
Abstract
Background
Thinopyrum intermedium (2n = 6x = 42) is an important wild perennial Triticeae species exhibiting many potentially favorable traits for wheat improvement. Wheat-Th. intermedium partial amphiploids serve as a bridge to transfer desirable genes from Th. intermedium into common wheat.
Results
Three octoploid Trititrigia accessions (TE261–1, TE266–1, and TE346–1) with good resistances to stripe rust, powdery mildew and aphids were selected from hybrid progenies between Th. intermedium and the common wheat variety ‘Yannong 15’ (YN15). Genomic in situ hybridization (GISH), fluorescence in situ hybridization (FISH) and multicolor GISH (McGISH) analyses demonstrated that the three octoploid Trititrigia possess 42 wheat chromosomes and 14 Th. intermedium chromosomes. The 14 alien (Th. intermedium) chromosomes belong to a mixed genome consisting of J-, JS- and St-genome chromosomes rather than a single J, JS or St genome. Different types of chromosomal structural variation were also detected in the 1A, 6A, 6B, 2D and 7D chromosomes via FISH, McGISH and molecular marker analysis. The identity of the alien chromosomes and the variationes in the wheat chromosomes in the three Trititrigia octoploids were also different.
Conclusions
The wheat-Th. intermedium partial amphiploids possess 14 alien chromosomes which belong to a mixed genome consisting of J-, JS- and St- chromosomes, and 42 wheat chromosomes with different structural variations. These accessions could be used as genetic resources in wheat breeding for the transfer of disease and pest resistance genes from Th. intermedium to common wheat.
Similar content being viewed by others
Background
Thinopyrum intermedium (Host) Barkworth & D.R. Dewey [syn. Agropyron intermedium (Host) Beauvor and Elytrigia intermedia (Host) Nevski] (2n = 6x = 42; genome formula EeEeEbEbStSt, JJJSJSStSt, EEVVStSt or JrJrJvsJvsStSt) is considered as a segmental autoallohexaploid, and its genome constitution hanging in doubt is a research hot subject in Triticeae crop research. The presence of S genome (later changed to St by Wang et al. [1]) in Th. intermedium was first reported by Liu and Wang [2] and confirmed by Zhang et al. [3], leading to the genome formula JJJJStSt or EEEEStSt, where J genome is from Th. bessarabicum (2n = 14, JJ or EbEb), St genome is from Pseudoroegneria strigosa (2n = 14, StSt) and E genome is from Th. elongatum (2n = 14, EE or EeEe). Also by using genomic in situ hybridization, Chen et al. considered that the genome constitution of Th. intermedium is JJJSJSStSt where JS might be a modified J or E [4]. Ji et al. concluded that the genome constitution of Th. intermedium is EeEeEbEbStSt by using multicolor GISH (McGISH) [5]. Analyzing the chloroplast trnL-F sequence, granule-bound starch synthase I (GBSSI) and GISH, Mahelka et al. (2011) inferred that genomes of Th. intermedium are related to Ps. strigosa (StSt), Dasypyrum villosum (2n = 14, VV), and a complex origin from Th. elongatum (EE) and Aegilops tauschii (2n = 14, DD) [6]. JrJrJvsJvsStSt was proposed by Wang et al., who made use of EST-SSR markers to analyze the genome evolution of Th. intermedium, and Jr and Jvs refer to ancestral genomes of Je(E) and Jb(J), respectively [7].
Th. intermedium is described as one of the most important perennial Triticeae species, possessing many potentially favorable traits for wheat improvement [8,9,10,11]. Due to its crossability with common wheat, the transfer and utilization of Th. intermedium disease resistance genes, including the leaf rust resistance gene Lr38 [12]; the stem rust resistance gene Sr44 [13]; the stripe rust resistance gene Yr50 [14]; and the barley yellow dwarf resistance genes Bdv2 [15], Bdv3 [16] and Bdv4 [17, 18], have been accomplished.
Trititrigia octoploids were developed from hybrid progenies between Th. intermedium and common wheat and inherited numerous excellent traits from Th. intermedium, such as resistance to powdery mildew, leaf rust, stem rust and stripe rust [19, 20]. Furthermore, it is easier to successfully cross common wheat with Trititrigia octoploids than with Th. intermedium. Thus, Trititrigia octoploids are valuable germplasm to hybridize with common wheat, continually producing wheat-alien addition, substitution and translocation lines that can transfer beneficial traits from alien species into wheat [9, 21, 22].
Many Trititrigia octoploids have been produced in the past five decades and used as intermediary parents to facilitate the transfer of excellent genes from Th. intermedium to wheat. These Trititrigia octoploids include TAF46 [23], 78,829 [24], the ‘Zhong’ series of Trititrigia octoploids [25] and the TE series of Trititrigia octoploids [20, 21, 26,27,28]. The TE series of Trititrigia octoploids was developed from hybrid progenies between Th. intermedium and the common wheat variety ‘Yannong 15’ (YN15) and inherited many characteristics from Th. intermedium, such as large spikes, multi-florets, multi-tillers, and good resistance to stripe rust, powdery mildew and leaf rust. Thus, this TE series of Trititrigia octoploids represent an excellent genetic resource for wheat improvement [21, 26,27,28,29].
Genomic in situ hybridization (GISH) is generally used to distinguish alien chromosomes in the common wheat background [30, 31]. When Pseudoroegneria strigosa (2n = 2x = 14, StSt) genomic DNA was used as a probe in GISH analysis of Th. intermedium, the karyotype of Th. intermedium chromosomes could be classified into three groups. The first group is the St genome, completely labeled by probe signals, the second group is the JS genome labeled only in the centromere areas, and the third group is the J genome labeled only in the subtelomeric position [4, 32]. As GISH with the St-genome DNA probe could effectively distinguish the three sub-genome of Th. intermedium, it was widely used in the identification of Th. intermedium-derived wheat germplasms [19, 21, 28, 33].
Fluorescence in situ hybridization (FISH) and multicolor GISH (McGISH) are generally used to distinguish different chromosomes, analyze the chromosomal constitution and detect chromosomal variations in cytological studies [19, 20, 34, 35]. The available FISH probes that reveal fine banding patterns in wheat chromosomes include the Aegilops tauschii clone pAs1 [36, 37], the rye clone pSc119.2 [38], the GAA-satellite sequence [39], T. aestivum clone pTa535 [40] and many other oligonucleotides [41, 42]. Wheat chromosomes can be clearly distinguished using a combination of FISH probes including the pAs1 clone and the GAA-satellite sequence. Chromosome painting using this combination of FISH probes has been achieved in the common wheat cultivar ‘Chinese Spring’ (CS) [43, 44] in which all chromosomes of A, B and D subgenomes were labeled with specific signals, with the signals on the B- and D-genome chromosomes being particularly abundant. Through combined McGISH and FISH analysis, the positions and characteristics of chromosomal structural variations in such materials can be revealed.
In this study, the chromosomal constitution and chromosomal variations in three Trititrigia octoploids (TE261–1, TE266–1 and TE346–1) exhibiting good resistance to stripe rust, powdery mildew and aphids were investigated. The genetic characteristics, types of chromosome variation of the three Trititrigia octoploids were revealed using GISH, FISH, McGISH and molecular markers.
Results
Chromosomal constitution of the three Trititrigia octoploids
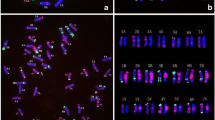
The root tip chromosome analysis showed that chromosome number of TE261–1, TE266–1 and TE346–1 was 2n = 56. Observation of meiotic chromosomes in PMCs revealed that most chromosomes in the observed cells formed 28 bivalents at meiotic metaphase I, 14 alien chromosomes formed 7 bivalents were observed in GISH analysis, indicating high cytological stability (Supplemental Fig. 1, Supplemental Table 1).
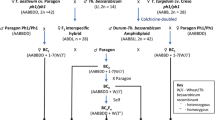
GISH, FISH and McGISH were used to identify the chromosomal constitution of TE261–1, TE266–1 and TE346–1. The GISH (Fig. 1-A1) and FISH (Fig. 1-A2) results indicated that TE261–1 contained 42 wheat chromosomes and 14 Th. intermedium chromosomes (Fig. 2), including one pair of St-genome chromosomes that were completely labeled with probe signals, one pair of J-genome chromosomes labeled only in the telomeres, three pairs of JS-genome chromosomes with obvious labeling in centromere areas, one pair of acrocentric chromosomes from the JS genome, and one pair of J-St translocated chromosomes. The JS acrocentric chromosomes (Fig. 2, Supplemental Fig. 1-A2), were first discovered in the TE series of Trititrigia octoploids.

GISH, FISH and McGISH patterns of TE261–1, TE266–1, TE346–1 and YN15. GISH patterns of TE261–1 (A1), TE266–1 (B1) and TE346–1 (C1): St (Ps. strigosa)-genome DNA labeled with Texas-Red-5-dCTP was used as the probe, and YN15 genome DNA was employed as the block. FISH patterns of TE261–1 (A2), TE266–1 (B2), TE346–1 (C2) and YN15 (D1): red signals represent (GAA)8 labeled with 5′ TAMRA, and green signals represent pAs1 labeled with fluorescein-12-dUTP. McGISH patterns of TE261–1 (A3), TE266–1 (B3), TE346–1 (C3) and YN15 (D2): A genomes are labeled with fluorescein-12-dUTP; D genomes are labeled with Texas-Red-5-dCTP; and B-genome DNA (gray) is used for blocking. A-D (7D) translocation are indicated by red asterisks in McGISH patterns (A3, B3 and C3); A-D (2A) translocation are indicated by yellow asterisks in TE266–1 (B3); and 2A, 2D and 7D are marked in the McGISH patterns of YN15 (D2)

Alien chromosome GISH patterns of the three Trititrigia octoploids. St-genome DNA labeled with Texas-Red-5-dCTP was used as the probe, and YN15 genome DNA was employed as the block, JS-a represents acrocentric chromosomes from the JS genome
The results revealed that TE266–1 also contained 42 wheat chromosomes and 14 Th. intermedium chromosomes (Fig. 1-B1, B2, B3 and Fig. 2), including two pairs of St-genome chromosomes, one pair of J-genome chromosomes, three pairs of JS-genome chromosomes, and one pair of J-St translocated chromosomes. As shown in Fig. 1-C1, C2, C3 and Fig. 2, TE346–1 also contained 42 wheat chromosomes and 14 Th. intermedium chromosomes, including one pair of St-genome chromosomes, one pair of J-genome chromosomes, four pairs of JS-genome chromosomes and one pair of J-St translocated chromosomes.
Variation in the wheat chromosomes in the three Trititrigia octoploids
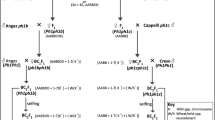
FISH analysis were used to identify different wheat chromosomes of TE261–1, TE266–1, TE346–1 and YN15 while using labled (GAA)8 (red signals) and pAs1 (green signals) as probes. McGISH analysis were also conducted while using labeled A (green signals) and D (red signals) genomes as probes, B-genome DNA (gray) is used for blocking. The wheat chromosome variations in the three Trititrigia octoploids compared with their common wheat parent YN15 based on FISH and McGISH patterns. For the A-genome chromosomes, the hybridization signals of pAs1 are not visible on 1A in TE261–1, TE266–1 and TE346–1 (Fig. 3D). An absence of (GAA)8 signals on 6AL and additional pAs1 signals at the 6AS subtelomeric position were observed in TE261–1 and TE346–1, while only the absence of (GAA)8 signals on 6AL was observed in TE266–1 (Fig. 3D).

FISH patterns for comparison of YN15 with the three Trititrigia octoploids. In the FISH patterns comparing YN15 with TE261–1 (A), TE266–1 (B), and TE346–1 (C), the results for one pair of chromosomes for YN15 are on the left, and those for the Trititrigia octoploids are on the right. D: FISH and McGISH patterns for the comparison of YN15 with three Trititrigia octoploids on various chromosomes. The patterns in a, b, c and d indicate the corresponding chromosomes of YN15, TE261–1, TE266–1 and TE346–1, respectively. The lower right part of Fig. D shows the McGISH patterns of chromosome 2D and 7D in YN15 and three Trititrigia octoploids
Compared with YN15, the signals on the 6B chromosome in TE261–1, TE266–1 and TE346–1 also changed (Fig. 3A-D). Interspersed green signals for the pAs1 probe were detected on 6BS of TE261–1 and TE266–1, and the length of the 6BS chromosomes appeared to be extended in TE266–1 and TE346–1. Faint red (GAA)8 signals were also observed on the 6BL chromosomes in the three Trititrigia octoploids.
The signals of the D-genome chromosomes in TE261–1, TE266–1 and TE346–1 appeared to vary significantly. FISH and McGISH results indicated that in TE266–1, the 2D chromosomes were replaced with one pair of A-D translocated chromosomes; the main body of the A-D translocated chromosomes was derived from 2A chromosomes according to the pAs1 signals, while part of the long arm was replaced with a 2D chromosome segment (Fig. 3-B, 3-D).
The FISH results showed that the original strong pAs1 signals on 7DS or 7DL in YN15 were replaced with faint (GAA)8 and pAs1 signals in TE261–1, TE266–1 and TE346–1. The McGISH results indicated that the partial long arm of the 7D chromosome in TE261–1 was replaced with the chromosome segment of the A-genome chromosome (Fig. 1-A3, Fig. 3-D). However, the translocated A-genome chromosome segment was located on the short arm in TE266–1 and TE346–1, not the long arm as in TE261–1. The source of the A-genome chromosome segment was uncertain due to the lack of a specific FISH signal in this A-genome chromosome segment.
Specific molecular markers of the wheat chromosomes were employed to verify the chromosomal variation and detect the variation type in the three Trititrigia octoploids. The specific band (~ 300 bp) of YN15 amplified with the specific marker Xmag3124, which was located on 1A, was not observed in the three Trititrigia octoploids (Fig. 4 and Supplemental Fig. 4). The specific band (~ 150 bp) of YN15 detected by marker GPW4344, which was located on 6A (Supplemental Fig. 2 and Supplemental Fig. 4), were also absent in the three Trititrigia octoploids, and a 6A specific band (~ 300 bp) absent in TE346–1 were detected by GPW7465. The 6B specific band (~ 200 bp) detected by the marker Xgwm219 showed a 10 bp insertion in three Trititrigia octoploids (Supplemental Fig. 2 and Supplemental Fig. 4). The results for several specific markers, such as Xgdm77, Xgdm35, Barc095, Ppd-D1, and Xwmc245, located on the 2D chromosomes (Fig. 4, Supplemental Fig. 2 and Supplemental Fig. 4) demonstrated great variation, i.e., the absent of specific bands in the 2D chromosomes in TE266–1, but the 2D chromosomes of TE261–1 and TE346–1 remained unchanged. Two specific bands (240, 200 bp) absent in three Trititrigia octoploids were detected by Barc172 and Barc053 located on the 7D chromosomes, (Fig. 4, Supplemental Fig. 2 and Supplemental Fig. 4), respectively.

Results of amplification of the wheat chromosome-specific markers Xmag3124 (1A), Ppd-D1 (2D) and BARC172 (7D). M: Marker DL2000, A: CS, N1: CS Nulli-tetrasomes N1AT1D, N2: N2DT2B, N3: N7DT7B, B: YN15, C: Th. intermedium, 1: TE261–1, 2: TE266–1, 3: TE346–1
The sequences of the primers used for these 16 chromosome-specific markers were employed to run local BLAST searches against the database of the T. aestivum (Chinese Spring, CS) whole-genome sequence. Eleven markers were screened according to screening criteria including the appropriate chromosomes, initial position, terminal position and degree of strict matching (Supplemental Table 2). For example, the chromosomes mapped by BLAST should be matched the specific chromosomes detected by PCR; the higher matching degree means the better option, and the 3′ of primer must be matched; the absolute value of margin between forward primer initial position and reverse primer terminal, should match the size of specific bands; and last, the appropriate position mapped by the markers were picked out. Compared BLAST with PCR results after screening, the size of specific bands amplified with majority markers were similar (Fig. 4), but the 7D specific band (~ 240 bp) detected by Barc053 was different from BLAST result (295 bp), and difference exist in Ppd-D1 (R1) result of PCR (250 bp) and BLAST (413 bp). The difference could be attributed to the sequence diversity between CS and YN15. The screened markers were then labeled at the specific location of the corresponding chromosomes using MapGene2Chromosomes v2, according to their physical position on the chromosomes (Supplemental Fig. 3). The positions of the variations detected with markers were consistent with the FISH signals on the 6A, 6BL, 2D and 7D chromosomes, while the variation at the 1AL telomere was detected with markers, rather than FISH signals.
Phenotypic evaluation of the three Trititrigia octoploids
The reactions to stripe rust, powdery mildew and aphids in TE261–1, TE266–1 and TE346–1 were evaluated at the seedling and adult plant stages using the wheat parents YN15 and Th. intermedium as controls (Table 1). At the seedling stage, TE261–1, TE266–1, and TE346–1 were immune to the stripe rust race CYR32 and resistant to the powdery mildew race E09. At the adult stage, all three Trititrigia octoploids showed good resistance to stripe rust and powdery mildew in the field. TE261–1 and TE346–1 also showed moderate resistance to aphids at the adult stage, while TE266–1 and YN15 were susceptible. As the common wheat parent YN15 is susceptible to stripe rust, powdery mildew and aphids, whereas Th. intermedium is immune to all three, we deduce that the resistance of Trititrigia is derived from Th. intermedium.
Discussion
Th. intermedium is a valuable perennial species of Triticeae for wheat improvement due to its resistance to a number of wheat diseases and pests as well as its stress tolerance and high crossability with various Triticum species [8,9,10,11, 45]. Partial wheat-Th. intermedium amphiploids showing high cross-compatibility with wheat are desirable ‘bridge’ materials for transferring desirable genes from Th. intermedium to common wheat [19, 20].
Chromosome counting in metaphase spreads revealed that the chromosome number of these three octoploids of Trititrigia was consistently 2n = 56. Molecular cytogenetic analysis then revealed that the three Trititrigia accessions all carry 42 wheat chromosomes and 14 Th. intermedium chromosomes and that the 14 alien (Th. intermedium) chromosomes are composed of a mixed genome consisting of J-, JS- and St-genome chromosomes rather than a single J, JS or St genome, similar to other reported of Trititrigia octoploids [21, 46, 47]. The mechanism underlying the formation of the mixed genome is uncertain, and with development of new techniques and methods, such as oligo-FISH painting system [48] further research will contribute to elucidating the homologous relationship and integrality between the J, JS and St genomes and the origin and evolution of Th. intermedium.
Two types of structural variation in the alien chromosomes were detected: J-St translocated chromosomes, which were present in all three Trititrigia octoploids, and JS acrocentric chromosomes in TE261–1 (Fig. 2), which might have formed via the centromere breakage of one pair of JS-genome chromosomes. J-St translocated chromosomes also exist in many TE series of Trititrigia octoploids [21, 26, 28], but JS acrocentric chromosomes were first discovered in the TE series of Trititrigia octoploids. The formation of these two types of variations might occur through chromosome breakage and fusion when F1 hybrids obtained through distant hybridization are backcrossed with hexaploid wheat [34].
Although the alien chromosomal constitution of three Trititrigia octoploids was different, some of the alien chromosomes were probably identical according to the GISH and FISH signals. For example, the J-genome chromosomes with the satellite sequence and the J-St translocated chromosomes in these three Trititrigia octoploids were similar. The St chromosomes of TE261–1 were also similar to the St-1 chromosomes of TE266–1, and the St chromosomes of TE346–1 were similar to the St-2 chromosomes of TE266–1 (Fig. 2).
In addition to the structural variation in the alien chromosomes, structural variations, such as the variations detected in the 1A, 6A, 6B, 2D and 7D chromosomes, occurred in the wheat chromosomes as well. The results indicated that during the formation of partial amphiploids, the wheat chromosomes underwent various types of chromosomal recombination due to chromosome segments introgressed from Th. intermedium into the common wheat chromosomes, although the introgressed segments were too small to detect via GISH. The structural variations may have been generated via recombination between different wheat chromosomes, such as homoeologous chromosomal recombination between A-, B-, and D-genome chromosomes, influenced by the Th. intermedium chromosomes. The variation in chromosomal patterns of repetitive sequences (e.g. chromosome 6B) could be also caused by repetitive sequence reorganization (amplification/loss). After hybridization, genomes can undergo genetic and epigenetic changes (genomic shock) which can reorganize their structure.
Similar chromosomal variations were detected in the 1A and 6A chromosomes of the three Trititrigia and might have been derived from the same origin in the earlier generations of backcrosses. Additionally, there were distinct variations in the 6B, 2D and 7D chromosomes of these Trititrigia octoploids, such as pAs1 signals at different positions of 6BS in TE266–1 and TE346–1 and the 2A-2D translocation in TE266–1. The variations in the 7D chromosomes of the three Trititrigia octoploids were also different. McGISH indicated that the partial 7DS chromosomes of TE266–1 and TE346–1 were replaced with an A-genome chromosome segment, but the corresponding position in TE261–1 was on the 7DL chromosome. The original position replaced in 7DL or 7DS was difficult to discern via FISH because the signals on the two arms of the 7D chromosomes were similar. The results of molecular marker analyses also supported the results of FISH and McGISH in TE261–1. Additional work will be needed to obtain strong evidence indicating how these chromosomal recombinations occurred as well as the effect of these chromosomal variations.
The drastic and abundant chromosome variations were detected in these three Trititrigia octoploids, a kind of generic hybrid between wheat and Th. intermedium. However, we guess the more drastic variations might exist in early-generation of Trititrigia octoploids, and then backcrosses with YN15 resulted in reducing variability and increasing stability. We hypothesized that Th. intermedium, as a natural generic hybrid, possessed drastic chromosomal variation causing difficulty to confirm the progenitor diploid species.
Although FISH results using general probes consisting of (GAA)8 and pAs1 were more abundant, colorful and characteristic to distinguish most common wheat chromosomes, it was still insufficient to identify the alien chromosomes in present amphiploids. Li et al. had reported an efficient Oligo-FISH painting system for revealing chromosome rearrangements and polyploidization in Triticeae [48], while using novel comparative genome-based oligo painting FISH procedure. These would be beneficial and helpful for determining the alien Thinopyrum chromosomes in these amphiploids in the future studies.
Phenotypic evaluation indicated the successful transfer of stripe rust and powdery mildew resistance from Th. intermedium to these three Trititrigia octoploids and that TE261–1 also exhibited moderate resistance to aphids. In addition, the chromosomal constitution of TE261–1, TE266–1 and TE346–1 is different from that of other TE series of Trititrigia octoploids reported previously [21, 26, 28]. Therefore, these accessions could be employed as bridge parents for crossing with wheat to develop addition, substitution or translocation lines to generate new rust-, powdery mildew- and aphid-resistant germplasms for wheat breeding.
Conclusions
Three octoploid Trititrigia accessions (TE261–1, TE266–1, and TE346–1) with good resistances to diseases and aphids were characterized by GISH, FISH, McGISH and molecular marker analyses. Molecular cytogenetic analysis revealed that these Trititrigia accessions all carry 42 wheat chromosomes and 14 Th. intermedium chromosomes, and the 14 alien (Th. intermedium) chromosomes are composed of a mixed genome consisting of J-, JS- and St-genome chromosomes rather than a single J, JS or St genome. Different structural variations occurred in the wheat chromosomes, such as the variations detected in the 1A, 6A, 6B, 2D and 7D chromosomes as well. Results indicated that the wheat-Th. intermedium partial amphiploids possess 14 alien chromosomes which belong to a mixed genome consisting of J-, JS- and St- chromosomes, and 42 wheat chromosomes with different structural variations.
Methods
Plant materials
Three Trititrigia octoploid accessions (TE261–1, TE266–1, and TE346–1) were selected from BC1F8 of common wheat Yannong15 crossed with Th. intermedium, and selfed for some generations. Th. intermedium from the original accession at the Northwest Institute of Botany, Chinese Academy of Sciences, was provided by the academician Li ZS; Pseudoroegneria strigose, Triticum urartu Thum. ex Gandil. (2n = 2x = 14, AA), Aegilops speltoides Tausch. (2n = 2x = 14, BB), Aegilops tauschii Coss. (2n = 2x = 14, DD) and the CS nulli-tetrasomic accessions N1AT1B, N6AT6B, N2DT2A, N7DT7A were provided by the researcher Li LH from the Institute of Crop Sciences, Chinese Academy of Agricultural Sciences.
DNA extraction, PCR amplification and marker analysis
The CTAB method was used to extract total genomic DNA from the tender leaves [49] of Pseudoroegneria strigosa (St genome), Triticum urartu (A genome), Aegilops speltoides (B genome) and Aegilops tauschii (D genome). In total, 176 pairs of SSR primers [50, 51] (https://wheat.pw.usda.gov) were used to identify chromosomal variations. PCR amplification was performed according to Beales et al. [52], and marker screening was conducted by means of polyacrylamide gel electrophoresis (PAGE). The selected markers were analyzed with BLAST (Basic Local Alignment Search Tool) based on the database containing the Triticum aestivum (CS) whole genome sequence (IWGSC RefSeq v1.0, https://wheat-urgi.versailles.inra.fr/morgoth/Seq-Repository/BLAST) and then labeled on the corresponding chromosomes using MapGene2Chromosomes v2 (http://mg2c.iask.in/mg2c_v2.0/).
Chromosome slide preparation, meiotic preparation, GISH, FISH and McGISH
Fresh root tips were collected from germinating seeds treated with nitrous oxide (N2O) for 2 h [53] and then immersed in 90% glacial acetic acid to fix cell division. The preparation of root tip cell chromosome slides and analysis was performed referring to the method of Han [46]. When the flag leaf of wheat was spread, young spikes were sampled and anthers at metaphase I (MI) of meiosis were fixed in Carnoy’s solution. The meiotic chromosomes were prepared from pollen mother cells (PMCs) by knocking and pressing anthers in 45% acetic acid, after which the chromosome configuration in PMC metaphase I (MI) was observed.
St-genome DNA and D-genome DNA were labeled with Texas-Red-5-dCTP. The clone pAs1 (GenBank: D30736.1) is a repetitive DNA sequence (1015 bp) from Aegilops tauschii [37, 54]. The pAsl clone and A-genome DNA were labeled with fluorescein-12-dUTP using the nick translation method. Oligonucleotides (GAA)8 with 5′ TAMRA were synthesized by Sangon Biotech (Shanghai, China). The procedures for GISH, FISH and McGISH; signal detection; and image collection were done according to Han et al. [55] and Cui et al. [56].
The reaction volume for in situ hybridization was 10 μL per slide. GISH was conducted using a probe of 50 ng St-genome DNA, with a block of 6000 ng YN15 DNA; FISH was performed with 5 ng (GAA)8 and 200 ng pAs1; and McGISH was carried out with probes of 50 ng A-genome DNA and 100 ng D-genome DNA and a block of 8000 ng B-genome DNA diluted to the desired total volume with ddH2O. The in situ hybridization program consisted of denaturation at 95 °C for 5 min and hybridization at 55 °C for 6 h. The fluorescent signals of FITC (Excitation Filter EX465–495, Dichroic Mirror DM505, Barrier Filter BA515–555), TRITC (Excitation Filter EX540/25, Dichroic Mirror DM565, Barrier Filter BA605/55) and DAPI (Excitation Filter EX340–380, Dichroic Mirror DM400, Barrier Filter BA435–485) were detected, and images were collected using a NIKON eclipse Ni-U fluorescence microscope; the images were processed using NIS-Elements BR 4.00.12 software. After rinsing with 2X SSC solution, the slides were used for in situ hybridization again.
Evaluation of resistance to powdery mildew, stripe rust and aphids
The powdery mildew and stripe rust resistance of plants in the seedling stage were tested in a plant incubator. The wheat variety Huixianhong was employed for inoculation of the Blumeria graminis f. sp. tritici (Bgt) pathotype E09 and the Puccinia striiformis f. sp. tritici (Pst) pathotype CYR32. Thirty seeds of the test materials were grown in seedling plates and placed in a plant incubator under 12 h light (25,000 lx) at 25 °C and 12 h of darkness at 17 °C, with 70% relative humidity. Huixianhong and YN15 were used as controls. Inoculation with E09 and CYR32 was achieved by smearing at the one-leaf stage, and the gradient of infection was investigated when Huixianhong had been completely infected; these experiments were repeated three times. The powdery mildew and stripe rust resistances of adult-stage plants were tested via natural infection in an open field. The resistance evaluation criteria for powdery mildew and stripe rust followed the methods of Sheng [57] and Li et al. [58]. In the scale of powdery mildew resistance, 0 is immune; 0; is nearly immune with necrotic flecks; 1 is highly resistant with small lesions (< 1 mm) and thin mycelium; 2 is moderately resistant with small lesions (< 1 mm) and a slightly thick mycelium; 3 is moderately susceptible with lesions (> 1 mm) and a thick mycelium; and 4 is highly susceptible with lesions (> 1 mm) and a consecutive thick mycelium. In the scale of stripe rust resistance, 0 is immune; 0; is nearly immune with tiny flecks; 1 is highly resistant with yellowish-white flecks; 2 is moderately resistant with necrotic plaques around small spore stripes; 3 is moderately susceptible with fading around spore stripes; and 4 is highly susceptible with large spore stripes.
For the identification of aphid resistance, natural aphids were employed, and the evaluation was performed during the wheat grain-filling stage. The test materials were sown on two 1.4-m-long rows with YN15 as CK, Th. intermedium planting in pot were put nearby to evaluate the aphid resistance. The investigation of aphid injury and the classification criteria for aphid resistance followed the method of Liu et al. [59]. The number of aphids on 10 seriously aphid-damaged stems was determined for all materials; then, using this number as the numerator and the average number of aphids on YN15 as the denominator, the ratio was computed as the evaluation criterion: 0 is immune with no aphids; 1 is highly resistant with a ratio between 0.01 and 0.30; 2 is moderately resistant with a ratio between 0.31 and 0.6; 3 is slightly resistant with a ratio between 0.61 and 0.90; 4 is slightly susceptible with a ratio between 0.91 and 1.2; 5 is moderately susceptible with a ratio between 1.21 and 1.5; and 6 is highly susceptible, with a ratio > 1.5.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- CS:
-
Chinese Spring
- FISH:
-
Fluorescence in situ hybridization
- GISH:
-
Genomic in situ hybridization
- McGISH:
-
Multicolor GISH
- YN15:
-
Yannong 15
References
Wang R, Von Bothmer R, Dvorak J, Fedak G, Linde-Laursen I, Muramatsu M. Genome symbols in the Triticeae (Poaceae). Herbarium Publications. 1994;2:29–34. https://digitalcommons.usu.edu/herbarium_pubs/20/.
Liu ZW, Wang RR. Genome analysis of Elytrigia caespitosa, Lophopyrum nodosum, Pseudoroegneria geniculata ssp. scythica, and Thinopyrum intermedium (Triticeae: Gramineae). Genome. 1993;36(1):102–11.
Wang RR, Zhang XY. Characterization of the translocated chromosome using fluorescence in situ hybridization and random amplified polymorphic DNA on two Triticum aestivum- Thinopyrum intermedium translocation lines resistant to wheat streak mosaic or barley yellow dwarf virus. Chromosom Res. 1996;4(8):583–7.
Chen Q, Conner RL, Laroche A, Thomas JB. Genome analysis of Thinopyrum intermedium and Thinopyrum ponticum using genomic in situ hybridization. Genome. 1998;41(4):580–6.
Wanquan J, Xiuzhuang X, Qiuying W, Changyou W, Fedak G, Guangtian L. GISH analysis of Thinopyrum intermedium. Acta Botan Boreali-Occiden Sin. 2001;21(3):401–5.
Mahelka V, Kopecký D, Paštová L. On the genome constitution and evolution of intermediate wheatgrass (Thinopyrum intermedium: Poaceae, Triticeae). BMC Evol Biol. 2011;11(1):127.
Wang RR, Larson SR, Jensen KB, Bushman BS, DeHaan LR, Wang S, Yan X. Genome evolution of intermediate wheatgrass as revealed by EST-SSR markers developed from its three progenitor diploid species. Genome. 2015;58(2):63–70.
Gupta P: Use of alien genetic variation for wheat improvement. In: Molecular breeding for sustainable crop improvement. Berlin: Springer; 2016:1–30.
Han FP, Fedak G, Benabdelmouna A, Armstrong K, Ouellet T. Characterization of six wheat × Thinopyrum intermedium derivatives by GISH, RFLP, and multicolor GISH. Genome. 2003;46(3):490–5.
Li H, Wang X. Thinopyrum ponticum and Th. intermedium: the promising source of resistance to fungal and viral diseases of wheat. J Genet Genomics. 2009;36(9):557–65.
Wang RR-C: Agropyron and psathyrostachys. In: Wild crop relatives: genomic and breeding resources. Berlin: Springer; 2011:77–108.
Friebe B, Zeller FJ, Mukai Y, Forster BP, Bartos P, McIntosh RA. Characterization of rust-resistant wheat-Agropyron intermedium derivatives by C-banding, in situ hybridization and isozyme analysis. Theor Appl Genet. 1992;83(6–7):775–82.
Friebe B, Jiang J, Raupp WJ, McIntosh RA, Gill BS. Characterization of wheat-alien translocations conferring resistance to diseases and pests: current status. Euphytica. 1996;91(1):59–87.
Liu J, Chang Z, Zhang X, Yang Z, Li X, Jia J, Zhan H, Guo H, Wang J. Putative Thinopyrum intermedium-derived stripe rust resistance gene Yr50 maps on wheat chromosome arm 4BL. Theor Appl Genet. 2013;126(1):265–74.
Banks PM, Larkin PJ, Bariana HS, Lagudah ES, Appels R, Waterhouse PM, Brettell RI, Chen X, Xu HJ, Xin ZY, et al. The use of cell culture for subchromosomal introgressions of barley yellow dwarf virus resistance from Thinopyrum intermedium to wheat. Genome. 1995;38(2):395–405.
Sharma H, Ohm H, Goulart L, Lister R, Appels R, Benlhabib O. Introgression and characterization of barley yellow dwarf virus resistance from Thinopyrum intermedium into wheat. Genome. 1995;38(2):406–13.
Ayala-Navarrete L, Tourton E, Mechanicos AA, Larkin PJ. Comparison of Thinopyrum intermedium derivatives carrying barley yellow dwarf virus resistance in wheat. Genome. 2009;52(6):537–46.
Lin ZS, Huang DH, Du LP, Ye XG, Xin ZY. Identification of wheat-Thinopyrum intermedium 2Ai-2 ditelosomic addition and substitution lines with resistance to barley yellow dwarf virus. Plant Breed. 2006;125(2):114–9.
Kruppa K, Turkosi E, Mayer M, Toth V, Vida G, Szakacs E, Molnar-Lang M. McGISH identification and phenotypic description of leaf rust and yellow rust resistant partial amphiploids originating from a wheat x Thinopyrum synthetic hybrid cross. J Appl Genet. 2016;57(4):427–37.
Wang Y, Wang H. Characterization of three novel wheat-Thinopyrum intermedium addition lines with novel storage protein subunits and resistance to both powdery mildew and stripe rust. J Genet Genomics. 2016;43(1):45–8.
Bao Y, Wu X, Zhang C, Li X, He F, Qi X, Wang H. Chromosomal constitutions and reactions to powdery mildew and stripe rust of four novel wheat-Thinopyrum intermedium partial amphiploids. J Genet Genomics. 2014;41(12):663–6.
Tang S, Li Z, Jia X, Larkin P. Genomic in situ hybridization (GISH) analyses of Thinopyrum intermedium, its partial amphiploid Zhong 5, and disease-resistant derivatives in wheat. Theor Appl Genet. 2000;100(3–4):344–52.
Brettell RIS, Banks PM, Cauderon Y, Chen X, Cheng ZM, Larkin PJ, Waterhouse PM. A single wheatgrass chromosome reduces the concentration of barley yellow dwarf virus in wheat. Ann Appl Biol. 1988;113(3):599–603.
Zhang XY, Koul A, Petroski R, Ouellet T, Fedak G, Dong YS, Nwang RR. Molecular verification and characterization of BYDV-resistant germ plasms derived from hybrids of wheat with Thinopyrum ponticum and Th. intermedium. Theor Appl Genet. 1996;93(7):1033–9.
Sun S. Research on Triticum Agropyron Form and species formation. Sci Agric Sin(in Chinese). 1980;6:1–9.
Bao Y, Li X, Liu S, Cui F, Wang H. Molecular cytogenetic characterization of a new wheat-Thinopyrum intermedium partial amphiploid resistant to powdery mildew and stripe rust. Cytogenet Genome Res. 2009;126(4):390–5.
Liu S-B, Wang H-G, Zhang X-Y, Li X-F, Li D-Y, Duan X-Y, Zhou Y-L. Molecular cytogenetic identification of a wheat-Thinopyron intermedium(host) Barkworth & DR Dewey partial Amphiploid resistant to powdery mildew. J Integr Plant Biol. 2005;47(6):726–33.
Qi X, Bao Y, Li X, Qian Z, Wang R, Wu K, Wang H. Cytological identification and chromosome constitution analyses of ten octoploid Trititrigia accessions. Acta Agron Sin. 2017;43(7):967–73.
Wang H, Liu S, Li X, Gao J. Feng D-s, Chen D: breeding and identification of six octoploid trititrigia. J Triticeae Crops. 2006;26(4):6–10.
Cai X, Jones SS, Murray TD. Molecular cytogenetic characterization of Thinopyrum genomes conferring perennial growth habit in wheat-Thinopyrum amphiploids. Plant Breed. 2001;120(1):21–6.
Le HT, Armstrong K, Miki B. Detection of rye DNA in wheat-rye hybrids and wheat translocation stocks using total genomic DNA as a probe. Plant Mol Biol Report. 1989;7(2):150–8.
Chen Q, Friebe B, Conner R, Laroche A, Thomas J, Gill B. Molecular cytogenetic characterization of Thinopyrum intermedium-derived wheat germplasm specifying resistance to wheat streak mosaic virus. Theor Appl Genet. 1998;96(1):1–7.
Chen Q, Conner RL, Li HJ, Sun SC, Ahmad F, Laroche A, Graf RJ. Molecular cytogenetic discrimination and reaction to wheat streak mosaic virus and the wheat curl mite in Zhong series of wheat--Thinopyrum intermedium partial amphiploids. Genome. 2003;46(1):135–45.
Guo X, Shi Q, Wang J, Hou Y, Wang Y, Han F. Characterization and genome changes of new amphiploids from wheat wide hybridization. J Genet Genomics. 2015;42(8):459–61.
Zheng Q, Luo Q, Niu Z, Li H, Li B, Xu SS, Li Z. Variation in chromosome constitution of the Xiaoyan series partial Amphiploids and its relationship to stripe rust and stem rust resistance. J Genet Genomics. 2015;42(11):657–60.
Rayburn AL, Gill BS. Molecular identification of the D-genome chromosomes of wheat. J Hered. 1986;77(4):253–5.
Rayburn AL, Gill BS. Isolation of a D-genome specific repeated DNA sequence from Aegilops squarrosa. Plant Mol Biol Report. 1986;4(2):102–9.
Mukai Y, Nakahara Y, Yamamoto M. Simultaneous discrimination of the three genomes in hexaploid wheat by multicolor fluorescence in situ hybridization using total genomic and highly repeated DNA probes. Genome. 1993;36(3):489–94.
Pedersen C, Rasmussen SK, Linde-Laursen I. Genome and chromosome identification in cultivated barley and related species of the Triticeae (Poaceae) by in situ hybridization with the GAA-satellite sequence. Genome. 1996;39(1):93–104.
Komuro S, Endo R, Shikata K, Kato A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome. 2013;56(3):131–7.
Du P, Zhuang L, Wang Y, Yuan L, Wang Q, Wang D. Dawadondup, tan L, Shen J, Xu H et al: development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes. Genome. 2017;60(2):93–103.
Tang Z, Yang Z, Fu S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J Appl Genet. 2014;55(3):313–8.
Cabrera A, Martin A, Barro F. In-situ comparative mapping (ISCM) of Glu-1 loci in Triticum and Hordeum. Chromosom Res. 2002;10(1):49–54.
Pedersen C, Langridge P. Identification of the entire chromosome complement of bread wheat by two-colour FISH. Genome. 1997;40(5):589–93.
Jiang J, Friebe B, Gill BS. Recent advances in alien gene transfer in wheat. Euphytica. 1994;73(3):199–212.
Han F, Liu B, Fedak G, Liu Z. Genomic constitution and variation in five partial amphiploids of wheat--Thinopyrum intermedium as revealed by GISH, multicolor GISH and seed storage protein analysis. Theor Appl Genet. 2004;109(5):1070–6.
Yu Z, Wang H, Xu Y, Li Y, Lang T, Yang Z, Li G. Characterization of chromosomal rearrangement in new wheat—Thinopyrum intermedium addition lines carrying Thinopyrum—specific grain hardness genes. Agronomy. 2019;9:18.
Li G, Zhang T, Yu Z, Wang H, Yang E, Yang Z. An efficient Oligo-FISH painting system for revealing chromosome rearrangements and polyploidization in Triticeae. Plant J. 2020. https://doi.org/10.1111/tpj.15081.
Li G, Quiros CF. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in brassica. Theor Appl Genet. 2001;103(2–3):455–61.
Roder MS, Korzun V, Wendehake K, Plaschke J, Tixier MH, Leroy P, Ganal MW. A microsatellite map of wheat. Genetics. 1998;149(4):2007–23.
Song QJ, Shi JR, Singh S, Fickus EW, Costa JM, Lewis J, Gill BS, Ward R, Cregan PB. Development and mapping of microsatellite (SSR) markers in wheat. Theor Appl Genet. 2005;110(3):550–60.
Beales J, Turner A, Griffiths S, Snape JW, Laurie DA. A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor Appl Genet. 2007;115(5):721–33.
Kato A. Air drying method using nitrous oxide for chromosome counting in maize. Biotechnic Histochem. 2009;74(3):160–6.
Nagaki K, Tsujimoto H, Isono K, Sasakuma T. Molecular characterization of a tandem repeat, Afa family, and its distribution among Triticeae. Genome. 1995;38(3):479–86.
Han F, Gao Z, Birchler JA. Reactivation of an inactive centromere reveals epigenetic and structural components for centromere specification in maize. Plant Cell. 2009;21(7):1929–39.
Cui Y, Zhang Y, Qi J, Wang H, Wang RR, Bao Y, Li X. Identification of chromosomes in Thinopyrum intermedium and wheat Th. intermedium amphiploids based on multiplex oligonucleotide probes. Genome. 2018;61(7):515–21.
Sheng B. Using infection type records the wheat powdery mildew at seedling stage. Plant Prot. 1988;1:49.
Li ZF, Zheng TC, He ZH, Li GQ, Xu SC, Li XP, Yang GY, Singh RP, Xia XC. Molecular tagging of stripe rust resistance gene YrZH84 in Chinese wheat line Zhou 8425B. Theor Appl Genet. 2006;112(6):1098–103.
Liu X, Wang Y, Sang L, Xiang J, Ji W. Relationship between morphological characters of wheat germplasm and their resistance to Sitobion avenae (F.). J Triticeae Crops. 2006;6:24–8.
Acknowledgments
Not applicable.
Funding
This work was supported by the National Key Research and Development Program of China (2016YFD0102000), the National Natural Science Foundation of China (31671675). The funders Xingfeng Li and Honggang Wang conceived and designed this research. The funding bodies didn’t play a role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author information
Authors and Affiliations
Contributions
YC, PX performed the experiments, data analysis and manuscript writing. XQ, YB, HW participated in some experiments and data analysis. RRCW helped in the manuscript preparation and data analysis. XL designed the project and contributed to the writing of the manuscript and approved the final manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
There is no ethics approval and consent to participate in this manuscript.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplemental Fig. 1.
Chromosomal configuration and GISH results for PMC MI in TE261–1, TE266–1 and TE346–1.
Additional file 2: Supplemental Fig. 2.
Results of the amplification of wheat chromosome-specific markers.
Additional file 3: Supplemental Fig. 3.
Specific molecular marker map indicating chromosomal variation; chr indicates the corresponding chromosome.
Additional file 5: Supplemental Table 1.
Chromosomal configurations of TE261–1, TE266 and TE346–1 at PMC MI.
Additional file 6: Supplemental Table 2.
Sequence alignment results for the screened wheat chromosome-specific molecular markers.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cui, Y., Xing, P., Qi, X. et al. Characterization of chromosome constitution in three wheat - Thinopyrum intermedium amphiploids revealed frequent rearrangement of alien and wheat chromosomes. BMC Plant Biol 21, 129 (2021). https://doi.org/10.1186/s12870-021-02896-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-021-02896-9