
Abstract
Background
Acclimatization to winter conditions is an essential prerequisite for the survival of small birds in the northern temperate zone. Changes in photoperiod, ambient temperature and food availability trigger seasonal physiological and behavioral acclimatization in many passerines. Seasonal trends in metabolic parameters are well known in avian populations from temperate environments; however, the physiological and biochemical mechanisms underlying these trends are incompletely understood. In this study, we used an integrative approach to measure variation in the thermogenic properties of the male Silky Starling (Sturnus sericeus) at different levels or organization, from the whole organism to the biochemical. We measured body mass (M b), basal metabolic rate (BMR), energy budget, the mass of selected internal organs, state 4 respiration and cytochrome c oxidase (COX) activity in the heart, liver and muscle.
Methods
Oxygen consumption was measured using an open-circuit respirometry system. The energy intake of the birds were then determined using an oxygen bomb calorimeter. Mitochondrial state 4 respiration and COX activity in heart, liver and pectoral muscle were measured with a Clark electrode.
Results
The results suggest that acclimatization to winter conditions caused significant change in each of the measured variables, specifically, increases in M b, organ mass, BMR, energy intake and cellular enzyme activity. Furthermore, BMR was positively correlated with body mass, energy intake, the mass of selected internal organs, state 4 respiration in the heart, liver and muscle, and COX activity in the heart and muscle.
Conclusions
These results suggest that the male Silky Starling’s enhanced basal thermogenesis under winter conditions is achieved by making a suite of adjustments from the whole organism to the biochemical level, and provide further evidence to support the notion that small birds have high phenotypic plasticity with respect to seasonal changes.
Similar content being viewed by others


Background
The thermogenic properties of animals are closely related to energy utilization (Webster and Weathers 2000; Vézina et al. 2006, 2007), energy distribution (Dawson and Marsh 1986), life-history strategy and evolutionary history (Diamond 1998; Wiersma et al. 2007; Swanson 2010). Furthermore, the thermogenesis of birds reflects their physiological capacity to adapt to environmental change (Swanson 1991a; McKechnie 2008; Zheng et al. 2013a, 2014a). Many investigators have studied the relationship between survival and the thermoregulation in birds and small mammals in tropical and temperate regions (Weathers 1979; Wikelski et al. 2003; Wiersma et al. 2007; Zheng et al. 2008a; Smit and McKechnie 2010; Swanson 2010; Nilsson and Nilsson 2016; Petit et al. 2017). In temperate regions, cold temperatures and reduced food availability affect the energy budgets of winter-active birds. In response, birds have evolved a range of morphological, physiological and behavioral mechanisms to enable them to adapt to the greater seasonal variation in climate experienced in temperate regions (Swanson 1990, 1991b; Zheng et al. 2008b, 2014b). Phenotypic flexibility refers to phenotypic changes that are reversible, temporary, and repeatable (Piersma and Drent 2003; Starck and Rahmaan 2003; Tieleman et al. 2003; McKechnie 2008). The study of phenotypic flexibility has become a central topic in evolutionary ecology and resident endothermic animals in temperate climates are ideal subjects for associated natural experiments (McKechnie et al. 2006; Liknes and Swanson 2011; Swanson et al. 2014). One example of phenotypic flexibility is the seasonal phenotypes developed by some small birds in response to seasonal climatic variation (Swanson 2010). Many small birds inhabiting seasonal environments increase body mass with winter acclimatization (McKechnie and Swanson 2010). The winter increase in body mass could result from increases in fat deposits and/or lean mass and thereby influence thermogenic demands (Cooper 2007; Zheng et al. 2014a). The winter phenotype of such birds is characterized by improved cold tolerance and enhanced capacity for thermogenesis, increases in both basal metabolic rate (BMR; minimum metabolic rate) and summit metabolic rate (M sum; maximal thermoregulatory metabolic rate) (Swanson 1990, 1991a; McKechnie and Wolf 2004; Swanson and Garland 2009; McKechnie and Swanson 2010). BMR, a widely-accepted benchmark of metabolic expenditure in endotherms, is correlated with the energetic cost of thermoregulation and (McKechnie and Wolf 2004; McNab 2009). BMR is thought to increase in winter because of the higher metabolic cost of supporting thermogenic tissues (McKechnie 2008; Zheng et al. 2013a, 2014a; Swanson et al. 2017), whereas elevated M sum is directly correlated with improved cold tolerance in winter birds (Swanson 2001; Swanson and Liknes 2006).
Thermogenesis, or thermoregulation, in small birds is an example of a flexible trait that can be seasonally modulated through adjustments at several levels of organization (McKechnie 2008; Swanson 2010; Petit et al. 2014). At the organismal level, winter acclimatized birds, often, but not always, have a higher BMR than summer acclimatized birds (Swanson 1990, 1991a; Zheng et al. 2008a, 2014a). At the organ level, alternation in the size, or mass, of metabolically active tissues and organs, such as the liver, kidneys, digestive tract, or muscles, can influence BMR (Brand et al. 2003; Zheng et al. 2008b, 2014a; Liknes and Swanson 2011). At the biochemical level, changes in catabolic enzyme activity, and or, the capacity for metabolic substrate transport, can influence the mass-specific metabolic activity of organs, thereby affecting organismal metabolic rates (Swanson 1991b; Liu et al. 2008; Zheng et al. 2008b, 2014a; Liknes and Swanson 2011). Variation in cellular metabolic intensity is often measured by examining variation in state 4 respiration (reflecting oxidative phosphorylation capacity) or cytochrome c oxidase (COX) activity (a key regulatory enzyme of oxidative phosphorylation) (Zheng et al. 2008b, 2014a; Swanson 2010; Zhou et al. 2016; Hu et al. 2017). Seasonal changes in thermogenesis at the organismal level have been well documented in several species under natural conditions (Dawson and Carey 1976; Southwick 1980; Liknes and Swanson 1996; Smit and McKechnie 2010; Zheng et al. 2008b, 2014a; Wu et al. 2015). For instance, body mass and BMR in Chinese Bulbuls (Pycnonotus sinensis) were higher in spring and winter than in summer. The dry masses of several nutritional organs and mitochondrial protein content, state 4 respiration, and COX activity in liver and muscle were all heightened in winter relative to other seasons, and this enhanced performance has been linked to regulatory changes in nutritional and exercise organ masses and liver and muscle cellular aerobic capacity interact to promote seasonal metabolic flexibility in Chinese Bulbuls (Zheng et al. 2008b). Similarly, Eurasian Tree Sparrows (Passer montanus) living at high latitude exhibit greater thermogenic capacities compared with their low latitude counterparts. Consistently, state 4 respiration and COX activity in liver and muscle were remarkably higher with latitude. These results are consistent with a pronounced latitudinal phenotypic flexibility mediated through physiological and biochemical adjustments in Eurasian Tree Sparrows (Zheng et al. 2014b). Because changes in thermogenic regulation allow the organisms to rapidly and reversibly respond to environmental variation, the study in thermoregulation can provide insights into the mechanistic underpinnings of phenotypic flexibility and the adaptive modification of complex physiological traits, the hierarchical levels of physiological traits is well illustrated of physiological adjustment that enhance thermogenic performance (Swanson 2010; Swanson et al. 2017; Maria et al. 2015; Zhou et al. 2016).
The Silky Starling (Sturnus sericeus; Passeriformes, Sturnidae) is a resident species in vast areas of southern and southeastern China, South Korea, North Vietnam and the Philippines (MacKinnon and Phillipps 2000). Its preferred habitat is broadleaf, and conifer-broadleaf mixed forest, but it can also be found in orchards and farmland. The Silky Starling is omnivorous and primarily feeds on insects, fruits and seeds. Studies have found that male Silky Starlings have a larger body mass than females, and a higher body temperature, higher thermal conductance, and a lower BMR with a relatively wide thermal neutral zone (Zhang et al. 2006; Zhao et al. 2013). The Silky Starling also has high metabolic water production (MWP) and evaporative water loss (EWL) (Bao et al. 2014). However, understanding seasonal changes in thermogenesis at the organismal level in this species has been hampered by the lack of research on cellular responses to seasonal change. To resolve this problem we investigated a broad range of thermogenic responses to temperature in the Silky Starling, from the cellular to the organismal level. In addition, we measured M b, BMR and the energy budget of individual birds, as well as changes in the mass of the liver, heart, pectoral muscle and digestive tract (gizzard, small intestine and rectum). We also measured cellular level state 4 respiration and COX activity in the heart, liver, and pectoral muscle. We hypothesized that the Silky Starling would acclimatize to winter conditions by increasing M b, BMR, energy budget and internal organs mass. At the cellular level, we predicted that winter acclimatized birds would increase their BMR by activating state 4 respiration and increasing COX activity in the heart, liver, and pectoral muscle.
Methods
Study site
This study was carried out in Wenzhou City, Zhejiang Province (27°29′N, 120°51′E, 14 m in elevation), China. Eighteen male Silky Starlings (male Silky Starling has whitish head contrasting with female) were live-trapped in forest between July 2013 and January 2014. Nine birds were caught during summer (hereafter referred to as “summer birds”) and nine in winter (hereafter “winter birds”). The climate of Wenzhou is warm-temperate with an average annual rainfall of 1500 mm spread across all months, and slightly more precipitation during spring and summer. Mean ambient temperature ranged from 31.3 ± 0.2 °C in summer (July 2013) to 8.6 ± 0.4 °C in winter (January 2014). Daily minimum and maximum ambient temperatures vary greatly, ranging from 25.4 to 38.6 °C in summer, and from 0.9 to 14.8 °C in winter (Wenzhou Bureau of Meteorology). M b to the nearest 0.1 g was determined immediately upon capture with an electronic balance (Sartorius BT25S, Germany). Birds were then transported to the laboratory and kept outdoors for 1 or 2 days in 50 cm × 30 cm × 20 cm cages under natural photoperiod and temperature before physiological measurements were taken (Zhang et al. 2006). Food and water were supplied ad libitum. All experimental procedures were approved by the Animal Care and Use Committee of the Wenzhou University (Protocol 20130012).
Measurement of metabolic rate
Birds’ metabolic rates were estimated by measuring their oxygen consumption in an open-circuit respirometry system (AEI technologies S-3A/I, USA). Metabolic chambers were 1.5 L in volume, made of plastic, and provided with a perch for the bird to stand on (Smit and McKechnie 2010). Chamber temperature was regulated by a temperature-controlled cabinet (Artificial climatic engine BIC-300, China) capable of regulating temperature to ± 0.5 °C. Water vapour and CO2 were scrubbed from the air passing through the chamber in a silica gel/soda lime/silica column before passing through the oxygen analyzer. We measured the oxygen content of excurrent gas from metabolic chambers with an oxygen sensor (AEI technologies N-22M, USA). We used a flow control system (AEI technologies R-1, USA) to set the flow of excurrent gas. The pump was located downstream of the metabolic chamber and air pulled through the chamber at 300 mL/min by the pump during metabolic rate measurements (Zheng et al. 2014a; Wu et al. 2015). This maintained a fractional concentration of O2 in the respirometry chamber of about 20%, calibrated to ± 1% accuracy with a general purpose thermal mass flow-meter (TSI 4100 Series, USA) (McNab 2006). Oxygen consumption rates were measured at 25 ± 0.5 °C, which is within the thermal neutral zone of the Silky Starling (Bao et al. 2014). Baseline O2 concentrations were obtained before and after each test (Li et al. 2010; Wu et al. 2015). All measurements of gas exchange were obtained during the resting-phase of birds’ circadian cycles (between 20:00 and 24:00) in darkened chambers when individual birds could reasonably be expected to be post-absorptive. Food was removed 4 h before each measurement to minimize the heat increment associated with feeding (Zhang et al. 2006; Bao et al. 2014). We first ensured that birds were perching calmly in the chamber and started recording oxygen consumption at least 1 h later. Each animal was generally in the metabolic chamber for at least 2 h. The data obtained were used to calculate 5 min running means of instantaneous oxygen consumption over the entire test period using equation 2 of Hill (1972). The lowest 5 min mean recorded over the test period was considered the resting metabolic rate (Smit and McKechnie 2010). All values for oxygen consumption were expressed as mL O2/h and corrected to STPD conditions (Christians 1999; Schmidt-Nielsen 1997). Body temperature (T b) was measured during metabolic measurements with a lubricated thermocouple inserted into the cloaca, the output of which was digitized by an Oakton thermocouple meter (Eutech Instruments, Singapore). M b was measured to the nearest 0.1 g before and after experiments; mean M b was used in calculations. All measurements were made daily between 20:00 and 24:00.
Energy budget
The energy intake of the birds caught in summer and winter was measured over 3 days according to previously established methods (Klaassen et al. 2004; Wu et al. 2014). During each test, subjects were housed individually in cages, where food was provided in excess of the animals’ needs and water was provided ad libitum. The uneaten food and feces were collected after the 3-day period, separated manually and oven-dried at 65 °C for at least 72 h. The caloric contents of the dried food and feces were then determined using a C 2000 oxygen bomb calorimeter (IKA Instrument, Germany). Gross energy intake (GEI), fecal energy (FE), digestible energy intake (DEI) and digestibility of energy were calculated according to Li and Wang (2005) and Wu et al. (2014):
Measurements of organ masses
Birds were euthanized by cervical dislocation at the end of the experiment and their heart, liver, pectoral muscle, gizzard, small intestine and rectum extracted and weighed to the nearest 0.1 mg. Part of the heart, liver and muscle was used to investigate state 4 respiration and COX activity (Zheng et al. 2008b, 2014a), but the other internal organs, including the remainder of the heart, liver and pectoral muscle, were dried to a constant mass over 2 days at 65 °C and weighed to the nearest 0.1 mg (Williams and Tieleman 2000; Liu and Li 2006).
Measurements of tissue state 4 respiration and cytochrome c oxidase (COX) activity
State 4 respiration in the heart, liver and pectoral muscle was measured at 30 °C in 1.96 mL of respiration medium (225 mM sucrose, 50 mM Tris/HCl, 5 mM MgCl2, 1 mM EDTA and 5 mM KH2PO4, pH 7.2) with a Clark electrode (Hansatech Instruments LTD., England, DW-1), essentially as described by Estabrook (1967). State 4 respiration was measured over a 1 h period under substrate dependent conditions, with succinate as the substrate (Zheng et al. 2008b, 2010, 2013b). COX activity in the heart, liver and pectoral muscle were measured polarographically at 30 °C using a Clark electrode according to Sundin et al. (1987). We express state 4 respiration and COX activity measurements in terms of whole organ activity (µmol O2/min per organ) (Wiesinger et al. 1989; Zheng et al. 2013b, 2014a; Zhou et al. 2016).
Statistics
Data were analyzed using SPSS (version 19.0). The normality of all variables was assessed using the Kolmogorov–Smirnov test and any non-normally distributed data were normalized by log-transformation prior to statistical analysis. Direct comparisons of the M b and body temperature of starlings acclimatized to either winter or summer were made with Student t test. Because digestive efficiency consists of percentage data, these are not typically normally distributed, so we used arcsin-square root transformed prior to statistical analysis. Changes in metabolic rate, GEI, FE and DEI were analyzed by ANCOVA with tarsus length as a covariate. Differences in state 4 respiration and COX activity between birds caught in winter and summer were evaluated with Student t test where appropriate. Least-squares linear regression was used to test for allometric correlations between the log masses of different organs, log BMR and log M b. For organ mass allometric regressions we used body mass minus wet organ mass for the organ in question to avoid statistical problems with part-whole correlations (Christians 1999). Residuals were calculated from allometric equations and linear regression was used to determine if log organ mass residuals were significantly correlated with those of log BMR. Least-squares linear regression was also used to evaluate the relationship between log BMR and log M b, and between log BMR, log state 4 respiration and log COX. All results were expressed as mean ± SEM; p < 0.05 was considered statistically significant.
Results
Body mass (M b) and basal metabolic rate (BMR)
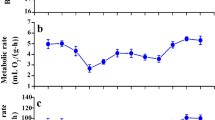
Winter birds had significantly higher M b and BMR than summer birds (M b, t 16 = 3.850, p < 0.01, Fig. 1a; BMR, t 16 = 6.446, p < 0.001, Fig. 1b); the BMR of winter birds exceeded that of their summer counterparts by 33%. After adjusting for body size by using tarsus length as a covariate, the BMR of winter birds averaged 262.2 ± 9.7 O2/h, still significantly higher than that of the summer birds (214.8 ± 9.7 O2/h; F 1,15 = 8.185, p < 0.05). There was a significant, positive correlation between log M b and log BMR (r 2 = 0.709, p < 0.001; Fig. 1c). There was no significant difference in body temperature between winter and summer birds (winter, 42.2 ± 0.5 °C; summer, 41.6 ± 0.5 °C; t 16 = 0.833, p > 0.05).

Body mass (a), resting metabolic rate (b), and the correlation between body mass and basal metabolic rate (c), of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China. Data are shown as mean ± SEM, **p < 0.01 and ***p < 0.001
Energy budget
GEI, FE and DEI were all significantly higher in winter birds than in summer birds (GEI: ANCOVA, F 1,15 = 11.744, p < 0.01, Fig. 2a: FE: ANCOVA, F 1,15 = 6.914, p < 0.05, Fig. 2b; DEI: ANCOVA, F 1,15 = 4.681, p < 0.05, Fig. 2c). GEI and DEI of winter birds were 27 and 24%, respectively, higher than those of summer birds. There was, however, no significant difference between winter and summer birds in digestibility (t 16 = 0.450, p = 0.659; Fig. 2d). There were positive correlations between log GEI and log M b (Fig. 3a), between log DEI and log M b (Fig. 3b), between log GEI and log BMR (Fig. 3c), and between log DEI and log BMR (Fig. 3d).

Gross energy intake (a), fecal energy (b), digestible energy intake (c), and digestibility (d), of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China. Data are shown as mean ± SEM, *p < 0.05 and **p < 0.01. GEI gross energy intake, FE fecal energy, DEI digestible energy intake

Correlation between the GEI, body mass and basal metabolic rate, and between the DEI, body mass and basal metabolic rate, of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China
Organ mass
Winter birds had a significantly heavier gizzard (ANCOVA, F 1,15 = 17.599, p < 0.01), smaller intestine (ANCOVA, F 1,15 = 12.303, p < 0.01), and total digestive tract (ANCOVA, F 1,15 = 17.567, p < 0.01), than summer birds (Fig. 4a). There were positive correlations between log gizzard mass and log GEI (Fig. 4b), and between log small intestine mass and log DEI (Fig. 4c). Winter birds also had a significantly heavier heart (ANCOVA, F 1,15 = 9.740, p < 0.01), liver (ANCOVA, F 1,15 = 28.146, p < 0.001), and pectoral muscle (ANCOVA, F 1,15 = 17.387, p < 0.01), than summer birds (Fig. 5a). The slopes of allometric relationships between log organ mass and log M b (minus wet mass of the organ) were positive for all organs; however, those for heart, pectoral muscle and gizzard were statistically significant (Table 1). The slopes of the statistically significant allometric regression lines were all > 1.0, indicating that the increase of organ mass with body size was greater than the increase in overall M b (Table 1). There was a significant, positive correlation between log BMR residuals and log organ mass residuals for all organs except the heart and small intestine. However, the correlation between BMR and heart mass residuals was nearly significant (Table 1).

Mass of the gizzard, small intestine, rectum and total gut (a), relationship between the GEI and gizzard mass (b), and between the DEI and small intestine mass (c), of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China. Data are shown as mean ± SEM, **p < 0.01. GEI gross energy intake, DEI digestible energy intake

Dry mass (a), state 4 respiration (b), and cytochrome c oxidase activity (c), in the heart, liver and pectoral muscle of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China. Data are shown as mean ± SEM, **p < 0.01 and ***p < 0.001
Tissue state 4 respiration and COX activity
Winter birds had significantly higher state 4 respiration in the heart (mass-specific, t 16 = 4.829, p < 0.001; organ, t 16 = 4.266, p < 0.01), liver (mass-specific, t 16 = 2.846, p < 0.05; organ, t 16 = 3.489, p < 0.01), and pectoral muscle (mass-specific, t 16 = 8.284, p < 0.001; whole tissue, t 16 = 6.759, p < 0.001) (Fig. 5b), compared to summer birds. Winter birds also had significantly higher COX activity in the heart (mass-specific, t 16 = 2.920, p < 0.05; organ, t 16 = 2.591, p < 0.05) and pectoral muscle (mass-specific, t 16 = 4.759, p < 0.001; whole tissue, t 16 = 3.305, p < 0.01), but not in the liver (mass-specific, t 16 = 1.265, p = 0.224; organ, t 16 = 1.407, p = 0.178) (Fig. 5c), relative to summer birds. There were significant, positive correlations between log BMR and log state 4 respiration in the heart, liver and pectoral muscle (Fig. 6a, c, e), and between log BMR and log COX activity in the heart and pectoral muscle (Fig. 6b, f). Although the correlation between log BMR and log COX activity in the liver was not statistically significant, it was nearly so (p = 0.07, Fig. 6d).

Correlations between state 4 respiration and basal metabolic rate (BMR) in the heart (a), liver (c) and pectoral muscle (e), and between cytochrome c oxidase activity and BMR in the heart (b), liver (d) and pectoral muscle (f), of Silky Starlings (Sturnus sericeus) captured in either winter or summer in Wenzhou City, China
Discussion
The results of this study indicated that silky starlings captured in winter have significantly higher M b, energy budget, organ mass, metabolic rate, and respiratory enzyme activity, than those captured in summer. This suggests that the thermogenic response of Silky Starlings to winter conditions is achieved by a range of adjustments from the organismal to cellular level.
Seasonal variation in BMR, M b, and energy budget
Many, but not all, small birds inhabiting temperate latitudes in the Northern hemisphere increase their BMR in winter relative to that in summer (McKechnie 2008; Smit and McKechnie 2010; Wu et al. 2015). For example, Cooper (2000) showed that the BMR of captive, winter-acclimatized Mountain Chickadees (Poecile gambeli) was higher than that measured in summer. Zheng et al. (2008b) also found that Eurasian Tree Sparrows captured in winter had higher BMR than those captured in summer. Similar results have also been observed in other free-living species (Rising and Hudson 1974; Southwick 1980; Liknes et al. 2002). Our data indicate that the BMR of the Silky Starling is 33% higher in winter than in summer. The functional significance of the winter increase in BMR is likely that it supports the maintenance costs of the thermogenic machinery. Seasonal variation in M b, especially in small birds, is considered to be an adaptive strategy important for survival (Chamane and Downs 2009; Cooper 2000; Zhao et al. 2015). Our results suggest that the Silky Starling is, on average, 18% heavier in winter than in summer. This increased winter M b could reflect both increased thermogenic capacity and the accumulation of energy reserves (Dawson et al. 1983; Cooper 2007). An increase in M b during winter is often due to increases in fat deposits, and or, the mass of metabolically active tissues (Williams and Tieleman 2000; Zheng et al. 2008b). In addition to the seasonal variation in M b, seasonal differences also occurred for some organ masses, so variation in these factors also likely contributes to the seasonal differences in BMR (Zheng et al. 2013a). An increased energy intake and budget can compensate for the increased energy expenditure associated with thermogenesis in cold conditions (Williams and Tieleman 2000). M b is an important indicator of energy balance (Doucette and Geiser 2008). A positive balance results from an adaptive increase in energy intake that periodically exceeds energy output as birds replenish their fat reserves and metabolic organ mass (Swanson 1991a; Cooper 2007; Liu et al. 2008; Zheng et al. 2008b, 2014a). Many studies have found evidence of significant seasonal variation in daily energy intake within species (Kelly 1998; Webster and Weathers 2000; Guillemette and Butler 2012; Hegemann et al. 2012). In this study, winter birds had 27% higher GEI, and 29% higher DEI, compared to summer birds, and the strong positive correlation between M b and BMR (see “Energy budget”), suggesting that enhancement of energy intake could be a general response to higher energy demands in small endotherms (Stokkan et al. 1986; Wu et al. 2014).
Seasonal variation in organ mass and tissue oxidative capacity
With respect to metabolic adjustments, the BMR of an animal is the sum of the metabolic rates of its organs and other metabolically active structures (Zheng et al. 2008a; Swanson 2010; Clapham 2012). The mechanisms underlying seasonal metabolic adjustments in birds may operate at multiple hierarchical levels. Several authors have suggested that much of the energy used in basal metabolism is consumed by visceral organs (Daan et al. 1990; Piersma et al. 1996; Burness et al. 1998), including the gizzard and small intestine (storage, digestion and absorption), the heart and lungs (oxygen transport), the liver (catabolism), and the kidneys (elimination of waste) (Kersten and Piersma 1987; Daan et al. 1990; Hammond et al. 2001). Although the liver, kidneys, heart and digestive tract represent less than 10% of total M b they contribute more than 60% of all energy expenditure while at rest (Clapham 2012). The results of this study indicate that winter birds had a heavier gizzard, digestive tract, heart, liver, and pectoral muscles, than summer birds, and masses of these organs also co-varied with BMR at seasons in a manner consistent with a regulatory role for seasonal RMR variation in this species (Table 1). The data for Silky Starlings in this study are thus consistent with previous data from small birds and, collectively, these studies suggest that seasonal changes in organ masses are important general contributors to RMR variation in small birds. In addition, the mass of the gizzard and small intestine, where food storage, digestion and absorption occur, were strongly correlated with GEI and DEI (Fig. 4b, c), suggesting that winter birds had increased their gizzard and small intestine mass in response to increased food intake.
State 4 respiration is clearly connected with BMR whereas COX activity is correlated with maximal capacity for thermogenesis (the increase in COX activity may simply be a function of mass-specific values and/or larger organ mass). We found that state 4 respiration in the heart (mass-specific and organ), liver (mass-specific and organ) and pectoral muscle (mass-specific and whole tissue) of winter birds was significantly higher than in summer birds. In addition, COX activity in the heart (mass-specific and organ) and muscle (mass-specific and whole tissue) of winter birds was significantly higher than in summer birds. Moreover, BMR was significantly correlated with state 4 respiration in the heart, liver and pectoral muscle, and with COX activity in the heart and pectoral muscle (Fig. 6). Although the correlation between BMR and COX activity in the liver was not statistically significant, it was nearly so (Fig. 6d). Similar results have been reported in other small passerines from the same region, such as the Chinese Bulbul (Zheng et al. 2010, 2014a) and Eurasian Tree Sparrow (Liu et al. 2008; Zheng et al. 2008a, 2014b), and other small birds in northern hemisphere like Little Bunting (Emberiza pusilla) (Zheng et al. 2013a), Black-capped Chickadee (Poecile atricapillus), House Sparrow (Passer domesticus) (Liknes and Swanson 2011), and in small mammals such as the Plateau Pika (Ochotona curzoniae) (Wang et al. 2006a), Root Vole (Microtus oeconomus) (Wang et al. 2006b) and Mongolian Gerbil (Meriones unguiculatus) (Zhang and Wang 2007). These results suggest that modulation of cellular thermogenesis at the tissue and organ levels are important aspects of seasonal acclimatization in the Silky Starling. These studies collectively present an increasingly comprehensive picture of seasonal adjustments in thermoregulation from the cellular to the organismal level.
Conclusions
Organisms are thought to acclimatize to seasonal variation in the environment through a range of physiological and biochemical adjustments (Zheng et al. 2008b). The results of this study suggest that Silky Starlings primarily acclimatize to winter conditions by increasing their M b, organ mass, BMR, energy intake, and tissue oxidative capacity. This suggests that a suite of adjustments, from the cellular to the organismal level, have the potential to contribute significantly to the thermogenic efforts of seasonal acclimatized birds. The observed seasonal changes in M b, basal thermogenesis (BMR) and tissue oxidative capacity in Silky Starlings may all be important to the ability of this temperate passerine to survive winter conditions (Liknes and Swanson 2011; Zheng et al. 2008b, 2014a).
References
Bao HH, Liang QJ, Zhu HL, Zhou XQ, Zheng WH, Liu JS. Metabolic rate and evaporative water loss in the silky starling (Sturnus sericeus). Zool Res. 2014;35:280–6.
Brand MD, Turner N, Ocloo A, Else PL, Hulbert AJ. Proton conductance and fatty acyl composition of liver mitochondria correlates with body mass in birds. Biochem J. 2003;376:741–8.
Burness GP, Ydenberg RC, Hochachka PW. Interindividual variability in body composition and resting oxygen consumption rates in breeding tree swallows Tachycineta bicolor. Physiol Zool. 1998;71:247–56.
Chamane SC, Downs CT. Seasonal effects on metabolism and thermoregulation abilities of the red-winged starling (Onychognathus morio). J Therm Biol. 2009;34:337–41.
Christians JK. Controlling for body mass effects: Is part-whole correlation important? Physiol Biochem Zool. 1999;72:250–3.
Clapham JC. Central control of thermogenesis. Neuropharmacology. 2012;63:111–23.
Cooper SJ. Seasonal energetics of mountain chickadees and juniper titmice. Condor. 2000;102:635–44.
Cooper SJ. Daily and seasonal variation in body mass and visible fat in mountain chickadees and juniper titmice. Wilson J Ornithol. 2007;119:720–4.
Daan S, Masman D, Groenewold A. Avian basal metabolic rates: their association with body composition and energy expenditure in nature. Am J Physiol. 1990;259:R333–40.
Dawson WR, Carey C. Seasonal acclimatization to temperature in cardueline finches I. Insulative and metabolic adjustments. J Comp Physiol B. 1976;112:317–33.
Dawson WR, Marsh RL. Winter fattening in the American Goldfinch and the possible role of temperature in its regulation. Physiol Zool. 1986;59:353–69.
Dawson WR, Marsh RL, Buttemer WA, Carey C. Seasonal and geographic variation of cold resistance in house finches. Physiol Zool. 1983;56:353–69.
Diamond JM. Evolution of biological safety factors: a cost/benefit analysis. In: Weibel ER, Taylor CR, Bolis L, editors. Principles of animal design. Cambridge: Cambridge University Press; 1998. p. 21–7.
Doucette LI, Geiser F. Seasonal variation in thermal energetics of the Australian owlet-nightjar (Aegotheles cristatus). Comp Biochem Physiol. 2008;151A:615–20.
Estabrook RW. Mitochondrial respiratory control and polarographic measurement of ADP/O ratio. In: Estabrook RW, Pullman ME, editors. Methods in enzymes, X. New York: Academic Press; 1967. p. 41–7.
Guillemette M, Butler PJ. Seasonal variation in energy expenditure is not related to activity level or water temperature in a large diving bird. J Exp Biol. 2012;215:3161–8.
Hammond KA, Szewczak J, Krόl E. Effects of altitude and temperature on organ phenotypic plasticity along an altitudinal gradient. J Exp Biol. 2001;204:1991–2000.
Hegemann A, Matson KD, Versteegh MA, Tieleman BI. Wild skylarks seasonally modulate energy budgets but maintain energetically costly inflammatory immune responses throughout the annual cycle. PLoS ONE. 2012;7:e36358.
Hill RW. Determination of oxygen consumption by use of the paramagnetic oxygen analyzer. J Appl Physiol. 1972;33:261–3.
Hu SN, Zhu YY, Lin L, Zheng WH, Liu JS. Temperature and photoperiod as environmental cues affect body mass and thermoregulation in Chinese bulbuls, Pycnonotus sinensis. J Exp Biol. 2017;220:844–55.
Kelly JP. Behavior and energy budgets of belted kingfishers in winter. J Field Ornithol. 1998;69:75–84.
Kersten M, Piersma T. High levels of energy expenditure in shorebirds; metabolic adaptations to an energetically expensive way of life. Ardea. 1987;75:175–87.
Klaassen M, Oltrogge M, Trost L. Basal metabolic rate, food intake, and body mass in cold- and warm-acclimated Garden Warblers. Comp Biochem Phys A. 2004;137A:639–47.
Li XS, Wang DH. Seasonal adjustments in body mass and thermogenesis in Mongolian gerbils (Meriones unguiculatus): the roles of short photoperiod and cold. J Comp Physiol B. 2005;175:593–600.
Li YG, Yang ZC, Wang DH. Physiological and biochemical basis of basal metabolic rates in Brandt’s voles (Lasiopodomys brandtii) and Mongolian gerbils (Meriones unguiculatus). Comp Biochem Phys A. 2010;157:204–11.
Liknes ET, Swanson DL. Seasonal variation in cold tolerance, basal metabolic rate, and maximal capacity for thermogenesis in white-breasted nuthatches Sitta carolinensis and downy woodpeckers Picoides pubescens, two unrelated arboreal temperate residents. J Avian Biol. 1996;27:279–88.
Liknes ET, Swanson DL. Phenotypic flexibility in passerine birds: seasonal variation of aerobic enzyme activities in skeletal muscle. J Therm Biol. 2011;36:430–6.
Liknes ET, Scott SM, Swanson DL. Seasonal acclimatization in the American goldfinch revisited: to what extent to metabolic rates vary seasonally? Condor. 2002;104:548–57.
Liu JS, Li M. Phenotypic flexibility of metabolic rate and organ masses among tree sparrows Passer montanus in seasonal acclimatization. Acta Zool Sin. 2006;52:469–77.
Liu JS, Li M, Shao SL. Seasonal changes in thermogenic properties of liver and muscle in tree sparrows Passer montanus. Acta Zool Sin. 2008;54:777–84.
MacKinnon J, Phillipps K. A field guide to the birds of China. London: Oxford University Press; 2000.
Maria S, Swanson DL, Cheviron ZA. Regulatory mechanisms of metabolic flexibility in the dark-eyed junco (Junco hyemalis). J Exp Biol. 2015;218:767–77.
McKechnie AE. Phenotypic flexibility in basal metabolic rate and the changing view of avian physiological diversity: a review. J Comp Physiol B. 2008;178:235–47.
McKechnie AE, Swanson DL. Sources and significance of variation in basal, summit and maximal metabolic rates in birds. Curr Zool. 2010;56:741–58.
McKechnie AE, Wolf BO. The allometry of avian basal metabolic rate: good predictions need good data. Physiol Biochem Zool. 2004;77:502–21.
McKechnie AE, Freckleton RP, Jetz W. Phenotypic plasticity in the scaling of avian basal metabolic rate. Proc R Soc Lond B. 2006;273:931–7.
McNab BK. The relationship among flow rate, chamber volume and calculated rate of metabolism in vertebrate respirometry. Comp Biochem Physiol A. 2006;145:287–94.
McNab BK. Ecological factors affect the level and scaling of avian BMR. Comp Biochem Physiol A. 2009;152:22–45.
Nilsson JF, Nilsson J-Å. Fluctuating selection on basal metabolic rate. Ecol Evol. 2016;6:1197–202.
Piersma T, Bruinzeel L, Drent R, Kersten M, Van der Meer J, Wiersma P. Variability in basal metabolic rate of a long-distance migrant shorebird (Red Knot, Calidris canutus) reflects shifts in organ sizes. Physiol Zool. 1996;69:191–217.
Piersma T, Drent J. Phenotypic flexibility and the evolution of organismal design. Trends Ecol Evol. 2003;18:228–33.
Petit M, Lewden A, Vézina F. How dose flexibility in body mass composition relate to seasonal changes in metabolic performance in a small passerine wintering at northern latitude? Physiol Biochem Zool. 2014;87:539–49.
Petit M, Clavijo-Baquet S, Vézina F. Increasing winter maximal metabolic rate improves intrawinter survival in small birds. Physiol Biochem Zool. 2017;90:166–77.
Rising JD, Hudson JW. Seasonal variation in the metabolism and thyroid activity of the black-capped chickadee (Parus atricapillus). Condor. 1974;76:198–203.
Schmidt-Nielsen K. Animal physiology: adaptation and environment. London: Cambridge University Press; 1997.
Smit B, McKechnie AE. Avian seasonal metabolic variation in a subtropical desert: basal metabolic rates are lower in winter than in summer. Funct Ecol. 2010;24:330–9.
Southwick EE. Seasonal thermoregulatory adjustments in white-crowned sparrows. Auk. 1980;97:76–85.
Starck JM, Rahmaan GHA. Phenotypic flexibility of structure and function of the digestive system of Japanese quail. J Exp Biol. 2003;206:1887–97.
Stokkan KA, Mortensen A, Blix AS. Food intake, feeding rhythm, and body mass regulation in Svalbard rock ptarmigan. Am J Physiol. 1986;251:R264–7.
Sundin U, Moore G, Nedergaard J, Cannon B. Thermogenin amount and activity in hamster brown fat mitochondria: effect of cold acclimation. Am J Physiol. 1987;252:R822–32.
Swanson DL. Seasonal variation in cold hardiness and peak rates of cold-induced thermogenesis in the dark-eyed junco (Junco hyemalis). Auk. 1990;107:561–6.
Swanson DL. Seasonal adjustments in metabolism and insulation in the dark-eyed junco. Condor. 1991a;93:538–45.
Swanson DL. Substrate metabolism under cold stress in seasonally acclimatized dark-eyed juncos. Physiol Zool. 1991b;64:1578–92.
Swanson DL. Are summit metabolism and thermogenic endurance correlated in winter—acclimatized passerine birds? J Comp Physiol B. 2001;171:475–81.
Swanson DL. Seasonal metabolic variation in birds: functional and mechanistic correlates. In: Thompson CF, editor. Current ornithology. Berlin: Springer; 2010. p. 75–129.
Swanson DL, Garland T Jr. The evolution of high summit metabolism and cold tolerance in birds and its impact on present-day distribution. Evolution. 2009;63:184–94.
Swanson DL, Liknes ET. A comparative analysis of thermogenic capacity and cold tolerance in small birds. J Exp Biol. 2006;209:466–74.
Swanson DL, McKechnie AE, Vézina F. How low can you go? An adaptive energetic framework for interpreting basal metabolic rate variation in endotherms. J Comp Physiol B. 2017;Suppl 1:1–18.
Swanson DL, Zhang Y, Liu JS, Merkord CL, King MO. Relative roles of temperature and photoperiod as drivers of metabolic flexibility in dark-eyed juncos. J Exp Biol. 2014;217:866–75.
Tieleman BI, Williams JB, Buschur ME, Brown CR. Phenotypic variation of larks along an aridity gradient: are desert birds more flexible? Ecology. 2003;84:1800–51.
Vézina F, Jalvingh K, Dekinga A, Piersma T. Acclimation to different thermal conditions in a northerly wintering shorebird is driven by body mass-related changes in organ size. J Exp Biol. 2006;209:3141–54.
Vézina F, Jalvingh KM, Dekinga A, Piersma T. Thermogenic side effects to migratory disposition in shorebirds. Am J Physiol. 2007;292:R1287–97.
Wang JM, Zhang YM, Wang DH. Seasonal thermogenesis and body mass regulation in plateau pikas (Ochotona curzoniae). Oecologia. 2006a;149:373–82.
Wang JM, Zhang YM, Wang DH. Seasonal regulations of energetics, serum concentrations of leptin, and uncoupling protein 1 content of brown adipose tissue in root voles (Microtus oeconomus) from the Qinghai-Tibetan plateau. J Comp Physiol B. 2006b;176:663–71.
Weathers WW. Climatic adaptation in avian standard metabolic rate. Oecologia. 1979;42:81–9.
Webster MD, Weathers WW. Seasonal changes in energy and water use by verdins, Auriparus flaviceps. J Exp Biol. 2000;203:3333–44.
Wiersma P, Muñoz-Garcia A, Walker A, Williams JB. Tropical birds have a slow pace of life. Proc Natl Acad Sci USA. 2007;104:9340–5.
Wiesinger H, Heldmaier G, Buchberger A. Effect of photoperiod and acclimation temperature on nonshivering thermogenesis and GDP-binding of brown fat mitochondria in the Djungarian hamster Phodopus s. sungorus. Pflugers Arch Eur J Physiol. 1989;413:667–72.
Wikelski M, Spinney L, Schelsky W, Scheuerlein A, Gwinner E. Slow pace of life in tropical sedentary birds: a common-garden experiment on four stonechat populations from different latitudes. Proc R Soc Lond B. 2003;270:2383–8.
Williams J, Tieleman BI. Flexibility in basal metabolic rate and evaporative water loss among hoopoe larks exposed to different environmental temperatures. J Exp Biol. 2000;203:3153–9.
Wu MS, Xiao YC, Yang F, Zhou LM, Zheng WH, Liu JS. Seasonal variation in body mass and energy budget in Chinese bulbuls (Pycnonotus sinensis). Avian Res. 2014;5:4.
Wu MX, Zhou LM, Zhao LD, Zhao ZJ, Zheng WH, Liu JS. Seasonal variation in body mass, body temperature and thermogenesis in the Hwamei, Garrulax canorus. Comp Biochem Physiol A. 2015;179:113–9.
Zhang YP, Liu JS, Hu XJ, Yang Y, Chen LD. Metabolism and thermoregulation in two species of passerines from south-eastern China in summer. Acta Zool Sin. 2006;52:641–7.
Zhang ZQ, Wang DH. Seasonal changes in thermogenesis and body mass in wild Mongolian gerbils (Meriones unguiculatus). Comp Biochem Physiol. 2007;148:346–53.
Zhao L, Zheng LY, Zhang W, Huang DF, Xu Y, Liu JS. Daily cyclic variation of metabolism and thermoregulation in the silky starling. Chin J Zool. 2013;48:269–77.
Zhao LD, Wang RM, Wu YN, Wu MS, Zheng WH, Liu JS. Daily variation in body mass and thermoregulation in male Hwamei (Garrulax canorus) at different seasons. Avian Res. 2015;6:4.
Zheng WH, Liu JS, Jang XH, Fang YY, Zhang GK. Seasonal variation on metabolism and thermoregulation in Chinese bulbul. J Therm Biol. 2008a;33:315–9.
Zheng WH, Li M, Liu JS, Shao SL. Seasonal acclimatization of metabolism in Eurasian tree sparrows (Passer montanus). Comp Biochem Physiol A. 2008b;151:519–25.
Zheng WH, Fang YY, Jang XH, Zhang GK, Liu JS. Comparison of thermogenic character of liver and muscle in Chinese bulbul Pycnonotus sinensis between summer and winter. Zool Res. 2010;31:319–27.
Zheng WH, Lin L, Liu JS, Xu XJ, Li M. Geographic variation in basal thermogenesis in little buntings: relationship to cellular thermogenesis and thyroid hormone concentrations. Comp Biochem Physiol A. 2013a;164:240–6.
Zheng WH, Lin L, Liu JS, Pan H, Cao MT, Hu YL. Physiological and biochemical thermoregulatory responses of Chinese bulbuls Pycnonotus sinensis to warm temperature: phenotypic flexibility in a small passerine. J Therm Biol. 2013b;38:483–90.
Zheng WH, Liu JS, Swanson DL. Seasonal phenotypic flexibility of body mass, organ masses, and tissue oxidative capacity and their relationship to RMR in Chinese bulbuls. Physiol Biochem Zool. 2014a;87:432–44.
Zheng WH, Li M, Liu JS, Shao SL, Xu XJ. Seasonal variation of metabolic thermogenesis in Eurasian tree sparrows Passer montanus over a latitudinal gradient. Physiol Biochem Zool. 2014b;87:704–18.
Zhou LM, Xia SS, Chen Q, Wang RM, Zheng WH, Liu JS. Phenotypic flexibility of thermogenesis in the Hwamei (Garrulax canorus): responses to cold acclimation. Am J Physiol. 2016;310:R330–6.
Authors’ contributions
JL and WZ provided the research idea and designed the experiments. ML, YS, HM and XJ conducted the experiments and collected the data. ML, YS and XJ finished the data analysis, compiled the results and wrote the first draft of the article. JL and WZ supervised the research and revised the draft. All authors read and approved the final manuscript.
Acknowledgements
We thank Dr. Ron Moorhouse revising the English of this MS. Thanks are also given to all the members of Animal Physiological Ecology Group, Institute of applied ecology of Wenzhou University, for their helpful suggestions. This study was financially supported by grants from the National Natural Science Foundation of China (No. 31470472), the National Undergraduate Innovation and Entrepreneurship Training Program and the Zhejiang Province “Xinmiao” Project.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, M., Sun, Y., Mao, H. et al. Seasonal phenotypic flexibility in body mass, basal thermogenesis, and tissue oxidative capacity in the male Silky Starling (Sturnus sericeus). Avian Res 8, 25 (2017). https://doi.org/10.1186/s40657-017-0083-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-017-0083-4