
Abstract
Background
Anthranilate is an aromatic amine used industrially as an intermediate for the synthesis of dyes, perfumes, pharmaceuticals and other classes of products. Chemical synthesis of anthranilate is an unsustainable process since it implies the use of nonrenewable benzene and the generation of toxic by-products. In Escherichia coli anthranilate is synthesized from chorismate by anthranilate synthase (TrpED) and then converted to phosphoribosyl anthranilate by anthranilate phosphoribosyl transferase to continue the tryptophan biosynthetic pathway. With the purpose of generating a microbial strain for anthranilate production from glucose, E. coli W3110 trpD9923, a mutant in the trpD gene that displays low anthranilate producing capacity, was characterized and modified using metabolic engineering strategies.
Results
Sequencing of the trpED genes from E. coli W3110 trpD9923 revealed a nonsense mutation in the trpD gene, causing the loss of anthranilate phosphoribosyl transferase activity, but maintaining anthranilate synthase activity, thus causing anthranilate accumulation. The effects of expressing genes encoding a feedback inhibition resistant version of the enzyme 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase (aroGfbr), transketolase (tktA), glucokinase (glk) and galactose permease (galP), as well as phosphoenolpyruvate:sugar phosphotransferase system (PTS) inactivation on anthranilate production capacity, were evaluated. In shake flask experiments with minimal medium, strains W3110 trpD9923 PTS- and W3110 trpD9923/pJLBaroGfbrtkt A displayed the best production parameters, accumulating 0.70–0.75 g/L of anthranilate, with glucose-yields corresponding to 28–46% of the theoretical maximum. To study the effects of extending the growth phase on anthranilate production a fed-batch fermentation process was developed using complex medium, where strain W3110 trpD9923/ pJLBaroGfbrtkt A produced 14 g/L of anthranilate in 34 hours.
Conclusion
This work constitutes the first example of a microbial system for the environmentally-compatible synthesis of anthranilate generated by metabolic engineering. The results presented here, including the characterization of mutation in the trpD gene from strain W3110 trpD9923 and the development of a fermentation strategy, establish a step forward towards the future improvement of a sustainable process for anthranilate production. In addition, the present work provides very useful data regarding the positive and negative consequences of the evaluated metabolic engineering strategies.
Similar content being viewed by others

Background
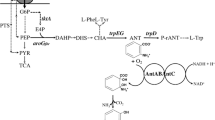
Anthranilate is an aromatic amine used as precursor for the synthesis of compounds having applications in the chemical, food and pharmaceutical industries. Current anthranilate manufacture methods are based on chemical synthesis using precursors derived from petroleum, such as benzene. Also, chemical synthesis of anthranilate is a multistep process requiring conditions of high temperature and pressure, which makes the process expensive for commercial use [1, 2]. Several microbial and plant species have the metabolic capacity to synthesize this aromatic compound, opening the possibility for generating sustainable technologies for anthranilate manufacture. This compound is a metabolic intermediate and therefore it is normally not accumulated. Anthranilate is an intermediate in the tryptophan biosynthetic pathway (Fig. 1). Carbon flow into the common aromatic pathway starts with the condensation of D-erythrose 4-phosphate (E4P) and phosphoenolpyruvate (PEP) to yield 3-deoxy-D-arabino-heptulosonate 7-phosphate (DAHP), in a reaction catalyzed by the enzyme DAHP synthase. After six more reactions, chorismate is synthesized, leading to a branch point where biosynthetic pathways for L-tryptophan (L-Trp), L-tyrosine (L-Tyr) and L-phenylalanine (L-Phe) originate. In Escherichia coli, the first two reactions in the L-Trp biosynthetic pathway are catalyzed by the enzyme complex anthranilate synthase-phosphoribosyl transferase (TrpE-TrpD). It is a multifunctional and heterotetrameric complex composed of two TrpE and two TrpD polypeptides (component I and II, respectively). Component I (TrpE) catalyses the conversion of chorismate and glutamine to anthranilate, glutamate and pyruvate. The anthranilate synthase activity is the result of aminase and amidotransferase activities that are encoded by trpE and the amino terminal region encoded by trpGD, respectively (Fig. 2a). Component II (TrpD) catalyses the transfer of the phosphoribosyl group of 5-phosphorylribose-l-pyrophosphate to anthranilate, forming N-phosphoribosylanthranilate. The carboxyl terminal region of TrpD has the anthranilate phosphoribosyl transferase activity [3, 4]. After five more metabolic steps, L-Trp is synthesized.

Metabolic network related to anthranilate biosynthesis in E. coli. Arrows with dashed lines indicate more than one enzymatic reaction. Metabolite symbols: G6P, glucose 6-phosphate; F6P, fructose 6-phosphate; F1,6DP fructose 1,6 diphosphate; G3P, glyceraldehyde 3-phosphate; Ru5P, ribulose 5-phosphate; R5P, ribose 5-phosphate; X5P, xylulose 5-phosphate; S7P, sedoheptulose 7-phosphate; PYR, pyruvate; PEP, phosphoenolpyruvate; E4P, erythrose 4-phosphate; DAHP, 3-deoxy-D-arabino-heptulosonate 7-phosphate; CHA, chorismate; PPA, prephenate; ANT, anthranilate; L-Gln, L-glutamine; L-Glu, L-glutamate; L-Phe, L-phenylalanine; L-Tyr, L-tyrosine; L-Trp, L-tryptophan. Protein and gene symbols: IICBGlc, glucose-specific integral membrane permease; TCA, tricarboxylic acid cycle; PTS, phosphotransferase transport system; tktA, transketolase; aroGfbr, feedback inhibition resistant DAHP synthase; trpED, anthranilate synthase-phosphoribosyl transferase complex.

Sequence determination of the trpEGD genes of E. coli trpD9923. (a) Organization of trpEGD genes of E. coli. (b) Comparison of the nucleotide and amino acid partial sequences of trpGD genes of E. coli MG1655 and E. coli trpD9923.
Early studies on the polarity of the L-Trp operon in E. coli enabled the identification of mutants that secreted anthranilate [5]. The characterization of one of the strains obtained by UV mutagenesis (W3110 trpD9923), revealed that the mutation was present in the trpD gene. These results suggest the feasibility of modifying E. coli to generate strains for anthranilate production. With the purpose of exploring a rational approach to improve the production capacity of strain W3110 trpD9923, in this work we characterized the mutation enabling anthranilate accumulation. In addition, we studied the effect of overexpressing genes encoding a feedback inhibition resistant DAHP synthase (aroGfbr), transketolase (tktA), glucokinase (glk) and galactose permease (galP) on anthranilate productivity and yield from glucose in strains having either active or inactive PEP:sugar phosphotransferase system (PTS).
Results
Characterization of E. coli W3110 trpD9923
The W3110 trpD9923 strain belongs to a set of E. coli mutants obtained after random mutagenesis by UV light exposure, having mutations in the first three genes of the tryptophan operon [5]. This report indicated that this strain is a tryptophan auxotroph which accumulates anthranilate. However, the specific mutation responsible for this phenotype and the anthranilate production capacity were not determined.
In order to characterize the mutation that causes anthranilate accumulation in this strain, the nucleotide sequence of the trpEGD genes was determined and compared to the corresponding sequence from E. coli MG1655 [6]. This analysis revealed a mutation at position 613, corresponding to the eighth codon of the anthranilate phosphoribosyl transferase domain of the anthranilate synthase component II (trpD), where a G to T transversion was detected, resulting in the generation of a stop codon (Fig. 2b). This mutation in trpD9923 results in the synthesis of a truncated anthranilate synthase component II protein, retaining the full glutamine amidotransferase domain and only seven of the 333 amino acid residues of the anthranilate phosphoribosyl transferase domain (Fig. 2b). This mutation in the trpD gene causes the loss of anthranilate phosphoribosyl transferase activity, but glutamine amidotransferase activity is not affected. Therefore, anthranilate can be synthesized in this strain, but it is not further metabolized to N-phosphoribosylanthranilate, thus causing anthranilate accumulation and tryptophan auxotrophy.
To determine the anthranilate production capacity of strain W3110 trpD9923, cultures were performed in shake flasks with M9 mineral medium supplemented with 20 μg/mL tryptophan and 10 g/L of glucose at 37°C. Under these conditions, this strain displayed a specific growth rate (μ) of 0.26 ± 0.04 h-1(Table 1), a maximum biomass concentration of 1.29 ± 0.03 gDCW/L in 16 h and no lag phase was observed (Fig. 3a). The specific glucose consumption rate (qGlc) was 0.34 ± 0.01 gGlc/gDCW·h. After a 12 h production phase, this strain accumulated 0.31 ± 0.01 g/L of anthranilate as the maximum concentration (Fig. 3c) with a specific anthranilate production rate (qAnt) of 0.02 ± 0.00 gAnt/gDCW·h and an anthranilate yield from glucose (YAnt/Glc) of 0.06 ± 0.01 gAnt/gGlc (Table 1).

Flask cultures of E. coli W3110 trpD9923 derivative strains for the production of anthranilate. (a) Growth curves, (b) glucose consumption, and (c) anthranilate production. (filled circle) W3110 trpD9923; (open circle) W3110 trpD9923 PTS-; (filled square) W3110 trpD9923 PTS-/pv5Glk5GalP; (X) W3110 trpD9923/pJLBaroGfbr; (open triangle) W3110 trpD9923/pJLBaroGfbrtkt A; (open square) W3110 trpD9923 PTS-/pJLBaroGfbrtkt A; (filled triangle) W3110 trpD9923 PTS-/pv5Glk5GalP/pJLBaroGfbrtkt A. Graphs show results from the mean of the triplicate experiments.
PTS inactivation in strain trpD9923
To improve the anthranilate production capacity of W3110 trpD9923, two different and complementary metabolic engineering strategies were applied: one involved increasing the availability of PEP and E4P; two metabolic precursors for anthranilate biosynthesis, and the other was based on redirecting carbon flow from central metabolism into the common aromatic pathway. Condensation of PEP and E4P is the first step in the aromatic amino acid biosynthesis pathway (Fig. 1). Several reported studies have demonstrated that PEP is a limiting precursor with regard to aromatics yield from glucose [7–9]. When E. coli is growing with glucose as the carbon source, PTS is the main activity that consumes PEP; therefore, it has been identified as a target for inactivation to increase aromatics production capacity [10–12]. In order to increase PEP biosynthetic availability in the cell, the PTS operon was inactivated in strain W3110 trpD9923 by transduction of the ΔptsHIcrr::kmR mutation, generating strain W3110 trpD9923 PTS-. Flask cultures with this PTS- strain using M9 mineral medium showed a significantly different growth profile compared to that observed with the PTS+ strain W3110 trpD9923. Cultures with strain W3110 trpD9923 PTS- showed a 10 h lag phase and the maximum biomass was 1.04 ± 0.05 gDCW/L in 44 h (Fig. 3a). This diminished growth capacity was evident by a 65% lower μ than that observed for W3110 trpD9923 (Table 1). Also, glucose consumption rate in the PTS- strain was 62% lower than the qGlc of the progenitor strain. W3110 trpD9923 PTS- displayed a 24 h anthranilate production phase, where the qAnt was 1.5-fold higher and the YAnt/Glc 2-fold higher than the corresponding values obtained in the PTS+ strain cultures (Table 1). As a result, the PTS- strain accumulated a 2.2-fold higher amount of anthranilate than W3110 trpD9923 (Fig. 3c). These results show that PTS inactivation caused a positive effect on anthranilate production capacity.
Increasing glucose transport capacity in strain W3110 trpD9923 PTS-
As expected, inactivation of PTS in W3110 trpD9923 caused a significant decrease in its qGlc, due to a reduced capacity to import this sugar [12]. As a result, its growth rate was severely affected. Thus, in order to increase the glucose transport capacity, strain W3110 trpD9923 PTS- was transformed with plasmid pv5Glk5GalP, which carries the genes glk and galP encoding glucokinase (Glk) and galactose permease (GalP), respectively. Expression of these two proteins has been shown to restore glucose import and phosphorylation activities; functions previously provided by the PTS [13]. Shake flask experiments with strain W3110 trpD9923 PTS-/pv5Glk5GalP under previously described conditions showed that expression of glk and galP in the PTS- strain caused a positive effect in glucose assimilation capacity; the observed qGlc was 1.8-fold higher than the value for the PTS- strain (Table 1). Also, the μ increased 2.2-fold with respect to that observed for the PTS- strain. Values for YAnt/Glc were similar to those in the PTS- strain and qAnt increased 1.3-fold (Table 1). However, the anthranilate production phase was reduced to 8 h due to faster glucose consumption; therefore, a lower anthranilate titer of 0.33 ± 0.01 g/L was reached at the end of the culture (Fig. 3c).
Redirection of glycolytic and pentose phosphate pathway precursors to the common aromatic amino acid biosynthetic pathway
As mentioned before, condensation of PEP and E4P generates DAHP by action of DAHP synthase (Fig. 1). However this enzyme is highly regulated by allosteric control. Thus, to increase cellular DAHP synthase activity, strain W3110 trpD9923 was transformed with plasmid pJLBaroGfbr [14], which harbors the aroGfbr gene encoding a feedback inhibition resistant mutant of DAHP synthase. Increased dosage of aroGfbr caused an increase in the qAnt and YAnt/Glc (3.5 and 2.7-fold, respectively), and 1.4-fold higher anthranilate accumulation in comparison with the parental strain. Increasing DAHP synthase activity causes a higher demand for E4P; therefore, to avoid a limitation for this intermediate, it is necessary to increase the activity of the enzyme that synthesizes it. A way to achieve this is through the high level expression of the enzyme transketolase, responsible for E4P production. Therefore, to evaluate the effect of the co-expression of tktA on anthranilate production, this gene was cloned downstream of the aroGfbr gene, generating the plasmid pJLBaroGfbrtktA. Co-expression of aroGfbr and tktA in strain W3110 trpD9923 did not affect significantly the μ, qGlc, and qAnt parameters, in comparison with strain W3110 trpD9923/ pJLBaroGfbr. However, the presence of tktA gene in the plasmid pJLBaroGfbr caused a 1.2-fold increase in YAnt/Glc, resulting in a 1.7-fold higher anthranilate final titer (Table 1) with respect to the strain expressing only aroGfbr. Although the final anthranilate titer accumulated by W3110 trpD9923/pJLBaroGfbrtktA is comparable to that produced by W3110 trpD9923 PTS-, the qAnt of the former strain is 2.3-fold higher (Table 1). The maximum theoretical yield (maxYAnt/Glc) of anthranilate from glucose is 0.435 gAnt/gGlc, considering this value, the YAnt/Glc from W3110 trpD9923/pJLBaroGfbrtktA strain corresponded to 46% of the maxYAnt/Glc.
Previous results demonstrated that the simultaneous expression of aroGfbr and tktA genes caused a 3.3-fold increase in YAnt/Glc and a 2.4-fold increase in the anthranilate titer in strain W3110 trpD9923, thus, in order to increase carbon flux into aromatic biosynthesis and E4P availability, strains W3110 trpD9923 PTS- and W3110 trpD9923 PTS-/pv5Glk5GalP were transformed with plasmid pJLBaroGfbrtktA. The presence of plasmid pJLBaroGfbrtktA in strain W3110 trpD9923 PTS- had a negative impact on the final biomass concentration corresponding to 37% of the strain lacking this plasmid. When compared to W3110 trpD9923 PTS-, no significant changes in qAnt and YAnt/Glc were detected. However the final anthranilate titer was 0.45 ± 0.02 g/L due to the lower biomass concentration (Fig. 3c). Transformation of strain W3110 trpD9923 PTS-/pv5Glk5GalP with plasmid pJLBaroGfbrtktA did not have a significant effect on its growth capacity and the qGlc. In contrast, the qAnt decreased 2-fold but the production phase was 2.5-fold longer than that from the isogenic strain lacking pJLBaroGfbrtktA, therefore, similar final anthranilate titers were produced by both strains (Table 1).
Fed-batch fermentor cultures for anthranilate production
Previous results indicated that anthranilate accumulation occurs mainly during the growth phase in all studied strains. Therefore, to study the effect of extending the growth phase on anthranilate production, all strains were cultured in a fermentor using a fed-batch system with complex medium where a total of 30 g/L yeast extract and 90 g/L glucose were fed in order to improve the final biomass concentration. As Figure 4 shows, all strains displayed growth, glucose consumption and anthranilate accumulation profiles similar to those observed in the flask cultures (Fig. 3). By using a fed-batch process, final biomass concentration was increased an average of 19-fold among all strains (Fig. 4a), when compared to shake-flask conditions (Fig. 3a), likewise, the anthranilate production phase and final anthranilate titer were increased an average of 1.6-fold and 19.4-fold (Fig. 4c), respectively. Analysis of kinetic parameters (Table 2) of all fermentor cultures demonstrated that W3110 trpD9923/pJLBaroGfbrtktA was the best anthranilate producer strain. It accumulated 14 g/L of anthranilate in 34 h with a YAnt/Glc of 0.20 ± 0.00 gAnt/gGlc, the highest values observed among all W3110 trpD9923 derivatives (Table 2). It should be noted that the YAnt/Glc values presented in Table 2 are useful only for comparison among strains grown in the fed batch conditions, since nutrients present in the yeast extract could provide precursors for anthranilate synthesis. With respect to acetic acid production, final titer in W3110 trpD9923 strain was 9.65 ± 2.17 g/L (Table 2). In contrast, a much lower amount of acetic acid (0.50 ± 0.1 g/L) was detected in the medium of W3110 trpD9923/pJLBaroGfbrtktA cultures. In addition, PTS inactivation caused a severe reduction in the production of acetic acid, as it was not detected in the supernatants of all PTS- strains (Table 2).

Fermentor cultures of E. coli W3110 trpD9923 derivative strains for the production of anthranilate. (a) Growth curves, (b) glucose consumption, and (c) anthranilate production. (filled circle) W3110 trpD9923; (open circle) W3110 trpD9923 PTS-; (filled square) W3110 trpD9923 PTS-/pv5Glk5GalP; (X) W3110 trpD9923/pJLBaroGfbr; (open triangle) W3110 trpD9923/pJLBaroGfbrtkt A; (open square) W3110 trpD9923 PTS-/pJLBaroGfbrtkt A; (filled triangle) W3110 trpD9923 PTS-/pv5Glk5GalP/pJLBaroGfbrtkt A. Graphs show results from the mean of the duplicate experiments.
Discussion
In this work, molecular characterization of the trpD9923 mutant allele demonstrated that UV-light treatment generated a nonsense mutation in the trpD gene. As a result of this mutation, gene trpD9923 encodes a truncated anthranilate synthase component II, strongly suggesting that the mutant protein retained glutamine amidotransferase activity and lost the anthranilate phosphoribosyl transferase function. This assumption is consistent with the observed phenotype of strain W3110 trpD9923 (anthranilate accumulation and L-Trp auxotrophy). The identification of the locus and the type of mutation present in strain W3110 trpD9923 will facilitate future efforts for the construction of anthranilate production strains by enabling the generation or transfer of this mutant allele to different microbial species.
Cultures in shake flask and fermentor allowed the characterization of strain W3110 trpD9923 and derivatives with genetic modifications expected to have an impact on anthranilate production capacity. Under the fed-batch conditions utilized in this work, strain W3110 trpD9923 produced 4.2 g/L of anthranilate. With the purpose of improving its performance as a production strain, W3110 trpD9923 was subjected to genetic modifications, following several metabolic engineering strategies expected to improve microbial strains for the production of aromatic amino acids, and more recently in the production of chorismate-derived fine chemicals [15–17]. A key target for improving aromatic amino acids production capacity is the modification of central metabolism to increase PEP and E4P availability [7–9]. Fifty percent of the PEP generated in glycolysis is spent in glucose uptake by the PTS. As the major PEP consuming activity in E. coli, PTS is the main target for inactivation to increase precursor availability for aromatic compounds [10, 11]. PTS inactivation in W3110 trpD9923 caused a 3.2-fold increase in the anthranilate titer in fed batch cultures. Also, PTS inactivation caused a severe reduction in acetic acid production in comparison with the PTS+ strain, eliminating the negative effect of acetate accumulation that is responsible for growth and productivity reduction [18–20].
However, an expected consequence of PTS inactivation was a reduction in qGlc, resulting in 60% lower growth rate of W3110 trpD9923 PTS-. It has been demonstrated that expression of galP and glk genes increases glucose internalization and glycolytic flux to fermentation products in PTS- mutants [10, 13, 21, 22]. In W3110 trpD9923 PTS- the presence of plasmid pv5Glk5GalP effectively increased glucose assimilation capacity as was evident by the higher values of μ and qGlc than those present in W3110 trpD9923 PTS-strain. However, in fed batch cultures, YAnt/Glc and anthranilate titer were reduced in W3110 trpD9923 PTS-/pv5Glk5GalP. A similar effect was reported by Chen et al. [23], where galP expression was ineffective in increasing the L-Phe titers in an E. coli PTS- strain. A possible explanation for this result is that glucose imported by GalP must be phosphorylated by glucokinase, using ATP as the phosphate donor, thus possibly having a negative impact on the cell's energy balance, growth capacity and productivity. An alternate explanation is that the lower qGlc alters carbon flux distribution, resulting in a negative impact on biosynthetic metabolism. This negative effect was evident by the affected growth capacity of PTS- strain and also by the lower values of YBiom/Glc observed in strains PTS- transformed with plasmid pv5Glk5GalP and/or pJLBaroGfbrtkt A (Tables 1 and 2). Metabolic flux redirection in several segments of central metabolism has been reported as a consequence of PTS inactivation and the corresponding lower glucose transport capacity in E. coli [24].
As mentioned, inactivation of PTS is a common modification to improve aromatics biosynthesis in E. coli. The PTS- strains are always further modified to increase their glucose transport capacity, either by isolating spontaneous mutants of by expressing genes encoding alternate glucose transport and phosphorylating activities [10, 22]. In this work, it was found that the residual glucose transport capacity of a PTS- strain is sufficient to allow relatively high anthranilate production capacity. In both shake flask and fermentor cultures, strain W3110 trpD9923 PTS- displayed the second highest final anthranilate titer of all studied strains; however, the productivity was low. When the qGlc and growth capacity were improved in this strain by the expression of galP and glk genes, the final anthranilate titer was reduced. These results suggest that fine-tuning the expression level of galP and glk could allow the development of PTS- production strains having both adequate growth and anthranilate production capacities.
In a wild type E. coli strain, carbon flow into the common aromatic pathway represents only 1.5% of the glucose uptake rate [24]. This is the result of tight regulation of the DAHP synthase isozymes that control carbon entry into this pathway [15]. To overcome this limitation, feedback resistant mutant versions of either one of the three DAHP synthase isozymes have been expressed in engineered aromatics production strains [8, 14, 25, 26]. In the present work, expression of the feedback resistant DAHP synthase aroGfbr in fermentor cultures resulted in a 1.6-fold higher amount of anthranilate accumulated in comparison to W3110 trpD9923 strain, also higher values of qAnt and YAnt/Glc were achieved. In addition to aroGfbr expression, overexpression of the non-oxidative pentose pathway enzyme; transketolase, has been shown to increase E4P availability [27]. The co-expression of aroGfbr and tktA genes in strain W3110 trpD9923 resulted in elevated titers of anthranilate; 14 g/L were obtained under fed-batch fermentor culture. Also, the highest qAnt and YAnt/Glc values of strain W3110 trpD9923/pJLBaroGfbrtkt A evidenced the elevated carbon flux redirection from central metabolism to the product-forming pathway via DAHP. Analysis of kinetic and fermentation parameters from flask cultures of W3110 trpD9923/pJLBaroGfbr and W3110 trpD9923/pJLBaroGfbrtkt A, when compared to W3110 trpD9923, enabled to determine that overexpression of aroGfbr and tktA contributes with 80% and 20% of the increase in YAnt/Glc, respectively. Co-expression of tktA in other E. coli engineered strains has shown a 30–40% increment in aromatic products yields [25, 28–30]. It was also observed that co-expression of aroGfbr and tkt A had an effect in acetic acid production. The expression of aroGfbr caused a 3-fold reduction in final acetic acid titer when compared to W3110 trpD9923. Remarkably, the presence of plasmid pJLBaroGfbrtkt A in strain W3110 trpD9923 caused a 19.3-fold reduction in the final acetate concentration. This result can be explained considering that redirection of PEP to the common aromatic pathway should reduce carbon flow to pyruvate, an intermediate that is both a direct and indirect precursor to acetic acid.
The presence of plasmid pJLBaroGfbrtkt A in all W3110 trpD9923 derivatives caused an increase in qAnt. However, the simultaneous presence of compatible plasmids pJLBaroGfbrtkt A and pv5Glk5GalP in W3110 trpD9923 PTS- resulted in low final biomass concentration, possibly caused by plasmid and gene expression metabolic burden. This negative effect was more pronounced in the trpD9923 PTS-/pJLBaroGfbrtkt A strain, possibly due to a carbon and energy limited condition caused by its lower glucose import capacity resulting from an inactive PTS. Co-expression of aroGfbr and tkt A genes caused a reduction in the YBiom/Glc of 11% in the PTS+ strain and 32% in the PTS-strain in comparison with their parental strains without pJLBaroGfbrtkt A plasmid (Tables 1 and 2). These results suggest that the lower anthranilate titers observed in the trpD9923 PTS-/pJLBaroGfbrtkt A strain is consequence of the susceptibility of the PTS- strain to the metabolic burden caused by gene overexpression.
The microbial synthesis of anthranilate has been previously described using a Bacillus subtillis strain resistant to sulfaguanidine and flourotryptophan [31]. It is reported that fermentor cultures with this strain using minimal medium, resulted in the production on 3.5 g/L of anthranilate and 25 g/L of acetoin after 60 h. In contrast, fermentor cultures using complex medium with strain W3110 trpD9923/pJLBaroGfbrtkt A produced 14 g/L of anthranilate in 34 h and a low level of acetate was detected (0.50 g/L).
The results presented in this work, including the characterization of mutation trpD9923 and the effects on strain productivity of specific genetic modifications, will enable further optimization work focused in exploring additional metabolic engineering strategies and process technology to improve the current E. coli-based production system for the environmentally-compatible synthesis of anthranilate. These efforts should include the evaluation of using environmentally-friendly raw materials such as lignocellulosic hydrolysates and other carbon sources; as glycerol, for the production of anthranilate in a sustainable process.
Methods
Strains and plasmids
Bacterial strains and plasmids used in this study are described in Table 3. E. coli strain W3110trpD9923 was obtained from the E. coli Genetic Stock Center (Yale University, New Haven, CT). E. coli W3110 trpD9923 strain is a mutant in the tryptophan operon obtained by treatment with ultraviolet radiation [3]; it is a tryptophan auxotroph. A PTS- derivative of trpD9923 was obtained by P1 vir phage transduction using PB11 (ΔptsH, ptsI, crr::KmR) strain as donor, as described by Flores et al [10].
Plasmid pJLBaroGfbr carries the aroGfbr gene encoding a feedback inhibition resistant mutant version of the enzyme DAHP synthase under transcriptional control of the lac UV5 promoter [14]. To co-express from this plasmid the gene encoding transketolase, the tktA gene including its native promoter region was amplified by PCR using chromosomal DNA of E. coli W3110 as template and the forward primer 5' GCGCAGCGGACGGGCGAGTAGATTGCGCA 3' and the reverse primer 5' CGCCTGTTCGTTATCTATTCCGCACGCGT CGCG 3', both primers contain the Fsp I site (in bold). The tktA PCR product was cloned into plasmid pJLBaroGfbr previously digested with Bst Z17I enzyme, to generate plasmid pJLBaroGfbrtktA. Plasmid pv5Glk5GalP carries the glk and galP genes, encoding glucokinase (Glk) and galactose permease (GalP), under transcriptional control of a trc-derived promoter [13].
Nucleotide sequence determination of trpED genes
Chromosomal DNA (200 ng) from strain W3110trpD9923 was used as template for PCR amplification using a set of primers designed with the Clone Manager v6.0 software (Scientific and Educational Software, Durham, NC). The primers were designed to bind to different regions of the trpED genes, allowing the determination of the full sequence. Primers used were the following: 5'TAGAGAATAACCATGGAAACACAAAAACCG3', 5'CGCGGATCCCGGTTTGCATCATTTACCCTCG3', 5'CGATTACCAGCAGGCCTCCGGTTGCAGCGTGGTGGCTGGCTCTAG3', 5'ATTCCAGTTCCATCCGGAATCC3', 5'ATCTCGTTCGGGTGCTCACC3', 5'CAGGAGAAAGCATCAGCACC3' and 5'GAGTTCGGTGGCGTAGTGCG3'. PCR reactions were carried out with the Elongase enzyme mix (Invitrogen, Carlsbad, CA) in accordance with the supplier recommendations. PCR products were analyzed for expected size and purified using a PCR purification kit (Marligen, BioScience, Ijamsville, MD). Nucleotide sequences were determined from PCR templates by the Taq FS Dye Terminator Cycle Fluorescence-Based Sequencing method, with an Applied Biosystems Model 377-18 sequencer (Foster City, CA).
Growth media, inoculum preparation and culture conditions
Cells were routinely grown in Luria Bertani (LB) broth or LB agar plates [32]. M9 mineral medium was used for flask cultures, containing 10 g/L glucose, 6 g/L Na2HPO4, 0.5 g/L NaCl, 3 g/L KH2PO4, 1 g/L NH4Cl, 246.5 mg/L MgSO4, 14.7 mg/L CaCl2 and 10 μg/mL vitamin B1, and supplemented with 20 μg/mL tryptophan. Medium for fermentor cultures contained 3 g/L Na2HPO4, 3 g/L KH2PO4, 1.7 g/L (NH4)2HPO4 and 1 mL/L of trace elements solution. This solution contains 27 g/L FeCl3, 2 g/L ZnCl3, CoCl2·6H2O, 2 g/L Na2MoO4·2H2O, 2 g/L CaCl2·2H2O, 0.5 g/L H3BO3 and 100 mL/L HCl.
Fermentor medium initially contained 10 g/L of yeast extract and 30 g/L of glucose. A total of two independent pulses containing 30 g/L glucose and 10 g/L yeast extract were added to the fermentor whenever glucose concentration in the medium decreased to 10 g/L. Each pulse contained 25 mL of 60% glucose solution and 25 mL of 20% yeast extract solution. Antibiotics were added to the corresponding cultures at a final concentration of 30 μg/mL spectomycin, 20 μg/mL tetracycline and 30 μg/mL kanamycin during selection, propagation and fermentation stages.
Inoculum preparation was started using strain samples from frozen vials that were cultured overnight at 37°C in M9 mineral medium plates supplemented with 0.2% of glucose and 20 μg/mL tryptophan, colonies from these plates were used to inoculate baffled shake flasks. For fermentor cultures, colonies from plates were grown in shake flasks with 50 mL LB medium, after overnight culture at 37°C a sample was used for inoculation.
Flask cultures were done in 250 mL flasks containing 50 mL of M9, inoculated at an initial optical density at 600 nm (OD600 nm) of 0.1 and incubated for 60 h at 37°C and 300 rpm in an orbital shaker (Series 25, New Brunswick Scientific, Inc., NJ).
Fermentor cultures were performed in 1 L stirred tank bioreactors (Applikon, The Netherlands), using a working volume of 500 mL. Cultures were inoculated at an initial OD600 nm of 0.5. pH was maintained at 7.0 by automatic addition of a 12.5% NH4OH solution. Temperature was controlled at 37°C. Airflow was set to 1 vvm. Dissolved oxygen tension was measured with a polarographic oxygen electrode (Applisens, Applikon) and maintained above 20% air saturation during all cultivation period by modifying the impeller speed.
For cultures of strains carrying plasmid pJLBaroGfbr, pJLBaroGfbrtktA and/or pv5Glk5GalP gene induction was started by adding IPTG to a final concentration of 0.1 mM at an OD600 nm of 0.6 for shake flask and 3.0 for fermentor cultures.
Kinetic parameters calculation
For the characterization of the strains used in this work, specific rates of growth (μ), glucose consumption (qGlc), anthranilate production (qAnt), yield of biomass on glucose (YBiom/Glc) and yield of anthranilate on glucose (YAnt/Glc) were determined. μ, qGlc and YBiom/Glc were calculated during exponential growth phase. Since growth rates and anthranilate production kinetics differed among studied strains, qAnt and YAnt/Glc were calculated considering only the anthranilate production phase, defined as the time period starting one sample (1 hour) before anthranilate is detected up to the point when a sharp decrease in anthranilate accumulation was observed. Flask cultures were performed in triplicate and fermentor cultures in duplicate. The values reported represent the mean of the experiments performed.
Analytical methods
Biomass concentration was measured as OD600 nm using a spectrophotometer (Beckman DU-70, Palo Alto, CA) and converted to dry cell weight (DCW) considering that 1 OD600 nm = 0.37 gDCW/L [13]. Samples taken during cultivation period were centrifugated at 10000 rpm for 2 min. Supernatant was filtered using 0.45 μm syringe-filter and stored at -20°C for subsequent analysis. Glucose was determined using an enzymatic analyzer (YSI 2700, YSI Life Sciences, OH). Acetate was determined by high performance liquid chromatography (HPLC) (Waters, Milford, MA), using an Aminex HPX-87H column (300 × 7.8 mm; Bio-Rad, Hercules, CA); running conditions were 5 mM H2SO4 as mobile phase, flow of 0.5 mL/min and temperature of 50°C. Detection was performed by photodiode array at 210 nm. Anthranilate was determined by HPLC (Agilent Technologies, Palo Alto, CA) using a Synergy Hydro C18 4 μm column (4.6 × 150 mm, Phenomenex, Torrance, CA); running conditions were 0.1% trifluoroacetic acid in 40% methanol as mobile phase, flow of 0.5 mL/min. Detection was performed by photodiode array at 330 nm. The maximum theoretical yield of anthranilate from glucose (maxYAnt/Glc) was determined by applying elementary mode flux analysis using METATOOL [33].
References
Bell FK: Process for the manufacture of anthranilic acid. US Patent 1, 492, 664. 1924
Kilpper G, Grimmer J: Continuous preparation of anthranilic acid. US Patent 4, 276, 433. 1992
Merino E, Jensen RA, Yanofsky C: Evolution of bacterial trp operons and their regulation. Curr Opin Microbiol. 2008, 11: 78-86. 10.1016/j.mib.2008.02.005.
Xie G, Keyhani NO, Bonner CA, Jensen RA: Ancient origin of the tryptophan operon and the dynamics of evolutionary change. Microbiol Mol Biol Rev. 2003, 67: 303-342. 10.1128/MMBR.67.3.303-342.2003.
Yanofsky C, Horn V, Bonner M, Stasiowski S: Polarity and enzyme functions in mutants of the first three genes of the tryptophan operon of Escherichia coli. Genetics. 1971, 69: 409-433.
Database of the analysis of the http://genolist.pasteur.fr/Colibri/
Berry A: Improving production of aromatic compounds in Escherichia coli by metabolic engineering. Trends Biotechnol. 1996, 14: 250-256. 10.1016/0167-7799(96)10033-0.
Gosset G, Yong-Xiao J, Berry A: A direct comparison of approaches for increasing carbon flow to aromatic biosynthesis in Escherichia coli. J Ind Microbiol. 1996, 17: 47-52. 10.1007/BF01570148.
Ikeda M: Towards bacterial strains overproducing L-tryptophan and other aromatics by metabolic engineering. Appl Microbiol Biotechnol. 2006, 69: 615-626. 10.1007/s00253-005-0252-y.
Flores N, Xiao J, Berry A, Bolivar F, Valle F: Pathway engineering for the production of aromatic compounds in Escherichia coli. Nature Biotechnol. 1996, 14: 620-623. 10.1038/nbt0596-620. 10.1038/nbt0596-620.
Gosset G: Improvement of Escherichia coli production strains by modification of the phosphoenolpyruvate:sugar phosphotransferase system. Microb Cell Fact. 2005, 4: 14- 10.1186/1475-2859-4-14.
Postma PW, Lengeler JW, Jacobson GR: Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol Rev. 1993, 57: 543-594.
Hernández-Montalvo V, Martínez A, Hernández-Chávez G, Bolívar F, Valle F, Gosset G: Expression of galP and glk in an Escherichia coli PTS mutant restores glucose transport and increases glycolytic flux to fermentation products. Biotechnol Bioeng. 2003, 83: 687-694. 10.1002/bit.10702.
Báez-Viveros JL, Osuna J, Hernández-Chávez G, Soberón X, Bolívar F, Gosset G: Metabolic engineering and protein directed evolution increase the yield of L-phenylalanine synthesized from glucose in Escherichia coli. Biotechnol Bioeng. 2004, 87: 516-524. 10.1002/bit.20159.
Bongaerts J, Krämer M, Müller U, Raeven L, Wubbolts M: Metabolic engineering for microbial production of aromatic amino acids and derived compounds. Metab Eng. 2001, 3: 289-300. 10.1006/mben.2001.0196.
Kern A, Tilley E, Hunter IS, Legisa M, Glieder A: Engineering primary metabolic pathways of industrial micro-organisms. J Biotechnol. 2007, 129: 6-29. 10.1016/j.jbiotec.2006.11.021.
Sprenger GA: From scratch to value: engineering Escherichia coli wild type cells to the production of L-phenylalanine and other fine chemicals derived from chorismate. Appl Microbiol Biotechnol. 2007, 75: 739-749. 10.1007/s00253-007-0931-y.
De Anda R, Lara AR, Hernández V, Hernández-Montalvo V, Gosset G, Bolívar F, Ramírez OT: Replacement of the glucose phosphotransferase transport system by galactose permease reduces acetate accumulation and improves process performance of Escherichia coli for recombinant protein production without impairment of growth rate. Metab Eng. 2006, 8: 281-290. 10.1016/j.ymben.2006.01.002.
Koh BT, Nakashimada U, Pfeiffer M, Yap MGS: Comparison of acetate inhibition on growth of host and recombinant Escherichia coli K-12 strains. Biotechnol Lett. 1992, 14: 1115-1118. 10.1007/BF01027012. 10.1007/BF01027012.
Turner C, Gregory ME, Turner MK: A study of the effect of specific growth rate and acetate on recombinant protein production of Escherichia coli JM107. Biotechnol Lett. 1994, 16: 891-896. 10.1007/BF00128620. 10.1007/BF00128620.
Báez JL, Bolívar F, Gosset G: Determination of 3-deoxy-D-arabino-heptulosonate 7-phosphate productivity and yield from glucose in Escherichia coli devoid of the glucose phosphotransferase transport system. Biotechnol Bioeng. 2001, 73: 530-535. 10.1002/bit.1088.
Yi J, Draths KM, Li K, Frost JW: Altered glucose transport and shikimate pathway product yields in E. coli. Biotechnol Prog. 2003, 19: 1450-1459. 10.1021/bp0340584.
Chen R, Yap WMGJ, Postma PW, Bailey JE: Comparative studies of Escherichia coli strains using different glucose uptake systems: metabolism and energetics. Biotechnol Bioeng. 1997, 56: 583-590. 10.1002/(SICI)1097-0290(19971205)56:5<583::AID-BIT12>3.0.CO;2-D.
Flores S, Gosset G, Flores N, de Graaf AA, Bolívar F: Analysis of carbon metabolism in Escherichia coli strains with an inactive phosphotransferase system by 13C labeling and NMR spectroscopy. Metab Eng. 2002, 4: 124-137. 10.1006/mben.2001.0209.
Lütke-Eversloh T, Stephanopoulos G: Combinatorial pathway analysis for improved L-tyrosine production in Escherichia coli: identification of enzymatic bottlenecks by systematic gene overexpression. Metab Eng. 2008, 10: 69-77. 10.1016/j.ymben.2007.12.001.
Patnaik R, Spitzer RG, Liao JC: Pathway engineering for production of aromatics in Escherichia coli: confirmation of stoichiometric analysis by independent modulation of AroG, TktA, and Pps activities. Biotechnol Bioeng. 1995, 46: 361-370. 10.1002/bit.260460409.
Frost JW: Enhanced production of common aromatic pathway compounds. US Patent 5, 168, 056. 1992
Berry A, Dodge TC, Pepsin M, Weyler W: Application of metabolic engineering to improve both the production and use of biotech indigo. J Ind Microbiol Biotechnol. 2002, 28: 127-133. 10.1038/sj.jim.7000228.
Jung YM, Lee JN, Shin HD, Lee YH: Role of tktA gene in pentose phosphate pathway on odd-ball biosynthesis of poly-beta-hydroxybutyrate in transformant Escherichia coli harboring phbCAB operon. J Biosci Bioeng. 2004, 98: 224-227.
Yakandawala N, Romeo T, Friesen AD, Madhyastha S: Metabolic engineering of Escherichia coli to enhance phenylalanine production. Appl Microbiol Biotechnol. 2008, 78: 283-291. 10.1007/s00253-007-1307-z.
Cooper B, Meyer J, Euler K: Production of anthranilic acid by a strain of Bacillus subtillis resistant to sulfaguanidine and fluorotryptophan. US Patent 5, 422, 256. 1995
Sambrook J, Fitsch EF, Maniatis T: Molecular Cloning: A Laboratory Manual. 1989, Cold Spring Harbor, Cold Spring Harbor Press
Schuster S, Dandekar T, Fell DA: Detection of elementary flux modes in biochemical networks: A promising tool for pathway analysis and metabolic engineering. Trends Biotechnol. 1999, 17: 53-60. 10.1016/S0167-7799(98)01290-6.
Acknowledgements
We thank Eugenio Meza Mora for his help with elementary mode analysis and Mercedes Enzaldo for her technical assistance. CONACyT D43243-Z grant supported this work. Victor E. Balderas Hernández is thankful for the postdoctoral fellowship support by the DGAPA-UNAM.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
VEBH carried out the production experiments and wrote the manuscript. AS, PS and NCV constructed the plasmids and strains. GHC assisted the determination of metabolites. JLBV, AM and FB participated in the design of the study. GG coordinated the study and wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Balderas-Hernández, V.E., Sabido-Ramos, A., Silva, P. et al. Metabolic engineering for improving anthranilate synthesis from glucose in Escherichia coli. Microb Cell Fact 8, 19 (2009). https://doi.org/10.1186/1475-2859-8-19
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2859-8-19