
Abstract
Despite efforts to control late blight in potatoes by introducing R pi -genes from wild species into cultivated potato, there are still concerns regarding the durability and level of resistance. Pyramiding R pi -genes can be a solution to increase both durability and level of resistance. In this study, two resistance genes, R Pi-mcd1 and R Pi-ber, introgressed from the wild tuber-bearing potato species Solanum microdontum and S. berthaultii were combined in a diploid S. tuberosum population. Individual genotypes from this population were classified after four groups, carrying no R pi -gene, with only R Pi-mcd1, with only R Pi-ber, and a group with the pyramided R Pi-mcd1 and R Pi-ber by means of tightly linked molecular markers. The levels of resistance between the groups were compared in a field experiment in 2007. The group with R Pi-mcd1 showed a significant delay to reach 50% infection of the leaf area of 3 days. The group with R Pi-ber showed a delay of 3 weeks. The resistance level in the pyramid group suggested an additive effect of R Pi-mcd1 with R Pi-ber. This suggests that potato breeding can benefit from combining individual R pi -genes, irrespective of the weak effect of R Pi-mcd1 or the strong effect of R Pi-ber.
Similar content being viewed by others


Introduction
Late blight, caused by Phytophthora infestans, is a ruthless disease causing great losses in potato yield. One of the solutions has been the use of chemicals. The International Potato Centre (CIP) estimates that about 1 billion dollars per year are spent on fungicides to control late blight in the US, Europe and developing countries (http://gilb.cip.cgiar.org/what-is-late-blight/economic-impact/social-impact-and-economic-importance-of-late-blight/). The loss in yield is nevertheless still considerable, and in addition, the environment suffers from the use of fungicides. In the 1900s, the first successful breeding efforts were undertaken to raise resistance levels of potato varieties by hybridization with wild potatoes (Salaman 1910). Eleven R pi -genes (R1–R11), introgressed from S. demissum, were the first ones to be used in potato cultivars. But unfortunately soon after the introduction of many potato cultivars with S. demissum R-genes in the 1950s and 1960s, the resistance proved not to be durable. For a period of time, the prevailing strategy to achieve a durable solution against P. infestans was thought to be the use of horizontal (race-non-specific) resistance instead of R-genes in potato breeding (Colon et al. 1995; Landeo et al. 1995; Tan et al. 2008; Turkensteen 1993). Potato genotypes showing a hypersensitive response to Phytophthora races were discarded, because the non-durable R-genes would only blur the view on the level of horizontal resistance. Breeding for late blight resistance would require “R pi -gene free” germplasm (Turkensteen 1993). This strategy has not provided any improvement, mostly because of the inevitable correlation between resistance and late maturity (Simko 2002). Furthermore, it might be a valid assumption that residual effects of defeated R pi -genes could be a factor in horizontal resistance (Stewart et al. 2003; Rauscher et al. 2008; Tan et al. 2008). These days the focus is back on R pi -gene mediated resistance (Allefs et al. 2005). Since the discovery of the S. demissum R-genes, a wide range of wild Solanum species have been identified as potential sources for more R pi -genes, e.g. species in the Dutch–German potato collection (Ross and Baerecke 1951; Van Soest et al. 1984). Eventually, the resistance genes R Pi-ber from S. berthaultii (Ewing et al. 2000), R Pi-mcd1 from S. microdontum (Tan et al. 2008) and four R pi -genes from S. bulbocastanum: RB/R Pi-blb1, R Pi-blb2, R Pi-blb3 and R Pi-abpt (Park et al. 2005a, b; Song et al. 2003; van der Vossen et al. 2003, 2005) have been characterised. Furthermore R pi -genes similar to R pi-blb1 have also been found in S. verrucosum (Liu and Halterman 2006) and in S. stoloniferum and S. papilla (Vleeshouwers et al. 2008). Currently, these R pi -genes are being deployed in breeding programs and one (R Pi-blb2) is present in the commercial potato varieties “Toluca” and “Bionica” after more than 40 years of traditional crossing and breeding efforts (Hermsen 1966). However, not all of the R pi -genes render absolute levels of resistance in the practical field situation. Worse is the expected durability of these newly identified R pi -genes, as there is no reason to assume a longer durability of the new R pi -genes than of R1–R11, especially since these new R pi -genes have to oppose faster evolution rates of P. infestans with the introduction of the A2 mating type in the 1980s.
Pyramiding (major) R-genes can be one solution to improve on both durability and level of resistance (Nelson 1972). Pyramiding is the accumulation of (R)-genes into a single genotype or cultivar and can be achieved using major R pi -genes, defeated R pi -genes, different alleles of one gene, or the same alleles (allele-dosage). Although the potato cultivars such as Pentland Dell (1961) and Escort (1982) demonstrate that breeders were involved in pyramiding a long time ago, a surprisingly limited number of scientific reports have been published quantifying the effects of pyramided R-genes in different plant-pathosystems. Barloy et al. (2007) showed a higher level of resistance against cereal cyst nematodes in wheat when pyramiding resistance genes CreX and CreY. Several groups have reported on the pyramiding of bacterial blight resistance genes in rice and observed higher resistance levels and obtained additionally, a broader spectrum of resistance (Huang et al. 1997; Singh et al. 2001; Yoshimura et al. 1995; Zhang et al. 2006). When pyramiding multiple QTLs involved in resistance against barley stripe rust, Richardson et al. (2006) reported higher levels of resistance as achieved with individual QTLs. Whereas most studies show an additive effect of pyramiding two R-genes, the study of Sharma et al. (2004) did not. Marker-assisted pyramiding of the brown planthopper resistance genes Bph1 and Bph2 on rice chromosome 12 resulted in a resistance level of the pyramided line equivalent to that of the Bph1-single introgression line.
Slightly different are the examples of studies on the effect of allele-dosage. Toxopeus (1957) observed no additional P. infestans resistance in duplex R 3 or triplex R 3 genotypes compared to simplex R 3 genotypes. Likewise, no effect of allele dosage was observed in a study with H 1 , an R-gene against Globodera rostochiensis in potato, where numbers of developed cysts were equal on genotypes containing simplex or multiplex copies of the H 1 gene (Brodie and Plaisted 1992).
In this paper, we describe a potato field trial in which the effect of pyramiding two P. infestans R-genes on late blight resistance is studied. The two R-genes R Pi-ber (chromosome 10) and R Pi-mcd1 (chromosome 4) used for pyramiding have been described in literature. R Pi-mcd1 is described in Tan et al. (2008). To study the effect of pyramiding, it is necessary that both used R pi -genes do not provide absolute resistance. From our own experience with R Pi-mcd1, an R-gene originating from S. microdontum, we knew that this R-gene gives a weak resistance to P. infestans in the field (Tan et al. 2008). Since both R-genes do not give absolute resistance, we deemed them suitable for studying putative quantitative differences in resistance when pyramiding these R-genes.
We developed a pyramiding population by crossing two diploid parents that were both heterozygous for one of the R pi -genes. In theory, the full-sib offspring can be divided into four groups, carrying no R pi -gene, with only R Pi-mcd1, with only R Pi-ber, and a group with the pyramided R Pi-mcd1 and R Pi-ber. The advantage of making one pyramiding population instead of comparing cultivars with different and combined R pi -genes, is that the genetic background of the groups in one population is on average the same. The four groups cannot be differentiated by late blight assays, unless the different resistance spectra (if any) of the R pi -genes can be used. However, because the genetic position of both R-genes R Pi-ber and R Pi-mcd1 are known, we could make use of molecular markers flanking the R-genes. These molecular markers proved an accurate tool to genotype the pyramiding population and assign the genotypes into the four groups: (1) without R pi -genes, (2) with R Pi-mcd1 present, (3) with R Pi-ber present, and (4) containing both R pi -genes R Pi-mcd1 + R Pi-ber.
Materials and methods
Plant material and Phytophthora resistance assays
Two segregating diploid populations were developed. The first population RH97-739 (see Fig. 1) was developed to verify the map position of R Pi-ber and to develop flanking DNA markers. The second population RH03-424 (see Fig. 2) was developed to identify four groups of descendants without, with either or both R pi -genes.

Pedigree of RPi-ber mapping population RH97-739 used to develop flanking markers

Pedigree of pyramiding population RH03-424 which combines none, either or both the R pi -genes from Solanum microdontum and S. berthaultii
The first population, which was used to characterise the R Pi-ber gene used in this experiment was derived from S. berthaultii gene bank accession CGN 17823/PI265858. This gene might differ from the R pi -gene from the S. berthaultii gene bank accession PI473331 (Ewing et al. 2000 and Rauscher et al. 2006). To verify that the position of both R pi -genes (R Pi-ber1 and R Pi-ber) is identical, a disease assay was performed with the interspecific mapping population RH97-739 (Fig. 1). Eighty-eight genotypes were tested in a field experiment in 2005, as well as in a detached leaf assay, whereas the population size for marker analysis was 79 descendants. The field assay was performed in two replications, in a randomised complete block design. Inoculation took place with a spore suspension of a complex P. infestans isolate IPO-82001 (race 1,2,3,4,5,6,7,10,11) as described by Vleeshouwers et al. (1999). Infection was scored 18 and 30 days post inoculation (dpi). The genotypes were scored in a qualitative fashion at 30 dpi and were classified as either resistant when infection was ≤50% of the average leaf area or susceptible when ≥95% of the average leaf area was infected. The genotypes that fell in-between <50 and >95% were excluded from the qualitative genetic analysis. The detached leaf assay was performed with leaves collected from greenhouse plants and were tested in two replications as described by Vleeshouwers et al. (1999). The cultivar Bintje and parents RH91-172-2 and 94-2031-01 were included as control plants. Complex isolates IPO-82001 (race 1,2,3,4,5,6,7,10,11) and IPO-90128 (race 1,3,4,6,7,8,10,11) were used to determine leaf resistance. Inoculation was performed as described in Vleeshouwers et al. (1999) except that the concentration of the inoculum was adjusted to 5 × 105 zoospores/ml. In case of discrepancy between field and detached leaf assay results the field data were deemed most reliable.
The second population, also referred to as the pyramiding population RH03-424 (n = 93), descended from a cross between R Pi-mcd1 donor RH90-038-21, a BC1 of S. microdontum accession BGRC 18302 (CGN21342), and R Pi-ber donor 94-2031-01, a BC1 of S. berthaultii accession BGRC 10063 (CGN 17823/PI265858). The pedigree of RH03-424 is presented in Fig. 2. The resistance gene R Pi-mcd1 is located on the short arm of chromosome 4 and gives a delay of infection to P. infestans (Tan et al. 2008).
In 2007, a field assay was performed on 93 individuals of pyramiding population RH03-424. The location of the field was the “Hoge Born”, in the area near Wageningen, NL on sandy soil. Experimental design was a randomised complete block design, in two replications. Per replication, 4 tubers per genotype were planted. Astarte, Bildtstar and Eersteling were used as standards. Tubers were planted on the 17th of April. Inoculation took place on the 26th of June with a spore suspension of a complex P. infestans isolate IPO-82001 (race 1,2,3,4,5,6,7,10,11) as described by Vleeshouwers et al. (1999). Foliage infection levels were scored 15 times during 6.5 weeks. The percentage of diseased leaf area was recorded on a scale comprising 16 classes, corresponding to 0, 1, 2, 5, 10, 20,…, 80, 90, 95, 99, 100 percent diseased leaf tissue. The four plants per genotype were scored as one experimental unit.
Maturity type was observed on a fungicide protected field experiment, on clay soil, in August of 2006 and 2007. Maturity type was scored approximately 130 days after planting on an ordinal scale ranging from 2 (green and vigorous; late to very late) to 6 (yellowing and plant is sagging) to 9 (dead; very early plant types). Four plants per genotype were treated as one experimental unit. Observations from 2006 and 2007 were averaged.
Maturity corrected resistance (MCR) was obtained essentially as described by Visker et al. (2003), where MCR is estimated from the initial resistance value using maturity as co-variate. MCR thus represents the difference between the observed resistance value and the resistance value as expected given its maturity. When resistance and maturity data from this population or from potato cultivars from the Dutch National List are plotted in a graph, the regression slope of their correlation is approximately −1. This means that one unit gain on the maturity scale is decreasing late blight resistance with one unit (Visker 2005). Therefore, MCR values (or residuals) can be obtained easily by subtracting the maturity value from the resistance value. To display MCR values in a graph along with uncorrected resistance, the MCR values were increased with a constant value that equals the average maturity of the population. (MCR = foliage infection − maturity value + average maturity).
Genomic DNA extraction
Genomic DNA was isolated from young leaf material, harvested from young greenhouse grown plants, as described by van der Beek et al. (1992). Fresh tissue was ground at room temperature in STE-extraction buffer with steel balls using a Retsch machine (Retsch Inc., Haan, Germany). All volumes of the DNA extraction procedure were adapted to 2 ml deep 96-wells Costar plates (Corning Inc., Corning, NY, USA).
DNA marker assays
The verification and mapping of the R Pi-ber gene was performed with CAPS markers CT214 (Rauscher et al. 2006) and TG63 and SCAR marker Q133. TG63F2/R2 primers were newly developed on TG63 DNA sequence of a genotype of S. berthaultii accession BGRC 10063 (CGN 17823/PI265858) amplified with TG63F1/R primers (Rauscher et al. 2006). Marker Q133, was newly developed using DNA sequence information (Genbank AF404451) of a Resistance Gene Analogue which maps to chromosome 10 of tomato (Pan et al. 2001). Genotyping of the resistance locus R Pi-mcd1 was performed with chromosome 4 specific CAPS marker TG339 (Tan et al. 2008). All primers were developed with Primer3 software (Rozen and Skaletsky 2000). Primers and PCR conditions are listed in Table 1. Subsequently, the flanking molecular markers were tested for usability on the parents of the pyramiding population because it might be possible that polymorphisms found in one population cannot be used in other populations. However, all markers were usable for genotyping RH03-424 with the restriction enzymes identified in the R Pi-mcd1 or R Pi-ber mapping population. The molecular markers allowed the pyramiding population to be divided into four groups: (1) without R pi -genes, (2) with R Pi-mcd1 present, (3) with R Pi-ber present, and (4) containing both R pi -genes R Pi-mcd1 + R Pi-ber pyramided.
Linkage map construction of R Pi-ber
The marker order of chromosome 10 specific markers Q133, TG63, CT214 including R Pi-ber was determined by RECORD (van Os et al. 2005) and visually inspected by graphical genotyping the raw data in MS-Excel using the conditional formatting of cells conditional to marker genotype and linkage phase. Map distances were calculated based on the frequency of recombination between the markers, and in view of the short distances no adjustment for putative double cross-overs was made.
Statistical analysis
Analyses were performed with MS-Excel and comprised the chi-square test for goodness-of-fit and the analysis of group differences in resistance level in the pyramiding population using a 2-sided t test assuming unequal variances.
Results
Mapping of R Pi-ber
The response of the genotypes of the R Pi-ber mapping population RH97-793 in the field assay could be scored in a discrete fashion, where 49 genotypes could be scored as resistant and 37 as susceptible. The vast majority of the susceptible clones reached infection within 18 dpi, whereas the resistant clones were largely without symptoms. Only at 30 dpi did the first few resistant genotypes start to display infection (see Fig. 3). Two genotypes could not be scored unambiguously. The detached leaf assay showed the same segregation pattern as compared with the field assay, except for four differences, probably misclassifications due to mildew infection in the detached leaf assay. Therefore, subsequent analyses have been based on results of the field assay. Chromosome 10 specific marker loci Q133, CT214 and TG63 are closely linked with the locus for resistance, at distances of 6.4, 5.1 and 1.3 cM, respectively, north of R Pi-ber. The order and distances of the markers towards R Pi-ber was determined by five recombinants, and one singleton marker observation.

Distribution of the percentage of infected leaf area at 30 days past infection of the offspring genotypes of the mapping population segregating for R Pi-ber. From 17 until 30 dpi the U-shaped distribution allows unambiguous classification of resistant and susceptible segregants
One of the five recombinants suggested the position of TG63 to be located north of R Pi-ber. While taking this marker order, another descendant, with an ambiguous resistance phenotype then resulted in a singleton. Only the alternative marker order with TG63 south of R Pi-ber would remove this friction, but would cause map friction in the recombinant with an unambiguous late blight phenotype. Therefore, the marker order as shown in Fig. 4 is the most plausible order as judged from the marker data.

Genetic map of part of chromosome 10 of paternal resistant parent 94-2031-01 including the location of R Pi-ber
Analysis of the pyramiding population RH03-424 with markers flanking R Pi-mcd1 and R Pi-ber
To identify which of either resistance genes are present in each offspring of the pyramiding population (n = 93), molecular markers flanking the R pi -genes were used. The pyramiding population RH03-424 could be differentiated into four groups: (1) without R pi -genes, (2) with R Pi-mcd1 present, (3) with R Pi-ber present, and (4) containing both R pi -genes R Pi-mcd1 + R Pi-ber pyramided (Table 2). The observed segregation pattern into the four groups deviated significantly from the expected 1:1:1:1 ratio (χ 2 = 16.7, P < 0.001). This deviation is largely due to an overrepresentation of resistant genotypes having the R Pi-ber allele (63:30), whereas the R Pi-mcd1 locus hardly deviates from a 1:1 segregating ratio 56:37 (χ 2 = 3.88; P = 0.05).
Evaluation of the resistance level in the offspring of the pyramiding population
The results described below will first address the effect of the individual genes (a) R Pi-mcd1 and (b) R Pi-ber, respectively, (c) their interaction and (d) the effects after correction for maturity type.
R pi -gene free group versus RPi-mcd group
One week after inoculation, the R pi -gene free group had an average foliage infection of 66% (Fig. 5a). The group of descendants with the R Pi-mcd1 allele showed infection as well, but to a lesser degree with 46% infection at this time point. The difference between the groups with and without the R Pi-mcd1 allele was significant from 7 until 14 dpi (2-sided t test, df = 24–27, P < 0.05), with the largest effect at 9 dpi (P = 0.0027). The difference was no longer significant at 17 dpi and later time points, when both groups with or without R Pi-mcd1 approached 100% infection.

Late blight disease progress curve showing the proportion of leaf area infected across 45 days post infection (dpi) of the four groups of pyramiding population RH03-424; without R pi -genes, with R Pi-mcd1 present, with R Pi-ber present, and containing both R pi -genes R Pi-mcd1 + R Pi-ber. Error bars indicate the standard error of the mean. a Uncorrected disease progress using values as observed in the field. b Disease progress values using maturity corrected resistance (MCR) values
Resistance level of R Pi-ber
The difference in infection level between the groups with or without the R Pi-ber allele is much larger compared to the effect caused by the R Pi-mcd1 allele (Fig. 5a). When the foliage of plants from the R pi -gene free group is more than 90% infected, the R Pi-ber bearing groups remain almost free from Phytophthora symptoms, with an average degree of infection below 5%. This absence of Phytophthora infection as the effect of the R Pi-ber allele, remains until 24 dpi, a full 3 weeks longer than the R pi -gene free group, as well as the R Pi-mcd1 group. R Pi-ber does not, however, provide absolute resistance. At 24 dpi, these plants also start to show late blight infection. In this experiment, the genotypes with R Pi-ber passed the level of 50% infection on average at 36 dpi, 6 times later than the R pi -gene free genotypes and 4.5 times later than genotypes with R Pi-mcd1.
Effect of pyramiding R Pi-mcd1 + R Pi-ber
From the first observation date, the presence of the R Pi-ber allele had a more noticeable effect as compared to the presence of the R Pi-mcd1 allele. The effect of pyramiding of the two resistance genes can only be observed when the plants having the R Pi-ber allele (with or without R Pi-mcd1) also showed infection. A significant contribution (2-sided t test, df = 45–48, P < 0.05) of the R Pi-mcd1 allele in plants with R Pi-ber was observed from 35 dpi and remains significant till 37 dpi, with the largest effect observed at day 35 (p = 0.026) (Fig. 5a). Considering that the genetic background of the R Pi-ber group and the pyramided groups are on average the same, we conclude that the difference in resistance level was due to the additive effect of the R Pi-mcd1 allele. The magnitude of this effect is comparable with the difference observed between the R pi -gene free group and the R Pi-mcd1 group: 50% infection was achieved 3 days later in the pyramided group than in the group containing only the R Pi-ber allele.
The effect of plant maturity on late blight infection
Within each group, there is considerable variation in the level of infection per genotype, where some genotypes containing the R Pi-ber allele may have a level of infection which is lower than some genotypes in the pyramided group. The infection of both the R Pi-ber groups begins at 24 dpi, which is 3 months after planting, long enough for first early cultivars to end their life cycle. Maturity types in the pyramiding population varied between 4 and 9 with a mean of 6.9 on a total scale of 1 to 9. Differences in maturity type could potentially influence the level of late blight resistance. The strong correlation between P. infestans foliage resistance and late plant maturity resulted in the use of maturity corrected resistance (MCR) (Bormann et al. 2004; Visker et al. 2004). Therefore, we re-analysed our late blight resistance observations using a maturity corrected dataset, where late blight resistance is compensated for the effect of maturity. MCR was calculated in this study by subtracting maturity values from leaf area infection values, but adding up the average value for maturity of the population, leaving the residuals. This is a simple and valid approach, because analysis of variance components showed that the variance contributed by maturity is of the same magnitude as the variance contributed by the R pi -gene composition.
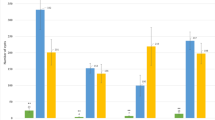
In Fig. 6, the effect is illustrated of this correction for maturity on late blight infection at 35 dpi. When comparing the maturity uncorrected (Fig. 6a) versus the maturity corrected (Fig. 6b) analysis, it becomes apparent that the correction for maturity slightly improves the difference in resistance levels between the R Pi-ber group and the R Pi-ber + R Pi-mcd1 group. In addition, the correction for maturity reduced the standard error of the group mean. Removing the heterogeneity in maturity between plants within the four groups thus allowed a more accurate assessment of late blight resistance differences due to R pi -gene composition. The correction for maturity also confirmed that quantitative variation in the observed level of infection was not only caused by R pi -genes but in part caused by maturity. Lastly, the maturity corrected analysis showed that the effect of pyramiding remains statistically significant for a much longer period, from 31 until 39 dpi (9 days) instead of 3 days (2-sided t test, df = 47–50, P < 0.05), with the largest effect at 35 dpi (P = 0.0014). In Fig. 5b, the progress in time of the maturity corrected foliage infection for the four groups is illustrated.

Foliage infection (in %) of offspring genotypes classified according to R pi -genes (a) Group averages of foliage infection at 35 dpi. b Group averages of maturity corrected resistance (MCR) at 35 dpi. Error bars indicate the standard error of the mean
Discussion
The individual R pi -genes used in this study have been described before in the literature. The phenotypic effects of resistance based on R Pi-mcd1 are described in Tan et al. (2008). It was then estimated that R Pi-mcd1 causes a delay of infection of about 1–2 weeks. In this experiment though, genotypes with R Pi-mcd1 pass the level of 50% infection on average 3 days later than R pi -gene free genotypes. The shorter delay of infection by R Pi-mcd1 in this experiment is likely caused by the relatively cold and humid weather favouring rapid late blight development in the summer of 2007.
Rauscher et al. (2006) mapped R Pi-ber 5.1 cM south of marker CT214 and 0.6 cM north of marker TG63. The resistance we observed in this study mapped to chromosome 10, close to marker TG63, but 1.3 cM south of marker TG63. Although the position of marker TG63 relative to R Pi-ber differs in this study with the marker order shown by Rauscher et al., we do not wish to draw strong conclusions on only one recombination event. Therefore, we cannot present conclusive evidence that this resistance is a different locus to the previously reported R Pi-ber. The difference in position of marker TG63 could be attributed to false positive/negative marker or phenotypic scoring which can make a large impact when dealing with a small population and a small number of markers. An alternative explanation could be that marker TG63 is located inside (and thus flanked by) a large cluster of R pi -gene homologues. The use of one accession or another accession with different functional members of the cluster could lead to alternative mapping orders. R Pi-ber, originating from S. berthaultii, was reported as a new R pi -gene by Ewing et al. (2000) and Rauscher et al. (2006). According to information on internet by Dr. J. Jones, there are two R pi -genes located on chromosome 10 of S. berthaultii named R Pi-ber1 and R Pi-ber2 (http://www.ayeaye.tsl.ac.uk/index.php?option=com_content&task=view&id=98&Itemid=59). They are both roughly in the same position as the previously identified R Pi-ber (Ewing et al. 2000; Rauscher et al. 2006). R Pi-ber1 may be the same gene as R Pi-ber. Because we were uncertain if the S. berthaultii resistance from our accession CGN 17823/PI265858) was the same locus as the previously identified R Pi-ber, we confirmed its map position using data from a field experiment. The data of this field experiment also confirmed that the resistance conferred by R Pi-ber is not absolute, but can be described as strong in effect.
The objective of this research was to pyramid two functional R pi -genes originating from wild potato species in a S. tuberosum background and to compare the resistance level of those pyramided R pi -genes with the level of the single R pi -genes. We combined R Pi-mcd1 with R Pi-ber. Both R pi -genes confer incomplete resistance to P. infestans but greatly differ in their effect. The R Pi-mcd1 gene offers a small but significant reduction of the infection, lasting only for a short period while the R Pi-ber resistance gene offers a strong effect. It took 36 days until 50% of the average leaf area was infected in plants that carry the R Pi-ber resistance gene. In the field trial, the plants with the R Pi-ber resistance gene did not confer immunity during the entire growth cycle. The failure of the resistance by the end of the growing cycle is an annually observed phenomenon, known from other trials and years using this genetic material (data not shown). This is best explained by the physiological aging of the plant. Pyramiding of R Pi-mcd1 and R Pi-ber resulted in a higher level of resistance. This is not an obvious conclusion that should have been expected before starting this experiment. In view of the large effect of the R Pi-ber gene until late in the growing season, and in view of the short delay of infection at the beginning of the infection period we could also have expected that effect of the R Pi-ber gene would entirely surpass the modest effect of the R Pi-mcd1 gene. The resistance level in the pyramid group showed an additive effect of R Pi-mcd1 with R Pi-ber without clear evidence for a ‘law of diminishing returns’.
In our experiment, we used a large population of full sibs. This sibling structure would on average nullify the interaction between a specific genetic background and the R pi -genes. One example of such an interaction is the influence of maturity type on resistance level, irrespective of R pi -gene composition. This experimental design proved to be effective, because it allowed us to compensate for maturity effects. In our experiment, the magnitude of the effect caused by the individual R pi -genes was slightly obscured by variation in maturity. When resistance levels were corrected for maturity type, the difference between pyramided and the single RPi-ber resistance levels stayed significant for a longer period of time: 9 days instead of 3 days.
The pyramiding of R pi -genes is a valid strategy if the anticipated benefits exceed the breeding efforts. It is debatable if an infection delay of a couple of days is valuable. In our pyramided group, the resistance level of R Pi-ber was complemented by the resistance level of R Pi-mcd1. R Pi-mcd1 is known to be effective for a limited period. Therefore, improvement of resistance levels would be more economic when using R pi -genes with stronger effects. Irrespective of the complexities of natural infection with late blight, our findings indicate that combining the R pi -genes R Pi-mcd1 and R Pi-ber in potato is useful. In view of the additivity of the effects of the R Pi-mcd1 and R Pi-ber genes, and the lack of evidence for the ‘law of diminishing returns’, we recommend the development and analysis of potato clones with additionally pyramided R pi -genes to investigate the hypothesis that resistance levels comparable to non-host resistance might be feasible (Heath 2001; Jeuken and Lindhout 2002).
More important than economic considerations, the strategy of pyramiding R pi -genes is valued because of epidemiological and evolutionary considerations. Besides raising the level of resistance, pyramiding R pi -genes might contribute to the durability of resistance. The pathogen would need double or multiple mutations to overcome the resistance. The durability of an R pi -gene can be perceived as the evolutionary potential (mutation and recombination) of the plant pathogen to avoid R pi -gene recognition. In that case R pi -gene durability is based on the evolution of effector molecules and the indispensability of their current function. But when discussing durability of single or pyramided R pi -genes, one should be aware that durability can not be predicted in advance, but can only be reflected upon by historical evaluation. To ensure a maximum lifespan of an R pi -gene though, it is important that single R pi -gene and pyramided plants are not deployed simultaneously. The single-gene plants would in that case provide a “stepping-stone” for pathogens to overcome each of the pyramided resistance genes (Zhao et al. 2003).
Cultivars with pyramided R pi -genes can be developed by traditional and time consuming breeding or with genetic modification. Genetic modification is assumed to be a fast way to insert two or more R pi -genes into an existing variety. A huge benefit is that undesirable linkage drag from the wild potato genome, causing for example an elevated glycoalkaloid content, is prevented. Most of the R-genes against P. infestans originate from crossable wild potato species. Therefore, the GM product should not be considered as transgenic but as cisgenic (Jacobsen and Schouten 2007). As long as public acceptance for genetic modification is lacking in the EU, classical introgression breeding is required. One positive aspect of the use of diploid progenitor lines is the high frequency of gene transfer of these genes to the tetraploid potato via 4X–2X crosses. On average 80% of the heterozygous R pi -gene in diploids will result in tetraploid offspring with a simplex R pi -gene due to First Division Restitution of the unreduced male-gametes (Hermsen 1984). Molecular markers can facilitate the selection of superior descendants by tracing the R pi -genes in the breeding process and removing linkage drag efficiently.
In this paper, we have tested the hypothesis whether or not pyramiding is a valid strategy. There was no a priori expectation on the added value of the weaker R Pi-mcd1 locus in the presence of the stronger R Pi-ber locus. Our results suggest that pyramiding can result in an additive effect of the individual genes on the level of resistance. In our opinion, potato breeding can benefit from combining individual R pi -genes.
References
Allefs JJHM, Muskens MWM, van der Vossen EAG (2005) Breeding for foliage late blight resistance in the genomics era. In: Haverkort AJ, Struik PC (eds) Potato in progress science meets practice. Wageningen Academic Publishers, Wageningen, pp 255–267
Barloy D, Lemoine J, Abelard P, Tanguy AM, Rivoal R, Jahier J (2007) Marker-assisted pyramiding of two cereal cyst nematode resistance genes from Aegilops variabilis in wheat. Mol Breed 20:31–40
Bormann CA, Rickert AM, Ruiz RAC, Paal J, Lubeck J, Strahwald J, Buhr K, Gebhardt C (2004) Tagging quantitative trait loci for maturity-corrected late blight resistance in tetraploid potato with PCR-based candidate gene markers. Mol Plant Microbe Interact 17:1126–1138
Brodie BB, Plaisted RL (1992) Effect of H1 gene dosage on the development of Globodera rostochiensis in potato. J Nematol 24:584
Colon LT, Turkensteen LJ, Prummel W, Budding DJ, Hoogendoorn J (1995) Durable resistance to late blight (Phytophthora-Infestans) in old potato cultivars. Eur J Plant Pathol 101:387–397
Ewing EE, Šimko I, Smart CD, Bonierbale MW, Mizubuti ESG, May GD, Fry WE (2000) Genetic mapping from field tests of qualitative and quantitative resistance to Phytophthora infestans in a population derived from Solanum tuberosum and Solanum berthaultii. Mol Breed 6:25–36
Heath MC (2001) Non-host resistance to plant pathogens: nonspecific defense or the result of specific recognition events? Physiol Mol Plant Pathol 58:53–54
Hermsen JGT (1966) Crossability, fertility and cytogenetic studies in Solanum acaule × Solanum bulbocastanum. Euphytica 15:149–155
Hermsen JG (1984) Mechanisms and genetic implication of 2n gamete formation. Iowa State J Res 58:421–434
Huang N, Angeles ER, Domingo J, Magpantay G, Singh S, Zhang G, Kumaravadivel N, Bennett J, Khush GS (1997) Pyramiding of bacterial blight resistance genes in rice: marker-assisted selection using RFLP and PCR. Theor Appl Genet 95:313–320
Jacobsen E, Schouten HJ (2007) Cisgenesis strongly improves introgression breeding and induced translocation breeding of plants. Trends Biotechnol 25:219–223
Jeuken M, Lindhout P (2002) Lactuca saligna, a non-host for lettuce downy mildew (Bremia lactucae), harbors a new race-specific Dm gene and three QTLs for resistance. Theor Appl Genet 105:384–391
Landeo JA, Gastelo M, Pinedo H, Flores F (1995) Breeding for horizontal resistance to late blight in potato free of R-genes. In: Dowley LJ, Bannon E, Cooke LR, Keane T, O’Sullivan E (eds) Phytophthora infestans 150. Boole Press, Dublin, pp 268–274
Liu Z, Halterman D (2006) Identification and characterization of RB-orthologous genes from the late blight resistant wild potato species Solanum verrucosum. Phys Mol Plant Pathol 69(4–6):230–239
Nelson RR (1972) Stabilizing racial populations of plant pathogens by use of resistance genes. J Environ Qual 1:220–227
Pan QL, Liu YS, Budai-Hadrian O, Sela M, Carmel-Goren L, Zamir D, Fluhr R (2001) Comparative genetics of nucleotide binding site-leucine rich repeat resistance gene homologues in the genomes of two dicotyledons: tomato and arabidopsis (vol 155, pg 309, 2001). Genetics 159:1867
Park T-H, Vleeshouwers VGAA, Hutten RCB, van Eck HJ, van der Vossen E, Jacobsen E, Visser RGF (2005a) High-resolution mapping and analysis of the resistance locus Rpi-abpt against Phytophthora infestans in potato. Mol Breed 16:33
Park TH, Gros J, Sikkema A, Vleeshouwers VGAA, Muskens M, Allefs S, Jacobsen E, Visser RGF, van der Vossen EAG (2005b) The late blight resistance locus Rpi-blb3 from Solanum bulbocastanum belongs to a major late blight R gene cluster on chromosome 4 of potato. Mol Plant Microbe Interact 18:722–729
Rauscher GM, Smart CD, Simko I, Bonierbale M, Mayton H, Greenland A, Fry WE (2006) Characterization and mapping of Rpi-ber, a novel potato late blight resistance gene from Solanum berthaultii. Theor Appl Genet 112:674–687
Rauscher GM, Mayton H, Bonierbale M, Simko I, Smart C, Grunwald NJ, Fry W (2008) Effect of isolate, environment and a defeated R-gene (RPi-ber) on quantitative resistance of potato to late blight. American Phytopathological Society Annual Meeting, Minneapolis, Minnesota, July 26–30. Phytopathology 98:S130
Richardson KL, Vales MI, Kling JG, Mundt CC, Hayes PM (2006) Pyramiding and dissecting disease resistance QTL to barley stripe rust. Theor Appl Genet 113:485–495
Ross H, Baerecke ML (1951) Über die Bedeutung der argentinischen Solanum-Arten simplicifolium, vernei, berthaultii, acaule une einiger Formen von S. andigenum für die Züchtung Krankheitsresistenter Kartoffeln. Zeitschrift für Pflanzenzüchtung 30:280–291
Rozen S, Skaletsky H (2000) Primer3 on the WWW for general users and for biologist programmers. In: Krawetz S, Misener S (eds) Bioinformatics Methods and Protocols: Methods in Molecular Biology. Humana Press, Totowa, pp 365–386
Salaman RN (1910) Inheritance of colour and other characters in the potato. J Genet 1:7–46
Sharma PN, Torii A, Takumi S, Mori N, Nakamura C (2004) Marker-assisted pyramiding of brown planthopper (Nilaparvata lugens Stal) resistance genes Bph1 and Bph2 on rice chromosome 12. Hereditas 140:61–69
Simko I (2002) Comparative analysis of quantitative trait loci for foliage resistance to Phytophthora infestans in tuber-bearing Solanum species. Am J Potato Res 79:125–132
Singh S, Sidhu JS, Huang N, Vikal Y, Li Z, Brar DS, Dhaliwal HS, Khush GS (2001) Pyramiding three bacterial blight resistance genes (Xa5, Xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor Appl Genet 102:1011–1015
Song JQ, Bradeen JM, Naess SK, Raasch JA, Wielgus SM, Haberlach GT, Liu J, Kuang HH, Austin-Phillips S, Buell CR, Helgeson JP, Jiang JM (2003) Gene RB cloned from Solanum bulbocastanum confers broad spectrum resistance to potato late blight. Proc Natl Acad Sci USA 100:9128–9133
Stewart HE, Bradshaw JE, Pande B (2003) The effect of the presence of R-genes for resistance to late blight (Phytophthora infestans) of potato (Solanum tuberosum) on the underlying level of field resistance. Plant Pathol 52:193–198
Tan MYA, Hutten RCB, Celis C, Park T-H, Niks RE, Visser RGF, van Eck HJ (2008) The Rpi-mcd1 locus from Solanum microdontum involved in resistance to Phytophthora infestans, causing a delay in infection, maps on potato Chromosome 4 in a cluster of NBS-LRR genes. Mol Plant Microbe Interact 21:909–918
Toxopeus HJ (1957) On the influence of extra R-genes on the resistance of the potato to the corresponding P-races of Phytophthora infestans. Euphytica 6:106
Turkensteen LJ (1993) Durable resistance of potatoes against Phytophthora infestans. In: Th Jacobs, Parlevliet JE (eds) Durability of disease resistance. Kluwer, Dordrecht, pp 115–124
van der Beek JG, Verkerk R, Zabel P, Lindhout P (1992) Mapping strategy for resistance genes in tomato based on RFLPs between cultivars: Cf9 (resistance to Cladosporium fulvum) on chromosome 1. Theor Appl Genet 84:106–112
van der Vossen E, Sikkema A, Hekkert BTL, Gros J, Stevens P, Muskens M, Wouters D, Pereira A, Stiekema W, Allefs S (2003) An ancient R gene from the wild potato species Solanum bulbocastanum confers broad-spectrum resistance to Phytophthora infestans in cultivated potato and tomato. Plant J 36:867–882
van der Vossen EAG, Gros J, Sikkema A, Muskens M, Wouters D, Wolters P, Pereira A, Allefs S (2005) The Rpi-blb2 gene from Solanum bulbocastanum is an Mi-1 gene homolog conferring broad-spectrum late blight resistance in potato. Plant J 44:208–222
van Os H, Stam P, Visser RG, van Eck HJ (2005) RECORD: a novel method for ordering loci on a genetic linkage map. Theor Appl Genet 112:30–40
Van Soest LJM, Schöber B, Tazelaar MF (1984) Resistance to Phytophthora infestans in tuber-bearing species of Solanum and its geographical distribution. Potato Res 27:393–411
Visker M (2005) Association between late blight resistance and foliage maturity type in potato; physiological and genetic studies. Ph.D. Thesis, Wageningen University. ISBN: 90-8504-149-x http://www.library.wur.nl/wda/dissertations/dis3709.pdf
Visker MHPW, Keizer LCP, Van Eck HJ, Jacobsen E, Colon LT, Struik PC (2003) Can the QTL for late blight resistance on potato chromosome 5 be attributed to foliage maturity type? Theor Appl Genet 106:317–325
Visker M, van Raaij HMG, Keizer LCP, Struik PC, Colon LT (2004) Correlation between late blight resistance and foliage maturity type in potato. Euphytica 137:311–323
Vleeshouwers VGAA, van Dooijeweert W, Keizer LCP, Sijpkes L, Govers F, Colon LT (1999) A laboratory assay for Phytophthora infestans resistance in various Solanum species reflects the field situation. Eur J Plant Pathol 105:241–250
Vleeshouwers VG, Rietman H, Krenek P, Champouret N, Young C, Oh SK, Wang M, Bouwmeester K, Vosman B, Visser RG, Jacobsen E, Govers F, Kamoun S, van der Vossen EAG (2008) Effector genomics accelerates discovery and functional profiling of potato disease resistance and Phytophthora infestans avirulence genes. PLoS ONE 3:e2875
Yoshimura S, Yoshimura A, Iwata N, Mccouch SR, Abenes ML, Baraoidan MR, Mew TW, Nelson RJ (1995) Tagging and combining bacterial-blight resistance genes in rice using RAPD and RFLP markers. Mol Breed 1:375–387
Zhang J, Li X, Jiang G, Xu Y, He Y (2006) Pyramiding of Xa7 and Xa21 for the improvement of disease resistance to bacterial blight in hybrid rice. Plant Breed 125:600–605
Zhao JZ, Cao J, Li YX, Collins HL, Roush RT, Earle ED, Shelton AM (2003) Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat Biotechnol 21:1493–1497
Acknowledgments
We gratefully acknowledge René Alles for technical assistance and Björn d’Hoop for critically reading the manuscript.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Bryan.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Tan, M.Y.A., Hutten, R.C.B., Visser, R.G.F. et al. The effect of pyramiding Phytophthora infestans resistance genes R Pi-mcd1 and R Pi-ber in potato. Theor Appl Genet 121, 117–125 (2010). https://doi.org/10.1007/s00122-010-1295-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-010-1295-8