
Abstract
5-Aminolevulinic acid (5-ALA), a non-proteinogenic five-carbon amino acid, has received intensive attentions in medicine due to its approval by the US Food and Drug Administration (FDA) for cancer diagnosis and treatment as photodynamic therapy. As chemical synthesis of 5-ALA performed low yield, complicated processes, and high cost, biosynthesis of 5-ALA via C4 (also called Shemin pathway) and C5 pathway related to heme biosynthesis in microorganism equipped more advantages. In C4 pathway, 5-ALA is derived from condensation of succinyl-CoA and glycine by 5-aminolevulic acid synthase (ALAS) with pyridoxal phosphate (PLP) as co-factor in one-step biotransformation. The C5 pathway involves three enzymes comprising glutamyl-tRNA synthetase (GltX), glutamyl-tRNA reductase (HemA), and glutamate-1-semialdehyde aminotransferase (HemL) from α-ketoglutarate in TCA cycle to 5-ALA and heme. In this review, we describe the recent results of 5-ALA production from different genes and microorganisms via genetic and metabolic engineering approaches. The regulation of different chassis is fine-tuned by applying synthetic biology and boosts 5-ALA production eventually. The purification process, challenges, and opportunities of 5-ALA for industrial applications are also summarized.
Similar content being viewed by others

Introduction and development of 5-ALA
5-aminolevulinic acid (5-ALA), an endogenous non-proteinogenic five-carbon amino acid, is the indispensable intermediate which involves in the tetrapyrrole biosynthesis and is the first compound from the porphyrin synthesis to heme. 5-ALA as the precursor of heme, chlorophylls, and vitamin B12 significantly affects the cell growth and metabolic flux. 5-ALA has been tested and applied as a prodrug for leukemia cells treatment (Malik and Lugaci 1987). In this therapy, 5-ALA was observed in the porphyrin metabolism, which 5-ALA converted to photosensitizer protoporphyrin IX (PPIX) and showed fluorescence signal when 5-ALA was accumulated inside the cell. PPIX would produce reactive oxygen species (ROS) under appropriate wavelength and causes cell damage by generating singlet oxygen. With PPIX accumulation, the tumor cell is visualized with emission of fluorescence. 5-ALA-induced photodynamic therapy and diagnosis had been reported since 1990 (Kennedy et al. 1990) and was approved by FDA in 2017.
On the other hand, therapy with 5-ALA not only possesses potential for topical, rapid and effective application but is also the non-toxic to human body (Wild et al. 2005). Therefore, 5-ALA has more advantages than using chemotherapy and surgeries (Peng et al. 1997a). When the reduced ferrochelatase activity in the tumor cells, the excess PPIX leads to formation of reactive oxygen species and thereby cell apoptosis when exposed to suitable wavelength (Peng et al. 1997b). Besides, 5-ALA can be used to monitor the difference between tumor and normal tissue by time (Krieg et al. 2002; Yang et al. 2015).
In addition to the application in medicine, 5-ALA has also been employed in the agriculture as it is harmless and belongs to biodegradable herbicide and insecticide, growth-promoting factor, thus enhancing the productivity of crops (Hotta et al. 1997). When plants are treated with high concentration of 5-ALA and exposed to light, the ROS produced by excess PPIX will oxidize unsaturated fatty acids, thereby damaging plants. The same mechanism was also observed when proper 5-ALA concentration was used for biodegradable insecticide (Sasikala et al. 1994; Sasaki et al. 2002). Although the ROS was generated at a low level under the normal condition, the balance was disturbed by environmental stress, such as salt, temperature, and drought, which would rapidly increase ROS thus to cause oxidative damage to lipids, proteins, and nucleic acids in the cell. As a key precursor in the biosynthesis of porphyrins, 5-ALA has been considered to increasing the yield of foliage and root. Previous studies showed that the yield and quality of plants and crops could be improved by 5-ALA (Hotta et al. 1997; Dolmans et al. 2003; Liu et al. 2013). As a conclusion, 5-ALA is able to promote seedling growth of plants by raising chlorophyll content and net photosynthetic rate to prevent stress damage (Naeem et al. 2012; Liu et al. 2014).
Till now, six methods of chemical synthesis for 5-ALA have been mentioned. 5-ALA can be synthesized from levulinic acid, 2-hydroxypyridine, furfural, furfurylamine, tetrahydrofurfurylamine, and succinic acid (Sasaki et al. 2002). Above all, the photo-oxidation of furfurylamine is regarded as the most practical method. However, with the awareness of environmental protection and sustainability, strategies related to biosynthesis of 5-ALA have attracted more attentions. Two main metabolic pathways, C4 and C5 pathway, for the biosynthesis of 5-ALA from the precursors on tricarboxylic acid (TCA) cycle in organisms have been reported (Fu et al. 2010; Kang et al. 2004, 2017). Interestingly, the C4 pathway is present in birds, mammals, yeast, and purple non-sulfur-photosynthetic bacteria, while the C5 pathway exists in algae, higher plants, and many bacteria, including Escherichia coli and archaea (Yang et al. 2016). In the C4 pathway, 5-ALA is derived from the condensation of succinyl-coenzyme A (succinyl-CoA) and glycine by the 5-ALA synthase (ALAS, encoded by the hemA gene) (Sasaki et al. 2002). On the other hand, 5-ALA formation involves three sequential enzymatic reaction catalyzed by glutamyl-tRNA synthetase, glutamyl-tRNA reductase, and glutamate-1-semialdehyde aminotransferase (encoded by gltX (Huang and Wang 1986; Schneegurt and Beale 1988), hemA (Li et al. 1989; Schauer et al. 2002), and hemL (Jahn et al. 1992; Wang et al. 1984) gene, respectively) from TCA cycle intermediate, glutamate in C5 route. The C4 pathway uses a unique 5-ALA synthase (ALAS) as single-step reaction is easier for regulation than that applies the C5 pathway, even most prokaryotes originally obtain this pathway. But C5 pathway is not limited or restricted by using glycine as the precursor. In the view of green, sustainable, and highly efficient production of chemicals with appropriate organisms, for example, Escherichia coli, is more favorable.
Sasaki et al. has reviewed the biosynthesis and application of 5-ALA in 2002 (Sasaki et al. 2002). They have pointed out the strategies and affecting factors for 5-ALA production in microorganisms. The recent advances in synthetic biology have been explored for 5-ALA biosynthesis. The microbial production of 5-ALA through biological and genetic approaches and challenges were also discussed recently (Kang et al. 2017). Herein, we will introduce the determinant factors and novel strategies for 5-ALA bioproduction via C4 and C5 pathway by genetic, synthetic, and metabolic approaches in the last decade. Moreover, the regulation between 5-ALA accumulation and heme, purification, challenges, and prospective of 5-ALA for industry are also discussed.
Production of 5-ALA via C4 pathway in Escherichia coli
In previous study, hemAC4 gene fragment was mostly amplified from purple non-sulfur bacteria, such as Rhodobacter sphaeroides and Rhodobacter capsulatus. ALAS is a PLP-dependent homodimeric enzyme which could catalyze the decarboxylation as well as the deprotonation (Liu et al. 2014). Co-factor PLP firstly binds to an active site lysine and therefore changes conformation endowing ALAS with ability to catalyze the condensation reaction. Next, glycine binds to ALAS-PLP complex accompanied with deprotonation and forms a quinonoid intermediate (Stojanovski et al. 2014). Subsequently, succinyl-CoA reacted with the quinonoid intermediate to generate an unstable compound. Active site histidine would catalyze the decarboxylation after release of CoA. Structure simulation and analysis have unveiled that the lysine and histidine residue at K248 and H142 of RcALAS are responsible for the affinity to co-factor and substrates. Stojanovski et al. have taken advantage of simulation to illustrate that the substrate specificity of ALAS was attributed to the conformational hindrance if ALAS-PLP complex bounded to amino acid except for glycine (Stojanovski et al. 2014).
As an enzyme belongs to α-oxoamine synthase (AOS) family, ALAS adapts asparagine instead of histidine and phenylalanine which are widely observed at analogous site in the structure. The unusual amino acid usage is assumed that histidine and phenylalanine may impede the 5-ALA release and even result in balance shift to reverse reaction (Stojanovski and Ferreira 2015).
Gene selection and enzyme activity
Choosing proper proteins to redirect the carbon flux to the target chemicals is highly significant because the enzyme from different organisms also diverged considerably in its specific activity, kinetic constants, and optimal conditions. An extremely active enzyme can catalyze the reaction more rapidly as well as transform the substrates to products more efficiently. 5-Aminolevulinic acid synthase (ALAS), a key enzyme of the C4 pathway, is encoded by hemA gene from different organisms, such as Agrobacterium radiobacter (Fu et al. 2010), Bradyrhizobium japonicum (Choi et al. 1999; Jung et al. 2004), Rhodobacter capsulatus (Kang et al. 2004; Lou et al. 2014; Yu et al. 2020), Paracoccus (Sato et al. 1985), Rattus norvegicus (Nakakuki et al. 1980), Rhodobacter sphaeroides (Kang et al. 2011a, b; Shih et al. 2021; Tan et al. 2020a; Tran et al. 2019), Rhodopseudomonas palustris (Ong et al. 2013; Zhang et al. 2013; Liu et al. 2016), and Saccharomyces cerevisiae (Volland and Felix 1984). The ALAS from different organisms used for 5-ALA production via C4 pathway in the genetic E. coli strains are summarized in Table 1. Among all, the specific activities of ALAS from R. sphaeroides, A. radiobacter, R. capsulatus, and R. palustris were 0.117, 0.151, 0.198, and 0.924 µmol/min/mg, respectively (Lou et al. 2014), while Meng et al. reported that the ALAS from Laceyella sacchari possessed the highest activity of 7.8 µmol/min/mg (Meng et al. 2015). Although the reaction system and approach from separate group would lead to different calculation, the comparison of ALAS activity from diverse strains is still useful. The activity and characteristic of enzymes have also been organized in BRENDA database (www.brenda-enzymes.org). From the database, the ALAS from R. palustris possesses higher activities than other organisms, which made it a promising candidate for high-efficient 5-ALA yield. Based on this concept, Zhu et al. have co-expressed R. palustris ALAS, catalase, and superoxide dismutase in E. coli, and finally achieved the highest 5-ALA production in record as 11.5 g/L in a 5-L bioreactor using fermentation (Zhu et al. 2019).
Codon optimization of ALAS
Codon optimization of ALAS is favorable to reach higher protein expression level, protein structures, and specific activity to further obtain higher 5-ALA production. In the central dogma, protein expression is the final step of several complicated processes, including regulation at the level of transcription, mRNA turnover, protein translation, and post-translational modifications leading to the formation of the stable and functional protein. Although there are only twenty amino acids in protein sequences, the codons specified for all the amino acids are sixty one, i.e., codon usage bias. Thus, the regulation at the level of transcription is highly relative to the codon usage bias (Kane 1995). For example, the heterologous protein expression showed the lower protein yield due to the rare codon usage bias or the insufficient of transfer RNA (tRNA) as the native codon is used. Consequently, for most sequenced genomes, synonymous codons are not equal-frequently used, as called “non-optimal codons,” leading to the low protein level due to the limiting available cognate tRNAs in the expression host. Kane also discussed the effect of rare codons for heterologous protein expression in E. coli, reporting that some codons would cause translational errors (Kane 1995). Angov reviewed the effect of codon bias on mRNA structure and gene translation, which further regulated the protein expression levels (Angov 2011). The approaches for codon usage analysis and optimization tools were also provided by using a software for calculation of codon adaptation index (CAI), which was an empirical measure of protein expressivity (Fuglsang 2003). Menzella evaluated “one amino acid one codon” and “codon randomization” for codon optimization, suggesting that more protein expression was observed from the sequence with codon randomization (Menzella 2011). Accordingly, the hemA gene from S. arizona was expressed after codon optimization and enhance 5-ALA accumulation in E. coli via C5 pathway (Zhang et al. 2015, 2018a). Li et al. also expressed codon-optimized ALAS from S. cerevisiae in E. coli and improved 5-ALA titer via C4 pathway (Li et al. 2016). Furthermore, Yu et al. showed the expression of codon-optimized R. capsulatus ALAS with chaperone would obtain more soluble ALAS and further produce high level of 5-ALA (Yu et al. 2020). However, the overly excessive protein expression level leads to the formation of insoluble products sequestered in inclusion bodies which hinder the function of proteins (Sørensen and Mortensen 2005). As the result, the modification of codon for regulation of optimal expression level should be the priority instead of reaching the highest protein production. Besides, the pRARE plasmid carrying tRNAs recognizing rare codons was also reported to be effective to improve heterologous protein expression (Burgess-Brown et al. 2008; Effendi et al. 2020). Moreover, the Rosetta(DE3) strain was acquired by introducing pRARE into E. coli BL21(DE3) for assisting the transcription. The ALAS from R. sphaeroides and A. radiobacter was expressed under T7 promoter in Rosetta(DE3) and more 5-ALA was reached with higher enzyme activity than that in BL21(DE3) (Fu et al. 2007, 2010; Yang et al. 2013). The activities of ALAS from A. radiobacter, R. sphaeroides and R. capsulatus were compared in Rosetta(DE3). Among the enzymes, ALAS from R. capsulatus showed the highest activity and 8.8 g/L of 5-ALA titer was achieved (Lou et al. 2014). Besides, GenScript company (https://www.genscript.com/tools/rare-codon-analysis) provides the online software, while Integrated DNA Technologies® supports codon optimization tool for DNA synthesis which meets the goal to obtain higher expression level.
Transporter and chaperone
As high concentration of 5-ALA is toxic to E. coli, it is necessary to avoid the 5-ALA accumulation in the cell. Even though cell can drain the metabolism out of the cell spontaneously, the cell still requires transporter to accelerate the export of 5-ALA. Several amino acid transport systems are identified in E. coli, while the efflux mechanism of 5-ALA is still an enigma (Zhu et al. 2020). Auspiciously, the structure of 5-ALA is similar to glycine; thus, an amino acid exporter would solve the problem. RhtA is capable of translocating a variety of amino acids and related compounds, such as dipeptide and amino acid analogs, and has been reported extremely powerful for different amino acid excretion (Diesveld et al. 2009). Yang et al. accomplished 5-ALA up to 14.7 g/L via application of RhtA in C. glutamicum (Yang et al. 2016). The YeaS is a leucine exporter, which also encodes to be RhtB transporter family. The over-expression of yeaS results in resistance of cells to leucine analogs, glycyl-L-leucine dipeptide, and other amino acids. However, the efficiency of YeaS is much less than RhtA (Kang et al. 2011a, b). Therefore, RhtA is an efficient transporter for 5-ALA till now.
Chaperone which assists the folding of protein is a critical part to be deliberated for the insoluble aggregates of the foreign protein expressed in E. coli. Recent report has demonstrated the optimal chaperone is powerful to improve the functional expression of recombinant horseradish peroxidase (Yang et al. 2019a, b). Considering the translational effect of ALAS and preventing the inclusion bodies forming is crucial for ALA production. The GroELS system includes GroEL and GroES, while DnaK system consists of DnaK, DnaJ, and GrpE are the most eminent chaperone systems (de Marco et al. 2007). GroEL is a cylindrical complex of double ring of subunits that composes of an ATP-binding domain, an intermediate hinge domain, and an apical domain in the three-dimensional structure as a protein-folding machinery in E. coli (Hayer-Hartl et al. 2016). The previous research has applied GroEL chaperone to enhance 5-ALA production with RcALAS from 1.2 g/L to 3.6 g/L due to the improvement of the soluble protein (Yu et al. 2020). DnaK, DnaJ, and GrpE are the members of Hsp70 chaperone family. At first, DnaK prevents and repairs the thermally induced protein damage in E. coli. Moreover, DnaK also binds with various abnormal proteins and protects RNA polymerase from thermal inactivation. On the other hand, DnaJ overturns the aggregation of inactive protein and target DnaK to the substrate. Shih et al. compared the 5-ALA production with the assistance of GroELS and DnaKJ, respectively (Shih et al. 2021). Although DnaK has been reported to refold other inactive proteins, the efficiency is not observable in 5-ALA production (Xie et al. 2003; Yang et al. 2019a, b; Yu et al. 2020). Till now, GroE system is more beneficial to ALAS folding and leading higher 5-ALA production. Meanwhile, it is worth mentioning that high 5-ALA concentration in medium may harm cell via generating ROS. Therefore, expressing catalase and superoxide dismutase is a promising strategy to enhance the tolerance of E. coli toward the oxidative stress caused by ROS from 5-ALA (Zhu et al. 2019). The effects of 5-ALA on cell integrity and morphology were demonstrated to deal with oxidative stress and enhance 5-ALA accumulation effectively in E. coli.
Medium and trace element
Thanks to the development of genetic engineering, biosynthesis is a more feasible and efficient strategy for 5-ALA production of chemicals in recent decades. In 1996, Van der Werf and Zeikus investigated the effect of different carbon sources and amino acids on 5-ALA production (Van der Werf and Zeikus 1996). Luria–Bertani broth (LB) and Terrific Broth (TB) composed of tryptone and yeast extract have been widely used as nutrient-rich media and have been applied for 5-ALA production (Choi et al. 2008; Lou et al. 2014; Van der Werf and Zeikus 1996). However, minimum medium that includes only glucose, inorganic nitrogen source, and mineral salts as a cost-effective medium has been more favorable for 5-ALA production in recent years (Choi et al. 2008; Kang et al. 2011a, b; Zhang et al. 2015). In the early study of medium effect, concentrations of precursors (i.e., glycine and succinic acid) have the great influences on 5-ALA production in recombinant E. coli BL21(DE3)-pLysS and achieved 1.3 g/L (Chung et al. 2005). Lin et al. have explored that not only glucose but also xylose could serve as inhibitor of HemB. With supply of glucose and xylose, the 5-ALA production yield reached to 7.3 g/L (Lin et al. 2009). Cui et al. demonstrated that yeast extract played an important role in 5-ALA production (Cui et al. 2019). Although 12 g/L yeast extract dramatically enhanced both cell growth and 5-ALA accumulation, lower concentration of yeast extract was more feasible for industrial bioproduction.
For ion effect, zinc, copper, and cobalt ion are definitely affecting the ALAS activity (Liu et al. 2016; Lou et al. 2014). HemB would be only activated by a quite low concentration of Mn2+ ion. Moreover, Mg2+ ion even activated HemB activity (Erskine et al. 1999; Nandi et al. 1968). Thus, concentrations of Mn2+ and Mg2+ ion are the critical factors for 5-ALA production. Cui also showed that 1 g/L MgSO4·7H2O and 0.01 g/L MnSO4·7H2O were moderate for 5-ALA production in E. coli (Cui et al. 2019). Reducing magnesium salts and adding calcium salts could be a feasible formula for enhancing 5-ALA production, since Ca2+ ion showed less influence on ALAS activity. Most recently, Fe2+ ion could be a tricky factor for increasing 5-ALA accumulation. With 30 µM Fe2+ supplement, the 5-ALA titer has significantly increased in R. sphaeroides (Tangprasittipap et al. 2007), while addition of 1 mM Fe2+ showed a dramatical decline in ALAS activity (Nakakuki et al. 1980). Therefore, defining the moderate Fe2+ ion concentration is a promising strategy to further enhance 5-ALA biosynthesis as well.
Gene deletion in the chromosome
As the genetic engineering tools are well developed, producing strains are no longer limited to use well-known hosts, for example, original E. coli BL21(DE3) or MG1655. Manipulation of several genes could be conducted to refine a strain into unique host which pushes and pulls the metabolic flux into product-oriented pathway.
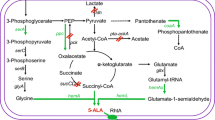
Recently, genes deletion has become a promising strategy for further promotion of 5-ALA. Noh et al. have revealed that deletion of sucA leaded to lower biomass but higher 5-ALA production in E. coli (Noh et al. 2017). However, deleting iclR, a transcriptional regulator of glyoxylate-related genes (aceBAK), would redistribute the metabolic flux from TCA cycle to glyoxylate route but reduce 5-ALA production due to down-regulating the isocitrate dehydrogenase activity. Afterward, Ding et al. knocked out sdhAB and sucCD to block conversion of succinyl-CoA into other down-stream metabolites (Ding et al. 2017). According to the results, only deletion of sucCD showed a significant increase in 5-ALA titer. On the other hand, deletion of recA (Cui et al. 2019) and endA (Zhang et al. 2019) which prevented homologous recombination by the hosts thus enhanced the genetic stability. Recently, Miscevic et al. also represented 5-ALA production from glycerol in E. coli through metabolic engineering (Miscevic et al. 2020). The hemB was depressed and the ldhA was deleted to dissimilate carbon flux toward the TCA cycle. Afterward, the genes sdhA and iclR encoding succinate dehydrogenase complex flavoprotein subunit A and transcriptional AceBAK operon repressor were knocked out to further enhance succinyl-CoA formation in the cell. With the double mutant of iclR and sdhA, higher 5-ALA production was achieved since the succinyl-CoA could be increase by the carbon flux at the succinate node via the glyoxylate shunt (Fig. 1).

The C4 pathway for 5-ALA and heme synthesis. The up and down arrows beside the gene indicate the up- or down-regulation of the gene which can increase 5-ALA accumulation
To sum up, the 5-ALA productions via C4 pathway in E. coli are listed in Table 2 and gene knock out is a double-edged concept. Each metabolite plays a role in the whole metabolism network. Thus, gene deletion will easily trigger the imbalance due to the metabolic flux changes and causes either poor cell growth or low productivity. The trade-off between cell growth and product titer must be considered. Also, replenishment or regeneration of crucial compounds should be a possible strategy to compensate the gene deficiency.
Production of 5-ALA by regulation of C5 pathway in E. coli
The rate-limiting enzyme in C5 pathway is HemA catalytic reaction. HemA catalyzes the reaction that convert the glutamyl-tRNA glutamate-1-semialdehyde (GSA). Interestingly, HemA and HemL would assemble a synergistic complex to protect highly reactive compound GSA (Lüer et al. 2005). After GSA formation, HemL, a PLP-dependent enzyme (Zhang et al. 2019), could catalyze the quick reaction to convert glutamate-1-semialdehyde to 5-ALA. In recent publications, hemA was mostly derived from Salmonella sp., while hemL was derived from E. coli for enhancing 5-ALA biosynthesis. To over-express gltX gene playing an intuitive way to enhance 5-ALA production, however, it may not work as expected. According to Su et al.s’ study, over-expression of gltX may even up-regulate hemB transcription and result in lower 5-ALA accumulation (Su et al. 2019). This phenomenon indicates strong feedback inhibition of gltX which is also controlled by the final product, heme.
Fine-tuning heme and regulation in the pathway
In 1970, the first research of 5-ALA production through C5 pathway by Chlorella was published in which the carbon dioxide was chosen as the main substrate (Beale 1970). Afterward, the biosynthesis of 5-ALA has been considered as the rate-limiting step for heme biosynthesis and is regulated in organisms (Verderber et al. 1997). Heme is a porphyrin derivative and essential hemoproteins which are important for oxygen transport, eliminating ROS and transferring electrons for energy generation (Mayfield et al. 2011). The transcriptional regulation of hemC, hemD, hemH, hemA, and hemM genes on heme synthesis pathway was firstly examined by McNicolas et al., revealing the regulation between genes and heme availability in E. coli (McNicolas et al. 1997). Kwon et al. over-expressed hemA from R. capsulatus to increase 5-ALA and down-stream genes, including hemB, hemC, hemD, and hemF genes from E. coli, hemE gene from Synechocystis sp., and hemH gene from Bacillus subtilis, to promote heme production (Kwon et al. 2003). The result showed the greatly positive correlation between 5-ALA and heme production. Also, down-regulation of hemB and hemH genes decreased the metabolic flux from 5-ALA to heme. Moreover, the regulatory of hemA and hemL, hemB, hemD, hemF, hemG, and hemH genes on heme synthesis pathway was investigated (Zhang et al. 2015). The regulation of transcription level of genes was also explored by quantitative real-time PCR, in which feedback inhibition of hemB by the intermediate PPIX was verified, suggesting that regulation of heme biosynthesis pathway genes is complicated. Zhao et al. confirmed that the C5 pathway obtained superior performance over C4 pathway for heme production, and further adjusted the metabolic flux involved in heme biosynthesis (Zhao et al. 2018).
The strategies for 5-ALA production in E. coli via C5 pathway in the past decade are organized in Table 3. A small mRNA, ryhB, was applied in E. coli to regulate the heme biosynthesis pathway and 5-ALA production (Li et al. 2014). The RyhB is able to reduce iron-binding proteins expression and has been proved to down-regulate transcription level of hemB and hemH; thus, the 5-ALA accumulation was enhanced to 116%. The regulatory of heme synthesis pathway was explored which relied on the transcriptional regulation of each gene in the pathway (Kang et al. 2011a, b). Kang et al. firstly over-expressed hemA from S. arizona, hemL, and threonine/homoserine exporter rhtA from E. coli, reaching 4.13 g/L of 5-ALA from glucose without any other co-substrate or inhibitor. At first, the up-regulation of hemD and hemF increased 5-ALA accumulation. Next, the over-expression of hemB, hemG, or hemH caused reduction on most of genes in the pathway, while over-expression of hemD resulted in significant increase of ALA. After applying modular optimization for expression cassette, the highest 5-ALA was achieved at 3.25 g/L. The iron was also found to be indispensable for cell growth and heme biosynthesis (Zhang et al. 2016). With the optimized iron concentration and co-expression of hemA, hemL, hemF, and hemD, the 5-ALA production achieved 4.05 g/L in batch fermentation. Su et al. employed CRISPRi system to fine-tune hemB on heme synthesis pathway (Su et al. 2019). The 6 CRISPRi targeting various regions of hemB were selected for down-regulation of the gene by 15, 19, 33, 36, 71, and 80% respectively. The optimized targeting site enhanced fourfold of 5-ALA compared to the original strain, thus providing a new insight into fine-tuning the heme biosynthesis pathway for 5-ALA accumulation.
The above references represented the great relationship between 5-ALA in C5 pathway and heme synthesis, and the regulation of transcriptional level on the pathway affected both 5-ALA and heme accumulation. Therefore, understanding the overall regulatory mechanism of heme biosynthesis pathway genes is vital for 5-ALA production.
Other regulations in TCA cycle
The metabolic engineering has been greatly benefited from the synthetic and system biology owing to the abundant advancement as it collaborated with metabolic regulation and has applied to optimize natural chemical production (Jones et al. 2015). Producing maximum chemical is the principal objective for balancing metabolic pathway, while the imbalance occurred in the host causing death to the cell (Yang et al. 2020). Since 5-ALA can be biologically produced via metabolic route in which the TCA cycle and heme synthesis pathway are involved, the regulation and redistribution of metabolic flux are crucial for its production in microorganisms.
Not only the heme synthesis pathway is crucial to 5-ALA accumulation but engineering metabolic flux from the upstream and TCA cycle is also important to its biosynthesis. The carbon flux from glucose was optimized by Noh et al. by over-expressing gltA and aceA with the deletion of sucA (Noh et al. 2017). Moreover, hemA from S. typhimurium with addition of two lysine codons was employed for its higher activity and stability. However, the deletion of sucA resulted in lower biomass and 5-ALA production, which came from the impaired TCA cycle of insufficient energy production. The over-expression of gltA was reported to show the negative effect on 5-ALA accumulation but a slight improvement in cell biomass and glucose consumption. Finally, the carbon flux on glyoxylate cycle was optimized by fine-tuning the transcription level of aceA, and 3.4 g/L of 5-ALA was obtained after the optimization of culture condition. Moreover, the tuning relationship between heme synthesis genes and 5-ALA production was also provided. Zhang et al. engineered multiple ribosome binding site (RBS) for hemA and hemL over-expression and replaced the promoter of hemB to weaken 5-ALA catabolism (Zhang et al. 2019). Besides, the co-factor PLP for ALAS was synthesized by over-expressing pdxH and the 5-ALA production was finally increased to 5.25 g/L with a two-stage fermentation. This is the first report implementing pdx gene for PLP enhancement to improve 5-ALA accumulation in E. coli. Xue et al. also fine-tuned the supplementation of PLP by integrating pdxY gene and has augmented the 5-ALA synthesis via C4 pathway (Xue et al. 2021). Aiguo and Meizhi introduced hemA1 (encoding glutamyl-tRNAGlu reductase) and its stimulator protein encoded by pgr7 genes from higher plant Arabidopsis thaliana into E. coli for overproduction of 5-ALA via C5 pathway (Aiguo and Meizhi 2019). By over-expressing hemA1 and pgr7 and up-regulating zwf, gnd, pgl, and rhtA, 7.64 g/L 5-ALA was produced from 10 g/L glutamate and 15 g/L glucose at 48 h. In order to obtain a stable and efficient strain for 5-ALA, chemically inducible chromosomal evolution (CIChE) strategy was carried out by Cui et al. for high gene copy expression in E. coli recently (Cui et al. 2019). The expression cassette containing hemA from S. arizona and hemL from E. coli was integrated onto MG1655 and successfully reached 98 copies with 2.72 g/L of 5-ALA. After a long-term adaptive evolution and the deletion of recA, the 5-ALA accumulation was improved and achieved 4.55 g/L at 72 h. The glutamyl-tRNA reductase encoded by hemA was also reported that its N-terminus played a critical role on its stability control, which has been improved by inserting lysine or arginine residues behind Thr2 (Zhang et al. 2017). A heme-responsive regulatory system was designed to containing a heme biosensor HrtR and CRISPRi for regulation of 5-ALA production as well as maintaining the intracellular heme homeostasis for cell growth (Zhang et al. 2020a). The metabolite-binding affinity of HrtR was modified by heme titration and dynamic simulation. They finally improved the 5-ALA production to 5.35 g/L by down-regulating the PBG formation with the regulatory.
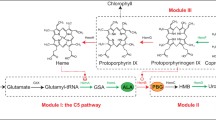
The total concept of 5-ALA production from C5 pathway is shown in Fig. 2. Although the production of 5-ALA via C5 pathway is more difficult than via C4 pathway, it is still important to explore the mechanism and regulation on heme synthesis pathway. Without the demand of substrates (i.e., glycine and succinate), producing 5-ALA via C5 pathway may be more economical and ecofriendly.

The C5 pathway for 5-ALA and heme synthesis. The up and down arrows beside the gene indicate the up- or down-regulation of the gene which can increase 5-ALA accumulation
Biosynthesis of 5-ALA in different microorganisms
Besides E. coli, other organisms have also been studied for more efficient 5-ALA production to achieve different benefits. By applying suitable strains, studies have reported to reach higher 5-ALA accumulation or deal with other issues at the same time. The reports provided the ideas and opportunities for 5-ALA in various organisms and application (Table 4).
Corynebacterium glutamicum
Corynebacterium glutamicum is one of the most important microorganisms for amino acid production, which utilizes C5 pathway for 5-ALA biosynthesis. The recent results of 5-ALA production using C. glutamicum are summarized in Table 4. Ramzi et al. firstly exploited C. glutamicum for 5-ALA production by over-expressing hemA from S. typhimurium and hemL from E. coli (Ramzi et al. 2015). Finally, the highest production was achieved at 2.2 g/L under penicillin induced in the fermentation. The 5-ALA synthesis and heme pathway in C. glutamicum were discussed by Yu et al., showing the similarity of regulation between C. glutamicum and E. coli (Yu et al. 2015). C. glutamicum was able to produce 1.79 g/L of 5-ALA with co-expression of hemA from S. arizona and E. coli hemL in cooperation with optimal iron concentration. To efficiently convert L-glutamate to 5-ALA via C5 pathway, hemA and hemL from S. typhimurium and E. coli were tandemly over-expressed in which glutamate transporter encoded by ncgl1221 was knocked out to prevent glutamate secretion and weaken hemB by replacing its RBS to improve 5-ALA production (Zhang and Ye 2018). Another observation was that the 5-ALA synthesis was improved by inactivating lysE and putP, which reduced the conversion of glutamate to arginine and proline. Thus, to block the secretion of these amino acids has ensured that glutamate is sufficient for conversion into glutamate and further toward 5-ALA. By the three single deletion strains, it could achieve higher 5-ALA titer compared to strain using original C. glutamicum S9114. Ko et al. devoted on the flux redistribution of the TCA cycle toward L-glutamate in C. glutamicum (Ko et al. 2019). As a result, the site-directed mutagenesis was applied in oxoglutarate dehydrogenase inhibitor (OdhI) to allow more glutamate accumulation by inhibiting 2-oxoglutarate dehydrogenase complex. The exporter rhtA from E. coli and the induction of trigger for glutamate from ethambutol enabled 2.9 g/L of 5-ALA production finally. Zhang et al. designed HrrSA regulatory system to fine-tune the RhtA expression for 5-ALA export in response to extracellular heme (Zhang et al. 2020b). The odhA encoding α-KG dehydrogenase was also modulated by temperature-induced promoter to optimize the expression for higher 5-ALA accumulation. Under precise control of dynamic metabolic engineering, 3.16 g/L 5-ALA was achieved through C5 pathway in this study.
The first study applied C4 pathway in C. glutamicum was reported by Feng et al. (2016). The hemA from R. sphaeroides was codon optimized and expressed in C. glutamicum to enhance 5-ALA accumulation. Several genes responsible for acetate and lactate formation were knocked out to centralize the carbon flux (i.e., ldhA, pqo, pta, ackA, and cat). Moreover, four genes encoding high-molecular-weight penicillin-binding proteins (HMW-PBPs, encoded by pbp1a, pbp1b, pbp2a, and pbp2b, respectively) were deleted to increase the permeability of cell wall, and further enhanced extracellular 5-ALA accumulation to 7.53 g/L. Since glycine is an important precursor for 5-ALA synthesis via C4 pathway, the biosynthesis pathway of serine and glycine was remodeled by Zou et al. to improve 5-ALA accumulation based on the multi-gene engineered C. glutamicum strain as aforementioned (Zou et al. 2017). The final 5-ALA concentration was 3.4 g/L by over-expressing serB, serC, mutant serA, and codon-optimized hemA from R. sphaeroides. Out of expectation, deletion of sdaA (encoding L-serine dehydratase, which converts L-serine into pyruvate) decreased biomass and 5-ALA titer due to decline the amount of pyruvate. Yang et al. evaluated several ALAS from different sources and observed a codon-optimized ALAS from R. capsulatus displaying the best potential (Yang et al. 2016). Although the TCA derivative metabolites showed no significant difference in sucCD deficient strain, the 5-ALA production indeed increased. With the co-expression of E. coli rhtA, the titer of 5-ALA was increased to 14.7 g/L through two-stage fermentation from glucose and glycine. However, Chen et al. found out that the hemA from R. palustris possessed higher activity in C. glutamicum than that from R. sphaeroides recently (Chen et al. 2020). The ALAS expression was further optimized for enhanced 5-ALA production via RBS engineering and the phosphoenolpyruvate carboxylase (PPC) was over-expressed to improve biosynthesis of oxaloacetate derivatives. Till now, the highest record of 5-ALA production is applying genetic C. glutamicum as the best strain, which produced 18.5 g/L after 39 h fed-batch fermentation (Chen et al. 2020).
Rhodobacter sphaeroides
Rhodobacter sphaeroides is also a suitable host for 5-ALA production since it naturally possesses 5-ALA biosynthesis via C4 pathway and has been reported to have strong activity of ALAS. Liu et al. firstly applied R. sphaeroides to produce 5-ALA and remove pollutants in wastewater in 2015 (Liu et al. 2015). The light intensity, pH, and concentration of trace elements were optimized to reach higher biomass, 5-ALA production, and chemical oxygen demand (COD) removal. Finally, 31.8 mg/l/h biomass production rate, 1.15 g/L 5-ALA, and 93.3% COD removal were achieved. Since Fe2+ dosage was found to be an important factor to 5-ALA production in according to previous report (Liu et al. 2015), the concentration of Fe2+ and its effect were discussed in 2018 (Liu et al 2018). The optimized dosage of Fe2+ for R. sphaeroides was 400 μmol/L and the mechanism revealed that Fe2+ vastly improved ATP production by up-regulating the nif gene expression. The increasing ATP could enhance the biomass and ALA yield to 4.02 g/L by supplying sufficient energy. In addition, the nifA and nifU gene expression displayed high consistency of co-transcription at the optimal Fe2+ dosage. The 5-ALA production from the wild-type R. sphaeroides was still with limitation even most of the studies used the RsALAS (Liu et al. 2015; Shih et al. 2021).
Saccharomyces cerevisiae
Apart from bacterium, Saccharomyces cerevisiae, a well-known model microorganism used for bioproduction of value-added compounds, has also been applied to produce 5-ALA. The C4 and C5 pathways were both enhanced in S. cerevisiae for the first time in 2018 (Zhang and Ye 2018b). By over-expressing hemL from S. cerevisiae, hemA from R. sphaeroides, and hemA and hemL from E. coli, the 5-ALA production reached to 525.8 mg/L with addition of glycine and succinate. Hara et al. improved 5-ALA production by over-expressing S. cerevisiae HEM1, ACO1, and ACO2 at the first time in 2019 (Hara et al. 2019). The ACO2 encoding aconitase was elucidated to be the rate-limiting enzyme in 5-ALA biosynthesis since iso-citrate was strictly limited in S. cerevisiae. Therefore, increasing gene expression of HEM1 and ACO2 effectively enhanced 5-ALA production by adding 40 mM levulinic acid, an inhibitor to 5-ALA dehydratase (ALAD). The solid-state fermentation (SSF) for 5-ALA was developed by Mao et al., reporting that SSF can be used to efficiently enrich feed food with 5-ALA at a low cost (Mao et al 2020). They achieved 63.8 mg/L 5-ALA production in flask cultivation by over-expressing HEM1 from S. cerevisiae. Finally, a titer of 225.63 mg/kg dry materials was achieved within 48 h through SSF after process optimization, exceeding the usual effective dosage reported in animal trials. This is the first report on combining the simultaneous saccharification and fermentation (SSF) and microbial 5-ALA production, which broadens the application of 5-ALA for feeds, but it still needs more efforts to improve the productivity.
Other genus: Streptomyces, Bacillus, and Shewanella
Streptomyces coelicolor, a gram-positive bacterium well known for the ability to synthesize antibiotics, is considered to be promising hosts for the production of bioactive molecules by expressing foreign genes. Moreover, S. coelicolor possesses better glucose utilization than E. coli and C. glutamicum. The first report to integrate hemA from R. sphaeroides into S. coelicolor genome to produce 5-ALA via C4 pathway was provided (Tran et al. 2019). Their result showed that glucose and yeast extract had strongly positive effect on 5-ALA accumulation in S. coelicolor. After optimization of casamino acid, peptone, malt extract, glycine, and succinic acid, they finally produced 137 mg/L 5-ALA in bioreactor culture. Although several studies stated that S. coelicolor was a suitable candidate for chemical production due to its low endogenous protease activity and efficient secretion of products as well as the lack of a strong restriction system (Anné et al. 2014), the difficulties on genetic manipulation made it hard to reach comparable yield of product with E. coli.
Bacillus subtilis, a generally recognized safe cell (GRAS), was also reported as a host for 5-ALA production via C5 pathway by Liu et al. (2020). The endogenous hemA and hemL were over-expressed effectively under maltose induction and reached 68.45 mg/L 5-ALA after 60-h cultivation. On the other hand, Yi and Ng also put effort in Shewanella oneidensis for 5-ALA production, showing the promising ability of S. oneidensis to produce 5-ALA after stepwise engineering. The expression level of hemD and hemF genes was firstly enhanced under T7 promoter in S. oneidensis, achieving 4.96-fold improvement on 5-ALA accumulation (Yi ang Ng 2020). The glycolysis pathway was regulated by CRISPRi system to augment the carbon flux into TCA cycle and increase 5-ALA synthesis with assistance of ALAS from R. capsulatus, finally reaching 207 mg/L (Yi ang Ng 2021). To sum up, the 5-ALA production requires effective genetic tools and genomic information for DNA manipulation; thus, E. coli and C. glutamicum are still favorable for 5-ALA biosynthesis.
Purification of 5-ALA from bioprocess and the applications
The biosynthesis of 5-ALA is known as sustainable cost-effective and eco-friendly process; however, the purification is complicated due to the final fermentation broths are containing saccharides, protein, amino acids, organic acids, and metal ions, which are produced during microbial growth. Specifically, glycine may affect the efficiency of crystallization of 5-ALA hydrochloride (5-ALA-HCl) (Okada et al. 2016). Therefore, it is necessary to remove the residual compounds prior to the crystallization by chromatography. Ion-exchange chromatography (IEC) with cation exchange resin (CER) is a common strategy to separate 5-ALA from crude solution (Fig. 3), which is also affected by initial 5-ALA concentration, pH, and eluant (Table 5). Venosa et al. employed a Dowex 50 × 8 (strongly acid hydrogen form) CER to separate 5-ALA from its derivatives with 1 M sodium acetate, reaching 90 ± 4% 5-ALA recovery while only 3 to 9% of the 5-ALA derivatives remained (Venosa et al. 2004). However, suitable condition for separating ALA using IEC for the highest yield is different due to the cultural conditions, such as type of medium and other substances aim at increasing growth. Tripetch et al. optimized the separation method of 5-ALA from Rhodobacter sphaeroides suspension to obtain the highest yield and low cost. First, 2% activated carbon was used in decolorization for 5 to 10 min at pH 5 with 95% efficiency. To purify 5-ALA from the suspension, a strong acid CER Dowex 50WX8 resin was used. The decolorized suspension was washed down through two steps: (1) 1 M sodium acetate (pH4.7); (2) 1 M sodium acetate (pH 3.8). As the result, the recovery was > 70% and the elution time was less than 1000 min (Tripetch et al. 2013). Okada et al. also applied a strongly acid CER Amberlite IR-120B® hydrogen form with 0.3 M ammonia to purify 5-ALA from the culture containing glycine and metabolites, achieving 100% recovery (Okada et al. 2016). Nevertheless, 5-ALA is highly unstable in the alkaline solution after desorption. Hence, 5-ALA solution was directly condensed from 60.3 g/L to 393 g/L by using a vacuum machine, mixed with hydrochloride acid and poor solvent as a precipitation method through dropping the 5-ALA-HCl solution into acetone to obtain crystals of 5-ALA-HCl with 99.5% purity. Lin et al. also condensed 5-ALA solution from 46.8 g/L to 570 g/L with the same approach before crystallization (Lin et al. 2014). The 5-ALA phosphate salt was produced through cooling down at 0 °C with the cooling rate 15 °C/h to form the 5-ALA phosphate crystal, possessing 91.9% recovery and 99.4% purity.

ALA ion exchange process. The resin is filled in the chromatography to adsorb 5-ALA and then washed out by eluent
Although the microbial production of 5-ALA is abundant, the reports about purification and crystallization of 5-ALA are rare. During the process, the absorption of 5-ALA on resin is efficient, but the low recovery is always caused by long reaction time from desorption and crystallization. Therefore, it is still challenged to purify in acceptable amount of 5-ALA from culture broth.
The 5-ALA with 99% purity is applied in the medical field, while the purity less than 2% is adapted for environmental and agricultural fields (Table 6). The conversion of pure 5-ALA to PPIX in organisms makes it a suitable prodrug for photodynamic therapy (PDT), which is promising treatments to cancer treatment (Nordmann and Michael 2020; Shinoda et al. 2021), dental infections (Amos-Tautua et al. 2019), and condyloma acuminatum (Yang et al. 2020). Moreover, 5-ALA has also applied for the treatment to acne and actinic keratosis, because the generation of porphyrins from 5-ALA boosts the perifollicular inflammatory reaction and activates the expression of keratinocyte-derived IL-8 (Amos-Tautua et al. 2019; Meierhofer et al. 2021).
The study of antibacterial photodynamic therapy (aPDT) used the low concentration of 5-ALA to environmental issue. The accumulated porphyrins from 0.3% of 5-ALA show the different structures with various metal ions and serve as efficient photosensitizer, which destroys bacteria, including sewage bacteriophage, bacterial endospores, and sewage fecal coliforms (Amos-Tautua et al. 2019). 5-ALA is also a biodegradable insecticide as it is a promising nematicide against plant-parasitic nematodes (Cheng et al. 2017). With 1 g/L of 5-ALA (i.e., < 0.1%) and under suitable light exposure, 5-ALA is an effective growth regulator for plants. Wu et al. revealed that 5-ALA remedied the damages of photosynthetic apparatus under salinity by down-regulating the heme content and augmenting the intermediates from the chlorophyll branch (Wu et al. 2018). Based on the advantages of 5-ALA as aforementioned, Farid et al. fed 5-ALA to the sunflowers and increased chlorophyll content and protein expression (Farid et al. 2018).
Prospective and challenges
As 5-ALA is an important intermediate in the heme synthesis pathway, the accumulation would affect heme biosynthesis which is critical to aerobic metabolism and cell growth. Heme has been widely used in the healthcare and dietary supplement industries as a bioavailable iron-supplying agent (Hoppe et al. 2013). Previous studies have reported the increasing 5-ALA level also showing the positive effect on heme production. The regulation of heme synthesis pathway not only benefits 5-ALA production but also provides prospective strategies for heme accumulation. However, the extraction and purification of 5-ALA and heme from bacterium are difficult. The exporter rhtA has been widely used for 5-ALA in several studies (Kang et al. 2011a, b; Ko et al. 2019; Yang et al. 2016; Yu et al. 2020; Zhang et al. 2020b), but secretion of heme to extracellular environment was only reported by Zhao et al., using the heme exporter ccmABC (Zhao et al. 2018). Besides heme, the corrin ring of vitamin B12 is also a major tetrapyrrolic product, that is generated from 5-ALA through heme synthesis pathway, which is one of the most fascinating molecules in the medicine (Martens et al. 2002). Interestingly, the glycine riboswitch was used to control 5-ALA production as a regulation between glycine and hemB gene to heme (Zhou et al. 2019). Controlling 5-ALA accumulation is a crucial and important direction. Moreover, improving the enzymatic activity via direct evolution with optimal screening platform may accelerate the 5-ALA production in the future (Tan et al. 2020b).
The next generation of green technology is considered in low-carbon footprint and more beneficially to environment. Therefore, the development of molecular biology, genetic, metabolic engineering, or synthetic biology is rapidly extending in bioindustry (Cho et al. 2020). To reach this goal, Tan and Ng have provided a new insight to a high-efficient 5-ALA production with low carbon release (Tan and Ng 2021). In the past ten years, biosynthesis of 5-ALA in microorganisms through metabolic engineering and system biology has drawn an intensive attention due to its high efficiency, sustainability, low cost, and reaching the goal of next-generation biotechnology. Scientists have dedicated to obtaining higher 5-ALA production by a stable strain, which leads to explore various strategies for metabolic regulation and enzymatic engineering. In C4 pathway, in which the 5-ALA production is mainly affected by the expression level and activity of ALAS, the codon sequence and protein structure are critical. Therefore, the optimization of codon and the chaperons play important roles for maximizing the performance of enzyme in E. coli. On the other hand, the balancing and regulation of heme synthesis pathway are crucial for 5-ALA accumulation via C5 pathway. Several results showed that there was no significant improvement with the over-expression or down-regulation of genes on heme pathway. Notably, the origins of hemA and hemL genes used for 5-ALA production were mostly with medium or low copy number.
Exploration of enzyme with higher activity was also feasible for further enhancement of 5-ALA biosynthesis via C5 pathway. In the near future, artificial ALAS with higher activity via C4 pathway will be a promising strategy to accelerate 5-ALA production. In addition, fermentation strategies coupling the feeding of substrates and regulation of metabolic flux could be oriented to more efficient 5-ALA production. In C5 pathway, centralizing carbon flux into heme synthesis pathway is a prospective strategy; however, the metabolic regulation needs to coordinate the 5-ALA and cell growth since the heme synthesis pathway is essential in microorganisms. On the other hand, exploring more different microorganisms for 5-ALA production, especially of lactic acid bacteria which are the major human microbiome, is an alternative approach for direct usage in food (Cho et al. 2020).
5-ALA must be extracted and purified from broth for practical application, especially in medical field; however, the processes are still less described. Meanwhile, instability of 5-ALA during the process is considerable. The operating pH value and temperature may likely result in degradation. 5-ALA belongs to the class of α-amino ketones which easily dimerizes under alkaline conditions (Bunke et al. 2000), and even is unstable in the physiological environment at − 20 °C (Gadmar et al. 2002). Moreover, 5-ALA is a hydrophilic compound and not easily be permeated into cell membrane and periplasm. To solve the above problems, the derivatization of 5-ALA to increasing its lipophilicity, stability in solution, and application field is necessary.
The previous studies of 5-ALA production especially across the E. coli, C. glutamicum, and some non-typical chassis are summarized in this review. Beyond the fermentation strategies, we highlight the enzyme performance which is critical and must be optimized at the beginning. Chaperone, transporter, and chemical effects according to medium components are also mentioned. Chromosomal engineering is an alternative approach to increase 5-ALA production with higher stability and the regulation of heme biosynthesis pathway by fine-tuning the genes and promoters for 5-ALA biosynthesis are crucial. To broaden the application of 5-ALA, the purification from culture broth is necessary and still requires much improvement. As the 5-ALA and heme are critical for medical applications, more technologies and efforts which including scale-up and product storage will be considered in the future.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- 5-ALA:
-
5-Aminolevulinic acid
- ALAS:
-
5-Aminolevulinic acid synthase
- AOS:
-
α-Oxoamine synthase
- CAI:
-
Codon adaptation index
- HMW-PBPs:
-
High-molecular-weight penicillin-binding proteins
- PBG:
-
Porphobilinogen
- PLP:
-
Pyridoxal phosphate
- PPC:
-
Phosphoenolpyruvate carboxylase
- PPIX:
-
Protoporphyrin IX
- ROS:
-
Reactive oxygen species
- PDT:
-
Photodynamic therapy
- IEC:
-
Ion-exchange chromatography
References
Aiguo Z, Meizhi Z (2019) Production of 5-aminolevulinic acid from glutamate by overexpressing HemA1 and pgr7 from Arabidopsis thaliana in Escherichia coli. World J Microbiol Biotechnol 35:175
Amos-Tautua BM, Songca SP, Oluwafemi OS (2019) Application of porphyrins in antibacterial photodynamic therapy. Molecules 24:2456
Angov E (2011) Codon usage: nature’s roadmap to expression and folding of proteins. Biotechnol J 6:650–659
Anné J, Vrancken K, Van Mellaert L, Van Impe J, Bernaerts K (2014) Protein secretion biotechnology in Gram-positive bacteria with special emphasis on Streptomyces lividans. Biochim Biophys Acta Mol Cell Res 1843:1750–1761
Beale SI (1970) The biosynthesis of δ-aminolevulinic acid in Chlorella. Plant Physiol 45:504–506
Bunke A, Zerbe O, Schmid H, Burmeister G, Merkle HP, Gander B (2000) Degradation mechanism and stability of 5-aminolevulinic acid. J Pharm Sci 89:1335–1341
Burgess-Brown NA, Sharma S, Sobott F, Loenarz C, Oppermann U, Gileadi O (2008) Codon optimization can improve expression of human genes in Escherichia coli: a multi-gene study. Protein Expr Purif 59:94–102
Chen J, Wang Y, Guo X, Rao D, Zhou W, Zheng P et al (2020) Efficient bioproduction of 5-aminolevulinic acid, a promising biostimulant and nutrient, from renewable bioresources by engineered Corynebacterium glutamicum. Biotechnol Biofuels 13:1–13
Cheng F, Wang J, Song Z, Cheng JE, Zhang D, Liu Y (2017) Nematicidal effects of 5-aminolevulinic acid on plant-parasitic nematodes. J Nematol 49:295
Cho SW, Yim J, Seo SW (2020) Engineering tools for the development of recombinant lactic acid bacteria. Biotechnol J 15:e1900344
Choi C, Hong BS, Sung HC, Lee HS, Kim JH (1999) Optimization of extracellular 5-aminolevulinic acid production from Escherichia coli transformed with ALA synthase gene of Bradyrhizobium japonicum. Biotechnol Lett 21:551–554
Choi HP, Lee YM, Yun CW, Sung HC (2008) Extracellular 5-aminolevulinic acid production by Escherichia coli containing the Rhodopseudomonas palustris KUGB306 hemA gene. J Microbiol Biotechnol 18:1136–1140
Chung SY, Seo KH, Rhee JI (2005) Influence of culture conditions on the production of extra-cellular 5-aminolevulinic acid (ALA) by recombinant E. coli. Process Biochem 40:385–394
Cui Z, Jiang Z, Zhang J, Zheng H, Jiang X, Gong K et al (2019) Stable and efficient biosynthesis of 5-aminolevulinic acid using plasmid-free Escherichia coli. J Agric Food Chem 67:1478–1483
de Marco A, Deuerling E, Mogk A, Tomoyasu T, Bukau B (2007) Chaperone-based procedure to increase yields of soluble recombinant proteins produced in E. coli. BMC Biotechnol 7:32
Diesveld R, Tietze N, Fürst O, Reth A, Bathe B, Sahm H, Eggeling L (2009) Activity of exporters of Escherichia coli in Corynebacterium glutamicum, and their use to increase L-threonine production. J Mol Microbiol Biotechnol 16:198–207
Ding W, Weng H, Du G, Chen J, Kang Z (2017) 5-Aminolevulinic acid production from inexpensive glucose by engineering the C4 pathway in Escherichia coli. J Ind Microbiol Biotechnol 44:1127–1135
Dolmans DE, Fukumura D, Jain RK (2003) Photodynamic therapy for cancer. Nat Rev Cancer 3:380–387
Effendi SSW, Tan SI, Chang CH, Chen CY, Chang JS, Ng IS (2020) Development and fabrication of disease resistance protein in recombinant Escherichia coli. Bioresour Bioprocess 7:1–10
Erskine PT, Norton E, Cooper JB, Lambert R, Coker A, Lewis G et al (1999) X-ray structure of 5-aminolevulinic acid dehydratase from Escherichia coli complexed with the inhibitor levulinic acid at 2.0 Å resolution. Biochemistry 38:4266–4276
Farid M, Ali S, Rizwan M, Ali Q, Saeed R, Nasir T et al (2018) Phyto-management of chromium contaminated soils through sunflower under exogenously applied 5-aminolevulinic acid. Ecotoxicol Environ Saf 151:255–265
Feng L, Zhang Y, Fu J, Mao Y, Chen T, Zhao X, Wang Z (2016) Metabolic engineering of Corynebacterium glutamicum for efficient production of 5-aminolevulinic acid. Biotechnol Bioeng 113:1284–1293
Fu W, Lin J, Cen P (2007) 5-Aminolevulinate production with recombinant Escherichia coli using a rare codon optimizer host strain. Appl Microbiol Biotechnol 75:777–782
Fu W, Lin J, Cen P (2010) Expression of a hemA gene from Agrobacterium radiobacter in a rare codon optimizing Escherichia coli for improving 5-aminolevulinate production. Appl Biochem Biotechnol 160:456–466
Fuglsang A (2003) Codon optimizer: a freeware tool for codon optimization. Protein Expr Purif 31:247–249
Gadmar ØB, Moan J, Scheie E, Ma LW, Peng Q (2002) The stability of 5-aminolevulinic acid in solution. J Photochem Photobiol B 67:187–193
Hara KY, Saito M, Kato H, Morikawa K, Kikukawa H, Nomura H et al (2019) 5-Aminolevulinic acid fermentation using engineered Saccharomyces cerevisiae. Microb Cell Fact 18:1–8
Hayer-Hartl M, Bracher A, Hartl FU (2016) The GroEL–GroES chaperonin machine: a nano-cage for protein folding. Trends Biochem Sci 41:62–76
Hoppe M, Brün B, Larsson MP, Moraeus L, Hulthén L (2013) Heme iron-based dietary intervention for improvement of iron status in young women. Nutr 29:89–95
Hotta Y, Tanaka T, Takaoka H, Takeuchi Y, Konnai M (1997) Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant Growth Regul 22:109–114
Huang DD, Wang WY (1986) Chlorophyll biosynthesis in Chlamydomonas starts with the formation of glutamyl-tRNA. Int J Biol Chem 261:13451–13455
Jahn D, Verkamp E, So D (1992) Glutamyl-transfer RNA:a precursor of heme and chlorophyll biosynthesis. Trends Biochem Sci 17:215–218
Jones JA, Toparlak ÖD, Koffas MA (2015) Metabolic pathway balancing and its role in the production of biofuels and chemicals. Curr Opin Biotechnol 33:52–59
Jung S, Yang K, Lee DE, Back K (2004) Expression of Bradyrhizobium japonicum 5-aminolevulinic acid synthase induces severe photodynamic damage in transgenic rice. Plant Sci 167:789–795
Kane JF (1995) Effects of rare codon clusters on high-level expression of heterologous proteins in Escherichia coli. Curr Opin Biotechnol 6:494–500
Kang DK, Kim SS, Chi WJ, Hong SK, Kim HK, Kim HU (2004) Cloning and expression of the Rhodobacter capsulatus hemA gene in E. coli for the production of 5-aminolevulinic acid. J Microbiol Biotechnol 14:1327–1332
Kang Z, Wang Y, Gu P, Wang Q, Qi Q (2011a) Engineering Escherichia coli for efficient production of 5-aminolevulinic acid from glucose. Metab Eng 13:492–498
Kang Z, Wang Y, Wang Q, Qi Q (2011b) Metabolic engineering to improve 5-aminolevulinic acid production. Bioeng Bugs 2:342–345
Kang Z, Ding W, Gong X, Liu Q, Du G, Chen J (2017) Recent advances in production of 5-aminolevulinic acid using biological strategies. World J Microbiol Biotechnol 33:200
Kennedy J, Pottier RH, Pross DC (1990) Photodynamic therapy with endogenous protoporphyrin: IX: basic principles and present clinical experience. J Photochem Photobiol B 6:143–148
Ko YJ, You SK, Kim M, Lee E, Shin SK, Park HM et al (2019) Enhanced production of 5-aminolevulinic acid via flux redistribution of tca cycle toward L-glutamate in Corynebacterium glutamicum. Biotechnol Bioprocess Eng 24:915–923
Krieg RC, Messmann H, Rauch J, Seeger S, Knuechel R (2002) Metabolic characterization of tumor cell–specific protoporphyrin IX accumulation after exposure to 5-aminolevulinic acid in human colonic cells. Photochem Photobiol 76:518–525
Kwon SJ, De Boer AL, Petri R, Schmidt-Dannert C (2003) High-level production of porphyrins in metabolically engineered Escherichia coli: systematic extension of a pathway assembled from overexpressed genes involved in heme biosynthesis. Appl Environ Microbiol 69:4875–4883
Li JM, Russell CS, Cosloy SD (1989) Cloning and structure of the hemA gene of Escherichia coli K-12. Gene 82:209–217
Li F, Wang Y, Gong K, Wang Q, Liang Q, Qi Q (2014) Constitutive expression of RyhB regulates the heme biosynthesis pathway and increases the 5-aminolevulinic acid accumulation in Escherichia coli. FEMS Microbiol Lett 350:209–215
Li T, Guo YY, Qiao GQ, Chen GQ (2016) Microbial synthesis of 5-aminolevulinic acid and its coproduction with polyhydroxybutyrate. ACS Synth Biol 5:1264–1274
Lin J, Fu W, Cen P (2009) Characterization of 5-aminolevulinate synthase from Agrobacterium radiobacter, screening new inhibitors for 5-aminolevulinate dehydratase from Escherichia coli and their potential use for high 5-aminolevulinate production. Bioresour Technol 100:2293–2297
Lin J, Lou J, Cen P (2014) Crystallization method of 5-aminolevulinic acid phosphate. CN103265444B, 5 November 2014.
Liu D, Wu L, Naeem MS, Liu H, Deng X, Xu L et al (2013) 5-Aminolevulinic acid enhances photosynthetic gas exchange, chlorophyll fluorescence and antioxidant system in oilseed rape under drought stress. Acta Physiol Plant 35:2747–2759
Liu S, Zhang G, Li X, Zhang J (2014) Microbial production and applications of 5-aminolevulinic acid. Appl Microbiol Biotechnol 98:7349–7357
Liu S, Li X, Zhang G, Zhang J (2015) Optimization of influencing factors on biomass accumulation and 5-aminolevulinic acid (ALA) yield in Rhodobacter sphaeroides wastewater treatment. J Microbiol Biotechnol 25:1920–1927
Liu S, Zhang G, Li J, Li X, Zhang J (2016) Effects of metal ions on biomass and 5-aminolevulinic acid production in Rhodopseudomonas palustris wastewater treatment. Water Sci Technol 73:382–388
Liu S, Zheng Z, Tie J, Kang J, Zhang G, Zhang J (2018) Impacts of Fe2+ on 5-aminolevulinic acid (ALA) biosynthesis of Rhodobacter sphaeroides in wastewater treatment by regulating nif gene expression. Res J Environ Sci 70:11–19
Liu J, Ye Z, Wu H, Liu J, Gong Y (2020) Overexpression of hemA and hemL in Bacillus subtilis promotes overexpression of 5-aminolevulinic acid. Indian J Anim Res 54.
Lou JW, Zhu L, Wu MB, Yang LR, Lin JP, Cen PL (2014) High-level soluble expression of the hemA gene from Rhodobacter capsulatus and comparative study of its enzymatic properties. J Zhejiang Univ Sci B 15:491–499
Lüer C, Schauer S, Möbius K, Schulze J, Schubert WD, Heinz DW et al (2005) Complex formation between glutamyl-tRNA reductase and glutamate-1-semialdehyde 2, 1-aminomutase in Escherichia coli during the initial reactions of porphyrin biosynthesis. Int J Biol Chem 280:18568–18572
Malik Z, Lugaci H (1987) Destruction of erythroleukaemic cells by photoactivation of endogenous porphyrins. Br J Cancer 56:589
Mao Y, Chen Z, Lu L, Jin B, Ma H, Pan Y, Chen T (2020) Efficient solid-state fermentation for the production of 5-aminolevulinic acid enriched feed using recombinant Saccharomyces cerevisiae. J Biotechnol 322:29–32
Martens JH, Barg H, Warren MA, Jahn D (2002) Microbial production of vitamin B12. Appl Microbiol Biotechnol 58:275–285
Mayfield JA, Dehner CA, DuBois JL (2011) Recent advances in bacterial heme protein biochemistry. Curr Opin Chem Biol 15:260–266
McNicholas PM, Javor G, Darie S, Gunsalus RP (1997) Expression of the heme biosynthetic pathway genes hemCD, hemH, hemM and hemA of Escherichia coli. FEMS Microbiol Lett 146:143–148
Meierhofer C, Silic K, Urban MV, Tanew A, Radakovic S (2021) The impact of occlusive vs non-occlusive application of 5-aminolevulinic acid (BF-200 ALA) on the efficacy and tolerability of photodynamic therapy for actinic keratosis on the scalp and face: a prospective within-patient comparison trial. Photodermatol Photoimmunol Photomed 37:56–62
Meng Q, Zhang Y, Ma C, Ma H, Zhao X, Chen T (2015) Purification and functional characterization of thermostable 5-aminolevulinic acid synthases. Biotechnol Lett 37:2247–2253
Menzella HG (2011) Comparison of two codon optimization strategies to enhance recombinant protein production in Escherichia coli. Microb Cell Fact 10:15
Miscevic D, Mao JY, Kefale T, Abedi D, Moo-Young M, Chou CP (2020) Strain engineering for high-level 5-aminolevulinic acid production in Escherichia coli. Biotechnol Bioeng. https://doi.org/10.1002/bit.27547
Naeem MS, Warusawitharana H, Liu H, Liu D, Ahmad R, Waraich EA et al (2012) 5-Aminolevulinic acid alleviates the salinity-induced changes in Brassica napus as revealed by the ultrastructural study of chloroplast. Plant Physiol Biochem 57:84–92
Nakakuki M, Yamauchi K, Hayashi N, Kikuchi G (1980) Purification and some properties of delta-aminolevulinate synthase from the rat liver cytosol fraction and immunochemical identity of the cytosolic enzyme and the mitochondrial enzyme. J Biol Chem 255:1738–1745
Nandi DL, Baker-Cohen KF, Shemin D (1968) δ-Aminolevulinic acid dehydratase of Rhodopseudomonas spheroides I. Isolation and properties. Int J Biol Chem 243:1224–1230
Noh MH, Lim HG, Park S, Seo SW, Jung GY (2017) Precise flux redistribution to glyoxylate cycle for 5-aminolevulinic acid production in Escherichia coli. Metab Eng 43:1–8
Nordmann NJ, Michael AP (2020) 5-Aminolevulinic acid radiodynamic therapy for treatment of high-grade gliomas: systematic review. Clin Neurol Neurosurg. 201:106430
Okada H, Tanaka T, Nomura T (2016) Process for producing 5-aminolevulinic acid hydrochloride. EP1927586B1, 27 April 2016.
Ong PY, Lee CT, Sarmidi MR, Awad HM, Chua LS, Razali F (2013) Production of extracellular 5-aminolevulinic acid by Rhodopseudomonas palustris in solid-state fermentation. Developments in sustainable chemical and bioprocess technology. Springer, Boston, pp 173–179
Peng Q, Warloe T, Berg K, Moan J, Kongshaug M, Giercksky KE, Nesland JM (1997a) 5-Aminolevulinic acid-based photodynamic therapy: clinical research and future challenges. Cancer 79:2282–2308
Peng Q, Berg K, Moan J, Kongshaug M, Nesland JM (1997b) 5-Aminolevulinic acid-based photodynamic therapy: principles and experimental research. Photochem Photobiol 65:235–251
Ramzi AB, Hyeon JE, Kim SW, Park C, Han SO (2015) 5-Aminolevulinic acid production in engineered Corynebacterium glutamicum via C5 biosynthesis pathway. Enzyme Microb Technol 81:1–7
Sasaki K, Watanabe M, Tanaka T (2002) Biosynthesis, biotechnological production and applications of 5-aminolevulinic acid. Appl Microbiol Biotechnol 58:23–29
Sasikala C, Ramana CV, Rao PR (1994) 5-aminolevulinic acid: a potential herbicide/insecticide from microorganisms. Biotechnol Prog 10:451–459
Sato K, Ishida K, Mutsushika O, Shimizu S (1985) Purification and some properties of δ-aminolevulinic acid synthases from Protaminobacter ruber and Rhodopseudomonas spheroides. Agric Biol Chem 49:3415–3421
Schauer S, Chaturvedi S, Randau L, Moser J, Kitabatake M, Lorenz S et al (2002) Escherichia coli glutamyl-tRNA reductase trapping the thioester intermediate. Int J Biol Chem 277:48657–48663
Schneegurt MA, Beale SI (1988) Characterization of the RNA required for biosynthesis of δ-aminolevulinic acid from glutamate: purification by anticodon-based affinity chromatography and determination that the UUC glutamate anticodon is a general requirement for function in ALA biosynthesis. Plant Physiol 86:497–504
Shih IT, Yi YC, Ng IS (2021) Plasmid-Free System and modular design for efficient 5-aminolevulinic acid production by engineered Escherichia coli. Appl Biochem Biotechnol 193:1–14
Shinoda Y, Kato D, Ando R, Endo H, Takahashi T, Tsuneoka Y, Fujiwara Y (2021) Systematic review and meta-analysis of in vitro anti-human cancer experiments investigating the use of 5-aminolevulinic acid (5-ALA) for photodynamic therapy. Pharmaceuticals 14:229
Sørensen HP, Mortensen KK (2005) Advanced genetic strategies for recombinant protein expression in Escherichia coli. J Biotechnol 115:113–128
Stojanovski BM, Ferreira GC (2015) Asn-150 of murine erythroid 5-aminolevulinate synthase modulates the catalytic balance between the rates of the reversible reaction. J Biol Chem 290:30750–30761
Stojanovski BM, Hunter GA, Jahn M, Jahn D, Ferreira GC (2014) Unstable reaction intermediates and hysteresis during the catalytic cycle of 5-aminolevulinate synthase implications from using pseudo and alternate substrates and a promiscuous enzyme variant. J Biol Chem 289:22915–22925
Su T, Guo Q, Zheng Y, Liang Q, Wang Q, Qi Q (2019) Fine-tuning of hemB using CRISPRi for increasing 5-aminolevulinic acid production in Escherichia coli. Front Microbiol 10:1731
Tan SI, Ng IS (2021) Stepwise optimization of genetic RuBisCO-equipped Escherichia coli for low carbon-footprint protein and chemical production. Green Chem. https://doi.org/10.1039/D1GC00456E
Tan SI, You SC, Shih IT, Ng IS (2020a) Quantification, regulation and production of 5-aminolevulinic acid by green fluorescent protein in recombinant Escherichia coli. J Biosci Bioeng 129:387–394
Tan SI, Yu PJ, Ng IS (2020b) CRISPRi-mediated programming essential gene can as a direct enzymatic performance evaluation & determination (DEPEND) system. Biotechnol Bioeng 117:2842–2851
Tangprasittipap A, Prasertsan P, Choorit W, Sasaki K (2007) Biosynthesis of intracellular 5-aminolevulinic acid by a newly identified halotolerant Rhodobacter sphaeroides. Biotechnol Lett 29:773–778
Tran NT, Pham DN, Kim CJ (2019) Production of 5-aminolevulinic acid by recombinant Streptomyces coelicolor expressing hemA from Rhodobacter sphaeroides. Biotechnol Bioprocess Eng 24:488–499
Tripetch P, Srzednicki G, Borompichaichartkul C (2013) Separation process of 5-aminolevulinic acid from Rhodobacter spaeroides for increasing value of agricultural product by ion exchange chromatography. Acta Hortic 1011:265–271
Van der Werf MJ, Zeikus JG (1996) 5-Aminolevulinate production by Escherichia coli containing the Rhodobacter sphaeroides hemA gene. Appl Environ Microbiol 62:3560–3566
Venosa DG, Fukuda H, Perotti C, Batlle A, Casas A (2004) A method for separating ALA from ALA derivatives using ionic exchange extraction. J Photochem Photobiol B: Biol 75:157–163
Verderber E, Lucast LJ, Van Dehy JA, Cozart P, Etter JB, Best EA (1997) Role of the hemA gene product and delta-aminolevulinic acid in regulation of Escherichia coli heme synthesis. J Bacteriol Res 179:4583–4590
Volland C, Felix F (1984) Isolation and properties of 5-aminolevulinate synthase from the yeast Saccharomyces cerevisiae. Eur J Biochem 142:551–557
Wang WY, Huang DD, Stachon D, Gough SP, Kannangara CG (1984) Purification, characterization, and fractionation of the δ-aminolevulinic acid synthesizing enzymes from light-grown Chlamydomonas reinhardtii cells. Plant Physiol 74:569–575
Wild PJ, Krieg RC, Seidl J, Stoehr R, Reher K, Hofmann C et al (2005) RNA expression profiling of normal and tumor cells following photodynamic therapy with 5-aminolevulinic acid–induced protoporphyrin IX in vitro. Mol Cancer Ther 4:516–528
Wu Y, Jin X, Liao W, Hu L, Dawuda MM, Zhao X et al (2018) 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front Plant Sci 9:635
Xie L, Hall D, Eiteman MA, Altman E (2003) Optimization of recombinant aminolevulinate synthase production in Escherichia coli using factorial design. Appl Microbiol Biotechnol 63:267–273
Xue C, Yu TH, Ng IS (2021) Engineering pyridoxal kinase PdxY-integrated Escherichia coli strain and optimization for high-level 5-aminolevulinic acid production. J Taiwan Inst Chem Eng 120:49–58
Yang J, Li Z, Fu W, Lin Y, Lin J, Cen P (2013) Improved 5-aminolevulinic acid production with recombinant Escherichia coli by a short-term dissolved oxygen shock in fed-batch fermentation. Chin J Chem Eng 21:1291–1295
Yang X, Palasuberniam P, Kraus D, Chen B (2015) Aminolevulinic acid-based tumor detection and therapy: molecular mechanisms and strategies for enhancement. Int J Mol Sci 16:25865–25880
Yang P, Liu W, Cheng X, Wang J, Wang Q, Qi Q (2016) A new strategy for production of 5-aminolevulinic acid in recombinant Corynebacterium glutamicum with high yield. Appl Environ Microbiol 82:2709–2717
Yang HJ, Lee KH, Lim HJ, Kim DM (2019) Tandem cell-free protein synthesis as a tool for rapid screening of optimal molecular chaperones. Biotechnol J 14:e1800523
Yang Y, Zhang Y, Zou X, Guo X, Lin H (2019b) Perspective clinical study on effect of 5-aminolevulinic acid photodynamic therapy (ALA-PDT) in treating condylomata acuminata in pregnancy. Photodiagn Photodyn Ther 25:63–65
Yang D, Park SY, Park YS, Eun H, Lee SY (2020) Metabolic engineering of Escherichia coli for natural product biosynthesis. Trends Biotechnol 38:745–765
Yi YC, Ng IS (2020) Establishment of toolkit and T7RNA polymerase/promoter system in Shewanella oneidensis MR-1. J Taiwan Inst Chem Eng 109:8–14
Yi YC, Ng IS (2021) Redirection of metabolic flux in Shewanella oneidensis MR-1 by CRISPRi and modular design for 5-aminolevulinic acid production. Bioresour Bioprocess 8:1–11
Yu X, Jin H, Liu W, Wang Q, Qi Q (2015) Engineering Corynebacterium glutamicum to produce 5-aminolevulinic acid from glucose. Microb Cell Fact 14:183
Yu TH, Yi YC, Shih IT, Ng IS (2020) Enhanced 5-aminolevulinic acid production by co-expression of codon-optimized hemA gene with chaperone in genetic engineered Escherichia coli. Appl Biochem Biotechnol 191:299–312
Zhang L, Chen J, Chen N, Sun J, Zheng P, Ma Y (2013) Cloning of two 5-aminolevulinic acid synthase isozymes HemA and HemO from Rhodopseudomonas palustris with favorable characteristics for 5-aminolevulinic acid production. Biotechnol Lett 35:763–768
Zhang J, Kang Z, Chen J, Du G (2015) Optimization of the heme biosynthesis pathway for the production of 5-aminolevulinic acid in Escherichia coli. Sci Rep 5:8584
Zhang J, Kang Z, Ding W, Chen J, Du G (2016) Integrated optimization of the in vivo heme biosynthesis pathway and the in vitro iron concentration for 5-aminolevulinate production. Appl Biochem 178:1252–1262
Zhang J, Weng H, Ding W, Kang Z (2017) N-terminal engineering of glutamyl-tRNA reductase with positive charge arginine to increase 5-aminolevulinic acid biosynthesis. Bioengineered 8:424–427
Zhang J, Rang Z, Qian S, Qiu L, Chen J, Du G (2018a) Construction of recombinant Saccharomyces cerevisiae for production of 5-aminolevulinic acid. J Food Sci Biotechnol 37:232–239
Zhang X, Zhang J, Xu J, Zhao Q, Wang Q, Qi Q (2018b) Engineering Escherichia coli for efficient coproduction of polyhydroxyalkanoates and 5-aminolevulinic acid. J Ind Microbiol Biotechnol 45:43–51
Zhang J, Weng H, Zhou Z, Du G, Kang Z (2019) Engineering of multiple modular pathways for high-yield production of 5-aminolevulinic acid in Escherichia coli. Bioresour Technol 274:353–360
Zhang B, Ye BC (2018) Pathway engineering in Corynebacterium glutamicum S9114 for 5-aminolevulinic acid production. 3 Biotech 8:247
Zhang J, Wang Z, Su T, Sun H, Zhu Y, Qi Q, Wang Q (2020) Tuning the binding affinity of heme-responsive biosensor for precise and dynamic pathway regulation. iScience 23(5):101067
Zhang C, Li Y, Zhu F, Li Z, Lu N, Li Y et al (2020) Metabolic engineering of an auto-regulated Corynebacterium glutamicum chassis for biosynthesis of 5-aminolevulinic acid. Bioresour Technol 318:124064
Zhao XR, Choi KR, Lee SY (2018) Metabolic engineering of Escherichia coli for secretory production of free haem. Nat Catal 1:720–728
Zhou L, Ren J, Li Z, Nie J, Wang C, Zeng AP (2019) Characterization and engineering of a Clostridium glycine riboswitch and its use to control a novel metabolic pathway for 5-aminolevulinic acid production in Escherichia coli. ACS Synth Biol 8:2327–2335
Zhu C, Chen J, Wang Y, Wang L, Guo X, Chen N et al (2019) Enhancing 5-aminolevulinic acid tolerance and production by engineering the antioxidant defense system of Escherichia coli. Biotechnol Bioeng 116:2018–2028
Zhu Y, Zhou C, Wang Y, Li C (2020) Transporter engineering for microbial manufacturing. Biotechnol J. 15:1900494
Zou Y, Chen T, Feng L, Zhang S, Xing D, Wang Z (2017) Enhancement of 5-aminolevulinic acid production by metabolic engineering of the glycine biosynthesis pathway in Corynebacterium glutamicum. Biotechnol Lett 39:1369–1374
Acknowledgements
The authors are grateful to the funding support received from the Ministry of Science and Technology in Taiwan.
Funding
The authors are grateful to the financial support received from the Ministry of Science and Technology in Taiwan (MOST 108-2221-E-006-004-MY3 and MOST 110-2221-E-006-030-MY3).
Author information
Authors and Affiliations
Contributions
Y-CY, I-TS, T-HY, and Y-JL prepared the draft and I-SN edited, supervised, and reviewed the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All the authors have read and agreed the ethics for publishing the manuscript.
Consent for publication
The authors approved the consent for publishing the manuscript.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yi, YC., Shih, IT., Yu, TH. et al. Challenges and opportunities of bioprocessing 5-aminolevulinic acid using genetic and metabolic engineering: a critical review. Bioresour. Bioprocess. 8, 100 (2021). https://doi.org/10.1186/s40643-021-00455-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-021-00455-6