
Abstract
An attempt was undertaken to comprehend how phenotypic markers, leaf tip necrosis (LTN), and pseudo black chaff (PBC), are associated with pleiotropic resistance genes in wheat (Triticum spp.). Pleiotropic resistance to stripe rust (Puccinia striiformis f. sp. tritici), leaf rust (P. triticina), stem rust (P. graminis f. sp. tritici) and powdery mildew (Blumeria graminis f. sp. tritici) was evaluated in 57 Egyptian spring wheat cultivars at seedling stage in greenhouse and at adult plant stage in field during 2021/22 and 2022/23. At seedling stage, pleiotropic resistance rated moderately resistance (MR) was observed in five cultivars, Misr-3 (all three rusts), Misr-4 (stripe rust, leaf rust, powdery mildew), Giza-168 (leaf rust, stem rust, powdery mildew), Sakha-94 and Sids-13 (leaf rust, stem rust). At adult plant stage, Misr-4 outperformed all other evaluated cultivars, exhibiting a high level of pleiotropic adult plant resistance (PAPR) against all diseases investigated. High levels of PAPR were also noticed against the three rusts in nine cultivars, Misr-3, Sakha-93, Sakha-94, Sakha-95, Giza-156, Giza-168, Giza-171, Gemmeiza-10 and Sids-13, and moderate levels against powdery mildew in Misr-3, Sakha-95, Giza-156. Four PAPR genes were characterized in cultivars based on phenotypic and molecular markers. Phenotypic markers (LTN and PBC) were observed in 23 cultivars. Molecular marker csLV34 indicated the presence of Lr34/Yr18/Sr57/Pm38/Ltn1 in six cultivars, whilst marker Xwmc44 indicated that Lr46/Yr29/Sr58/Pm39/Ltn2 was present in five cultivars. Marker Xcfd71 indicated the presence of Lr67/Yr46/Sr55/Pm46/Ltn3 in 19 cultivars. Gene Sr2/Yr30/Lr27/Pbc was identified with marker csSr2 in three cultivars. A complete association between LTN and molecular markers was recorded, while a strong association (r = 0.73) was recorded between PBC and molecular markers. Findings demonstrated the reliability of phenotypic markers in predicting pleiotropic resistance in wheat, which would facilitate marker-assisted selection in breeding programs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wheat is a stable food crop for around one-third of the world’s population, with a total area of around 219 million hectares and an annual production of around 760 million tonnes (FAOSTAT 2020). Wheat is prone to be attacked by many diseases. However, rusts and powdery mildew are considered the major constraints to global production of bread wheat (Triticum aestivum L.) and durum wheat (Triticum turgidum var. durum) (Shi et al. 1998; de Vallavieille-Pope et al. 2012; Juliana et al. 2015, Draz et al. 2019; Abou-Zied et al. 2023). Even with the continuous release of resistant wheat cultivars, rusts and powdery mildew constantly threaten sustainable wheat production in Egypt and the world because these cultivars lose their resistance within a few years (Chen et al. 2005; Shahin et al. 2020; Esmail et al. 2021). Yield losses due to stripe rust caused by Puccinia striiformis West. f. sp. tritici ranged from 10 to 70% depending on cultivar susceptibility, earliness of the infection, development rate and disease duration (Chen 2005). Such disease has affected most Egyptian wheat cultivars, resulting in a loss in grain yield ranging from 14 to 26% (El-Daoudi et al. 1996; Draz et al. 2018). Yield losses due to leaf rust caused by P. triticina Eriks., were previously estimated to reach 50% in Egypt (Abdel-Hak et al. 1980). Stem rust caused by P. graminis Pers. f. sp. tritici may cause 100% yield losses in susceptible wheat cultivars when conditions are favorable for the disease (Singh et al. 2002). In addition, epidemic cases of stem rust in 1947 and 1968 were reported in Egypt (Gomma 1968). Powdery mildew caused by Blumeria graminis (DC.) Speer f. sp. tritici causes 13–34% yield loss under low to moderate infection (Li et al. 2011) however, the loss may reach 50% under severe infection (Griffey et al. 1993). The yield losses have been reported in highly susceptible Egyptian cultivars up to 26.68% (Draz et al. 2019). The most effective, economical, and eco-friendly means of wheat disease management is the use of resistant cultivars (Chen 2005; Huerta-Espino et al. 2020). The nature of wheat resistance to rust and powdery mildew could be categorized as race-specific resistance (seedling resistance), also called all-stage resistance, which protects the plant at all growth stages, and as race-non-specific resistance, which is expressed at later stages of plant growth conferring adult plant resistance (APR) (Chen 2013). Race-specific resistance controlled by major genes in cultivars can be easily overcome by new virulent pathogen races, rendering resistance in the cultivars ineffective (McIntosh et al. 1995; Pretorius et al. 2000; Wanyera et al. 2006). On the other hand, race non-specific resistance dependent on the additive effects of minor gene accumulation associated with quantitative traits is characterized by durable, partial, and adult plant resistance (APR) expressed at the post-seedling stage with extended latent periods that delay the spread of the disease (Parlevliet 1975; Das et al. 1992,), and, therefore, it is considered as a promising trend of genetic resistance in wheat (Singh et al. 1998, 2005; Rehman et al. 2013). Among the cataloged resistance genes that have been widely deployed because of their exceptional durability to stripe rust, leaf rust, stem rust and powdery mildew, four genes i.e., Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc confer pleiotropic adult plant resistance (PAPR) (Dyck and Samborski 1979; Singh 1992; McIntosh et al. 2003; Spielmeyer et al. 2005; Vanegas et al. 2008; Singh et al. 2012, 2013; Herrera-Foessel et al. 2014).
The use of both phenotype (observable traits) and genotype (genetic characteristics) assessments is essential for evaluating wheat germplasm for resistance to rust and powdery mildew, and incorporating into breeding programs. Phenotype evaluation can be done through field observations and assessments, where the performance of different cultivars is compared under similar environmental conditions. By evaluating the phenotypes of wheat cultivars, it is possible to select those that show disease resistance. Genotype evaluation, on the other hand, focuses on the genetic makeup of the wheat cultivars. Molecular techniques, such as DNA markers and genetic sequencing, can be used to identify specific genes or gene variants associated with resistance to these diseases. By analyzing the genotypes of wheat cultivars, the presence or absence of genes linked to disease resistance can be determined.
The pleiotropic adult plant resistance (PAPR) genes have been reported to be associated with phenotypical or morphological markers in wheat. A phenotype called leaf tip necrosis (LTN), which is characterized at the post-flowering stage by necrosis of the flag leaf tips extending to a few centimeters along the leaf edges, is associated with the resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1 and Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 (Dyck 1987; Singh 1992; Herrera-Foessel et al. 2014). Leaf tip necrosis is an innate defense mechanism in resistant lines and occurs spontaneously even before pathogen challenges (Rubiales and Niks 1995; Hulbert et al. 2007). Besides LTN, another significant phenotype of black pigmentation is called pseudo-black chaff (PBC), which is associated with the PAPR gene Sr2/Yr30/Lr27/Pbc (McFadden 1939; Hare and McIntosh 1979; Kota et al. 2006). This gene has provided effective resistance for many decades to stem rust, stripe rust, leaf rust, and powdery mildew (Singh et al. 2000, 2005; Mago et al. 2011). Pseudo-black chaff occurs after anthesis around the glumes and the stem internodes (Kuspira and Unrau 1958), with degrees of expression varying depending on genotype and environment (Singh et al. 2008). Several mechanisms were supposed to explain the genetic association between PBC and durable resistance of Sr2/Yr30/Lr27/Pbc. The expression of the PBC phenotype in resistant plants might form physical or chemical challenges to the pathogen that delay the infection process and the two traits were inseparable even by recombination (Kota et al. 2006). High expression of PBC might lead to shrunk kernels that reduce yield and make it undesirable for the farmers (Hare and McIntosh 1979); therefore, selecting moderate levels of PBC in resistant lines with Sr2/Yr30/Lr27/Pbc is preferred for breeders. The delay mechanism of disease development through durable resistance is a form of partial resistance in wheat germplasm (Singh et al. 2012). Field evaluations of infection types and severities can be used to refer to the existence of APR in wheat varieties. The quantitative aspects of adult plant resistance in wheat cultivars have been described and estimated using disease severity and coefficient of infection (CI) values at a particular crop development stage (Saari and Wilcoxson 1974; Pathan and Park 2006).
The use of molecular markers is the most reliable way to confirm the presence of phenotypic traits (LTN and PBC) associated with pleiotropic resistance genes in wheat. Molecular markers of resistance genes have long been a major aim for marker-assisted selection and identification in different wheat germplasm. The STS marker csLV34 is a specific diagnostic tool closely linked to the Lr34/Yr18/Sr57/Pm38/Ltn1 locus for a multi-pathogen resistance trait (Lagudah et al. 2006). Suenaga et al. (2003) determined that the microsatellite locus Xwmc44 is located 5.6 cM proximal to the putative QTL for Lr46/Yr29/Sr58/Pm39/Ltn2. The SSR marker Xcfd71 has been reported to be closely linked to Lr67/Yr46/Sr55/Pm46/Ltn3 (Hiebert et al. 2010). Mago et al. (2011) developed a CAPS marker named csSr2 to be an accurate and reliable marker linked to Sr2/Yr30/Lr27/Pbc. The association between phenotypic traits (LTN and PBC) and pleiotropic resistance genes still needs further study to ascertain how well these traits predict pleiotropic resistance in wheat, which would facilitate marker-assisted selection. Hence, this study aimed to screen Egyptian spring wheat cultivars for pleiotropic minor gene-based resistance against rusts and powdery mildew based on phenotypic and molecular markers.
Materials and methods
Plant material
A total of 57 Egyptian spring wheat cultivars listed in Table 1 were screened for seedling and adult plant resistance against rusts and powdery mildew under field conditions during two growing seasons 2021/22 and 2022/23. Three wheat genotypes, RL6058, RL6077 and Pavon-F76 listed in Table 2 each carrying pleiotropic resistance genes and phenotypic traits, i.e., Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc were provided by International Maize and Wheat Improvement Center (CIMMYT) and served as reference material. A universal susceptible wheat cultivar “Morocco” served as the susceptible check for disease evaluation.
Seedling resistance evaluation
Evaluation of seedling resistance of the wheat genotypes (Tables 1 and 2) against rusts and powdery mildew was carried out in the greenhouse of the Wheat Disease Research Department, Sakha Agricultural Research Station, Plant Pathology Research Institute, ARC, Egypt. Collections of pathotypes (races) prevalent in Egypt that were earlier described in our recent studies for yellow rust (Esmail et al. 2021), leaf rust, powdery mildew (Draz et al. 2022) and stem rust (Abou-Zeid et. al. 2023) were used separately for inoculation process. Each experiment was performed in a completely randomized design with three replicates (pots) for each cultivar. In separate tests, inoculation was performed using urediniospores of P. striiformis f. sp. tritici, P. triticina, P. graminis f. sp. tritici and conidiospores of B. graminis f. sp. tritici according to the methods described by Stubbs (1988), Roelfs et al. (1992), Stakman et al. (1962), and Browder (1972), respectively. The spore inoculum of each pathogen was multiplied and propagated on 8-day-old seedlings of the universal susceptible wheat cultivar “Morocco” to obtain enough inoculum for evaluation. The inoculated seedlings were incubated overnight in a dark dew chamber under high relative humidity (RH) at 10 °C for stripe rust and at 18–20 °C for leaf rust, stem rust and powdery mildew. After incubation, the inoculated seedlings were transferred to the greenhouse conditioned and maintained at 13–15 °C (stripe rust), 19–22 °C (leaf rust), 20–25 °C (stem rust), and 20–22 °C (powdery mildew) under 90–100% RH and 102.6 µmol m−2 s−1 light intensity with a 16/8 h day/night photoperiod. After spore propagation, 8-day-old seedlings of the tested cultivars were inoculated separately with spores of each pathogen according to the methods described above. Mildew infection types (ITs) were recorded ten days after inoculation, using the 0–9 classification standard of Leath and Heun (1990). Rust infection types were scored two weeks after inoculation, using the 0–9 scale for stripe rust (McNeal et al. 1971), the 0–4 classification standard for leaf rust and stem rust according to Roelfs et al. (1992) and Stakman et al. (1962), respectively. Seedling responses were expressed as R = Resistant (stripe rust ITs 0, 1, 2; leaf and stem rust ITs 0, 1; mildew ITs 0–3), MR = Moderately Resistant (stripe rust ITs 3–5; leaf and stem rust ITs 2; mildew ITs 4), MS = Moderately Susceptible (stripe rust ITs 6, 7; leaf and stem ruts ITs 3; Mildew ITs 5, 6), S = Susceptible (stripe rust ITs 8; leaf and stem rust ITs 4; mildew ITs 7, 8), HS = Highly Susceptible (Stripe rust and mildew ITs 9).
Adult plant resistance evaluation
The wheat genotypes (Tables 1 and 2) were screened against rusts and powdery mildew in the open field at the Experimental Farm of Sakha Agricultural Research Station, Plant Pathol Res. Inst., ARC, Egypt during two growing seasons 2021/22 and 2022/23. The experiment was designed in a randomized complete block design (RCBD) with three replicates. Cultivars were sown in experimental units (plots) containing three rows 3 m long and 30 cm apart and 5 g seed/row. All recommended cultural practices in commercial wheat fields were applied. Adult plant resistance was evaluated under field natural infection at growth stages GS 50–90 (Zadoks et al. 1974). The level of APR was determined based on the average coefficient of infection (ACI) values. For rust, host responses, R; Resistant, MR; Moderately Resistant, MS; Moderately Susceptible, or S; Susceptible were scored according to Roelfs et al. (1992). Disease severity was expressed as the percentage coverage of leaves with rust pustules following Cobb’s scale modified by Peterson et al. (1948). The final rust severity (FRS) was computed as disease severity (DS) for each cultivar in comparison with a susceptible cultivar “Morocco” (Das et al. 1993). For powdery mildew, a double-digit (D1D2) scale was used for disease assessment (Draz et al. 2019). The scoring was denoted on the plants (20 plants/accession) using the 0–9 scale (Saari and Prescott 1975). The disease severity percentage was calculated according to Sharma and Duveiller (2007). The coefficient of infection (CI) was calculated according to Saari and Wilcoxson (1974) and Pathan and Park (2006), by multiplying the severity value by a constant value of 0.2, 0.4, 0.6, 0.8 and 1.0 for host response ratings of R, MR, MR-MS, MS or S, respectively. These were calculated as the average coefficient of infection (ACI) values over two growing seasons. Cultivars with ACI values of 0–20, 21–40 and 41–60 were regarded as possessing high, moderate and low levels of APR, respectively. Cultivars with ACI values above 60 were regarded as susceptible, although values between 61 and 80 could reflect the presence of minor genes.
Phenotypic marker characterization
Phenotypic markers (LTN and PBC) were evaluated in the tested wheat cultivars under field conditions during the two growing seasons 2021/22 and 2022/23. Phenotyping for LTN involved measuring the extent of LTN from the tip of the leaf up to a few centimeters along the edges at anthesis. A 0-to-4 scale adopted by (Juliana et al. 2015) was used where 0 = no LTN; 1 = slight LTN; 2 = medium LTN; 3 = high LTN; and 4 = very high LTN. Phenotyping for PBC was done at anthesis based on the presence of black pigmentation around the glumes and stem internodes (Kota et al. 2006; Kaur et al. 2009). A 0-to-4 scale was used where 0 = no pigmentation; 1 = slight pigmentation; 2 = medium pigmentation; 3 = high pigmentation; and 4 = very high pigmentation.
Molecular marker characterization
The wheat cultivars showing phenotypic markers were subjected to molecular assay to determine the presence/absence of associated resistance genes. Four reliable molecular markers, the STS marker csLV34 (Lagudah et al. 2006), the microsatellite marker Xwmc44 (Suenaga et al. 2003), Xcfd71 (Hiebert et al. 2010), and the CAPS marker csSr2 (Mago et al. 2011) were used to detect the resistance genes Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively. Genomic DNA was extracted from fresh leaf samples using the method of Doyle and Doyle (1987). PCR was performed in a total reaction volume of 25 μl in Techne, PROGENE Thermocycler according to the initial methods of Lagudah et al. (2006), Suenaga et al. (2003), Hiebert et al. (2010) and Mago et al. (2011) for Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively. The PCR amplification products of csSr2 marker were digested with the restriction enzyme Bsph1. Amplification products of 10 μl each sample were electrophoresed at 100 V for about 20 min. in 2.5% agarose gel stained with ethidium bromide. A 100 bp DNA ladder H3 RTU, Nippon Genetics Europe GmbH, served in calibration. Banding patterns were visualized using a UV-transilluminator, Herolab UVT 2020, Kurzwellig, and then photographed. A detail list of the molecular markers, primer sequence, amplification band, and PCR conditions are shown in Table 3.
Phenotypic and molecular diversity
Phenotypic diversity of phenotypic markers; leaf tip necrosis (Ltn1, Ltn2 and Ltn3) and pseudo-black chaff (Pbc), along with molecular diversity of molecular markers; csLV34, Xwmc44, Xcfd71 and csSr2, associated with pleiotropic resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively, were measured in twenty-three Egyptian wheat cultivars investigated. Phenotypic and molecular data obtained were analyzed based on Simpsonʼs index (HS) and Shannonʼs index (HSh) according to Groth and Roelfs (1987) using the PC software MVSP 3.1 (Kovach 1999).
Relative pleiotropic resistance index (RPRI)
The highest infection of a candidate genotype is set at 100 and all other genotypes are adjusted accordingly to calculate the country average relative percentage attack (CARPA) for each genotype. Relative pleiotropic resistance index (RPRI) was calculated based on 0–9 scale according to Aslam (1982) using the following formula:
RPRI = ((100 − PA)/100) × 9.
Where PA is a percentage attack of each disease derived from the calculated means of ACI data during two growing seasons.
The RPRI was classified based on the 0–9 scale, where 0 denotes most susceptible and 9 highly resistant. The desirable index value is 7 or above and the acceptable index is 6.
Genetic distance of wheat cultivars
The genetic distance of the tested Egyptian spring wheat cultivars was generated according to Nei & Li’s coefficient using the PC software MVSP 3.1 (Kovach 1999). Binary data generated from phenotypic and molecular data (presence/absence) associated with pleiotropic resistance genes in cultivars were used to construct a dendrogram. Cluster analysis was done using the unweighted pair group arithmetic mean (UPGMA) method.
Statistical analysis
Data were pooled from repeated testes and subjected to analysis of variance using SPSS software V22.0. Means were compared using LSD method at 0.05 significance (Steel and Torrie 1980). The association of phenotypic and molecular data was calculated using a correlation coefficient (r) according to Pearson and Hartley (1970).
Result
Pleiotropic seedling resistance
Seedling responses of the tested 57 Egyptian wheat cultivars revealed their susceptibility to rusts and powdery mildew (Table 4). However, ten cultivars (Misr-3, Misr-4, Sakha-94, Giza-168, Sids-13, Sids-14, Beniswef-1, Beniswef-4, Beniswef-5, Beniswef-7) were moderately resistant (MR) to leaf rust, and three cultivars (Misr-4, Giza-168, Gemmeiza-12) showed MR to powdery mildew. In comparison, five cultivars (Misr-3, Sakha-94, Giza-168, Giza-171, Sids-13) were MR to stem rust and only two cultivars (Misr-3, Misr-4) exhibited MR to stripe rust. Moreover, moderate susceptible (MS) responses were observed with 12 cultivars (Misr-1, Misr-2, Sakha-93, Giza-171, Sids-12, Shandweel-1, Shandweel-2, Nubaria-2, Beniswef-3, Beniswef-5, Sohag-3, Sohag-4) against leaf rust and 12 cultivars (Misr-1, Misr-2, Misr-3, Sakha-95, Giza-168, Giza-171, Gemmeiza-9, Gemmeiza-10, Gemmeiza-11, Beniswef-5, Beniswef-6, Sohag-4) against powdery mildew. While MS responses were recorded with eight cultivars to stem rust (Misr-4, Sakha-95, Gemmeiza-9, Sids-14, Shandweel-2, Nubaria-2, Beniswef-6, Sohag-4) and ten cultivars to stripe rust (Misr-4, Sakha-93, Sakha-94, Sakha-95, Giza-168, Giza-171, Shandweel-2, Nubaria-2, Beniswef-7, Sohag-4). It also observed that five cultivars, Misr-1, Sakha-69, Giza-160, Gemmeiza-11 and Sids-12, showed high susceptibility (HS) to stripe rust, while five cultivars Sakha-93, Sakha-94, Gemmeiza-1 and Gemmeiza-3 exhibited HS to powdery mildew. The remaining tested cultivars were rated as susceptible (S) to rusts and powdery mildew. Although, none of the tested wheat exhibited pleiotropic seedling resistance (PSR) with specific resistance response (R-MR) to all diseases. However, PSR (rated MR) against three diseases was observed in Misr-3 (three rusts), Misr-4 (stripe rust, leaf rust, powdery mildew) and Giza-168 (leaf rust, stem rust, powdery mildew). It was also recorded in two cultivars, Sakha-94 and Sids-13 against leaf rust and stem rust. The reference genotype RL6058 (Lr34/Yr18/Sr57/Pm38/Ltn1) showed MS to rusts, while it showed MR to powdery mildew. The other two reference genotypes RL6077 (Lr67/Yr46/Sr55/Pm46/Ltn3) and Pavon-F76 (Lr46/Yr29/Sr58/Pm39/Ltn2 and Sr2/Yr30/Lr27/Pbc) showed MS to all diseases.
Pleiotropic adult plant resistance
The levels of adult plant resistance of the tested wheat cultivars to rusts and powdery mildew were assessed based on the average coefficient of infection (ACI) calculated during both growing seasons, 2021/22 and 2022/23. Analysis of variance (Table 5) revealed highly significant differences between means calculated for genotypes (G) for all diseases, while no significant differences between seasons (S) as well as the interaction of genotype and season (G × S) for all diseases. In the data given in Table 6, high variations of ACI values among cultivars demonstrated variable levels of APR among cultivars. Cultivars with ACI values 0–20, 21–40, and 41–60 imply that they possess high, moderate, and low levels of APR, respectively. Among 57 Egyptian wheat cultivars tested, only 13 cultivars exhibited high levels of APR to stripe rust, the best of them, Misr-3, Misr-4, Sakha-93, Sakha-94, Giza-168, Giza-171, Sids-13 and Beniswef-6 which possessed ACI values up to 10.00. The data also indicated that 30 cultivars had high levels of APR to leaf rust; the best were Misr-1, Misr-2, Misr-3, Misr-4, Sakha-93, Sakha-94, Giza-168, Giza-171, Gemmeiza-10, Sids-7, Sids-13, Sids-14 recording ACT values up to 8.00. High levels of APR to stem rust were recorded in 26 cultivars; of them, Misr-3, Misr-4, Sakha-94, Sids-13, and Shandweel-1 showed best levels up to 7.66 ACI. As data revealed that only three cultivars (Misr-4, Sids-3, Sids-5) exhibited high levels of APR to powdery mildew. The best performance of cultivars exhibiting pleiotropic resistance was recorded with Misr-4 which possessed high levels of PAPR against all diseases. High levels of PAPR were also recorded against the three rusts in nine cultivars, Misr-3, Sakha-93, Sakha-94, Sakha-95, Giza-156, Giza-168, Giza-171, Gemmeiza-10 and Sids-13, and moderate levels against powdery mildew in Misr-3, Sakha-95, Giza-156 and Giza-168. Moderate levels of PAPR were observed against all diseases in six cultivars, Gemmeiza-1, Gemmeiza-12, Shandweel-2, Nubaria-2, Beniswef-7 and Sohag-4. The remaining cultivars showed variation levels with PAPR to at least two diseases except Giza-160. The reference genotype RL6058 (Lr34/Yr18/Sr57/Pm38/Ltn1) and Pavon-F76 (Lr46/Yr29/Sr58/Pm39/Ltn2 and Sr2/Yr30/Lr27/Pbc) exhibited high levels of pleiotropic adult plant resistance (PAPR) to the three rusts and moderate levels of APR to powdery mildew, while RL6077 (Lr67/Yr46/Sr55/Pm46/Ltn3) exhibited moderate levels of PAPR to all diseases.
Phenotypic markers
Phenotypic markers (LTN and PBC) were evaluated in the tested wheat cultivars after the heading stage under field conditions. The results for characterizing phenotypic markers of pleiotropic resistance genes exhibited different degrees as shown in Table 6. Among the tested 57 cultivars, only 23 cultivars, Misr-1, Misr-2, Misr-3, Misr-4, Sakha-93, Sakha-94, Sakha-95, Giza-168, Giza-171, Gemmeiza-9, Gemmeiza-10, Gemmeiza-11, Gemmeiza-12, Sids-12, Sids-13, Sids-14, Shandweel-1, Shandweel-2, Nubaria-2, Beniswef-1, Beniswef-5, Sohag-3 and Sohag-4, exhibited phenotypic markers LTN and PBC. They all exhibited LTN markers with different degrees, while only five cultivars, Misr-1, Misr-2, Sakha-93, Giza-168, and Shandweel-1, exhibited PBC markers. Degrees of LTN rated as 1 were observed with 19 cultivars, Misr-1, Misr-2, Misr-3, Misr-4, Sakha-93, Sakha-94, Sakha-95, Giza-168, Gemmeiza-9, Gemmeiza-10, Gemmeiza-11, Gemmeiza-12, Sids-12, Sids-13, Sids-14, Shandweel-2, Nubaria-2, Beniswef-5 and Sohag-3. Four cultivars, Misr-1, Misr-2, Shandweel-1, Beniswef-1, and Sohag-4, also exhibited LTN degrees rated as 2, while three cultivars, Misr-4, Sakha-94, and Sakha-95, rated LTN as 4. Only one cultivar (Giza-168) also exhibited LTN rated as 3. It is worth noting that Misr-1 and Misr-2 exhibited LTN degrees of 1 and 2, and Misr-4, Sakha-94, and Sakha-95 each exhibited LTN degrees of 1 and 4. Sids-13 exhibited both LTN degrees of 1 and 3. On the other hand, the PBC marker with a degree of 3 was recorded with only two cultivars, Misr-1 and Shandweel-1, while a degree of 2 was observed with Misr-2. A PBC degree 1 was observed with two cultivars, Sakha-93 and Giza-168. The last-mentioned five cultivars exhibited both phenotypic traits (LTN and PBC) in combination. The reference genotype RL6058 showed the LTN marker with a degree of 4, while the reference genotype RL6077 showed the LTN marker with a degree of 1. Pavon-F76 showed phenotypic traits (LTN and PBC) with a degree of 2 for LTN and a degree of 3 for PBC. No phenotypic traits were observed with the Morocco check cultivar free from these genes. It is also observed that the 23 cultivars that have phenotypic markers also possessed a level of PAPR with different degrees to rusts and powdery mildew.
Molecular markers
The Egyptian wheat cultivars, Misr-1, Misr-2, Sakha-8, Sakha-93, Sakha-94, Giza-156, Giza-167, Giza-168, Giza-171, Gemmeiza-9, Gemmeiza-10, Gemmeiza-11, Sids-12, Sids-13, Sids-14, Sandweel-1, Beniswef-1, Beniswef-4 which possessed phenotypic markers (LTN or PBC) were genotyped using molecular markers csLV34, Xwmc44, Xcfd71 and csSr2 to detect the PAPR genes Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively. DNA banding patterns illustrated in Figs. 1, 2, 3 and 4 and data in Table 7 revealed the presence/absence of resistance genes in tested cultivars. Out of the 23 cultivars, amplification products of csLV34 indicated the presence of Lr34/Yr18/Sr57/Pm38/Ltn1 in six cultivars, Misr-3, Sids-13, Sakha-94, Sakha-95, Giza-168 and Giza-171 with a fragment of 150-bp with a fragment of 150-bp as presented in the positive reference genotype RL6058 (Fig. 1). The allele with a PCR fragment of 229-bp resulted in the other cultivars indicated the absence of the gene. The marker Xwmc44 amplified DNA bands of 242-bp in five cultivars, Misr-1, Misr-2, Shandwel-1, Beniswef-1 and Sohag-4 which indicated the presence of Lr46/Yr29/Sr58/Pm39/Ltn2 as in the positive reference Pavon-F76. Other fragments of allele indicated the absence of the gene (Fig. 2). The marker Xcfd71 amplified DNA bands of 214-bp in 19 cultivars which indicated the presence Lr67/Yr46/Sr55/Pm46/Ltn3 in Misr-1, Misr-2, Misr-3, Misr-4, Sakha-93, Sakha-94, Sakha-95, Giza-168, Gemmiza-9, Gemmiza-10, Gemmiza-11, Gemmiza-12, Sids-12, Sids-13, Sids-14, Shandweel-2, Nubaria-2, Beniswef-5, and Sohag-3 as in reference genotype RL6077 (Fig. 3). Molecular marker csSr2 was found to be validated to resistance gene Sr2/Yr30/Lr27/Pbc in three cultivars, Misr-1, Misr2 and Shandweel-1 resulting in three fragments of 172-bp, 112-bp, and 53-bp as presented in the positive reference Pavon-F76. Cultivars with no or other fragments imply that they lack the resistance gene (Fig. 4). The two cultivars, Misr-1and Misr-2, showed a combination of three genes Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27, while a combination of two genes Lr34/Yr18/Sr57/Pm38/Ltn1 and Lr67/Yr46/Sr55/Pm46/Ltn3 was present in five cultivars, Misr-3, Sakha-94, Sakha-95, Giza-168 and Sids-13. Lr46/Yr29/Sr58/Pm39/Ltn2 and Sr2/Yr30/Lr27/Pbc was combined in only one cultivar Shandweel-1.

PCR amplification products of the STS marker csLV34 indicate the presence of Lr34/Yr18/Sr57/Pm38/Ltn1 in six cultivars, Misr-3 (lane 2), Sids-13 (lane 4), Sakha-94 (lane 16), Sakha-95 (lane 17), Giza-168 (lane18) and Giza-171 (lane 19) with a fragment of 150-bp as presented in the positive genotype RL6058 (lane 24). A fragment of 229-bp indicates the absence of the gene. M, DNA ladder at 100-bp

PCR amplification products of the SSR marker Xwmc44 indicate the presence of Lr46/Yr29/Sr58/Pm39/Ltn2 in five cultivars, Misr-1 (lane 5), Misr-2 (lane 10), Shandwel-1 (lane 21), Beniswef-1 (lane 22), Sohag-4 (lane 23) with a fragment of 242-bp as presented in the positive genotype Pavon-F76 (lane 24). M, DNA ladder at 100-bp

PCR amplification products of the SSR marker Xcfd71 indicate the presence of Lr67/Yr46/Sr55/Pm46/Ltn3 in 19 wheat cultivars, Misr-1 (lane 1), Misr-2 (lane 2), Misr-3 (lane 3), Misr-4 (lane 4), Sakha-93 (lane 5), Sakha-94 (lane 6), Sakha-95 (lane 7), Giza-168 (lane 8), Gemmiza-9 (lane 9), Gemmiza-10 (lane 11), Gemmiza-11 (lane 13), Gemmiza-12 (lane 14), Sids-12 (lane 15), Sids-13 (lane 16), Sids-14 (lane 17), Shandweel-2 (lane 18), Nubaria-2 (lane 21), Beniswef-5 (lane 22) and Sohag-3 (lane 23) with a fragment of 214-bp as presented in the positive reference RL6077 (lane 24). M, DNA ladder at 100-bp

PCR amplification products of the CAPS marker csSr2 indicate the presence of Sr2/Yr30/Lr27/Pbc in three cultivars, Misr-1 (lane 2), Misr-2 (lane 3), Shandweel-1 (lane 9) with three unique fragments of 172-bp, 112-bp, and 53-bp as presented in the positive check cultivar Pavon-F76 (lane 1). M, DNA ladder at 100-bp
Association of phenotypic and molecular markers
Data in Table 7 and Fig. 5 revealed the association of phenotypic and molecular markers characterized in 23 Egyptian wheat cultivars. The obtained data indicated a significant association between phenotypic and molecular markers, indicating the presence of the resistance gene in all cultivars except only two cultivars, Sakha-93 and Giza-168, for PBC. The 23 cultivars which exhibited LTN and/or PBC indicated that the phenotypic markers were strongly associated with molecular markers for genes Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2 Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc. Only two cultivars, Sakha-93 and Giza-168, with slight PBC, had no successful association with molecular markers and, therefore, resistance gene Sr2/Yr30/Lr27/Pbc. LTN phenotype degree could be classified into three categories, Ltn1, Ltn2 and Ltn3. In which, Ltn1 could be characterized by high (degree 3) and very high (degree 4) LTN phenotype, while Ltn2 could be characterized by medium (degree 2) LTN phenotype and Ltn3 could be characterized by slight LTN phenotype (degree 1). It was observed that the medium degree of LTN phenotype was associated with Lr46/Yr29/Sr58/Pm39/Ltn2 in five cultivars, Misr-1, Misr-2, Shandwel-1, Beniswef-1, Sohag-4 as in reference genotype Pavon-F76. LTN phenotype degrees rated high and very high were associated with Lr34/Yr18/Sr57/Pm38/Ltn1 in six cultivars, Misr-3, Sids-13, Sakha-94, Sakha-95, Giza-168 and Giza-171 as in reference genotype RL6058. Slight LTN phenotypes were associated with Lr67/Yr46/Sr55/Pm46/Ltn3 in 19 cultivars as in reference genotype RL6077. Different degrees of PBC phenotype, whether a high pigmentation (degree 3) in cultivars Misr-1 and Shandweel-1 or medium pigmentation (degree 2) in Misr-2 were associated with the presence of Sr2/Yr30/Lr27/Pbc, while slight pigmentation (degree 1) in Sakha-93 and Giza-168 had no association with the resistance gene Sr2/Yr30/Lr27/Pbc. It could be concluded that the wheat cultivars with pleiotropic resistance genes, individually or in combination, all possessed phenotypic markers and PAPR. A correlation coefficient (r) indicated a complete association between molecular markers csLV34, Xwmc44, and Xcfd71 and Ltn1, Ltn2, and Ltn3, respectively (r = 1). A strong association (r = 0.7348) was recorded between marker csSr2 and Pbc.

Phenotypic markers, leaf tip necrosis (Ltn1, Ltn2 and Ltn3) and pseudo-black chaff (Pbc) of pleiotropic resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, characterized in twenty-three Egyptian wheat cultivars investigated
Phenotypic and molecular diversity
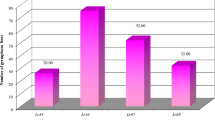
Phenotypic and molecular data obtained were analyzed based on Simpsonʼs index (HS) and Shannonʼs index (HSh). Data in Fig. 6 illustrated phenotypic diversity of phenotypic markers; leaf tip necrosis (Ltn1, Ltn2 and Ltn3) and pseudo-black chaff (Pbc), along with molecular diversity of molecular markers; csLV34, Xwmc44, Xcfd71 and csSr2, associated with pleiotropic resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively, in twenty-three Egyptian wheat cultivars investigated. Simpsonʼs index (HS) indicated diversity from 0.75 to 0.95, while Shannonʼs index (HSh) ranged from 1.38 to 2.99. The diversities observed between each of phenotypic markers, Ltn1, Ltn2 and Ltn3, and molecular markers, csLV34, Xwmc44 and Xcfd71, respectively, were identical. However, it was a little different between Pbc (0.80 HS, 1.60 HSh) and csSr2 (0.75 HS, 1.38 HSh). The highest diversity was recorded with Ltn3 and Xcfd71 (0.95 HS, 2.99 HSh each).

Phenotypic diversity of phenotypic markers; leaf tip necrosis (Ltn1, Ltn2 and Ltn3) and pseudo-black chaff (Pbc), along with molecular diversity of molecular markers; csLV34, Xwmc44, Xcfd71 and csSr2, associated with pleiotropic resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively, characterized in twenty-three Egyptian wheat cultivars investigated
Genetic distance of wheat cultivars
The Nei & Li’s coefficient generated the genetic distance of 23 Egyptian wheat cultivars based on phenotypic and genotyping data (Fig. 7). Clustering separated cultivars into two sub-clusters at a genetic distance of 0.185, each of them consisting of two sub-sub-clusters at a close genetic distance 0.556 and 0.549, respectively. The first sub-cluster consisted of two sub-sub-clusters, the first grouped 11 cultivars with the same genetic distance of 1.0, i.e., Sohag-3, Beniswef-5, Nubaria-2, Shandweel-1, Sids-14, Sids-12, Gemmeiza-9, Gemmeiza-10, Gemmeiza-11, Gemmeiza-12 and Misr-1, additional to Sakha-93, which diverged individually at a genetic distance of 0.8. The other sub-sub-cluster consisted of five cultivars with the same genetic distance, i.e., Sids-13, Giza-168, Sakha-94, Sakha-95 and Misr-3 additional to Giza-171, which diverged individually at a genetic distance of 0.667. The second sub-cluster consisted of two sub-sub-clusters; one of them consisted of two cultivars with the same distance, i.e., Sohag-4 and Beniswef-1, while the other contained two cultivars, Misr-1 and Misr-2 at the same distance and Shandweel-1, which diverged individually at a distance of 0.8.

Dendrogram of Nei & Li's genetic distance constructed for Egyptian wheat cultivars based on phenotypic and molecular data, clustering with bootstrap values
Relative pleiotropic resistance index (RPRI)
Data in Table 8 distinguished 23 Egyptian wheat cultivars based on the relative pleiotropic resistance index (RPRI). Four cultivars exhibited desirable RPRI (7 or more) or acceptable (6 and less than 7) against all diseases, i.e., Misr-4, Sakha-95, Shandweel-2 and Sohag-4. Moreover, nine cultivars exhibited desirable/acceptable RPRI against the three rusts, i.e., Misr-3, Sakha-93, Sakha-94, Giza-168, Giza-171, Gemmeiza-9, Gemmeiza-10, Beniswef-1 and Beniswef-5. Three cultivars exhibited desirable/acceptable RPRI against three diseases, i.e., Misr-2 (stripe rust, leaf rust, powdery mildew), Gemmeiza-11 and Shandweel-1 (leaf rust, stem rust, powdery mildew) and Nubaria-2 (stripe rust, stem rust, powdery mildew). Sids-14 exhibited desirable RPRI against two rusts of leaf and stem. The remaining cultivars exhibited desirable/acceptable RRI against one disease, i.e., leaf rust (Misr-1, Gemmeia-12, Sohag-3), and stem rust (Sids-12).
Discussion
The concept of pleiotropic disease resistance in wheat cultivars containing multiple resistance genes remains encouraged in many breeding programs worldwide, particularly at CIMMYT, emphasized by Norman Borlaug (Huerta-Espino et al. 2020). Many wheat cultivars have been released by CIMMYT, which carries high levels of resistance to stripe rust, leaf rust, stem rust, and powdery mildew. In the present study, we attempt to screen spring wheat cultivars possessing multiple disease-resistance genes based on phenotypic and molecular markers. Four pleiotropic resistance genes, Lr34/Yr18/Sr57/Pm38/Ltn1 (RL6058), Lr67/Yr46/Sr55/Pm46/Ltn3 (RL6077), Lr46/Yr29/Sr58/Pm39/Ltn2 and Sr2/Yr30/Lr27/Pbc (Pavon-F76) provided by CIMMYT, were used as reference genotypes in the current study. A total of 57 Egyptian spring wheat cultivars were investigated for seedling and adult plant resistance against rusts and powdery mildew during two growing seasons, 2021/22 and 2022/23. Seedling responses emphasized the variation in infection types with a high level of susceptibility among the cultivars. Seedling pleiotropic resistance (rated MR) was observed in Misr-3 (three rusts), Misr-4 (stripe rust, leaf rust, powdery mildew), Giza-168 (leaf rust, stem rust, powdery mildew), Sakha-94 and Sids-13 (leaf rust and stem rust). These findings would give us the ground to say that the last-mentioned cultivars possess race-specific resistance controlled by major resistance genes as well as the components of slow rusting or durable resistance (race-none-specific resistance) at a seedling stage, which is controlled by minor genes. Seedling resistance and its components in Egyptian wheat cultivars against rusts and powdery mildew have also been described in several studies (El-Daoudi et al. 1983; El-Shamy and Mousa 2004; Shahin et al. 2020; Esmail et al. 2021; Draz et al. 2022; Abou-Zeid et al. 2023) which were in agreement with our results. The levels of APR are similar between cultivars, independent of whether the cultivars possess seedling resistance or susceptibility (Huerta-Espino et al. 2020).
The adult plant resistance (durable resistance) in wheat is based on the additive effect of partially resistant minor genes, usually polygenic and active at the adult plant stage. The varieties possessing minor gene-based resistance showed almost the same level of adult plant resistance against different races and remained effective in different climatic conditions. The varieties with major gene-based race-specific resistance did not have a long life and usually collapsed after a few years. Adult plant resistance could be estimated using disease severity and average coefficient of infection (ACI) values during the crop development stage (Saari and Wilcoxson 1974; Pathan and Park 2006). Our findings revealed variations in levels of adult plant resistance to rust and powdery mildew in Egyptian wheat cultivars tested during two growing seasons (2021/22 and 2022/23). Cultivars were most affected by powdery mildew where only three cultivars exhibited a high level of APR, followed by stripe rust (13 cultivars), stem rust (26 cultivars), and leaf rust (30 cultivars). The best performance of cultivars was recorded with Misr-4 which possessed high levels of PAPR against all diseases. High levels of PAPR were also noticed against the three rusts in nine cultivars, Misr-3, Sakha-93, Sakha-94, Sakha-95, Giza-156, Giza-168, Giza-171, Gemmeiza-10 and Sids-13, and moderate levels against powdery mildew in Misr-3, Sakha-95, Giza-156 and Giza-168. Moderate levels of PAPR were observed against all diseases in six cultivars, Gemmeiza-1, Gemmeiza-12, Shandweel-2, Nubaria-2, Beniswef-7 and Sohag-4. The susceptible check cultivar Morocco was full of diseases, rating ACI more than 80.00 for rusts and 69.33 for powdery mildew, where there was no chance of escape. Considerable efforts to screen adult plant resistance of Egyptian wheat cultivars against rusts and powdery mildew have been previously reported in several studies (Ashmawy et al. 2014; Draz et al. 2018, 2019; Shahin et al. 2018, 2020; Esmail et al. 2021; Abou-Zeid et al. 2023) that agreed with our results. Therefore, a mechanism based on the additive effects of partial resistance minor genes that probably exist in wheat varieties of Egypt would be useful in breeding for disease resistance. This kind of resistance is desirable, as it is long-lasting and more durable against the changing disease virulence patterns. This is supported by the consistent resistance response of the CIMMYT genotypes RL6058, RL6077, and Pavon-F76 (Singh et al. 1998, 2000) used as reference material in our study, as well as Egyptian cultivars, Sakha-93, Sakha-94, Giza-156, Giza-168, Giza-171, and Sids-14, hence high economic returns may be achieved from such kind of resistance. Hence, as per our findings, the last-mentioned cultivars showing the low frequency of disease severity with lower ACI values have pleiotropic resistance against three types of rusts and powdery mildew and can be utilized in breeding programs. CIMMWT genotypes possess resistance genes Lr34/Yr18/Sr57/Pm38/Ltn1 (RL6058), Lr67/Yr46/Sr55/Pm46/Ltn3 (RL6077), Lr46/Yr29/Sr58/Pm39/Ltn2 and Sr2/Yr30/Lr27/Pbc (Pavon-F76) (Singh et al. 2005; Kota et al. 2006). Each of these gene pairs is tightly linked or pleiotropic. Cultivars that exhibited lower ACI values at the adult plant stage have race-non-specific resistance (Singh et al. 2005).
Adult plant resistance is polygenic, and at least 2–3 controlling genes remain successful for a longer time (Dehghani and Moghaddam 2004). To address this issue, the phenotypic marker in conjugation with a molecular marker approach was utilized for identifying resistance genes in cultivars in the current study. Phenotypic markers associated with Lr34/Yr18/Sr57/Pm38Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 (LTN) and Sr2/Yr30/Lr27/Pbc (PBC) were observed in 23 Egyptian wheat cultivars. They all exhibited the phenotypic marker LTN, while the PBC marker was recorded with only five cultivars, Misr-1, Misr-2, Sakha-93, Giza-168 and Shandweel-1. These cultivars which possessed PAPR genes, exhibited different levels of APR to rusts and powdery mildew. However, they all exhibited desirable/acceptable to at least two diseases. Phenotypic markers associated with different resistance genes can be used in wheat breeding programs (Hare and McIntosh 1979; Singh 1992; Line 2002). In wheat lines with LTN phenotype, a reduction in leaf and stripe rust severities have been estimated up to 30.5 and 20.8%, respectively (Navabi et al. 2005). Application of phenotypic markers in the screening of wheat cultivars for resistance has been successfully achieved by Juliana et al. (2015), Hussain et al. (2015), Muthe et al. (2016) and Rahmatov et al. (2019).
PCR-based DNA markers have been used by many researchers to verify the presence of resistance genes in wheat (Prabhu et al. 2004; Lagudah et al. 2006; Dakouri et al. 2013). Therefore, molecular markers linked to the tested pleiotropic resistance genes were used to mark the presence or absence of the gene in Egyptian cultivars. Numerous genes conferring resistance to wheat rust and powdery mildew have been identified and used in wheat breeding. Since the 1990s, the wheat germplasm by CIMMYT having minor gene-based resistance has been developed and distributed worldwide (Singh et al. 2000). Four PAPR loci, Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc have been mapped (Suenaga et al. 2003). In Egypt, scientists identified the host material of Egyptian wheat for numerous resistance genes and reported its presence in some of the local genetic stocks (Abdelbacki et al. 2014; Fahmi et al. 2015; Elkot et al. 2016; El-Orabey et al. 2019; Shahin et al. 2020; Draz et al. 2022; Abou-Zeid et al. 2023). In this respect, wheat cultivars exhibited phenotypic markers and postulated to carry pleiotropic resistance genes were screened with molecular markers csLV34, Xwmc44, Xcfd71, and csSr2 for Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, respectively. In this study, molecular data revealed that most cultivars carry at least one or more of the tested resistance genes. Among the cultivars evaluated, there is a considerable variation in their response to rusts and powdery mildew across the years. The presence of a gene based on the molecular markers of a determined cultivar showed a higher level of resistance compared to a cultivar with the absence of a resistance factor. Lr34/Yr18/Sr57/Pm38/Ltn1 was found to be present in six cultivars, Misr-3, Sids-13, Sakha-94, Sakha-95, Giza-168 and Giza-171 based on phenotypic marker LTN (high—very high degree) and molecular marker csLV34 (150-bp) located on chromosome 7D. The six cultivars exhibited good levels of PAPR, ranging from moderate to high, along with moderate seedling resistance of Sakha-94 and Sids-13 to leaf and stem rusts. Two variants of allelic size 150-bp and 229-bp were identified. A strong association was observed with the presence of the Lr34/Yr18/Sr57/Pm38/Ltn1 gene and 150-bp allele. However, cultivars having gene and positive for csLv34 allele 150-bp were rare. The lineage of Lr34/Yr18/Sr57/Pm38/Ltn1 is tracked back to varieties Mentana and Ardito developed in Italy during the early 1990s. CIMMYT cultivar Jupateco-73-R with the resistance gene Lr34/Yr18/Sr57/Pm38 had effective and durable resistance. Gene Lr34/Yr18/Sr57/Pm38/Ltn1 has been used extensively in spring wheat grown in the US, and no complete virulence to this gene has been detected (Kolmer et al. 2008a, b). Lr34/Yr18/Sr57/Pm38/Ltn1, first described by Dyck (1987), has been shown to confer APR (German and Kolmer 1992) and co-segregate with leaf tip necrosis (LTN) (Sing 1992; Spielmeyer et al. 2005; Liang et al. 2006). These multipathotypic resistance traits have made the Lr34/Yr18/Sr57/Pm38/Ltn1 locus one of the highly valuable regions for disease resistance in wheat (Kolmer et al. 2008a, b). The STS marker csLV34 was found to be useful as a molecular marker and reported to be closely linked to Lr34/Yr18/Sr57/Pm38/Ltn1 (Lagudah et al. 2006; Hussain et al. 2015; Muthe et al. 2016; Rahmatov et al. 2019). The presence of Lr34/Yr18/Sr57/Pm38/Ltn1 was largely deduced by the phenotypic response of adult plants to leaf rust and stripe rust and the expression of a leaf tip necrosis phenotype (Kolmer et al. 2008a, b; Sivasamy et al. 2014; Huerta-Espino et al. 2020).
The pleiotropic gene Lr46/Yr29/Sr58/Pm39/Ltn2 was found to be present in Egyptian wheat cultivars, which exhibited LTN (medium) phenotype. It was genotyped by the molecular marker Xwmc44 that amplified a fragment of 242-bp in five cultivars, Misr-1, Misr-2, Shandwel-1, Beniswef-1 and Sohag-4. They all exhibited medium to high levels of PAPR along with moderate resistance of seedlings. This marker Xwmc44 has been reported with the presence of Lr46/Yr29/Sr58/Pm39/Ltn2 in our reference cultivar Pavon-F76 and described its association with LTN phenotype in different cultivars (Suenaga et al. 2003; Rosewarne et al. 2006, 2010; Shah et al. 2010; Skowrońska et al. 2019). Singh et al. (1998) identified Lr46/Yr29/Sr58/Pm39/Ltn2 in the cultivar Pavon-76 and found it located on chromosome 1BL. This gene has been described as one of the most utilized wheat PAPR loci known to confer broad-spectrum resistance to four biotrophic diseases, stripe rust, leaf rust, stem rust (William et al. 2003; Singh et al. 1998, 2013), and powdery mildew (Lillemo et al. 2008). Martinez et al. (2001) reported that infected adult plants carrying Lr46/Yr29/Sr58/Pm39/Ltn2 have a longer latency period as compared to plants without this gene. The plants with this gene show a higher abortion rate of fungal colonies without any chlorotic or necrotic effects and decrease the colony size. Leaf tip necrosis, a phenotypic trait associated with Lr34/Yr18/Sr57/Pm38/Ltn1 and Lr46/Yr29/Sr58/Pm39/Ltn2, was observed among cultivars that showed the highest resistance for leaf rust and yellow rust (Hussain et al. 2015; Skowrońska et al. 2019).
Molecular data also revealed that Lr67/Yr46/Sr55/Pm46/Ltn3 was found to be present in the majority of Egyptian wheat cultivars, which exhibited LTN (slight) phenotype. The marker Xcfd71 amplified 214-bp in 19 cultivars, indicating the presence of Lr67/Yr46/Sr55/Pm46/Ltn3. All of them exhibited moderate to high levels of PAPR. The presence of another resistant gene(s) could be the possible reason for plant resistance against disease, such as Lr67/Yr46/Sr55/Pm46/Ltn3, which is similar in many characteristics to Lr34/Yr18/Sr57/Pm38/Ltn1 (Spielmeyer et al. 2013). The gene Lr67 has been previously identified in Egyptian wheat cultivars but using different molecular markers (Abdelbaki et al. 2014; El-Orabey et al. 2019) that agreed with our data. Moreover, in current study Lr67/Yr46/Sr55/Pm46/Ltn3 additional to Lr34/Yr18/Sr57/Pm38/Ltn1 and Lr46/Yr29/Sr58/Pm39/Ltn2 were identified in newly released cultivars such as Misr-4, Shandweel-2 and Nubaria-2. The presence of Sr2/Yr30/Lr27/Pbc in three Egyptian wheat cultivars, Misr-1, Misr-2, and Shandweel-1, was detected in the current study using the CAPS marker named csSr2 marker, which amplified three fragments 172, 112 and 53-bp tightly linked to Sr2/Yr30/Lr27/Pbc (Mago et al. 2011). These cultivars exhibited a PBC phenotype and moderate to high PAPR. Unlike results by Elkot et al. (2016) who reported the presence of Sr2 in cultivar Giza-168 but using another marker (Xgwm533) which amplified 120-bp. We did not use this marker because it is complicated as there are two different Xgwm533 on 3BS and several lines that did not carry Sr2/Yr30/Lr27/Pbc also amplified a 120-bp product for Xgwm533, which was another SSR allele that differed in sequence (Spielmeyer et al. 2003). Rahmatov et al. (2019) described the association of phenotypic marker PBC and molecular marker csSr2 marker with the PAPR gene Sr2/Yr30/Lr27/Pbc. The gene Sr2/Yr30/Lr27/Pbc located on chromosome 3BS has been transferred to hexaploid wheat from tetraploid emmer wheat cultivar “Yaroslav” in 1920 and used in breeding for around 60 years for durable resistance, including resistance to Ug99. Sr2/Yr30/Lr27/Pbc was detected in semi-dwarf CIMMYT cultivars, such as Pavon-F76, Parula, Kingbird and Dollarbird, exhibited maximum disease severity of 10%-15% with moderately resistant responses. Ellis et al. (2014) reported that the PAPR genes of Sr2/Yr30/Lr27/Pbc and Lr34/Yr18/Sr57/Pm38/Ltn1 in combination with seedling resistance genes have provided durable resistance in several wheat breeding programs. Sr2/Yr30/Lr27/Pbc, in combination with other minor genes, confers effective pleiotropic resistance to stem rust, stripe rust, and leaf rust (Singh et al. 2005 and 2008; Mago et al. 2011; Huerta-Espino et al. 2020).
It might also be beneficial to explore the phenotypic and molecular diversity observed within the studied wheat cultivars. Phenotypic diversity reflects the observable traits of the wheat plants, which can be influenced by both genetic and environmental factors. By considering both genetic and phenotypic diversity, a more holistic understanding of the wheat cultivars and their ability to adapt and improve can be gained. In the current study, identical diversities were observed between each of phenotypic markers, Ltn1, Ltn2 and Ltn3 and each of molecular markers, csLV34, Xwmc44 and Xcfd71, respectively. However, it was a little different between Pbc and csSr2. The highest diversity was recorded with Ltn3, followed by Ltn1 and Ltn2. It suggests a strong correlation between the observable traits and the underlying genetic makeup of the individuals or cultivars. The observation of identical phenotypic and molecular diversity indicates a tight link between the observable traits and the genetic variation present within the wheat cultivars. This correlation suggests that the genetic differences among individuals or cultivars directly contribute to the variation in their phenotypic characteristics. Such a strong association between phenotypic and molecular diversity underscores the importance of genetic factors in shaping the observable traits of wheat plants. This correlation highlights the potential for using molecular markers as proxies for phenotypic traits in breeding programs, enabling breeders to predict and select desired traits based on the genetic profile of the plants. The alignment between phenotypic and molecular diversity streamlines the breeding process by allowing breeders to target specific genetic variants associated with desirable traits. Breeders can leverage molecular markers linked to key phenotypic traits to accelerate the development of improved wheat cultivars with enhanced yield, quality, and resilience to stresses. Additionally, understanding the genetic basis of phenotypic variation facilitates the identification of novel alleles or genetic combinations that could further enhance the adaptive potential and sustainability of wheat production systems.
The tested 23 wheat cultivars that have phenotypic and molecular markers were distinguished based on phenotypic and molecular data using Nei & Li’s coefficient. A complete or high genetic similarity among most cultivars was observed. However, these cultivars also exhibited high genetic diversity based on phenotypic and molecular data. High genetic diversities in the aforementioned cultivars have been previously reported individually for resistance genes of rusts and powdery mildew (El-Orabey et al. 2019; Esmail et al. 2021; Draz et al. 2022; Abou-Zeid et al. 2023). Out of the tested 23 wheat cultivars, 18 cultivars with pleiotropic resistance genes exhibited desirable/acceptable RPRI against at least two diseases. The development of strong resistance in wheat to rusts and powdery mildew at a regional scale requires a high genetic diversity of cultivars in the resistance genes. Breeders should be encouraged to utilize horizontal resistance genes in cultivars in particular that have pleiotropic effects because vertical resistance tends to erode with changes in pathogen virulence (Navabi et al. 2000; Esmail et al. 2021).
The pedigree of wheat genotypes clear that the PAPR genes Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3 and Sr2/Yr30/Lr27/Pbc, have been introgressed into the Egyptian wheat germplasm through local breeding programs from Mexican germplasm provided annually by CIMMYT, Mexico. For instance, Lr34/Yr18/Sr57/Pm38/Ltn1 in Sakha-94 derived from OPATA-85/RAYON-89//KAUZ, MEX were coming from different origins of Mexican germplasm whether OPATA-85 and/or KAUZ, MEX as both carry the gene, while the last-mentioned cultivar likely to be responsible for that gene in Sids-13. Likewise, Lr46/Yr29/Sr58/Pm39/Ltn2 present in Egyptian cultivars also appears to be coming from Mexican germplasm, for instance, Pastor in the pedigree of Misr-1, as well as Pavon-F-76 present in the pedigree of Shandweel-1. The presence of Lr34/Yr18/Sr57/Pm38/Ltn1 and Lr46/Yr29/Sr58/Pm39/Ltn2 in many newly released CIMMYT germplasm was also attributed to cultivar Frontana (Huerta-Espino et al. 2020). Misr-1 and Misr-2 are breeding cultivars originally developed by CIMMYT during the last decade in Ethiopia to possess Sr2/Yr30/Lr27/Pbc for resistance to the Ug99 race (Singh et al. 2011). CIMMYT genotypes have been reported to possess Lr67/Yr46/Sr55/Pm46/Ltn3, such as Chapingo-53, Mayo-54, Yaqui-53, Marroqui-588, Chapingo-48 and Yaqui-50 (Huerta-Espino et al. 2020). These genotypes were found in several crosses in the pedigree of Egyptian wheat cultivars. The presence of Sr2/Yr30/Lr27/Pbc in Shandweel-1 is likely to be attributed to the CIMMYT cultivar Pavon-F76 which carries that gene and is present in the pedigree of that local cultivar Shandweel-1. It is worth noting that the lineage indicates the presence of many other genes with major or minor effectors in the cultivars that may support resistance. Even though many major genes have been detected in various Egyptian wheat cultivars in our previous studies, their efficacy against the diverse pathotypes is limited due to race specificity. For instance, race-specific gene Sr31/Yr9/Lr26/Pm8 was found to be present in many Egyptian cultivars (Shahin et al. 2020), but it was ineffective for resistance to stripe rust and leaf rust as well as that gene (Sr31) is among susceptible genes of a quick detection set for Ug99 race and, therefore, it must be restricted from the use alone in breeding for resistance. The susceptibility of three Egyptian wheat cultivars, Misr-1, Gemmeiza-11, and Sids-12, to stripe rust in the current study could be attributed to the presence of race-specific gene Yr9 (Shahin et al. 2020). By contrast, APR is generally considered long-lasting, but its known genes represent a minority (Chen 2005; Ellis et al. 2014). Our findings emphasized that the multiple disease resistance in cultivars was primarily attributed to the studied genes linked to phenotypic traits, or it may be attributed to the presence of additional not yet cataloged resistance genes.
We observed two sets of observations comparing phenotypic and molecular data with cultivar performance in the field; the first type included those genotypes whose molecular data for the existence of resistance genes corresponded well with cultivar performance in the field. Likewise, the cultivars did not show the presence of resistance genes and were susceptible or low on-field APR. Consequently, the absence of this gene, as revealed by marker data, corresponded well with field expression data. In the second observation group, the two data sets did not match each other, where some of the cultivars, such as Misr-1 and Shandweel-1, indicated the existence of resistance genes in the molecular assay, while they had low APR rates for stripe rust in the field. Such discrepancy in the field results may be due to random mutations; the suppression or evolution of new pathotypes may also be the reason for the inability of cultivars to acclimate to the virulence (Dakouri et al. 2013). Two cultivars, Sakha-93 and Giza-168, did not reveal the presence of resistance gene Sr2/Yr30/Lr27/Pbc locus when screened through molecular markers; however, in the field, those lines exhibited slight PBC and low to moderate resistance against some diseases. This discrepancy has been previously reported in wheat cultivars Cappelle and Desprez, where the presence of Lr34/Yr18/Sr57/Pm38/Ltn1 was observed with an association of leaf and stripe rust resistance (McIntosh 1992) but was not detected using a molecular marker (Lagudah et al. 2009). The failure probability of PCR amplification of the particular band also could cause discrepancies between field and molecular marker data.
The combination of phenotype and genotype evaluation allows for a comprehensive understanding of the resistance mechanisms in wheat cultivars. This information can then be used in breeding programs to select and cross cultivars with desirable traits, ultimately leading to the development of improved wheat varieties that are resistant to rust and powdery mildew. It is important to note that both phenotype and genotype evaluations should be conducted in conjunction to ensure accurate and reliable results. The integration of these approaches in breeding programs enhances the efficiency and success rates of developing disease-resistant wheat cultivars.
Conclusion
The strategy of pleiotropic adult plant resistance in wheat cultivars is the best way to achieve long-lasting resistance under the changing pattern of rust and powdery mildew races in wheat-growing areas. In this study, phenotypic and molecular markers were successfully applied to detect the presence of known PAPR genes Lr34/Yr18/Sr57/Pm38/Ltn1, Lr46/Yr29/Sr58/Pm39/Ltn2, Lr67/Yr46/Sr55/Pm46/Ltn3, and Sr2/Yr30/Lr27/Pbc in Egyptian wheat cultivars. This approach can help predict the field resistance and durability of cultivars and aid decisions in selecting parents for future breeding and development of resistant cultivars. The outstanding cultivars, such as Misr-3, Misr-4, Sakha-94, Sakha-95, Giza-168, Giza-171, Shandweel-2, Nubaria-2 and Sids-13, having a good level of pleiotropic resistance may be used in a breeding program to transfer its resistance character to the adapted wheat cultivars. Integrating genomic information with detailed phenotypic data can provide deeper insights into the genetic architecture of complex traits and facilitate the development of more precise and efficient breeding strategies.
References
Abdelbacki AMM, Omara RI, Najeeb MA, Soliman NE (2014) Identification of leaf rust resistant genes Lr9, Lr25, Lr28, Lr29 and Lr67 in ten Egyptian wheat cultivars using molecular markers. Biotechnol Res Int 2:89
Abdel-Hak TM, EL-Sherif NA, Bassiouny AA, Shafik II, EL-Dauadi YH (1980) Control of wheat leaf rust by systemic fungicides. In: Proceedings of the Fifth European and Mediterranean Cereal Rusts Conference, Bari, Italy (p. 255-266).
Abou-Zeid MA, Mabrouk OI, Draz IS, SaadElDin HI, Safhi FA, ALshamrani SM, Esmail SM (2023) Stem rust appraisal of local and global wheat germplasm with a new virulence threat to resistance genes. Australasian Plant Pathol 52:67–87
Ashmawy MA, El-Orabey WM, Abu Aly AA, Shahin AA (2014) Losses in grain yield of some wheat cultivars infected with powdery mildew. Egypt J Phytopathol 42:71–82
Aslam M (1982) Uniform procedure for development and release of improved wheat varieties. Mimeograph, ARC, Islamabad
Browder LE (1972) A multi-culture inoculation system for study of host-parasite relationships. Plant Dis Rep 56:847–849
Chen XM (2005) Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can J Plant Pathol 27:314–337. https://doi.org/10.1080/07060660509507230
Chen XM (2013) Review article: High-temperature adult-plant resistance, key for sustainable control of stripe rust. Am J Plant Sci 4:608–627. https://doi.org/10.4236/ajps.2013.43080
Dakouri A, McCallum BD, Radovanovic N, Cloutier S (2013) Molecular and phenotypic characterization of seedling and adult plant leaf rust resistance in a world wheat collection. Mol Breed 32:663–677. https://doi.org/10.1007/s11032-013-9899-8
Das MK, Rajaram S, Mundt CC, Kronstad WE, Singh RP (1992) Inheritance of slow rusting resistance in wheat. Crop Sci 32:1452–1456. https://doi.org/10.2135/cropsci1992.0011183X003200060028x
Das MK, Rajaram S, Ktonstad WK, Mundt CC, Singh RP (1993) Associations and genetics of three components of slow rusting in leaf rust of wheat. Euphytica 68(112):99–109. https://doi.org/10.1007/BF00024159
de Vallavieille-Pope C, Ali S, Leconte M, Enjalbert J, Delos M, Rouzet J (2012) Virulence dynamics and regional structuring of Puccinia striiformis f. sp. tritici in France between 1984 and 2009. Plant Dis 96:131–140. https://doi.org/10.1094/PDIS-02-11-0078
Dehghani H, Moghaddam M (2004) Genetic analysis of the latent period of stripe rust in wheat seedlings. J Phytopathol 152:325–330. https://doi.org/10.1111/j.1439-0434.2004.00848.x
Draz IS, Esmail SM, Abou-Zeid M, Hafez YM (2018) Changeability in stripe rust infection and grain yield of wheat associated to climatic conditions. Env Biodiv Soil Security 2:143–153. https://doi.org/10.21608/jenvbs.2019.6674.1040
Draz IS, Esmail SM, Abou-Zeid MA, Essa TA (2019) Powdery mildew susceptibility of spring wheat cultivars as a major constraint on grain yield. Ann Agric Sci 64:39–45. https://doi.org/10.1016/j.aoas.2019.05.007
Draz IS, Elkot AF, Abdelrhim AS (2022) Allelism and resistance loci of powdery mildew and leaf rust in Egyptian hexaploid bread wheat. Cereal Res Commun 50:85–93. https://doi.org/10.1007/s42976-021-00163-z
Dyck PL (1987) The association of a gene for leaf rust resistance with the chromosome 7D suppressor of stem rust resistance in common wheat. Genome 29:467–469. https://doi.org/10.1139/g87-081
Dyck PL, Samborski DJ (1979) Adult plant resistance in PI250413, an introduction of common wheat. Can J Plant Sci 59:329–332. https://doi.org/10.4141/cjps79-053
El-Daoudi YH, Nazim M, Sherif SE, Shenoda Ikhals S, Bassiouni AA (1983) Slow-rusting resistance to stem rust in some Egyptian wheat cultivars. Egyptian Major Crop Improvement Project. Wheat Pathologist Report, June 1983, pp 11
El-Daoudi YH, Ikhlas Shafik, Ghamem EH, Abu El Naga SA, Sherif SO, Khalifa MMO, Mitkees RA, Bassiouni AA (1996) Stripe rust occurrence in Egypt and assessment of grain yield loss in 1995. In: Proceedings Du Symposium Regional Sur les Maladies des Cerales et des Legumineuses Alimentaries 11–14 Nov 1996, Rabat, Maroc
Elkot AFA, Abd El-Aziz MH, Aldrussi IA, El-Maghraby MA (2016) Molecular identification of some stem rust and yellow rust resistance genes in Egyptian wheat and some exotic genotypes. Assiut J Agric Sci 47(4):124–135
Ellis JG, Lagudah E, Spielmeyer W, Dodds P (2014) The past, present and future of breeding rust resistant wheat. Front Plant Sci 5:1–13. https://doi.org/10.3389/fpls.2014.00641
El-Orabey WM, Hamwieh A, Ahmed SM (2019) Molecular markers and phenotypic characterization of adult plant resistance genes Lr 34, Lr 46, Lr 67 and Lr 68 and their association with partial resistance to leaf rust in wheat. J Genet 98:82. https://doi.org/10.1007/s12041-019-1122-1
El-Shamy MM, Mousa MM (2004) Detection of partial resistance components of leaf rust in four Egyptian wheat cultivars. J Agric Res 30(2):116–170
Esmail SM, Draz IS, Ashmawy MA, El-Orabey WM (2021) Emergence of new aggressive races of Puccinia striiformis f. sp. tritici causing yellow rust epiphytotic in Egypt. Physiol Mol Plant Pathol 114:101612. https://doi.org/10.1016/j.pmpp.2021.101612
Fahmi AI, El-Shehawi AM, El-Orabey WM (2015) Leaf rust resistance and molecular identification of Lr34 gene in Egyptian wheat. J Microb Biochem Technol 7(6):338–343. https://doi.org/10.4172/1948-5948.1000236
FAOSTAT (2020). Food and Agriculture Organization of the United Nations (FAO) FAOSTAT. Crop production summary. https://www.fao.org/faostat/en/#data/QCL
German SE, Kolmer JA (1992) Effect of gene Lr34 in the enhancement of resistance to leaf rust of wheat. Theor Appl Genet 84:97–105. https://doi.org/10.1007/BF00223987
Gomma ASA (1968) Report of the Egyptian wheat varieties and their development. Egyptian Society of Genetics Science, Cairo
Griffey CA, Das MK, Stromberg EL (1993) Effectiveness of adult-plant resistance in reducing grain yield loss to powdery mildew in winter wheat. Plant Dis 77(6):618–622
Groth JV, Roelfs AP (1987) The concept and measurement of phenotypic diversity in Puccinia graminis on wheat. Phytopathology 77:1395–1399
Hare RA, McIntosh RA (1979) Genetic and cytogenetic studies of durable adult-plant resistances in ‘Hope’ and related cultivars to wheat rusts. Z Pflanzenzucht 83:350–367
Herrera-Foessel SA, Singh RP, Lillemo M, Huerta-Espino J, Bhavani S, Singh S et al (2014) Lr67/Yr46 confers adult plant resistance to stem rust and powdery mildew in wheat. Theor Appl Genet 127:781–789. https://doi.org/10.1007/s00122-013-2256-9
Hiebert CW, Thomas JB, McCallum BD, Humphreys DG, DePauw RM, Hayden MJ, Mago R, Schnippenkoetter W, Spielmeyer W (2010) An introgression on wheat chromosome 4DL in RL6077 (Thatcher*6/PI 250413) confers adult plant resistance to stripe rust and leaf rust (Lr67). Theor Appl Genet 121:1083–1091. https://doi.org/10.1007/s00122-010-1373-y
Huerta-Espino J, Singh R, Crespo-Herrera LA, Villaseñor-Mir HE, Rodriguez-Garcia MF, Dreisigacker S, Barcenas-Santana D, Lagudah E (2020) Adult plant slow rusting genes confer high levels of resistance to rusts in bread wheat cultivars from Mexico. Front Plant Sci 11:824. https://doi.org/10.3389/fpls.2020.00824
Hulbert SH, Bai J, Fellers JP, Pacheco MG, Bowden RL (2007) Gene expression patterns in near isogenic lines for wheat rust resistance gene Lr34/Yr18. Phytopathology 97(9):1083–1093. https://doi.org/10.1094/PHYTO-97-9-1083
Hussain M, Khan MA, Hussain M, Javed N, Khaliq I (2015) Application of phenotypic and molecular markers to combine genes for durable resistance against rust virulences and high yield potential in wheat. Int J Agric Biol 17:421–430
Juliana P, Rutkoski JE, Poland JA, Singh RP, Murugasamy S, Natesan S, Barbier H, Sorrells ME (2015) Genome-wide association mapping for leaf tip necrosis and pseudo-black chaff in relation to durable rust resistance in wheat. The Plant Genome 8(2):1–12. https://doi.org/10.3835/plantgenome2015.01.0002
Kaur J, Bansal UK, Khanna R, Saini RG, Bariana HS (2009) Molecular mapping of stem rust resistance in HD2009/WL711 recombinant inbred line population. Int J Plant Breed 3:28–33
Kolmer JA, Singh RP, Garvin DF, Viccars L, William HM, Huerta-Espino J, Ogbonnaya FC, Raman H, Orford S, Bariana HS, Lagudah ES (2008a) Analysis of the Lr34/Yr18 rust resistance region in wheat germplasm. Crop Sci 48:1841–1852. https://doi.org/10.2135/cropsci2007.08.0474
Kolmer JA, Singh RP, Gravin DF, Vicaars L, William HM, Huerta-Espino J, Ogbonnayya FC, Raman H, Orford S, Bariana HS, Lagudha ES (2008b) Analysis of Lr34/Yr18 rust resistance region in wheat germplasm. Crop Sci 48:1841–1852. https://doi.org/10.2135/cropsci2007.08.0474
Kota R, Spielmeyer W, McIntosh RA, Lagudah ES (2006) Fine genetic mapping fails to dissociate durable stem rust resistance gene Sr2 from pseudo-black chaff in common wheat (Triticum aestivum L.). Theor Appl Genet 112:492–499. https://doi.org/10.1007/s00122-005-0151-8
Kovach WL (1999) MVSP-A multivariate statistical Package for Windows, ver. 3.1. Kovach Computing Services, Pentraeth, Wales, UK, 137
Kuspira J, Unrau J (1958) Determination of the number and dominance relationships of genes on substituted chromosomes in common wheat Triticum aestivum L. Can J Plant Sci 38:199–205. https://doi.org/10.4141/cjps58-033
Lagudah ES, McFadden H, Singh RP, Huerta-Espino J, Bariana HS, Spielmeyer W (2006) Molecular genetic characterization of the Lr34/Yr18 slow rusting resistance gene region in wheat. Theor Appl Genet 114:21–30. https://doi.org/10.1007/s00122-006-0406-z
Lagudah ES, Krattinger SG, Herrera-Foessel S, Singh RP, Huerta-Espino J, Spielmeyer W, Brown-Guedira G, Selter LL, Keller B (2009) Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor Appl Genet 119:889–898. https://doi.org/10.1007/s00122-009-1097-z
Leath S, Heun M (1990) Identification of powdery mildew resistance genes in cultivars of soft red winter wheat. Plant Dis 74:747–752. https://doi.org/10.1094/PD-74-0747
Li H, Wang X, Song F, Wu C, Wu X, Zhang N, Zhon Y, Zhang X (2011) Response to powdery mildew and detection of resistance genes in wheat cultivars from China. Acta Agron Sin 37:943–954. https://doi.org/10.1016/S1875-2780(11)60026-6
Liang SS, Savenaga K, He ZH, Wang ZL, Liu HY, Wang DS, Singh RP, Sourdille P, Xia YC (2006) Quantitative trait loci mapping for adult-plant resistance to powdery mildew in bread wheat. Phytopathology 96:784–789. https://doi.org/10.1094/PHYTO-96-0784
Lillemo M, Asalf B, Singh RP, Huerta-Espino J, Chen XM, He ZH, Bjørnstad A (2008) The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theor Appl Genet 116(8):1155–1166. https://doi.org/10.1007/s00122-008-0743-1
Line RF (2002) Stripe rust of wheat and barley in North America: a retrospective historical review. Ann Rev Phytopathol 40:75–118. https://doi.org/10.1146/annurev.phyto.40.020102.111645
Mago R, Brown-Guedira G, Dreisigacker S, Breen J, Jin Y, Singh RP, Appels R, Lagudah ES, Ellis J, Spielmeyer W (2011) An accurate DNA marker assay for stem rust resistance gene Sr2 in wheat. Theor Appl Genet 122(4):735–744. https://doi.org/10.1007/s00122-010-1482-7
Martinez F, Nicks RE, Singh RP, Rubiales D (2001) Characterization of Lr46, a gene conferring partial resistance to wheat leaf rust. Hereditas 135:111–114. https://doi.org/10.1111/j.1601-5223.2001.00111.x
McFadden ES (1939) Brown necrosis, a discolouration associated with rust infection in certain rust resistant wheats. J Agric Res 58:805–819
McIntosh RA (1992) Close genetic linkage of genes conferring adult-plant resistance to leaf rust and stripe rust in wheat. Plant Pathol 41:523–527. https://doi.org/10.1111/j.1365-3059.1992.tb02450.x
McIntosh RA, Wellings CR, Park RF (1995) Wheat rusts: an atlas of resistance genes. CSIRO, Australia
McIntosh RA, Yamazaki Y, Devos KM, et al. (2003). Catalogue of gene symbols for wheat. In: Pogna NE, Romano M, Pogna A, Galterio G, editors. In: Proceedings of the 10th International Wheat Genetics Symposium; 2003 Sep 1–6; Istituto Sperimentale per la Cerealicoltura; Paestum, Italy
McNeal FH, Konzak CF, Smith EP, Tate WS, Russell TS (1971) A uniform system for recording and processing cereal data. USDA Agric Res Serv Bull 42:34–121
Muthe ST, Kulwal PL, Gadekar DA, Jadhav AS (2016) Molecular marker based detection of leaf rust resistance gene Lr34 in Indian bread wheat (Triticum aestivum L.). Australas Plant Pathol 45(4):369–376. https://doi.org/10.1007/s13313-016-0423-6
Navabi A, Singh RP, Tewaril JP, Briggs KG (2000) Polygenic inheritance of adult plant resistance to stripe rust (Puccinia striiformis) and leaf rust (P. triticina) in spring wheat. International Symposium Durable Rust Resistance Key to Sustainable Agriculture, pp: 4−32. Nov. 28 – Dec., 1, 2000
Navabi A, Tewari JP, Singh RP, McCallum B, Laroche A, Briggs KG (2005) Inheritance and QTL analysis of durable resistance to stripe and leaf rusts in an Australian cultivar, Triticum aestivum 'Cook'. Genome 48(1):97–107. https://doi.org/10.1139/g04-100
Njau PN, Bhavani S, Huerta-Espino J, Keller B, Singh RP (2013) Identification of QTL associated with durable adult plant resistance to stem rust race Ug99 in wheat cultivar ‘Pavon 76.’ Euphytica 190:33–44. https://doi.org/10.1007/s10681-012-0763-4
Parlevliet JE (1975) Partial resistance of barley to leaf rust, Puccinia hordei. I. Effect of cultivar and development stage on latent period. Euphytica 24:21–27. https://doi.org/10.1007/BF00147164
Pathan AK, Park RF (2006) Evaluation of seedling and adult plant resistance to leaf rust in European wheat cultivars. Euphytica 149:327–342. https://doi.org/10.1007/s10681-005-9081-4
Pearson ES, Hartley HO (1970) Biometrika tables for statisticians, 3rd edn. Cambridge University Press, Cambridge
Peterson RF, Campbell AB, Hannah AE (1948) A diagrammatic scale for estimating rust severity on leaves and stems of cereals. Can J Genet Cytol 26:496–500. https://doi.org/10.1139/cjr48c-033
Prabhu KV, Gupta SK, Charpe A, Koul S (2004) SCAR marker tagged to the alien leaf rust resistance gene Lr19 uniquely marking the Agropyron elongatum-derived gene Lr24 in wheat: a revision. Plant Breed 123:417–420. https://doi.org/10.1111/j.1439-0523.2004.00971.x
Pretorius ZA, Singh RP, Wagoire WW, Payne TS (2000) Detection of virulence to wheat stem rust resistance gene Sr31 in Puccinia graminis f. sp. tritici in Uganda. Plant Dis 84:203. https://doi.org/10.1094/PDIS.2000.84.2.203B
Rahmatov M, Otambekova M, Muminjanov H, Rouse MN, Hovmøller MS, Nazari K, Steffenson BJ (2019) Characterization of stem, stripe and leaf rust resistance in Tajik bread wheat accessions. Euphytica 215(55):1–22. https://doi.org/10.1007/s10681-019-2377-6
Rehman AU, Sajjad M, Khan SH, Ahmad N (2013) Prospects of wheat breeding for durable resistance against brown, yellow and black rust fungi. Int J Agric Biol 15:1209–1220
Roelfs AP, Singh RP, Saari EE (1992) Rust Diseases of Wheat: Concepts and Methods of Disease Management. CIMMYT, Mexico, p 81
Rosewarne GM, Singh RP, Huerta-Espino J, William HM, Bouchet S, Cloutier S, McFadden H, Lagudah ES (2006) Leaf tip necrosis, molecular markers and b1-proteasome subunits associated with the slow rusting resistance genes Lr46/Yr29. Theor Appl Genet 112:500–508. https://doi.org/10.1007/s00122-005-0153-6
Rosewarne GM, Singh RP, William W, Huerta-Espino J (2010) Identification of phenotypic and molecular markers associated with slow rusting resistance gene Lr46. In: 11th international cereal rusts and powdery mildews conference, Norwich, 22–27 September 2004
Rubiales D, Niks RE (1995) Characterization of Lr34 a major gene conferring nonhypersensitive resistance to wheat leaf rust. Plant Dis 79:1208–1212. https://doi.org/10.1094/PD-79-1208
Saari EE, Prescott JM (1975) A scale for appraising the foliar intensity of wheat diseases. Plant Dis Rep 59:377–380
Saari EE, Wilcoxson RD (1974) Plant disease situation of high yielding durum wheat in Asia and Africa. Ann Rev Phytopathol 2:49–68. https://doi.org/10.1146/annurev.py.12.090174.000405
Shah SJA, Imtiaz M, Hussain S (2010) Phenotypic and molecular characterization of wheat for slow rusting resistance against Puccinia striiformis Westend. f. sp. tritici. J Phytopathol 158:393–402. https://doi.org/10.1111/j.1439-0434.2009.01631.x
Shahin AA, Hasan MA, Abou-Zeid MA (2018) Adult plant resistance to stem rust and molecular marker analysis of some Egyptian and exotic bread wheat genotypes. J Plant Prot Path 9(12):901–907
Shahin AA, Draz IS, Esmail SM (2020) Race specificity of stripe rust resistance in relation to susceptibility of Egyptian wheat cultivars. Egypt J Phytopathol 48:1–13. https://doi.org/10.21608/EJP.2020.107601
Sharma RC, Duveiller E (2007) Advancement toward new spot blotch resistant wheats in South Asia. Crop Sci 47:961–968. https://doi.org/10.2135/cropsci2006.03.0201
Shi AN, Leath S, Murphy JP (1998) A major gene for powdery mildew resistance transferred to common wheat from wild einkorn wheat. Phytopathol 88:144–147. https://doi.org/10.1094/PHYTO.1998.88.2.144
Singh RP (1992) Association between gene Lr34 for leaf rust resistance and leaf tip necrosis in wheat. Crop Sci 32:874–878. https://doi.org/10.2135/cropsci1992.0011183X003200040008x
Singh RP, Mujeeb-Kazi A, Huerta-Espino J (1998) Lr46: a gene conferring slow rusting resistance to leaf rust in wheat. Phytopathology 88:890–894. https://doi.org/10.1094/PHYTO.1998.88.9.890
Singh RP, Huerta-Espino J, Rajaram S (2000) Achieving near-immunity to leaf rust and stripe rust in wheat by combining slow rusting resistance genes. Acta Phytopathol Entomol 35:133–139
Singh RP, Huerta-Espino J, Roelfs AP (2002) The wheat rusts. In: Curtis BC, Rajaram S, Gomez Macpherson H (eds) Bread wheat: improvement and production Plant Production and Protection Series no. 30. FAO, Rome, pp 317–330
Singh RP, Huerta-Espino J, William HM (2005) Genetics and breeding for durable resistance to leaf and stripe rusts in wheat. Turk J Agric 29:121–127
Singh RP, Hodson DP, Huerta-Espino J, Jin Y, Njau P, Wanyera R, Herrera-Foessel SA, Ward RW (2008) Will stem rust destroy the world’s wheat crop? Adv Agron 98:271–309. https://doi.org/10.1016/S0065-2113(08)00205-8
Singh RP, Herrera-Foessel S, Huerta-Espino L, Bariana H, Bansal U, Mccallum B, Hiebert C, Bhavani S, Singh S, Lan C, Lagudah E (2012) Lr34/Yr18/Sr57/Pm38/Bdv1/Ltn1 Confers slow rusting, adult plant resistance to Puccinia graminis tritici. In: W.-Q. Chen, editor, Proc. of the 13th Int. Cereal Rusts and Powdery Mildews Conf., Beijing, China. 28 Aug.–1 Sept. 2012, China Agricultural Science and Technology Press, Beijing
Singh RP, Herrera-Foessel SA, Huerta-Espino J, Lan CX, Basnet BR, Bhavani S et al. (2013) Pleiotropic gene Lr46/Yr29/Pm39/Ltn2 confers slow rusting, adult plant resistance to wheat stem rust fungus. In: Proceedings of the 2013 Technical Workshop: Borlaug Global Rust Initiative, New Delhi
Singh RP, Hodson DP, Huerta‒Espino J, Jin Y, Bhavani S, Njau P, Herrera-Foessel SA, Singh PK, Singh S, Govindan V (2011) The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu Rev Phytopathol 49(1):465–481. https://doi.org/10.1146/annurev-phyto-072910-095423
Sivasamy M, Aparna M, Kumar J, Jayaprakash P, Vikas VK, John P, Nisha R, Sivan SK, Punniakotti E (2014) Phenotypic and molecular confirmation of durable adult plant leaf rust resistance (APR) Genes Lr34+, Lr46+ and Lr67+ linked to leaf tip necrosis (LTN) in select registered Indian wheat (T. aestivum) genetic stocks. Cereal Res Commun 42:262–273. https://doi.org/10.1556/CRC.2013.0054
Skowrońska R, Kwiatek M, Tomkowiak A, Nawracała J (2019) Development of multiplex PCR to detect slow rust resistance genes Lr34 and Lr46 in wheat. J Appl Genet 60:301–304. https://doi.org/10.1007/s13353-019-00520-z
Spielmeyer W, Sharp PJ, Lagudah ES (2003) Identification and validation of markers linked to broad-spectrum stem rust resistance gene Sr2 in wheat Triticum aestivum L. Crop Sci 43:333–336. https://doi.org/10.2135/cropsci2003.3330
Spielmeyer W, McIntosh RA, Kolmer J, Lagudah ES (2005) Powdery mildew resistance and Lr34/Yr18 genes for durable resistance to leaf and stripe rust cosegregate at a locus on the short arm of chromosome 7D of wheat. Theor Appl Genet 111:731–735. https://doi.org/10.1007/s00122-005-2058-9
Spielmeyer W, Mago R, Wellings C, Ayliffe M (2013) Lr67 and Lr34 rust resistance genes have much in common, they confer broad spectrum resistance to multiple pathogens in wheat. BMC Plant Biol 13:96. https://doi.org/10.1186/1471-2229-13-96
Stakman EC, Stewart DM, Loegering WQ (1962) Identification of physiologic races of Puccinia graminis tritici. ARS, USDA, Agric. Res. Serv. Bull. E-617. 53
Steel RGD, Torrie JH (1980) Principles and Procedures of Statistics, 2nd ed. Mc Graw Hill Book Company, New York.
Stubbs RW (1988) Pathogenicity analysis of yellow (stripe) rust of wheat and its significance in a global context. Breeding strategies for resistance to the rusts of wheat. Eds N.W. Simmonds and S. Rajaram (Mexico: CIMMYT D.F.), ISBN (1988), 96–127
Suenaga K, Singh RP, Huerta-Espino J, William HM (2003) Microsattelite markers for gene Lr34/Yr18 and other quantitative trait loci for leaf rust and stripe rust resistance in bread wheat. Phytopathology 93:881–889. https://doi.org/10.1094/PHYTO.2003.93.7.881
Vanegas CDG, Garvin DF, Kolmer JA (2008) Genetics of stem rust resistance in the spring wheat cultivar Thatcher and the enhancement of stem rust resistance by Lr34. Euphytica 159:391–401. https://doi.org/10.1007/s10681-007-9541-0
Wanyera R, Kinyua MG, Jin Y, Singh RP (2006) The spread of stem rust caused by Puccinia graminis f. sp. tritici, with virulence on Sr31 in wheat in Eastern Africa. Plant Dis 90:113. https://doi.org/10.1094/PD-90-0113A
William HM, Singh RP, Huerta-Espino J, Ortiz-Islas S, Hoisington D (2003) Molecular marker mapping of leaf rust resistance gene Lr46 and its association with stripe rust gene Yr29 in wheat. Phytopathology 93(2):153–159
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for the growth stages of cereals. Weed Res 14:415–421. https://doi.org/10.1111/j.1365-3180.1974.tb01084.x
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The authors declare that no funds, grants, or other support were received for setting up the experiment and conducting all the assays discussed in the study as well as during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
Conceptualization: ISD, SME; methodology and validation: ISD, SME, DAK; formal analysis: ISD, DAK; writing original draft preparation: ISD; writing review and editing: ISD, SME, DAK; administration: ISD, SME; All authors reviewed the manuscript and approved the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial competing interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Draz, I.S., Esmail, S.M. & Komeil, D.A. Phenotypic and molecular characterization of pleiotropic resistance to rusts and powdery mildew in spring wheat. Euphytica 220, 99 (2024). https://doi.org/10.1007/s10681-024-03362-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-024-03362-x