
Abstract
Porcine epidemic diarrhea virus (PEDV) infects the intestine of young pigs, but effective measures for prevention and treatment are lacking. N-Acetylcysteine (NAC) has been shown to reduce endotoxin-induced intestinal dysfunction. This study was conducted with the PEDV-infected neonatal piglet model to determine the effect of NAC supplementation on intestinal function. Thirty-two 7-day-old piglets were randomly allocated to one of four treatments in a 2 × 2 factorial design consisting of two liquid diets (0 or 50 mg/kg BW NAC supplementation) and oral administration of 0 or 104.5 TCID50 (50% tissue culture infectious dose) PEDV. On day 7 of the trial, half of the pigs (n = 8) in each dietary treatment received either sterile saline or PEDV (Yunnan province strain) solution at 104.5 TCID50 per pig. On day 10 of the trial, d-xylose (0.1 g/kg BW) was orally administrated to all pigs. One hour later, jugular vein blood samples were collected, and then all pigs were killed to obtain the small intestine. PEDV infection increased diarrhea incidence, while reducing ADG. PEDV infection also decreased plasma d-xylose concentration, small intestinal villus height, mucosal I-FABP and villin mRNA levels but increased mucosal MX1 and GCNT3 mRNA levels (P < 0.05). Dietary NAC supplementation ameliorated the PEDV-induced abnormal changes in all the measured variables. Moreover, NAC reduced oxidative stress, as indicated by decreases in plasma and mucosal H2O2 levels. Collectively, these novel results indicate that dietary supplementation with NAC alleviates intestinal mucosal damage and improves the absorptive function of the small intestine in PEDV-infected piglets.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
N-Acetylcysteine (also known as N-acetyl cysteine, N-acetyl-l-cysteine or NAC) is a source of sulfhydryl groups in cells and this sulfur-containing compound plays a critical role in animal physiology (Hou et al. 2013; Wu et al. 2004). For several decades, NAC has been used in clinical practice, for example, as a mucolytic agent and a curative for acetaminophen intoxication, doxorubicin cardiotoxicity, ischemia–reperfusion cardiac injury, acute respiratory distress syndrome, bronchitis, radiocontrast-induced nephropathy, chemotherapy-induced toxicity, HIV/AIDS, heavy metal toxicity and virus infection (Samuni et al. 2013; Berk et al. 2013; Radomska-Lesniewska and Skopinski 2012; Sadowska 2012; Anderson et al. 2011; Baker et al. 2009; Millea 2009; Dodd et al. 2008; Fishbane 2008; Atkuri et al. 2007; Fishbane et al. 2004; Zafarullah et al. 2003; Cotgreave 1997).
NAC is rapidly metabolized by the small intestine to produce glutathione (Hou et al. 2012, 2013) and it cannot be detected in the blood and other tissues of animals without intake from supplementation (Wu 2009). Enterocytes, colonocytes, and other intestinal cells readily transport NAC and convert it into l-cysteine, which helps to maintain the integrity, growth, and function of the intestinal mucosa (Hou et al. 2015; Wu et al. 2013; Wu 2014). We recently demonstrated that dietary supplementation with 500 mg/kg NAC alleviated the intestinal damage and dysfunction in LPS-infected piglets (Hou et al. 2012, 2013, 2015), which was mediated through the activation of multiple signaling pathways involving PI3K/Akt/mTOR, EGFR, TLR4/NF-κB, AMPK, and type I IFN pathways (Yi et al. 2017).
Porcine epidemic diarrhea virus (PEDV) belongs to the Coronaviridae family (Bertolini et al. 2016), and is the causative agent for porcine epidemic diarrhea (PED), a disease which was first described in England in the 1970s (Thomas et al. 2015). Since then, PED outbreaks have become challenging problems in pig producing countries all over the world, causing large economic losses (Bertolini et al. 2016). PEDV strains spread rapidly and causes acute diarrhea, vomiting, dehydration and high mortality (70–100%) in seronegative neonatal piglets (Jung and Saif 2015; Li et al. 2016; Bowman et al. 2015). As a result of the significant impact of PEDV, the US pork industry lost almost 10% of its domestic pig population after only a 1-year-epidemic period, amounting to approximately 7 million piglets (Jung and Saif 2015). Since late 2010, severe PED epizootics caused by new variants of PEDV have affected millions of pigs in the People’s Republic of China (Lv et al. 2016).
PEDV strains are highly enteropathogenic and can acutely infect villous epithelial cells of the entire intestine, although the jejunum and ileum are the primary target sites. PEDV infection causes acute, severe atrophic enteritis accompanied by viremia that leads to profound diarrhea and vomiting, followed by extensive dehydration, and is a major cause of death in nursing piglets (Jung and Saif 2015).
In previous studies, we have demonstrated that dietary NAC supplementation could reduce inflammation and oxidative stress, alleviate damage to the small intestine (Hou et al. 2013, 2015) and acetic acid-induced colitis in piglets (Wang et al. 2013). Therefore, we hypothesized that NAC could ameliorate PEDV-infected intestinal injury and could be used as a curative for the treatment of PED. This study was conducted to test this hypothesis and elucidate the underlying mechanisms. Findings of this study will provide vital clues for the prevention and treatment of PED in pigs and possibly other animal species with viral infection.
Materials and methods
Animal care and diets
The animal use protocol for this research was approved by the Animal Care and Use Committee of Hubei Province, China. All pigs used in this experiment were born naturally at term (114 days of gestation). Thirty-two crossbred healthy piglets (Duroc × Landrace × Yorkshire, 7-day-old neonatal pigs) initially weighing 2.86 ± 0.3 kg were used in this experiment. Pigs of the same gender were used for keeping animal uniformity. Pigs were penned individually (1.80 × 1.10-m pens) in a temperature-controlled nursery barn (28–30 °C). They were housed individually in pens. NAC was obtained from Sigma Chemicals (Sigma Chemical Inc., St. Louis, MO, USA). The experimental basal diet (liquid milk replacer), which was formulated to meet requirements of all nutrients for suckling piglets, was obtained from Wuhan Anyou Feed Co., Ltd (Wuhan, China). Prior to feeding, the milk replacer (powder) was dissolved in warm water (45–55 °C) to form a liquid feed (dry matter content of 20%), as described by Wang et al. (2014). Pigs were fed the liquid feed every 3 h between 8:00 a.m. and 8:00 p.m. This feeding strategy (more frequent provision of food at a small amount for each feeding) was adopted to prevent intestinal necrosis and improve the efficiency of utilization of dietary nutrients for piglet growth.
Experimental design
Throughout the study, pigs had free access to drinking water and were housed individually. They were fed NAC-supplemented liquid diet or the control liquid diet for 7 days before PEDV challenge (16 pigs/group). Immediately after PEDV challenge, the experiment was changed to a 2 × 2 factorial design. The main factors consisted of diet (0 or 50 mg/kg BW NAC supplementation in diet; +NAC or −NAC) and challenge (PEDV or saline administration; +PEDV or −PEDV). On day 7 of the trial, half of the pigs (n = 8) in each dietary treatment were administered orally with either PEDV (PEDV epidemic strain in Yunnan province) solution (kindly provided by Dr. He in College of Veterinary Medicine in Huazhong Agricultural University) at the dose of 104.5 TCID50 (50% tissue culture infectious dose) per pig (PEDV infection) or the same volume of sterile saline (Control). Each treatment group contained 8 piglets. NAC (powder) or PEDV (solution) was dissolved in sterile saline. The dose of PEDV (104.5 TCID50) was chosen to induce infection in accordance with our previous study. To exclude a possible effect of PEDV-induced reduction in food intake on the piglet intestine, all the piglets were pair-fed during days 0–10 of the trial. On day 10 of the trial, 10% d-xylose (1 mL/kg BW) was orally administrated to piglets, and the d-xylose absorptive test was performed to measure intestinal absorption capacity and mucosal integrity (Haeney et al. 1978). At 1 h after the administration of d-xylose, blood samples were collected, and then all pigs were killed under sodium pentobarbital anesthesia (50 mg/kg BW, iv) to obtain the small intestine (Hou et al. 2010, 2011a, b; Deng et al. 2009).
Blood sample collection
On day 10 of the trial, 1 h after the oral administration of d-xylose, blood samples were collected from anterior vena cava into heparinized vacuum tubes (Becton-Dickinson Vacutainer System, Franklin Lake, NJ, USA), as described by Hou et al. (2010). Blood samples were centrifuged at 3000 rpm for 10 min at 4 °C to obtain plasma (Hou et al. 2010; Tan et al. 2009), which was stored at −80 °C until analysis.
Intestinal sample collection
The pig abdomen was opened immediately from the sternum to the pubis, and the whole gastrointestinal tract was immediately exposed (Hou et al. 2010; Li et al. 2009). The small intestine was dissected free of the mesentery and placed on a chilled stainless steel tray. The 3- and 10-cm segments were cut at mid-jejunum and mid-ileum, respectively (Hou et al. 2010; Wang et al. 2008a). The 3-cm intestinal segments were flushed gently with ice-cold phosphate buffered saline (PBS, pH 7.4) and then placed in 10% fresh, chilled formalin solution for histological measurements (Hou et al. 2010; Nofrarías et al. 2006).
Intestinal segments (10-cm in length) were opened longitudinally and the contents were flushed with ice-cold PBS (Hou et al. 2010; Wang et al. 2008a). Mucosa was collected by scraping using a sterile glass microscope slide at 4 °C (Hou et al. 2010; Wang et al. 2009), rapidly frozen in liquid nitrogen, and stored at −80 °C until analysis. All samples were collected within 15 min after killing.
Blood biochemical measurements in plasma
Concentrations of biochemical parameters in plasma were measured with corresponding kits using a Hitachi 7060 Automatic Biochemical Analyzer (Hitachi, Japan) (Wang et al. 2016).
Determination of d-xylose in plasma
d-xylose in plasma was determined as described by Hou et al. (2010). Briefly, 50 μL of the collected plasma was added to 5 mL of the phloroglucinol color reagent solution (Sigma Chemical Inc., St. Louis, MO, USA) and heated at 100 °C for 4 min. The samples were allowed to cool to room temperature in a water bath. A xylose standard solution was prepared by dissolving d-xylose in saturated benzoic acid (prepared in deionized water) to obtain 0, 0.7, 1.3, and 2.6 mmol/L. The xylose standard solution or the sample was added to the color reagent solution. Absorbance of the resultant mixture at 554 nm was measured using a spectrophotometer (Model 6100, Jenway LTD., Felsted, Dunmow, CM6 3LB, Essex, England, UK). The standard solution of 0 mmol/L d-xylose was considered as the blank.
Determination of diamine oxidase (DAO) activity in plasma
Diamine oxidase (DAO) activities in plasma were determined using spectrophotometry as described by Hosoda et al. (1989). The assay mixture (3.8 mL) contained 3 mL of phosphate buffer (0.2 M, pH 7.2), 0.1 mL (0.004%) of horseradish peroxidase solution (Sigma Chemicals), 0.1 mL of o-dianisidine-methanol solution [0.5% of o-dianisidine (Sigma Chemicals) in methanol], 0.5 mL of plasma, and 0.1 mL of substrate solution (0.175% of cadaverine dihydrochloride, Sigma Chemicals). This mixture was incubated for 30 min at 37 °C, and absorbance at 436 nm was measured to indicate DAO activity (Hou et al. 2011a). Protein was measured as described by Hou et al. (2010). DAO was used as a marker of intestinal injury (Luk et al. 1980).
Levels of anti-oxidative enzymes and oxidation-relevant products in plasma and intestinal mucosa
Plasma, jejunal and ileal mucosae were used for the analyses of anti-oxidative enzymes and related products. Catalase (CAT) activities, malondialdehyde (MDA) and hydrogen peroxide (H2O2) concentrations were determined as described by Hou et al. (2013). Activities of CAT, as well as MDA and H2O2 were determined using commercially available kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s protocols. Assays were performed in triplicate.
Measurement of mucosal protein, DNA, and RNA
The protein, DNA, and RNA were extracted from the jejunal and ileal mucosae, using the TRI REAGENT-RNA/DNA/Protein isolation reagent, and their concentrations were determined colorimetrically (Hou et al. 2010, 2012). Intestinal mucosal protein was analyzed according to the method of Lowry et al. (1951). DNA was analyzed fluorimetrically using the method of Prasad et al. (1972). RNA was determined by spectrophotometry using a modified Schmidt–Tannhauser method as described by Munro and Fleck (1969). For measurement of mucosal protein, intestinal mucosal samples (~0.1 g) were homogenized with a tissue homogenizer in 1 mL of ice-cold PBS–EDTA buffer (0.05 mol/L Na3PO4, 2.0 mol/L NaCl, 2 mmol/L EDTA, pH 7.4) and the homogenates were centrifuged at 12,000×g for 10 min at 4 °C to obtain the supernatant fluid for assays. For measurement of small intestinal DNA and RNA levels, the mucosa was homogenized (~2 min) in a 100-fold volume of ice-cold saline (0.9%) and the homogenate was centrifuged at 1800×g for 10 min at 4 °C to obtain the supernatant fluid for analysis.
Intestinal morphology
Intestinal segments for morphological analysis were dehydrated and embedded in paraffin, sectioned at 4 µm, and stained with hematoxylin and eosin (Hou et al. 2010, 2012; Luna 1968). Morphological measurements were performed with a light microscope (American Optical Co., Scientific Instrument Div., Buffalo, NY, USA). Villus height (the distance from the villus tip to crypt mouth) and width (the distance of the widest villi), crypt depth (the distance from the crypt mouth to the base) were measured using a linear ocular micrometer with a computer-assisted morphometric system (BioScan Optimetric, BioScan Inc., Edmonds, WA, USA). Only vertically oriented villi and crypts were measured (Hou et al. 2010; Nofrarías et al. 2006). Values are expressed as means from ten adjacent villi. Villus:crypt ratio and villous surface area (VSA) were calculated.
Real-time PCR measurement
MX1, GCNT3, I-FABP and Villin mRNA levels in jejunal and ileal mucosae were quantified using real-time PCR. Total RNA isolation, quantification, cDNA synthesis, and real-time PCR were carried out as previously described (Hou et al. 2013). The primer pairs used are shown in Table 1.
Total RNA was quantified by using the NanoDrop® ND-2000 UV–VIS spectrophotometer (Thermo Scientific, Wilmington, DE, USA) at an OD of 260 nm, and the purity was assessed by determining the OD260/OD280 ratio. All the samples had an OD260/OD280 ratio above 1.8, corresponding to 90–100% pure nucleic acids. Meanwhile, the integrity of RNA in each sample was assessed using 1% denatured agarose gel electrophoresis. RNA was used for RT-PCR analysis when it had a 28 S/18 S rRNA ratio ≥1.8 (Hou et al. 2013).
Total RNA was reverse-transcribed using the PrimeScript® RT reagent Kit with gDNA Eraser (Takara, Dalian, China) according to the manufacturer’s instruction. cDNA was synthesized and stored at −20 °C until use. The RT-PCR analysis of gene expression was performed using primers for MX1, GCNT3, I-FABP, Villin, and ribosomal protein L4 (RPL4) (Table 1) and the SYBR® Premix Ex Taq™ (Takara, Dalian, China) on an Applied Biosystems 7500 Fast Real-Time PCR System (Foster City, CA, USA). The total volume of the PCR reaction system was 50 µL. Amplification products were verified by melting curves, agarose gel electrophoresis, and direct sequencing. Results were analyzed by the cycle threshold (C T) method (Fu et al. 2010).
The standard curves were generated using relative concentration versus the C T. The linear correlation coefficient of all genes were >0.995. Based on the slopes of the standard curves, the amplification efficiencies of the standard ranged from 90 to 110% (derived from the formula efficiency = 101/−slope − 1) (Nygard et al. 2007). Moreover, we tested other housekeeping genes (GAPDH and β-actin) through analyzing gene stability as described by Vandesompele et al. (2002), and found that RPL4 expression was more stable than other housekeeping genes in the intestinal mucosa; therefore, we used RPL4 as the normalizer in the calculation of relative mRNA levels for target genes. Each biological sample was run in triplicate (Hou et al. 2013).
Statistical analysis
All experimental data for each group were analyzed by ANOVA using the GLM procedure of SPASS17.0 appropriate for a 2 × 2 factorial design (SPSS Inc. Chicago, IL, USA). The statistical model consisted of the effects of diet (+NAC vs. −NAC) and challenge (saline vs. PEDV) and their interactions. Data were expressed as means ± pooled SEM. When a significant NAC × PEDV interaction or a trend for NAC × LPS interaction occurred, post hoc testing was performed using the Bonferroni’s multiple comparison test. When variances were heterogeneous, data were log-transformed before ANOVA was conducted. Differences were considered significant when P ≤ 0.05. Probabilities which were >0.05 but <0.10 were considered as trends.
Results
Average daily weight gain of piglets
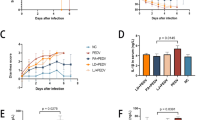
Data on the average daily weight gain (ADG) of piglets are summarized in Table 2. During days 0–7 of the trial (pre-challenge), there were no differences in the ADG of pigs fed the control and NAC-supplemented diets (P > 0.1). During days 7–10 of the trial (post-challenge), PEDV infection decreased the ADG (P < 0.05). No PEDV and NAC interaction was observed. Importantly, PEDV infection resulted in the occurrence of diarrhea in all (100%) pigs (P < 0.05), compared with pigs without PEDV infection (5% of the incidence of diarrhea) (data not shown).
Plasma biochemical parameters
Data on plasma biochemical parameters are summarized in Table 3. Compared with the non-infected pigs, the PEDV-infected pigs had higher concentrations of TP, ALB, TG, BUN, CL and GGT in plasma (P < 0.05) and tended to have higher plasma ALT (P = 0.064), and had a lower plasma TBIL (P < 0.05). However, pigs fed NAC had lower plasma ALT, AST and TBIL than the control pigs (P < 0.05). There were significant PEDV and NAC (or PEDV × NAC) interactions and crosstalk in affecting plasma ALT, AST and TBIL (P < 0.05) such that the responses of these parameters to the PEDV challenge were lower in those pigs fed the NAC compared with the PEDV-infected pigs fed the saline (−NAC), whereas there was no difference in these parameters in saline (−PEDV) supplemented pigs, and the data indicated that NAC supplementation attenuated the elevated levels of plasma ALT and AST in the PEDV-infected pigs (Table 3).
Blood DAO activity and d-xylose concentrations
Data on blood DAO activity and d-xylose concentrations are summarized in Table 4. The PEDV-infected pigs had a lower concentration of plasma d-xylose and a higher activity of plasma DAO than non-infected pigs (P < 0.05) (Table 4). However, pigs fed the NAC diet had a higher concentration of plasma d-xylose than the control pigs (P < 0.05). NAC supplementation enhanced plasma d-xylose concentration in both control pigs and the PEDV-infected pigs (P < 0.05).
Levels of anti-oxidative enzymes and oxidation-relevant products in plasma and intestinal mucosa
Data on CAT activities, MDA and H2O2 concentrations are summarized in Table 5. Compared with non-infected (−PEDV) pigs, PEDV-infected pigs had lower CAT activities in the plasma and ileum (P < 0.05), and higher MDA concentrations in the jejunum and ileum (P < 0.05), and H2O2 concentrations in the plasma and ileum (P < 0.05). There were no PEDV and NAC interactions in CAT activity in the jejunum, and H2O2 concentrations in the plasma, jejunum or ileum. However, the pigs fed the NAC diet had a higher jejunal CAT activity (P < 0.05), had lower plasma and jejunal H2O2 concentrations (P < 0.05), and tended to decrease ileal H2O2 concentration (P = 0.083). Of interest, dietary NAC supplementation reduced plasma and jejunal H2O2 concentrations in PEDV-infected pigs (Table 5).
Concentrations of protein, DNA and RNA in the small intestinal mucosa
Data on protein, DNA and RNA concentrations are summarized in Table 6. PEDV infection decreased total RNA concentration in the jejunum and ileum (P < 0.05) and RNA/DNA ratios in the ileum (P < 0.05). There were PEDV and NAC interactions in jejunal and ileal protein concentrations (P < 0.05) and ileal protein/DNA ratios (P < 0.05). Our results also showed that dietary NAC supplementation increased jejunal protein concentrations, ileal protein concentrations, and ileal protein/DNA ratios in PEDV-infected pigs, whereas NAC had no effect on these variables in non-infected pigs. No PEDV and NAC interaction was observed in jejunal RNA/DNA or ileal protein/DNA ratios. Thus, dietary supplementation of NAC increased jejunal RNA/DNA and ileal protein/DNA ratios (P < 0.05). Additionally, it was observed that under PEDV challenge dietary supplementation of NAC increased the ileal RNA/DNA ratio (P = 0.076) (Table 6).
Intestinal morphology
Data on the small intestinal morphology are summarized in Table 7. PEDV infection decreased villus height, villus height/crypt depth ratios, and villous surface area in the jejunum and ileum (P < 0.001), and increased jejunal crypt depth (P < 0.05). There were PEDV × NAC interactions in villus height and crypt depth in the jejunum (P < 0.05), villous surface area in the jejunum and ileum (P < 0.05), and a trend for ileal villus height (P = 0.060); the data indicated NAC supplementation could elevate villus height and villous surface area in jejunum and ileum in non-infected (−PEDV) pigs, whereas NAC had no influence on these variables in PEDV-infected pigs. Of note, NAC supplementation could reduce jejunal crypt depth in PEDV-infected pigs, but not in non-infected (−PEDV) pigs. No PEDV × NAC interaction was observed for villus height/crypt depth ratios in the jejunum and ileum. Dietary supplementation of NAC increased villus height/crypt depth ratios in the jejunum and ileum (P < 0.05), compared with the control (−NAC).
MX1, GCNT3, I-FABP and Villin mRNA levels in jejunal and ileal mucosae
PEDV infection increased MX1 and GCNT3 mRNA levels in the jejunum and ileum (P < 0.05), and decreased I-FABP and villin mRNA levels in the jejunum and ileum (P < 0.05) (Table 8). There were PEDV × NAC interactions in ileal I-FABP mRNA levels (P < 0.05) and jejunal and ileal villin mRNA levels (P < 0.001). NAC supplementation increased ileal I-FABP mRNA levels, as well as jejunal and ileal villin mRNA levels in PEDV-infected (+PEDV) pigs, whereas NAC had no effect on these variables in non-infected (−PEDV) pigs. No PEDV × NAC interaction was observed for MX1 or GCNT3 in the jejunum or ileum. Dietary supplementation of NAC decreased MX1 and GCNT3 mRNA levels in the jejunum and ileum (P < 0.05), compared with the control (−NAC). Specifically, dietary NAC supplementation attenuated the elevations of mRNA levels for MX1 in the jejunum and ileum, GCNT3 in the ileum of PEDV-infected pigs. Additionally, dietary NAC attenuated the reductions of I-FABP in the ileum, and villin in the jejunum and ileum of PEDV-infected pigs (Table 8).
Discussion
Based on the findings that NAC can exert beneficial effects in treating inflammatory bowel disease in animal models and human clinical trials (Hou et al. 2012, 2013, 2015; Wang et al. 2013; Cetinkaya et al. 2005), we investigated the protective effect of NAC on intestinal morphology and barrier function in PEDV-infected piglets because PEDV is deemed to be highly enteropathogenic. We were prompted to do this study in view of recent PED outbreaks in certain countries, including China and USA (Bertolini et al. 2016). An effective method for alleviating PED will be extremely beneficial for the global swine industry and will also have important implications for the prevention and treatment of virus-induced intestinal dysfunction in human infants and calves.
PEDV can infect pigs of all ages and causes acute and watery diarrhea, vomiting, dehydration, and high mortality in neonatal piglets (Lv et al. 2016). In the current study, a PEDV infection model was successfully developed with neonatal pigs by oral administration of 104.5 TCID50 PEDV per pig. The PEDV-infected pigs exhibited the symptoms of diarrhea and vomiting. In fact, oral administration of PEDV decreased the ADG of the piglets and resulted in a diarrhea incidence of 100%. PEDV infections were reported to cause acute, severe atrophic enteritis accompanied by viremia (Jung and Saif 2015). In keeping with this notion, we found that PEDV infection increased some plasma biochemical parameters in pigs, such as ALT, TP, ALB, TG, BUN, CL and GGT. Results of the present study indicated that PEDV infection readily resulted in systemic inflammation in piglets.
Elevations in plasma transaminases (ALT and AST) serve as a sensitive indicator of damage in tissues, particularly the liver (Yi et al. 2014; Chen et al. 2013). In addition, plasma TBIL level is also a valuable parameter of hepatic injury (Chen et al. 2013). Interestingly, dietary NAC supplementation alleviated the PEDV-induced increases in plasma ALT and AST activities, as we reported previously (Yi et al. 2014). Our new findings suggest that NAC supplementation could attenuate the damaged the hepatic architecture and function induced by PEDV infection and indicate a preventive effect of NAC against PEDV infection.
Intestinal biochemical indices, such as protein, RNA and DNA concentrations, as well as RNA/DNA and protein/DNA ratios, can be used to assess intestinal development (Hou et al. 2012; Fasina et al. 2007; Iji et al. 2001; Jeurissen et al. 2002). Mucosal protein and DNA contents are important indicators for cell metabolism (Liu et al. 2008). DNA concentration reflects the rate of mitosis in the production of new columnar epithelial cells, whereas RNA/DNA ratios indicate cellular efficiency and protein/DNA ratios implicate the efficiency of cellular protein synthesis (Fasina et al. 2007). In the present study, dietary supplementation of NAC restored the protein concentration in the jejunal and ileal mucosae, as well as protein/DNA ratios in the ileal mucosa of PEDV-infected piglets. In addition, regardless of PEDV infection, NAC increased the protein/DNA ratios in the jejunal mucosa (Table 6). These results demonstrate that dietary NAC stimulates the growth of the small intestinal mucosa under PEDV infection, which is in agreement with of our previous observations that dietary supplementation of NAC restored DNA concentration, as well as RNA/DNA and protein/DNA ratios in the jejunal mucosa of LPS-challenged piglets (Hou et al. 2012).
PEDV acutely infects villous epithelial cells of the entire intestine causing severe villous shortening (Jung and Saif 2015). Intestinal epithelial cells function as a physiological and structural barrier (Khurana and George 2008). Villus height, crypt depth, and villous surface area are biomarkers of gross intestinal morphology (Liu et al. 2012). Thus, an increase in villus height, villous surface area, and/or villus/crypt ratio points to an improvement of nutrient digestion and absorption (Hou et al. 2010; Montagne et al. 2003; Wu et al. 1996). In the current study, the PEDV infection decreased villus height, villus height/crypt depth ratios, and villous surface area, yet increased crypt depth in the jejunum and ileum (Table 7). These results indicate that PEDV induced a variety of morphologic alterations in the digestive tract and an increase in mucosal permeability. Notably, in this regard, we observed that NAC supplementation enhanced villus height, the ratio of villus height-to-crypt depth, and villous surface area, and decreased crypt depth in the jejunum and ileum (Table 7). These findings support the notion that NAC beneficially alleviates the PEDV-induced damage of the intestinal structure. Our findings are consistent with those of Hou et al. (2012) for lipopolysaccharide-challenged weanling pigs fed a diet supplemented with 500 mg NAC/kg.
Previous studies has reported that NAC improves the barrier function and the ability of the small intestine to absorb nutrients (Hou et al. 2012) and is potentially beneficial as a complementary agent in treating ulcerative colitis (Cetinkaya et al. 2005). Consistent with this view, in the current study, dietary supplementation with NAC prevented the PEDV-induced decrease in villin expression in jejunal and ileal mucosae (Table 8). Because villin is a marker of villus cell differentiation (West et al. 1988; Moll et al. 1987), enhanced villin expression implies that more villus cells undergo differentiation (Wang et al. 2008b; Gröne et al. 1986). This result indicates a protective effect of NAC against PEDV-induced enterocyte death and further supports the notion that dietary supplementation with NAC is effective in preventing intestinal injury in neonates.
An intact intestinal barrier plays a central role in preventing the penetration of luminal bacteria and dietary allergens into the mucosa (Liu et al. 2012; Martín-Venegas et al. 2006). Intestinal barrier function can be commonly assessed by many indices, such as d-xylose concentration and DAO activity in plasma (Liu et al. 2012; Hou et al. 2012). Plasma d-xylose concentration is a simple, specific, and sensitive indicator of intestinal absorption capacity. In healthy pigs, d-xylose is readily absorbed by the small intestine. However, under conditions of malabsorption, the entry of d-xylose from the intestinal lumen to the portal vein is impaired, thereby reducing d-xylose concentrations in both blood and urine (Hou et al. 2012; Mansoori et al. 2009). Additionally, plasma DAO can serve as a biomarker for the severity of mucosal injury (Hou et al. 2011b, 2012; Luk et al. 1983). In mammals, DAO is abundantly expressed in the upper part of the intestinal mucosa. Under certain circumstances, intestinal mucosal cells undergo necrosis and are sloughed off into the intestinal lumen, leading to an increase in the circulating levels of DAO (Li et al. 2002). Similarly, intestinal fatty acid-binding protein (I-FABP) is also detectable in the circulation during enterocyte death (Coufal et al. 2016). I-FABP locates mainly in small intestine enterocytes, and is released into the blood stream after intestinal ischemia and cell disruption (Cheng et al. 2015; Pelsers et al. 2005). Therefore, I-FABP is considered as another marker of intestinal cell damage. In the present study, we observed that PEDV decreased plasma d-xylose concentration, while increasing DAO activity in the plasma. Moreover, PEDV resulted in a lower I-FABP expression in the small intestine. These results indicate that PEDV induces intestinal cell damage and impairs intestinal function. Importantly, NAC supplementation improved intestinal function, as indicated by increased plasma d-xylose concentration (Table 4) and ileal mucosal I-FABP mRNA levels (Table 8). In addition, we found that NAC supplementation augmented the entry of orally administered d-xylose into the systemic circulation (Table 4), which is consistent with our previous observation (Hou et al. 2010, 2012). Furthermore, we observed that NAC supplementation elevated I-FABP mRNA levels in the ileum of PEDV-infected pigs (Table 8). Taken together, these data indicate that NAC can attenuate intestinal injury and improve intestinal function under PEDV infection.
Elevation of reactive oxygen species (ROS) production is well recognized as an essential factor in the pathogenesis of GI mucosal diseases, including peptic ulcers, inflammatory bowel disease (IBD), and GI cancers (Chu et al. 2016; Bhattacharyya et al. 2014). As a scavenger of free radicals, NAC interacts with oxidants and protects cells against oxidative damage (Hou et al. 2013; Zafarullah et al. 2003). In the current study, levels of MDA (malondialdehyde) and H2O2 (hydrogen peroxide) were elevated in the plasma and small intestine of PEDV-infected pigs, while CAT (catalase) activities in the plasma and intestine were decreased (Table 5). MDA is an important indicator of oxidative damage (Wang et al. 2013; Hou et al. 2011a) and can serve as a useful biomarker of in vivo oxidative stress (Hou et al. 2014; Fang et al. 2002). ROS, such as H2O2, are produced primarily by the mitochondria in cells as a by-product of normal metabolism during the conversion of molecular oxygen and hydrogen ions to water (Hou et al. 2013; Fang et al. 2002). However, cells possess defense mechanisms against ROS and other oxidants. H2O2 is degraded to water by anti-oxidative enzymes including CAT, which catalyzes the reduction of H2O2 to water (Wu et al. 2004). In the present study, we found that NAC supplementation increased CAT activity in the jejunum and decreased H2O2 concentration in the plasma and jejunum of PEDV-infected pigs (Table 5). Thus, it appears that NAC exerts its antioxidant effects in pigs at both the systemic and intestinal levels, which is in agreement with our previous results from the studies involving lipopolysaccharide-challenged weanling pigs (Hou et al. 2013).
The MX (myxovirus resistance 1, interferon-induced p78) proteins are major effector molecules that prevent influenza-infected animals from developing severe phenotypes (Godino and Fernández 2016). MX1 is an important downstream effector of type I interferons characterized by a tripartite GTP-binding domain and leucine zipper motif (AlFadhli et al. 2016). The MX1 gene has been shown to possess the disease-resistance function against a variety of RNA viruses that include influenza A in several species, including pigs, Sus scrofa (Godino and Fernández 2016). In addition, MX1 plays a major role in apoptosis and cytokine-mediated cell signaling (AlFadhli et al. 2016) and its expression is strongly induced by viral infections (Haller et al. 2007; Yan et al. 2014). We observed that PEDV increased the expression of MX1 in the jejunum and ileum (Table 8), which can be attenuated by NAC supplementation. This novel observation provides a new solution to treating patients with virus-induced intestinal dysfunction.
Altered glycosylation is considered as a cancer hallmark. Expression of mucin-type core 2 1,6-N-acetylglucosaminyltransferase enzyme (C2GnT-M), encoded by the GCNT3 gene, is detected in the colon, kidney, pancreas, small intestine, trachea and stomach. C2GnT-M has been reported to possess tumor suppressor properties (González-Vallinas et al. 2014). In our study, GCNT3 expression was up-regulated in the jejunal and ileal mucosae of PEDV-infected pigs. Intriguingly, NAC supplementation suppressed GCNT3 expression in the jejunal and ileal mucosae of the pigs. Because little is known about the association of GCNT3 with PEDV infection, further studies are warranted to determine if GCNT3 inhibitors could function as anti-PEDV drugs.
In conclusion, dietary NAC supplementation exerts beneficial effects on improving intestinal integrity and absorption in PEDV-infected neonatal piglets. The effects of NAC are associated with the following changes: (1) alleviated small intestinal injury and improved the absorptive function (indicated by the alleviation of the PEDV-induced damage of the small intestinal structure, increases in plasma d-xylose concentrations, protein concentration, RNA/DNA ratios, and protein/DNA ratios, and up-regulation of I-FABP and villin expression in the small intestinal mucosa); (2) reduced oxidative stress (indicated by the decrease of H2O2 concentration in the plasma and small intestinal mucosa); and (3) decreases in the expression of MX1 and GCNT3 in the small intestinal mucosa. These findings not only aid in the understanding the role of NAC in PEDV-infected neonatal piglets, but also have important implications for the improvement of infant nutrition in humans and other animals during viral infection.
Abbreviations
- CAT:
-
Catalase
- C T :
-
Cycle threshold
- H2O2 :
-
Hydrogen peroxide
- I-FABP:
-
Intestinal fatty acid-binding protein
- MDA:
-
Malondialdehyde
- MX1:
-
Myxovirus resistance 1
- NAC:
-
N-Acetylcysteine
- PEDV:
-
Porcine epidemic diarrhea virus
- SEM:
-
Standard error of the mean
- TCID50:
-
50% tissue culture infectious dose
References
AlFadhli S, Al-Mutairi M, Al Tameemi B, Nizam R (2016) Influence of MX1 promoter rs2071430 G/T polymorphism on susceptibility to systemic lupus erythematosus. Clin Rheumatol 35:623–629
Anderson SM, Park ZH, Patel RV (2011) Intravenous N-acetylcysteine in the prevention of contrast media-induced nephropathy. Ann Pharmacother 45:101–107
Atkuri KR, Mantovani JJ, Herzenberg LA, Herzenberg LA (2007) N-acetylcysteine—a safe antidote for cysteine/glutathione deficiency. Curr Opin Pharmacol 7:355–359
Baker WL, Anglade MW, Baker EL, White CM, Kluger J, Coleman CI (2009) Use of N-acetylcysteine to reduce post-cardiothoracic surgery complications: a meta-analysis. Eur J Cardiothorac Surg 35:521–527
Berk M, Malhi GS, Gray LJ, Dean OM (2013) The promise of N-acetylcysteine in neuropsychiatry. Trends Pharmacol Sci 34:167–177
Bertolini F, Harding JC, Mote B, Ladinig A, Plastow GS, Rothschild MF (2016) Genomic investigation of piglet resilience following porcine epidemic diarrhea outbreaks. Anim Genet. doi:10.1111/age.12522
Bhattacharyya A, Chattopadhyay R, Mitra S, Crowe SE (2014) Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol Rev 94:329–354
Bowman AS, Nolting JM, Nelson SW, Bliss N, Stull JW, Wang Q, Premanandan C (2015) Effects of disinfection on the molecular detection of porcine epidemic diarrhea virus. Vet Microbiol 179:213–218
Cetinkaya A, Bulbuloglu E, Kurutas EB, Ciralik H, Kantarceken B, Buyukbese MA (2005) Beneficial effects of N-acetylcysteine on acetic acid-induced colitis in rats. Tohoku J Exp Med 206:131–139
Chen F, Liu Y, Zhu H, Hong Y, Wu Z, Hou Y, Li Q, Ding B, Yi D, Chen H (2013) Fish oil attenuates liver injury caused by LPS in weaned pigs associated with inhibition of TLR4 and nucleotide-binding oligomerization domain protein signaling pathways. Innate Immun 19:504–515
Cheng S, Yu J, Zhou M, Tu Y, Lu Q (2015) Serologic intestinal-fatty acid binding protein in necrotizing enterocolitis diagnosis: a meta-analysis. Biomed Res Int 2015:156704
Chu FF, Esworthy RS, Doroshow JH, Grasberger H, Donko A, Leto TL, Gao Q, Shen B (2016) Deficiency in Duox2 activity alleviates ileitis in GPx1- and GPx2-knockout mice without affecting apoptosis incidence in the crypt epithelium. Redox Biol 11:144–156
Cotgreave IA (1997) N-acetylcysteine: pharmacological considerations and experimental and clinical applications. Adv Pharmacol 38:205–227
Coufal S, Kokesova A, Tlaskalova-Hogenova H, Snajdauf J, Rygl M, Kverka M (2016) Urinary intestinal fatty acid-binding protein can distinguish necrotizing enterocolitis from sepsis in early stage of the disease. J Immunol Res 2016:5727312
Deng D, Yin YL, Chu WY, Yao K, Li TJ, Huang RL, Liu ZQ, Zhang JS, Wu G (2009) Impaired translation initiation activation and reduced protein synthesis in weaned piglets fed a low-protein diet. J Nutr Biochem 20:544–552
Dodd S, Dean O, Copolov DL, Malhi GS, Berk M (2008) N-acetylcysteine for antioxidant therapy: pharmacology and clinical utility. Expert Opin Biol Ther 8:1955–1962
Fang YZ, Yang S, Wu G (2002) Free radicals, antioxidants, and nutrition. Nutrition 18:872–879
Fasina YO, Moran ET, Ashwell CM, Conner DE, Leslie M, Mckee SR (2007) Effect of dietary gelatin supplementation on the expression of selected enterocyte genes, intestinal development and early chick performance. Int J Poult Sci 6:944–951
Fishbane S (2008) N-acetylcysteine in the prevention of contrast-induced nephropathy. Clin J Am Soc Nephrol 3:281–287
Fishbane S, Durham JH, Marzo K, Rudnick M (2004) N-acetylcysteine in the prevention of radiocontrast-induced nephropathy. J Am Soc Nephrol 15:251–260
Fu WJ, Stromberg AJ, Viele K, Carroll RJ, Wu G (2010) Statistics and bioinformatics in nutritional sciences: analysis of complex data in the era of systems biology. J Nutr Biochem 21:561–572
Godino RF, Fernández AI (2016) New polymorphism of the influenza virus resistance Mx1 gene in Iberian domestic pigs. Postdoc J 4:15–19
González-Vallinas M, Molina S, Vicente G, Zarza V, Martín-Hernández R, García-Risco MR, Fornari T, Reglero G, Ramírez de Molina A (2014) Expression of microRNA-15b and the glycosyltransferase GCNT3 correlates with antitumor efficacy of Rosemary diterpenes in colon and pancreatic cancer. PLoS One 9(6):e98556
Gröne HJ, Weber K, Helmchen U, Osborn M (1986) Villin—a marker of brush border differentiation and cellular origin in human renal cell carcinoma. Am J Pathol 124:294–302
Haeney MR, Culank LS, Montgomery RD, Sammons HG (1978) Evaluation of xylose absorption as measured in blood and urine: a one-hour blood xylose screening test in malabsorption. Gastroenterology 75:393–400
Haller O, Kochs G, Weber F (2007) Interferon, Mx, and viral countermeasures. Cytokine Growth Factor Rev 18(5–6):425–433
Hosoda N, Nishi M, Nakagawa M, Hiramatsu Y, Hioki K, Yamamoto M (1989) Structural and functional alterations in the gut of parenterally or enterally fed rats. J Surg Res 47:129–133
Hou YQ, Wang L, Ding BY, Liu YL, Zhu HL, Liu J, Li YT, Wu X, Yin YL, Wu G (2010) Dietary α-ketoglutarate supplementation ameliorates intestinal injury in lipopolysaccharide-challenged piglets. Amino Acids 39:555–564
Hou YQ, Wang L, Ding BY, Liu YL, Zhu HL, Liu J, Li YT, Kang P, Yin YL, Wu G (2011a) α-Ketoglutarate and intestinal function. Fron Biosci Landmark 16:1186–1196
Hou YQ, Yao K, Wang L, Ding BY, Fu DB, Liu YL, Zhu HL, Liu J, Li YT, Kang P, Yin YL, Wu G (2011b) Effects of α-ketoglutarate on energy status in the intestinal mucosa of weaned piglets chronically challenged with lipopolysaccharide. Br J Nutr 106:357–363
Hou YQ, Wang L, Zhang W, Yang ZG, Ding BY, Zhu HL, Liu YL, Qiu YS, Yin YL, Wu G (2012) Protective effects of N-acetylcysteine on intestinal functions of piglets challenged with lipopolysaccharide. Amino Acids 43:1233–1242
Hou YQ, Wang L, Yi D, Ding BY, Yang ZG, Li J, Chen X, Qiu YS, Wu G (2013) N-acetylcysteine reduces inflammation in the small intestine by regulating redox, EGF and TLR4 signaling. Amino Acids 45:513–522
Hou YQ, Wang L, Yi D, Ding BY, Chen X, Wang QJ, Zhu HL, Liu YL, Yin YL, Gong J, Wu G (2014) Dietary supplementation with tributyrin alleviates intestinal injury in piglets challenged with intrarectal administration of acetic acid. Br J Nutr 111:1748–1758
Hou YQ, Wang L, Yi D, Wu G (2015) N-acetylcysteine and intestinal health: a focus on its mechanism of action. Front Biosci 20:872–891
Iji PA, Saki A, Tivey DR (2001) Intestinal development and body growth of broiler chicks on diets supplemented with non-starch polysaccharides. Anim Feed Sci Technol 89:175–188
Jeurissen SHM, Lewis F, van der Klis JD, Mroz Z, Rebel JMJ, ter Huurne AAHM (2002) Parameters and techniques to determine intestinal health of poultry as constituted by immunity, integrity and functionality. Curr Issues Intest Microbiol 3:1–14
Jung K, Saif LJ (2015) Porcine epidemic diarrhea virus infection: etiology, epidemiology, pathogenesis and immunoprophylaxis. Vet J 204:134–143
Khurana S, George SP (2008) Regulation of cell structure and function by actin-binding proteins: villin’s perspective. FEBS Lett 582:2128–2139
Li JY, Yu Y, Hu S, Sun D, Yao YM (2002) Preventive effect of glutamine on intestinal barrier dysfunction induced by severe trauma. World J Gastroenterol 8:168–171
Li P, Kim SW, Li XL, Datta S, Pond WG, Wu G (2009) Dietary supplementation with cholesterol and docosahexaenoic acid affects concentrations of amino acids in tissues of young pigs. Amino Acids 37:709–716
Li W, van Kuppeveld FJ, He Q, Rottier PJ, Bosch BJ (2016) Cellular entry of the porcine epidemic diarrhea virus. Virus Res 226:117–127
Liu Y, Huang J, Hou Y, Zhu H, Zhao S, Ding B, Yin Y, Yi G, Shi J, Fan W (2008) Dietary arginine supplementation alleviates intestinal mucosal disruption induced by Escherichia coli lipopolysaccharide in weaned pigs. British J Nutrition 100(3):552–560
Liu Y, Chen F, Odle J, Lin X, Jacobi SK, Zhu H, Wu Z, Hou Y (2012) Fish oil enhances intestinal integrity and inhibits TLR4 and NOD2 signaling pathways in weaned pigs after LPS challenge. J Nutr 142(11):2017–2024
Lowry OH, Rosebrough NJ, Farr AL (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Luk GD, Bayless TM, Baylin SB (1980) Diamine oxidase (histaminase). A circulating marker for rat intestinal mucosal maturation and integrity. J Clin Invest 66:66–70
Luk GD, Bayless TM, Baylin SB (1983) Plasma postheparin diamine oxidase. Sensitive provocative test for quantitating length of acute intestinal mucosal injury in the rat. J Clin Invest 71:1308–1315
Luna LG (1968) Manual of histologic staining methods of the armed forces institute of pathology, 3rd edn. McGraw-Hill Book Company, New York, p 258
Lv C, Xiao Y, Li XD, Tian K (2016) Porcine epidemic diarrhea virus: current insights. Virus Adapt Treat 2016:1–12
Mansoori B, Nodeh H, Modirsanei M, Rahbari S, Aparnak P (2009) d-Xylose absorption test: a tool for the assessment of the effect of anticoccidials on the intestinal absorptive capacity of broilers during experimental coccidiosis. Anim Feed Sci Technol 148:301–308
Martín-Venegas R, Roig-Pérez S, Ferrer R, Moreno JJ (2006) Arachidonic acid cascade and epithelial barrier function during Caco-2 cell differentiation. J Lipid Res 47:1416–1423
Millea PJ (2009) N-acetylcysteine: multiple clinical applications. Am Fam Physician 80:265–269
Moll R, Robine S, Dudouet B, Louvard D (1987) Villin: a cytoskeletal protein and a differentiation marker expressed in some human adenocarcinomas. Virchows Arch B Cell Pathol Incl Mol Pathol 54:155–169
Montagne L, Pluske JR, Hampson DJ (2003) A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim Feed Sci Technol 108:95–117
Munro HN, Fleck A (1969) Analysis of tissues and body fluids for nitrogenous constituents. In: Munro HN (ed) Mammalian protein metabolism. Academic press, NY, pp 465–483
Nofrarías M, Manzanilla EG, Pujols J, Gibert X, Majó N, Segalés J, Gasa J (2006) Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J Anim Sci 84:2735–2742
Nygard AB, Jørgensen CB, Cirera S, Fredholm M (2007) Selection of reference genes for gene expression studies in pig tissues using SYBR greenqPCR. BMC Mol Biol 8:67
Pelsers MMAL, Hermens WT, Glatz JFC (2005) Fatty acid-binding proteins as plasma markers of tissue injury. Clin Chim Acta 352(1–2):15–35
Prasad AS, DeMouchelle E, Koniuch D (1972) A simple fluorimetric method for the determination of RNA and DNA in tissue. J Lab Clin Med 80:598–602
Radomska-Lesniewska DM, Skopinski P (2012) N-acetylcysteine as an antioxidant and anti-inflammatory drug and its some clinical applications. Central Eur J Immun 37:57–66
Sadowska AM (2012) N-acetylcysteine mucolysis in the management of chronic obstructive pulmonary disease. Ther Adv Respir Dis 6:127–135
Samuni Y, Goldstein S, Dean OM, Berk M (2013) The chemistry and biological activities of N-acetylcysteine. Biochim Biophys Acta 1830:4117–4129
Tan B, Li XG, Kong XF, Huang RL, Ruan Z, Yao K, Deng ZY, Xie MY, Shinzato I, Yin YL, Wu G (2009) Dietary l-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids 37:323–331
Thomas JT, Chen Q, Gauger PC, Giménez-Lirola LG, Sinha A, Harmon KM, Madson DM, Burrough ER, Magstadt DR, Salzbrenner HM, Welch MW, Yoon KJ, Zimmerman JJ, Zhang J (2015) Effect of porcine epidemic diarrhea virus infectious doses on infection outcomes in naïve conventional neonatal and weaned pigs. PLoS One 10:e0139266
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F (2002) Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 3:RESEARCH0034
Wang JJ, Chen LX, Li DF, Yin YL, Wang XQ, Li P, Dangott LJ, Hu WX, Wu G (2008a) Intrauterine growth restriction affects the proteomes of the small intestine, liver and skeletal muscle in newborn pigs. J Nutr 138:60–66
Wang Y, Srinivasan K, Siddiqui MR, George SP, Tomar A, Khurana S (2008b) A novel role for villin in intestinal epithelial cell survival and homeostasis. J Biol Chem 283:9454–9464
Wang XQ, Ou DY, Yin JD, Wu G, Wang JJ (2009) Proteomic analysis reveals altered expression of proteins related to glutathione metabolism and apoptosis in the small intestine of zinc oxide-supplemented piglets. Amino Acids 37:209–218
Wang QJ, Hou YQ, Yi D, Wang L, Ding BY, Chen X, Long MH, Liu YL, Wu G (2013) Protective effects of N-acetylcysteine on acetic acid-induced colitis in a porcine model. BMC Gastroenterol 13:133
Wang WW, Dai ZL, Wu ZL, Lin G, Jia SC, Hu SD, Dahanayaka S, Wu G (2014) Glycine is a nutritionally essential amino acid for maximal growth of milk-fed young pigs. Amino Acids 46:2037–2045
Wang L, Yi D, Hou YQ, Ding BY, Li K, Li BC, Zhu HL, Liu YL, Wu G (2016) Dietary supplementation with a-ketoglutarate activates mTOR signaling and enhances energy status in skeletal muscle of lipopolysaccharide-challenged piglets. J Nutr 146:1514–1520
West AB, Isaac CA, Carboni JM, Morrow JS, Mooseker MS, Barwick KW (1988) Localization of villin, a cytoskeletal protein specific to microvilli, in human ileum and colon and in colonic neoplasms. Gastroenterology 94:343–352
Wu G (2009) Amino acids: metabolism, functions, and nutrition. Amino Acids 37:1–17
Wu G (2014) Dietary requirements of synthesizable amino acids by animals: a paradigm shift in protein nutrition. J Anim Sci Biotechnol 5:34
Wu G, Meier SA, Knabe DA (1996) Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J Nutr 126:2578–2584
Wu G, Fang YZ, Yang S, Lupton JR, Turner ND (2004) Glutathione metabolism and its implications for health. J Nutr 134:489–492
Wu G, Wu ZL, Dai ZL, Yang Y, Wang WW, Liu C, Wang B, Wang JJ, Yin YL (2013) Dietary requirements of “nutritionally nonessential amino acids” by animals and humans. Amino Acids 44:1107–1113
Yan Q, Yang H, Yang D, Zhao B, Ouyang Z, Liu Z, Fan N, Ouyang H, Gu W, Lai L (2014) Production of transgenic pigs over-expressing the antiviral gene Mx1. Cell Regen (Lond) 3(1):11
Yi D, Hou YQ, Wang L, Ding BY, Yang ZG, Li J, Long MH, Liu YL, Wu G (2014) Dietary N-acetylcysteine supplementation alleviates liver injury in lipopolysaccharide-challenged piglets. Br J Nutr 111:46–54
Yi D, Hou YQ, Xiao H, Wang L, Zhang Y, Chen HB, Wu T, Ding BY, Hu CAA, Wu G (2017) N-Acetylcysteine improves intestinal function in lipopolysaccharides-challenged piglets through multiple signaling pathways. Amino Acids (Accepted)
Zafarullah M, Li WQ, Sylvester J, Ahmad M (2003) Molecular mechanisms of N-acetylcysteine actions. Cell Mol Life Sci 60:6–20
Acknowledgements
This research was jointly supported by the National Natural Science Foundation of China (31572416, 31372319, and 31402084), the Hubei Provincial Key Project for Scientific and Technical Innovation (2014ABA022), Hubei Provincial Research and Development Program (2010BB023), Natural Science Foundation of Hubei Province (2013CFA097, 2012FFB04805 and 2011CDA131), Research and Innovation Initiatives of WHPU (2016J08), the Hubei Hundred Talent Program, and Texas A&M AgriLife Research (H-8200).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics statement
All experimental procedures were approved by the Institutional Animal Care and Use Committee at Wuhan Polytechnic University.
Informed consent
All authors read and approved the final manuscript.
Additional information
Handling Editors: C.-A.A. Hu, Y. Yin, Y. Hou, G. Wu, Y. Teng.
Rights and permissions
About this article

Cite this article
Wang, L., Zhou, J., Hou, Y. et al. N-Acetylcysteine supplementation alleviates intestinal injury in piglets infected by porcine epidemic diarrhea virus. Amino Acids 49, 1931–1943 (2017). https://doi.org/10.1007/s00726-017-2397-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-017-2397-2