
Abstract
Aims
Intercropping increases aboveground and belowground crop productivity, suggesting potential for carbon sequestration. Here we determined whether intercropping affects decomposition of soil organic matter (SOM) and root litter.
Methods
We measured in the laboratory and the field the breakdown of SOM, root litter of maize, wheat, or faba bean, litter mixtures, and a standard substrate (compost) in soils from a long term intercropping experiment.
Results
Soil organic matter from intercrop plots decomposed faster than SOM from monocrop plots, but compost decomposed at similar rates in different soils. Faster SOM decomposition was associated with lower soil C:N ratio. Root litter mixtures of maize and wheat decomposed as expected from single litters, but litter mixture of maize and faba bean decomposed faster than expected, both in the laboratory and in the field. Root litter decomposed slowly in maize/wheat intercrop soil compared to the two monocropped soils in the laboratory, but the effect was absent in the field.
Conclusions
Intercropping increases SOM decomposition, presumably through reduced SOM recalcitrance resulting from lower C:N ratio, higher litter input and better N retention. Depending on the crop combination, also non-additive effects of root litter mixing can enhance organic matter decomposition in intercropping soils.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Intercropping is an agricultural practice in which two or more crop species or varieties are grown together in the same field. It is widely practiced by smallholder farmers in developing countries because it enhances the amount of biomass and crop yield that is harvested from a piece of land (Willey 1979; Lithourgidis et al. 2011). This higher production is attributed to an increase in total resource capture due to complementarity between the intercropped species (Hinsinger et al. 2011), and to reduced impacts of pests, diseases and weeds (Liebman and Dyck 1993; Trenbath 1993; Zhu et al. 2000). The greater biomass production aboveground is mirrored belowground (Ghosh et al. 2006; Yang et al. 2010; Li et al. 2011; Cong et al. 2014a) and a recent study showed that intercropping increases soil organic carbon (C) stocks (Cong et al. 2014a). However, the increase in C stocks over 7 years of intercropping (4 %) was smaller compared to the estimated increases in yearly root litter input in intercrops as compared to monocrops (23 %) (Cong et al. 2014a). The comparatively small increase in C stocks suggests that increased root C input is perhaps in part compensated by larger C losses through decomposition of soil organic matter (SOM) and/or fresh root litter. In this study we test this hypothesis.
Intercropping could potentially affect the rate of decomposition of organic matter in soil via two pathways. The first pathway is via the greater diversity and quantity of the root litter added to the soil in intercropping. A second pathway could be through changes in the abiotic or biotic components of the decomposition environment, as a result of cumulative changes in the soil ecosystem resulting from intercropping over the longer term.
Effects of litter diversity and quantity
Plant diversity results in increased diversity of the litter. Studies manipulating the diversity of litter often reported the so-called “litter mixing effects”: mixing litters accelerated or inhibited decomposition compared to the average decomposition of the mixture components (Bardgett and Shine 1999; Hättenschwiler et al. 2005). These phenomena are often observed when the components of litter vary in their quality as a substrate for decomposition (Wardle et al. 1997). For example, nitrogen (N) rich litter can accelerate decomposition of N poor litter (Pérez-Harguindeguy et al. 2008; Vos et al. 2013; Handa et al. 2014). This is possible because N transfer from N rich litter to N poor litter relieves N limitation in the decomposer community, thus accelerating decomposition of the N poor litter (Seastedt 1984). Alternatively, when one of the litter component contains secondary compounds (e.g., polyphenols), mixing litters can inhibit microbial growth and activity, consequently impeding decomposition of the other mixture component (Schimel et al. 1998). Since residues of annual crops in agroecosystems are usually low in secondary compounds (Cadisch and Giller 1997), inhibition probably plays a minor role in intercrops.
The quantity of litter may also affect decomposition, and litter quantity may be related to plant diversity, especially in the long run, due to positive relationships and feedbacks between diversity, nutrient retention and productivity (Cong et al. 2014a, b). Several long-term biodiversity experiments in grassland ecosystems have shown higher rates of SOM decomposition in species-diverse plots than in monocultures (Zak et al. 2003; Dijkstra et al. 2005; Cong et al. 2014b). Soil organic matter consists of a variety of plant and animal residues at various stages of decomposition and different levels of recalcitrance towards further decomposition. Dijkstra et al. (2005) attributed higher decomposition rate of SOM largely to the greater soil C inputs resulting from increased biomass production in species-diverse plots. A greater litter input results in a rejuvenation of SOM stock, increasing the proportion of relatively young, labile C in total soil C, thereby reducing the overall recalcitrance of SOM, and accelerating SOM decomposition. This mechanism might play a role in intercropping, because intercrops have greater aboveground biomass (Willey 1979; Lithourgidis et al. 2011) and root biomass (Ghosh et al. 2006; Yang et al. 2010; Li et al. 2011; Cong et al. 2014a) than monocultures. In a recent 7-year study, we found that intercropping decreased the soil C/N ratio compared to the corresponding monocrops, regardless of the presence of legumes (Cong et al. 2014a). A lower C/N ratio is in general associated with higher rates of decomposition (Booth et al. 2005). Hence, intercropping could enhance SOM decomposition as a result of cumulative reductions in the recalcitrance of SOM. This hypothesis is as yet untested.
Decomposition environment effect
Another possible mechanism for intercropping effects on organic matter decomposition is alteration of the decomposition environment through changes in abiotic and biotic attributes of the soil ecosystem. Such changes may be expected because intercropping increases soil carbon and soil nitrogen (Cong et al. 2014a). Diversity effects on the decomposition environment may be explored by incubating a standard substrate in soils from different diversity treatments. Grassland biodiversity experiments have shown inconsistent effects of plant diversity on decomposition of standard substrates (e.g., senesced leaves), ranging from faster decomposition (Hector et al. 2000; Knops et al. 2001; Spehn et al. 2005; Vogel et al. 2013), no effect (Spehn et al. 2000; Scherer-Lorenzen 2008) to slower decomposition (Fornara et al. 2009). Plant diversity has frequently been found to affect the soil microbial decomposer community, either through a more diverse litter composition (Chung et al. 2007), or through higher biomass production (Zak et al. 2003), or through soil fauna-driven effects on soil structure (Stephan et al. 2000). Effects of plant diversity on organic matter decomposition are therefore variable.
Little is known of intercropping effects on organic matter decomposition. Song et al. (2007) and He et al. (2013) reported that intercropping modifies soil microbial properties, but did not report on organic matter decomposition. Li et al. (2005) reported that intercropping decreased concentrations of soil nitrate in the soil through enhanced N uptake, which could decelerate decomposition of substrates with a high C/N ratio. Oliveira et al. (2002) and Vachon and Oelbermann (2011) found faster decomposition of N poor residues by mixing with N rich residues of legume crops in intercrops. But neither of them unravels the above-mentioned potential mechanisms underlying the results and no studies have been done on mixtures without legumes.
In this study, we aim at separating the intercropping-induced effects of litter diversity and quantity from those of the decomposition environment using soil samples from a 7-year field experiment with maize, wheat and faba bean as monocrops and maize/wheat and maize/faba bean as intercrops. First, we incubated these five soils in the laboratory without any amendments to investigate whether intercropping affects the decomposition of the native SOM. Secondly, we assessed whether there were differences in decomposition environment between these soils by analysing decomposition of a single substrate, compost. Third, we used a factorial design to study the decomposition of single and mixed root litter of maize, wheat and faba bean in soils on which these or other species had been grown. Experiments were carried out both in the laboratory using CO2 efflux measurements and in the field using litter bags.
Materials and methods
Soil sampling
Soils were taken from a long-term field experiment that was initiated in 2003 in Wuwei, Gansu, China, to study effects of intercropping and rotation on productivity and soil properties (Li et al. 2007). The average annual temperature at the site is 8.9 °C. The climate is arid with a total yearly rainfall of 168 ± 8 mm and potential evaporation of 2021 mm. Soil at the site is a sandy loam, and classified as an Aridisol. The experiment comprises thirteen crop systems based on maize, wheat and faba bean. The experiment is laid out as a completely randomized block design with three blocks. For the experiments reported here, we used soil from five continuous cropping systems: (1) maize (Zea mays L.) monocropping (hereafter referred to as “Maize”), (2) wheat (Triticum aestivum L.) monocropping (“Wheat”), (3) faba bean (Vicia faba L.) monocropping (“Faba bean”), (4) maize/wheat intercropping (“Maize/Wheat”) and (5) maize/faba bean intercropping (“Maize/Faba bean”). In the intercropping treatments, the two species were cultivated yearly with each of the two crops growing on exactly the same place. The maize/wheat and maize/faba bean intercropping combinations are examples of a C3/C4 cereal combination and of a C3 cereal/C3 legume combination, respectively. These combinations result in different complementary resource acquisition strategies of the intercropped species and are therefore interesting cases exemplifying crop diversity effects on decomposition.
Maize monocrop was grown at a row distance of 40 cm, wheat monocrop at 13.3 cm and faba bean monocrop at 20 cm. The maize/wheat intercrop was planted in 1.6 m-wide strips that consisted of an 80 cm maize strip (2 rows at 40 cm distance) and an 80 cm wheat strip (6 rows at 13.3 cm row distance). The distance between adjacent maize and wheat rows was 26.7 cm. The maize/faba bean intercrop was planted in 1.6 m-wide strips that consisted of two rows maize at 40 cm row distance and four rows faba bean at 20 cm row distance. The distance between adjacent maize and faba bean rows was 30 cm. The intercrops thus had a relative density of both species of 50 % as compared to the monocrops. Wheat and faba bean were sown in late March and harvested early July (wheat) or late July (faba bean). Maize was sown in mid-April and harvested early October. Wheat and faba bean monocropping received flood irrigation five times during the growing season, while the other treatments (all including maize) were irrigated two more times after harvest of wheat or faba bean to meet the water demand of maize. Each plot received 225 kg ha−1 N year−1 as urea and 40 kg ha−1 year−1 P2O5 as triple superphosphate. The amount of N fertilization was chosen to ensure that N supply is sufficient to meet the requirement of the most N-demanding crop, i.e., maize. Two thirds of N were applied and all P fertilizers were applied at moldboard tillage (down to 20 cm depth). The remaining N fertilizer was top-dressed at wheat flowering in late May. Aboveground crop residues were removed after harvest for feed or fuel.
Soil samples (0–20 cm depth) were taken on 1 July 2010, when wheat was at dough stage, faba bean in pod-filling stage, and maize in vegetative stage (collar of 10th leaf visible). Twenty cores per plot were taken in the middle between two rows in monocropping treatments. For intercropping, we took twenty cores in the middle between the rows of the two crop species. Sampling time and place were such that root-root interactions between intercropped species are most intense (Li et al. 2006) and microorganisms are probably most active. Soil samples were bulked per plot, transported to the lab, sieved (2 mm) to remove visible fresh plant material and macrofauna, and stored at 4 ° C until further processing. A subsample was air-dried for 3 days and treated with HCl to remove inorganic C (Midwood and Boutton 1998). The acid-treated soil samples were ground in a ball mill prior to analysis.
Fresh roots (<2 mm diameter) of wheat, maize and faba bean were collected in June 2011 in monocrop plots, washed to remove soil particles, oven-dried at 70 °C for 48 h, and cut into pieces of approximately 1 cm length. C and N mass fractions of soil and root samples were determined prior to incubation by a vario MACRO Elemental Analyser (Elementar Company, Germany). C and N mass fractions of soil samples taken at the start of the experiment in 2003 were determined as a reference, using the same procedures.
Decomposition experiments
We conducted three experiments to examine intercropping effects on decomposition of SOM, compost (standard substrate) and root litter.
Experiment 1: Decomposition of SOM and compost in the laboratory
In the first experiment, we determined whether intercropping had resulted in differences between soils in the decomposition of the native organic matter. Soil samples were taken from the five treatments in the long-term field experiment as described. Measurements of CO2 production were done during incubation of the soils in the lab. The comparison of SOM decomposition cannot distinguish between differences in decomposition that are due to the nature of the SOM itself or to differences in the decomposition environment. Therefore a set of treatments with compost was added in soils from the five treatments with three replicates. The compost was a highly humified organic matter with a C mass fraction of 171 g/kg, an N mass fraction of 10.6 g/kg and a C/N ratio of 16. It was produced from urban waste (vegetables, fruits and garden wastes). In a similar aerobic lab incubation, this compost decomposed at a rate similar to that of soil organic matter (Rietberg 2009). If soils would decompose compost at a different rate, this would signal differences in the decomposition environment. It is not possible to separate the CO2 production from compost from that originating from SOM and priming effects of adding compost to the soil cannot be ruled out (Blagodatskaya and Kuzyakov 2011). Here, we refer to the difference in CO2 production between soils with and without compost as the compost-induced CO2 production, which includes a priming effect if it exists. Effects of the decomposition environment are identified using this “additional” CO2 production, induced by adding compost.
Standard amounts of moist soil (50 g on dry weight basis) were placed in 250 mL-bottles that were closed with a gas-permeable cotton plug and incubated in the laboratory at 20 °C. In half of the bottles soil was mixed with 0.5 g oven-dried (70 °C) compost. Soil moisture content was kept at 60 % of water holding capacity by supplying deionized water every 2 weeks. The production of CO2 was measured at 2, 4, 8 and 16 weeks after filling the bottles, avoiding the first 2 weeks during which priming effects may be strong (Nottingham et al. 2009; Pascault et al. 2013). Prior to these measurements, the bottles were flushed with compressed air for 15 min and closed with an air-tight lid and then incubated in the dark at 20 °C for 4 h. The increase in CO2 concentration after this 4 h incubation was monitored using a photo-acoustic gas monitor (INNOVA 1412, AirTech Instruments, Ballerup, Denmark). Compost-induced CO2 production was calculated as the difference between the CO2 efflux produced by compost-amended soil and the CO2 efflux from soil alone.
Experiment 2: Decomposition of root litter in the laboratory
A second experiment addressed CO2 production from root litter in the laboratory, and aimed at identifying differences in decomposition rate between single and mixed litters and between decomposition in soils originating from monocrop plots and decomposition in soils from intercrop treatments. We used a full factorial design with “soil origin” (i.e., decomposition environment) and “root litter” as factors. This factorial experiment combined two monocrop soils and one intercrop soil with two single litters and a root litter mixture in all 3 × 3 combinations. Each soil treatment had three replicates. The experiment was executed for two crop combinations: maize-wheat and maize-faba bean (Table 1).
A standard amount of moist soil (50 g on dry weight basis) was mixed with 0.5 g oven-dried (70 °C) root litter. The amount of root litter added to soil ensured sufficient C for microbial respiration during incubation. Mixtures contained 0.25 g root litter of each species. The incubation and monitoring procedures were the same as described for Experiment 1. Root-induced CO2 production was calculated as the difference between the CO2 efflux from root-amended soil and the CO2 efflux from soil alone as measured in Experiment 1.
Experiment 3: Decomposition of root litter in the field
Root litter decomposition was studied in the field in Wuwei, Gansu, China, in 2011 and 2012 by monitoring root mass loss in litterbags over a period of 12 months. The factorial design of experiment 2 (Table 1) was replicated in situ. Bags (5 × 5 cm; 250 μm nylon mesh) were filled with 1 g (dry weight basis) of oven-dried root samples. The root mixtures contained 0.5 g root litter of each species. A total of 306 litterbags (seventeen root × soil combinations, two litter bags for each combination, three field blocks and three exposure durations: 17*2*3*3 = 306) were buried in the soil at 10 cm depth in the long-term experiment on 15 June 2011. The bags were placed in the middle between two rows in monocrop treatments, and in the middle between rows of different crop species in intercrops. The litter bags were excavated at exactly 2, 6, and 12 months after burial, and oven-dried (70 °C, 48 h). Decomposing root litter was gently retrieved by carefully brushing away adhering soil particles and other extraneous material, and then weighed. Root mass loss was calculated by subtracting the dry mass of roots remaining at the sampling time from the initial mass. The results from the two litter bags from each combination were averaged prior to further analysis.
Quantitative comparison of decomposition rates
To quantitatively describe the decomposition process and compare treatment effects, we fitted mathematical models to measured CO2 effluxes in laboratory trials (Experiments 1 and 2). A simple first-order exponential decay model did not fit the data satisfactorily. We therefore applied the Yang and Janssen model (Yang and Janssen 2000), which has a time-dependent relative decomposition rate:
where Y t and Y 0 are the amounts of organic C present at time t and time 0, respectively. K 1 is the average relative decomposition rate during the first unit of time after t = 0, and S measures the speed at which the relative decomposition rate decreases over time. Greater values of S mean a faster decline in decomposition rate. This model accounts for a slowing down of the decomposition rate over time as the labile components of a substrate decompose first, and more recalcitrant components make up an increasing proportion of the material over time. Measured CO2 production over the time of incubation (4 h) was converted to daily C efflux, ΔCO2-C, by multiplication with six (24 h/4 h) and molecular mass ratio (C/CO2) of 12/44.
The CO2-C production at any time t is given by the first derivative of Eq. 1:
Equation 2 was fitted to ΔCO2-C, solving for K 1 and S, using non-linear regression (SPSS 20.0). Ordinary sums of squares (R2) was used as goodness of fit criterion. Fitting was done for each individual plot in the field from which the soil originated, resulting in a mean and standard error of fitted model parameters for each soil origin and each substrate. The calibrated parameters \( {K}_1 \) and S were used to calculate the Cumulative amount of C Decomposed (CCD, g CO2-C kg−1 soil) of the sample over 112 days of incubation. If the Yang and Janssen model did not describe the data satisfactorily, we used linear interpolation between subsequent measurements to calculate CCD (Muller-Stover et al. 2012). CCD was expressed as a percentage of the initial amount of C in the sample:
where C0 is the amount of C in the sample at t = 0 (g C kg−1 soil).
Calculation of expected decomposition
The expected percentage decomposition, under the assumption of only additive effects of intercropping, of native organic matter in a soil sample from an intercrop of species A and B was calculated as the weighted mean decomposition of SOM from plots with monocrops of A and B as:
where CDA (%) and CDB (%) are the percentage of C decomposed from the soil samples from monocrops species A and species B, respectively, while CA and CB are the organic C contents (g C kg−1 soil) of soils of monocrops species A and B.
Equation 4 was again used to calculate expected root litter decomposition in the field (Experiment 2). While the shares of the two species in the root sample were 50/50 on dry weight basis, the higher root C concentration of faba bean resulted in a slightly higher contribution of this species to total amount of C in the root mixture.
The expected root mass loss (%) from a root litter mixture of species A and B in the field (Experiment 3) was calculated as the simple average of root mass loss (%) of monocrop species A and B because the mixture consisted of equal amounts of component root litters.
In all experiments, expected amounts were calculated separately for each block to enable statistical analysis.
Statistical analyses
One-way analysis of variance (ANOVA) was used to compare C and N mass fractions and C/N ratio of soils and roots. Repeated-measures analysis of variance was used to analyse the effect of intercropping on the time course of CO2 production from native SOM (Experiment 1). Two-way ANOVA was used to determine the effects of compost addition and decomposition environment on SOM decomposition (Experiment 1). Two-way ANOVA was again used to evaluate the main and interactive effects of root litter type and decomposition environment on the percentage of C decomposed, CD (%), after 112 days of incubation in the laboratory (Experiment 2) and on root mass loss after 1 year in the field (Experiment 3). Once a main effect was found significant and there was no interaction between the two factors, Tukey’s post hoc test was used for pair-wise comparisons. One-way ANOVA was further used to test whether the rate of decomposition of mixed litter is different from the expected values calculated from the single litters (i.e., presence of a litter mixing effect). All data met the ANOVA assumptions of normality and homogeneity of variance.
Results
C and N content and C/N ratio of soil and root samples
Soil C and N content had significantly increased in all cropping systems after 7 years, but there were no significant differences in the C and N content and C/N ratio among treatments in 2010 (Table 2). We conducted a two-way ANOVA with intercropping (expected vs. observed) and crop combination (maize/wheat and maize/faba bean) as two factors. Expected C or N content in an intercrop soil was calculated as the average of the two monocrop soils within the same block. We found that intercropping increased soil C content slightly (2 %) but not significantly (P = 0.16, data not shown) compared to expected. However, intercropping significantly increased soil N content, by 8 % (P = 0.008) compared to expected, leading to decreased soil C/N ratio (P = 0.013). These effects were independent of intercropping combination (P for interaction: 0.83 for soil C; 0.87 for soil N; 0.34 for soil C/N ratio) and therefore not related to the presence of a legume (faba bean). This implies that the SOM in intercrop plots was enriched in N, potentially facilitating its decomposition.
Root C and N content and C/N ratio were significantly different among maize, wheat and faba bean (P = 0.022 for root C; P = 0.002 for root N; P = 0.007 for root C/N ratio; Table 2). Maize and wheat roots had similar C and N contents, but C content was somewhat higher and N content slightly but significantly higher in faba bean root than in wheat or maize roots. The C/N ratio of faba bean root was lower than of maize or wheat roots.
Experiment 1: Decomposition of SOM and compost in the laboratory
Production of CO2 as a result of SOM decomposition from most of the soil samples from the long term intercropping experiment tended to first increase, then decrease during incubation under laboratory conditions (Fig. 1a). There were significant differences in the time courses among the soils from the five cropping systems (Cropping system × time in repeated measures ANOVA: P = 0.01): the CO2 production was significantly higher from soils originating from the two intercrop treatments than from maize monocrop at 28 and 112 days but no significant treatment effects were observed at 14 and 56 days.

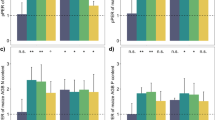
Decomposition of soil organic matter during laboratory incubation of soils from intercrop plots (Maize/Wheat and Maize/Faba bean) and from monocrop plots (Maize, Wheat or Faba bean) (Experiment 1). a Time course of CO2 production; b Soil organic C decomposed (%) between day 14 and day 112; c Expected and observed soil organic C decomposed (%) and d the relationship between soil organic C decomposed (%) and soil C/N ratio. Expected values were calculated as the weighted average soil organic C decomposed (%) of monocrop soils with the same crop species, using Eq. 4 (See Mat. & Meth.). Data are means ± SEM, n = 3. Asterisks refer to significant differences between treatments (P < 0.05). Means with the same letter are not significantly different (Tukey’s post hoc test, P < 0.05)
We used linear interpolation between observed CO2 fluxes at different times to calculate the percentage of C decomposed over the incubation period (Eq. 3) because the Yang and Janssen model gave an unsatisfactory fit to the CO2 production data (R2 ranging from 0.10 to 0.72, with a mean R2 of 0.39). The percentage of C decomposed was significantly higher in soil samples from maize/wheat and maize/faba bean intercrop plots than from maize plots (P = 0.03; Fig. 1b). The soil samples from wheat and faba bean plots were intermediate.
Regardless of crop combination (Intercropping × combination: P = 0.65), intercropping significantly increased decomposition of native SOM (P = 0.01; Fig. 1c): The observed C decomposed (%) was 1.3 times the expected in maize/wheat intercropping and 1.5 times the expected in maize/faba bean intercropping, where the expected percentage of C decomposed for the intercropping samples was calculated on the basis of the CO2 production measured in the samples from monocrops (Eq. 4). Furthermore, we found that SOM decomposition was negatively related to soil C/N ratio (R2 = 0.66; P < 0.001; Fig. 1d). The relationship was determined by two monoculture maize plots that both had a relatively high C/N ratio and slow SOM decomposition.
To examine whether the effect of intercropping on SOM decomposition acted through the decomposition environment, we added compost to soil samples. The total CO2 production from soil plus compost increased significantly (by 23 %) due to addition of compost (P < 0.01; Fig. 2), independent of the origin of the soil (Cropping system × compost: P = 0.92). The extra CO2 production due to compost addition was similar across the soils from the five cropping systems (P = 0.35), indicating that there were no differences between the soils in their ability to facilitate decomposition of a standard substrate. These results indicate that the intercropping affected decomposition of native SOM due to attributes of the organic matter. Differences in native SOM decomposition between treatments were associated with (but not fully explained by) C/N ratio of the soil.

Cumulative CO2 production between day 14 and day 112 during laboratory incubation of soils from intercrop plots (Maize/Wheat and Maize/Faba bean) and from monocrop plots (Maize, Wheat or Faba bean) with and without compost amendment (Experiment 1). Data are means ± SEM, n = 3
Experiment 2: Decomposition of root litter in the laboratory
Interactive effects of root litter diversity and soil environment were studied by measuring CO2 production in the laboratory upon adding different litters to different soils, and subtracting the CO2 production resulting from native SOM decomposition as quantified in Experiment 1. Root-induced CO2 production rates were fitted to the decomposition model by Yang and Janssen (2000) to calculate total decomposition over time by integration, and compare time courses. The model fitted the data well, with R2 ranging from 0.94 to 1.00, with a mean R2 of 0.99. The model with parameters K 1 and S calibrated per each treatment and replicate was then used to estimate the cumulative amount of root C decomposed over 112 days of incubation, expressed as a percentage of the added root C.
In the maize-wheat crop combination, we found that both root litter and soil significantly affected the percentage of C decomposed (P < 0.001 for root litter type; P < 0.001 for decomposition environment) without interaction (P = 0.99). The percentage of C decomposed was higher for maize than for wheat roots, and the root mixture of maize and wheat was intermediate (Fig. 3a). There was no significant difference between observed percentage of C decomposed of the root mixture of maize and wheat, and the expected value calculated from the single litters (P = 0.89), indicating absence of a litter mixing effect on decomposition. The percentage of root C decomposed was lower in maize/wheat intercrop soil than in maize or wheat monocrop soil (Fig. 3a; P < 0.001), independent of root litter type (Litter type × decomposition environment: P = 0.99). Thus maize/wheat intercropping slowed down root decomposition through a decomposition environment effect.

Decomposition of root litter during 112 days of incubation in the laboratory (Experiment 2) in full factorial experiments crossing soils from monocrop or intercrop plots with single or mixed root litters for (a) the maize/wheat combination and (b) the maize/faba bean combination. Expected root C decomposed (%) is calculated as the weighted average decomposition of single root litters using Eq. 4 (See Mat. & Meth.). Data are means ± SEM, n = 3
In the maize-faba bean crop combination, root litter type significantly affected the percentage of C decomposed (P < 0.001), while decomposition environment did not affect it (P = 0.10). The percentage of C decomposed was lower for maize than for faba bean roots or the root mixture of maize and faba bean (Fig. 3b). The observed decomposition rate of the mixture of maize and faba bean roots was slightly (2 %) and significantly (P = 0.02) greater than expected from the single root litters. The accelerating effect of the root mixture was independent from the decomposition environment (interaction: P = 0.47).
In both the maize/wheat and maize/faba bean intercropping combinations, similar results as presented above for 112 days of incubation were obtained with 14, 28 and 56 days of incubation.
Experiment 3: Decomposition of root litter in the field
The laboratory studies on decomposition of pure or mixed root litters of maize, wheat and faba bean in five different soil backgrounds were replicated in the field in Gansu. In the maize-wheat crop combination (Fig. 4a), root mass loss (%) after 1 year significantly differed between root litter types (P < 0.001), similarly as in the laboratory experiment (Experiment 2): maize roots decomposed faster than wheat roots, and the root mixture of maize and wheat decomposed at an intermediate rate in accordance with expectation (P = 0.89). This indicates litter mixing did not affect root decomposition. The absence of a litter mixing effect was consistent across different soils (Litter mixing × decomposition environment: P = 0.85). Decomposition environment did not have a significant effect on root litter decomposition (P = 0.19), regardless of root litter type (Litter type × decomposition environment: P = 0.47).

Decomposition of root litter in litter bags in the field (Experiment 3) in full factorial experiments crossing soils from monocrop or intercrop plots with single or mixed root litters, expressed as root mass loss (%) after 12 months in (a) the maize/wheat combination and (b) maize/faba bean combination. Expected root mass loss (%) of root litter mixtures is calculated as the simple average root mass loss of single litters in the same soil. Data are means ± SEM, n = 3
In the maize/faba bean combination (Fig. 4b), maize roots and the mixture of maize and faba bean roots had greater root mass loss than faba bean roots (P < 0.001). The maize/faba bean root mixture decomposed faster (6 %) than expected from the single litters (P = 0.002), independent of the decomposition environment (Litter mixing × decomposition environment: P = 0.56). This suggests a positive litter mixing effect in root mixture of maize and faba bean. Decomposition environment did not affect root litter decomposition (P = 0.57).
Discussion
Intercropping effects on SOM decomposition
Our study provides, to our best knowledge, the first evidence that intercropping accelerates the decomposition of soil organic matter (SOM). The observed C decomposed (%) was 1.3 times the expected in maize/wheat intercropping and 1.5 times the expected in maize/faba bean intercropping. Faster decomposition of SOM in intercropping soil can explain why intercropping increased soil C content only slightly whereas root biomass sampling indicated substantial increases in root biomass in intercrops as compared to sole crops (Cong et al. 2014a). Only one previous study compared decomposition rate of SOM in monocrop and intercrop soils (Rusinamhodzi et al. 2009), finding no significant differences. The contrast between the positive finding in our study and the negative finding in the study of Rusinamhodzi et al. (2009) may be the length of the intercropping history of the soil, which was 7 years in our study and only 2 years in the other study. It is possible that the enhancing effect of intercropping on decomposition develops over time as carbon and nitrogen are sequestered in the soil (Cong et al. 2014a). A similar finding was made in a grassland biodiversity study (Cong et al. 2014b).
Intercropping enhanced SOM decomposition most likely through changes in its quality. We found that adding compost produced similar increases in CO2 efflux regardless of cropping systems from which the soils originated, indicating that the capacity of the soils’ microbial communities to decompose recalcitrant organic substances was similar among the five treatments. Our results do not support the hypothesis that intercropping affects SOM decomposition through changing the decomposition environment.
One plausible reason for faster decomposition in intercrop is the 8 % higher organic N content in the soil at only marginally greater C content. The decreased C/N ratio is likely to facilitate decomposition (Booth et al. 2005). Indeed, our result showed that lower soil C/N ratio was associated with higher decomposition rate (Fig. 1d). This finding would parallel results from natural grasslands, in which enhanced organic matter decomposition is attributed to higher N availability to the decomposer community in more species rich plots (Hooper et al. 2005; Cong et al. 2015). Remarkably, and in contrast to many biodiversity studies (but see Cong et al. 2014b), reduction in soil C/N ratio and acceleration of SOM decomposition in our intercrop samples did not depend on the presence of a legume (Table 2, Fig. 1c). A reduction of N losses through leaching may drive a reduction in soil C/N ratio and, as a result, increase storage of N in SOM in intercrops (Cong et al. 2014a).
Greater root biomass production in intercrops compared to monocrops might also cause a change in SOM quality (Ghosh et al. 2006; Yang et al. 2010; Li et al. 2011; Cong et al. 2014a). The resulting greater root litter C input in soil would lead to a faster rejuvenation of the SOM pool with more labile SOM (since the start of the experiment, 7 years ago) resulting in a higher relative decomposition rate. Similar to the above-mentioned reduction in C/N ratio, this would extend the accelerating effect of plant species diversity on long-term soil organic C cycling found in nutrient poor natural grasslands (Dijkstra et al. 2005; Reid et al. 2012; Cong et al. 2014b) to nutrient rich agroecosystems.
Intercropping effect on root litter decomposition
Our results demonstrate that intercropping affected root litter decomposition both via litter mixing and decomposition environment effects. But the direction and magnitude of these effects depended on the crop combination and the incubation conditions. In the maize/faba bean combination, we found a positive litter mixing effect: root litter mixtures of maize and faba bean decomposed 2 % faster than expected on the basis of the decomposition of single litters during incubation under laboratory conditions (Fig. 3b) and 6 % faster than expected under field conditions (Fig. 4b). However, we did not find any non-additive effects of mixing roots of maize and wheat, either during incubation in the laboratory, or under field conditions (Fig. 3a, 4a). Our results with these two crop combinations are consistent with the general picture that non-additive litter mixing effects through the substrate diversity pathway occur mainly when the two component species differ in resource quality, such as maize and faba bean root litters which differ in C/N ratio (Table 2). This is in line with results on leaf litters in natural ecosystems (Wardle et al. 1997; Hättenschwiler et al. 2005; Pérez-Harguindeguy et al. 2008; Vos et al. 2013; Handa et al. 2014).
We did not find any effect of decomposition environment on the decomposition of root litter (Fig. 3b, 4b) in the maize/faba bean combination. The effect of decomposition environment was inconsistent for the maize/wheat combination: in the laboratory, the maize/wheat intercrop soil slowed down the decomposition rate of maize or wheat root litter by 32 % compared to the two monocropped soils (Fig. 3a) while this effect was not observed in the field (Fig. 4a). This deceleration of root litter decomposition in the lab may have been due to a lower mineral N availability in soils collected directly from the maize/wheat intercrop field plots than from the monocrop plots (demonstrated for the same long-term experiment by Li et al. 2005). The absence of this effect in the field (Fig. 4a) may have been related to another compensating decomposition environment factor related to intercropping management under field conditions: Intercropped plots (and plots with maize monocrop) received two additional irrigations compared to faba bean and wheat monocrops, to meet maize’s water demand, resulting in a higher soil moisture content in the intercropping treatments than the average moisture content in the corresponding monocrop plots. In the arid climate of the field experiment, this may have increased decomposition of root litter in plots with maize.
This study focused on exploring the effect of intercropping on decomposition of SOM or root litter in spots where root mixing is most intense, as soils were sampled in the middle between adjacent rows of the two crop species. So the effect size for a whole field and over a whole growing season may be smaller than what we measured in the field, also because the incubation temperature in the laboratory experiment (20 °C) was higher than in the average soil temperature in the field.
Concluding remarks
We conclude that intercropping enhanced SOM decomposition through a reduction in SOM recalcitrance. As a consequence, the soil C sequestration potential of intercropping via enhanced biomass input is partly counteracted by faster C cycling. A litter-mixing effect accelerates decomposition of mixtures of maize and faba bean root litter, but not of mixtures of maize and wheat root litter. An intercropping-related decomposition environment effect was only found for the maize/wheat intercropping combination, but the direction of the effect depended on the experimental conditions.
References
Bardgett RD, Shine A (1999) Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands. Soil Biol Biochem 31:317–321
Blagodatskaya E, Kuzyakov Y (2011) Priming effects in relation to soil conditions – mechanisms. In: Glinski J, Horabik J, Lipiec J (eds) Encyclopedia of agrophysics. Springer, Dordrecht, pp 657–667
Booth MS, Stark JM, Rastetter E (2005) Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data. Ecol Monogr 75:139–157
Cadisch G, Giller KE (1997) Driven by nature: plant litter quality and decomposition. CAB International, Wallingford
Chung HG, Zak DR, Reich PB, Ellsworth DS (2007) Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function. Glob Chang Biol 13:980–989
Cong W-F, Hoffland E, Long L, Six J, Sun JH, Bao XG, Zhang FS, Van der Werf W (2014a) Intercropping enhances soil carbon and nitrogen. Glob Chang Biol. doi:10.1111/gcb.12738
Cong W-F, Van Ruijven J, Mommer L, De Deyn GB, Berendse F, Hoffland E (2014b) Plant species richness promotes soil carbon and nitrogen stocks in grasslands without legumes. J Ecol 102:1163–1170
Cong W-F, Van Ruijven J, Van der Werf W, De Deyn GB, Mommer L, Berendse F, Hoffland E (2015) Plant species richness leaves a legacy of enhanced root litter-induced decomposition in soil. Soil Biol Biochem 80:341–348
Dijkstra FA, Hobbie SE, Reich PB, Knops JMH (2005) Divergent effects of elevated CO2, N fertilization, and plant diversity on soil C and N dynamics in a grassland field experiment. Plant Soil 272:41–52
Fornara DA, Tilman D, Hobbie SE (2009) Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J Ecol 97:48–56
Ghosh PK, Manna MC, Bandyopadhyay KK, Ajay TAK, Wanjari RH, Hati KM, Misra AK, Acharya CL, Rao AS (2006) Interspecific interaction and nutrient use in soybean/sorghum intercropping system. Agron J 98:1097–1108
Handa IT, Aerts R, Berendse F, Berg MP, Bruder A, Butenschoen O, Chauvet E, Gessner MO, Jabiol J, Makkonen M, McKie BG, Malmqvist B, Peeters ETHM, Scheu S, Schmid B, Van Ruijven J, Vos VCA, Hättenschwiler S (2014) Consequences of biodiversity loss for litter decomposition across biomes. Nature 509:218–221
Hättenschwiler S, Tiunov A, Scheu S (2005) Biodiversity and litter decomposition in terrestrial ecosystems. Annu Rev Ecol Evol Syst 36:191–218
He Y, Ding N, Shi JC, Wu M, Liao H, Xu JM (2013) Profiling of microbial PLFAs: Implications for interspecific interactions due to intercropping which increase phosphorus uptake in phosphorus limited acidic soils. Soil Biol Biochem 57:625–634
Hector A, Beale AJ, Minns A, Otway SJ, Lawton JH (2000) Consequences of the reduction of plant diversity for litter decomposition: effects through litter quality and microenvironment. Oikos 90:357–371
Hinsinger P, Betencourt E, Bernard L, Brauman A, Plassard C, Shen JB, Tang XY, Zhang FS (2011) P for two, sharing a scarce resource: soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol 156:1078–1086
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Naeem S, Schmid B, Setala H, Symstad AJ, Vandermeer J, Wardle DA (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Knops JMH, Wedin D, Tilman D (2001) Biodiversity and decomposition in experimental grassland ecosystems. Oecologia 126:429–433
Li WX, Li L, Sun JH, Guo TW, Zhang FS, Bao XG, Peng A, Tang C (2005) Effects of intercropping and nitrogen application on nitrate present in the profile of an Orthic Anthrosol in Northwest China. Agric Ecosyst Environ 105:483–491
Li L, Sun JH, Zhang FS, Guo TW, Bao XG, Smith FA, Smith SE (2006) Root distribution and interactions between intercropped species. Oecologia 147:280–290
Li L, Li SM, Sun JH, Zhou LL, Bao XG, Zhang HG, Zhang FS (2007) Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc Natl Acad Sci U S A 104:11192–11196
Li L, Sun JH, Zhang FS (2011) Intercropping with wheat leads to greater root weight density and larger below-ground space of irrigated maize at late growth stages. Soil Sci Plant Nutr 57:61–67
Liebman M, Dyck E (1993) Crop rotation and intercropping strategies for weed management. Ecol Appl 3:92–122
Lithourgidis AS, Dordas CA, Damalas CA, Vlachostergios DN (2011) Annual intercrops: an alternative pathway for sustainable agriculture. Aust J Crop Sci 5:396–410
Midwood AJ, Boutton TW (1998) Soil carbonate decomposition by acid has little effect on δ13C of organic matter. Soil Biol Biochem 30:1301–1307
Muller-Stover D, Hauggaard-Nielsen H, Eriksen J, Ambus P, Johansen A (2012) Microbial biomass, microbial diversity, soil carbon storage, and stability after incubation of soil from grass-clover pastures of different age. Biol Fertil Soils 48:371–383
Nottingham AT, Griffiths H, Chamberlain PM, Stott AW, Tanner EVJ (2009) Soil priming by sugar and leaf-litter substrates: a link to microbial groups. Appl Soil Ecol 42:183–190
Oliveira CA, Scotti MRMML, Purcino HA, Vasconcellos CA, Marriel IE, Sá NMH (2002) Decomposition of Arachis pintoi litter intercropped with forage grass in “Cerrado” soil in the dry and wet seasons. Biol Fertil Soils 36:405–410
Pascault N, Ranjard L, Kaisermann A, Bachar D, Christen R, Terrat S, Mathieu O, Leveque J, Mougel C, Henault C, Lemanceau P, Pean M, Boiry S, Fontaine S, Maron PA (2013) Stimulation of different functional groups of bacteria by various plant residues as a driver of soil priming effect. Ecosystems 16:810–822
Pérez-Harguindeguy N, Blundo CM, Gurvich DE, Díaz S, Cuevas E (2008) More than the sum of its parts? Assessing litter heterogeneity effects on the decomposition of litter mixtures through leaf chemistry. Plant Soil 303:151–159
Reid JP, Adair EC, Hobbie SE, Reich PB (2012) Biodiversity, nitrogen deposition, and CO2 affect grassland soil carbon cycling but not storage. Ecosystems 15:580–590
Rietberg P (2009) Does biochar affect decomposition of organic matter? MSc thesis. Wageningen University, the Netherlands.
Rusinamhodzi L, Murwira HK, Nyamangara J (2009) Effect of cotton-cowpea intercropping on C and N mineralisation patterns of residue mixtures and soil. Aust J Soil Res 47:190–197
Scherer-Lorenzen M (2008) Functional diversity affects decomposition processes in experimental grasslands. Funct Ecol 22:547–555
Schimel J, Cates R, Ruess R (1998) The role of balsam poplar secondary chemicals in controlling soil nutrient dynamics through succession in the Alaskan taiga. Biogeochemistry 42:221–234
Seastedt T (1984) The role of microarthropods in decomposition and mineralization processes. Annu Rev Entomol 29:25–46
Song YN, Zhang FS, Marschner P, Fan FL, Gao HM, Bao XG, Sun JH, Li L (2007) Effect of intercropping on crop yield and chemical and microbiological properties in rhizosphere of wheat (Triticum aestivum L.), maize (Zea mays L.), and faba bean (Vicia faba L.). Biol Fertil Soils 43:565–574
Spehn EM, Joshi J, Schmid B, Alphei J, Korner C (2000) Plant diversity effects on soil heterotrophic activity in experimental grassland ecosystems. Plant Soil 224:217–230
Spehn EM, Hector A, Joshi J, Scherer-Lorenzen M, Schmid B, Bazeley-White E, Beierkuhnlein C, Caldeira MC, Diemer M, Dimitrakopoulos PG, Finn JA, Freitas H, Giller PS, Good J, Harris R, Hogberg P, Huss-Danell K, Jumpponen A, Koricheva J, Leadley PW, Loreau M, Minns A, Mulder CPH, O’Donovan G, Otway SJ, Palmborg C, Pereira JS, Pfisterer AB, Prinz A, Read DJ, Schulze ED, Siamantziouras ASD, Terry AC, Troumbis AY, Woodward FI, Yachi S, Lawton JH (2005) Ecosystem effects of biodiversity manipulations in European grasslands. Ecol Monogr 75:37–63
Stephan A, Meyer AH, Schmid B (2000) Plant diversity affects culturable soil bacteria in experimental grassland communities. J Ecol 88:988–998
Trenbath BR (1993) Intercropping for the management of pests and diseases. Fields Crops Res 34:381–405
Vachon K, Oelbermann M (2011) Crop residue input and decomposition in a temperate maize-soybean intercrop system. Soil Sci 176:157–163
Vogel A, Eisenhauer N, Weigelt A, Scherer-Lorenzen MS (2013) Plant diversity does not buffer drought effects on early-stage litter mass loss rates and microbial properties. Glob Chang Biol 19:2795–2803
Vos VCA, Van Ruijven J, Berg MP, Peeters ETHM, Berendse F (2013) Leaf litter quality drives litter mixing effects through complementary resource use among detrivores. Oecologia 173:269–280
Wardle DA, Bonner KI, Nicholson KS (1997) Biodiversity and plant litter: experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos 79:247–258
Willey RW (1979) Intercropping – its importance and research needs. Part I, competition and yield advantages. Fields Crops Abstr 32:1–10
Yang HS, Janssen BH (2000) A mono-component model of carbon mineralization with a dynamic rate constant. Eur J Soil Sci 51:517–529
Yang CH, Chai Q, Huang GB (2010) Root distribution and yield responses of wheat/maize intercropping to alternate irrigation in the arid areas of northwest China. Plant Soil Environ 56:253–262
Zak DR, Holmes WE, White DC, Peacock AD, Tilman D (2003) Plant diversity, soil microbial communities, and ecosystem function: are there any links? Ecology 84:2042–2050
Zhu YY, Chen HR, Fan JH, Wang YY, Li Y, Chen JB, Fan JX, Yang SS, Hu LP, Leung H, Mew TW, Teng PS, Wang ZH, Mundt CC (2000) Genetic diversity and disease control in rice. Nature 406:718–722
Acknowledgments
We are grateful to Jaap Nelemans, Willeke van Tintelen, Liesje Mommer, Fusuo Zhang, Jianhao Sun, Xingguo Bao and Zhigang Wang for technical assistance and suggestions, and to Jasper van Ruijven for comments on a previous version of the manuscript.
Compliance with Ethical Standards
This research was financially supported by grants from the Natural Science Foundation of China: The Key Sino-Dutch Joint Research Project (Project No. 31210103906), the Innovative Group Grant of the Natural Science Foundation of China (Project No. 31121062) and the National Basic Research Program of China (Project No. 2011CB100405). W-F.C. had a scholarship for a PhD at Wageningen University from the China Scholarship Council.
Conflict of Interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Per Ambus.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Cong, WF., Hoffland, E., Li, L. et al. Intercropping affects the rate of decomposition of soil organic matter and root litter. Plant Soil 391, 399–411 (2015). https://doi.org/10.1007/s11104-015-2433-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2433-5