
Abstract
Background
Cryptosporidium parvum is an important zoonotic parasitic disease worldwide, but the molecular mechanisms of the host–parasite interaction are not fully understood. Noncoding microRNAs (miRNAs) are considered key regulators of parasitic diseases. Therefore, we used microarray, qPCR, and bioinformatic analyses to investigate the intestinal epithelial miRNA expression profile after Cryptosporidium parvum infection.
Results
Twenty miRNAs were differentially expressed after infection (four upregulated and 16 downregulated). Further analysis of the differentially expressed miRNAs revealed that many important cellular responses were triggered by Cryptosporidium parvum infection, including cell apoptosis and the inflammatory and immune responses.
Conclusions
This study demonstrates for the first time that the miRNA expression profile of human intestinal epithelium cells is altered by C. parvum infection. This dysregulation of miRNA expression may contribute to the regulation of host biological processes in response to C. parvum infection, including cell apoptosis and the immune responses. These results provide new insight into the regulatory mechanisms of host miRNAs during cryptosporidiosis, which may offer potential targets for future C. parvum control strategies.
Similar content being viewed by others

Background
Cryptosporidium is a genus of protozoan parasites that infect the gastrointestinal epithelium and other mucosal surfaces of their hosts, which include humans and domestic and wild animals worldwide [1,2,3]. Cryptosporidium is a major cause of moderate to severe diarrhea in children younger than 2 years, particularly infants, and is second only to rotavirus in this regard [4]. Morbidity and mortality are associated with cryptosporidial infections in patients with acquired immunodeficiency syndrome (AIDS) and young children [1, 5]. Cryptosporidium parvum is one of the species most commonly involved in human cryptosporidial infections [1]. Despite intensive efforts over the past 30 years, the precise molecular pathogenic mechanisms of cryptosporidial infection are not fully understood.
MicroRNAs (miRNAs) are endogenous noncoding small RNAs (22 nucleotides) that regulate gene expression at the posttranscriptional level and play important roles in the regulation of diverse pathological processes, including cell proliferation, differentiation, metabolism, the immune response, and apoptosis [6,7,8]. The miRNAs in mammalian cells have been shown to play crucial roles in the cellular responses to infection by diverse pathogens, including viruses, parasites, and bacteria [9,10,11]. Increasing evidence suggests that host miRNAs help the host to clear parasites by regulating the toll-like receptor 4 (TLR4) signaling pathways and the release of antimicrobial peptides [12, 13]. For example, C. parvum infection reduces the expression of the let-7 family miRNAs in biliary epithelial cells, which increases SNAP23 expression, coordinating the release of exosomes carrying antimicrobial-peptide in response to C. parvum infection [13]. The upregulation of miR-27b expression enhances the anti-C. parvum responses of biliary epithelial cells [14]. Some evidence suggests that host miRNAs are also used by C. parvum to enhance its own survival. For example, suppressor of cytokine signaling (SOCS) proteins are negative regulators of cytokine signaling [15]. Cryptosporidium parvum infection downregulates the expression of miR-98 and let-7 to induce SOCS protein expression in biliary epithelial cells [16]. The upregulation of miR-21 after C. parvum infection inhibits the activation of TLR4/NF-κB signaling in biliary epithelial cells by targeting PDCD4, a proinflammatory protein that promotes the activation of NF-κB [17]. Therefore, studying the changes in the host miRNA expression profile after C. parvum infection will extend our understanding of the interaction between C. parvum and its host.
There has been no report of the changes in the miRNA expression profile in the intestinal epithelium after C. parvum infection. Therefore, the objective of this study was to analyze the intestinal epithelial miRNA expression profile after C. parvum infection using microarray and bioinformatics analyses in an in vitro model using human ileocecal HCT-8 adenocarcinoma cells.
Methods
Cell and parasite
Human ileocecal adenocarcinoma (HCT-8) cells (American Type Culture Collection, Manassas, VA) were cultured and maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum, 4 mmol/L l-glutamine, 100 U/mL penicillin, and 100 U/mL streptomycin at 37 °C in a humidified 5% CO2 incubator.
Neonatal calves were purchased from Ruiya Animal Husbandry Co., Ltd. (Zhengzhou, China). Oocysts of C. parvum (IId subtype) were maintained in infected neonatal calves, purified from their feces, and stored at 4 °C in 2.5% potassium dichromate until use. Before use, the oocysts were washed with phosphate-buffered saline (PBS), treated with 10% hypochlorite for 10 min at 4 °C, and washed again [18]. The infected calf was killed by anesthesia (113 mg sodium pentobarbital/kg, Intravenous injection) and carcass was treated harmlessly by a specialized agency of our college.
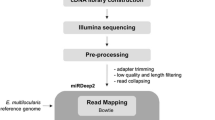
The infection assays have been reported previously [18]. Briefly, cell monolayers in six-well cell culture dishes were inoculated with 100 μL of purified oocysts (1 × 107/well) in DMEM, that the oocyst:host cell ratio is 5:1. Heat-inactivated (100 °C, 5 min) oocysts were used as the controls. After 2.5 h, oocysts and sporozoites remaining in the supernatant were removed and replaced with DMEM containing 2% fetal bovine serum, 4 mmol/L l-glutamine, 100 U/mL penicillin, 100 g/mL streptomycin, 0.3 g/L bicarbonate, and 0.02 g/L taurocholate. The plates were incubated at 37 °C in a humidified 5% CO2 incubator.
Total RNA isolation and quality control
At 4 and 12 h postinfection, the cells were washed three times with PBS, and 1 mL of TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) was added to each well. The total RNA from three uninfected separate monolayers and three infected separate monolayers were isolated according to the manufacturer’s protocol.
The quality of the RNA was measured with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA). RNA purity was assessed from the absorbance (A) of the solutions at various wavelengths: 2.0 > A260/A280 > 1.8 and A260/A230 > 2.0 were considered to indicate high purity. RNA integrity was determined from the 28S:18S rRNA ratio determined with an electrophoretic analysis.
Microarray analysis
miRNA microarrays based on miRBase v20.0 (http://www.mirbase.org/) were used to study the expression profiles of 2555 mature miRNAs. Microarray chips are synthesized by LC Sciences and have patent rights. The chips are made by a flexible, in situ synthesis method using conventional chemistry. The technology encompasses a novel high-throughput biopolymer synthesis chemistry carried out using a new class of microfluidic reaction devices and an advanced digital light synthesizer apparatus and is developed for carrying out picoliter scale chemical and/or biochemical reactions. Briefly, Hybridization was performed using the μParaflo® microfluidic array technology (LC Sciences, Hangzhou, China), according to the manual. The assay started from 4 to 8 μg total RNA sample were 3′-extended with a poly(A) tail using poly(A) polymerase. An oligonucleotide tag that binding to Cy3 dyes in later chip hybridization was then ligated to the poly(A) tail. Hybridization was performed overnight on a μParaflo microfluidic chip using a micro-circulation pump (Atactic Technologies). On the microfluidic chip, each detection probe consisted of a chemically modified nucleotide coding segment complementary to target microRNA (from miRBase, http://www.mirbase.org/). The detection probes were made by in situ synthesis using PGR (photogenerated reagent) chemistry. The hybridization melting temperatures were balanced by chemical modifications of the detection probes. Hybridization used 100 μL 6xSSPE buffer (0.90 M NaCl, 60 mM Na2HPO4, 6 mM EDTA, pH 6.8) containing 25% formamide at 34 °C. After RNA hybridization, oligonucleotide tag-conjugating Cy3 dye were circulated through the microfluidic chip for dye staining. Fluorescence images were collected using a laser scanner (GenePix 4000B, Molecular Device) and digitized using Array-Pro image analysis software (Media Cybernetics). Data were analyzed by first subtracting the background and then normalizing the signals using a LOWESS filter (Locally-weighted Regression).
Each miRNA has a fold change in the signals detected in infected cells relative to the signals detected in uninfected cells and the P value that calculated with Student’s t test. Differentially expressed miRNAs was selected based on a fold change of > 2 or < − 2 and a P value of ≤0.05.
Validation of microarray data with quantitative PCR (qPCR) analysis
miRNAs were prepared with an All-in-One™ miRNA qRT–PCR Detection Kit (GeneCopoeia, Rockville, MD, USA), according to the manufacturer’s instructions. Briefly, the extracted RNA was reverse-transcribed in the presence of a poly(A) polymerase and an oligo-dT adaptor. Quantitative PCR was then performed with SYBR Green detection using a forward primer specific for the mature miRNA sequence and a universal adaptor reverse primer. The forward primers used for the qPCRs are shown in Table 1. Each RNA was run in triplicate wells with three biological replicates, including three uninfected separate monolayers and three infected separate monolayers. The cycling protocol was: 95 °C for 10 min, followed by 40 cycles of 95 °C for 10 s and 60 °C for 20 s, with a final extension at 72 °C for 20 s. When all cycles were finished, a melting-curve analysis was performed. The baseline and cycle threshold (Ct) values were automatically determined for all plates with the Roche LightCycler® 96 Real-Time PCR System. The miRNA expression levels in each sample were normalized to the expression of the housekeeping gene U6 [19,20,21]. The real-time PCR results were analyzed and expressed as the relative Ct value using the 2−ΔΔCt method [22]. The mean and standard deviation (M ± SD) was calculated from three independent biological trials. Differences between infected and control group were measured using the t-test by SPSS 17.0 with P < 0.05 indicating significance.
Predicted target genes of differentially expressed miRNAs, gene ontology (GO) and Kyoto encyclopedia of genes and genomes
(KEGG) pathway analyses
Despite an abundance of online software for predicting human microRNA targets, many human miRNAs are only included in TargetScanHuman7.1 and miRDB. Therefore, the target genes of the miRNAs were predicted with the two online programs TargetScanHuman7.1 (http://www.targetscan.or g/vert_71/) [23] and miRDB (http://www.mirdb.org/cgi-bin/ search.cgi) [24]. The target genes of the differentially expressed miRNAs were analyzed in terms of their GO categories and KEGG pathways using the Database for Annotation, Visualization and Integrated Discovery (DAVID) (https://david.ncifcrf.gov/), a gene annotation tool [25].
miRNA–target gene network analysis
The miRNA- predicted target genes regulatory network was constructed with the miRNA-target gene pairs using the Cytoscape software(www.cytoscape.org) [26,27,28], based on the interactions of the miRNAs and genes [29], Briefly, a text file containing miRNAs and predicted target genes was imported in Cytoscape software and optimized the visual features of nodes and edges, such as shape and color. With an miRNA–target gene network, we can identify the crucial miRNA which have the highest connectivity to other miRNAs. The higher the connectivity of a miRNA was, the more important it was in the network. Also, we can find the key genes which were the common target genes of the differentially expressed miRNAs.
Results
Differentially expressed miRNAs
A total of 2555 mature human miRNA sequences were detected in C. parvum-infected and uninfected HCT-8 cells using miRNA microarrays (Sanger miRBase v20.0). A dendrogram produced with a hierarchical clustering analysis of the miRNAs differentially expressed in C. parvum-infected and -uninfected cells is shown in Fig. 1. In total, 20 miRNAs (four upregulated and 16 downregulated) were significantly differentially expressed in the infected HCT-8 cells compared to uninfected cells. These differentially expressed miRNAs (with fold changes ≥2 and P ≤ 0.05) are shown in Table 2 with P values. From this table, it is clear that downregulated miRNAs were more frequent than upregulated miRNAs (P < 0.05). Among these miRNAs, hsa-miR-942-5p was most strongly upregulated and hsa-miR-4689 was most strongly downregulated.

Expression profiling of miRNAs in HCT-8 cells after C. parvum infection. Expression profiles of miRNAs in HCT-8 cells at 4 h after C. parvum infection. The horizontal axis indicates samples of non-infected cells (n = 3; Control 1, 2, and 3) and cells after exposure to live C. parvum for 12 h (n = 3, C. parvum 1, 2, and 3) (a). Expression profiles of miRNAs in HCT-8 cells at 12 h after C. parvum infection. The horizontal axis indicates samples of non-infected cells (n = 3; Control 1, 2, and 3) and cells after exposure to live C. parvum for 12 h (n = 3, C. parvum 1, 2, and 3) (b)
Validation of miRNA microarray data with qPCR
Six selected miRNAs and the housekeeping U6 mRNA were assayed with qPCR to confirm the expression profiles identified with the microarray analysis. The expression patterns determined with qPCR agreed well with those determined from the microarray data (Fig. 2).

qPCR confirmation of the miRNA microarray data. qPCR confirmation of the miRNAs differentially expressed at 4 h after C. parvum infection (a). qPCR confirmation for the miRNAs differentially expressed at 12 h after C. parvum infection (b). Data are the means and standard errors of the means of triplicate experiments
Predicting the target genes of differentially expressed miRNAs
We predicted the target genes for all the miRNAs differentially expressed in HCT-8 cells after C. parvum infection. Target genes were predicted for only 18 of the 20 differentially expressed miRNAs using two different online software programs, TargetScan and miRDB.
GO analysis of differentially expressed miRNAs
To further understand the functions of the differentially expressed miRNAs, a GO analysis of the target genes was performed. DAVID gene annotation was applied to explain the biological effects of the differentially expressed miRNAs based on their target genes. The GO annotations of differentially expressed miRNAs (with P < 0.05) are shown in Fig. 3. The GO-specific functions mainly involved biological processes (e.g., transcription initiation from the RNA polymerase II promoter, apoptotic process, and immune response), cell components (e.g., membrane, actin cytoskeleton, focal adhesion, plasma membrane, cell surface, and apical part of the cell), and molecular functions (e.g., protein binding, ATP binding, calcium ion binding). Among the GO annotations (with P < 0.05), hsa-miR-34b-5p, hsa-miR-942-5p, hsa-miR-3591-3p, hsa-miR-18b-3p and hsa-miR-3976 were involved in the regulation of apoptotic or autophagy processes, hsa-miR-34b-5p and hsa-miR-3591-3p were involved in the regulation of the immune response (Additional file 1: Figure S1). Thus, the GO analysis of the differentially expressed miRNA target genes revealed that some have potentially important biological functions in regulating the host against C. parvum infection.

GO analysis of targets of differentially expressed miRNAs at 4 h and 12 h respectively. According to gene count, top 19 GO terms (P < 0.05) are shown. GO analysis of targets of differentially expressed miRNAs at 4 h (a). GO analysis of targets of differentially expressed miRNAs at 4 h (b)
KEGG pathway analysis of differentially expressed miRNAs
The main KEGG pathway annotations of the target genes (with P < 0.05) are shown in Table 3. Among these KEGG pathways, several were associated with the immune response (e.g., cytokine receptor interaction, chemokine signaling pathway, and inflammatory mediator regulation of TRP channels), apoptosis (e.g., MAPK signaling pathway and ERBB signaling pathway), cytoskeleton (e.g., actin cytoskeleton and WNT signaling pathway), and cell adhesion (e.g., cell adhesion molecules).
Regulatory network of miRNAs and target genes
To investigate the interactions between the miRNAs and their target genes, we performed an miRNA–target gene network analysis (Fig. 4). From the network of the miRNA and their target genes at 4 h after C. parvum infection (Fig. 4a), we found among seven differentially expressed miRNAs at 4 h hsa-miR-34b-5p and hsa-miR-3591-3p have a connection with six differentially expressed miRNAs. Among eleven differentially expressed miRNAs at 12 h, hsa-miR-942-5p, hsa-let-7b-3p and hsa-miR-6721-5p have a connection with ten differentially expressed miRNAs(Fig. 4b). According to these datasets, the key miRNAs among the differentially expressed miRNAs were hsa-miR-34b-5p, hsa-miR-3591-3p, hsa-miR-942-5p, hsa-let-7b-3p, and hsa-miR-6721-5p Their common target genes of the key miRNAs at 12 h were shown in Fig. 4c.

miRNA–target gene network. Network of miRNAs and their target genes at 4 h after C. parvum infection (a). Network of miRNAs and target genes at 12 h after C. parvum infection (b). Network of the key miRNAs and target genes at 12 h after C. parvum infection (c). In the diagram of the miRNA–gene network, the ellipses represent the miRNAs, the rectangles represent the target genes, and triangles represent the key target genes, with the interaction between them are represented by lines. The crucial miRNAs has the highest connectivity which refers to the number of edges possessed by one miRNA. The key genes are the common target genes of differentially expressed miRNAs
Discussion
Increasing evidence suggests that miRNAs regulate gene expression at the posttranscriptional level and thus affect a variety of biological processes, including the pathogenesis and progression of various diseases [30]. miRNAs also play important roles in the regulation of complex parasite–host interactions [10, 12, 14, 15]. Cryptosporidium parvum-induced miRNA expression in human biliary epithelial cells has already been investigated and analyzed [10]. However, there has been no report of the miRNA expression profile of human intestinal epithelial cells infected with C. parvum. Therefore, we analyzed the changes in the miRNA the profile of HCT-8 cells at 4 h and 12 h after C. parvum infection using microarray technology. Our results show that most miRNAs were not significantly differentially expressed in the infected HCT-8 cells compared to uninfected cells. However, some differentially expressed miRNAs were detected (four upregulated and 16 downregulated), implying that C. parvum infection alters the miRNA profile of intestinal epithelial cells. However, the differentially expressed miRNAs in the infected HCT-8 cells differed from those in biliary epithelial cells [10]. The changes in these specific miRNAs induced by C. parvum might be involved in the regulation of human intestinal epithelial cells in response to C. parvum infection. This is the first report to describe the alterations in miRNA expression induced by C. parvum infection in human intestinal epithelial cells, and extends our understanding of the interplay between C. parvum and its host at the miRNA level. The identification of the functions of these miRNA sheds new light on the molecular mechanisms triggered by C. parvum infection. Therefore, in this study, GO and KEGG analyses were used to identify the functions of the predicted target genes involved in C. parvum infection. Cell apoptosis is one of the ancient antiparasite strategies used by a host after its invasion by a parasite [31,32,33,34]. Research has shown that the early phase of C. parvum infection results in the apoptosis of HCT-8 cells [35]. However, miRNA-mediated cell apoptosis after C. parvum infection has not been reported. Our results suggest that the functions of hsa-miR-34b-5p, hsa-miR-942-5p, hsa-miR-18b-3p, hsa-miR-3976 and hsa-miR-3591-3p, most of which (not hsa-miR-942-5p) were downregulated after C. parvum infection, regulate apoptotic processes. Several reports have shown that hsa-miR-34b, hsa-miR-18a, and hsa-miR-942 regulate cell apoptosis in response to microbial infection. For example, upregulated hsa-miR-18a in macrophages after Toxoplasma gondii infection leads to the survival of the infected macrophages by targeting BIM [36]. Upregulated hsa-miR-30c-1 in human macrophages is involved in an anti-apoptosis response to T. gondii infection [37]. The downregulation of hsa-miR-34a induces cell apoptosis in Influenza A virus-infected A549 cells [38], and the downregulation of hsa-miR-942 enhances the apoptosis of HLCZ01 cells in response to Hepatitis C virus infection [39].
Other host antiparasite strategies are the immune responses [40,41,42]. As well as apoptosis, we found that hsa-miR-3591-3p and hsa-miR-34b-5p regulate the epithelial immune responses. Several reports have shown that hsa-miR-34a regulate host immune responses in response to microbial infection. For example, hsa-miR-34a negatively regulate the innate immune response to viral infections by targeting interferon β [43]. hsa-miR-3591-3p could increases interleukin 6 mRNA levels in THP-1 cells in response to lipopolysaccharide stimulation [44]. These data suggest that hsa-miR-3591-3p, hsa-miR-34b-5p, hsa-miR-942-5p, hsa-miR-18b-3p and hsa-miR-3976 may be involved in cell apoptosis or the immune responses to C. parvum infection. Hsa-miR-34b-5p, hsa-miR-3591-3p, hsa-miR-942-5p, hsa-let-7b-3p, and hsa-miR-6721-5p were also identified as the key miRNAs among the miRNAs differentially expressed after C. parvum infection. Therefore, the functions of hsa-let-7b-3p, hsa-miR-3591-3p, hsa-miR-34b-5p, hsa-miR-942-5p, hsa-miR-6721-5p, hsa-miR-18b-3p, hsa-miR-3976, and hsa-miR-3121-5p during C. parvum infection require further investigation.
There were some limitations to this study that should be addressed. First, the time points in the early phase of C. parvum infection was relatively limited and microRNA expression profile of HCT-8 cells in the later phase of C. parvum infection should be further analyzed to better understand the interplay between C. parvum and its host at the miRNA level. Second, we only assessed microRNA regulation in a single cell line, so the results should be confirmed with primary intestinal epithelial cells or another human intestinal epithelial cells line. Last, we should decrease the dose of oocysts used to infect the monolayer, because the high dose of oocysts may potentially have triggered a stress response in the host cell, the differentially regulated miRNA could reflect such a response and be unrelated to the actual C. parvum infection.
Conclusions
In this study, we determined the miRNA expression profile of human intestinal epithelial cells infected by C. parvum for the first time, and demonstrated that the dysregulation of these miRNAs may regulate the host’s biological processes in response to C. parvum infection, including cell apoptosis and the immune responses. These findings extend our understanding of the miRNA regulatory network that operates during the host–C. parvum interaction. They also provide clues for further research into the regulatory mechanisms of miRNAs in human intestinal epithelial cells in response to C. parvum infection, which may identify potential targets for future C. parvum control strategies.
Abbreviations
- DAVID:
-
Database for annotation, visualization and integrated discovery
- DMEM:
-
Dulbecco’s modified Eagle’s medium
- GO:
-
Gene Ontology
- KEGG:
-
Kyoto encyclopedia of genes and genomes
- SOCS:
-
Suppressor of cytokine signaling
References
Ryan U, Fayer R, Xiao L. Cryptosporidium species in humans and animals: current understanding and research needs. Parasitology. 2014;141(13):1667–85.
Hunter PR, Hadfield SJ, Wilkinson D, Lake IR, Harrison FC, Chalmers RM. Subtypes of Cryptosporidium parvum in humans and disease risk. Emerg Infect Dis. 2007;13(1):82–8.
Ryan U, Power M. Cryptosporidium species in Australian wildlife and domestic animals. Parasitology. 2012;139(13):1673–88.
Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the global enteric multicenter study, GEMS): a prospective, case–control study. Lancet. 2013;382(9888):209–22.
Striepen B. Parasitic infections: time to tackle cryptosporidiosis. Nature. 2013;503(7475):189–91.
Bueno MJ, Pérez de Castro I, Malumbres M. Control of cell proliferation pathways by microRNAs. Cell Cycle. 2008;7(20):3143–8.
Kedde M, Agami R. Interplay between microRNAs and RNA-binding proteins determines developmental processes. Cell Cycle. 2008;7(7):899–903.
O'Connell RM, Rao DS, Chaudhuri AA, Baltimore D. Physiological and pathological roles for microRNAs in the immune system. Nat Rev Immunol. 2010;10(2):111–22.
Piedade D, Azevedo-Pereira JM. The role of microRNAs in the pathogenesis of herpesvirus infection. Viruses. 2016;8(6):156–87.
Zhou R, Hu G, Liu J, Gong AY, Drescher KM, Chen XM. NF-kappaB p65-dependent transactivation of miRNA genes following Cryptosporidium parvum infection stimulates epithelial cell immune responses. PLoS Pathog. 2009;5(12):e1000681.
Staedel C, Darfeuille F. MicroRNAs and bacterial infection. Cell Microbiol. 2013;15(9):1496–507.
Chen XM, Splinter PL, O'Hara SP, LaRusso NF. A cellular micro-RNA, let-7i, regulates toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. J Biol Chem. 2007;282(39):28929–38.
Hu G, Gong AY, Roth AL, Huang BQ, Ward HD, Zhu G, Larusso NF, Hanson ND, Chen XM. Release of luminal exosomes contributes to TLR4-mediated epithelial antimicrobial defense. PLoS Pathog. 2013;9(4):e1003261.
Zhou R, Gong AY, Eischeid AN, Chen XM. miR-27b targets KSRP to coordinate TLR4-mediated epithelial defense against Cryptosporidium parvum infection. PLoS Pathog. 2012;8(5):e1002702.
Sato Y, Koshizuka T, Ishibashi K, Hashimoto K, Ishioka K, Ikuta K, Yokota SI, Fujii N, Suzutani T. Involvement of herpes simplex virus type 1 UL13 protein kinase in induction of SOCS genes, the negative regulators of cytokine signaling. Microbiol Immunol. 2017;61(5):159–67.
Hu G, Zhou R, Liu J, Gong AY, Chen XM. MicroRNA-98 and let-7 regulate expression of suppressor of cytokine signaling 4 in biliary epithelial cells in response to Cryptosporidium parvum infection. J Infect Dis. 2010;202(1):125–35.
Sheedy FJ, Palsson-McDermott E, Hennessy EJ, Martin C, O'Leary JJ, Ruan Q, Johnson DS, Chen Y, O'Neill LA. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat Immunol. 2010;11(2):141–7.
Verdon R, Keusch GT, Tzipori S, Grubman SA, Jefferson DM, Ward HD. An in vitro model of infection of human biliary epithelial cells by Cryptosporidium parvum. J Infect Dis. 1997;175(5):1268–72.
Hu M, Xia M, Chen X, Lin Z, Xu Y, Ma Y, Su L. MicroRNA-141 regulates Smad interacting protein 1 (SIP1) and inhibits migration and invasion of colorectal Cancer cells. Dig Dis Sci. 2010;55(8):2365–72.
Saito Y, Liang G, Egger G, Friedman JM, Chuang JC, Coetzee GA, Jones PA. Specific activation of microRNA-127 with downregulation of the proto–oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell. 2006;9(6):435–43.
Li R, Qian N, Tao K, You N, Wang X, Dou K. MicroRNAs involved in neoplastic transformation of liver cancer stem cells. J Exp Clin Cancer Res. 2010;29:169.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(–Delta Delta C(T)) method. Methods. 2001;25(4):402–8.
Agarwal V, Bell GW, Nam J, Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. elife. 2015;4:e05005.
Wong N, Wang X. miRDB: an online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015;43:D146–52.
Huang d W, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37:1–13.
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–504.
Pahl MC, Derr K, Gäbel G, Hinterseher I, Elmore JR, Schworer CM, Peeler TC, Franklin DP, Gray JL, Carey DJ, et al. MicroRNA expression signature in humanabdominal aortic aneurysms. BMC Med Genet. 2012;5:25.
Mokutani Y, Uemura M, Munakata K, Okuzaki D, Haraguchi N, Takahashi H, Nishimura J, Hata T, Murata K, Takemasa I, et al. Down-regulation of microRNA-132 is associated with poor prognosis of colorectal Cancer. Ann Surg Oncol. 2016;23(5):599–608.
Ji J, Shi J, Budhu A, Yu Z, Forgues M, Roessler S, Ambs S, Chen Y, Meltzer PS, Croce CM, et al. MicroRNA expression, survival, and response to interferon in liver cancer. N Engl J Med. 2009;361(15):1437–47.
Li J, Tan S, Kooger R, Zhang C, Zhang Y. MicroRNAs as novel biological targets for detection and regulation. Chem Soc Rev. 2014;43(2):506–17.
Chen XM, Levine SA, Tietz P, Krueger E, McNiven MA, Jefferson DM, Mahle M, LaRusso NF. Cryptosporidium parvum is cytopathic for cultured human biliary epithelia via an apoptotic mechanism. Hepatol. 1998;28(4):906–13.
McCole DF, Eckmann L, Laurent F, Kagnoff MF. Intestinal epithelial cell apoptosis following Cryptosporidium parvum infection. Infect Immun. 2000;68(3):1710–3.
Chen XM, Gores GJ, Paya CV, LaRusso NF. Cryptosporidium parvum induces apoptosis in biliary epithelia by a Fas/Fas ligand-dependent mechanism. Am J Phys. 1999;277(1):599–608.
Widmer G, Corey EA, Stein B, Griffiths JK, Tzipori S. Host cell apoptosis impairs Cryptosporidium parvum development in vitro. J Parasitol. 2000;86(5):922–8.
Liu J, Deng M, Lancto CA, Abrahamsen MS, Rutherford MS, Enomoto S. Biphasic modulation of apoptotic pathways in Cryptosporidium parvum infected human intestinal epithelial cells. Infect Immun. 2009;77(2):837–49.
Cai Y, Chen H, Mo X, Tang Y, Xu X, Zhang A, Lun Z, Lu F, Wang Y, Shen J. Toxoplasma gondii inhibits apoptosis via a novel STAT3- miR-17-92-Bim pathway in macrophages. Cell Signal. 2014;26(6):1204–12.
Cai Y, Chen H, Jin L, You Y, Shen J. STAT3-dependent transactivation of miRNA genes following Toxoplasma gondii infection in macrophage. Parasit Vectors. 2013;6:356.
Fan N, Wang J. MicroRNA 34a contributes to virus-mediated apoptosis through binding to its target gene Bax in influenza a virus infection. Biomed Pharmacother. 2016;83:1464–70.
Yang D, Meng X, Xue B, Liu N, Wang X, Zhu H. MiR-942 mediates hepatitis C virus–induced apoptosis via regulation of ISG12a. PLoS One. 2014;9(4):e94501.
McDonald V, Korbel DS, Barakat FM, Choudhry N, Petry F. Innate immune responses against Cryptosporidium parvum infection. Parasite Immunol. 2013;35(2):55–64.
Pantenburg B, Dann SM, Wang HC, Robinson P, Castellanos-Gonzalez A, Lewis DE, White AC Jr. Intestinal immune response to human Cryptosporidium sp. infection. Infect Immun. 2008;76(1):23–9.
Petry F, Jakobi V, Tessema TS. Host immune response to Cryptosporidium parvum infection. Exp Parasitol. 2010;126(3):304–9.
Witwer KW, Sisk JM, Gama L, Clements JE. MicroRNA regulation of IFN-beta protein expression: rapid and sensitive modulation of the innate immune response. J Immunol. 2010;184(5):2369–76.
Woo MY, Yun SJ, Cho O, Kim K, Lee ES, Park S. MicroRNAs differentially expressed in Behçet disease are involved in interleukin-6 production. J Inflamm. 2016;13:22.
Acknowledgments
The authors thank Fuchang Yu, Jianke Cao, and Junqiang Li for their help with the animal infection experiment.
Funding
This study was supported by the Key Program of the National Natural Science Foundation of China (31330079), the National Natural Science Foundation of China (31672548, 31302079), and National Key Research and Development Program of China (2017YFD0501305, 2017YFD0500405). The funding bodies had no role in the design of the study, collection, analysis, interpretation of data and in the writing of the manuscript.
Availability of data and materials
All the data supporting our findings is contained within the manuscript. Full details of the miRNA microarray analyses were deposited in the Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) public database with the associated platform accession number GPL23111. The raw data are available through GEO under Accession: GSE95365. The entire microarray data set was MIAME compliant.
Author information
Authors and Affiliations
Contributions
LoZ oversaw the entire project, including experimental design, reviewing the manuscript and submission, CW performed all experimental procedures including animal work, acquisition and analyses of data, drafting the manuscript, LL and HZ edited the manuscript and performed bioinformatic analyses, LuZ, ZZ, JH performed animal work and revised the manuscript, RW, FJ, SZ and CN performed bioinformatic and statistical analyses and contributed intellectually and critically to the manuscript. All the authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research was conducted according to the Chinese Laboratory Animal Administration Act (1988) after the review and approval of its protocol by the Research Ethics Committee of Henan Agricultural University. No specific permits were required, and no endangered or protected species were involved.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Figure S1. GO analysis of targets of differentially expressed miRNAs involved in the regulation of apoptotic processes and the immune response. (PDF 521 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wang, C., Liu, L., Zhu, H. et al. MicroRNA expression profile of HCT-8 cells in the early phase of Cryptosporidium parvum infection. BMC Genomics 20, 37 (2019). https://doi.org/10.1186/s12864-018-5410-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-018-5410-6