene. 1997, 14: 899-913. 10.1038/sj.onc.1200914
Yang BS, Hauser CA, Henkel G, Colman MS, Van Beveren C, Stacey KJ, Hume DA, Maki RA, Ostrowski MC: Ras-mediated phosphorylation of a conserved threonine residue enhances the transactivation activities of c-Ets1 and c-Ets2. Mol Cell Biol. 1996, 16: 538-547.
Petersen JM, Skalicky JJ, Donaldson LW, McIntosh LP, Alber T, Graves BJ: Modulation of transcription factor Ets-1 DNA binding: DNA-induced unfolding of an alpha helix. Science. 1995, 269: 1866-1869.
Rabault B, Ghysdael J: Calcium-induced phosphorylation of ETS1 inhibits its specific DNA binding activity. J Biol Chem. 1994, 269: 28143-28151.
Cowley DO, Graves BJ: Phosphorylation represses Ets-1 DNA binding by reinforcing autoinhibition. Genes Dev. 2000, 14: 366-376.
Wasylyk C, Kerckaert JP, Wasylyk B: A novel modulator domain of Ets transcription factors. Genes Dev. 1992, 6: 965-974.
Hagman J, Grosschedl R: An inhibitory carboxyl-terminal domain in Ets-1 and Ets-2 mediates differential binding of ETS family factors to promoter sequences of the mb-1 gene. Proc Natl Acad Sci U S A. 1992, 89: 8889-8893.
Hahn SL, Wasylyk B: The oncoprotein v-Ets is less selective in DNA binding than c-Ets-1 due to the C-terminal sequence change. Oncogene. 1994, 9: 2499-2512.
Lim F, Kraut N, Framptom J, Graf T: DNA binding by c-Ets-1, but not v-Ets, is repressed by an intramolecular mechanism. Embo J. 1992, 11: 643-652.
Garvie CW, Pufall MA, Graves BJ, Wolberger C: Structural analysis of the autoinhibition of Ets-1 and its role in protein partnerships. J Biol Chem. 2002, 277: 45529-45536. 10.1074/jbc.M206327200
Wang H, McIntosh LP, Graves BJ: Inhibitory module of Ets-1 allosterically regulates DNA binding through a dipole-facilitated phosphate contact. J Biol Chem. 2002, 277: 2225-2233. 10.1074/jbc.M109430200
Pognonec P, Boulukos KE, Gesquiere JC, Stehelin D, Ghysdael J: Mitogenic stimulation of thymocytes results in the calcium-dependent phosphorylation of c-ets-1 proteins. Embo J. 1988, 7: 977-983.
Fisher RJ, Fivash M, Casas-Finet J, Erickson JW, Kondoh A, Bladen SV, Fisher C, Watson DK, Papas T: Real-time DNA binding measurements of the ETS1 recombinant oncoproteins reveal significant kinetic differences between the p42 and p51 isoforms. Protein Sci. 1994, 3: 257-266.
Gegonne A, Punyammalee B, Rabault B, Bosselut R, Seneca S, Crabeel M, Ghysdael J: Analysis of the DNA binding and transcriptional activation properties of the Ets1 oncoprotein. New Biol. 1992, 4: 512-519.
Yang C, Shapiro LH, Rivera M, Kumar A, Brindle PK: A role for CREB binding protein and p300 transcriptional coactivators in Ets-1 transactivation functions. Mol Cell Biol. 1998, 18: 2218-2229.
Giles RH, Peters DJ, Breuning MH: Conjunction dysfunction: CBP/p300 in human disease. Trends Genet. 1998, 14: 178-183. 10.1016/S0168-9525(98)01438-3
Brown CE, Lechner T, Howe L, Workman JL: The many HATs of transcription coactivators. Trends Biochem Sci. 2000, 25: 15-19. 10.1016/S0968-0004(99)01516-9
Wasylyk C, Schlumberger SE, Criqui-Filipe P, Wasylyk B: Sp100 interacts with ETS-1 and stimulates its transcriptional activity. Mol Cell Biol. 2002, 22: 2687-2702. 10.1128/MCB.22.8.2687-2702.2002
Tian G, Erman B, Ishii H, Gangopadhyay SS, Sen R: Transcriptional activation by ETS and leucine zipper-containing basic helix-loop-helix proteins. Mol Cell Biol. 1999, 19: 2946-2957.
Liu H, Grundstrom T: Calcium Regulation of GM-CSF by Calmodulin-Dependent Kinase II Phosphorylation of Ets1. Mol Biol Cell. 2002, 13: 4497-4507. 10.1091/mbc.E02-03-0149
Giese K, Kingsley C, Kirshner JR, Grosschedl R: Assembly and function of a TCR alpha enhancer complex is dependent on LEF-1-induced DNA bending and multiple protein-protein interactions. Genes Dev. 1995, 9: 995-1008.
Goetz TL, Gu TL, Speck NA, Graves BJ: Auto-inhibition of Ets-1 is counteracted by DNA binding cooperativity with core-binding factor alpha2. Mol Cell Biol. 2000, 20: 81-90.
Kim WY, Sieweke M, Ogawa E, Wee HJ, Englmeier U, Graf T, Ito Y: Mutual activation of Ets-1 and AML1 DNA binding by direct interaction of their autoinhibitory domains. Embo J. 1999, 18: 1609-1620. 10.1093/emboj/18.6.1609
Sato M, Morii E, Komori T, Kawahata H, Sugimoto M, Terai K, Shimizu H, Yasui T, Ogihara H, Yasui N, Ochi T, Kitamura Y, Ito Y, Nomura S: Transcriptional regulation of osteopontin gene in vivo by PEBP2alphaA/CBFA1 and ETS1 in the skeletal tissues. Oncogene. 1998, 17: 1517-1525. 10.1038/sj.onc.1202064
Wotton D, Ghysdael J, Wang S, Speck NA, Owen MJ: Cooperative binding of Ets-1 and core binding factor to DNA. Mol Cell Biol. 1994, 14: 840-850.
Bassuk AG, Leiden JM: A direct physical association between ETS and AP-1 transcription factors in normal human T cells. Immunity. 1995, 3: 223-237.
Logan SK, Garabedian MJ, Campbell CE, Werb Z: Synergistic transcriptional activation of the tissue inhibitor of metalloproteinases-1 promoter via functional interaction of AP-1 and Ets-1 transcription factors. J Biol Chem. 1996, 271: 774-782. 10.1074/jbc.271.2.774
Pei H, Yordy JS, Leng Q, Zhao Q, Watson DK, Li R: EAPII interacts with ETS1 and modulates its transcriptional function. Oncogene. 2003, 22: 2699-2709. 10.1038/sj.onc.1206374
Thomas RS, Tymms MJ, McKinlay LH, Shannon MF, Seth A, Kola I: ETS1, NFkappaB and AP1 synergistically transactivate the human GM-CSF promoter. Oncogene. 1997, 14: 2845-2855. 10.1038/sj.onc.1201125
Wasylyk B, Wasylyk C, Flores P, Begue A, Leprince D, Stehelin D: The c-ets proto-oncogenes encode transcription factors that cooperate with c-Fos and c-Jun for transcriptional activation. Nature. 1990, 346: 191-193. 10.1038/346191a0
Jayaraman G, Srinivas R, Duggan C, Ferreira E, Swaminathan S, Somasundaram K, Williams J, Hauser C, Kurkinen M, Dhar R, Weitzman S, Buttice G, Thimmapaya B: p300/cAMP-responsive element-binding protein interactions with ets-1 and ets-2 in the transcriptional activation of the human stromelysin promoter. J Biol Chem. 1999, 274: 17342-17352. 10.1074/jbc.274.24.17342
Paumelle R, Tulasne D, Kherrouche Z, Plaza S, Leroy C, Reveneau S, Vandenbunder B, Fafeur V, Tulashe D: Hepatocyte growth factor/scatter factor activates the ETS1 transcription factor by a RAS-RAF-MEK-ERK signaling pathway. Oncogene. 2002, 21: 2309-2319. 10.1038/sj.onc.1205297
Seidel JJ, Graves BJ: An ERK2 docking site in the Pointed domain distinguishes a subset of ETS transcription factors. Genes Dev. 2002, 16: 127-137. 10.1101/gad.950902
Baillat D, Begue A, Stehelin D, Aumercier M: ETS-1 transcription factor binds cooperatively to the palindromic head to head ETS-binding sites of the stromelysin-1 promoter by counteracting autoinhibition. J Biol Chem. 2002, 277: 29386-29398. 10.1074/jbc.M200088200
Tolon RM, Castillo AI, Jimenez-Lara AM, Aranda A: Association with Ets-1 causes ligand- and AF2-independent activation of nuclear receptors. Mol Cell Biol. 2000, 20: 8793-8802. 10.1128/MCB.20.23.8793-8802.2000
Blumenthal SG, Aichele G, Wirth T, Czernilofsky AP, Nordheim A, Dittmer J: Regulation of the human interleukin-5 promoter by Ets transcription factors. Ets1 and Ets2, but not Elf-1, cooperate with GATA3 and HTLV-I Tax1. J Biol Chem. 1999, 274: 12910-12916. 10.1074/jbc.274.18.12910
Elvert G, Kappel A, Heidenreich R, Englmeier U, Lanz S, Acker T, Rauter M, Plate K, Sieweke M, Breier G, Flamme I: Cooperative interaction of hypoxia-inducible factor-2alpha (HIF-2alpha) and Ets-1 in the transcriptional activation of vascular endothelial growth factor receptor-2 (Flk-1). J Biol Chem. 2003, 278: 7520-7530. 10.1074/jbc.M211298200
Dittmer J, Pise-Masison CA, Clemens KE, Choi KS, Brady JN: Interaction of human T-cell lymphotropic virus type I Tax, Ets1, and Sp1 in transactivation of the PTHrP P2 promoter. J Biol Chem. 1997, 272: 4953-4958. 10.1074/jbc.272.8.4953
McCracken S, Leung S, Bosselut R, Ghysdael J, Miyamoto NG: Myb and Ets related transcription factors are required for activity of the human lck type I promoter. Oncogene. 1994, 9: 3609-3615.
Postigo AA, Sheppard AM, Mucenski ML, Dean DC: c-Myb and Ets proteins synergize to overcome transcriptional repression by ZEB. Embo J. 1997, 16: 3924-3934. 10.1093/emboj/16.13.3924
Dudek H, Tantravahi RV, Rao VN, Reddy ES, Reddy EP: Myb and Ets proteins cooperate in transcriptional activation of the mim-1 promoter. Proc Natl Acad Sci U S A. 1992, 89: 1291-1295.
Bassuk AG, Anandappa RT, Leiden JM: Physical interactions between Ets and NF-kappaB/NFAT proteins play an important role in their cooperative activation of the human immunodeficiency virus enhancer in T cells. J Virol. 1997, 71: 3563-3573.
Garvie CW, Hagman J, Wolberger C: Structural studies of Ets-1/Pax5 complex formation on DNA. Mol Cell. 2001, 8: 1267-1276.
Maier H, Colbert J, Fitzsimmons D, Clark DR, Hagman J: Activation of the early B-cell-specific mb-1 (Ig-alpha) gene by Pax-5 is dependent on an unmethylated Ets binding site. Mol Cell Biol. 2003, 23: 1946-1960. 10.1128/MCB.23.6.1946-1960.2003
Wheat W, Fitzsimmons D, Lennox H, Krautkramer SR, Gentile LN, McIntosh LP, Hagman J: The highly conserved beta-hairpin of the paired DNA-binding domain is required for assembly of Pax-Ets ternary complexes. Mol Cell Biol. 1999, 19: 2231-2241.
Augustijn KD, Duval DL, Wechselberger R, Kaptein R, Gutierrez-Hartmann A, van der Vliet PC: Structural characterization of the PIT-1/ETS-1 interaction: PIT-1 phosphorylation regulates PIT-1/ETS-1 binding. Proc Natl Acad Sci U S A. 2002, 99: 12657-12662. 10.1073/pnas.192693499
Lindemann RK, Braig M, Ballschmieter P, Guise TA, Nordheim A, Dittmer J: Protein kinase Calpha regulates Ets1 transcriptional activity in invasive breast cancer cells. Int J Oncol. 2003, 22: 799-805.
Sampath J, Sun D, Kidd VJ, Grenet J, Gandhi A, Shapiro LH, Wang Q, Zambetti GP, Schuetz JD: Mutant p53 cooperates with ETS and selectively up-regulates human MDR1 not MRP1. J Biol Chem. 2001, 276: 39359-39367. 10.1074/jbc.M103429200
Xu D, Wilson TJ, Chan D, De Luca E, Zhou J, Hertzog PJ, Kola I: Ets1 is required for p53 transcriptional activity in UV-induced apoptosis in embryonic stem cells. Embo J. 2002, 21: 4081-4093. 10.1093/emboj/cdf413
Lindemann RK, Ballschmieter P, Nordheim A, Dittmer J: Transforming growth factor beta regulates parathyroid hormone-related protein expression in MDA-MB-231 breast cancer cells through a novel Smad/Ets synergism. J Biol Chem. 2001, 276: 46661-46670. 10.1074/jbc.M105816200
Rekdal C, Sjottem E, Johansen T: The nuclear factor SPBP contains different functional domains and stimulates the activity of various transcriptional activators. J Biol Chem. 2000, 275: 40288-40300. 10.1074/jbc.M006978200
Block KL, Shou Y, Poncz M: An Ets/Sp1 interaction in the 5'-flanking region of the megakaryocyte-specific alpha IIb gene appears to stabilize Sp1 binding and is essential for expression of this TATA-less gene. Blood. 1996, 88: 2071-2080.
Dittmer J, Gegonne A, Gitlin SD, Ghysdael J, Brady JN: Regulation of parathyroid hormone-related protein (PTHrP) gene expression. Sp1 binds through an inverted CACCC motif and regulates promoter activity in cooperation with Ets1. J Biol Chem. 1994, 269: 21428-21434.
Gegonne A, Bosselut R, Bailly RA, Ghysdael J: Synergistic activation of the HTLV1 LTR Ets-responsive region by transcription factors Ets1 and Sp1. Embo J. 1993, 12: 1169-1178.
Han B, Liu N, Yang X, Sun HB, Yang YC: MRG1 expression in fibroblasts is regulated by Sp1/Sp3 and an Ets transcription factor. J Biol Chem. 2001, 276: 7937-7942. 10.1074/jbc.M007470200
Kavurma MM, Bobryshev Y, Khachigian LM: Ets-1 positively regulates Fas ligand transcription via cooperative interactions with Sp1. J Biol Chem. 2002, 277: 36244-36252. 10.1074/jbc.M200463200
Rameil P, Lecine P, Ghysdael J, Gouilleux F, Kahn-Perles B, Imbert J: IL-2 and long-term T cell activation induce physical and functional interaction between STAT5 and ETS transcription factors in human T cells. Oncogene. 2000, 19: 2086-2097. 10.1038/sj.onc.1203542
Hahn SL, Wasylyk B, Criqui-Filipe P, Criqui P: Modulation of ETS-1 transcriptional activity by huUBC9, a ubiquitin-conjugating enzyme. Oncogene. 1997, 15: 1489-1495. 10.1038/sj.onc.1201301
Sieweke MH, Tekotte H, Jarosch U, Graf T: Cooperative interaction of ets-1 with USF-1 required for HIV-1 enhancer activity in T cells. Embo J. 1998, 17: 1728-1739. 10.1093/emboj/17.6.1728
Li R, Pei H, Watson DK, Papas TS: EAP1/Daxx interacts with ETS1 and represses transcriptional activation of ETS1 target genes. Oncogene. 2000, 19: 745-753. 10.1038/sj.onc.1203385
Sieweke MH, Tekotte H, Frampton J, Graf T: MafB is an interaction partner and repressor of Ets-1 that inhibits erythroid differentiation. Cell. 1996, 85: 49-60.
Gu TL, Goetz TL, Graves BJ, Speck NA: Auto-inhibition and partner proteins, core-binding factor beta (CBFbeta) and Ets-1, modulate DNA binding by CBFalpha2 (AML1). Mol Cell Biol. 2000, 20: 91-103.
Dwivedi PP, Omdahl JL, Kola I, Hume DA, May BK: Regulation of rat cytochrome P450C24 (CYP24) gene expression. Evidence for functional cooperation of Ras-activated Ets transcription factors with the vitamin D receptor in 1, 25-dihydroxyvitamin D(3)-mediated induction. J Biol Chem. 2000, 275: 47-55. 10.1074/jbc.275.1.47
Fleischman LF, Holtzclaw L, Russell JT, Mavrothalassitis G, Fisher RJ: ets-1 in astrocytes: expression and transmitter-evoked phosphorylation. Mol Cell Biol. 1995, 15: 925-931.
Fleischman LF, Pilaro AM, Murakami K, Kondoh A, Fisher RJ, Papas TS: c-Ets-1 protein is hyperphosphorylated during mitosis. Oncogene. 1993, 8: 771-780.
Coffer P, de Jonge M, Mettouchi A, Binetruy B, Ghysdael J, Kruijer W: junB promoter regulation: Ras mediated transactivation by c-Ets-1 and c-Ets-2. Oncogene. 1994, 9: 911-921.
Wasylyk B, Hagman J, Gutierrez-Hartmann A: Ets transcription factors: nuclear effectors of the Ras-MAP-kinase signaling pathway. Trends Biochem Sci. 1998, 23: 213-216. 10.1016/S0968-0004(98)01211-0
Kawasaki H, Schiltz L, Chiu R, Itakura K, Taira K, Nakatani Y, Yokoyama KK: ATF-2 has intrinsic histone acetyltransferase activity which is modulated by phosphorylation. Nature. 2000, 405: 195-200. 10.1038/35012097
Czuwara-Ladykowska J, Sementchenko VI, Watson DK, Trojanowska M: Ets1 is an effector of the transforming growth factor beta (TGF-beta) signaling pathway and an antagonist of the profibrotic effects of TGF-beta. J Biol Chem. 2002, 277: 20399-20408. 10.1074/jbc.M200206200
Ballschmieter P, Braig M, Lindemann RK, Nordheim A, Dittmer J: Splicing variant DeltaVII-Ets1 is downregulated in invasive Ets1-expressing breast cancer cells. Int J Oncol. 2003, 22: 849-853.
Anderson MK, Hernandez-Hoyos G, Diamond RA, Rothenberg EV: Precise developmental regulation of Ets family transcription factors during specification and commitment to the T cell lineage. Development. 1999, 126: 3131-3148.
Barton K, Muthusamy N, Fischer C, Ting CN, Walunas TL, Lanier LL, Leiden JM: The Ets-1 transcription factor is required for the development of natural killer cells in mice. Immunity. 1998, 9: 555-563.
Koskela K, Lassila O: Single-Cell Analysis of Ets-1 Transcription Factor Expression during Lymphocyte Activation and Apoptosis. Scand J Immunol. 2003, 57: 56-61. 10.1046/j.1365-3083.2003.01192.x
Bulanova EG, Budagyan VM, Yarilin AA, Mazurenko NN: Expression of protooncogenes during lymphocyte activation by growth factors. Biochemistry (Mosc). 1997, 62: 1021-1025.
Bhat NK, Thompson CB, Lindsten T, June CH, Fujiwara S, Koizumi S, Fisher RJ, Papas TS: Reciprocal expression of human ETS1 and ETS2 genes during T-cell activation: regulatory role for the protooncogene ETS1. Proc Natl Acad Sci U S A. 1990, 87: 3723-3727.
Clausen PA, Athanasiou M, Chen Z, Dunn KJ, Zhang Q, Lautenberger JA, Mavrothalassitis G, Blair DG: ETS-1 induces increased expression of erythroid markers in the pluripotent erythroleukemic cell lines K562 and HEL. Leukemia. 1997, 11: 1224-1233. 10.1038/sj.leu.2400735
Tanaka K, Oda N, Iwasaka C, Abe M, Sato Y: Induction of Ets-1 in endothelial cells during reendothelialization after denuding injury. J Cell Physiol. 1998, 176: 235-244. 10.1002/(SICI)1097-4652(199808)176:2<235::AID-JCP2>3.0.CO;2-P
Naito S, Shimizu S, Maeda S, Wang J, Paul R, Fagin JA: Ets-1 is an early response gene activated by ET-1 and PDGF-BB in vascular smooth muscle cells. Am J Physiol. 1998, 274: C472-C480.
Hultgardh-Nilsson A, Cercek B, Wang JW, Naito S, Lovdahl C, Sharifi B, Forrester JS, Fagin JA: Regulated expression of the ets-1 transcription factor in vascular smooth muscle cells in vivo and in vitro. Circ Res. 1996, 78: 589-595.
Yasuda M, Ohzeki Y, Shimizu S, Naito S, Ohtsuru A, Yamamoto T, Kuroiwa Y: Stimulation of in vitro angiogenesis by hydrogen peroxide and the relation with ETS-1 in endothelial cells. Life Sci. 1999, 64: 249-258. 10.1016/S0024-3205(98)00560-8
Goetze S, Kintscher U, Kaneshiro K, Meehan WP, Collins A, Fleck E, Hsueh WA, Law RE: TNFalpha induces expression of transcription factors c-fos, Egr-1, and Ets-1 in vascular lesions through extracellular signal-regulated kinases 1/2. Atherosclerosis. 2001, 159: 93-101. 10.1016/S0021-9150(01)00497-X
Stenmark KR, Mecham RP: Cellular and molecular mechanisms of pulmonary vascular remodeling. Annu Rev Physiol. 1997, 59: 89-144. 10.1146/annurev.physiol.59.1.89
Naito S, Shimizu S, Matsuu M, Nakashima M, Nakayama T, Yamashita S, Sekine I: Ets-1 upregulates matrix metalloproteinase-1 expression through extracellular matrix adhesion in vascular endothelial cells. Biochem Biophys Res Commun. 2002, 291: 130-138. 10.1006/bbrc.2002.6418
Iwasaka C, Tanaka K, Abe M, Sato Y: Ets-1 regulates angiogenesis by inducing the expression of urokinase-type plasminogen activator and matrix metalloproteinase-1 and the migration of vascular endothelial cells. J Cell Physiol. 1996, 169: 522-531. 10.1002/(SICI)1097-4652(199612)169:3<522::AID-JCP12>3.3.CO;2-Q
Chen Z, Fisher RJ, Riggs CW, Rhim JS, Lautenberger JA: Inhibition of vascular endothelial growth factor-induced endothelial cell migration by ETS1 antisense oligonucleotides. Cancer Res. 1997, 57: 2013-2019.
Oda N, Abe M, Sato Y: ETS-1 converts endothelial cells to the angiogenic phenotype by inducing the expression of matrix metalloproteinases and integrin beta3. J Cell Physiol. 1999, 178: 121-132. 10.1002/(SICI)1097-4652(199902)178:2<121::AID-JCP1>3.3.CO;2-6
Pourtier-Manzanedo A, Vercamer C, Van Belle E, Mattot V, Mouquet F, Vandenbunder B: Expression of an Ets-1 dominant-negative mutant perturbs normal and tumor angiogenesis in a mouse ear model. Oncogene. 2003, 22: 1795-1806. 10.1038/sj.onc.1206215
Tomita N, Morishita R, Taniyama Y, Koike H, Aoki M, Shimizu H, Matsumoto K, Nakamura T, Kaneda Y, Ogihara T: Angiogenic property of hepatocyte growth factor is dependent on upregulation of essential transcription factor for angiogenesis, ets-1. Circulation. 2003, 107: 1411-1417. 10.1161/01.CIR.0000055331.41937.AA
Fujimoto J, Aoki I, Toyoki H, Khatun S, Sato E, Tamaya T: Expression of ETS-1 Related to Angiogenesis in Uterine Endometrium during the Menstrual Cycle. J Biomed Sci. 2003, 10: 320-327. 10.1159/000070097
Luton D, Sibony O, Oury JF, Blot P, Dieterlen-Lievre F, Pardanaud L: The c-ets1 protooncogene is expressed in human trophoblast during the first trimester of pregnancy. Early Hum Dev. 1997, 47: 147-156. 10.1016/S0378-3782(96)01775-6
Laurent-Huck FM, Egles C, Kienlen P, Stoeckel ME, Felix JM: Expression of the c-ets1 gene in the hypothalamus and pituitary during rat development. Brain Res Dev Brain Res. 1996, 97: 107-117. 10.1016/S0165-3806(96)00134-4
Redlich K, Kiener HP, Schett G, Tohidast-Akrad M, Selzer E, Radda I, Stummvoll GH, Steiner CW, Groger M, Bitzan P, Zenz P, Smolen JS, Steiner G: Overexpression of transcription factor Ets-1 in rheumatoid arthritis synovial membrane: regulation of expression and activation by interleukin-1 and tumor necrosis factor alpha. Arthritis Rheum. 2001, 44: 266-274. 10.1002/1529-0131(200102)44:2<266::AID-ANR43>3.3.CO;2-7
Wernert N, Justen HP, Rothe M, Behrens P, Dreschers S, Neuhaus T, Florin A, Sachinidis A, Vetter H, Ko Y: The Ets 1 transcription factor is upregulated during inflammatory angiogenesis in rheumatoid arthritis. J Mol Med. 2002, 80: 258-266. 10.1007/s00109-001-0316-0
Ito M, Nakayama T, Naito S, Matsuu M, Shichijo K, Sekine I: Expression of Ets-1 transcription factor in relation to angiogenesis in the healing process of gastric ulcer. Biochem Biophys Res Commun. 1998, 246: 123-127. 10.1006/bbrc.1998.8585
Wautier MP, Boval B, Chappey O, Enjolras O, Wernert N, Merland JJ, Wautier JL: Cultured endothelial cells from human arteriovenous malformations have defective growth regulation. Blood. 1999, 94: 2020-2028.
Naito T, Razzaque MS, Nazneen A, Liu D, Nihei H, Koji T, Taguchi T: Renal expression of the Ets-1 proto-oncogene during progression of rat crescentic glomerulonephritis. J Am Soc Nephrol. 2000, 11: 2243-2255.
Wernert N, Gilles F, Fafeur V, Bouali F, Raes MB, Pyke C, Dupressoir T, Seitz G, Vandenbunder B, Stehelin D: Stromal expression of c-Ets1 transcription factor correlates with tumor invasion. Cancer Res. 1994, 54: 5683-5688.
Fujimoto J, Aoki I, Toyoki H, Khatun S, Tamaya T: Clinical implications of expression of ETS-1 related to angiogenesis in uterine endometrial cancers. Ann Oncol. 2002, 13: 1605-1611. 10.1093/annonc/mdf334
Tokuhara K, Ogata Y, Nakagawa M, Shirouzu K: Ets-1 expression in vascular endothelial cells as an angiogenic and prognostic factor in colorectal carcinoma. Int Surg. 2003, 88: 25-33.
Fujimoto J, Aoki I, Toyoki H, Khatun S, Tamaya T: Clinical implications of expression of ETS-1 related to angiogenesis in uterine cervical cancers. Ann Oncol. 2002, 13: 1598-1604. 10.1093/annonc/mdf248
Span PN, Manders P, Heuvel JJ, Thomas CM, Bosch RR, Beex LV, Sweep CG: Expression of the transcription factor Ets-1 is an independent prognostic marker for relapse-free survival in breast cancer. Oncogene. 2002, 21: 8506-8509. 10.1038/sj.onc.1206040
Davidson B, Reich R, Goldberg I, Gotlieb WH, Kopolovic J, Berner A, Ben-Baruch G, Bryne M, Nesland JM: Ets-1 messenger RNA expression is a novel marker of poor survival in ovarian carcinoma. Clin Cancer Res. 2001, 7: 551-557.
Sasaki H, Yukiue H, Moiriyama S, Kobayashi Y, Nakashima Y, Kaji M, Kiriyama M, Fukai I, Yamakawa Y, Fujii Y: Clinical significance of matrix metalloproteinase-7 and Ets-1 gene expression in patients with lung cancer. J Surg Res. 2001, 101: 242-247. 10.1006/jsre.2001.6279
Pande P, Mathur M, Shukla NK, Ralhan R: Ets-1: a plausible marker of invasive potential and lymph node metastasis in human oral squamous cell carcinomas. J Pathol. 1999, 189: 40-45. 10.1002/(SICI)1096-9896(199909)189:1<40::AID-PATH405>3.0.CO;2-#
Sacchi N, de Klein A, Showalter SD, Bigi G, Papas TS: High expression of ets-1 gene in human thymocytes and immature T leukemic cells. Leukemia. 1988, 2: 12-18.
Kitange G, Shibata S, Tokunaga Y, Yagi N, Yasunaga A, Kishikawa M, Naito S: Ets-1 transcription factor-mediated urokinase-type plasminogen activator expression and invasion in glioma cells stimulated by serum and basic fibroblast growth factors. Lab Invest. 1999, 79: 407-416.
Kitange G, Tsunoda K, Anda T, Nakamura S, Yasunaga A, Naito S, Shibata S: Immunohistochemical expression of Ets-1 transcription factor and the urokinase-type plasminogen activator is correlated with the malignant and invasive potential in meningiomas. Cancer. 2000, 89: 2292-2300. 10.1002/1097-0142(20001201)89:11<2292::AID-CNCR18>3.0.CO;2-0
Behrens P, Rothe M, Wellmann A, Krischler J, Wernert N: The Ets-1 transcription factor is up-regulated together with MMP 1 and MMP 9 in the stroma of pre-invasive breast cancer. J Pathol. 2001, 194: 43-50. 10.1002/path.844
Watabe T, Yoshida K, Shindoh M, Kaya M, Fujikawa K, Sato H, Seiki M, Ishii S, Fujinaga K: The Ets-1 and Ets-2 transcription factors activate the promoters for invasion-associated urokinase and collagenase genes in response to epidermal growth factor. Int J Cancer. 1998, 77: 128-137. 10.1002/(SICI)1097-0215(19980703)77:1<128::AID-IJC20>3.0.CO;2-9
Si X, Liu Z: Expression and significance of cell cycle-related proteins Cyclin Dl, CDK4, p27, E2F-l and Ets-1 in chondrosarcoma of the jaws. Oral Oncol. 2001, 37: 431-436. 10.1016/S1368-8375(00)00091-9
Ito Y, Takeda T, Okada M, Matsuura N: Expression of ets-1 and ets-2 in colonic neoplasms. Anticancer Res. 2002, 22: 1581-1584.
Nakayama T, Ito M, Ohtsuru A, Naito S, Sekine I: Expression of the ets-1 proto-oncogene in human colorectal carcinoma. Mod Pathol. 2001, 14: 415-422.
Mukherjee T, Kumar A, Mathur M, Chattopadhyay TK, Ralhan R: Ets-1 and VEGF expression correlates with tumor angiogenesis, lymph node metastasis, and patient survival in esophageal squamous cell carcinoma. J Cancer Res Clin Oncol.
Saeki H, Kuwano H, Kawaguchi H, Ohno S, Sugimachi K: Expression of ets-1 transcription factor is correlated with penetrating tumor progression in patients with squamous cell carcinoma of the esophagus. Cancer. 2000, 89: 1670-1676. 10.1002/1097-0142(20001015)89:8<1670::AID-CNCR4>3.0.CO;2-J
Kanda K, Nakayama T, Onizuka S, Tomioka T, Kanematsu T: Expression of the Ets-1 proto-oncogene is linked to cell differentiation of human hepatocellular carcinoma. Hepatogastroenterology. 2002, 49: 747-751.
Ito Y, Miyoshi E, Takeda T, Sakon M, Tsujimoto M, Yokosaki Y, Monden M, Matsuura N: ets-1 expression in extrahepatic bile duct carcinoma and cholangiocellular carcinoma. Oncology. 2000, 58: 248-252. 10.1159/000012108
Takanami I, Takeuchi K, Karuke M: Expression of ETS-1 is correlated with urokinase-type plasminogen activator and poor prognosis in pulmonary adenocarcinoma. Tumour Biol. 2001, 22: 205-210.
Behrens P, Rothe M, Florin A, Wellmann A, Wernert N: Invasive properties of serous human epithelial ovarian tumors are related to Ets-1, MMP-1 and MMP-9 expression. Int J Mol Med. 2001, 8: 149-154.
Ito T, Nakayama T, Ito M, Naito S, Kanematsu T, Sekine I: Expression of the ets-1 proto-oncogene in human pancreatic carcinoma. Mod Pathol. 1998, 11: 209-215.
Nakayama T, Ito M, Ohtsuru A, Naito S, Nakashima M, Fagin JA, Yamashita S, Sekine I: Expression of the Ets-1 proto-oncogene in human gastric carcinoma: correlation with tumor invasion. Am J Pathol. 1996, 149: 1931-1939.
Tsutsumi S, Kuwano H, Asao T, Nagashima K, Shimura T, Mochiki E: Expression of Ets-1 angiogenesis-related protein in gastric cancer. Cancer Lett. 2000, 160: 45-50. 10.1016/S0304-3835(00)00559-0
Sasaki H, Kobayashi Y, Tanahashi M, Yukiue H, Yano M, Kaji M, Kiriyama M, Fukai I, Yamakawa Y, Fujii Y: Ets-1 gene expression in patients with thymoma. Jpn J Thorac Cardiovasc Surg. 2002, 50: 503-507.
Nakayama T, Ito M, Ohtsuru A, Naito S, Nakashima M, Sekine I: Expression of the ets-1 proto-oncogene in human thyroid tumor. Mod Pathol. 1999, 12: 61-68.
Naito S, Shimizu K, Nakashima M, Nakayama T, Ito T, Ito M, Yamashita S, Sekine I: Overexpression of Ets-1 transcription factor in angiosarcoma of the skin. Pathol Res Pract. 2000, 196: 103-109.
Rowe A, Propst F: Ets-1 and Ets-2 protooncogene expression in theca cells of the adult mouse ovary. Exp Cell Res. 1992, 202: 199-202.
Kilpatrick LM, Kola I, Salamonsen LA: Transcription factors Ets1, Ets2, and Elf1 exhibit differential localization in human endometrium across the menstrual cycle and alternate isoforms in cultured endometrial cells. Biol Reprod. 1999, 61: 120-126.
Knittel T, Kobold D, Dudas J, Saile B, Ramadori G: Role of the Ets-1 transcription factor during activation of rat hepatic stellate cells in culture. Am J Pathol. 1999, 155: 1841-1848.
Delannoy-Courdent A, Mattot V, Fafeur V, Fauquette W, Pollet I, Calmels T, Vercamer C, Boilly B, Vandenbunder B, Desbiens X: The expression of an Ets1 transcription factor lacking its activation domain decreases uPA proteolytic activity and cell motility, and impairs normal tubulogenesis and cancerous scattering in mammary epithelial cells. J Cell Sci. 1998, 111 (Pt 11): 1521-1534.
Maroulakou IG, Bowe DB: Expression and function of Ets transcription factors in mammalian development: a regulatory network. Oncogene. 2000, 19: 6432-6442. 10.1038/sj.onc.1204039
Kola I, Brookes S, Green AR, Garber R, Tymms M, Papas TS, Seth A: The Ets1 transcription factor is widely expressed during murine embryo development and is associated with mesodermal cells involved in morphogenetic processes such as organ formation. Proc Natl Acad Sci U S A. 1993, 90: 7588-7592.
Maroulakou IG, Papas TS, Green JE: Differential expression of ets-1 and ets-2 proto-oncogenes during murine embryogenesis. Oncogene. 1994, 9: 1551-1565.
Raouf A, Seth A: Ets transcription factors and targets in osteogenesis. Oncogene. 2000, 19: 6455-6463. 10.1038/sj.onc.1204037
Chen JH, Jeha S, Oka T: Negative regulatory elements in the human ETS1 gene promoter. Oncogene. 1993, 8: 133-139.
Majerus MA, Bibollet-Ruche F, Telliez JB, Wasylyk B, Bailleul B: Serum, AP-1 and Ets-1 stimulate the human ets-1 promoter. Nucleic Acids Res. 1992, 20: 2699-2703.
Seth A, Papas TS: The c-ets-1 proto-oncogene has oncogenic activity and is positively autoregulated. Oncogene. 1990, 5: 1761-1767.
So EN, Crowe DL: Characterization of a retinoic acid responsive element in the human ets-1 promoter. IUBMB Life. 2000, 50: 365-370. 10.1080/152165400300089394
Raouf A, Li V, Kola I, Watson DK, Seth A: The Ets1 proto-oncogene is upregulated by retinoic acid: characterization of a functional retinoic acid response element in the Ets1 promoter. Oncogene. 2000, 19: 1969-1974. 10.1038/sj.onc.1203505
Oikawa M, Abe M, Kurosawa H, Hida W, Shirato K, Sato Y: Hypoxia induces transcription factor ETS-1 via the activity of hypoxia-inducible factor-1. Biochem Biophys Res Commun. 2001, 289: 39-43. 10.1006/bbrc.2001.5927
Goetze S, Kintscher U, Kim S, Meehan WP, Kaneshiro K, Collins AR, Fleck E, Hsueh WA, Law RE: Peroxisome proliferator-activated receptor-gamma ligands inhibit nuclear but not cytosolic extracellular signal-regulated kinase/mitogen-activated protein kinase-regulated steps in vascular smooth muscle cell migration. J Cardiovasc Pharmacol. 2001, 38: 909-921. 10.1097/00005344-200112000-00013
Lindemann RK, Nordheim A, Dittmer J: Interfering with TGFbeta-induced Smad3 nuclear accumulation differentially affects TGFbeta-dependent gene expression. Mol Cancer. 2003, 2: 20- 10.1186/1476-4598-2-20
Igarashi T, Abe M, Oikawa M, Nukiwa T, Sato Y: Retinoic acids repress the expression of ETS-1 in endothelial cells. Tohoku J Exp Med. 2001, 194: 35-43.
Jorcyk CL, Garrett LJ, Maroulakou IG, Watson DK, Green JE: Multiple regulatory regions control the expression of Ets-1 in the developing mouse: vascular expression conferred by intron I. Cell Mol Biol (Noisy-le-grand). 1997, 43: 211-225.
Fisher AA, Ye D, Sergueev DS, Fisher MH, Shaw BR, Juliano RL: Evaluating the specificity of antisense oligonucleotide conjugates. A DNA array analysis. J Biol Chem. 2002, 277: 22980-22984. 10.1074/jbc.M203347200
Bories JC, Willerford DM, Grevin D, Davidson L, Camus A, Martin P, Stehelin D, Alt FW: Increased T-cell apoptosis and terminal B-cell differentiation induced by inactivation of the Ets-1 proto-oncogene. Nature. 1995, 377: 635-638. 10.1038/377635a0
Muthusamy N, Barton K, Leiden JM: Defective activation and survival of T cells lacking the Ets-1 transcription factor. Nature. 1995, 377: 639-642. 10.1038/377639a0
Speck NA, Terryl S: A new transcription factor family associated with human leukemias. Crit Rev Eukaryot Gene Expr. 1995, 5: 337-364.
Marziali G, Perrotti E, Ilari R, Lulli V, Coccia EM, Moret R, Kuhn LC, Testa U, Battistini A: Role of Ets-1 in transcriptional regulation of transferrin receptor and erythroid differentiation. Oncogene. 2002, 21: 7933-7944. 10.1038/sj.onc.1205925
Aringer M, Hofmann SR, Frucht DM, Chen M, Centola M, Morinobu A, Visconti R, Kastner DL, Smolen JS, O'Shea JJ: Characterization and analysis of the proximal Janus Kinase 3 promoter. J Immunol. 2003, 170: 6057-6064.
Topol LZ, Tatosyan AG, Ascione R, Thompson DM, Blair DG, Kola I, Seth A: C-ets-1 protooncogene expression alters the growth properties of immortalized rat fibroblasts. Cancer Lett. 1992, 67: 71-78.
Nakada M, Yamashita J, Okada Y, Sato H: Ets-1 positively regulates expression of urokinase-type plasminogen activator (uPA) and invasiveness of astrocytic tumors. J Neuropathol Exp Neurol. 1999, 58: 329-334.
Grevin D, Chen JH, Raes MB, Stehelin D, Vandenbunder B, Desbiens X: Involvement of the proto-oncogene c-ets 1 and the urokinase plasminogen activator during mouse implantation and placentation. Int J Dev Biol. 1993, 37: 519-529.
Sato Y, Abe M, Tanaka K, Iwasaka C, Oda N, Kanno S, Oikawa M, Nakano T, Igarashi T: Signal transduction and transcriptional regulation of angiogenesis. Adv Exp Med Biol. 2000, 476: 109-115.
Jiang Y, Xu W, Lu J, He F, Yang X: Invasiveness of hepatocellular carcinoma cell lines: contribution of hepatocyte growth factor, c-met, and transcription factor Ets-1. Biochem Biophys Res Commun. 2001, 286: 1123-1130. 10.1006/bbrc.2001.5521
Brooks PC, Silletti S, von Schalscha TL, Friedlander M, Cheresh DA: Disruption of angiogenesis by PEX, a noncatalytic metalloproteinase fragment with integrin binding activity. Cell. 1998, 92: 391-400.
Donahue JP, Sugg N, Hawiger J: The integrin alpha v gene: identification and characterization of the promoter region. Biochim Biophys Acta. 1994, 1219: 228-232. 10.1016/0167-4781(94)90278-X
Kita D, Takino T, Nakada M, Takahashi T, Yamashita J, Sato H: Expression of dominant-negative form of Ets-1 suppresses fibronectin-stimulated cell adhesion and migration through down-regulation of integrin alpha5 expression in U251 glioma cell line. Cancer Res. 2001, 61: 7985-7991.
Tamagnone L, Comoglio PM: Control of invasive growth by hepatocyte growth factor (HGF) and related scatter factors. Cytokine Growth Factor Rev. 1997, 8: 129-142. 10.1016/S1359-6101(97)00007-5
Gambarotta G, Boccaccio C, Giordano S, Ando M, Stella MC, Comoglio PM: Ets up-regulates MET transcription. Oncogene. 1996, 13: 1911-1917.
Horikawa T, Sheen TS, Takeshita H, Sato H, Furukawa M, Yoshizaki T: Induction of c-Met proto-oncogene by Epstein-Barr virus latent membrane protein-1 and the correlation with cervical lymph node metastasis of nasopharyngeal carcinoma. Am J Pathol. 2001, 159: 27-33.
Taniyama Y, Morishita R, Hiraoka K, Aoki M, Nakagami H, Yamasaki K, Matsumoto K, Nakamura T, Kaneda Y, Ogihara T: Therapeutic angiogenesis induced by human hepatocyte growth factor gene in rat diabetic hind limb ischemia model: molecular mechanisms of delayed angiogenesis in diabetes. Circulation. 2001, 104: 2344-2350.
Ozaki I, Zhao G, Mizuta T, Ogawa Y, Hara T, Kajihara S, Hisatomi A, Sakai T, Yamamoto K: Hepatocyte growth factor induces collagenase (matrix metalloproteinase-1) via the transcription factor Ets-1 in human hepatic stellate cell line. J Hepatol. 2002, 36: 169-178. 10.1016/S0168-8278(01)00245-8
Dittmer J, Gitlin SD, Reid RL, Brady JN: Transactivation of the P2 promoter of parathyroid hormone-related protein by human T-cell lymphotropic virus type I Tax1: evidence for the involvement of transcription factor Ets1. J Virol. 1993, 67: 6087-6095.
Wysolmerski JJ, Broadus AE: Hypercalcemia of malignancy: the central role of parathyroid hormone-related protein. Annu Rev Med. 1994, 45: 189-200. 10.1146/annurev.med.45.1.189
Akino K, Ohtsuru A, Kanda K, Yasuda A, Yamamoto T, Akino Y, Naito S, Kurokawa M, Iwahori N, Yamashita S: Parathyroid hormone-related peptide is a potent tumor angiogenic factor. Endocrinology. 2000, 141: 4313-4316. 10.1210/en.141.11.4313
Yin JJ, Selander K, Chirgwin JM, Dallas M, Grubbs BG, Wieser R, Massague J, Mundy GR, Guise TA: TGF-beta signaling blockade inhibits PTHrP secretion by breast cancer cells and bone metastases development. J Clin Invest. 1999, 103: 197-206.
dit Faute MA, Laurent L, Ploton D, Poupon MF, Jardillier JC, Bobichon H: Distinctive alterations of invasiveness, drug resistance and cell-cell organization in 3D-cultures of MCF-7, a human breast cancer cell line, and its multidrug resistant variant. Clin Exp Metastasis. 2002, 19: 161-168. 10.1023/A:1014594825502
Ko JH, Miyoshi E, Noda K, Ekuni A, Kang R, Ikeda Y, Taniguchi N: Regulation of the GnT-V promoter by transcription factor Ets-1 in various cancer cell lines. J Biol Chem. 1999, 274: 22941-22948. 10.1074/jbc.274.33.22941
Dennis JW, Granovsky M, Warren CE: Glycoprotein glycosylation and cancer progression. Biochim Biophys Acta. 1999, 1473: 21-34. 10.1016/S0304-4165(99)00167-1
Hatta M, Fukamizu A: PODs in the nuclear spot: enigmas in the magician's pot. Sci STKE. 2001, 2001: PE1-
Salomoni P, Pandolfi PP: The role of PML in tumor suppression. Cell. 2002, 108: 165-170.
Ohtani N, Zebedee Z, Huot TJ, Stinson JA, Sugimoto M, Ohashi Y, Sharrocks AD, Peters G, Hara E: Opposing effects of Ets and Id proteins on p16INK4a expression during cellular senescence. Nature. 2001, 409: 1067-1070. 10.1038/35059131
Seeler JS, Dejean A: SUMO: of branched proteins and nuclear bodies. Oncogene. 2001, 20: 7243-7249. 10.1038/sj.onc.1204758
Chakrabarti SR, Sood R, Nandi S, Nucifora G: Posttranslational modification of TEL and TEL/AML1 by SUMO-1 and cell-cycle-dependent assembly into nuclear bodies. Proc Natl Acad Sci U S A. 2000, 97: 13281-13285. 10.1073/pnas.240315897
Lincoln DW, Phillips PG, Bove K: Estrogen-induced Ets-1 promotes capillary formation in an in vitro tumor angiogenesis model. Breast Cancer Res Treat. 2003, 78: 167-178. 10.1023/A:1022904624054
Sato Y, Teruyama K, Nakano T, Oda N, Abe M, Tanaka K, Iwasaka-Yagi C: Role of transcription factors in angiogenesis: Ets-1 promotes angiogenesis as well as endothelial apoptosis. Ann N Y Acad Sci. 2001, 947: 117-123.
Lelievre E, Lionneton F, Soncin F, Vandenbunder B: The Ets family contains transcriptional activators and repressors involved in angiogenesis. Int J Biochem Cell Biol. 2001, 33: 391-407. 10.1016/S1357-2725(01)00025-5
Wakiya K, Begue A, Stehelin D, Shibuya M: A cAMP response element and an Ets motif are involved in the transcriptional regulation of flt-1 tyrosine kinase (vascular endothelial growth factor receptor 1) gene. J Biol Chem. 1996, 271: 30823-30828. 10.1074/jbc.271.48.30823
Teruyama K, Abe M, Nakano T, Takahashi S, Yamada S, Sato Y: Neurophilin-1 is a downstream target of transcription factor Ets-1 in human umbilical vein endothelial cells. FEBS Lett. 2001, 504: 1-4. 10.1016/S0014-5793(01)02724-7
Lelievre E, Mattot V, Huber P, Vandenbunder B, Soncin F: ETS1 lowers capillary endothelial cell density at confluence and induces the expression of VE-cadherin. Oncogene. 2000, 19: 2438-2446. 10.1038/sj.onc.1203563
Zhang C, Kavurma MM, Lai A, Khachigian LM: Ets-1 protects vascular smooth muscle cells from undergoing apoptosis by activating p21WAF1/Cip1: ETS-1 regulates basal and inducible p21WAF1/Cip1 transcription via distinct CIS-acting elements in the p21WAF1/Cip1 promoter. J Biol Chem. 2003, 278: 27903-27909. 10.1074/jbc.M304328200
Teruyama K, Abe M, Nakano T, Iwasaka-Yagi C, Takahashi S, Yamada S, Sato Y: Role of transcription factor Ets-1 in the apoptosis of human vascular endothelial cells. J Cell Physiol. 2001, 188: 243-252. 10.1002/jcp.1112
Li R, Pei H, Papas T: The p42 variant of ETS1 protein rescues defective Fas-induced apoptosis in colon carcinoma cells. Proc Natl Acad Sci U S A. 1999, 96: 3876-3881. 10.1073/pnas.96.7.3876
Lie-Venema H, Gittenberger-de Groot AC, van Empel LJ, Boot MJ, Kerkdijk H, de Kant E, DeRuiter MC: Ets-1 and Ets-2 transcription factors are essential for normal coronary and myocardial development in chicken embryos. Circ Res. 2003, 92: 749-756. 10.1161/01.RES.0000066662.70010.DB
Kwee L, Baldwin HS, Shen HM, Stewart CL, Buck C, Buck CA, Labow MA: Defective development of the embryonic and extraembryonic circulatory systems in vascular cell adhesion molecule (VCAM-1) deficient mice. Development. 1995, 121: 489-503.
Yang JT, Rayburn H, Hynes RO: Cell adhesion events mediated by alpha 4 integrins are essential in placental and cardiac development. Development. 1995, 121: 549-560.
Umetani M, Nakao H, Doi T, Iwasaki A, Ohtaka M, Nagoya T, Mataki C, Hamakubo T, Kodama T: A novel cell adhesion inhibitor, K- reduces the endothelial VCAM-1 induction by inflammatory cytokines, acting through the regulation of GATA. Biochem Biophys Res Commun. 7174, 272: 370-374. 10.1006/bbrc.2000.2784.
Yamamoto H, Flannery ML, Kupriyanov S, Pearce J, McKercher SR, Henkel GW, Maki RA, Werb Z, Oshima RG: Defective trophoblast function in mice with a targeted mutation of Ets2. Genes Dev. 1998, 12: 1315-1326.
Cheng YH, Handwerger S: Identification of an enhancer of the human activating protein-2alpha gene that contains a critical ets1 binding site. J Clin Endocrinol Metab. 2003, 88: 3305-3311. 10.1210/jc.2002-021831
Cohen LE: Genetic regulation of the embryology of the pituitary gland and somatotrophs. Endocrine. 2000, 12: 99-106. 10.1385/ENDO:12:2:99
de la Hoya M, Vila V, Jimenez O, Castrillo JL: Anterior pituitary development and Pit-1/GHF-1 transcription factor. Cell Mol Life Sci. 1998, 54: 1059-1066. 10.1007/s000180050234
Schweppe RE, Gutierrez-Hartmann A: Pituitary Ets-1 and GABP bind to the growth factor regulatory sites of the rat prolactin promoter. Nucleic Acids Res. 2001, 29: 1251-1260. 10.1093/nar/29.5.1251
Bradford AP, Brodsky KS, Diamond SE, Kuhn LC, Liu Y, Gutierrez-Hartmann A: The Pit-1 homeodomain and beta-domain interact with Ets-1 and modulate synergistic activation of the rat prolactin promoter. J Biol Chem. 2000, 275: 3100-3106. 10.1074/jbc.275.5.3100
Sun P, Loh HH: Transcriptional regulation of mouse delta-opioid receptor gene: role of Ets-1 in the transcriptional activation of mouse delta-opioid receptor gene. J Biol Chem. 2001, 276: 45462-45469. 10.1074/jbc.M104793200
Franchini G: Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 1995, 86: 3619-39.
Gitlin SD, Dittmer J, Shin RC, Brady JN: Transcriptional activation of the human T-lymphotropic virus type I long terminal repeat by functional interaction of Tax1 and Ets1. J Virol. 1993, 67: 7307-16.
Posada R, Pettoello-Mantovani M, Sieweke M, Graf T, Goldstein H: Suppression of HIV type 1 replication by a dominant-negative Ets-1 mutant. AIDS Res Hum Retroviruses. 2000, 16: 1981-1989. 10.1089/088922200750054710
Goekjian PG, Jirousek MR: Protein kinase C inhibitors as novel anticancer drugs. Expert Opin Investig Drugs. 2001, 10: 2117-2140.
Plasterk RH: RNA silencing: the genome's immune system. Science. 2002, 296: 1263-1265. 10.1126/science.1072148
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Dittmer, J. The Biology of the Ets1 Proto-Oncogene. Mol Cancer 2, 29 (2003). https://doi.org/10.1186/1476-4598-2-29
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-4598-2-29