
Abstract
Tardigrades in lichens have been poorly studied with few papers published on their ecology and diversity so far. The aims of our study are to determine the (1) influence of habitat heterogeneity on the densities and species diversity of tardigrade communities in lichens as well as the (2) effect of nutrient enrichment by seabirds on tardigrade densities in lichens. Forty-five lichen samples were collected from Spitsbergen, Nordaustlandet, Prins Karls Forland, Danskøya, Fuglesongen, Phippsøya and Parrøya in the Svalbard archipelago. In 26 samples, 23 taxa of Tardigrada (17 identified to species level) were found. Twelve samples consisted of more than one lichen species per sample (with up to five species). Tardigrade densities and taxa diversity were not correlated with the number of lichen species in a single sample. Moreover, the densities of tardigrades was not significantly higher in lichens collected from areas enriched with nutrients by seabirds in comparison to those not enriched. The incorporation of previously published data on the tardigrades of Spitsbergen into the analysis showed that tardigrade densities was significantly higher in moss than it was in lichen samples. We propose that one of the most important factors influencing tardigrade densities is the cortex layer, which is a barrier for food sources, such as live photosynthetic algal cells in lichens. Finally, the new records of Tardigrada and the first and new records of lichens in Svalbard archipelago are presented.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Terricolous lichens are one of the main components of the vegetation of polar regions (Węgrzyn et al. 2011). The Arctic climate, with its low annual temperatures, reduces vascular plant colonization (Olech et al. 2011). Lichens are pioneer organisms, and some of them have the ability to assimilate atmospheric nitrogen, which allows them to colonize areas completely devoid of vegetation due to a lack of nutrients in the soil (Millbank 1978; Rychert et al. 1978; Belnap 2002; Węgrzyn and Wietrzyk 2015). Moreover, lichens provide an important habitat for various invertebrates including water bear species (Ramazzotti and Maucci 1983; Nelson 2002; Nelson et al. 2015).
Tardigrada, also known as water bears, is a phylum, whose representatives inhabit various ecosystems, from the deepest seas to the highest parts of mountains (Ramazzotti and Maucci 1983; Kaczmarek et al. 2015). Terrestrial tardigrades inhabit mosses, lichens, hepatics, rosette-type higher plants, algae, leaf litter and soil, while aquatic species inhabit either freshwater or marine habitats (e.g., Ramazzotti and Maucci 1983; Dastych 1988). Currently, more than 1200 tardigrade species have been described (Degma and Guidetti 2007; Vicente and Bertolani 2013; Degma et al. 2009–2016).
Various aspects of the diversity of lichen-dwelling micrometazoans have been discussed by different authors (e.g., Gerson and Seaward 1977; Bates et al. 2012; Šatkauskienė 2012; Hauck et al. 2014). First, Gerson and Seaward (1977) presented a complex discussion of the associations between invertebrates and lichens. The authors underlined the poor knowledge of the interactions between these lichens and their accompanying biota (i.e., micrometazoans) and suggested avenues for future research. Recent papers concerning tardigrades in lichens include taxonomical, ecological or physiological aspects (e.g., Greven 1972; Morgan and Lampart 1986; Rebecchi et al. 2006; Guil et al. 2009). In the scientific browsers Web of Science and Scopus, only 67/81 papers use the keywords “Tardigrada” and “lichens,” which is low in comparison to the keywords “Tardigrada” and “moss,” with 121/143 papers (http://apps.webofknowledge.com/, https://www.scopus.com/ accessed on 21 July 2016). Although studies of Arctic tardigrades have taken place since the early the twentieth century (Scourfield 1897) and the first comprehensive monographs were published more than 3 decades ago (Marcus 1936; Ramazzotti and Maucci 1983), lichens are less studied than mosses in terms of their tardigrade ecology. Most of the previous ecological studies have focused on the distribution of tardigrades within moss samples (e.g., Nelson 1975; Nelson and Adkins 2001; Jönsson 2003; Degma et al. 2011; Guil et al. 2009) and only Morgan and Lampard (1986), Miller et al. (1988), Bartels and Nelson (2007) and Guil et al. (2009) have presented studies on the relationship between lichens and tardigrades. Morgan and Lampard (1986) found seven species of tardigrade in four species of lichen. Miller et al. (1988) found four species of tardigrade in eight species of lichen. Bartels and Nelson (2007) found 35 tardigrade species in lichens and estimated the species richness of water bears, providing a maximum estimation of 37–45 tardigrade species in lichens. Lastly, Guil et al. (2009) described the species diversity and abundance of tardigrades in lichens collected from trees and rocks and found 17 tardigrade species.
Assemblages of invertebrates in the Arctic are usually more abundant in seabird colony areas because of the higher nutrient content (Zmudczyńska-Skarbek et al. 2015). It is well known that seabirds in polar regions strongly influence the composition of the ground and in many ways enhance the abiotic characteristics of the soil (Myrcha and Tatur 1991; Simas et al. 2007). In the Arctic, this results in greater plant diversity in the areas of bird cliffs than in the places where birds are absent (Dubiel and Olech 1992; Węgrzyn et al. 2011). As a consequence, such areas of greater plant diversity and biomass, mainly mosses, also provide a habitat for numerous invertebrate species (Zmudczyńska et al. 2012), including Tardigrada (Zawierucha et al. 2016).
Ecological analyses of the tardigrades from the Arctic tundra have been conducted by Dastych (1985), Johansson et al. (2013), Zawierucha et al. (2015a, b) and Zawierucha et al. (2016), who surveyed the influence of altitude, type of bedrock, seabird guano and substratum on tardigrade distribution, abundance, and species diversity. In this article, we provide novel information about Arctic tardigrade densities, species diversity and distribution with respect to environmental variables in Svalbard lichen samples, thus increasing our knowledge about this poorly studied tardigrade habitat.
Materials and methods
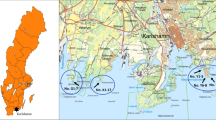
In total, we analyzed 45 samples collected in the summer of 2011 from Hornsund, Spitsbergen (Zawierucha et al. 2016), and in 2013 from Spitsbergen (Nissenfjella), Nordaustlandet, Prins Karls Forland, Danskøya, Fuglesongen, Phippsøya and Parrøya. Lichen samples were collected along a latitudinal gradient from the southern to the northern part of the Svalbard archipelago (Fig. 1), and samples were collected from 14 to 72 m a.s.l. Twenty samples were collected from areas influenced by seabird colonies (areas enriched in guano) and 23 samples from areas not influenced by seabirds. For two samples the presence/absence of seabird enrichment is unknown because this information was not noted during sampling. For a list of the samples and their GPS coordinates, see Online Resource 1.

Map of Svalbard with sampling sites
The materials were stored in small paper bags, which allowed them to dry slowly, and were then weighed within a precision of ±0.01 g.
Statistical analysis
The number of tardigrades found in each sample was extrapolated for 10 g of dry mass of the vegetation to calculate the total density. Pearson correlation coefficients were used to assess the relationship between the tardigrade species diversity (number of taxa) and their densities in the lichens (determined in this study). Incorporating previously published data (Zawierucha et al. 2015a), we used the non-parametric Mann–Whitney test to test for the difference in the total density of water bears between lichens collected from areas enriched in nutrients (N = 33) and areas not enriched (N = 23) by seabird colonies. The Mann–Whitney test was also used to check for differences in Tardigrada abundance between moss (N = 56) and lichen samples (N = 56). Mann–Whitney tests were used because of the distinct deviations from normal distributions of the data; Shapiro–Wilk test, p < 0.05. For this comparison we combined previous data published by Zawierucha et al. (2015a) and Zawierucha et al. (2016) and randomly selected moss samples for comparison. Statistical analyses were performed with STATISTICA 10.0 (StatSoft, Inc., 2011).
Species identification
Tardigrada
All samples were treated following the standard methods for tardigrade examination (e.g., Dastych 1980; Ramazzotti and Maucci 1983; Stec et al. 2015). Each sample was soaked in water for ca. 5 h. Water containing tardigrades and detritus particles was then decanted and examined under a stereomicroscope. All extracted specimens, exuviae and eggs were mounted on microscope slides in Hoyer’s medium and were then examined with a phase contrast microscope (PCM) (Olympus BX41) associated with an ART-CAM 300 MI photo camera and QuickPhoto Camera 2.3 software. Species were identified with the keys of Ramazzotti and Maucci (1983) and later original descriptions, re-descriptions and keys of Dastych (1985, 1988), Binda (1988), Fontoura and Pilato (2007), Kaczmarek and Michalczyk (2009) and Michalczyk et al. (2012a b). Nomenclature and new generic names follow Bertolani et al. (2014) and Vecchi et al. (2016). All the microscope slides were deposited in the Department of Animal Taxonomy and Ecology at Adam Mickiewicz University in Poznań, Poland.
Lichens
Lichens were identified by traditional taxonomic methods and standard microscopy. Cladonia species were determined using thin-layer chromatography (TLC) in solvent C, following the technique of Orange et al. (2001). Apothecial handmade sections were mounted in water, and their anatomical structures were measured in ethyl alcohol. Spot test reactions for apothecial pigments were examined in KOH and NaClO. For species determination, the following guides were used: Thomson (1984, 1997), Brodo et al. (2001), Smith et al. (2009) and Wirth (2013). The nomenclature follows the MycoBank Fungal Database (MycoBank 2015).
Results
In total, 321 specimens of Tardigrada belonging to 23 taxa (with 17 determined to species level) were found in 26 samples (42% of all samples collected). Analyzed lichen material included 20 lichen taxa (Online Resource 1). Due to the lack of tardigrade eggs, some of the specimens were determined to the species group level (see Online Resource 2). The most frequent species in the samples were Hypsibius pallidus Thulin, 1911 (present in nine samples), Diphascon pingue pingue (Marcus, 1936) (present in four samples) and Adropion prorsirostre (Thulin, 1928) (present in four samples). The highest number of tardigrades calculated per 10 g of dry material was 720 specimens (for samples analyzed in this study), which were found in a mixed lichen sample of Cladonia arbuscula (Wallr.) Flot. and Cladonia rangiferina (L.) Weber ex F.H. Wigg. The highest tardigrade diversity was found in two samples, each with four Tardigrada species (see Online Resource 2). It is worth noting that lichens with the highest tardigrade densities have a characteristic cottony-tomentose thallus surface composed of loose hyphae, and thus they do not have a typical cortical layer (e.g., Cladonia arbuscula, C. mitis, C. rangiferina). The average number of tardigrades calculated per 10 g of dry material for samples was 66 individuals.
Twenty samples were collected from areas that were enriched in nutrients because of seabird colonies, with 13 of these samples containing tardigrades (ca. 65 %). In comparison, 23 samples were collected from areas that were not influenced by seabirds, with 11 of these samples containing tardigrades (ca. 47.8 %) (see Online Resource 2). For two samples, data on the influence of guano were not available. Including previously published data, tardigrades in lichens were not more abundant in the samples collected from areas that were enriched in nutrients because of seabird colonies than in the samples collected from non-enriched areas (Mann–Whitney, p = 0.13). Furthermore, regardless of the nutrient status of lichens, tardigrades were more abundant in moss samples (Mann–Whitney, p < 0.005) (Fig. 2). The diversity of lichens species in single samples ranged from one to five species. Twelve of the 45 lichen samples consisted of more than one lichen species. Tardigrade densities and taxa diversity were not significantly related to the number of lichen species in a single sample (Pearson, p = 0.72, r = − 0.05 and p = 0.82, r = 0.04, respectively).

Comparison of total Tardigrada density between vegetation types (abundances per 10 g of dry material). The error bar intervals represent the 95% confidence interval for the estimate of the mean (based on the standard error)
Statistics (mean, minimum, maximum and standard error) of tardigrade density for combined lichen samples analyzed during this study as well as those of Zawierucha et al. (2015a) are presented in Table 1.
Tardigrades for the islands of Danskøya, Fuglesongen, Phippsøya and Parrøya are new records (Online Resource 2). The presences of tardigrades A. prorsirostre, D. p. pingue and H. pallidus from Prins Karls Forland, A. prorsirostre, Echiniscus merokensis merokensis Richters, 1904, and Macrobiotus crenulatus Richters, 1904, from Nordaustlandet are new records for these islands (see Online Resource 2). The first record of the lichen Stereocaulon nanodes Tuck. from Svalbard is reported, along with new records of lichens from Prins Karls Forland (Cladonia amaurocrocea, C. macroceras, C. mitis, C. rangiferina), Danskøya (Cladonia crispata var. cetrariformis), Fuglesongen (Cetraria hepatizon, Cladonia crispata var. cetrariformis, Sphaerophorus globosus, Stereocaulon alpinum) and Phippsøya (Cladonia crispata var. cetrariformis); see Online Resource 1.
Discussion
Lichens are pioneers, but are also the most fragile and fastest disappearing organisms in the Arctic because they are affected by trampling and winter grazing by reindeer and face competition from vascular plants (Cornelissen et al. 2001; Storeheier et al. 2002; Węgrzyn et al. 2011). This study is an attempt to investigate factors such as habitat heterogeneity and nutrient enrichment of lichens by seabirds in terms of tardigrade density and species diversity in lichen thalli. Morgan and Lampard (1986) previously presented a study on the relationship between supralittoral lichens and tardigrades from Great Cumbrae Island in the Firth of Clyde, UK, and found seven species of tardigrades in four species of lichen along seashore. Unfortunately, the authors did not present their methods of calculation for the tardigrades in the samples; thus a comparison with the present study is not possible. Bartels and Nelson (2007) focused on species diversity estimators and predicted a lower number of tardigrade species in lichens than in mosses in Great Smoky National Park, USA. Guil et al. (2009) reported that Echiniscus species are more abundant in mosses and lichens collected from rocks than in leaf litter or in mosses and lichens collected from trees in the Sierra de Guadarrama, Central Spain. Moreover, Guil et al. (2009) noted significantly higher abundance and species richness in lichens collected from rocks because of the contribution of heterotardigrades (Guil et al. 2009). In our study the most frequent water bears were eutardigrades, and only three (not frequent) heterotardigrade species were found.
Despite the fact that lichens can constitute the only source of energy in cold desert areas, such as polar regions (Gerson and Seaward 1977), previous studies on Arctic tardigrades either did not focus on species diversity and ecology in lichens or noted a low number of specimens in lichens (e.g., Zawierucha et al. 2016). Studies conducted on the distribution and abundance of tardigrades in the Hornsund fjord (Spitsbergen) showed that one of the most significant factors influencing tardigrade abundance was vegetation type, with mosses having the most abundant individuals (Zawierucha et al. 2016).
Faunistic data for Svalbard are available only from limited areas (Węglarska 1965; Kaczmarek et al. 2012). Until now, extensive faunistic data on tardigrades in the Spitsbergen have only been published by Dastych (1985) and for the Svalbard archipelago by Zawierucha et al. (2013), with the authors analyzing different kinds of habitat samples, but predominantly mosses. The most frequently recorded species in the samples collected from our study were H. pallidus (present in 9 out of 26 samples) and D. p. pingue and A. prorsirostre (both present in 4 out of 26 samples) (Online Resource 2). Bartels and Nelson (2007) found 35 taxa of tardigrades in lichens collected from Great Smoky National Park. In contrast, only five of the same species were recorded in our study: Adropion s. scoticum (Murray, 1905), Diphascon p. pingue (Marcus, 1936), Hypsibius dujardini (Doyère, 1840), Platicrista angustata (Murray, 1905) and Mesobiotus harmsworthi harmsworthi (Murray, 1907). Guil et al. (2009) found 17 and 14 species in lichens collected from rocks and trees, respectively. Echiniscus bisetosus, E. canadensis, E. m. merokensis and R. oberhaeuseri were highly abundant in lichen samples from Spain (Guil et al. 2009). Only one species, E. m. merokensis, was recorded in both Svalbard and Spain. Tardigrade species composition in different biogeographical zones may be influenced by kind of substratum, general climatic conditions (e.g., humidity and temperature) or topographic features.
In the Arctic, seabirds are the link between marine and terrestrial ecosystems. They feed in the sea and nest on the land where they produce guano, which results in greater plant diversity in the areas surrounding bird colonies (Isaksen and Gavrilo 2000; Stempniewicz et al. 2007). Jennings (1976a, b) showed that in samples collected around vertebrate guano deposits from seabirds and seals, tardigrade densities are higher than in areas not affected by these nutrients in the Antarctic. However, Porazinska et al. (2002) and Smykla et al. (2012) showed that plant diversity and thus tardigrade abundance are decreased on flat regions of the Antarctic because of the high-salinity guano and trampling by penguins. Conversely, in the Arctic, seabird guano positively affected the biomass of mosses in which tardigrade abundance is higher than it is in lichens (Zawierucha et al. 2016). The positive influence of the Arctic seabirds on invertebrate densities and biomass has also been confirmed for other invertebrate groups (e.g., Zmudczyńska et al. 2012, 2015). However, to the best of our knowledge comparative tardigrade abundance in lichens with and without guano has never been studied previously. Here we show that the effect of seabird nutrients on tardigrade abundance in lichens was not significant and tardigrades were more abundant in mosses than in lichens.
The 12 collected samples consisted of more than 1 lichen species per sample (with up to 5 species per sample). In previous ecological studies, the diversity of invertebrates increased with the heterogeneity of the environment on various scales, e.g., forest age classes and complex and simple forms of plants (Niemelä et al. 1996; Benton et al. 2003; Taniguchi et al. 2003). But here, the tardigrade species diversity and abundance were not correlated to the number of lichen species in a single sample. It is possible that the biodiversity of tardigrades is affected by the availability of food sources, such as live photosynthetic algal cells in lichens. Water bears may feed on living, photosynthesizing eukaryotic cells (e.g., Altiero and Rebecchi 2001; Guidetti et al. 2012). Therefore, it is possible that the greater number of water bears noted in bryophytes than in lichens may be associated with easier access to photosynthetic cells in mosses. Alternatively, it may be related to the shape of the lichen (e.g., foliose, fruticose or crustose) and exposure to sunlight.
Conclusion
The results of this study underline the difficulties of surveying habitat associations of tardigrade species with single species of lichens in the Arctic. The 12 collected samples consisted of more than 1 lichen species per sample (with up to 5 species per sample). Contrary to expectation, the tardigrade taxa diversity and densities did not increase with the number of lichen species in a single sample. Additionally, we show that the nutrients from seabird colonies did not influence tardigrade densities in lichens; generally, tardigrades were more abundant in mosses than in lichens. In our study, we have presented higher numbers of tardigrades associated with some of the lichens, mainly Cladonia arbuscula, C. mitis and C. rangiferina (see Online Resource 1). These lichens, in addition to S. alpinum, do not have a thick surface layer and thus potentially provide the tardigrades with easier access to the photobiont layer and algal cells of the lichen. Usually, the most abundant tardigrades within samples are herbivores, with buccal tube morphology as discussed by Guidetti et al. (2012) and Guil and Sanchez-Moreno (2013). Thus, the availability of algal cells could influence tardigrade abundance in lichens. Nevertheless, factors such as pH, AMP (anti-microbial proteins) and the presence of other invertebrates (predators) in the sample should also be considered in future studies of the High Arctic lichen tardigrade assemblages.
References
Altiero T, Rebecchi L (2001) Rearing tardigrades: results and problems. Zool Anz 240:217–221. doi:10.1078/0044-5231-00028
Bartels PJ, Nelson DR (2007) An evaluation of species richness estimators for tardigrades of the Great Smoky Mountains National Park, Tennessee and North Carolina, USA. J Limnol 66:104–110
Bates ST, Berg-Lyons D, Lauber CL, Walters WA, Knight R, Fierer N (2012) A preliminary survey of lichen associated eukaryotes using Pyrosequencing. Lichenologist 44(1):137–146
Belnap J (2002) Nitrogen fixation in biological soil crusts from southeast Utah, USA. Biol fert soils 35(2):128–135
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18(4):182–188
Bertolani R, Guidetti R, Marchioro T, Altiero T, Rebecchi L, Cesari M (2014) Phylogeny of Eutardigrada: New molecular data and their morphological support lead to the identification of new evolutionary lineages. Mol Phylogenet Evol 76:110–126
Binda MG (1988) Redescrizione di Macrobiotus echinogenitus Richters, 1904 e sul valore di buona specie di Macrobiotus crenulatus Richters, 1904 (Eutardigrada). Animalia 15:201–210
Brodo IM, Sharnoff SD, Sharnoff S (2001) Lichens of North America. Yale University Press, New Haven
Cornelissen JHC, Callaghan TV, Alatalo JM, Michelsen A, Graglia E, Hartley AE, Hik DS, Hobbie SE, Press MC, Robinson CH, Henry GHR, Shaver GR, Phoenix GK, Gwynn Jones D, Jonasson S, Chapin FS, Molau U, Neill C, Lee JA, Melillo JM, Sveinbjörnsson B, Aerts R (2001) Global change and arctic ecosystems: is lichen decline a function of increases in vascular plant biomass? J Ecol 89(6):984–994
Dastych H (1980) Niesporczaki (Tardigrada) Tatrzańskiego Parku Narodowego. Monografie Fauny Polski 9:1–232
Dastych H (1985) West Spitsbergen Tardigrada. Acta Zool Crac 28:169–214
Dastych H (1988) The tardigrada of Poland. Monografie Fauny Polski 16:1–255
Degma P, Guidetti R (2007) Notes on the current checklist of tardigrada. Zootaxa 1579:41–53
Degma P, Bertolani R, Guidetti R (2009–2016) Actual checklist of Tardigrada species. http://www.tardigrada.modena.unimo.it/miscellanea/Actual%20checklist%20of%20Tardigrada.pdf, pp. 40. Accessed 03 May 2015
Degma P, Katina S, Sabatovičová L (2011) Horizontal distribution of moisture and Tardigrada in a single moss cushion. J Zool Syst Evol Res 49:71–77
Dubiel E, Olech M (1992) Ornithocoprophilous plant communities of the southern slope of Ariekammen (Hornsund region, Spitsbergen) Landscape, Life World and Man in High Arctic. Institute of Ecology PAS, Warszawa. pp 167–175
Fontoura P, Pilato G (2007) Diphascon (Diphascon) faialense sp. n. a new species of Tardigrada (Eutardigrada, Hypsibiidae) from Azores and a key to the species of the D. pingue group. Zootaxa 1589:47–55
Gerson URI, Seaward MRD (1977) Lichen—invertebrate associations. In: Seaward MRD (ed) Lichen ecology. Academic Press, London, pp 70–112
Greven H (1972) Tardigraden des nördlichen Sauerlandes. Zool Anz 189:368–381
Guidetti R, Altiero T, Marchioro T, Sarzi Amade L, Avdonina AM, Bertolani R, Rebecchi L (2012) Form and function of the feeding apparatus in Eutardigrada (Tardigrada). Zoomorphology 131:127–148
Guil N, Sanchez-Moreno S (2013) Fine–scale patterns in micrometazoans: tardigrade diversity, community composition and trophic dynamics in leaf litter. Syst Biodivers 11:181–193
Guil N, Sánchez-Moreno S, Machordom A (2009) Local biodiversity patterns in micrometazoans: are tardigrades everywhere? Syst Biodivers 7:259–268
Hauck M, Dulamsuren C, Bayartogtokh B, Ulykpan K, Burkitbaeva DU, Otgonjargal E, Titov SV, Enkhbayar T, Sundetpaev AK, Beket U, Leuschner C (2014) Relationships between the diversity patterns of vascular plants, lichens and invertebrates in the Central Asian forest-steppe ecotone. Biodivers Conserv 23:1105–1117
Isaksen K, Gavrilo MV (2000) Little Auk Alle alle. In: Anker-Nilssen T, Bakken V, Strøm H, GolovkinAN, Bianki VV, Tatarinkova I P (eds) The status of marine birds breeding in the barents sea region. Norsk Polarinstitutt Rapportserie Nr. 113, Norwegian Polar Institute, Tromsø
Jennings PG (1976a) The Tardigrada of Signy Island, South Orkney Islands, with a note on the Rotifera. Brit Antarcts Surv B 44:1–25
Jennings PG (1976b) Tardigrada from the Antarctic Peninsula and Scotia Ridge region. Brit Antarcts Surv B 44:77–95
Johansson C, Miller WR, Linder ET, Adams BJ, Boreliz-Alvarado E (2013) Tardigrades of Alaska: distribution patterns, diversity and species richness. Polar Res 32:1–11
Jönsson KI (2003) Population density and species composition of moss-living tardigrades in a boreo-nemoral forest. Ecography 26:356–364
Kaczmarek Ł, Zawierucha K, Smykla J, Michalczyk Ł (2012) Tardigrada of the Revdalen (Spitsbergen) with the descriptions of two new species: Bryodelphax parvuspolaris (Heterotardigrada) and Isohypsibius coulsoni (Eutardigrada). Polar Biol 35:1013–1026
Kaczmarek Ł, Michalczyk Ł, McInnes SJ (2015) Annotated zoogeography of non-marine Tardigrada. Part II: South America. Zootaxa 3923(1):1–107
Kaczmarek Ł, Michalczyk Ł (2009) Redescription of Hypsibius microps Thulin, 1928 and H. pallidus Thulin, 1911 (Eutardigrada: Hypsibiidae) based on the type material from the Thulin collection. Zootaxa 2275:60–68
Marcus E (1936). Tardigrada. Das Tierreich 66:1–340
Michalczyk Ł, Wełnicz W, Frohme M, Kaczmarek Ł (2012a) Redescriptions of three Milnesium Doye`re, 1840 taxa (Tardigrada: Eutardigrada: Milnesiidae), including the nominal species for the genus. Zootaxa 3154:1–20
Michalczyk Ł, Wełnicz W, Frohme M, Kaczmarek Ł (2012b) Corrigenda of Zootaxa 3154: 1–20 Redescriptions of three Milnesium Doye`re, 1840 taxa (Tardigrada: Eutardigrada: Milnesiidae),including the nominal species for the genus. Zootaxa 3154:1–20
Millbank JW (1978) The contribution of nitrogen fixing lichens to the nitrogen status of their environment. Environmental role of nitrogen-fixing blue-green algae and asymbiotic bacteria. Ecol Bull 26:260–265
Miller J, Horne P, Heatwole H, Miller WR, Bridges L (1988) A survey of the terrestrial Tardigrada of the Vestfold Hills, Antarctica. Hydrobiologia 165:197–208
Morgan CI, Lampard DJ (1986) Supralittoral lichens as a habitat for tardigrades. Glasgow Nat 21:127–138
MycoBank (2015) MycoBank. http://www.mycobank.org/. Accessed 15 March 2015
Myrcha A, Tatur A (1991) Ecological role of the current and abandoned penguin rookeries in the land environment of the maritime Antarctic. Pol. Polar Res 12:3–24
Nelson DR (1975) Ecological distribution of Tardigrades on Roan Mountain, Tennessee—North Carolina. In: Higgins RP (ed.) Proceedings of the first international symposium on Tardigrades. Mem Ist ital Idrobiol 32:225–276
Nelson DR (2002) Current status of the Tardigrada: evolution and ecology. Integr Comp Biol 42:652–659 doi:10.1093/icb/42.3.652
Nelson DR, Adkins RG (2001) Distribution of tardigrades within a moss cushion: do tardigrades migrate in response to changing moisture conditions? Zool Anz 240:493–500
Nelson DR, Guidetti R, Rebecchi L (2015) Phylum Tardigrada. In: Thorp J, Rogers DC (eds) Ecology and General Biology: Thorp and Covich’s Freshwater Invertebrates, Academic Press, London, pp. 347–380
Niemelä J, Haila Y, Punttila P (1996) The importance of small-scale heterogeneity in boreal forests: variation in diversity in forest-floor invertebrates across the succession gradient. Ecography 19:352–368
Olech M, Węgrzyn M, Lisowska M, Słaby A, Angiel P (2011) Contemporary changes in vegetation of polar regions. Papers on Global Change. IGBP 18:35–51
Orange A, James PW, White FJ (2001) Microchemical Methods for the Identification of Lichens. London, British Lichen Society, 101
Porazińska DL, Wall DH, Wirginia RA (2002) Invertebrates in ornithogenic soils on Ross Island, Antarctica. Polar Biol 25:569–574
Ramazzotti G, Maucci W (1983) II Phylum Tardigrada (III. Edizione riveduta e aggiornata). Mem Ist Ital Idrobiol 41:1–1016
Rebecchi L, Guidetti R, Borsari S, Altiero T, Bertolani R (2006) Dynamics of long-term anhydrobiotic survival of lichen-dwelling tardigrades. Hydrobiologica 558:23–30
Rychert RC, Skujins J, Sorensen D, Porcella D (1978) Nitrogen fixation by lichens and free-living microorganisms in deserts. In: West NW, Skujins J (eds) Nitrogen in desert ecosystems 9. Dowden, Hutchinson, Ross, Stroudsburg, pp. 20–30
Šatkauskienė I (2012) Microfauna of Lichen (Xanthoria parietina) in Lithuania: diversity patterns in polluted and non-polluted sites. Balt For 18:255–262
Scourfield DJ (1897) Contributions to the non-marine fauna of Spitsbergen. Part I. Preliminary notes, and reports on the Rhizopoda, Tardigrada, Entomostraca, & c. Proceedings of the Zoological Society of London 65:784–792.
Simas FNB, Schaefer CEGR, Melo VF, Albuquerque-Filho MR, Michel RFM, Pereira VV, Gomes MRM, Costa LM (2007) Ornithogenic cryosols from Maritime Antarctica: Phosphatization as a soil forming process. Geoderma 138:191–203
Smith CW, Aptroot A, Coppins BJ, Fletcher A, Gilbert OL, James PW, Wolseley PA (2009) The Lichens of Great Britain and Ireland. The British Lichen Society, Department of Botany, The Natural History Museum. ISBN 978 09540418 8 5. Londyn.
Smykla J, Iakovenko N, Devetter M, Kaczmarek Ł (2012) Diversity and distribution of tardigrades in soils of Edmonson Point (Northern Victoria Land, continental Antarctica). Czech Polar Rep 2:61–70
StatSoft, Inc. Team (2011) STATISTICA (data analysis software system), version 10. Oklahoma. StatSoft Inc, Tulsa
Stec D, Smolak R, Kaczmarek Ł, Michalczyk Ł (2015) An integrative description of Macrobiotus paulinae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: hufelandi group) from Kenya. Zootaxa 4052(5):501–526
Stempniewicz L, Błachowiak-Samołyk K, Węsławski JM (2007) Impact of climate change on zooplankton communities seabird populations and arctic terrestrial ecosystem—a scenario. Deep Sea Res Part 2 Top Stud Oceanogr 54:2934–2945. doi:10.1016/j.dsr2.2007.08.012
Storeheier PV, Mathiesen SD, Tyler NJ, Olsen MA (2002) Nutritive value of terricolous lichens for reindeer in winter. Lichenologist 34(3):247–258
Taniguchi H, Nakano S, Tokeshi M (2003) Influences of habitat complexity on the diversity and abundance of epiphytic invertebrates on plants. Freschwater biol 48(4):718–728
Thomson JW (1984) American arctic lichens I: the macrolichens. New York, Columbia University Press, p 512.
Thomson JW (1997) American arctic lichens II: the microlichens. University of Wisconsin Press, Madison, p 675
Vecchi M, Cesari M, Bertolani R, Jönsoon IK, Guidetti R (2016) Integrative systematic studies on tardigrades from Antarctica identify new genera and new species within Macrobiotoidea and Echiniscoidea. Invertebr Syst 30(4):303–322
Vicente F, Bertolani R (2013) Considerations on the taxonomy of the Phylum Tardigrada. Zootaxa 3626:245–248
Węglarska B (1965) Die Tardigraden (Tardigrada) Spitzbergens. Act Zool Cracov 11:43–51
Węgrzyn M, Wietrzyk P (2015) Phytosociology of snowbed and exposed ridge vegetation of Svalbard. Polar Biol 38 (11):1905–1917
Węgrzyn M, Lisowska M, Olech M, Osyczka P (2011) Changes in vegetation. In: Ziaja W. (ed) Transformation of the natural environment in Western Sørkapp Land (Spitsbergen) since the 1980s. Jagiellonian University Press, Kraków, pp 69–81
Wirth W (2013) Die Flechten Deutschland. Eugen Ulmer, Stuttgart
Zawierucha K, Coulson J, Michalczyk Ł, Kaczmarek Ł (2013) Current knowledge on the Tardigrada of Svalbard with the first records of water bears from Nordaustlandet (High Arctic). Polar Res 32:20886. doi:10.3402/polar.v32i0.20886
Zawierucha K, Smykla J, Michalczyk Ł, Gołdyn B, Kaczmarek Ł (2015a) Distribution and diversity of Tardigrada along altitudinal gradients in the Hornsund, Spitsbergen (Arctic). Polar Res 34:24168. doi:10.3402/polar.v34.24168
Zawierucha K, Cytan J, Smykla J, Wojczulanis-Jakubas K, Kaczmarek Ł, Kosicki JZ, Michalczyk Ł (2015b) Seabird guano boosts body size of water bears (Tardigrada) inhabiting the Arctic tundra. Polar Biol 38:579–582
Zawierucha K, Zmudczyńska-Skarbek K, Kaczmarek Ł, Wojczulanis-Jakubas K (2016) The influence of a seabird colony on abundance and species composition of water bears (Tardigrada) in Hornsund (Spitsbergen, Arctic). Polar Biol 39:713–723
Zmudczyńska K, Olejniczak I, Zwolicki A, Iliszko L, Convey P, Stempniewicz L (2012) Influence of allochtonous nutrients delivered by colonial seabirds on soil collembolan communities on Spitsbergen. Polar Biol 35:1233–1245
Zmudczyńska-Skarbek K, Zwolicki A, Convey P, Barcikowski M, Stempniewicz L (2015) Is ornithogenic fertilisation important for collembolan communities in Arctic terrestrial ecosystems? Polar Res 34:25629. 10.3402/polar.v34.25629
Acknowledgements
We are grateful to journal editor Dieter Piepenburg for valuable remarks that improved the manuscript. We are also grateful to three anonymous reviewers for their comments on the manuscript. We want to thank Jakub Kosicki (Adam Mickiewicz University in Poznań) for statistical consultation. Special thanks go to Ariel Deutsch (Brown University, USA) and Marta Kołowrotkiewicz for proofreading the article. We are grateful to Adam Nawrot (forScience Fundation) for helping during sampling. Sampling was conducted under permit RIS no. 6390. The study was supported by the Polish Ministry of Science and Higher Education via the ‘‘Diamond Grant’’ program (grant no. DIA 2011035241 to KZ). KZ is a beneficiary of National Science Center scholarship for PhD no. 2015/16/T/NZ8/00017.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Zawierucha, K., Węgrzyn, M., Ostrowska, M. et al. Tardigrada in Svalbard lichens: diversity, densities and habitat heterogeneity. Polar Biol 40, 1385–1392 (2017). https://doi.org/10.1007/s00300-016-2063-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-016-2063-2