
Abstract
Stripe rust, caused by Puccinia striiformis f. sp. tritici (Pst), is one of the most important diseases on wheat in China. To assess resistance in wheat cultivars and breeding lines in China, 330 leading cultivars and 164 advanced breeding lines were evaluated with stripe rust. In the greenhouse tests, seedlings of the entries were inoculated separately with several Pst pathotypes. In the field tests, the entries were evaluated for stripe rust resistance in Yangling, Shaanxi Province artificially inoculated and in Tianshui, Gansu Province under natural infection of Pst. The oversummering/wintering and spring epidemic zones of resistance genes were postulated using molecular markers for Yr5, Yr9, Yr10, Yr15, Yr17, Yr18, and Yr26, in combination with resistance spectra. Out of the 494 wheat entries, 16 (3.24 %) entries had all-stage resistance (ASR) in all race tests, 99 (20.04 %) had adult-plant resistance (APR), 28 (5.67 %) were considered to have slow-rusting (SR), and 351 (71.05 %) were susceptible to one or more races in both seedling and adult-plant stages. Advanced breeding lines had a higher percentage (37.2 %) of resistant entries (The sum of ASR, APR and SR) than leading cultivars (24.85 %). Among the epidemic regions, southern Gansu had a higher percentage of resistant entries than any other regions. Based on stripe rust reactions and molecular markers, two cultivars were found to possibly have Yr5 while no entries have Yr10 or Yr15. Resistance genes Yr9, Yr17, Yr18, and Yr26 were found in 134 (29.4 %), 45 (9.1 %), 10 (2 %), and 15 (3 %) entries, respectively.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Stripe (yellow) rust, caused by Puccinia striiformis Westend. f. sp. tritici Erikss. (Pst), is the most destructive disease in all winter wheat growing regions in the northwestern, southwestern, and northern China, as well as in the spring wheat areas in the northwestern China (Li and Zeng 2002; Wan et al. 2004; Chen et al. 2009). The disease can cause up to 100 % crop loss, though commonly in the range of 10–70 % (Chen 2005). Severe epidemics in 1950, 1964, 1990, and 2002 in China resulted in yield losses of 6.0, 3.2, 1.8, and 1.3 million tons, respectively (Wan et al. 2007; Chen et al. 2009). In recent years, major yield losses were prevented by widespread and timely applications of fungicides based on accurate monitoring and prediction of disease epidemics (Kang et al. 2010). Growing resistant cultivars is considered the most effective, low-cost, and environmentally safe approach to control stripe rust (Roebbelen and Sharp 1978; Line and Chen 1995). The use of resistant cultivars substantially depends upon the availability and careful deployment of resistance genes. However, a prerequisite for use of stripe rust resistance genes (abbreviate to Yr) in breeding is detailed knowledge on which Yr genes present in released wheat cultivars.
To date, 53 Yr genes have been officially named (McIntosh et al. 2012); most confer race-specific all-stage resistance, and some confer adult-plant or high-temperature adult-plant (HTAP) resistance (Chen 2005; Lin and Chen 2007, 2009). However, the rapid development of new Pst races often circumvents resistance in cultivars (Line and Qayoum 1992). Races of Pst in China have been intensively studied (Li and Zeng 2002; Wan et al. 2004; Chen et al. 2009). Race CYR32 detected in 1994 and CYR33 found in 1997 have become the most predominant races (Chen et al. 2009), which have rendered wheat cultivars with Yr9, Yr3b, Yr4b and YrSu susceptible. Cultivars with one or more of these genes have been widely grown, accounting for 90 % of total winter wheat area in the entire country (Kang et al. 2010). Survey data from 1999 to 2009 show that on average about 4 million hectares can be affected by stripe rust annually (Kang et al. 2010). After these genes becoming ineffective to the predominant races, cultivars and lines with Yr24 (=Yr26) were widely grown in Sichuan and Gansu provinces (Li et al. 2006). A new race, CH42, was identified by Liu et al. (2010) on previously resistant wheat cultivar Chuanmai 42, which has Yr24, during the 2008–2009 crop season in Sichuan. The race is virulent on wheat lines with Yr10 that is effective against all previously identified Chinese Pst races. Based on recent evaluations in China, genes Yr5, Yr15, Yr18, Yr36, Yr39 and Yr41, as well as Yr12, Yr13, Yr14, Yr16, and some temporarily designated genes are still effective and could be used in breeding programs, while Yr1, Yr2, Yr3, Yr4, Yr6, Yr7, Yr8, Yr9, Yr10, Yr17, Yr20, Yr21, Yr22, Yr24/Yr26, Yr25, Yr27 and Yr29 are ineffective to the currently prevalent or new races (Wan et al. 2004, 2007). Wheat stripe rust has remained at high levels for the last decade and continues posing a threat to wheat production across the entire country. Based on epidemiological considerations, it has been suggested that different resistance genes for controlling wheat stripe rust should be deployed in the defined over-summering, over-wintering, and eastern spring epidemic regions (Fig. 1, Li and Zeng 2002; Wan et al. 2007). To use this approach, it is important to know what genes are already used in cultivars and breeding programs in different epidemiological regions.

Northwest-Central-Middle and Low Reaches of Yangtze River stripe rust epidemic regions in China. 1 Stripe rust core oversummering region in southern Gansu, including Tianshui and Wudu city. 2 overwintering regions, including Sichuan Basin, central and southern Shaanxi, eastern Gansu, northwestern Hubei and southern Henan. 3 spring epidemic region, including Shanxi, Hebei, Shandong, Anhui, Jiangsu, Beijing, most of Henan and a fraction of Zhejiang and Inner Mongolia
In order to know stripe rust resistance in China, wheat cultivars and lines were collected from various epidemic regions and tested with five selected Pst races in a greenhouse and in fields in Shaanxi and Gansu under artificial inoculation and natural infection, respectively. Molecular markers for resistance genes Yr9, Yr17 and Yr18 that have been intensively used in wheat breeding programs both in China and the world (Singh et al. 2000; Zhou et al. 2004; Tanguy et al. 2005); Yr5 and Yr15 that are still effective against the Chinese Pst population (Sharma-Poudyal et al. 2013); and Yr10 and Yr24/Yr26 that have recently become ineffective in China (Liu et al. 2010) were used to determine their presence in Chinese wheat cultivars and breeding lines in combination with their phenotype data of the greenhouse and field tests.
Materials and methods
Plant materials
A total of 494 wheat entries, including 330 leading cultivars and 164 advanced breeding lines, were collected from 13 provinces (Fig. 1. Gansu, Shaanxi, Sichuan, Ningxia, Hubei, Henan, Shanxi, Hebei, Shandong, Anhui, Jiangsu, Beijing and Inner Mongolia) covering most important wheat produced regions in China from 2003 to 2010. In addition, 8 wheat lines with different Yr genes were used in the seedling tests in a greenhouse (Table 1). The cultivar ‘Mingxian 169′ was used as a universal susceptible control.
Pst isolates
Five Chinese Pst races (CYR23, CYR29, CYR32, CYR33 and CH42, Table 1) were selected to test the cultivars and lines for resistance in seedling stage in a greenhouse, and nine races (CYR23, CYR25, CYR29, CYR31, CYR32, CYR33, CH42, Su11-4 and Su11-7) were used in the field experiments. CYR23, CYR25, CYR29, CYR31, and CYR32 were from our collection in the Institute of Plant Pathology of Northwest A&F University, while CYR33, Su11-4 and Su11-7 were kindly provided by Prof. Xiangchun Zhou, Plant Protection Institute, Gansu Academy of Agricultural Sciences. CH42 was provided by Prof. Yunliang Peng, Plant Protection Institute of Sichuan Academy of Agricultural Sciences. Urediniospores of all races were increased from a single urediniospore and confirmed by testing on the set of wheat cultivars used to differentiate Chinese Pst races (Chen et al. 2009).
Seedling tests
Seedling tests were conducted two replicates in a greenhouse. About 10 seeds of the 494 entries were inoculated with urediniospores (mixed with talc at a ratio of approximately 1:20) at two-leaf stage, incubated at 10 °C for 24 h in dark and grown at 10–18 °C with 16 h light (About 10,000 lx of fluorescent light)/8 h dark. Minxian 169 was used as a susceptible check. Infection types were recorded three times at a three-day interval starting about 15 days after inoculation, according to the 0–9 scale (0–3 considered is resistant, 4–6 considered is intermediate, 7–9 considered is susceptible) described by Line and Qayoum (1992).
Field tests using artificial inoculation
Field tests, according to complete randomized block design with two replicates, were conducted during the 2009, 2010 and 2011 cropping seasons at the experimental farm of Northwest A&F University, Yangling, Shaanxi. Seeds of the 494 lines were planted in two rows after every 20 entries as a susceptible cultivar check. Susceptible variety Mingxian 169 was planted at both ends of the nursery as a stripe rust spreader. During late February to early March, sporulating seedlings of Mingxian 169 that had been inoculated with a mixture of urediniospores of the eight selected races in the greenhouse were transplanted throughout in the experimental field. The ratio of the urediniospores among the races forming each-year inoculation was determined according to their frequencies in the previous years. The percentages of the eight races used to make the urediniospore mixtures for the 2009–2010 test were CYR33 (30 %), CYR32 (30 %), Su11-7 (10 %), Su11-4 (10 %), CYR31 (5 %), CYR29 (5 %), CYR25 (5 %) and CYR23 (5 %). Because of the increasing frequency of CH42, it was added to the mixture in the 2011 test. The percentages of the nine races in the 2011 experiment were CYR33 (25 %), CYR32 (25 %), CH42 (10 %), Su11-7 (10 %), Su11-4 (10 %), CYR31 (5 %), CYR29 (5 %), CYR25 (5 %) and CYR23 (5 %). The nurseries were managed using the common practices in the region, except that the field was irrigated twice at the critical period of infection to ensure the adequate level of disease. Infection type and severity of top three leaves were recorded twice according to Li (2002) and Line and Qayoum (1992) in the middle and late May when the plants reached boot and milk stages, and the disease severity (DS) on susceptible check reached 30 and 80 %, respectively.
Field tests under natural infection
All entries were planted in a field near Wangjia Village (latitude 34°27′N, longitude 105°56’E, altitude 1,697 m), Tianshui, Gansu Province, about 300 km away from Yangling, Shaanxi during the 2008–2012 cropping seasons. The experimental design was the same as described above for the artificial inoculation experiments. The nursery was managed using the common practices in the region and without artificial inoculation. Seeds were sowed in middle September. Infection types of seedlings were recorded in late November and early December when the plants were at tillering stage and both infection type and severity data were recorded in late May and early June when the crops were at boot and milk stages, respectively, and disease severity on susceptible check reached to 40 and 80 %, respectively.
DNA extraction
Plants were grown in a greenhouse up to the two-leaf stage without light to obtain etiolated leaf tissue for efficiently isolating genomic DNA. A total of 100 mg of leaf tissue was collected from each line, immediately frozen in liquid nitrogen. Genomic DNA was extracted from leaves as described by Weining et al. (1994).
PCR amplification and electrophoresis
Primers of the markers for identifying specific Yr genes were listed in Table 2 and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). PCR amplification and electrophoresis were carried out as described for each gene in the reference. The PCR reactions of different primer pairs were briefly described in Table 3.PCR reactions were performed in a Bio-Rad ® S1000 Thermal Cycler With 96-Well Fast Reaction Module. Mg-free 10× PCR buffer, MgCl2 and Taq DNA polymerase were bought from Fermentas (Canada); dNTPs was bought from Roche (German). For the primer pairs STS7/8 and URIC/LN2, after amplification, PCR product was digested with DpnII (New England Biolabs, USA) as the described by Chen et al. (2003) with slight modification. Briefly, the reaction mixture for enzymatic digestion contained 10 μl of PCR product, 1U of restriction enzyme DpnII (New England Biolabs, Beverly, MA) and 2 μl of 10× buffer for DpnII (New England Biolabs). Samples were incubated at 37 °C for 4 h and the digested products were separated in an agarose gel. For the agarose gel, after amplification 3 μl of 6× DNA loading dye (Fermentas) was added to the PCR product. Then 8 μl of the PCR product and loading buffer mixture for each sample was loaded for electrophoresis in the particular concentration of the agarose gel in 1× TAE buffer under the voltage of 5 V/cm for about 30 min followed by staining with 0.5 μg/mL ethidium bromide for 20 min and visualized with UV light using the gel imaging system (Molecular Imager® Gel Doc™ XR+ System, Bio-Rad, USA). For polyacrylamide gel electrophoresis (PAGE), after amplification 3 μl of formamide loading buffer was added to the PCR product. After 5 min denaturation at 94 °C and immediately ice-bath for 5 min, about 4 μl of the PCR product and loading buffer mixture for each sample was loaded for electrophoresis in a 5 % denaturing polyacrylamide gel and the gel was silver stained as described by An et al. (2009).
Data analysis
Entries with IT ≤6 both in the greenhouse and Tianshui seedling tests were treated as all-stage resistant (ASR); those that were susceptible (ITs 7-9) to at least one race in the seedling stage but resistant (IT ≤6, severity ≤30 %) to all tests in the adult-plant stage of the field tests were considered having adult plant resistance (APR), while those that had a high infection type (ITs 7-8) but low severity (≤ 30 %) in the field tests at adult-plant stage were considered having slow-rusting (SR). Entries with ITs 7-9 in all tests and more than 30 % severity were considered susceptible (S).
Results
Stripe rust resistance evaluation
Supplement Table 1 shows infection types and severity of the 494 entries in all greenhouse and field tests. Based on the seedling and field reactions, the 494 entries could be classified into four groups. The first group consisted of 4 advanced lines and 12 cultivars that were resistant to stripe rust in all tests. These lines and cultivars have effective all-stage resistance in China. Whether they have APR and different from each other could not be determined with the data from this study. Ninety-nine (20 %) lines were resistant in all field tests and susceptible to at least one test in the seedling stage showed APR. Entries in this group have great effectiveness to control stripe rust for its durable and non-race-specific resistance to stripe rust. The third group consisted of 28 (5.7 %) lines that have high infection type (ITs 7-8) but low severity (≤30 %) in the field tests at adult-plant stage. This group was considered having slow-rusting which is expressed quantitatively, so generally considered to confer durable resistance. These effects conferred by the genes in this group typically too weak to be adequate in stripe rust control, thereby requiring combination with multiple genes to provide adequate protection against stripe rust. The last group consisted of 351 (71 %) entries that were susceptible to at least one of the races in both seedling and adult-plant stage (Table 4). This group should not be used in wheat production. If used in production for any agronomic reasons, fungicide application is likely needed. Of the last group, 62 entries (12.55 % of the total 494 entries) were susceptible in all tests, indicating no efficient gene resistance to any of the races used in this study.
Comparison of resistant entries between leading cultivars and advanced lines
The number and frequency of ASR, APR, SR and S entries of leading cultivars were 12 (3.64 %), 48 (14.55 %), 22 (6.66 %) and 248 (75.15 %), respectively. The number and frequency of ASR, APR, SR and S entries of advanced breeding lines were 4 (2.44 %), 51 (31.10 %), 6 (3.66 %) and 103 (62.80 %), respectively (Table 4). The higher percentage of APR and lower percentage of ASR in the advanced breeding lines than those of leading cultivars indicate that breeding programs have taken more effort to use non-race-specific durable resistance for developing stripe rust resistant cultivars.
Stripe rust resistance in different epidemic regions
The data shown in Fig. 2 suggested great variations in resistance among the three epidemic regions. Out of the 28 entries in the oversummering region in southern Gansu, 4 (14.29 %) had ASR, 9 (32.14 %) had APR and 6 (21.43 %) had SR. Of the 264 entries from the overwintering regions, 12(4.62 %), 64 (24.62 %), and 22 (8.46 %) had ASR, APR, and SR, respectively. Of the 226 entries from the spring epidemic region, only 1 (0.44 %), 30 (13.27 %) and 2 (0.88 %) were found to have ASR, APR and SR, respectively. The percent of resistance in the oversummering region was the highest, indicating that the breeding programs in Gansu have taken more effort in developing wheat cultivars with resistance to stripe rust.

Percentage of wheat entries in different groups of reactions to stripe rust in three epidemic regions. #Region 1 oversummering region in Southern Gansu, Region 2 overwintering regions and Region 3 eastern spring epidemic region. (##) Among the 494 entries of wheat, 20 cultivars were exist in different regions (8 cultivars were both exist in region 1 and 2, 12 cultivars were both exist in region 2 and 3), so the number of wheat entries counted for different regions was 514. * ASR all-stage resistance (Entries with IT ≤6 both in the greenhouse and Tianshui seedling tests), APR adult-plant resistance [Entries that were susceptible (ITs 7-9) to at least one race in the seedling stage but resistant (IT ≤ 6, severity ≤30 %) to all tests in the adult-plant stage of the field tests], SR slow resistance [Entries that had a high infection type (ITs 7-8) but low severity (≤30 %) in the field tests at adult-plant stage] and S susceptible (Entries with ITs 7-9 in all tests and more than 30 % severity)
Identification of Yr genes using molecular markers
Genes Yr5 and Yr15 confer race-specific all-stage resistance but still effective against all races in China. Similar to the lines with Yr5 and Yr15, 10 cultivars (Lantian 22, Lantian 18, Mianmai 41, Shaanmai 139, Zhongliang 95-85, Hanmai 13, Chuannong 11, Chuanmai 52, Hanmai 5 and Lantian 21) were resistant in the seedling tests to all tested races (CYR23, CYR29, CYR32, CYR33 and CH42) and also in all field tests. Genes Yr10 and Yr24/Yr26 confer race-specific all-stage resistance and still effective against CYR32 and CYR33 but have become ineffective (IT = 7 or 8) against CH42. Therefore, 55 cultivars that were resistant to both CYR32 and CYR33 were tested with markers for Yr10, Yr24 or Yr26. Genes Yr9, Yr17 and Yr18 that were ineffective against CYR32 and CYR33 in the seedling tests were tested for presence/absence in all entries with their molecular markers. The presence and absence of molecular markers for Yr5, Yr9, Yr10, Yr15, Yr17, Yr18 and Yr26 in the tested wheat entries are shown in Supplement Table 1.
Identification of Yr5
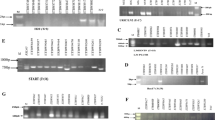
Yr5 was originally derived from Triticum spelta album (TSA), an hexaploid spelta wheat. Two sequence-tagged site (STS) markers S19M93-100 F/R (Smith et al. 2007) and STS7/8 which were then developed into cleaved amplified polymorphic sequence (CAPS) markers on the basis of a DpnII restriction site (Chen et al. 2003) were used to detect Yr5. The S19M93-100 F/R primers amplified a single 100 bp band in AvSYr5NIL (Yr5); and the band was absent in wheat lines without Yr5, (Fig. 3a). The STS7/8 primers amplified a 478-bp band in AvSYr5NIL and a 472-bp band in Avocet S without Yr5. The difference could not be visualized in a 1.5 % agarose gel. After digestion of the STS7/8 PCR products with the DpnII restriction enzyme resulted in five restriction fragments in the Avocet S*6/Yr5 (308, 77, 63, 20, and 12 bp), and six restriction fragments (181, 122, 77, 63, 20, and 12 bp) in AVS and other non-Yr5 genotypes. The differences between the 308-bp fragment in the resistant lines and the 181-bp fragment in the susceptible lines were easy to visualize in agarose gels (Fig. 3b). After evaluated by these two primers, cultivars Lantian 22 and Zhongliang 95-85 were postulated to carry Yr5.

Electrophoretogram of different primers. a, b identification of Yr5, AVS/6*Yr5 is the NIL known to carry the Yr5 gene, AVS is the reference line known didn’t carry the the Yr5 gene. c, d identification of Yr9, AVS/6*Yr9 is the NIL known to carry the Yr9 gene, AVS is the reference line known didn’t carry the the Yr9 gene. e, f identification of Yr17, AVS/6*Yr17 is the NIL known to carry the Yr17 gene, AVS is the reference line known didn’t carry the the Yr17 gene. g, h identification of Yr18, Jupiteco R and Chinese Spring are the reference lines known to carry the Yr18 gene, AVS is the reference line known didn’t carry the the Yr18 gene. i, j identification of Yr26, 92R137 and R55 are the reference lines known to carry the Yr26 gene, AVS is the reference line known didn’t carry the the Yr26 gene. Note M1: 250 bp DNA Ladder-II (Generay Biotech (Shanghai) Co., Ltd.), bands: 2000, 1000, 750, 500, 250, 100 bp. M2: pUC19 DNA/MspI (HpaII) Marker (Frementas life sciences), bands: 501, 489, 404, 331, 242, 190, 147, 111, 110, 67, 34, 26 bp. M3: 50 bp Ladder DNA Marker (Biomed, Beijing), bands: 500, 400, 300, 250, 200, 150, 100, 50 bp
Identification of Yr9
Yr9 is located on the short arm of rye chromosome 1R, which was transferred to wheat through chromosomal translocation of 1B/1R. The primer pairs AF1/AF4 (Francis et al. 1995) and H20 (Liu et al. 2008) were chosen to detect Yr9. For primers AF1/AF4, clearly a single product of size 1.5 kb was produced whenever the entries have the rye chromatin (Fig. 3c). Primer pair H20 F/R amplified a 1,598-bp fragment (Fig. 3d). Based on the PCR tests with these two primer pairs, 127 (25.71 %) entries were found to have Yr9. While among these 127 entries, 120 showed resistance to CYR23 which similar to the near-isogenic line of Yr9, 4 (Xinmai 208, Shandong 664, Xiaoyan 389 and Zhoumai 9) showed susceptible to CYR23 and 3 had no data. Among 120 entries, 47 were resistant to more races than the near-isogenic line of Yr9, indicating that these lines might carry other seedling resistance genes.
Identification of Yr10
Yr10 was originally found in wheat line PI 178383 and located in the short arm of chromosome 1B. Singh et al. (2009) designed two primer pairs (Yr10 F/R and Yr10 F1/R1) based on the Yr10 sequence and produced markers completely linked to Yr10. Only Moro, AvSYr10NIL and PI 178383 had the Yr10 marker and none of the Chinese wheat entries had the Yr10 markers (the figure is not shown).
Identification of Yr15
Yr15, derived from Triticum dicoccoides, was located in chromosome 1BS (McIntosh et al. 1996). Murphy et al. (2009) identified that Xbarc8 and Xgwm413 appeared to be completely linked with Yr15 in a population of 136 BC7F4 lines. These markers were not found in the 10 ASR entries (the figure not shown), suggesting that none of the 494 Chinese wheat entries have Yr15.
Identification of Yr17
Rust resistance genes Yr17, Lr37 (a leaf rust resistance gene) and Sr38 (a stem rust resistance gene) are located within a segment of Triticum ventricosum chromosome 2NS translocated to the short arm of wheat chromosome 2A (Helguera et al. 2003). The primers URIC and LN2 amplified two fragments of 285 bp (N-allele) and 275 bp (A-allele). Digestion of the PCR products with restriction enzyme DpnII facilitated the differentiation between these two bands. The undigested 285-bp band corresponded to the N genome PCR product and the 166- and 109-bp fragments corresponded to the A genome digested PCR product (Fig. 3e). The VENTRIUP-LN2 primers were 2NS specific, and the 259-bp PCR amplification product was observed only in plants carrying the 2NS translocation (Fig. 3f). The marker identified 45 (9.11 %) entries to have Yr17. Among these 45 entries, 28 showed resistance to CYR23 and CH43, similar to the near-isogenic line of Yr17 (resistance to both CYR23 and CH42, susceptible to all the three races CYR29, CYR32, CYR33), 8 were susceptible to either CYR23 or CH42 and 8 were susceptible to both CYR23 and CH42 which seemed to be false positive with the molecular marker, and 1 had no integrated data. Among 28 entries, 22 showed broader resistance spectra (5 resistance to CYR29, 4 resistance to CYR33, 2 resistance to both CYR29 and CYR32, 6 resistance to both CYR29 and CYR32, 2 resistance to both CYR32 and CYR33, 3 resistance to all the three races CYR29, CYR32 and CYR33,) than the near-isogenic line of Yr17, suggesting that they carry additional all-stage resistance genes.
Identification of Yr18
The Yr18/Lr34/Pm38 locus confers partial and durable APR against leaf rust, stripe rust and powdery mildew. Lagudah et al. (2009) developed two primer pairs for tracking both the +Y18 and the –Yr18 alleles. The first multiplex reaction was developed based on primer combinations L34DINT9F/L34PLUSR and csLV34 F/R, which resulted in the amplification of two bands (517 bp and 150 bp) indicating the presence of Yr18 and one band (229 bp) indicates absence of Yr18 (Fig. 3 g). The second multiplex reaction was developed based on primer combinations L34SPF/L34DINT13R2 and L34DINT9F/L34MINUSR. This reaction amplified two bands of different sizes that can be easily resolved in a 1 % agarose gel: a 751-bp fragment specific for the +Yr18 allele and a 523-bp fragment specific for the –Yr18 allele (Fig. 3h). Of the 494 entries, 10 (2.02 %) were found to carry Yr18, including cultivars Jing 3310, Zhongliang 17, Strampelli, Q042, Jinan 9, Jing 5642, Lunxuan 715, Lantian 11, Lantian 12 and Xinong 928. The near-isogenic line of Yr18 was susceptible to all the races in the seedling stage, whereas Strampelli, Lunxuan 715, Lantian 11 and Lantian 12 were resistant to more races, indicating that these four entries carry additional all-stage resistance genes. In the field test, near-isogenic line of Yr18 showed a lower disease severity than other susceptible entries at the beginning, but at the third note-taking the disease severity abruptly increased to 60 % and more. The observation was consistent with the previous reports that Yr18 confers slow-rusting (Lagudah et al. (2009).
Identification of Yr26
Yr26, carried by Triticum aestivum-Haynaldia villosa translocation lines, is located on chromosome 1B. Using primer WE173 developed by Wang et al. (2008), a 451-bp band in 92R137 and R55 (Yr26 gene donors) and a 730 bp band in Avocet S without Yr26 were detected (Fig. 3i). The WE210 primer pair amplified a 500-bp band in Avocet S, but not in 92R137 and R55 (Fig. 3j). Amplification of the 55 entries that were resistant to both CYR32 and CYR33 but susceptible to CH42 identified 15 (27.27 %) cultivars possessing Yr26.
Discussion
Stripe rust resistance in Chinese wheat cultivars and breeding lines
The epidemic regions in the Northwest, the North and -the Middle and Low Reaches of Yangtze River cover 13 provinces, autonomous regions, and municipalities, and produces 80 % of the total wheat grain in China. The majority (71 %) of the 494 wheat entries found to be susceptible indicates a great need for developing stripe rust resistant cultivars. Only 23 entries (4.7 %) were resistant against all five major races (CYR23, CYR29, CYR32, CYR33 and CH42) tested in the greenhouse. The number of entries with ASR decreased to 16 when the seedling data in the Tianshui field experiments were used. These results indicated that 7 of the 23 entries with all-stage resistance were no longer resistant to the natural races population in the oversummering zone of Tianhui. There were 105 entries identified to have APR style when compared their reactions in the field experiments in Yangling inoculated with 8 or 9 races and the susceptible reactions in the greenhouse seedling tests with five races. The number of entries with all-stage resistance was reduced to 96 when the seedling and adult-plant reactions in the field experiments at Tianshui were compared. The nine entries becoming susceptible at Tianshui may have race-specific adult-plant resistance that might not effective against races in Tianshui or have a low level of HTAP resistance which might not fully express under the relatively low temperatures.
The proportion of resistant entries of advanced lines (37.2 %) was more than that of leading cultivars (24.85 %) and the proportion of APR entries in the advanced lines (31.1 %) was more than twice of that in leading cultivars (14.55 %). The higher percentage of resistant entries in advanced lines than leading cultivars indicates that the breeding programs have care more about stripe rust resistance and more preferred to use non-race specific resistance. Currently, CYR32 and CYR33 are the predominant races in China. Therefore, the fact that the entries which were resistant to these two races (such as 92R and Guinong series) became susceptible to the new race CH42 has led to a low level of ASR in advanced lines. The small number of resistant lines and cultivars limits the rational deployment of cultivars with different resistance genes.
The eastern and southern Gansu province and the northwestern Sichuan province are key regions for stripe rust to complete its year-round disease cycle. Southern Gansu including the Tianshui and Wudu regions is the most important core oversummering region and is one of the most important selection zone of new races (Zeng and Luo 2006; Li and Zeng 2002). Four cultivars in this region have ASR to the current Pst population; two of them with Yr5 and two with unknown genes. Two of nine APR cultivars carry Yr17 gene but do not have Yr18. In this region at the seedling stage, 85.71 % (including APR, SR and susceptibility entries) cultivars were susceptible to stripe rust. Growing these cultivars could produce a large quantity of inoculum that might spread to other regions. For its core status in the stripe rust epidemic cycle in China, major yield losses should be prevented by widespread and timely applications of fungicides based on accurate monitoring and prediction of disease epidemics.
The regions of Sichuan Basin, central and southern Shaanxi, eastern Gansu, northwestern Hubei and southern Henan are stripe rust overwintering regions. In these regions, the pathogen can overwinter and even reproduce during the winter time in some warm areas. The Sichuan Basin overwintering region, including Chengdu, Mianyang, Guangyuan, Deyang, Suining, Ziyang, Neijiang, Meishan, Leshan, Yaan, Nanchong, Bazhong and Guangan is the most important overwinter region. The regions of eastern Gansu, southern Ningxia, central Shaanxi, and southern Shanxi are the bridge regions of the pathogen spreading from southern Gansu and the Sichuan basin to the spring epidemic region to the east. In this region, cultivars with highly effective resistance in the seedling stage as well as the adult-plant stage should be used to reduce the inoculum from the oversummering region in autumn. Actually only 3 (2.21 %) cultivars have effective ASR in this region. By inoculating 92 regional cultivars from the Gansu, Sichuan, Henan and Hebei provinces with mixture of CYR33, CYR32, CYR31, CYR30, CYR29, CYR17, Su-1 and CH42 in the field test, Wang et al. (2013) also found the resistance of wheat cultivars to stripe rust in Gansu and Sichuan was much stronger than Henan and Hebei. The region of southern Shaanxi-northwestern Hubei-southern Henan, including Ankang, Hanzhong, Xiangfan, Jingmen, Shiyan, Xinyang and Nanyang, is the pathway of stripe rust spreading from Gansu, Shaanxi and Sichuan to the large wheat production area of Henan, Hebei and Shandong provinces. To control the epidemic of stripe rust, effective ASR genes which different from the ASR genes that used in the oversummer region should be used in the overwinter region.
The spring epidemic region is the most important wheat production region in China (Zhuang 2003). The proportion of resistant cultivars is the lowest as193 (85.3 %) cultivars were found to be susceptible. Because rust inoculum is from the overwinter and oversummer regions and the disease usually develops in the late stage, cultivars with APR, HTAP, or slow rusting should be recommended to plant in this region.
It is more desirable to use effective ASR genes combined with durable APR genes in all of these regions. Using resistant cultivars with multiple or different resistance genes is capable of reducing the build-up of inoculum. If varieties become susceptible, they should be withdrawn from production as quickly as possible. For any agronomic reasons, fungicides should be applied on time to control stripe rust when susceptible cultivars are grown under disease favorable conditions.
Marker-assisted selection
Yr5 and Yr15 are still effective against stripe rust in China. Only two entries have Yr5. Yr9 became ineffective when race CYR29 appeared in the field in 1983. Yr10 and Yr26 are effective to the predominant races CYR32 and CYR33 but susceptible to new race CH42. Yr9 was transferred to wheat through chromosomal translocation of 1B/1R and linkage with Lr26, Sr31 and Pm8 resistance genes and the cultivars which carry Yr9 have a good productivity and adaptability (Zhou et al. 2004). Yr9 was widely used in breeding programs in 1970s to 1980s in China (Wan et al. 2004). In the present study, 127 of the 494 entries were found to have Yr9. When the molecular markers were used to detect Yr10 and Yr15, the targeted bands were only observed in positive controls and none of the Chinese wheat entries have these genes. Zhuang (2003) thought that during the history of the wheat breeding in China there were no evidence of using Yr10 and Yr15. Yr26 was found in several T. aestivum-H. villosa 6VS/6AL translocation lines (92R89, 92R90, 92R137, 92R149 and 92R178), developed by the Cytogenetics Institute, Nanjing Agricultural University. The marker data for the presence of Yr26 in the identified entries were in consistence with their pedigrees. Of the 55 entries with ASR to the four previously and currently predominant races (CYR23, CYR29, CYR32 and CYR33), 15 varieties have Yr26 and 10 of them were from the Sichuan Basin, suggesting that the gene have been overused. When tested with the newly emerging race, CH42 that is virulent to Yr26, only three of the 15 were still resistant, suggesting that Yr26 cannot be used singly.
The Yr17, Lr37 and Sr38 rust resistance genes, which confer resistance in wheat against stripe rust, leaf rust, and stem rust, respectively, have been used in breeding programs in different parts of the world. The combination of an N specific marker and an A specific CAPS marker developed by Helguera et al. (2003) provides more flexibility to manipulate these genes in breeding programs than previously reported PCR markers. Based on the presence of these markers, 45 out of the 494 entries were found to have Yr17.
Five allele-specific markers (cssfr1–cssfr5) were developed by Lagudah (2009) based on a 3 bp deletion in exon 11 of the Yr18-gene. Cssfr3 and cssfr5 are probably the best primer combinations for marker assisted selection because it allows detection by simple agarose gel electrophoresis. Ten entries were found to have Yr18 using these primers. Yang (2008)identified Lr34/Yr18 genes in 231 Chinese wheat cultivars and 422 landraces using the STS marker csLV34 which found with a frequency of 6.1 % and 89.6 % in cultivars and landraces, respectively. The result that higher frequency in landraces than cultivars in Yang’s study and present study indicates an erosion of this gene resulting from preferring using dominant R genes which were easier to select and breed into existing crop cultivars in Chinese breeding programs. Though these primers are developed based on the gene sequence, they seem not to be prefect markers for the susceptible control variety Mingxian 169 that was detected to be positive by these markers. After sequencing the region spanning introns 9 ton 13 of the Lr34/Yr18 gene (about 751 bp) of Chinese Spring and Mingxian169, there was no difference (data not shown). The result indicates that Mingxian 169 may have a different allele at the Yr18 locus. The complete sequence of the Mingxian 169 allele at the Yr18 locus may needed to test the hypothesis.
Conclusion
The results revealed a low number of ASR cultivars and a low diversity of resistance genes in Chinese cultivars. After analyzing the pedigree of 55 entries which showed ASR to the four epidemic races (CYR23, CYR29, CYR32 and CYR33), the Guinong and 92R series are the most frequently used resistance sources in wheat breeding lines and currently grown cultivars. Once resistance of Guinong series and Yr26 are overcome by a new race, there will be severe epidemics and consequent yield losses. The continuing use of Guinong series and Yr26 is a major concern as Yr26 has become ineffective against a newly emerging race. Therefore, identifying new genes for effective all-stage resistance and non-race specific resistance and use those in breeding programs to develop wheat cultivars with diverse and effective resistance genes are very urgent for preventing major stripe rust epidemics. When Sr33, an major Puccinia graminis f. sp. tritici (Pgt) resistance gene was combined with Sr2, an adult plant, partial, non-race specific Pgt gene that has provided durable resistance for more than 70 years to stem rust (Periyannan et al. 2013).
Abbreviations
- Pst:
-
Puccinia striiformis Westend. f. sp. tritici Erikss
- CYR:
-
Chinese yellow rust
- ASR:
-
All-stage resistance
- APR:
-
Adult-plant resistance
- SR:
-
Slow-rusting
- Yr:
-
Yellow rust (stripe rust) resistance genes
- PAGE:
-
Polyacrylamide gel electrophoresis
- IT:
-
Infection type
- DS:
-
Disease severity
- TSA:
-
Triticum spelta album
- STS:
-
Sequence-tagged site
- CAPS:
-
Cleaved amplified polymorphic sequence
- HTAP:
-
High-temperature adult-plant resistance
References
An Z, Xie L, Cheng H, Zhou Y, Zhang Q, He X, Huang H (2009) A silver staining procedure for nucleic acids in polyacrylamide gels without fixation and pretreatment. Analyt Biochem 391:77–79
Chen XM (2005) Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can J Plant Pathol 27(3):314–317
Chen XM, Soria MA, Yan GP, Sun J, Dubcovsky J (2003) Development of sequence tagged site and cleaved amplified polymorphic sequence markers for wheat stripe rust resistance gene Yr5. Crop Sci 43:2058–2064
Chen WQ, Wu LR, Liu TG, Xu SC, Jin SL, Peng YL, Wang BT (2009) Race dynamics, diversity, and virulence evolution in Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust in China from 2003 to 2007. Plant Dis 93:1093–1101
Francis HA, Leitch AR, Koebner RMD (1995) Conversion of a RAPD-generated PCR product, containing a novel dispersed repetitive element, into a fast and robust assay for the presence of rye chromatin in wheat. Theor Appl Genet 90:636–642
Helguera M, Khan IA, Kolmer J, Lijavetzky D, Zhong-qi L, Dubcovsky J (2003) PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Sci 43:1839–1847
Kang Z, Zhao J, Han D, Zhang H, Wang X, Wang C, Han Q, Guo J, Huang L (2010) Status of wheat rust research and control in China. In: BGRI 2010 Technical Workshop Oral Presentations, 2010. p 50
Lagudah E, McFadden H, Singh R, Huerta-Espino J, Bariana H, Spielmeyer W (2006) Molecular genetic characterization of the Lr34/Yr18 slow rusting resistance gene region in wheat. Theor Appl Genet 114:21–30
Lagudah E, Krattinger S, Herrera-Foessel S, Singh R, Huerta-Espino J, Spielmeyer W, Brown-Guedira G, Selter L, Keller B (2009) Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor Appl Genet 119:889–898
Li ZQ, Zeng SM (2002) Wheat rust in China. China agriculture press, Beijing
Li G, Li Z, Yang W, Zhang Y, He Z, Xu S, Singh R, Qu Y, Xia X (2006) Molecular mapping of stripe rust resistance gene YrCH42 in Chinese wheat cultivar Chuanmai 42 and its allelism with Yr24 and Yr26. Theor Appl Genet 112(8):1434–1440
Lin F, Chen XM (2007) Genetics and molecular mapping of genes for race-specific all-stage resistance and non-race-specific high-temperature adult-plant resistance to stripe rust in spring wheat cultivar alpowa. Theor Appl Genet 114:1277–1287
Lin F, Chen XM (2009) Quantitative trait loci for non-race-specific, high-temperature adult-plant resistance to stripe rust in wheat cultivar express. Theor Appl Genet 118:631–642
Line RF, Chen X (1995) Successes in breeding for and managing durable resistance to wheat rusts. Plant Dis 79:1254–1255
Line RF, Qayoum A (1992) Virulence, aggressiveness, evolution, and distribution of races of Puccinia striiformis (the cause of stripe rust of wheat) in North America. U.S. Department of Agriculture Techchnical Bulletin No. 1788
Liu C, Yang Z-J, Li G-R, Zeng Z-X, Zhang Y, Zhou J-P, Liu Z-H, Ren Z-L (2008) Isolation of a new repetitive DNA sequence from Secale africanum enables targeting of Secale chromatin in wheat background. Euphytica 159:249–258
Liu TG, Peng YL, Chen WQ, Zhang ZY (2010) First detection of virulence in Puccinia striiformis f. sp. tritici in China to resistance genes Yr24 (=Yr26) present in wheat cultivar Chuanmai 42. Plant Dis 94:1163
McIntosh RA, Silk J, The TT (1996) Cytogenetic studies in wheat XVII. Monosomic analysis and linkage relationships of gene Yr15 for resistance to stripe rust. Euphytica 89:395–399
McIntosh RA, Dubcovsky J, Rogers WJ, Morris C, Appels R, Xia XC (2012) Catalogue of gene symbols for wheat: 2012 supplement. http://www.shigen.nig.ac.jp/wheat/komugi/genes/symbolClassList.jsp
Murphy LR, Santra D, Kidwell K, Yan G, Chen X, Campbell KG (2009) Linkage maps of wheat stripe rust resistance genes Yr5 and Yr15 for use in marker-assisted selection. Crop Sci 49:1786–1790
Periyannan S, Moore J, Ayliffe M, Bansal U, Wang X, Huang L, Deal K, Luo M, Kong X, Bariana H, Mago R, McIntosh R, Dodds P, Dvorak J, Lagudah E (2013) The Gene Sr33, an ortholog of barley Mla genes, encodes resistance to wheat stem rust race Ug99. Science 341(6147):786–788. doi:10.1126/science.1239028
Roebbelen G, Sharp EL (1978) Mode of inheritance, interaction and application of genes conditioning resistance to yellow rust. Fortschritte der Pflanzenzuechtung 9:1–88
Sharma-Poudyal D, Chen X, Wan A, Zhan G, Kang Z, Cao S, Jin S, Morgounov A, Akin B, Mert Z (2013) Virulence characterization of international collections of the wheat stripe rust pathogen, Puccinia striiformis f. sp. tritici. Plant Dis 97:379–386
Singh R, Datta D, Priyamvada, Singh S, Tiwari R ((2009)) A diagnostic PCR based assay for stripe rust resistance gene Yr10 in wheat. Acta Phytopathologica et Entomologica Hungarica 44:11–18
Singh RP, Huerta-Espino J, Rajaram S (2000) Achieving near-immunity to leaf and stripe rusts in wheat by combining slow rusting resistance genes. Acta Phytopathologica et Entomologica Hungarica Hungary 35:133–139
Smith PH, Hadfield J, Hart NJ, Koebner RMD, Boyd LA (2007) STS markers for the wheat yellow rust resistance gene Yr5 suggest a NBS–LRR-type resistance gene cluster. Genome 50:259–265
Tanguy AM, Coriton O, Abélard P, Dedryver F, Jahier J (2005) Structure of Aegilops ventricosa chromosome 6Nv, the donor of wheatgenes Yr17, Lr37, Sr38, and Cre5. Genome 48:541–546
Wan A, Zhao Z, Chen X, He Z, Jin S, Jia Q, Yao G, Yang J, Wang B, Li G, Bi Y, Yuan Z (2004) Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis 88:896–904
Wan AM, Chen XM, He ZH (2007) Wheat stripe rust in China. Aust J Agric Res 58:605–619
Wang C, Zhang Y, Han D, Kang Z, Li G, Cao A, Chen P (2008) SSR and STS markers for wheat stripe rust resistance gene Yr26. Euphytica 159:359–366
Wang D, Feng J, Wang F, Lin R, Xu S (2013) Genetic diversity and stripe rust resistance of regional trial wheat cultivars in four provinces in 2010–2011. Plant Protection 39(1):21–28
Weining S, Ko L, Henry RJ (1994) Polymorphisms in the α-amy1 gene of wild and cultivated barley revealed by the polymerase chain reaction. Theor Appl Genet 89:509–513
Yang W, Yang F, Liang D, He Z, Shang X, Xia X (2008) Molecular characterization of slow-rusting genes Lr34/Yr18 in Chinese wheat cultivars. Acta Agronomica Sinica 34:1109–1113
Zeng S, Luo Y (2006) Long-Distance spread and interregional epidemics of heat stripe Rust in China. Plant Dis 90:980–988
Zhou Y, Zhong HH, Zhang GS, Xia LQ, Chen XM, Gao YC, Jing ZB, Yu GJ ((2004)) Utilization of 1BL/1RS translocation in wheat breeding in China. Acta Agronomica Sinica 30:531–535
Zhuang QS (2003) Wheat improvement and pedigree analysis in China. China agriculture press, Beijing
Acknowledgments
This study was financially supported by the National Basic Research Program of China (No.2013CB127700), the National Major Project of Breeding for New TransgenicOrganisms in China (2011ZX08002-001), the earmarked fund for Modern Agro-industry Technology Research System (No. CARS-3-1-11) and the 111 Project from the Ministry of Education of China (B07049).
Author information
Authors and Affiliations
Corresponding author
Additional information
Qing-Dong Zeng and De-Jun Han contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Zeng, QD., Han, DJ., Wang, QL. et al. Stripe rust resistance and genes in Chinese wheat cultivars and breeding lines. Euphytica 196, 271–284 (2014). https://doi.org/10.1007/s10681-013-1030-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-013-1030-z