
Abstract
Aims
Zinc deficiency is a common micronutrient deficiency in plants growing in many different regions of the world and is associated with disturbances in uptake and accumulation of mineral nutrients. Despite many published data on physiological factors affecting ion accumulation in Zn deficient plants, there is very little information about the genetic factors underlying this. We aim to identify genetic loci involved in mineral accumulation and plant performance under Zn deficiency.
Methods
Genetic loci were identified using the genetically segregating Ler × Cvi recombinant inbred line (RIL) population grown under Zn deficient conditions. Lines were analysed for the concentrations of Zn, Fe, Mn, K, Ca, Mg, P, Cu, S and Al in shoot dry matter. The same was done for the same lines grown under Zn sufficient conditions.
Results
We found considerable heritable variation for most mineral concentrations. In general, there was a positive correlation between mineral concentrations. For Zn only condition-dependent QTLs were identified, while for most other mineral concentrations both condition-dependent and -independent QTLs were identified. Several QTLs co-localize, including co-localization to loci controlling shoot biomass and to mineral concentration loci found previously in this and other RIL populations.
Conclusions
There are different genetic loci controlling Zn accumulation under deficient and sufficient Zn supply. Only for few minerals, their accumulation is controlled by Zn-supply-specific loci.
Similar content being viewed by others

Introduction
Plants require essential minerals, including zinc (Zn), and are at the same time an important dietary source of essential minerals for humans. Zinc deficiency is a common micronutrient deficiency in plants growing in different regions of the world including India, China, Australia, Pakistan and Turkey (Alloway 2008; Cakmak 2000, 2008; Sillanpaeae 1982). Turkey is a major wheat-producing country, where nearly 45 % of the production area is located on highly calcareous soils in Central Anatolia (Cakmak et al. 1996). In this region, Zn deficiency represents a critical plant nutrition problem, substantially limiting wheat production (Cakmak et al. 1999a; Cakmak et al. 1996). Soil Zn deficiency in general affects both the nutritional quality (e.g. low Zn contents of seeds and grains) and the yield of food crops. It is therefore not surprising that soil Zn deficiency and human Zn deficiency are often closely associated (Cakmak 2008). Understanding the genetics of how plants deal with Zn deficiency, comprising the acquisition of Zn from the soil, distribution of Zn throughout the plant and storage of Zn, under Zn-limited conditions, while maintaining homeostasis of Zn and other mineral nutrients, is an important issue for breeding for Zn-efficient crops.
Zinc efficiency is defined as the ability of a plant genotype to maintain growth and yield well on a Zn-limited soil (Graham 1984). Different mechanisms are involved in Zn efficiency in plants, including enhanced Zn bioavailability in the rhizosphere by the release of root exudates, increased root Zn uptake and better internal Zn utilization (Cakmak et al. 1999b; Rengel et al. 1999). While differences in Zn efficiency have been determined for several crop species, there are relatively few studies assessing the genetic basis of these differences among genotypes of the same species in Zn deficient and -sufficient conditions (Genc et al. 2009; Wissuwa et al. 2006; Wu et al. 2007). A firm understanding of the genetic regulation of mineral homeostasis is also required to accelerate breeding for crop bio-fortification, which is a method of breeding crops for increased nutritional value (www.harvestplus.org) (Bouis 2002).
Quantitative trait locus (QTL) analysis is a powerful method to identify the genetic factors involved in controlling a trait (Alonso-Blanco et al. 2009). It provides information on the chromosomal location of the target loci without prior knowledge of the genes related to the trait, which can be conveniently used to identify QTLs affecting mineral homeostasis in crops (Ghandilyan et al. 2006). The A. thaliana Ler × Cvi Recombinant Inbred Line (RIL) immortal mapping population (Alonso-Blanco et al. 1998), derived from a cross between the Landsberg erecta (Ler) lab strain and the Cape Verde Islands (Cvi) accession, has been used extensively to study the genetic basis of multiple traits in Arabidopsis (recently reviewed by (Alonso-Blanco et al. 2009)) including mineral content in seeds (Vreugdenhil et al. 2004; Waters and Grusak 2008). QTL analyses of mineral concentrations in seeds, rosettes and roots were conducted in Arabidopsis Ler × Kond, Ler × An-1 and Ler × Eri populations grown under various conditions (Ghandilyan et al. 2009b).
In order to identify genetic factors involved in mineral homeostasis of Arabidopsis under natural Zn deficient conditions, the Ler × Cvi RIL population was grown on Zn deficient soil from Central Anatolia, supplemented with low or adequate Zn to complement the otherwise extreme deficiency, and mineral QTLs were determined.
Materials and methods
Plant material and growing conditions
Arabidopsis thaliana lab strain Landsberg erecta (Ler, N20; Nottingham Arabidopsis Stock Centre, www.arabidopsis.info) and accession Cape Verde Islands (Cvi, N8580; Nottingham Arabidopsis Stock Centre, www.arabidopsis.info), and the recombinant inbred line (RIL) population derived from the cross between these two accessions (RIL Ler × Cvi) (Alonso-Blanco et al. 1998), were used for the experiments. The parents and population were grown on naturally Zn deficient soil, with minor or adequate Zn supplementation, in a climate-controlled greenhouse. The Zn deficient and nutrient-poor soil originated from Eskisehir, Central Anatolia, Turkey. This was an alkaline (pH 8.1 in dH2O) and calcareous (12 % CaCO3) soil with a low organic matter content (1.1 %). The diethylenetriamine pentaacetic acid (DTPA)-extractable mineral concentrations were 0.13 mg.kg-1 for Zn, 1.84 mg.kg-1 for Fe, 4.91 mg.kg-1 for Mn, and 0.89 mg.kg-1 for Cu. The NaHCO3-extractable P concentration was 3.63 mg.kg-1, and the ammonium acetate-extractable concentrations of K and Mg were 478 mg.kg-1 and 424 mg.kg-1, respectively.
After three-day stratification at 5 °C, imbibed seeds were directly sown on to the prepared soil. The experiment was designed as a three-pot-replicate experiment in which each pot contained five plants. Replications were randomized within the plot. The soil was amended with essential nutrients prior to the sowing of the seeds: 200 mg/kg N as calcium nitrate [Ca(NO3)2], 100 mg/kg P as potassium di-hydrogen phosphate (KH2PO4), 25 mg/kg S as potassium sulfate (K2SO4) and 3 mg/kg Fe as Fe-EDTA. Pots were supplied separately with Zn (ZnSO4) as follows: 0.25 mg Zn/kg soil for low Zn conditions (Zn deficient) and 3 mg Zn/kg soil for adequate Zn conditions (Zn sufficient). Each pot contained 300 g of the fertilized soil. The soil surface of each pot was covered with perforated black nylon sheets. Each nylon sheet had five holes, uniformly dispersed. Several stratified and imbibed seeds were sown into each hole. The covers were used in order to reduce evaporative water loss from the soil and minimize soil contamination on plant shoots. The pots were placed in trays and watered from the bottom.
At harvesting time, the whole shoots, including the bolts if plants were bolting at the time of harvest were harvested and dried. Plant samples have been analyzed for mineral nutrients reported by inductively coupled plasma optical emission spectrometer (ICP-OES; Vista-Pro Axial; Varian Pty Ltd, Mulgrave, Australia) after their acid digestion in a closed-vessel microwave system. The details of the acid digestion and other steps are described by Aciksoz et al. (2011)
The Zn deficiency tolerance index (%) was determined by using the following equation: (shoot dry weight under Zn deficient conditions/shoot dry weight under Zn sufficient conditions) * 100. Zinc efficiency was calculated using the following equation: shoot Zn concentration under Zn deficiency growth conditions/shoot Zn concentration under Zn sufficient growth conditions. So-called KRAT ([K]/([Ca] + [Mg] Ratio) values were determined according to Larson and Mayland (2007). These are important for grazing animals, which stand the risk of grass tetany or hypomagnesaemia with KRAT values over 2.2. See Table 1 for abbreviations of all investigated traits.
Statistical tests and QTL mapping
For all statistical analyses, the statistical package SPSS version 15.0 was used. Differences in mean trait values of the genotypes were analysed by Univariate Analysis of Variance using the Dunnett’s pairwise multiple comparison t-tests in the General Linear model module of the package. For each analysis, trait values were used as dependent variables, and genotypes were used as fixed factors. Two-sided tests were performed with a significance threshold level of 0.05. Independent samples t-test of the package was used to determine mean differences between two individual lines. Correlation analyses were performed by calculating the Pearson or Spearman correlation coefficients.
The QTL mapping was performed using the computer program MapQTL version 5.0, which is based on composite interval mapping (http://www.kyazma.nl). LOD score thresholds distinguishing relevant QTLs are based on 1,000 permutation tests. Epistatic or QTL × QTL interactions occur when either the effect of one QTL is dependent on the presence of an allele at another locus or when each locus by itself appears to have no effect on the trait, yet when two loci are considered together there is an effect. A complete pairwise search for epistatic interactions for each trait (P < 0.001, determined by Monte Carlo simulations) was done using the EPISTAT statistical package (Chase et al. 1997).
Results
Variations in plant mineral concentrations
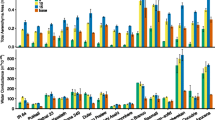
Shoot Zn, Fe, Mn, K, Ca, Mg, P, Cu, S and Al concentrations differed considerably between the RILs when grown on Zn deficient or Zn sufficient soil (Fig. 1). Shoot mineral concentrations varied 2.3- to 4.8-fold under Zn deficiency and 2.4- to 5.6-fold under Zn sufficiency (Table 2). As expected, shoot Zn concentrations were significantly lower under Zn deficiency when compared to the Zn sufficient conditions, while the concentrations of Mn, Ca, Mg were significantly higher under Zn deficiency. Zinc availability in the growth medium clearly affected the Zn status, but it did not significantly affect the Fe concentration (Fig. 2), even though Zn and Fe homeostasis are partly using similar transporters.

Frequency distributions of the concentrations of Zn, Fe, Mn, Cu, Al (in ppm) and K, Ca, Mg, P and S (in %) in shoots of the Ler × Cvi RILs grown on soil under Zn deficiency (light) and sufficiency (dark) conditions. Arrows indicate the levels in the Ler (thick arrow) and Cvi (thin arrow) parental lines

Average concentrations of shoot Zn, Fe, Mn, Cu, Al (in ppm) and K, Ca, Mg, P and S (in %) (± SE) of all Ler × Cvi RILs grown on soil under Zn deficiency (light) and Zn sufficiency (dark) conditions. * indicate significant differences (P < 0.05) between both Zn supply conditions
The fold difference for Zn, i.e. the concentration of Zn in the RIL with the highest concentration divided by concentration of Zn in the RIL with lowest Zn concentration, was higher under Zn deficiency (3.1) than under Zn sufficiency (2.5). For Fe, the fold difference was higher at sufficient Zn (5.6) compared to deficient Zn (3.4). These observations that mineral concentrations and fold differences differ between both Zn supply conditions, implies that there will be genetic variation for mineral concentration, which depends on genotype-environment interactions.
Correlation between traits
Positive correlations were found for all shoot mineral concentrations, except for Zn, when comparing Zn deficient and sufficient conditions (Table 3). This means that genetic factors controlling the homeostasis of minerals other than Zn are not severely affected by Zn availability in the growth substrate. Shoot Zn and Fe concentrations are positively correlated, and both are also positively correlated with shoot Cu concentrations. Shoot biomass (SB) is negatively correlated with the shoot Zn concentration, probably due to a dilution effect of Zn at increased growth and it is positively correlated with shoot K concentration, but not with Fe concentration. The correlations between shoot biomass and shoot mineral concentrations are similar for both growing conditions. The correlation between shoot biomass under Zn deficiency and Zn sufficiency is only moderate, suggesting a considerable genotype x environment interaction.
QTL analyses
The proportion of phenotypic variation in the population that is attributed to genetic variation was estimated by calculating the broad-sense heritability values for all traits (Fig. 3). Heritability values vary considerably for traits, with high heritability for Fe, Mn, Ca, K, Mg and P concentrations, and low heritability for S and Cu concentrations. The heritability values depend on Zn supply. For shoot Zn and Fe concentrations, heritability was higher under Zn sufficient conditions, whereas for Cu concentrations, it was higher under Zn deficient conditions. High heritability values are generally a good indication for the ability to detect QTLs controlling the traits, which is why a QTL analysis was performed of shoot mineral concentrations in response to Zn supply.

Heritability values (in percentages) and the percentage of the total phenotypic variance explained by identified QTLs (Total Explained Variance) for mineral accumulation and shoot biomass (SB) in the Ler × Cvi RIL population grown under Zn deficiency (“Deficiency”) or sufficiency (“Control”) conditions
At least one significant QTL was identified for each mineral concentration (Fig. 4, Tables 4 and 5). Whereas 12 QTLs were found to co-locate for the same mineral when comparing the two growth conditions, there were also 26 QTLs that were only detected in one condition. In addition to QTLs with additive effects, 21 epistatic interactions between loci were identified (Table 6). Most of these interactions had relatively small phenotypic effects, and often affected the concentration of one mineral, in only one of the QTL allele pair combinations (Table 6). The total phenotypic variances explained by the QTLs represented a relatively small part of the heritability values, which means only part of the genetic variation could be assigned to QTLs while the remainder went undetected. Two hotspots for co-locating QTLs were found, one at the top of chromosome 1, around marker AXR-1, and the other at the upper region of chromosome 5. Most of these co-located loci correspond to minerals for which shoot concentrations were found to be correlated (Table 3). The identified co-locations consisted of a mix of macro- and micro-elements.

Genetic map of the Ler × Cvi RIL population with identified QTLs (including 1- and 2-LOD confidence intervals; resp. thick and narrow bars) for shoot mineral concentrations of plants grown on soil under Zn deficiency (dark boxes, ending on “d”) and Zn sufficiency (light boxes, ending on “c”) conditions. See Table 1 for abbreviations of the traits
Based on K, Ca and Mg concentrations, KRAT values were determined. QTLs for KRAT values under Zn deficiency mapped to chromosome 4, while a QTL for KRAT values under Zn sufficiency mapped to chromosome 3. This is in accordance with the K, Ca and Mg concentration differences between different growth conditions, which resulted in significantly higher KRAT values under Zn sufficiency.
Discussion
Variations in shoot mineral concentrations between genotypes can have many reasons, since many biological processes affect shoot mineral accumulation and for each process several genes are involved. Minerals need to be mobilized in the rhizosphere, where some may have to be reduced in order to be taken up by roots; minerals are then stored in root cell vacuoles or loaded into xylem for transport to shoots; in the shoot, they will need to be distributed over different tissues, and sequestered to the designated organelles (Clemens 2001; Waters and Sankaran 2011). Considering that there are large variations in available nutrients in soils, plant genotypes have developed adaptive root mechanisms to accommodate optimal chemical availability and root uptake of nutrients (Marschner 2011). Throughout the plant, various ligands are present to chelate minerals for either detoxification or mobilization, such as organic acids (citrate, malate), phytochelatins, histidine or nicotianamine (Haydon and Cobbett 2007). In case of Zn deficiency stress, plant genotypes can activate root uptake and root-to-shoot transport of Zn and change root morphology for better Zn acquisition from soil (Broadley et al. 2007; Rengel 2001). Understanding the regulation and the differences in mineral homeostasis in plants grown under different growing conditions would help to identify relevant genes involved in Zn acquisition from soils and Zn accumulation in plant tissues, for further breeding or genetic modification purposes.
For the first time we used natural Zn deficient soil to investigate genetic variation for mineral concentrations in Arabidopsis, which is different from previous work using this model species to investigate the genetic basis of mineral homeostasis. Because of the high pH and the high CaCO3 level of the soil, mineral bioavailability in general is limited and without any additional nutrient amendment, the soil we used is too poor to allow proper plant growth. We expected that differences in Zn bioavailability in soil would cause differences in mobilization, uptake, translocation and sequestration levels of Zn and other minerals, considering the shared affinities of mineral transporters and chelates, eventually leading to different mineral concentrations in various plant tissues (Clemens et al. 2002; Maser et al. 2001). We used the well-known Arabidopsis Ler × Cvi immortal RIL mapping population to identify the genetic loci controlling regulation of mineral homeostasis under Zn deficiency and compare it with the genetics of mineral homeostasis under Zn sufficiency. This population has been used before for genetic analysis of mineral accumulation (Vreugdenhil et al. 2004; Waters and Grusak 2008), and thus allows the comparison of identified QTLs over different growing conditions and tissues.
Our results shows that the genetic regulation of shoot mineral concentrations differs considerably between growth conditions. The relatively low explained variances for most of the considered traits are probably due to the presence of many genetic factors, each with relatively small allelic effects. These are difficult to detect, given the size of the population. Several of the identified QTLs were growth condition specific. Major QTLs for shoot Zn concentrations were mapped to chromosomes 3 and 5, the locus mapping to the top of chromosome 3 affecting the shoot Zn concentration under sufficient supply and the locus on chromosome 5 involved in the shoot Zn concentration under Zn deficiency. As there was no obvious correlation between shoot Zn concentrations when comparing both growth conditions, and since the identified QTLs did not co-locate, there is specific genetic variation for genes controlling shoot Zn concentrations depending on Zn availability. Both of these loci co-locate with loci detected before in the same population for Zn concentration in seeds (Vreugdenhil et al. 2004; Waters and Grusak 2008) and in other populations for Zn concentration in seeds or rosette leaves (Ghandilyan et al. 2009a, b). It is tempting to conclude that this indicates genetic variation of the same genes, but it may also indicate different genes. Only cloning and confirmation of the cloned loci by reciprocal transformation of contrasting alleles, can verify this. The locus on chromosome 5 co-locates with QTLs for Fe, Mg and Ca concentrations, found for both Zn sufficient and deficient conditions. In the same region, QTLs were previously found for Fe and Mg concentrations in seeds (Waters and Grusak 2008). The confidence interval of the Zn concentration locus overlaps with those for seed Zn concentration QTLs previously found in the Ler × Cvi and Ler × An-1 populations (Ghandilyan et al. 2009b; Vreugdenhil et al. 2004), but since the QTL peaks are not mapping in close vicinity, this QTL appears to be different from the one detected previously. Overlap of this QTL with the Fe concentration QTL can indicate genetic variation for one gene involved in both Zn and Fe homeostasis, which is not unexpected as they share part of their molecular regulation (Colangelo and Guerinot 2004; van de Mortel et al. 2006). The locus on chromosome 3 appears to be more specific to Zn. It maps to a region close to the FRD3 gene (At3g08040; www.arabidopsis.org), but when fine-mapping the locus for Zn concentration in seeds (Vreugdenhil et al. 2004), FRD3 was not mapped to the fine-mapping interval (Du and Aarts, unpublished results).
Although the confidence intervals for the identified QTLs are large, there are several candidate genes in the intervals, that could account for the QTLs, e.g. FER2 (ferritin), MTP5 (a vacuolar cation importer), ZIP1 (a cell membrane Zn transporter), CAX2, CAX9 (low-affinity calcium transporters), CHX3, CHX9, CHX18, CHX19, CHX24 (cation/H+ exchangers), CNGC19, CNGC20 (ion channels), MGT3 (a magnesium transporter) and FRO2 (ferric reductase), but equally many, including the well-known HMA4 involved in Zn root to shoot translocation, that are not within the confidence intervals. However, without further fine-mapping it is not possible to indicate any of these as more or less likely candidates.
In general, different traits can be genetically correlated due to pleiotropy (one gene affecting different traits) or due to close linkage of different genes with different functions (Jiang and Zeng 1995). Since several correlations were found when comparing concentrations of different minerals, the co-locations of QTLs for these concentrations suggest some sharing of mechanisms controlling homeostasis. Not all QTLs identified in the present study co-located with the QTLs identified previously for Arabidopsis seeds in the same population (Vreugdenhil et al. 2004; Waters and Grusak 2008). The reason for this apparent discrepancy could be the difference in growing conditions (nutrient amended Zn deficient calcareous soil vs. peat-based potting soil) as well as the fact that regulation of mineral homeostasis depends on organ type as well (Ghandilyan et al. 2009b).
We found a negative correlation between shoot Zn concentrations and shoot biomass on Zn deficient and sufficient conditions. Previously, we also found a negative correlation between organ dry weight and Zn concentration in other populations and growing conditions (Ghandilyan et al. 2009b). The same phenomenon was observed in other species (Morgounov et al. 2007; Shi et al. 2008), and is probably caused by a dilution effect. When plants grow, they will have to spread the accumulated minerals over a larger volume, thus decreasing the mineral concentration. The same we found previously for other minerals, such as in the Ler × An-1 population, which was grown in water deficit and optimal conditions (Ghandilyan et al. 2009a), in which clusters of mineral concentration QTLs often co-segregated with dry weight QTLs. One obvious region for such co-segregation of QTLs is the top of chromosome 1, indicating QTLs for shoot biomass and for several mineral concentrations (Fig. 4). This region contains the CRY2 gene, encoding for the CRYPTOCHROME 2 blue light receptor (El-Assal et al. 2001). When comparing wild type and mutant plants, we found that this gene affects seed mineral concentrations via variation in yield or yield-associated parameters and flowering time (Ghandilyan and Aarts, unpublished results). Our observation that two major QTLs, for shoot Zn concentrations in both growing conditions, did not co-locate with the identified QTLs for shoot biomass, is assuring evidence that there are QTLs additional to growth QTLs that control shoot Zn accumulation.
The concentrations of other minerals was also affected by Zn deficiency, but these concentrations were less dependent on Zn availability. That is probably also the reason that many QTLs were found under both Zn sufficient and deficient conditions. One example is the QTL for shoot P concentration mapped to the top of chromosome 3. Previously, QTLs for phosphorus and phytate concentrations in different plant organs were detected at this position in several populations (Bentsink et al. 2003; Ghandilyan et al. 2009a, b; Vreugdenhil et al. 2004; Waters and Grusak 2008).
Overall, this analysis identified several shoot mineral concentration QTLs in Arabidopsis, adding to our knowledge on genetic variation for mineral homeostasis. Although some QTLs were found to be specific for one of the two Zn supply conditions, many were not, meaning that an alteration in Zn supply does not necessarily have a considerable effect on the homeostasis of other mineral nutrients.
References
Aciksoz S, Yazici M, Ozturk L, Cakmak I (2011) Biofortification of wheat with iron through soil and foliar application of nitrogen and iron fertilizers. Plant Soil 349:215–225
Alloway B (2008) Zinc in soils and crop nutrition. In IZA Publications
Alonso-Blanco C, Peeters AJ, Koornneef M, Lister C, Dean C, van den Bosch N, Pot J, Kuiper MT (1998) Development of an AFLP based linkage map of Ler, Col and Cvi Arabidopsis thaliana ecotypes and construction of a Ler/Cvi recombinant inbred line population. Plant J 14:259–271
Alonso-Blanco C, Aarts MGM, Bentsink L, Keurentjes JJB, Reymond M, Vreugdenhil D, Koornneef M (2009) What has natural variation taught us about plant development, physiology, and adaptation? Plant Cell 21:1877–1896
Bentsink L, Yuan K, Koornneef M, Vreugdenhil D (2003) The genetics of phytate and phosphate accumulation in seeds and leaves of Arabidopsis thaliana, using natural variation. Theor Appl Genet 106:1234–1243
Bouis HE (2002) Plant breeding: a new tool for fighting micronutrient malnutrition. J Nutr 132:491S–494
Broadley MR, White PJ, Hammond JP, Zelko I, Lux A (2007) Zinc in plants. New Phytol 173:677–702
Cakmak I (2000) Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol 146:185–205
Cakmak I (2008) Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant Soil 302:1–17
Cakmak I, Yilmaz A, Kalayci M, Ekiz H, Torun B, Erenoglu B, Braun HJ (1996) Zinc deficiency as a critical problem in wheat production in Central Anatolia. Plant Soil 180:165–172
Cakmak I, KalaycI M, Ekiz H, Braun HJ, KilInç Y, Yilmaz A (1999a) Zinc deficiency as a practical problem in plant and human nutrition in Turkey: a NATO-science for stability project. Field Crop Res 60:175–188
Cakmak I, Tolay I, Ozdemir A, Ozkan H, Ozturk L, Kling CI (1999b) Differences in zinc efficiency among and within diploid, tetraploid and hexaploid wheats. Ann Bot 84:163–171
Chase K, Adler FR, Lark KG (1997) EPISTAT: a computer program for identifying and testing interactions between pairs of quantitative trait loci. Theor Appl Genet 94:724–730
Clemens S (2001) Molecular mechanisms of plant metal tolerance and homeostasis. Planta 212:475–486
Clemens S, Palmgren MG, Krämer U (2002) A long way ahead: understanding and engineering plant metal accumulation. Trends Plant Sci 7:309–315
Colangelo EP, Guerinot ML (2004) The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell 16:3400–3412
El-Assal SE-D, Alonso-Blanco C, Peeters AJM, Raz V, Koornneef M (2001) A QTL for flowering time in Arabidopsis reveals a novel allele of CRY2. Nat Genet 29:435–440
Genc Y, Verbyla AP, Torun AA, Cakmak I, Willsmore K, Wallwork H, McDonald GK (2009) Quantitative trait loci analysis of zinc efficiency and grain zinc concentration in wheat using whole genome average interval mapping. Plant Soil 314:49–66
Ghandilyan A, Vreugdenhil D, Aarts MGM (2006) Progress in the genetic understanding of plant iron and zinc nutrition. Physiol Plant 126:407–417
Ghandilyan A, Barboza L, Tisné S, Granier C, Reymond M, Koornneef M, Schat H, Aarts MGM (2009a) Genetic analysis identifies quantitative trait loci controlling rosette mineral concentrations in Arabidopsis thaliana under drought. New Phytol 184:180–192
Ghandilyan A, Ilk N, Hanhart C, Mbengue M, Barboza L, Schat H, Koornneef M, El-Lithy M, Vreugdenhil D, Reymond M, Aarts MGM (2009b) A strong effect of growth medium and organ type on the identification of QTLs for phytate and mineral concentrations in three Arabidopsis thaliana RIL populations. J Exp Bot 60:1409–1425
Graham RD (1984) Breeding for nutritional characteristics in cereals. Advances in Plant Nutrition 1:57–102
Haydon MJ, Cobbett CS (2007) Transporters of ligands for essential metal ions in plants. New Phytol 174:499–506
Jiang C, Zeng ZB (1995) Multiple trait analysis of genetic mapping for quantitative trait loci. Genetics 140:1111–1127
Larson S, Mayland H (2007) Comparative mapping of fiber, protein, and mineral content QTLs in two interspecific Leymus wildrye full-sib families. Molecular Breeding 20:331–347
Marschner P (2011) Mineral nutrition of higher plants. Academic Press Inc. pp 672
Maser P, Thomine S, Schroeder JI, Ward JM, Hirschi K, Sze H, Talke IN, Amtmann A, Maathuis FJ, Sanders D, Harper JF, Tchieu J, Gribskov M, Persans MW, Salt DE, Kim SA, Guerinot ML (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126:1646–1667
Morgounov A, Gómez-Becerra HF, Abugalieva A, Dzhunusova M, Yessimbekova M, Muminjanov H, Zelenskiy Y, Ozturk L, Cakmak I (2007) Iron and zinc grain density in common wheat grown in Central Asia. Euphytica 155:193–203
Rengel Z (2001) Genotypic differences in micronutrient use efficiency in crops. Communications in Soil Science and Plant Analysis 32:1163–1186
Rengel Z, Batten GD, Crowley DE (1999) Agronomic approaches for improving the micronutrient density in edible portions of field crops. Field Crop Res 60:27–40
Shi R, Li H, Tong Y, Jing R, Zhang F, Zou C (2008) Identification of quantitative trait locus of zinc and phosphorus density in wheat (Triticum aestivum L.) grain. Plant Soil 306:95–104
Sillanpaeae M (1982) Micronutrients and the nutrient status of soils: a global study. FAO Soils Bulletin, no.48, Rome, Italy
van de Mortel JE, Almar Villanueva L, Schat H, Kwekkeboom J, Coughlan S, Moerland PD, Loren V, van Themaat E, Koornneef M, Aarts MGM (2006) Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol 142:1127–1147
Vreugdenhil D, Aarts MGM, Koornneef M, Nelissen H, Ernst WHO (2004) Natural variation and QTL analysis for cationic mineral content in seeds of Arabidopsis thaliana. Plant Cell Environ 27:828–839
Waters BM, Grusak MA (2008) Quantitative trait locus mapping for seed mineral concentrations in two Arabidopsis thaliana recombinant inbred populations. New Phytol 179:1033–1047
Waters BM, Sankaran RP (2011) Moving micronutrients from the soil to the seeds: Genes and physiological processes from a biofortification perspective. Plant Sci 180:562–574
Wissuwa M, Ismail AM, Yanagihara S (2006) Effects of zinc deficiency on rice growth and genetic factors contributing to tolerance. Plant Physiol 142:731–741
Wu J, Schat H, Sun R, Koornneef M, Wang X, Aarts MGM (2007) Characterization of natural variation for zinc, iron and manganese accumulation and zinc exposure response in Brassica rapa L. Plant Soil 291:167–180
Acknowledgments
We gratefully acknowledge the help of Corrie Hanhart in preparing the seeds of the RIL population and the support of COST Actions 859 (Phytotechnologies to promotesustainable land use and improve food safety) and FA0905 (Mineral improved crop production for healthy food and feed) to support this work with grants to AG and UBK. We thank Maarten Koornneef for critically reading this manuscript and for providing valuable comments for its improvement.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Michael A. Grusak.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Ghandilyan, A., Kutman, U.B., Kutman, B.Y. et al. Genetic analysis of the effect of zinc deficiency on Arabidopsis growth and mineral concentrations. Plant Soil 361, 227–239 (2012). https://doi.org/10.1007/s11104-012-1334-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-012-1334-0