
Abstract
Lincomycin A is a clinically important antibiotic produced by Streptomyces lincolnensis that is used against gram-positive bacteria. To increase the yield of lincomycin A, a calcium gluconate feeding strategy was studied in a 15 L bioreactor. The results showed that the addition time of calcium gluconate was optimal during the late fermentation process to ensure a higher yield of lincomycin A. The optimum addition was continuous feeding at a speed of 0.0638 g/L/h from 111 to 158 h, which can increase the lincomycin A titer to 9160 mg/L, 41.3% higher than that without gluconate feeding. Enzyme activities of the central metabolic pathways, accumulation of intermediate metabolites, NADPH and NADH concentrations, and NADPH/NADH ratio were determined to investigate the mechanism of enhanced lincomycin A production by calcium gluconate addition. The activities of key enzymes of the pentose phosphate pathway (PPP) (glucose 6-phosphate dehydrogenase) and TCA cycle (isocitrate dehydrogenase) were enhanced by approximately twofold. A higher ratio of NADPH/NADH was observed in the fermentation process with the optimized feeding strategy providing sufficient reducing power. These data indicated that more flux flows through the PPP and the TCA cycle to provide more precursors, ATP and reducing power to support the synthesis of lincomycin A. The results showed a new strategy to improve the production of lincomycin A by manipulating the flux through the PPP and the TCA cycle.

Similar content being viewed by others

Introduction
Produced by Streptomyces lincolnensis, lincomycin is a clinically important lincosamide antibiotic with potent activity against gram-positive bacteria (Mason and Dietz 1964; Koberska et al. 2008). Its semisynthesized derivative clindamycin has a similar antimicrobial spectrum and greater antibiotic activity, even having antiprotozoal activity (Smieja 1998; Obonyo and Juma 2012). Lincomycin A is the main product during S. lincolnensis fermentation, with trace amounts of lincomycin B, which has only 25% of the antibacterial activity of lincomycin A (Kucers et al. 1997).
Lincomycin consists of a C8 backbone glycosyl moiety, methylthiolincosamide (MTL), and an amino moiety, 4-propyl-l-proline (PPL). The two components are synthesized separately and subsequently condensed through an amide bond (Lin et al. 2014; Sasaki et al. 2012; Floss and Beale 2014) (Fig. 1). The condensation of MTL and PPL is unusual in that two bacterial thiols play constructive role in the biosynthesis of lincomycin A: thiol ergothioneine (EGT) acts as a carrier to template the assembly of GDP-activated MTL and PPL moiety, and thiol mycothiol (MSH) is the sulfur donor for lincomycin maturation through two unusual S-glycosylation (Zhao et al. 2015; Zhang et al. 2018). MTL is generated via a transaldol reaction catalyzed by LmbR to form GDP-d-α-d-octose as the key intermediate in the MTL biosynthetic pathway (Sasaki et al. 2012; Lin et al. 2014). The substrate of LmbR is d-fructose 6-phosphate or sedoheptulose 7-phosphate as the C3 donor and d-ribose 5-phosphate as the C5 acceptor from the pentose phosphate pathway (PPP) (Sasaki et al. 2012). The PPL biosynthetic pathway contains eight successive reactions that convert tyrosine to PPL catalyzed by six enzymes: LmbB2, -B1, -W, -A, -X, and -Y (Peschke et al. 1995; Neusser et al. 1998; Jiraskova et al. 2016). It is known that PPL is derived from tyrosine, which is synthesized from shikimic acid, which is also an intermediate metabolite from the PPP synthesized by eight successive enzymatic reactions (Neusser et al. 1998). In the PPL biosynthesis pathway, LmbY is an F420-dependent reductase, which is proposed to catalyze two reduction steps to convert the intermediate (DH)2-PPL to PPL (Jiraskova et al. 2016). F420 is a cofactor of a riboflavin analog that participates in NADPH-mediated reductions in several Archaea and Mycobacteria (Kern et al. 1983; Bashiri et al. 2010).

Diagram for lincomycin biosynthesis
To improve lincomycin A production, many studies have been performed, including strain screening (Meng et al. 2013; Li 2013; Huang et al. 2017), optimization of medium composition (Xue et al. 2009; Lee et al. 2014) and process conditions (Li et al. 2009, 2013), and the genetic engineering of related functional genes (Pang et al. 2015) in S. lincolnensis. The above studies focused on dissolved oxygen concentration, pH control, consumption of nutrients, or rational breeding to screen strains with more productivity. Several metabolites in the PPP and redox reactions are involved in the biosynthesis of lincomycin. However, the effect of PPP on lincomycin production during fermentation is uncertain.
In this work, we developed an optimal strategy of adding calcium gluconate during the lincomycin fermentation process in a 15 L bioreactor. The effects of calcium gluconate on lincomycin production were studied by examining enzyme activity, intermediate metabolite accumulation, and NADPH and NADH concentration.
Media and methods
Strain and media
Streptomyces lincolnensis 18-8 was used throughout this work. The composition of seed medium was as follows (g/L): glucose 10, soybean meal 10, starch 20, corn steep liquor 30, (NH4)2SO4 1.5, and CaCO3 5, pH 7.2. For lincomycin A production, the following fermentation medium were used (g/L): glucose 28, starch 2, soybean meal 20, (NH4)2SO4 4.5, NaNO3 4.5, KH2PO4 0.5, NaCl 5, corn steep liquor 10, and CaCO3 8, pH 7.0. The composition of feeding medium was as follows (g/L): soybean meal 60, corn steep liquor 90, and CaCO3 1.25, pH 7.0.
Culture condition
Batch cultivation was conducted in a 15 L stirred bioreactor (Guoqiang Bioengineering Equipment Co., Ltd, Shanghai, China) with an initial working volume of 10 L. The inoculation volume was 10% (v/v) of the medium volume. The fermentation temperature and agitation speed were maintained at 30 °C and 550 rpm, respectively, and the aeration rate was 1.0 vvm. During the fermentation process, the pH was monitored on-line and controlled at 7.0 by 1 M H2SO4 and NH4OH solutions. The feed batch or continuous feeding strategy was adopted for feeding of calcium gluconate. The feeding condition were as follows: x g/L calcium gluconate was added at 0 h, then (4 − x) g/L was added totally at a specified time, or continuously in bioreactor during the specific fermentation time period. Samples were taken at 8 h intervals for off-line analysis.
Analytical methods
The culture samples taken at different times were centrifuged at 4000 rpm for 15 min to collect the mycelia. The precipitated mycelia were washed three times and dried to constant weight at 105 °C to measure the dry cell weight (DCW). The supernatant pH value, lincomycin titer, and concentration of reducing sugars and gluconic acid were determined after collecting the supernatant. All parameters were determined in triplicate.
The reducing sugar concentration was determined by direct titration using the Fehling method (Yao et al. 2017). The gluconic acid concentration was determined by the colorimetric method at 420 nm after oxidation by periodate (Jiang et al. 1997). The production of lincomycin A was assayed by high-performance liquid chromatography (HPLC) using a Diamonsil plus C18 column (250 × 4.6 mm, 5 μm; Dikma Corporation, Beijing, China). The column temperature was maintained at 30 °C, and UV detection was set at 210 nm. A mobile phase containing 50 mM ammonium acetate:methanol (3:2, v/v) was used at a flow rate of 0.4 mL/min (Li et al. 2007).
Preparation of cell-free extracts from mycelia was as follows. Briefly, mycelia were washed twice with distilled water and then cracked in an ice-water bath with a sonicator (Scientz Inc. Ningbo, China) for 5 min at 120 W. Cell fragments were removed by centrifugation at 12,000 rpm for 30 min. Protein concentrations of the cell-free extract were measured by the method of Lowry et al. (1951) with bovine serum albumin as the standard.
The activity of glucokinase (GK), glucose 6-phosphate (G6P) dehydrogenase (G6PD) and isocitrate dehydrogenase (ICD) in cell-free extracts was measured spectrophotometrically by monitoring the reduction of NADP+ directly or in an enzyme coupling reaction (Maitra 1970; Olano et al. 1995; Lee 1982). The assay mixture of glucose kinase contained 50 mM Tris–HCl, pH 7.0, 20 mM glucose, 25 mM MgCl2, 0.5 mM NADP+, 1 mM ATP, and 0.7 U/mL G6PD, and enzyme extract. The assay mixture of G6PD contained 50 mM Tris–HCl, pH 8.0, 1 mM G6P, 10 mM MgCl2, 0.5 mM NADP+, and enzyme extract. The assay mixture of isocitrate dehydrogenase contained 50 mM Tris–HCl, pH 7.6, 5 mM MgCl2, 3 mM isocitrate, 0.5 mM NADP+, and enzyme extract. All assays were started by the addition of up to 50 µL of cell extract. Units of enzyme activity were expressed as nmol NADP+ reduced per min per mg total cell protein at 30 °C.
Metabolite analysis
To prepare samples for metabolite analyses, a fast quenching method was used (Zhao et al. 2014). Briefly, samples (20 mL) were quickly transferred into 80 mL of quenching solution (glycerol: 0.23 M NaCl solution (3:2 v/v), then centrifuged at 14,000 rpm for 20 min at − 10 °C to collect S. lincolnensis cells. The intracellular metabolites were extracted using freeze–thaw methanol extraction and then centrifuged at 14,000 rpm for 10 min at − 10 °C. The supernatant was collected and vacuum dried using a SpeedVac SPD 131DDA solvent evaporation system (Thermo Scientific, Waltham, MA). Samples were dissolved in 50 μL of methoxamine hydrochloride (20 mg/mL in pyridine) and incubated at 70 °C for 50 min. Then, 80 μL of N-methyl-N-(trimethylsilyl) trifluoroacetamide (MSTFA) was added, and the sample was incubated at 70 °C for 50 min for trimethylsilylation (Ding et al. 2009). GC–MS analysis of intermediates was performed as described (Zhao et al. 2014).
Determination of NADPH or NADH
The NADPH and NADH concentrations were determined with an NADPH or NADH detection kit (Keming, Suzhou, China) using an enzyme cycling assay for determination. The fermentation samples were centrifuged immediately at 12,000 rpm for 1 min. After removal of the supernatants, 0.2 M NaOH was added to the pellets. The samples were placed in a 50 °C water bath for 10 min and then on ice to cool them to 0 °C. The extracts were neutralized by adding 0.1 M HCl dropwise while vortexing. The cellular debris were removed by centrifuging at 12,000 rpm for 5 min. Supernatants were collected for the measurement of intracellular NADH and NADPH by a sensitive NAD(P)-dependent dehydrogenase reactions assay with 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) as a terminal electron acceptor and phenazine ethosulfate (PES) as an electron carrier (Hou et al. 2010). The NAD(P)-dependent dehydrogenase was ADH for NADH and G6PDH for NADPH, respectively. The absorbance was measured at 570 nm.
Results and discussion
Influence of gluconate addition on lincomycin production and the optimization of gluconate addition
Previous experiments have shown that the addition of gluconate at the beginning and in late processes during fermentation enhances the production of secondary metabolites (Zhuang et al. 2018). We hypothesize that feeding gluconate will also improve the yield of lincomycin A. To study the optimal conditions of gluconate addition in a 15 L bioreactor, a strategy of two additions, one at the beginning and one at 127 or 147 h of fermentation, was investigated (Table 1, modes 1 and 2). The strategy was to select the appropriate feeding time during the late fermentation process to maintain the vitality of mycelia. With regard to lincomycin A production and cell growth, the additions at 127 and 147 h led to 18% and 20% higher levels of lincomycin A compared to those of the control, and the DCW increased almost 10% compared to that of the control (Table 1).
Considering the concentration of gluconate added at the beginning of the fermentation process, we compared the effects of decreasing the gluconate concentration at the beginning of fermentation (Table 1, modes 2–5), with the total amount constant at 4 g/L. The results showed that the production of lincomycin A increased with decreasing concentration of gluconate at the beginning of fermentation, while at the same time, the cell growth decreased. This result indicated that decreasing the concentration of gluconate could slow the growth of S. lincolnensis while promoting the production of lincomycin A, especially the starting titer of lincomycin. The optimal added concentration of gluconate was 0.6 g/L at the beginning with the other 3.4 g/L added at 147 h of fermentation. The titer of lincomycin A reached 8012 mg/L, 23% higher than that of the control.
Next, the optimal speed of continuous gluconate addition was studied (Table 1, Fig. 2). After adding 0.6 g/L gluconate at the beginning of fermentation, the other 3.4 g/L was added continuously from 110 h of fermentation at various speeds with a total amount of gluconate of 3.4 g/L. The final titer of lincomycin A was 9160 ± 316 mg/L, which was 41.3% higher than that of the control (p < 0.01). The calculated specific synthesis rate of lincomycin (qp) was enhanced to 80 mg/L/h, 60% higher than that of the control (50 mg/L/h).

Time course of LinA production with various feeding modes. Filled square: control, filled circle: mode 6, filled triangle: mode 7, filled star: mode 8, filled diamond : mode 9. Each point represents the mean (n = 3) ± standard deviation
Influence of gluconate addition on carbon and nitrogen source consumption
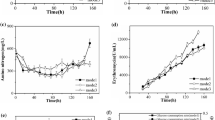
Glucose is used as the main carbon source during lincomycin fermentation, and there is evidence that calcium gluconate addition influences the consumption of glucose through the regulation of key enzymes in the glucose metabolic pathway during inosine and l-histidine fermentation (Guo et al. 2008; Zhao and Zhang 2008). The consumption of glucose and gluconate was explored in this work (Fig. 3a, b). The consumption rate of glucose in mode 8 was 0.08 g/L/h, 33% faster than that of the control (0.06 g/L/h) (p < 0.05) after gluconate addition. The intracellular accumulation of glucose was also determined and showed the same trend of faster consumption of glucose. The intracellular glucose concentration of mode 8 decreased to 3.45 μmol/g DCW, whereas the control remained at 13.29 μmol/g DCW (Fig. 3c; p < 0.01). The addition of gluconate can maintain the concentration of gluconate at 1.5–2.0 g/L, indicating the accumulation of gluconic acid (Fig. 3b). The addition of gluconate can also promote nitrogen utilization; the concentration of ammonium ion was approximately 100 mg/L, while that of the control was approximately 300 mg/L (Fig. 3d).

Time course changes of glucose (Glc) consumption rate (a), calcium gluconate concentration (b), intracellular Glc concentration (c), and extracellular ammonium ion concentration (d). Black: mode 8. Red: control. Each point represents the mean (n = 3) ± standard deviation
Influence of gluconate addition on enzyme activity
The central metabolic enzyme activity was measured during the lincomycin fermentation process (Fig. 4). The enzyme activity of GK (key enzyme of the EMP pathway) was similar to that of the control, which indicated that gluconate addition had little effect on the EMP pathway (Fig. 4a). G6PD is a key enzyme in the PPP. In the presence of gluconate, the enzyme activity of G6PD was significantly enhanced after 110 h, reached a maximum (8.35 U/mg protein) at 140 h that was almost 6 times the level of the control (1.28 U/mg protein) and was maintained at a stable high level in the following cultivation, indicating that the flux of the PPP increased after gluconate addition (Fig. 4b). The PPP can provide intermediate metabolites for lincomycin A synthesis, including ribose 5-phosphate (R5P), fructose 6-phosphate (F6P) and sedoheptulose 7-phosphate (S7P). ICD is a key enzyme in the TCA cycle. The results showed that the enzyme activity of ICD was significantly enhanced after 110 h, and the maximum activity reached 5.1 U/mg protein at 160 h, 4 times the control value (1.4 U/mg protein) (p < 0.01) (Fig. 4c). The enhanced TCA cycle enzyme activity suggested increased flux of the TCA cycle, and more ATP was synthesized to maintain the production of lincomycin A.

Time course of enzyme activities of glucokinase (GK) (EMP) (a), glucose 6-phosphate dehydrogenase (G6PD) (PPP) (b) and isocitrate dehydrogenase (ICD) (TCA cycle) (c). Black: mode 8. Red: control. Each point represents the mean (n = 3) ± standard deviation. **Represents p < 0.01
Influence of gluconate addition on the intracellular accumulation of metabolites
Intracellular accumulation of metabolites was measured during the lincomycin fermentation process (Table 2). Because the direct metabolites of OCT-8P and PPL of lincomycin biosynthesis were either found at very low concentrations or hard to be detected (Kameník et al. 2009), so we detected the common metabolites in the glucose metabolism to show the relationship of primary metabolism and secondary metabolism. Glucose, the substrate for microorganism growth and metabolism, succinate, fumarate, malate, and α-KG, the metabolites in TCA cycle, as well as G6P, R5P, and S7P for the metabolites in EMP and PPP, were assayed. Although these metabolites are not directly related to lincomycin biosynthesis, they are indirectly related to the production of OCT-8P, tyrosine and PPL (Sasaki et al. 2012). In the presence of gluconate, the intracellular accumulation of α-ketoglutaric acid (α-KG) decreased significantly to 4.23 μmol/g DCW at 144 h, only 6% of the control (67.17 μmol/g DCW) (p < 0.01). Moreover, the intracellular concentration of malic acid also reached a minimum (0.86 μmol/g DCW), accounting for only 11.9% of the control (7.17 μmol/g DCW) (p < 0.05). The intracellular concentrations of fumaric acid and succinic acid were also reduced, accounting for 21% and 2% of the control, respectively. Combined with the increased ICD enzyme activity, it was revealed that the flux of the TCA cycle was enhanced to provide more flux toward α-KG, while the decreased intracellular concentration of the intermediate metabolite α-KG indicated that more α-KG was taken away to synthesize a precursor of lincomycin, such as proline or PPL (Fig. 1). On the other hand, isocitrate was rerouted to malate via glyoxylate, and the accumulation of malate was decreased, whereas the corresponding accumulation of fumarate and succinate were decreased. It is speculated that more flux went from malate to oxaloacetate, leading to S-adenosylmethionine (SAM) (Fig. 1), which is essential for the methylation of lincomycin precursors. It is worth noting that the accumulation of G6P, R5P, and S7P were not detected, indicating the limiting of intracellular metabolites of the PPP.
Influence of gluconate addition on the intracellular accumulation of reducing power (NADH/NADPH)
Intracellular accumulation of NADH or NADPH was measured during the lincomycin fermentation process (Fig. 5). NADH/NAPH represents an important metabolic regulation of glucose metabolism, and NADPH is also an important reducing agent in the synthesis of assimilates such as lincomycin (Hou et al. 2010). In the presence of gluconate, the intracellular accumulation of NADPH was enhanced significantly to 11.92 nmol/g DCW at 144 h, almost 30% higher compared to that of the control (5.22 nmol/g DCW) (p < 0.01) (Fig. 5a). The intracellular concentration of NADH was almost the same as that of the control at 144 h (Fig. 5b), but the value of the NADPH to NADH ratio was significantly different (Fig. 5c). The value reached 1.57, which was 1.18 times higher than that of the control (0.72) (p < 0.05) (Fig. 5c), indicating that the addition of gluconate provides more of the reducing power of NADPH for the cell to support lincomycin synthesis. Because NADPH is mainly derived from the PPP, the enhanced PPP enzyme activity of G6PD and almost no accumulation of PPP metabolites might mean that the production of metabolic precursors is insufficient to support the synthesis of lincomycin, suggesting that PPP precursors may be one of the limiting factors for lincomycin biosynthesis.

Time course of intracellular NADPH (a) or NADH (b) concentrations and their ratio (c). Black: mode 8. Red: control. Each point represents the mean (n = 3) ± standard deviation. **Represents p < 0.01
Conclusion
In the present work, a strategy of continuous supplementation of gluconate leading to increased lincomycin A production accompanied by various responses of enzyme activity, intermediate metabolite and NADPH/NADH levels was achieved. We presented an effective example of enhanced antibiotic production during Streptomyces fermentation via manipulation of a biosynthetic precursor. Importantly, the developed strategy for the improved production of this highly valuable antibiotic may be helpful for large-scale fermentation.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- MTL:
-
methylthiolincosamide
- PPL:
-
N-methylated 4-propyl-l-proline
- α-KG:
-
α-ketoglutarate
- SAM:
-
S-adenosyl-l-methionine
- G6P:
-
glucose 6-phosphate
- R5P:
-
ribose 5-phosphate
- S7P:
-
sedoheptulose 7-phosphate
- GK:
-
glucokinase
- G6PD:
-
glucose 6-phosphate dehydrogenase
- ICD:
-
isocitrate dehydrogenase
References
Bashiri G et al (2010) Metabolic engineering of cofactor F420 production in Mycobacterium smegmatis. PLoS ONE 12:e15803
Ding MZ et al (2009) Comparative metabolomics analysis on industrial continuous and batch ethanol fermentation processes by GC-TOF-MS. Metabolomics 5:229–238
Floss HG, Beale JM (2014) Biosynthetic studies on antibiotics. Angew Chem Int Ed 43:1–20
Guo YX et al (2008) Effect of calcium gluconate on activities of key enzymes in glucose metabolic pathway during inosine fermentation. Chin J Pharm 39:254–257
Hou J et al (2010) Metabolic impact of increased NADH availability in Saccharomyces cerevisive. Appl Environ Microbiol 76:851–859
Huang WF et al (2017) Breeding of high yield Streptomyces linconinensis producing strain. Chin J Antibiot 42:647–651
Jiang J et al (1997) Study on colorimetric determination of gluconic acid. J Qiqhar Light Ind Inst 13:18–21
Jiraskova P et al (2016) New concept of the biosynthesis of 4-alkyl-l-proline procursors of lincomycin, hormaomycin, and pyrrolobenzodiazepines: could a γ-glutamyltransferase cleave the C–C bond? Front Microbiol 7:1–14
Kamenik et al (2009) HPLC-fluorescence detection method for determination of key intermediates of the lincomycin biosynthesis in fermentation broth. Anal Bioanal Chem 393:1779–1787
Kern R et al (1983) Isolation and structural identification of a chromophoric coenzyme F420 fragment from culture fluid of Methanobacterium thermoautotrophicum. Arch Microbiol 136:191–193
Koberska M et al (2008) Sequence analysis and heterologous expression of the lincomycin biosynthetic cluster of the type strain Streptomyces lincolnensis ATCC 25466. Folia Microbiol 53:395–401
Kucers A et al (1997) The use of antibiotics: a clinical review of antibacterial, antifungal and antiviral drugs. Oxford University Press, Oxford
Lee CY (1982) Glucose 6-phosphate dehydrogenase from mouse. Methods Enzymol 89:252–257
Lee Y et al (2014) Optimization of cultivation medium and fermentation parameters for lincomycin production by Streptomyces lincolnensis. Biotechnol Bioprocess Eng 19:1014–1021
Li JR (2013) Breeding of high lincomycin-producing strain. Chin J Antibiot 38:815–819
Li XB et al (2007) Improved industrial fermentation of lincomycin by phosphorus feeding. Proc Biochem 44:662–668
Li X et al (2009) Study on the metabolic characteristics of Streptomyces lincolnensis and regulation of pH during lincomycin fermentation. Chin J Antibiot 34:215–218
Li X et al (2013) Effects of flow field on the metabolic characteristics of Streptomyces lincolnensis in the industrial fermentation of lincomycin. J Biosci Bioeng 115:27–31
Lin CI et al (2014) In vitro characterization of LmbK and LmbO: identification of GDP-d-erythro-α-d-gluco-octose as a key intermediate in lincomycin A biosynthesis. J Am Chem Soc 136:1249–1265
Lowry OH et al (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193:265–275
Maitra PK (1970) A glucokinase from Saccharomyces cerevisiae. J Biol Chem 245:2423–2431
Mason DJ, Dietz A (1964) Lincomycin, a new antibiotic. Discovery and biological properties. Antimicrob Agents Chemother 115:554–559
Meng N et al (2013) Rational screening high-producing lincomycin strain. Bull Ferment Sci Technol 42:9–12
Neusser D et al (1998) The genes lmbB1 and lmbB2 of Streptomyces lincolnensis encode enzymes involved in the conversion of l-tyrosine to propylproline during the biosynthesis of the antibiotic lincomycin A. Arch Microbiol 169:322–332
Obonyo CO, Juma EA (2012) Clindamycin plus quinine for treating uncomplicated falciparum malaria: a systematic review and meta-analysis. Malar J 11:2–11
Olano J et al (1995) Kinetics and thermostability of NADP+-isocitrate dehydrogenase from Cephalosporium acremonium. Appl Environ Microbiol 61:2326–2334
Pang AP et al (2015) Co-overexpression of lmbW and metK led to increased lincomycin A production and decreased byproduct lincomycin B content in an industrial strain of Streptomyces lincolnensis. J Appl Microbiol 119:1064–1074
Peschke U et al (1995) Molecular characterization of the lincomycin-production gene cluster of Streptomyces linclonensis 78-11. Mol Microbiol 16:1137–1156
Sasaki E et al (2012) Construction of the octose 8-phosphate intermediate in lincomycin A biosynthesis: characterization of the reactions catalyzed by LmbR and LmbN. J Am Chem Soc 134:17432–17435
Smieja M (1998) Current indications for the use of clindamycin: a critical review. Can J Infect Dis 9:22–28
Xue ZL et al (2009) Optimization of culture medium for lincomycin production by response surface methodology. Chin J Antibiot 34:277–280
Yao KY et al (2017) Influence of dextrins on the production of spiramycin and impurity components by Streptomyces ambofaciens. Folia Microbiol 44:1–8
Zhang DZ et al (2018) Biosynthesis of lincosamide antibiotics: reactions associated with degradation and detoxification pathways play a constructive role. Acc Chem Res 51:1496–1506
Zhao JY, Zhang WG (2008) Effect of calcium gluconate on l-histidine fermentation and the fermentation conditions optimization. Biotechnol Bull 18:77–79
Zhao C et al (2014) Evaluation of metabolome sample preparation methods regarding leakage reduction for the oleaginous yeast Yarrowia lipolytica. Biochem Eng J 82:63–70
Zhao QF et al (2015) Metabolic coupling of two small-molecule thiols programs the biosynthesis of lincomycin A. Nature 518:115–119
Zhuang ZH et al (2018) Optimization of lincomycin production and component by response surface methodology. Chin J Antibiot 43:1049–1054
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China Youth Fund (No. 21506056); and the Open Funding Project of the State Key Laboratory of Bioreactor Engineering.
Author information
Authors and Affiliations
Contributions
SG and ZZ designed the experiments. ZZ performed bioreactor experiments with participation by CY. ZZ and LZ determined the concentration of intermediate metabolites, NADPH and NADH. ZZ prepared the draft with assistance from SG and QW. DZ and QM gave suggestions about the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Zhuang, Z., Zhang, L., Yang, C. et al. Enhanced lincomycin A production by calcium gluconate feeding in fermentation of Streptomyces lincolnensis. Bioresour. Bioprocess. 6, 31 (2019). https://doi.org/10.1186/s40643-019-0266-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-019-0266-4