
Abstract
Global plastic production has increased exponentially since manufacturing commenced in the 1950’s, including polymer types infused with diverse additives and fillers. While the negative impacts of plastics are widely reported, particularly on marine vertebrates, impacts on microbial life remain poorly understood. Plastics impact microbiomes directly, exerting toxic effects, providing supplemental carbon sources and acting as rafts for microbial colonisation and dispersal. Indirect consequences include increased environmental shading, altered compositions of host communities and disruption of host organism or community health, hormone balances and immune responses. The isolation and application of plastic-degrading microbes are of substantial interest yet little evidence supports the microbial biodegradation of most high molecular weight synthetic polymers. Over 400 microbial species have been presumptively identified as capable of plastic degradation, but evidence for the degradation of highly prevalent polymers including polypropylene, nylon, polystyrene and polyvinyl chloride must be treated with caution; most studies fail to differentiate losses caused by the leaching or degradation of polymer monomers, additives or fillers. Even where polymer degradation is demonstrated, such as for polyethylene terephthalate, the ability of microorganisms to degrade more highly crystalline forms of the polymer used in commercial plastics appears limited. Microbiomes frequently work in conjunction with abiotic factors such as heat and light to impact the structural integrity of polymers and accessibility to enzymatic attack. Consequently, there remains much scope for extremophile microbiomes to be explored as a source of plastic-degrading enzymes and microorganisms. We propose a best-practice workflow for isolating and reporting plastic-degrading taxa from diverse environmental microbiomes, which should include multiple lines of evidence supporting changes in polymer structure, mass loss, and detection of presumed degradation products, along with confirmation of microbial strains and enzymes (and their associated genes) responsible for high molecular weight plastic polymer degradation. Such approaches are necessary for enzymatic degraders of high molecular weight plastic polymers to be differentiated from organisms only capable of degrading the more labile carbon within predominantly amorphous plastics, plastic monomers, additives or fillers.
Similar content being viewed by others


Global plastic pollution
The first plastic to be produced in commercial quantities, Bakelite, was invented in the early 1900s. A scarcity of resources and a need to enhance technologies following the First World War drove the development of new and improved synthetic materials, including plastics. Plastics now constitute a large and diverse group of materials made from combinations of synthetic and semi-synthetic polymer materials, frequently incorporating additives which aid the manufacture and performance of the final product, such as plasticisers, antioxidants and flame retardants [1]. Plastics are predominantly derived from fossil fuels (e.g. oil or natural gas), although they may also be made from renewable resources (e.g. ‘bio-based’ plastics derived from corn starch or sugar beet); plastics such as polyethylene terephthalate (PET) may be synthesized from either source and are sometimes referred to as ‘drop-in’ plastics. With the onset of mass consumerism in the 1960s and a move away from the use of traditional natural materials to more versatile plastics, plastics are now an integral part of our everyday lives. Plastic production has increased exponentially since the 1950s, with an estimated 8300 million metric tonnes of virgin plastic being produced to date and an expected annual production rate of 1100 t by 2050 [2].
Despite the large variety of polymers available, just eight make up 95% of all primary plastics ever made, with polypropylene and polyethylene comprising 45% of global production [2]. The primary use of plastic is for packaging (36%), followed by use in building and construction (16%) [3]. Currently, the dominant polymer types are entirely fossil-fuel based and are not biodegradable in a timescale relevant for their end-of-life management. Fossil-fuel based biodegradable polymers such as polycaprolactone (PCL) and polybutylene adipate terephthalate (PBAT) are not currently used at large scale. In fact, less than 1% of polymers are bio-based, and of those 44.5% are ‘drop-in’ polymers which share the same properties of their fossil fuel-based versions, i.e., they are considered non-degradable [4]. Of the almost 360 million tonnes of plastic produced annually, only a small fraction (~ 1%) is bio-based [4].
At their end-of-life, there are essentially three fates for plastics: recycling; incineration and discarding. To date, end-of-life management of plastic products has not kept pace with rapid increases in production, resulting in widespread environmental contamination. Globally, it is estimated that only 10% of plastics are recycled and 14% incinerated; the remaining 76% goes to landfills or enters the natural environment [2]. Recent modelling estimates that under current rates of loss, with no changes to management practices and in conjunction with the anticipated increase in production, 710 million tonnes of plastic waste will have cumulatively entered the environment by 2040 [5]. Whilst large plastic waste normally comes to mind when discussing leakage to the environment, the natural wear and tear of items, such as ropes, clothing and tyres, sheds small fragments during use, facilitating the passive transport of smaller plastic fragments into the environment. These fragments, when less than 5 mm are referred to as microplastics, or nanoplastics if less than 1 μm [6]. Microplastic leakage is expected to increase by 1.3 – 2.5 times by 2040 under a business-as-usual scenario and equates to approximately 3 million trillion pieces [5]. This widespread ingress of plastics into the environment means they are distributed across the globe in many different forms and in all ecosystems so far investigated; from rivers and streams [7, 8] to deep ocean trenches [9, 10], mountain tops [11], and from the tropics [12] to the poles [13].
Microbial impacts of global plastic pollution
The recent death of a Cuvier’s beaked whale in the Philippines with 40 kg of plastic waste in its stomach [14] and the necropsy of a young sperm whale on a Scottish beach yielding 100 kg of refuse [15] caught global media attention and scientists continue to report impacts of plastic waste on a wide range of species [16,17,18]. More than 800 animal species are already shown to have been affected by plastic pollution, and with an increasing number, from detritivorous sea snails [19] to apex marine predators [20, 21], being found to have internalised plastics. Globally, Wilcox, et al. [22] predict that as many as 90% of all seabirds ingest plastics. Post-mortem images of plastics spilling from the guts of dissected marine animals are causing us to reconsider unsustainable plastic use, yet the impacts of plastic pollution on most smaller organisms remain less well studied. Certainly, negative consequences of plastics have been reported for meiofauna such as Daphnia magna [23] and Caenorhabditis elegans nematodes [24], largely attributed to toxicological impacts, or blockage of the digestive system and related reductions in feeding rates. In contrast, the impact of plastics on environmental communities of microorganisms is rather less well researched.
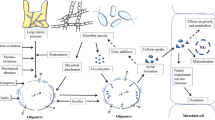
The term ‘microbiome’ describes the combined genetic material, or community, of microorganisms inhabiting a particular environment. While researchers continue to explore diverse microbiomes, including of soil, marine, freshwater, atmosphere and subsurface environments, the term ‘microbiome’ is perhaps predominently used to describe research into the microbiome of the gastrointestinal tract (the so-called ‘gut microbiome’). Since environmental plastics can concentrate in the digestive tracts of organisms from diverse trophic levels [25,26,27] they have the potential to impact the gut microbiome. However, due to their widespread environmental distribution, impacts of plastic pollution further extend to the microbiomes of diverse, non-host associated environments (which hereafter we refer to as the ‘environmental microbiome’). The direct impact of plastics on gut and environmental microbiomes are multiple (Fig. 1). (i) Some plastics and/or their associated additives provide organic carbon sources metabolizable by certain microorganisms. However, the microbial degradation of most plastics is restricted to only a few taxa [28], remains slow, and in many cases is unproven or disputed. Indeed, there remains a paucity of evidence for the microbial degradation of dominant plastic polymers, including polypropylene, polystyrene, polyethylene, nylon and polyvinyl chloride [29]. For these reasons, the impacts of plastics on microbial communities as a source of additional carbon are likely to be minimal, particularly in natural environments where alternative labile carbon and energy sources dominate. A notable exception to this may be following plastic consumption by certain insects where microbial degradation is postulated to be enhanced via ‘prior-processing’ by enzymes present within the gut [30]; this hypothesis however remains unproven. (ii) To a large degree, pure plastic polymers are chemically benign, having little toxic impact. However, industrial plastics contain additives including flame retardants (e.g., polychlorinated biphenyls and polychlorinated naphthalenes), plasticisers (e.g., bisphenol A) and UV stabilisers (e.g., benzotriazoles), some of which are demonstrated to impact microbial community composition and functioning. For example, plastic leachates from high-density polyethylene (HDPE) and polyvinylchloride (PVC) exert toxic effects on Prochlorococcus spp., impairing cell growth and population density in a dose-dependent manner [31]. Prochlorococcus is among the most numerous of photosynthetic organisms on Earth [32], responsible for perhaps ~ 10% of ocean net primary production [33]; in this regard, plastic pollution has demonstrated potential to impact major global microbial processes. Consumption of plasticisers including bisphenol A [34] may similarly cause dysbiosis of the gut microbiome, impacting host health. (iii) Plastics may also change microbial communities by impacting rates and extents of dispersal, since they provide a surface for microbial attachment and thereby can aid the transport of microbial cells, including pathogens, both around the globe and into the gut. In comparison to these direct impacts of plastics on microbiomes, far less is understood about their indirect impacts. Plastics and their additives can impact the health of host organisms with consequences for the gut microbiota that is intrinsic to the wellbeing of higher animals [35].

Schematic highlighting the diversity of direct and indirect impacts of plastics for gut and environmental microbiome communities and possible microbial solutions for the remediation of plastic waste
In this review, we highlight recent knowledge on the direct and indirect impacts of plastics on the health and functioning of environmental microbiomes, including of the gut. We further consider how the impacts of plastics may be mitigated and also manipulated to enhance both rates and extents of plastic degradation.
Impacts of plastics on the gut microbiome
Plastics cause a variety of undesirable mechanical, chemical and biological impacts on the species that ingest them. The consumption of plastics, either directly or via trophic level transfer [25], has multiple direct consequences, reducing appetite, impacting feeding activity and decreasing body weight [36], fitness [37] and fecundity [38]. In severe cases, the accumulation of large plastic masses may block the gastrointestinal tract; this has been recorded as a cause of death in diverse species including cetaceans [39, 40], turtles [41] and birds [42]. Smaller fractions of plastic may also bioaccumulate in the body, mostly in the gut, although translocation of plastics via the haemolymph and haemocytes of filter feeders is reported [26, 43], including to organs such as the liver and kidneys [44, 45]; this implies an ability for microplastics to cross the gut epithelial lining following ingestion and enter the circulatory system. Avio, et al. [43] explored the impact of polyethylene and polystyrene microplastics on the Mediterranean mussel (Mytilus galloprovincialis). Following 7 days of exposure to the plastic, histological analysis revealed aggregates of plastic in the intestinal lumen, epithelium and tubules. Further, increased DNA strand breakages provide evidence of genotoxic impacts, possibly caused by the greater production of reactive oxygen species (ROS) in response to microplastics. Nucleotide-binding oligomerization domain-like, or NOD-like receptor signalling pathways were enriched in M. galloprovincialis exposed to microplastics; these receptors recognise pathogenic factors entering the cell via phagocytosis and activate inflammatory responses. These findings support a growing body of evidence that micro- and nanoplastics cross biological barriers to promote immune and inflammatory responses [45, 46]. Where microplastics impact host immunity, this can further cause changes in gut microbial community composition and functioning. Oxidative states caused by inflammation can encourage the dominance of more resistant bacterial groups and, if associated with a rise in anaerobic respiratory terminal electron acceptors, may support the growth of anaerobic taxa such as members of the Enterobacteriaceae [47]. The gut microbiome influences not only the host immune system, but also contributes to digestion and the provision of essential nutrients [48], the degradation of harmful substances [49] and pathogen control within the gut [50]. The consumption and translocation of microplastics among bodily tissues therefore has far reaching consequences for the homeostasis normally maintained between a host and its microbiome.
While the physical presence of plastics demonstrably impacts the microbiome-gut-immune axis, additives which leach from plastic polymers have further consequences. Plasticisers are the largest group of plastic additives [51], particularly phthalates which may concentrate in bodily tissues to induce multiple adverse effects. For example, diethyl-hexyl phthalate (DEHP) causes antiestrogenic properties in fish hindering the development of reproductive organs [52], presumably due to competition with endogenous oestrogens for the receptor, and dibutyl phthalates delay gonad development and functioning in mammals [53] and amphibians [54]. The presence of bisphenol A (BPA) in the environment is predominantly due to it being a constitutive monomer of polycarbonate plastics, although it is also commonly added to PVC as a plasticiser. BPA has feminising impacts in fish, reducing male sperm quality, delaying and inhibiting ovulation in females [55] and in cases of high-concentration exposure, can induce intersex states [56]. Impacts on many other organisms are reported; BPA influences thyroid functioning and larval development in amphibians [57], early embryo development in marine bivalves [58] and reproductive birthweights and altered oestrous cyclicity in mammals [59, 60]. Plastics also adsorb organic pollutants such as polychlorinated biphenyl (PCB) from their environment [61, 62]; these contaminants may be transferred to the biological tissues of organisms such as birds following plastic ingestion [51]. While concentrations of plastic-associated contaminants are unlikely to be a major contributor to environmental concentrations of contaminants such as PCBs [63], a variety of plastic-associated compounds must be considered when assessing the impacts of plastic pollution on host-microbiome interactions [64].
The impacts of plastic additives on the gut microbiome remains little explored, although Adamovsky, et al. [65] assessed the consequences of environmentally relevant concentrations of the widely used plasticiser DEHP [66] on zebrafish. DEHP caused dysbiosis of the gut microbiota [67], and assessment of the gastrointestinal transcriptome revealed the up-regulation of T cells thought to play key roles in pathogen neutralisation by maintaining the integrity of the intestinal epithelia, while downregulating neuropeptide Y, a hormone which can modify immune activity by regulating T cell function. Analysis of the gut microbiome implicated several microbial metabolites that may contribute to immune and intercellular communication, including decreased L-glutamine in males and D-fructose 6-phosphate in females. Following DEHP exposure, Adamovsky, et al. [65] thereby identified the impact of microbial bioactive metabolites on host immune system dysregulation. Further negative impacts are reported. For example, the abundance of Mogibacteriaceae, Sutterella spp. and Clostridiales bacteria is increased within female mice exposed to BPA [68], presumably due to disrupted regulation of the sex hormones testosterone and oestrogen, implicating BPA for causing sex-dependent changes in the gut microbiome. The exposure of animals to plasticisers and plastic precursors including BPA are confirmed to impact intestinal microbial profiles in multiple studies [69,70,71], sometimes favouring microbial markers of dysbiosis such as a community dominance by Proteobacteria [72]. Nevertheless, understanding of cause and effect in host-microbiome interactions remains limited.
As we will later describe, microplastics are potential vectors of pests and pathogens around the globe via ocean currents, but so too may they vector pathogens into the gut. Microbial attachment to plastic particles can enhance both microbial dispersal and survival, as biofilms offer protection from environmental stress and enhanced opportunities for the sharing of beneficial traits via horizontal gene transfer. Pathogens such as Vibrio parahaemolyticus, which causes septicaemia and gastroenteritis in humans, have been identified in marine plastic-associated biofilm communities [73] and ingestion of such organisms hitchhiking on plastics might cause disease. However, even if not pathogenic, ingested organisms can influence gut community composition if they are capable of competing for resources within the gut [74]. Although the rich taxonomic and functional diversity of ‘plastisphere’ microbial communities has recently been unveiled [75], the role of plastics for microbial dispersal and colonisation of the gut remains poorly studied and understood.
Impacts of plastics on the environmental microbiome
In terrestrial environments, the mere presence of plastics exerts physical impacts directly impacting microbial communities. For example, agricultural plastic mulch films applied to enhance short-term crop productivity cover perhaps ~ 20 million hectares of farmland worldwide [76] and are a significant source of terrestrial plastic contamination [77]. While most research has focused on the impact of synthetic plastic films, the microbial consumption of biodegradable plastics is noted to have profound impacts on soil microbial communities [78]. Once embedded in the soil, plastics impact soil-water interactions by increasing water content [79], a major determinant of soil microbial community composition and functioning [80, 81]. By altering the availability of water, the physical impact of plastics on the soil environmental microbiome may be substantial [82]; the consequence of other physical impacts, such as increased shading by plastics which has been hypothesised to reduce aquatic photosynthesis, remain largely unsupported [83, 84].
The presence of plastic has direct chemical consequences for environmental microbial communities. Readily biodegradable plastics such as polylactic acid (PLA) contribute available carbon and in some cases significantly increase microbial biomass and enzyme activity [85]. The presence of such plastics in soils alter community composition, enriching the abundance and activity of certain taxa (e.g., members of the Ascomycota fungi [86]). The impact of more recalcitrant plastics remains less well understood, although even where degradation is slow, plasticising agents and additives such as phthalate acid esters may nevertheless leach, reaching elevated concentrations within receiving environments [87] and cause significant shifts in microbial community composition, abundance and enzyme activity [88, 89]. Although plastic additives are not always observed to impact environmental microbiomes at environmentally relevant concentrations [90], the sheer diversity of plastic additives used [91] means their impacts are yet to be fully understood. Of particular interest, Tetu, et al. [31] investigated the consequences of plastic leachate from HDPE bags and PVC matting on marine Prochlorococcus and confirmed that exposure to even the lowest dilution (approximately 1.6 g L− 1 and 0.125 g L− 1, respectively) of HDPE and PVC from 5-day old leachate impaired Prochlorococcus growth. Further, the transcription of genes associated with primary production was highly impacted, indicating that exposure to leachate from common plastic items has the capacity to impair the photosynthesis of the most dominant marine organisms.
Through the ubiquitous interactions between microorganisms and macroscopic plants and animals [92, 93], plastics and their associated compounds exert multiple indirect biological impacts on environmental microbiomes. For example, plants can be impacted as they take up plastics such as polystyrene via their roots, altering root length, weight and oxidative stress responses, possibly by the disruption of cell wall pores and cell-to-cell connections used for nutrient transport [94, 95]. Plant taxonomy and health play an important role in shaping soil and rhizosphere microbiomes, impacting the quantity and quality of root exudates [96] and the potential of plants to recruit specific members of the soil microbiome and promote the expression of genes, including those required for chemotaxis and biofilm formation [97]. Where observed, the impacts of plastics on the composition and health of plant and animal communities will likely have significant influences on environmental microbiomes, but to date insufficient evidence exists to suggest a strong link. Impacts on macroorganisms are rarely detected at environmentally relevant concentrations of microplastic; Judy, et al. [98] found no evidence of any impact of microplastics on wheat seedling emergence and production, or on the mortality or behaviour of earthworm and nematode populations.
While much research has focused on the impacts of plastics on microbial communities in situ, environmental plastics also influence rates and extents of microbial dispersal among environments. Buoyant plastics such as polyethylene, polypropylene and polystyrene, are transported over long distances by winds and oceanic currents [99] whereas non-buoyant plastics such as PET and PLA may act as a vector to transport surface-associated microbes to deeper water [100]. Microbial groups, including toxic microalgae [101] and potential human [75] and animal pathogens [102] have been detected associated with marine and freshwater plastics [73, 103] along with diverse antibiotic-resistant taxa [104]. Plastics are further postulated to vector pathogens through wastewater treatment plants [105] and pest species via ballast water [106]. Microbial communities colonising environmental plastics likely aid larval settlement and colonisation by species including bryozoans and polychaete worms, thereby assisting the movement of invasive marine macroorganisms around the globe [107]. Thus, in addition to supporting or retarding the growth of certain taxa, environmental plastics likely play significant roles in the dispersal of both microbes and higher organisms across diverse spatial scales and habitat types. Interestingly, the microbial colonisation of plastics can also impact particle buoyancy and transport [108, 109].
Assessing diverse plastisphere communities via amplicon and metagenome DNA sequencing
The development of molecular methods, including high-throughput DNA sequencing technology, is increasing our knowledge of the diverse nature of plastic-associated microbiomes. Although no taxa are known to only, or even to predominantly colonise plastic surfaces, multiple studies have demonstrated how the microbiomes of plastic debris differ from those present in the surrounding environment [110,111,112,113], with an overrepresentation in the plastisphere of bacterial phyla such as the Proteobacteria, Bacteriodetes [114] and Cyanobacteria [115] and fungi such as Chytridiomycota [113]. Nevertheless, with studies on the community composition of plastisphere microbiomes still in their infancy, it remains unclear the extent to which a core plastisphere community exists and the degree to which this differs from comparable microbiome communities in the same environment.
The specificity of plastisphere communities has been investigated in comparison to communities growing on inert surfaces such as glass and ceramic with varying results. A study by Oberbeckmann, et al. [116] using 16S rRNA gene amplicon sequencing for taxonomic analysis found no significant difference between the pelagic microbial communities associated with PET plastic bottles and glass microscope slides (as a control) deployed for 5-6 weeks. Pinto, et al. [117] also found that the overall community assembly on glass was similar among biofilms developing on HDPE, LDPE and PP over a period of up to 2 months, with families such as Flavobacteriaceae, Phyllobacteriaceae, Planctomycetaceae and Rhodobacteraceae being highly abundant across all surfaces. Such findings (also see Dang, et al. [118]) lead us to assume that there may be no specific plastic-associated communities. However, despite finding no differences in the total composition of communities growing on glass, HDPE, LDPE and PP (noting that significant differences were however observed for communities on PVC), Pinto, et al. [117] identified a subset of these communities incubated after immersion into seawater for up to 2 months, which was nonetheless responsive to the characteristics of individual plastic polymers or their additives (also see Ogonowski, et al. [119] and Kelly, et al. [7]). A higher relative abundance of the bacterial family Rhodobacteraceae discriminated communities growing on HDPE and Sphingomonadaceae for communities growing on LDPE, as compared to glass. Using a longer period of incubation, Kirstein et al. [120] found that after 15 months in a natural seawater flow-through system, biofilms from HDPE, LDPE, PP, PS, PET, PLA, styrene-acrylonitryle (SAN), polyurethane prepolymer (PESTUR) and PVC were significantly different to communities formed on glass. While communities on PVC were noticeable for having a high abundance (> 5%) of the bacterial genus Flexithrix, differences in the abundances of other plastic-specific taxa were largely attributed to variation in the presence and abundance of less dominant OTUs, suggesting that rarer species form specific associations with certain plastic types [121]. Also supporting the notion that less dominant members of the community may respond more specifically to the presence of different plastics, Erni-Cassola, et al. [122] demonstrated that during two-day incubations, weathered LDPE was enriched with a distinct community (particularly members of Roseobacter-, Oleiphilus- and Aestuariibacter-like taxa) from untreated PE and glass. However, this distinction was not detectable after 9 days, suggesting that substrate-specific microbes present in the plastisphere are quickly masked as the community matured and putative plastic-specific taxa were outnumbered. Interestingly, while significant differences in microbial community composition are not consistently reported among communities developing on different plastics, different plastic colours have recently been implicated as a significant determinant of plastisphere microbial community structure and functional diversity [123].
To date, a majority of studies assessing the formation and development of plastisphere communities have been conducted in the laboratory using different types of plastic of various condition (e.g., from ‘virgin’ plastics specifically manufactured for a study [124] to post-consumer plastics such as discarded bags and PET bottles [116]). Considering the longevity of plastic debris in the environment, the relatively short lengths of most lab-based studies may not be enough to explore the full degradative potential of the plastisphere microbiome. Environmental plastics hosting mature plastisphere microbiomes provide an alternative way to investigate the many factors that can influence plastisphere formation, such as plastic composition, age and condition. However, characterisation of aged microplastics, which dominate the marine plastisphere in terms of abundance, is often restricted as the biomass recovered from environmental microplastics is frequently very low, limiting abilities to recover sufficient nucleic acids for sequence analysis. As a consequence, there remain many unanswered questions regarding the plastisphere of aged environmental microplastics in particular.
As our knowledge of microorganisms present in the plastisphere is growing, there are still important questions that remain unanswered. (i) Which microorganisms act as pioneer species when the plastic is first introduced into the environment, and do the priority effects of early colonisation affect the overall composition and metabolic potential of the microbial community later on? These questions are of particular importance since the enrichment of plastic-degrading organisms may predominantly occur during early stages of colonisation, before the labile substrates generated from weathering are depleted and these plastic-specific microbes are dominated by more generalist biofilm-dwelling taxa [122]. (ii) Does there exist a core global community of plastic-degrading taxa, or do they exhibit substantial geographic or habitat-specific biogeography? (iii) If core members of the plastisphere vary in abundance between plastic types and biofilm maturity, can the presence and abundance of certain microorganisms indicate the approximate type and age of plastic debris? Answers to these questions will assist our ability to identify plastic-specific microorganisms from different regions, biomes, on different plastics and at different stages of plastic aging and degradation. Additionally, such knowledge likely increases our ability to use microbial community DNA to inform on the environmental impact of plastics (for example by adopting the approach of Hermans, et al. [125]).
As highlighted by Wright, et al. [126], many studies have characterised the plastisphere through taxonomic analyses [112, 117, 121, 122], however, there remains a lack of knowledge surrounding the functional potential of these communities. Bryant et al. [115] were among the first to explore the metabolic potential of the plastisphere microbiome using shotgun metagenomics, hypothesising that the genomes of plastic-associated taxa would be more distinct and exhibit increased metabolic activity compared to free-living bacteria in the surrounding marine water. Compared to those of the picoplankton community, their study revealed an increased abundance of genes encoding for chemotaxis and nitrogen fixation as well as several putative genes for xenobiotic biodegradation in plastic-associated communities. This included a gene encoding for 2,4-dichlorophenol 6-monooxygenase, a hydroxylase associated with the degradation of chlorinated aromatic pollutants [127] sometimes produced from polymer and plastic additive pyrolysis [128]. Similarly, the study revealed an increased abundance of multiple genes encoding for ring-cleaving enzymes, such as protocatechuate 3,4-dioxygenase and particularly homogentisate 1,2-dioxygenase, previously linked with styrene and polycyclic aromatic hydrocarbon degradation [129]. Whilst Bryant, et al. [115] were unable to confirm if microbes within the plastisphere are able to degrade the plastic polymer, the increased abundance of genes encoding for the degradation of several xenobiotics may assist identification of new plastic-degrading enzymes, and also the taxa expressing and utilising these enzymes. In common with previous studies, Pinnell & Turner [130] found the community composition of fossil fuel-derived PET-associated biofilms to be indistinguishable from those growing on ceramic beads deployed at the sediment-water interface of a coastal lagoon; in contrast, microbial communities associated with bio-based PHA pellets were dominated by sulphate-reducing organisms. Metagenomic analysis of the bioplastic-associated communities revealed substantial phylogenetic diversification of one depolymerase in particular, polyhydroxybutyrate (PHB) depolymerase, alongside an almost 20-fold increase in abundance of the depolymerase genes, suggesting they are widely distributed within the biofilm. An increased abundance of genes associated with sulphate reduction and plastic degradation, such as depolymerases, esterases and sulphate reductases, were also reported. Thus, while bio-based plastics continue to be perceived as an environmentally friendly alternative, if sedimentary inputs are large enough, the authors speculate that microbial responses could impact benthic biogeochemical cycling through the stimulation of sulphate reducers.
It is likely that communities work together to access plastic-derived carbon; the genes encoding for the degradation of alkanes, for example, are distributed among diverse assemblages of hydrocarbonoclastic organisms [131]. A greater understanding of the dynamics of plastic-associating communities may be achieved by determining co-occurrence patterns and associations among different organisms and genes. Toxic and poorly labile carbon substrates have been observed to strongly favour facilitation among microbial species such that they can each grow and degrade these substrates better in order to survive [132]. Where taxa or gene products are presumed to play a beneficial role in plastic degradation, correlated increases in their abundance across multiple samples as indicated by network analysis (e.g. see Gatica, et al. [133]) might identify other organisms and molecular pathways that could benefit from the community response to plastic contaminants.
Mitigation of plastic pollution by the gut microbiome
Recently, several insect species (particularly the larvae of darkling beetles, wax moths and meal moths) have garnered interest for their ability to consume and degrade a diversity of plastic polymers. For example, larvae of the Indian meal moth Plodia interpunctella can ingest and appear capable of degrading polystyrene [134] as do larvae of yellow and giant mealworms Tenebrio molitor and Zophrobas morio, respectively [135, 136]. Larvae of the greater [137] and lesser wax moths (Galleria mellonella and Achroia grisella [138]) are similarly reported to degrade polyethylene and polystyrene, respectively. Isotope analysis provides evidence that carbon from plastics such as PE is incorporated into the biomass of invertebrates [139]. Despite the findings of these and other studies, it nevertheless remains uncertain the extent to which either the higher organism or its associated microbiome contribute toward plastic polymer degradation. Further, the extent to which these biodegradative processes may be accelerated by synergistic effects of the host-microbiome remains unclear (Fig. 2).

Evidence for a role for insects, host-associated microbes, or host-independent, free-living microbes in plastic degradation. Degradation of the plastic polymer may be detected by a variety of methods, including: [i] mass loss of plastic such as clear zone development around colonies on plastic-infused/overlaid agar, [ii] altered plastic surface properties (e.g., visible by scanning electron microscopy) and [iii] generation of degradation products (e.g., CO2, polymer metabolites detected by Fourier-transform infrared spectroscopy or high-performance liquid chromatography)
Many organisms consume plastic incidentally and gain no nutritional value from its consumption; plastic has been found in abundance within the guts of diverse organisms from seabirds [22] and fish [140] to marine and freshwater worms [36, 141] and zooplankton [142]. Although the ingestion of plastics by species including the common earthworm Lumbricus terrestris is associated with reductions in plastic size distribution [143], in many cases, demonstration of plastic degradation, e.g. by conversion to CO2 or incorporation of plastic-associated carbon into animal biomass, is unsubstantiated [144]. Similarly, the ‘consumption’ of plastics by mealworms and wax moth larvae has gained much attention [30, 145], but confirmation of plastic degradation by the hosts’ gut-derived enzymes, independent of the hosts’ microbiome, requires further confirmation [146]. In most cases, it remains to be seen whether the host derives any nutritional benefits from plastic as a source of energy; without stronger evidence of more complete degradation in the gut, plastic fragments may merely be generated via mechanical processes (e.g. chewing) and ejected into the environment. To confirm plastic degradation by macroinvertebrates, studies in germ-free organisms (i.e., those lacking a microbiome) are desirable, noting the physiological homeostasis of organisms such as T. molitor are impacted by related changes in digestive enzyme expression by axenic cultures [147]. Another approach is to track the fate of radiolabelled (e.g. 13C, 14C) plastic polymer via incorporation into the cellular biomass or respiration products of consumer invertebrates [139], preferably in the absence of host microbial taxa to also eliminate the possibility of trophic carbon transfer. The lack of evidence to date for plastic degradation by germ-free larvae instead supports that microbiota are important drivers of plastic degradation within the invertebrate gut.
Since diverse putative plastic-degrading microbial taxa have now been described, including isolates from gut microbiota [28], it is hypothesised that the enzymes of gut-associated microbial taxa, rather than the enzymes of the host per se, perform most, if not all, plastic degradation by plastic-consuming invertebrate taxa. In a series of experiments, Cassone, et al. [148] provide multiple lines of evidence for the degradation of LDPE by the intact microbiome of G. mellonella larvae. The larvae of G. mellonella readily consume beeswax, which in some aspects is similar to plastics such as PE, being comprised of a diverse mixture of long-chain hydrocarbons. Hence, plastic consumption propensity may be related to the structural or chemical similarity of plastics to their preferred food source. PE-fed caterpillars had a far greater abundance of gut-associated microorganisms as compared to starved individuals, or even to organisms fed a natural diet of honeycomb, suggesting their microbiota could benefit from the abundance of PE in the gut. Antibiotic-treated caterpillars fed PE also excreted only half the concentration of ethylene glycol compared to untreated animals. Since ethylene glycol is a putative by-product of PE metabolism [30] this was used to imply a direct role of the gut microbiome for PE degradation. The inhibition of plastic depolymerisation following antibiotic treatment has now been observed in numerous studies, indicating that the host organism alone is poorly able to utilise plastic as a carbon or energy source, or is at least in part reliant on its microbiome as a source of plastic-degrading enzymes [135, 136, 144, 148]. Providing further evidence for a microbial role in plastic degradation, Cassone, et al. [148] isolated and grew bacteria from the gut (identified as Acinetobacter sp.) on carbon-free media, supplemented with PE fragments. A further observation was that the Acinetobacter sp. was only capable of degrading plastics at a very slow rate when isolated from the gut, providing evidence that plastic degradation is maximised by synergisms occurring between the host and its gut microbiome community, although the importance of community microbial interactions cannot be disregarded. Nevertheless, the extent to which the larvae impact the structure of the plastic polymer or associated additives, or enhances beneficial functional attributes of its gut microbiota currently remains unclear.
Prior to the study of Cassone, et al. [148], multiple authors had already isolated putative plastic-degrading bacteria from the insect gut microbiome. Yang, et al. [144] isolated the bacterium Exiguobacterium sp. Strain YT2 from the gut of styrofoam-fed mealworms and demonstrated its ability to grow on polystyrene film as a sole carbon source, associated with changes in the surface topography and hydrophobicity of the plastic. Mass loss of polystyrene combined with decreases in molecular weight and the release of water-soluble degradation products were used as further evidence to highlight the capacity for gut-associated microbes to degrade plastics (noting that Danso, et al. [29] question if sufficient evidence is available to confirm degradation of the high-molecular weight polymer, i.e. the polystyrene itself, rather than styrene monomers incorporated within the polymer matrix). Similar studies implicate Aspergillus flavus, Bacillus sp. YP1 and Enterobacter asburiae YT1 isolated from insect gut microbiomes as being capable of PE degradation [134, 149]. While such findings identify a possible role for gut-associated microbes to degrade plastic, organisms isolated from non-host environments are similarly capable of plastic degradation and could be exploited for their biodegradation capacity.
Mitigation of plastic pollution by the environmental microbiome
The first evidence that free-living environmental taxa contribute to plastic degradation was only published circa 30 years after the first commercial plastic production, in 1974, when Fields, et al. [150] showed that the fungus Aureobasidium pullulans was capable of PCL degradation. Since then, the number of microorganisms suggested as capable of plastic biodegradation has increased considerably. A recent study by Gambarini, et al. [28] reports over 400 publications describing the degradation of 72 different plastic types by 436 species of fungi and bacteria. Presumptive plastic-degrading microbes identified to date belong to five bacterial and three fungal phyla. Among the bacterial phyla, Proteobacteria (n = 133), Actinobacteria (n = 88), and Firmicutes (n = 60) have the greatest number of reported species, while Bacteroidetes (n = 3) and Cyanobacteria (n = 2) have far fewer. The fungal phyla include Ascomycota (n = 118), Basidiomycota (n = 19), and Mucoromycota (n = 13) (Fig. 3).

Number of putative plastic-degrading organisms reported by Gambarini, et al. [28], classified at the level of phylum level. The number following the phylum name represents the number of species from that specific phylum that are reported as plastic-degraders
As outlined earlier, a small number of plastic-degrading microbes have been isolated from plant- and animal-associated microbiomes [149, 151, 152]. However, most isolates reported in the literature were derived from soil [153, 154] or from waste processing sites such as composting facilities [155] and landfills [156]. An additional source comprises bacteria and fungi already deposited in culture collections [157]. All major synthetic polymers have species reported to degrade them, for instance PE [158, 159], PET [160, 161], PP [162], PS [163], PU [164] and PVC [165]. However, the strength of evidence for degradation varies by plastic type. To date, PET biodegradation has been studied the most comprehensively. A notable example includes the PET-degrading bacterium, Ideonella sakaiensis, isolated from sediment in the vicinity of a Japanese bottle recycling plant [161]. I. sakaiensis is the first organism for which the degradation of PET was well-described and the enzymatic degradation of PET elucidated, characterised [166] and enhanced [167]. Conversely, there is only weak evidence for the biodegradation of synthetic polymers such as nylon, PP, PS and PVC. For instance, nylon-oligomer biodegradation by the bacterium Agromyces sp. KY5R has been shown by Yasuhira, et al. [168] and the genes and corresponding enzymes responsible for the biodegradation activity have been identified; however, biodegradation of the plastic polymer (i.e. not just monomers and oligomers) is yet to be confirmed.
Bioprospecting for novel mechanisms of plastic degradation
Currently, there is a lack of information necessary to critically validate many reports of plastic degradation by microbial taxa or communities or to accurately reproduce the research. For instance, many reports provide no information regarding polymer composition and omit details of fillers and additives that may be present in polymer composites. Therefore, it is frequently not possible to differentiate between the microbial degradation of plastic polymers or their additives. The strength of the degradation evidence is also greatly dependent on the techniques applied, which can be divided into three main categories, those detecting: (i) changes in the polymer structure, (ii) physical loss of plastic mass and (iii) the generation of plastic metabolites. The strongest evidence of plastic biodegradation is likely achieved using a combination of techniques from all three categories. However, analysis of the dataset of Gambarini, et al. [28], which compiled data from 408 studies, revealed that of the microorganisms reported to degrade plastics, 48% of reports were based on assays relating to only one of these categories, 39% used techniques that covered two categories, and just 10% used techniques that covered all three (Fig. 4).

Percentage of studies using evidence for plastic degradation by microbial species based on: (i) changes in polymer structure (blue), (ii) physical loss of plastic mass (red), or (iii) detection of plastic metabolites (green), or these techniques in combination. Data were compiled using the
Most reports of plastic degradation by microbial isolates do not go on to explore the genes and enzymes responsible for the reported activity. In fact, only around 14% of the microorganisms reported to degrade plastic have the gene sequences conferring the degradation activity elucidated [28]. This represents a major shortcoming since knowledge of the relevant biochemical and molecular data provides the capability to advance the plastic biodegradation field enormously, allowing the search for new putative plastic-degrading genes in novel microbiomes by comparison to enzyme data banked in structural and molecular databases. Crucial information and procedures related to the reported plastic degraders are frequently missing or incomplete in the current literature, for example, the location and conditions of isolation of the plastic-degrading isolate, strength of evidence for degradation, accurate taxonomic classification, and a lack of deposited strains in culture banks. By not addressing these points adequately, reports of plastic degradation, possibly in a majority of studies undertaken to date, must be treated with caution.
To exploit the broad phenotypic diversity that may already be present in natural populations, future advances in plastic biodegradation will likely benefit from isolation of novel microorganisms from diverse microbiome communities. This calls for consideration of the sampling environment and likely growth requirements of organisms within the microbiome, the plastic type of interest and the empirical tests required to delineate growth-linked biodegradation of the polymer. By reviewing the current literature, we provide a ‘best practice’ workflow of methods necessary to describe the pathways of growth-linked plastic biodegradation, beginning with appropriately characterising the plastisphere microbiome and concluding with the identification of plastic biodegradation genes and pathways (Table 1).
Based on protein mutagenic and structural analysis studies [166], alongside homology database searches [28], it is likely that certain microorganisms already possess plastic degradation genes but do not express them in situ, and/or derive energy from more readily utilisable carbon sources when available. By incorporating inert controls (e.g., glass or ceramic surfaces), we may be able to distinguish between genes acquired and expressed for the process of plastic-degradation, from those normally expressed in biofilm communities (i.e. including where plastic is not present). Yoshida et al., [161] demonstrated that I. sakaiensis possesses two genes encoding enzymes which degrade PET (IsPETase and IsMHETase). However, they did not address if the IsPETase might be used by the organism for other functions, or whether it was being used in situ to degrade PET within the PET recycling plant from which the organism was originally isolated. Structural analyses of the IsPETase revealed that the enzyme has a wider active-site cleft compared to ancestral cutinase homologs [166]. Narrowing the active-site cleft via mutation of active-site amino acids improved crystalline PET degradation, indicating that the IsPETase was not fully optimised for PET metabolism. This, in conjunction with the initial isolations focusing on amorphous PET (1.9% crystalline) instead of the more crystalline PET abundant in bottle recycling plants (15.7% crystalline; Yoshida et al. [161]) suggests that the origin of the first I. sakaiensis isolate from a recycling plant might be coincidental.
Mere changes in polymer mechanical properties and physical structure, even when observed in concert with microbial biomass production, are insufficient evidence to confirm polymer biomineralisation by microbial isolates [172]. Physical losses of plastic mass should also be reported. Plastics can be incorporated into growth media as plastic films, powders or granules, and emulsifications. The first two approaches are primarily used to identify physical changes in polymer structure and the accumulation of biomass as first lines of evidence for plastic degradation (Table 1; Fig. 4). Evidence of polymer degradation from plastic films or polymer granules predominantly requires changes in polymer roughness, the formation of holes or cracks, fragmentation or color changes, confirmed using visual methods such as scanning electron microscopy (SEM), Fourier-transform infrared spectroscopy (FTIR) [173] or atomic force microscopy [174]. However, visual changes in surface structure, changes in plastic mass and mechanical properties do not provide direct evidence of biodegradation [175] because these physical changes cannot be distinguished from abiotic degradation. Where biodegradation is demonstrated it is likely that microbiomes work in conjunction with abiotic factors to impact the structural integrity of polymers [176]. Most polymers are too large to transverse the cell membranes and must be initially depolymerised (e.g. by heat, visible and non-visible spectrum light and oxygen) [177]. Additionally, measuring changes in the surface structure or molecular weight of plastics does not discriminate between the degradation of polymers or their additives [172]. Therefore, in addition to plastic film and granule-infused media, we recommended that biomass accumulation on plastic surfaces and changes to polymer structure should be accompanied by the detection of plastic metabolites to describe growth-linked biodegradation.
A common method for assessing microbial plastic metabolism is by observing clear zones in agar containing emulsified plastic [175, 178]. However, emulsifications are usually limited to amorphous or lower molecular weight plastics while environmental waste plastics such as nylon, PE and PET typically have a higher molecular weight, limiting the analysis of these pollutant plastics. In addition, solvents and surfactants widely used to form plastic emulsions are themselves documented to be degraded by microorganisms [179, 180]. Therefore, observation of clearance zones in culture media containing plastic emulsions should ideally be associated with other empirical tests, such as observations of incorporation of radiolabeled carbon from the polymer backbone into microbial biomass. Because plastic typically comprises the predominant or only carbon source in plastic metabolism assays, only small amounts of evolved CO2 are typically required to be detected to indicate polymer metabolism [175]. In addition to CO2, other plastic metabolites hypothesised to be produced during plastic degradation (e.g. the production of mono-(2-hydroxyethyl) terephthalate during PET hydrolysis) may be identified using methods such as liquid/light Chromatography-Mass Spectrometry, which detects multiple compounds in a single analytical run [181]. This approach was employed to implicate the role of a putative depolymerase in PHB degradation by Aspergillus fumigatus [182]. Similarly, HPLC-mediated detection of the PET-degradation metabolites MHET and terephthalate provided evidence for IsPETase involvement in PET degradation [183]. These methods, combined with approaches employed to detect changes in polymer structure and metabolism (Fig. 4) provide powerful evidence for confirming plastic biodegradation.
Knowledge of genes known to be associated with plastic degradation provides a strong tool to identify new degraders and genes among microbiome communities. For instance, Danso, et al. [29] developed a hidden Markov model (HMM) to search genome and metagenome databases for the presence of potential PET hydrolases. The authors used the sequences from nine different enzymes with verified activity on PET-based substrates and identified 504 possible PET hydrolase candidate genes. Studies such as this, and the work of Gambarini, et al. [28], indicates a huge potential for mining molecular databases for plastic degradation-conferring genes (PDGs). One useful approach to verify PDGs experimentally is by heterologous expression of the microbiome-derived candidate genes in a host that lacks degradation capacity in the absence of the introduced gene, followed by confirmation of the plastic-degrading phenotype of the transformant. Heterologous expression in hosts such as Escherichia coli has been used to verify plastic degradation-conferring phenotypes of PDGs encoding putative PHB-depolymerases, esterases, cutinases, carboxylesterase and PET hydrolases from a wide variety of bacteria, and some fungi [29, 184,185,186]. Overexpression in heterologous hosts is also a valuable tool for purifying high levels of enzyme for in vitro assays or studying enzyme crystal structure. Another approach is to disrupt or silence the candidate PDGs in the endogenous background and assess the effect this has on the plastic degradation phenotype. Mining metagenomes using the candidate gene approach does not inform on the discovery of completely novel determinants, or accessory factors that have not been previously described. Under this scenario, genotype-phenotype-based studies of individual degrading strains are still important to identify novel determinants, using methods such as DNA library screens in heterologous hosts, random mutagenesis or differential transcript expression. However, once PDGs are identified, interrogating metagenomes of closely related species for conserved alleles can inform on important residues and functional domains to exploit for genetic enhancement of plastic degradation traits.
Manipulating microbiomes to enhance rates and extents of plastic degradation
Different strategies may be employed to overcome the challenges of isolating microorganisms capable of efficient and/or fast plastic degradation. For example, higher temperatures can increase the flexibility of both amorphous [187, 188] and crystalline domains of the polymer chain [189,190,191], thereby improving their accessibility to enzymatic attack [188]. In this regard, thermophile microbiomes represent a promising source of enzymes because they will likely be more thermostable. In one study, the most thermostable enzyme tested (a leaf-branch compost cutinase (LCC) obtained from an uncultured bacterium [186]) had the highest PET depolymerization rates at 65 °C [192]. Degradation rates were further increased after improving enzyme thermostability through site-specific mutagenesis. To date however, only ~ 10% of isolated plastic degradation studies report polymer degradation at temperatures ≥50 °C and only a small fraction (~ 0.5%) of these have been isolated from extreme environments such as hot springs, composts and anaerobic digesters [28]. There would appear to be significant scope for mining thermophile and extremophile microbiomes as a promising source of putative plastic degrading enzymes and microorganisms.
The higher genotypic and phenotypic diversity present in microbial communities compared with single microbial strains may mean that communities are more efficient degraders of xenobiotic pollutants [193]. As such, artificial consortia created by selecting a small number of plastic degrading microorganisms within an already existing consortium (i.e., using a top-down approach [194]), or combining separately isolated microbial strains (i.e., using a bottom-up approach [162]) may be a useful strategy for improving plastic biodegradation. Alternatively, directed mutagenesis to improve gene expression and enzyme function, along with metabolic engineering and synthetic biology tools, could be exploited to obtain more efficient plastic-degrading consortia. Specifically, the introduction or modification of interspecific microbial interactions (such as intercellular communication via metabolite exchange) could be used to create consortia with improved biodegradation traits [195, 196]. Additionally, the segmentation of metabolic pathways among strains such that each organism produces an intermediate compound that can be used by the next organism in the pathway can be used to reduce the metabolic burden on any one organism. Because only limited information is available regarding genes and enzymes involved in plastic biodegradation [28], an improved understanding of degradation pathways by single strains and multi-strain co-degradation pathways is first required to facilitate this approach.
Conclusions
The impacts of global plastic pollution on microbiomes are diverse, ranging from the direct consequences of toxic leachates on microbial community health and activity to the indirect effects of plastics on host organisms and environments. Many hundreds of microbial species, genes and enzymes are implicated in plastic degradation. For a small number of particularly bio-based plastics, such as PLA, clear evidence is presented for their microbial degradation. However, for the majority of commercial plastics, evidence for microbial degradation remains weak, with studies failing to confirm microbial growth on the synthetic polymer. To ensure the correct identification of plastic-degrading taxa and enzymes, facilitating their improvement by environmental, biotic and genetic manipulation, multiple lines of evidence for plastic degradation should be presented. Ideally this will include evidence of changes in the polymer structure, mass loss and detection of degradation products, along with confirmation of the microbial strain and putative plastic-degrading enzymes and associated genes. Such details are essential for organisms and enzymes capable of plastic degradation to be reliably differentiated from those only capable of degrading the more labile carbon within predominantly amorphous plastics, plastic monomers, fillers and additives.
Abbreviations
- CAM:
-
Contact angle measurement
- DRIFT spectroscopy:
-
Diffuse reflectance infrared Fourier transform spectroscopy
- DSC:
-
Differential scanning calorimetry
- EDS:
-
Energy dispersive spectroscopy
- EFM:
-
Epi-fluorescence microscopy
- FCM:
-
Flow cytometry
- FTIR:
-
Fourier-transform infrared spectroscopy
- GC-FID:
-
Gas chromatography with flame ionization detection
- GC-MS:
-
Gas chromatography-mass spectrometry
- GPC:
-
Gel permeation chromatography
- HDPE:
-
High-density polyethylene
- HPLC:
-
High-performance liquid chromatography
- IR spectroscopy:
-
Infrared spectroscopy
- LDPE:
-
Low-density polyethylene
- LLDPE:
-
Linear low-density polyethylene
- MALDI:
-
Matrix assisted laser desorption/ionization
- MHET:
-
Mono-2-hydroxyethyl terephthalate
- MHETase:
-
A hydrolase enzyme which cleaves MHET
- MS:
-
Mass spectrometry
- NMR:
-
Nuclear magnetic resonance
- PBAT:
-
Polybutylene adipate terephthalate
- PCL:
-
Polycaprolactone
- PE:
-
Polyethylene
- PEA:
-
Polyesteracetal
- PEG:
-
Polyethylene glycol
- PES:
-
Polyestersulfone
- PESU:
-
Polyethersulfone
- PET:
-
Polyethylene terephthalate
- PHA:
-
Polyhydroxyalkanoate
- PHB:
-
Polyhydroxybutyrate
- PHBV:
-
Poly (3-hydroxybutyrate-co-3-hydroxyvalerate)
- PHO:
-
Polyhydroxyoctanoate
- PLA:
-
Polylactic acid
- PMCL:
-
Poly (4-methyl-ε-caprolactone)
- PS:
-
Polystyrene
- PP:
-
Polypropylene
- PU:
-
Polyurethane
- PVA:
-
Polyvinyl alcohol
- PVC:
-
Polyvinylchloride
- QCM-D:
-
Quartz crystal microbalance with dissipation monitoring
- RDS:
-
Rheometrics dynamic spectrometer
- SEC:
-
Size-exclusion chromatography
- SEM:
-
Scanning electron microscopy
- TOC:
-
Total organic carbon
- TGA:
-
Thermogravimetric analysis
- TPA:
-
Terephthalic acid
- UPLC:
-
Ultra-performance liquid chromatography
- WAXS:
-
Wide angle X-ray scattering
- XPS:
-
X-ray photoelectron spectroscopy
- XRD:
-
X-ray powder diffraction
References
Lithner D, Larsson A, Dave G. Environmental and health hazard ranking and assessment of plastic polymers based on chemical composition. Sci Total Environ. 2011;409(18):3309–24.
Geyer R. Chapter 2 - Production, use, and fate of synthetic polymers. In: Letcher TM, editor. Plastic Waste and Recycling. Oxford: Academic Press; 2020:13–32.
Geyer R, Jambeck JR, Law KL. Production, use, and fate of all plastics ever made. Sci Adv. 2017;3(7):e1700782.
European Bioplastics. Bioplastics market data 2018. Berlin: European Bioplastics; 2018.
Lau WWY, Shiran Y, Bailey RM, Cook E, Stuchtey MR, Koskella J, et al. Evaluating scenarios toward zero plastic pollution. Science. 2020;369(6510):1455-61.
Toussaint B, Raffael B, Angers-Loustau A, Gilliland D, Kestens V, Petrillo M, et al. Review of micro- and nanoplastic contamination in the food chain. Food Addit Contam Part A. 2019;36(5):639–73.
Kelly JJ, London MG, Oforji N, Ogunsola A, Hoellein TJ. Microplastic selects for convergent microbiomes from distinct riverine sources. Freshwater Sci. 2020;39(2):281–91.
Hoellein TJ, McCormick AR, Hittie J, London MG, Scott JW, Kelly JJ. Longitudinal patterns of microplastic concentration and bacterial assemblages in surface and benthic habitats of an urban river. Freshwater Sci. 2017;36(3):491–507.
Chiba S, Saito H, Fletcher R, Yogi T, Kayo M, Miyagi S, et al. Human footprint in the abyss: 30 year records of deep-sea plastic debris. Mar Policy. 2018;96:204–12.
Van Cauwenberghe L, Vanreusel A, Mees J, Janssen CR. Microplastic pollution in deep-sea sediments. Environ Pollut. 2013;182:495–9.
Bergmann M, Mützel S, Primpke S, Tekman MB, Trachsel J, Gerdts G. White and wonderful? Microplastics prevail in snow from the Alps to the Arctic. Sci Adv. 2019;5(8):eaax1157.
Lamb JB, Willis BL, Fiorenza EA, Couch CS, Howard R, Rader DN, et al. Plastic waste associated with disease on coral reefs. Science. 2018;359(6374):460–2.
Laganà P, Caruso G, Corsi I, Bergami E, Venuti V, Majolino D, et al. Do plastics serve as a possible vector for the spread of antibiotic resistance? First insights from bacteria associated to a polystyrene piece from King George Island (Antarctica). Int J Hyg Environ Health. 2019;222(1):89–100.
Borunda A. This young whale died with 88 pounds of plastic in its stomach. Natl Geogr Mag. 2019 (March 18). https://www.nationalgeographic.com/environment/2019/03/whale-dies-88-pounds-plastic-philippines/#close.
Beachum L. Dead sperm whale had 220 pounds of garbage in its stomach, including rope, plastic and gloves. Wahington DC: The Washington Post; 2019.
Brentano R, Petry MV. Marine debris ingestion and human impacts on the pygmy sperm whale (Kogia breviceps) in southern Brazil. Mar Pollut Bull. 2020;150:5.
Moore RC, Loseto L, Noel M, Etemadifar A, Brewster JD, MacPhee S, et al. Microplastics in beluga whales (Delphinapterus leucas) from the eastern Beaufort Sea. Mar Pollut Bull. 2020;150:7.
Deudero S, Alomar C. Mediterranean marine biodiversity under threat: reviewing influence of marine litter on species. Mar Pollut Bull. 2015;98(1-2):58–68.
Li RL, Zhang SP, Zhang LL, Yu KF, Wang SP, Wang YH. Field study of the microplastic pollution in sea snails (Ellobium chinense) from mangrove forest and their relationships with microplastics in water/sediment located on the north of Beibu gulf. Environ Pollut. 2020;263:8.
Nelms SE, Parry HE, Bennett KA, Galloway TS, Godley BJ, Santillo D, et al. What goes in, must come out: combining scat-based molecular diet analysis and quantification of ingested microplastics in a marine top predator. Methods Ecol Evol. 2019;10(10):1712–22.
Santos RG, Andrades R, Demetrio GR, Kuwai GM, Sobral MF, Vieira JDS, et al. Exploring plastic-induced satiety in foraging green turtles. Environ Pollut. 2020;265(Pt B):114918.
Wilcox C, Van Sebille E, Hardesty BD. Threat of plastic pollution to seabirds is global, pervasive, and increasing. Proc Natl Acad Sci U S A. 2015;112(38):11899–904.
Jemec A, Horvat P, Kunej U, Bele M, Krzan A. Uptake and effects of microplastic textile fibers on freshwater crustacean Daphnia magna. Environ Pollut. 2016;219:201–9.
Lei L, Wu S, Lu S, Liu M, Song Y, Fu Z, et al. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci Total Environ. 2018;619-620:1–8.
Farrell P, Nelson K. Trophic level transfer of microplastic: Mytilus edulis (L.) to. Carcinus maenas (L.). Environ Pollut. 2013;177:1–3.
Browne MA, Dissanayake A, Galloway TS, Lowe DM, Thompson R. Ingested microscopic plastic translocates to the circulatory system of the mussel, Mytilus edulis (L). Environ Sci Technol. 2008;42(13):5026–31.
Nelms SE, Galloway TS, Godley BJ, Jarvis DS, Lindeque PK. Investigating microplastic trophic transfer in marine top predators. Environ Pollut. 2018;238:999–1007.
Gambarini V, Pantos O, Kingsbury JM, Weaver L, Handley KM, Lear G. An updated phylogenetic distribution fo plastic-degrading organisms and enzymes. mSystems. 2020;.
Danso D, Chow J, Streit WR. Plastics: environmental and biotechnological perspectives on microbial degradation. Appl Environ Microbiol. 2019;85(19):14.
Bombelli P, Howe CJ, Bertocchini F. Polyethylene bio-degradation by caterpillars of the wax moth galleria mellonella. Curr Biol. 2017;27:R283–R93.
Tetu SG, Sarker I, Schrameyer V, Pickford R, Elbourne LDH, Moore LR, et al. Plastic leacates impair growth and oxygen production in Prochlorococcus, the ocean’s most abundant photosynthetic bacteria. Commun Biol. 2019;2:article No.: 184.
Partensky F, Hess WR, Vaulot D. Prochlorococcus, a marine photosynthetic prokaryote of global significance. Microbiol Mol Biol Rev. 1999;63(1):106–27.
Flombaum P, Gallegos JL, Gordillo RA, Rincón J, Zabala LL, Jiao N, et al. Present and future global distributions of the marine cyanobacteria Prochlorococcus and Synechococcus. Proc Natl Acad Sci U S A. 2013;110(24):9824–9.
Feng D, Zhang H, Jiang X, Zou J, Li Q, Mai H, et al. Bisphenol A exposure induces gut microbiota dysbiosis and consequent activation of gut-liver axis leading to hepatic steatosis in CD-1 mice. Environ Pollut. 2020;265(Pt A):114880.
Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009;9(5):313–23.
Wright SL, Rowe D, Thompson RC, Galloway TS. Microplastic ingestion decreases energy reserves in marine worms. Curr Biol. 2013;23(23):R1031–R3.
Besseling E, Wegner A, Foekema EM, van den Heuvel-Greve MJ, Koelmans AA. Effects of microplastic on fitness and PCB bioaccumulation by the lugworm Arenicola marina (L.). Environ Sci Technol. 2013;47(1):593–600.
Lee K-W, Shim WJ, Kwon OY, Kang J. Size-dependent effects of micro polystyrene particles in the marine copepod Tigriopus japonicus. Environ Sci Technol. 2013;47:11278–83.
Jacobsen JK, Massey L, Gulland F. Fatal ingestion of floating debris by two sperm whales (Physeter macrocephalus). Mar Pollut Bull. 2010;60:765–7.
Secci ER, Zarzur S. Plastic debris ingested by a Blainville’s beaked whale, Mesoplodon densirotris, washed ashore in Brazil. Aquat Mamm. 1999;25:21–4.
Wilcox C, Puckridge M, Schuyler QA, Townsend K, Hardesty BD. A quantitative analysis linking sea turtle mortality and plastic debris ingestion. Sci Rep. 2018;8:12536.
Ryan PG, Moore CJ, van Franeker JA, Moloney CL. Monitoring the abundance of plastic debris in the marine environment. Philos Trans Phys Sci Eng. 2009;364:1999–2012.
Avio CG, Gorbi S, Milan M, Benedetti M, Fattorini D, d’Errico G, et al. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ Pollut. 2015;198:211–22.
Collard F, Gilbert B, Compère P, Eppe G, Das K, Jauiaux T, et al. Microplastics in the livers of European anchovies (Engraulis encrasicolus, L.). Environ Pollut. 2017;229:1000–5.
Deng Y, Zhang Y, Lemos B, Ren H. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci Rep. 2017;7:46687.
von Moos N, Burkhardt-Holm P, Köhler A. Uptake and effects of microplastics on cells and tissue of the blue mussel Mytilus edulis L. after an experimental exposure. Environ Sci Technol. 2012;46(20):11327–35.
Winter SE, Bäumler AJ. Dysbiosis in the inflamed intestine: chance favours the prepared microbe. Gut Microbes. 2014;5:71–3.
Jing T-Z, Qi F-H, Wang Z-Y. Most dominant roles of insect gut bacteria: digestion, detoxification, or essential nutrient provision? Microbiome. 2020;8(1):38.
Velmurugan G, Ramprasath T, Swaminathan K, Mithieux G, Rajendhran J, Dhivakar M, et al. Gut microbial degradation of organophosphate insecticides-induces glucose intolerance via gluconeogenesis. Genome Biol. 2017;18(1):8.
Kamada N, Chen GY, Inohara N, Núñez G. Control of pathogens and pathobionts by the gut microbiota. Nat Immunol. 2013;14(7):685–90.
Teuten EL, Saquing JM, Knappe DRU, Barlaz MA, Jonsson S, Bjorn A, et al. Transport and release of chemicals from plastics to the environment and to wildlife. Philos Trans R Soc B-Biol Sci. 2009;364(1526):2027–45.
Kim E-J, Kim J-W, Lee S-K. Inhibition of oocyte development in Japanese medaka (Oryzias latipes) exposed to di-2-ethylhexyl phthalate. Environ Int. 2002;28(5):359–65.
Yin L, Yan LP, He B, Fang YJ, Liu XY, Duan CG, et al. The toxic effects of a plasticizer, dibutyl phthalate, on rat testis. Int J Clin Exp Pathol. 2016;9(11):11246–53.
Ohtani H, Miura I, Ichikawa Y. Effects of dibutyl phthalate as an environmental endocrine disruptor on gonadal sex differentiation of genetic males of the frog Rana rugosa. Environ Health Perspect. 2000;108(12):1189–93.
Lahnsteiner F, Berger B, Kletzl M, Weismann T. Effect of bisphenol a on maturation and quality of semen and eggs in the brown trout, Salmo trutta f. fario. Aquat Toxicol. 2005;75:213–24.
Mandich A, Bottero S, Benfenati E, Cevasco A, Erratico C, Maggioni S, et al. In vivo exposure of carp to graded concentrations of bisphenol a. Gen Comp Endocrinol. 2007;153(1):15–24.
Oehlmann J, Schulte-Oehlmann U, Kloas W, Jagnytsch O, Lutz I, Kusk KO, et al. A critical analysis of the biological impacts of plasticizers on wildlife. Philos Trans R Soc Lond Ser B Biol Sci. 2009;364(1526):2047–62.
Balbi T, Franzellitti S, Fabbri R, Montagna M, Fabbri E, Canesi L. Impact of bisphenol a (BPA) on early embryo development in the marine mussel Mytilus galloprovincialis: effects on gene transcription. Environ Pollut. 2016;218:996–1004.
Savabieasfahani M, Kannan K, Astapova O, Evans NP, Padmanabhan V. Developmental programming: differential effects of prenatal exposure to bisphenol-a or methoxychlor on reproductive function. Endocrinol. 2006;147:5956–66.
Rubin BS, Murray MK, Damassa DA, King JC, Soto AM. Perinatal exposure to low doses of bisphenol a affects body weight, patterns of estrous cyclicity, and plasma LH levels. Environ Health Perspect. 2001;109:675–80.
Rios LM, Jones PR, Moore C, Narayan UV. Quantitation of persistent organic pollutants adsorbed on plastic debris from the northern Pacific Gyre's "eastern garbage patch". J Environ Monit. 2010;12(12):2226–36.
Mato Y, Isobe T, Takada H, Kanehiro H, Ohtake C, Kaminuma T. Plastic resin pellets as a transport medium for toxic chemicals in the marine environment. Environ Sci Technol. 2001;35(2):318–24.
Ryan PG, Connell AD, Gardner BD. Plastic ingestion and PCBs in seabirds: is there a relationship? Mar Pollut Bull. 1988;19(4):174–6.
Li H-X, Getzinger GJ, Ferguson PL, Orihuela B, Zhu M, Rittschof D. Effects of toxic leachate from commercial plastics on larval survival and settlement of the barnacle Amphibalanus amphitrite. Environ Sci Technol. 2016;50(2):924–31.
Adamovsky O, Buerger AN, Vespalcova H, Sohag SR, Hanlon AT, Ginn PE, et al. Evaluation of microbiome-host relationships in the zebrafish gastrointenstinal system reveals adaptive immunity is a target of bis (2-ethylhexyl) phthalate (DEHP) exposure. Environ Sci Technol. 2020;54:5719–28.
Ceresana Market Research. Market Study: Plasticisers (5th Edition), vol. 2019. Germany: Ceresana Ltd; 2017. p. 350.
Chen LG, Guo YY, Hu CY, Lam PKS, Lam JCW, Zhou BS. Dysbiosis of gut microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol a: implications for host health in zebrafish. Environ Pollut. 2018;234:307–17.
Javurek AB, Spollen WG, Johnson SA, Bivens NJ, Bromert KH, Givan SA, et al. Effects of exposure to bisphenol a and ethinyl estradiol on the gut microbiota of parents and their offspring in a rodent model. Gut Microbes. 2016;7(6):471–85.
Lai K-P, Chung Y-T, Li R, Wan H-T, Wong CK-C. Bisphenol a alters gut microbiome: comparative metagenomics analysis. Environ Pollut. 2016;218:923–30.
Xu J, Huang GN, Nagy T, Teng Q, Guo TL. Sex-dependent effects of bisphenol a on type 1 diabetes development in non-obese diabetic (NOD) mice. Arch Toxicol. 2019;93(4):997–1008.
DeLuca JAA, Allred KF, Menon R, Riordan R, Weeks BR, Jayaraman A, et al. Bisphenol-a alters microbiota metabolites derived from aromatic amino acids and worsens disease activity during colitis. Exp Biol Med. 2018;243(10):864–75.
Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012;13:R79.
Kirstein IV, Kirmizi S, Wichels A, Garin-Fernandez A, Erler R, Löder M, et al. Dangerous hitchhikers? Evidence for potentially pathogenic Vibrio spp. on microplastic particles. Mar Environ Res. 2016;120:1–8.
Fackelmann G, Sommer S. Microplastics and the gut microbiome: how chronically exposed species may suffer from gut dysbiosis. Mar Pollut Bull. 2019;143:193–203.
Zettler ER, Mincer TJ, Amaral-Zettler LA. Life in the “plastisphere”: microbial communities on plastic marine debris. Environ Sci Technol. 2013;47(13):7137–46.
Steinmetz Z, Wollmann C, Schaefer M, Buchmann C, David J, Troeger J, et al. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation? Sci Total Environ. 2016;550:690–705.
Huang Y, Liu Q, Jia W, Yan C, Wang J. Agricultural plastic mulching as a source of microplastics in the terrestrial environment. Environ Pollut. 2020;260:114096.
Serrano-Ruiz H, Martin-Closas L, Pelacho AM. Biodegradable plastic mulches: impact on the agricultural biotic environment. Sci Total Environ. 2021;750:141228.
Machado AAD, Lau CW, Kloas W, Bergmann J, Bacheher JB, Faltin E, et al. Microplastics can change soil properties and affect plant performance. Environ Sci Technol. 2019;53(10):6044–52.
Drenovsky RE, Vo D, Graham KJ, Scow KM. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb Ecol. 2004;48(3):424–30.
Davidson EA, Belk E, Boone RD. Soil water Contant and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob Chang Biol. 1998;4(2):217–27.
Qi YL, Ossowicki A, Yang XM, Lwanga EH, Dini-Andreote F, Geissen V, et al. Effects of plastic mulch film residues on wheat rhizosphere and soil properties. J Hazard Mater. 2020;387:7.
Sjollema SB, Redondo-Hasselerharm P, Leslie HA, Kraak MHS, Vethaak AD. Do plastic particles affect microalgal photosynthesis and growth? Aquat Toxicol. 2016;170:259–61.
Besseling E, Wang B, Lürling M, Koelmans AA. Nanoplastic affects growth of S. obliquus and reproduction of D. magna. Environ Sci Technol. 2014;48:12336–43.
Li C, Moore-Kucera J, Lee J, Corbin A, Brodhagen M, Miles C, et al. Effects of biodegradable mulch on soil quality. Appl Soil Ecol. 2014;79:59–69.
Muroi F, Tachibana Y, Kobayashi Y, Sakurai T. Kasuya K-i. influences of poly (butylene adipate-co-terephthalate) on soil microbiota and plant growth. Poly Degrad Stab. 2016;129:338–46.
Kong S, Ji Y, Liu L, Chen L, Zhao X, Wang J, et al. Diversities of phthalate esters in suburban agricultural soils and wasteland soil appeared with urbanization in China. Environ Pollut. 2012;170:161–8.
Kong X, Jin D, Jin S, Wang Z, Yin H, Xu M, et al. Responses of bacterial community to dibutyl phthalate pollution in a soil-vegetable ecosystem. J Hazard Mater. 2018;353:142–50.
Xie H-J, Shi Y-J, Zhang J, Cui Y, Teng S-X, Wang S-G, et al. Degradation of phthalate esters (PAEs) in soil and the effects of PAEs on soil microcosm activity. J Chem Technol Biotechnol. 2010;85(8):1108–16.
Cartwright CD, Thompson IP, Burns RG. Degradation and impact of phthalate plasticizers on soil microbial communties. Environ Toxicol Chem. 2009;19(5):1253–61.
Hahladakis JN, Velis CA, Weber R, Iacovidou E, Purnell P. An overview of chemical additives present in plastics: migration, release, fate and environmental impact during their use, disposal and recycling. J Hazard Mater. 2018;344:179–99.
Herrera Paredes S, Lebeis SL. Giving back to the community: microbial mechanisms of plant–soil interactions. Front Ecol. 2016;30(7):1043–52.
Bray N, Wickings K. The roles of invertebrates in the urban soil microbiome. Front Ecol Evol. 2019;7:359.
Jiang X, Chen H, Liao Y, Ye Z, Li M, Klobučar G. Ecotoxicity and genotoxicity if polystyrene microplastics on higher plant Vicia faba. Environ Pollut. 2019;250:831–8.
Kalčíková G, Žgajnar Gotvajn A, Kladnik A, Jemec A. Impact of polyethylene microbeads on the floating freshwater plant duckweed lemma minor. Environ Pollut. 2017;230:1108–15.
Zhang C, Liu GB, Xue S, Song ZL. Rhizosphere soil microbial activity under different vegetation types on the loess plateau, China. Geoderma. 2011;161(3-4):115–25.
Pérez-Jaramillo JE, Mendes R, Raaijmakers JM. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol Biol. 2016;90:635–44.
Judy JD, Williams M, Gregg A, Oliver D, Kumar A, Kookana R, et al. Microplastics in municipal mixed-waste organic outputs induce minimal short to long-term toxicity in key terrestrial biota. Environ Pollut. 2019;252:522–31.
Lacerda ALDF, Rodrigues LDS, van Sebille E, Rodrigues FL, Ribeiro L, Secchi ER, et al. Plastics in sea surface waters around the Antarctic peninsula. Sci Rep. 2019;9:3977.
Audrézet F, Zaiko A, Lear G, Wood SA, Tremblay LA, Pochon X. Biosecurity implications of drifting marine plastic debris: current knowledge and future research. Mar Pollut Bull. 2020:111835. https://doi.org/10.1016/j.marpolbul.2020.111835.
Casabianca S, Capellacci S, Giacobbe MG, Dell'Aversano C, Tartaglione L, Varriale F, et al. Plastic-associated harmful microalgal assemblages in marine environment. Environ Pollut. 2019;244:617–26.
Goldstein MC, Carson HS, Eriksen M. Relationship of diversity and habitat area in North Pacific plastic-associated rafting communities. Mar Biol. 2014;161:1441–53.
McCormick A, Hoellein TJ, Mason SA, Schluep J, Kelly JJ. Microplastic is an abundant and distinct microbial habitat in an urban river. Environ Sci Technol. 2014;48:11863–71.
Moore RE, Millar BC, Moore JE. Antimicrobial resistance (AMR) and marine plastics: can food packaging litter act as a dispersal mechanism for AMR in oceanic environments? Mar Pollut Bull. 2020;150:7.
Bowley J, Baker-Austin C, Porter A, Hartnell R, Lewis C. Oceanic hitchhikers – Assessing pathogen risks from marine microplastic. Trends Microbiol. 2020; https://doi.org/10.1016/j.tim.2020.06.011.
Naik RK, Naik MM, D'Costa PM, Shaikh F. Microplastics in ballast water as an emerging source and vector for harmful chemicals, antibiotics, metals, bacterial pathogens and HAB species: a potential risk to the marine environment and human health. Mar Pollut Bull. 2019;149:110525.
Barnes DKA. Biodiversity - invasions by marine life on plastic debris. Nature. 2002;416:808–9.
Hoellein TJ, Shogren AJ, Tank JL, Risteca P, Kelly JJ. Microplastic deposition velocity in streams follows patterns for naturally occurring allochthonous particles. Sci Rep. 2019;9:3740.
Kooi M, van Nes EH, Scheffer M, Koelmans AA. Ups and downs in the ocean: effects of biofouling on vertical transport of microplastics. Environ Sci Technol. 2017;51(14):7963–71.
Debroas D, Mone A, Ter Halle A. Plastics in the North Atlantic garbage patch: A boat-microbe for hitchhikers and plastic degraders. Sci Total Environ. 2017;599-600:1222–32.
Oberbeckmann S, Kreikemeyer B, Labrenz M. Environmental factors support the formation of specific bacterial assemblages on microplastics. Front Microbiol. 2017;8:2709.
Dussud C, Meistertzheim AL, Conan P, Pujo-Pay M, George M, Fabre P, et al. Evidence of niche partitioning among bacteria living on plastics, organic particles and surrounding seawaters. Environ Pollut. 2018;236:807–16.
Kettner MT, Rojas-Jimenez K, Oberbeckmann S, Labrenz M, Grossart HP. Microplastics alter composition of fungal communities in aquatic ecosystems. Environ Microbiol. 2017;19(11):4447–59.
Oberbeckmann S, Loeder MGJ, Gerdts G, Osborn AM. Spatial and seasonal variation in diversity and structure of microbial biofilms on marine plastics in northern European waters. FEMS Microbiol Ecol. 2014;90(2):478–92.
Bryant JA, Clemente TM, Viviani DA, Fong AA, Thomas KA, Kemp P, et al. Diversity and activity of communities inhabiting plastic debris in the North Pacific Gyre. mSystems. 2016;1(3):e00024–16.
Oberbeckmann S, Osborn AM, Duhaime MB. Microbes on a bottle: substrate, season and geography influence community composition of microbes colonizing marine plastic debris. PLoS One. 2016;11(8):e0159289.
Pinto M, Langer TM, Huffer T, Hofmann T, Herndl GJ. The composition of bacterial communities associated with plastic biofilms differs between different polymers and stages of biofilm succession. PLoS One. 2019;14(6):e0217165.
Dang H, Li T, Chen M, Huang G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl Environ Microbiol. 2008;74(1):52–60.
Ogonowski M, Motiei A, Ininbergs K, Hell E, Gerdes Z, Udekwu KI, et al. Evidence for selective bacterial community structuring on microplastics. Environ Microbiol. 2018;20(8):2796–808.
Kirstein IV, Wichels A, Krohne G, Gerdts G. Mature biofilm communities on synthetic polymers in seawater - specific or general? Mar Environ Res. 2018;142:147–54.
Kirstein IV, Wichels A, Gullans E, Krohne G, Gerdts G. The Plastisphere - Uncovering tightly attached plastic "specific" microorganisms. PLoS One. 2019;14(4):e0215859–e.
Erni-Cassola G, Wright RJ, Gibson MI, Christie-Oleza JA. Early colonization of weathered polyethylene by distinct bacteria in marine coastal seawater. Microb Ecol. 2020;79(3):517–26.
Wen B, Liu J-H, Zhang Y, Zhang H-R, Gao J-Z, Chen Z-Z. Community structure and functional diversity of the plastisphere in aquaculture waters: does plastic color matter? Sci Total Environ. 2020;740:140082.
Harrison JP, Schratzberger M, Sapp M, Osborn AM. Rapid bacterial colonization of low-density polyethylene microplastics in coastal sediment microcosms. BMC Microbiol. 2014;14:232.
Hermans SM, Buckley HL, Case BS, Curran-Cournane F, Taylor M, Lear G. Using soil bacterial communities to predict physico-chemical variables and soil quality. Microbiome. 2020;8(1):79.
Wright RJ, Erni-Cassola G, Zadjelovic V, Latva M, Christie-Oleza JA. Marine plastic debris: a new surface for microbial colonization. Environ Sci Technol. 2020; 54(19):11657-72.
Ledger T, Pieper DH, Gonzalez B. Chlorophenol hydroxylases encoded by plasmid pJP4 differentially contribute to chlorophenoxyacetic acid degradation. Appl Environ Microbiol. 2006;72(4):2783–92.
Blankenship A, Chang DPY, Jones AD, Kelly PB, Kennedy IM, Matsumura F, et al. Toxic combustion by-products from the incineration of chlorinated hydrocarbons and plastics. Chemosphere. 1994;28(1):183–96.
Cao J, Lai Q, Yuan J, Shao Z. Genomic and metabolic analysis of fluoranthene degradation pathway in Celeribacter indicus P73T. Sci Rep. 2015;5:7741.
Pinnell LJ, Turner JW. Shotgun metagenomics reveals the benthic microbial community response to plastic and bioplastic in a coastal marine environment. Front Microbiol. 2019;1:1252.
Wang W, Shao Z. Enzymes and genes involved in aerobic alkane degradation. Front Microbiol. 2013;4:article 116.
Piccardi P, Vessman B, Mitri S. Toxicity drives facilitation between 4 bacterial species. Proc Natl Acad Sci U S A. 2019;116(32):15979–84.
Gatica J, Jurkevitch E, Cytryn E. Comparative metagenomics and network analyses provide novel insights into the scope and distribution of β-lactamase homologs in the environment. Front Microbiol. 2019;10:146.
Yang J, Yang Y, Wu WM, Zhao J, Jiang L. Evidence of polyethylene biodegradation by bacterial strains from the guts of plastic-eating waxworms. Environ Sci Technol. 2014;48(23):13776–84.
Peng BY, Su YM, Chen ZB, Chen JB, Zhou XF, Benbow ME, et al. Biodegradation of polystyrene by dark (Tenebrio obscurus) and yellow (Tenebrio molitor) mealworms (Coleoptera: Tenebrionidae). Environ Sci Technol. 2019;53(9):5256–65.
Yang Y, Wang JL, Xia ML. Biodegradation and mineralization of polystyrene by plastic-eating superworms Zophobas atratus. Sci Total Environ. 2020;708:7.
Lou Y, Ekaterina P, Yang SS, Lu BY, Liu BF, Ren NQ, et al. Biodegradation of polyethylene and polystyrene by greater wax moth larvae (galleria mellonella L.) and the effect of co-diet supplementation on the core gut microbiome. Environ Sci Technol. 2020;54(5):2821–31.
Kundungal H, Gangarapu M, Sarangapani S, Patchaiyappan A, Devipriya SP. Efficient biodegradation of polyethylene (HDPE) waste by the plastic-eating lesser waxworm (Achroia grisella). Environ Sci Pollut Res. 2019;26:18509–19.
Taipale SJ, Peltomaa E, Kukkonen JVK, Kainz MJ, Kautonen P, Tiirola M. Tracing the fate of microplastic carbon in the aquatic food web by compound-specific isotope analysis. Sci Rep. 2019;9:19894.
Lusher AL, McHugh M, Thompson RC. Occurrence of microplastics in the gastrointestinal tract of pelagic and demersal fish from the English Channel. Mar Pollut Bull. 2013;67(1-2):94–9.
Hurley RR, Woodward JC, Rothwell JJ. Ingestion of microplastics by freshwater tubifex worms. Environ Sci Technol. 2017;51(21):12844–51.
Cole M, Lindeque P, Fileman E, Halsband C, Goodhead R, Moger J. Microplastic ingestion by zooplankton. Environ Sci Technol. 2013;47(12):6646–55.
Huerta Lwanga E, Gertsen H, Gooren H, Peters P, Salánki T, van der Ploeg M, et al. Microplastics in the terrestial ecosystem: implications for Lumbricus terrestris (Oligochaeta, Lumbricidae). Environ Sci Technol. 2016;50:2685–91.
Yang Y, Yang J, Wu WM, Zhao J, Song YL, Gao LC, et al. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: part 2. Role of gut microorganisms. Environ Sci Technol. 2015;49(20):12087–93.
Brandon AM, El Abbadi SH, Ibekwe UA, Cho Y-M, Wu W-M, Criddle CS. Fate of hexabromocyclododecane (HBCD), a common flame retardant, in polystyrene-degrading mealworms: elevated HBCD levels in egested polymer but no bioaccumulation. Environ Sci Technol. 2020;54(1):364–71.
Kong HG, Kim HH, Chung JH, Jun J, Lee SB, Kim HM, et al. The galleria mellonella hologenome supports microbiota independent metabolism of long-chain hydrocarbon beeswax. Cell Rep. 2019;26:2451–64.
Vigneron A, Jehan C, Rigaud T, Moret Y. Immune defenses of a beneficial pest: The mealworm beetle, Tenebrio molitor. Front Physiol. 2019;10:138.
Cassone BJ, Grove HC, Elebute O, Villanueva SMP, LeMoine CMR. Role of the intestinal microbiome in low-density polyethylene degradation by caterpillar larvae of the greater wax moth, galleria mellonella. Proc R Soc B-Biol Sci. 2020;287(1922):9.
Zhang JQ, Gao DL, Li QH, Zhao YX, Li L, Lin HF, et al. Biodegradation of polyethylene microplastic particles by the fungus Aspergillus flavus from the guts of wax moth galleria mellonella. Sci Total Environ. 2020;704:8.
Fields RD, Rodriguez F, Finn RK. Microbial degradation of polyesters: Polycaprolactone degraded by P. pullulans. J Appl Polym Sci. 1974;18:3571–9.
Almeida EL, Carrillo Rincón AF, Jackson SA, Dobson ADW. In silico screening and heterologous expression of a polyethylene terephthalate hydrolase (PETase)-like enzyme (SM14est) with polycaprolactone (PCL)-degrading activity, from the marine sponge-derived strain Streptomyces sp. SM14. Front Microbiol. 2019;10:2187.
Koitabashi M, Noguchi MT, Sameshima-Yamashita Y, Hiradate S, Suzuki K, Yoshida S, et al. Degradation of biodegradable plastic mulch films in soil environment by phylloplane fungi isolated from gramineous plants. AMB Express. 2012;2(1):40.
Mergaert J, Webb A, Anderson C, Wouters A, Swings J. Microbial degradation of poly (3-hydroxybutyrate) and poly (3-hydroxybutyrate-co-3-hydroxyvalerate) in soils. Appl Environ Microbiol. 1993;59(10):3233–8.
Shahreza H, Sepahy AA, Hosseini F, Nejad RK. Molecular identification of Pseudomonas strains with polyethylene degradation ability from soil and cloning of alkB gene. Arch Pharm Pract. 2019;10:43–8.
Kleeberg I, Hetz C, Kroppenstedt RM, Müller RJ, Deckwer WD. Biodegradation of aliphatic-aromatic copolyesters by Thermomonospora fusca and other thermophilic compost isolates. Appl Environ Microbiol. 1998;64(5):1731–5.
Gajendiran A, Krishnamoorthy S, Abraham J. Microbial degradation of low-density polyethylene (LDPE) by Aspergillus clavatus strain JASK1 isolated from landfill soil. 3 Biotech. 2016;6(1):52.
da Luz JMR, Paes SA, Bazzolli DMS, Tótola MR, Demuner AJ, Kasuya MCM. Abiotic and biotic degradation of oxo-biodegradable plastic bags by Pleurotus ostreatus. PLoS One. 2014;9(11):e107438.
Montazer Z, Habibi Najafi MB, Levin DB. Microbial degradation of low-density polyethylene and synthesis of polyhydroxyalkanoate polymers. Can J Microbiol. 2019;65(3):224–34.
Skariyachan S, Manjunatha V, Sultana S, Jois C, Bai V, Vasist KS. Novel bacterial consortia isolated from plastic garbage processing areas demonstrated enhanced degradation for low density polyethylene. Environ Sci Pollut Res. 2016;23(18):18307–19.
Kawai F, Oda M, Tamashiro T, Waku T, Tanaka N, Yamamoto M, et al. A novel Ca2+-activated, thermostabilized polyesterase capable of hydrolyzing polyethylene terephthalate from Saccharomonospora viridis AHK190. Appl Microbiol Biotechnol. 2014;98(24):10053–64.
Yoshida S, Hiraga K, Takehana T, Taniguchi I, Yamaji H, Maeda Y, et al. A bacterium that degrades and assimilates poly (ethylene terephthalate). Science. 2016;351:1196–9.
Skariyachan S, Patil AA, Shankar A, Manjunath M, Bachappanavar N, Kiran S. Enhanced polymer degradation of polyethylene and polypropylene by novel thermophilic consortia of Brevibacillus sps. And Aneurinibacillus sp. screened from waste management landfills and sewage treatment plants. Poly Degrad Stab. 2018;149:52–68.
Tian L, Kolvenbach B, Corvini N, Wang S, Tavanaie N, Wang L, et al. Mineralisation of 14C-labelled polystyrene plastics by Penicillium variabile after ozonation pre-treatment. New Biotechnol. 2017;38:101–5.
Hung CS, Zingarelli S, Nadeau LJ, Biffinger JC, Drake CA, Crouch AL, et al. Carbon catabolite repression and Impranil polyurethane degradation in Pseudomonas protegens strain Pf-5. Appl Environ Microbiol. 2016;82(20):6080–90.
Sumathi T, Viswanath B, Sri Lakshmi A, SaiGopal DVR. Production of laccase by Cochliobolus sp isolated from plastic dumped soils and their ability to degrade low molecular weight PVC. Biochem Res Int. 2016;2016:9519527.
Austin HP, Allen MD, Donohoe BS, Rorrer NA, Kearns FL, Silveira RL, et al. Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc Natl Acad Sci U S A. 2018;115(19):E4350–E7.
Son HF, Cho IJ, Joo S, Seo H, Sagong H-Y, Choi SY, et al. Rational protein engineering of thermo-stable PETase from Ideonella sakaiensis for highly efficient PET degradation. ACS Catal. 2019;9(4):3519–26.
Yasuhira K, Uedo Y, Takeo M, Kato D-I, Negoro S. Genetic organization of nylon-oligomer-degrading enzymes from alkalophilic bacterium, Agromyces sp. KY5R. J Biosci Bioeng. 2007;104(6):521–4.
Montazer Z, Habibi-Najafi MB, Mohebbi M, Oromiehei A. Microbial degradation of UV-pretreated low-density polyethylene films by novel polyethylene-degrading bacteria isolated from plastic-dump soil. J Polym Environ. 2018;26(9):3613–25.
Novotný Č, Malachová K, Adamus G, Kwiecień M, Lotti N, Soccio M, et al. Deterioration of irradiation/high-temperature pretreated, linear low-density polyethylene (LLDPE) by Bacillus amyloliquefaciens. Int Biodeterior Biodegradation. 2018;132:259–67.
Hu X, Osaki S, Hayashi M, Kaku M, Katuen S, Kobayashi H, et al. Degradation of a terephthalate-containing polyester by thermophilic actinomycetes and Bacillus species derived from composts. J Polym Environ. 2008;16(2):103–8.
Oberbeckmann S, Labrenz M. Marine microbial assemblages on microplastics: Diversity, adaptation, and role in degradation; 2020. p. 209–32.
Watanabe T, Ohtake Y, Asabe H, Murakami N, Furukawa M. Biodegradability and degrading microbes of low-density polyethylene. J Appl Polym Sci. 2009;111(1):551–9.
Ikada E. Electron microscope observation of biodegradation of polymers. J Environ Polym Degrad. 1999;7(4):197–201.
Shah AA, Hasan F, Hameed A, Ahmed S. Biological degradation of plastics: a comprehensive review. Biotechnol Adv. 2008;26(3):246–65.
Mierzwa-Hersztek M, Gondek K, Kopeć M. Degradation of polyethylene and biocomponent-derived polymer materials: an overview. J Polym Environ. 2019;27(3):600–11.
Swift GJHobpHA, Amsterdam. Non-medical biodegradable polymers: environmentally degradable polymers. In: Domb AJ, Kost J, Wiseman DM, editors. Handbook of biodegradable polymers. Amsterdam: Harwood Academic Publishers; 1997. p. 473–511.
Nishida H, Tokiwa Y. Distribution of poly (β-hydroxybutyrate) and poly (ε-caprolactone) aerobic degrading microorganisms in different environments. J Environ Polym Degrad. 1993;1(3):227–33.
Li M, Shi Y, Li Y, Sun Y, Song C, Huang Z, et al. Shift of microbial diversity and function in high-efficiency performance biotrickling filter for gaseous xylene treatment. J Air Waste Manag Assoc. 2019;69(9):1059–69.
Scott MJ, Jones MN. The biodegradation of surfactants in the environment. Biochim Biophys Acta Biomembr. 2000;1508(1):235–51.
Pitt JJ. Principles and applications of liquid chromatography-mass spectrometry in clinical biochemistry. Clin Biochem Rev. 2009;30(1):19–34.
Jung H-W, Yang M-K, Su R-C. Purification, characterization, and gene cloning of an Aspergillus fumigatus polyhydroxybutyrate depolymerase used for degradation of polyhydroxybutyrate, polyethylene succinate, and polybutylene succinate. Poly Degrad Stab. 2018;154:186–94.
Seo H, Kim S, Son HF, Sagong H-Y, Joo S, Kim K-J. Production of extracellular PETase from Ideonella sakaiensis using sec-dependent signal peptides in E. coli. Biochem Biophys Res Commun. 2019;508(1):250–5.
Perz V, Bleymaier K, Sinkel C, Kueper U, Bonnekessel M, Ribitsch D, et al. Substrate specificities of cutinases on aliphatic-aromatic polyesters and on their model substrates. New Biotechnol. 2016;33(2):295–304.
Acero EH, Ribitsch D, Steinkellner G, Gruber K, Greimel K, Eiteljoerg I, et al. Enzymatic surface hydrolysis of PET: effect of structural diversity on kinetic properties of cutinases from Thermobifida. Macromolecules. 2011;44(12):4632–40.
Sulaiman S, Yamato S, Kanaya E, Kim J-J, Koga Y, Takano K, et al. Isolation of a novel cutinase homolog with polyethylene terephthalate-degrading activity from leaf-branch compost by using a metagenomic approach. Appl Environ Microbiol. 2012;78(5):1556.
Kawai F, Kawabata T, Oda M. Current knowledge on enzymatic PET degradation and its possible application to waste stream management and other fields. Appl Microbiol Biotechnol. 2019;103(11):4253–68.
Ronkvist ÅM, Xie W, Lu W, Gross RA. Cutinase-catalyzed hydrolysis of poly (ethylene terephthalate). Macromolecules. 2009;42(14):5128–38.
Marten E, Müller R-J, Deckwer W-D. Studies on the enzymatic hydrolysis of polyesters I. low molecular mass model esters and aliphatic polyesters. Poly Degrad Stab. 2003;80(3):485–501.
Marten E, Müller R-J, Deckwer W-D. Studies on the enzymatic hydrolysis of polyesters. II. Aliphatic–aromatic copolyesters. Poly Degrad Stab. 2005;88(3):371–81.
Müller R-J, Schrader H, Profe J, Dresler K, Deckwer W-D. Enzymatic degradation of poly (ethylene terephthalate): rapid hydrolyse using a hydrolase from T. fusca. Macromol Rapid Commun. 2005;26(17):1400–5.
Tournier V, Topham CM, Gilles A, David B, Folgoas C, Moya-Leclair E, et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature. 2020;580(7802):216–9.
Che S, Men Y. Synthetic microbial consortia for biosynthesis and biodegradation: promises and challenges. J Indust Microbiol Biotechnol. 2019;46(9-10):1343–58.
Zanaroli G, Di Toro S, Todaro D, Varese GC, Bertolotto A, Fava F. Characterization of two diesel fuel degrading microbial consortia enriched from a non acclimated, complex source of microorganisms. Microb Cell Factories. 2010;9(1):10.
McCarty NS, Ledesma-Amaro R. Synthetic biology tools to engineer microbial communities for botechnology. Trends Biotechnol. 2019;37(2):181–97.
Bernstein HC, Carlson RP. Microbial consortia engineering for cellular factories: in vitro to in silico systems. Comput Struct Biotechnol J. 2012;3:e201210017.
Funding
This work was conducted as part of the Aotearoa Impacts and Mitigation of Microplastics (AIM2) project, in receipt of funds from a New Zealand Ministry of Business, Innovation and Employment (MBIE) Endeavour Fund Grant (C03X1802). Additional support was also provided via a University of Auckland Doctoral Scholarship (to SM) and doctoral scholarship funds provided via the George Mason Centre for the Natural Environment (to VG).
Author information
Authors and Affiliations
Contributions
All authors contributed to the research, writing and editing of this manuscript and provider their full consent for publication. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
No ethics approvals or consents were required.
Competing interests
No competing interests are declared.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lear, G., Kingsbury, J.M., Franchini, S. et al. Plastics and the microbiome: impacts and solutions. Environmental Microbiome 16, 2 (2021). https://doi.org/10.1186/s40793-020-00371-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40793-020-00371-w