
Abstract
Gram-negative bacteria are known to subvert eukaryotic cell physiological mechanisms using a wide array of virulence factors, among which the type three-secretion system (T3SS) is often one of the most important. The T3SS constitutes a needle-like apparatus that the bacterium uses to inject a diverse set of effector proteins directly into the cytoplasm of the host cells where they can hamper the host cellular machinery for a variety of purposes. While the structure of the T3SS is somewhat conserved and well described, effector proteins are much more diverse and specific for each pathogen. The T3SS can remodel the cytoskeleton integrity to promote intracellular invasion, as well as silence specific eukaryotic cell signals, notably to hinder or elude the immune response and cause apoptosis. This is also the case in aquatic bacterial pathogens where the T3SS can often play a central role in the establishment of disease, although it remains understudied in several species of important fish pathogens, notably in Yersinia ruckeri. In the present review, we summarise what is known of the T3SS, with a special focus on aquatic pathogens and suggest some possible avenues for research including the potential to target the T3SS for the development of new anti-virulence drugs.
Similar content being viewed by others

1 Introduction
The demand for fish products has been increasing worldwide for multiple decades now, and with the output of fisheries largely stagnant, this demand has been mostly answered through the growth of aquaculture, particularly in developing countries [1]. Aquaculture has become the fastest growing animal food production sector [2, 3] and it is expected to almost double to reach 93.2 million tons in the next decade [4]. This increased production has been in part achieved by increasing the intensity of fish farming, which has led to an increase in the occurrence of infectious diseases, which represent a major limiting factor in aquaculture [5, 6].
Pathogenic bacteria, in particular gram-negative bacteria, are a major group among the pathogens associated with diseases in aquatic organisms and have evolved multiple features to influence their hosts and defend themselves against attackers. Bacteria are separated from their environment by a complex multi-layered envelope. In gram-negative species, this envelope is formed of three layers: the inner-membrane which is composed of a phospholipid bilayer, a thin peptidoglycan cell wall composed of repeating units of N-acetyl glucosamine and N-actyl muramic acid and an outer-membrane, a feature unique to gram-negative bacteria, composed of phospholipids and lipopolysaccharides [7]. The space between the inner and outer-membrane where the cell wall is located is termed the periplasm and can represent a large proportion, up to 40%, of the total volume of the bacterial cells [8]. In addition, many bacteria also secrete a glycocalyx in the form of capsules or slime-layers.
As the main role of this complex envelope is to separate the bacterial cytoplasm from the external environment, it is largely impermeable and movement of non-soluble compounds requires complex transport mechanisms. Notably, virulence factors are by their very definition active outside the bacterial cells and are therefore transferred through specific transport system from the bacterial cytoplasm toward the outer side of the envelope [9]. Several secretion systems have been linked to the export of virulence factors and classified into broad families, forming an ever-expanding list of numbered secretion systems that currently includes at least nine different types [10, 11], although more are very likely to be described in the future. Of course, not all types of secretion systems are present in all bacterial species. For example, the type 7 secretion system is mostly associated with members of the Mycobacterium genus and other gram-positive bacteria and no functional members of this family have yet been described in any species of gram-negative bacteria [9, 12].
Among these secretion systems, the best studied, or at least the one associated with the highest number of references in the PubMed database at the time of this writing, is the type 3 secretion system (T3SS). This interest can be explained by the important role of the T3SS in the virulence of many varied bacterial pathogens, including many important pathogens of fish and other aquatic animals.
Because of the importance of the T3SS in the establishment of disease by Gram-negative pathogens, an understanding of their mode of action is necessary for the understanding of many diseases. Moreover, they represent attractive targets in the development of anti-virulence therapy. In aquatic organisms notably, bacterial pathogens have been associated with several outbreaks and much of the bacterial species involved belong to the group of the gram negative, with several known to harbor a T3SS that is often required for the establishment of infections. However, despite this importance, much remains to be learned about the T3SS of many aquatic pathogens. Therefore, the present review was written with the goal to present an overview of what is currently known about the T3SS and list some of the areas still requiring more attention, with the hope that it might help further research on the subject. Since most of the research regarding the T3SS was conducted on pathogens of land animals, the present review will start with an overview of what is generally known about these T3SS, first in term of structure and then regarding their functions and effector proteins. Then, it will focus on what has been discovered specifically about the role of the T3SS in the establishment of diseases in aquatic organisms.
2 The T3SS
2.1 Structure of the Type III secretion system
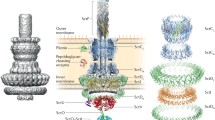
The T3SS either evolved from the bacterial flagellum or shares a direct relative with it, consequently, many structural proteins are homologous between the two apparatus [13,14,15,16]. While there are multiple distinct families of T3SS (Figure 1), the Ysc secretion system of the Yersinia genus is generally used as the model for our understanding of the T3SS. Moreover, while many species of bacteria share genes homologous to that belonging to the Ysc T3SS, they are often more closely related to T3SS from other genera (Additional file 1). Based on these similarities, seven categories (or families) of T3SS have been defined and most of the T3SS belong to one of these seven families [16]. In recent years, because of the presence of a number of homologous proteins, an effort has been conducted to standardise the names of the various proteins comprising the T3SS following a universal nomenclature based on homology [15, 17, 18]. In this nomenclature, the first 3 letters reflect the T3SS from which the protein originates, while the last letter, written in capital, indicates the specific protein and is identical between homologs. For example, the translocation protein AscE of Aeromonas hydrophila is homologous with the translocation proteins YscE in Yersinia sp. and SctE encoded by the pathogenicity island 1 (SPI-1) of Salmonella [19]. This is the terminology that we will apply in the next paragraphs.

(reproduced with permission from Troisfontaines and Cornelis [16]).
Phylogenetic tree of the best characterized T3SS families. The tree was prepared using aligned sequences from the Ribosomal Database Project II and the various sequences were allocated into 7 major families of T3SS based on homology
One way to categorise the different proteins composing the T3SS is between conserved and variable proteins. The former is represented by a set of about 15 to 20 conserved proteins [20], mostly playing a structural role, which are homologous between the different organisms harbouring a T3SS [18]. They are generally present in all T3SS and, in a way, define what constitutes this apparatus (Table 1). The second category of proteins is more variable and adapted to the specific bacterium and the role the T3SS play in its lifestyle. Specific effector proteins and corresponding chaperones are therefore most represented in this category.
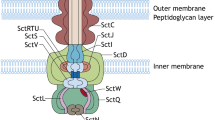
Another way to characterise this complex apparatus is based on the location of the different proteins. This allows dividing the T3SS into three regions: the cytoplasmic region, the trans-membrane region, also known as the basal body [21], and the extra-cellular region (Figure 2). The main role of the cytoplasmic region is to interact with proteins in the bacterial cytoplasm and to recruit effector proteins before directing them to the rest of the secretory apparatus. Consequently, this section of the T3SS is often characterised as the “sorting platform” or at the “export apparatus” of the T3SS [22]. This region is organised around a polymer formed by the protein YscV which complexes with the proteins YscR, YscS, and YscT [23]. In addition, YscQ, a homolog to the flagellar Motor C-ring is generally considered part of the export apparatus [24]. Energy for the protein export is provided through the ATPase YscN, which forms a hexameric ring and is regulated through YscL [25]. The sorting platform recognises specific substrate to be exported via the T3SS. These substrates are marked by a leader sequence followed by a specific translocation signal, however these are highly variable and ill-defined: unstructured N-terminal sequence appear common in all T3SS substrates but the length of these sequence vary greatly between molecules and some classes of proteins require additional signals to be recognised by the T3SS [26].

Schematic of a typical T3SS. A Schematic of the use of the bacterial type 3 secretion system. B Common organization and structure of the T3SS. The colors of the proteins correspond to the various areas of the T3SS. C Nomenclature of T3SS components displaying both the universal nomenclature and the corresponding protein on the SPI-1 T3SS. Reproduced with permission and minor modifications from Wagner et al. [20].
The trans-membrane region is composed of two rings forming an export channel through the inner and outer bacterial membranes. The channel in the inner membrane is composed of two proteins and is considered homologous to the flagellar M-ring. YscJ is a lipoprotein, located on the periplasmic side of the inner-membrane and co-purified with YscD [27, 28], which has both cytoplasmic and periplasmic domains [29]. Mutation of the forkhead-associated-domain of YscD was shown to result in silencing of the T3SS in Yersinia pseudotuberculosis [29]. YscC belongs to the family of secretin-proteins, it protrudes into the periplasm and forms a pore in the bacterial outer-membrane allowing for protein export into the rest of the T3SS [30]. It is theorised, based on observation performed on T3SS component proteins tagged with fluorescent proteins, that it is the insertion of YsC into the bacterial outer-membrane by the pilotin YscW that initiates assembly of the T3SS, at least as far as the YsC family is concerned, and that the rest of the T3SS is built around YscC [28, 31]. The role of the trans-membrane region is to transport the effector proteins across the bacterial cell membranes. In addition, it is also involved in the secretion of the extra-cellular components of the T3SS on the other side of the cell membranes where they will form the extra-cellular part of the T3SS [32].
Finally, the extra-cellular region can be further separated between the needle that bridges the extra-cellular space between the bacterial and the host cell membrane and the tip and translocon of the T3SS that are inserted into the host cell to act as the injection apparatus and move the effector proteins across the cell membrane. The needle is very conserved and composed of hundreds of repeats of the protein YscF [33]. The tip of the T3SS is formed by the protein LcrV, also known as the V antigen, a polymorphic protein which is absent in many T3SS families and whose function remains unclear [34, 35], although it has been suggested that it plays a role in manipulating the TLR2 mediated immune response. In addition, it is known that YopD/LcrV interactions play a role in the secretion through the T3SS [36] and LcrV are required for the insertion of the other two proteins, YopB and YopD [37]. These two proteins are called translocators and together, they form an integral multimeric 500 to 700 kDa complex referred to as the translocon that acts as a pore into the host membrane, allowing for the delivery of effector proteins directly into the cytoplasm [32, 38, 39]. For this reason, the T3SS is sometime called the “injectisome” [20].
As mentioned above, this description is based on our understanding of the Ysc secretion system, which is likely the best characterised of all T3SS and, while the basic structure is conserved, other T3SS families exist that differs from this model in some respect. For example, the assembly sequences for the basal body and the export apparatus differ between the Salmonella Spa and the Yersinia YsC secretion systems [40].
2.2 Chaperones of the T3SS
Alongside these structural components, another critical part of the T3SS is the associated chaperones. These chaperones are generally small amphiphilic molecules [41] and are often encoded adjacent to the effector proteins that they serve. They are generally not secreted themselves but may be necessary for secretion and therefore for the T3SS to perform its function [42]. Chaperones can play multiple roles but are mostly known to direct the correct folding of proteins and protect them from untimely degradation, oligomerization and unwanted interactions with proteins in the bacterial membrane [43]. For example, the Inv protein in Salmonella directs the folding of cognate secreted proteins in an ATP-dependent manner [42]. A major role of chaperones is to direct their substrate toward the ATPase of the T3SS that will propel them through the T3SS [42, 44]. Chaperones of the T3SS are generally divided within three classes based on their substrate with class I chaperones specialising in the translocation of effector proteins, class II chaperones translocating proteins that assemble to form the secretion pore in the host target membrane, and class III chaperones specialising in the secretion of extracellular filamentous proteins [42, 45].
In enteropathogenic Escherichia coli, co-immunoprecipitation assays have shown that the chaperone CesT interacts with both the translocated intimin receptor (Tir) and the Mitochondrial-associated protein (Map). Both gel overlays following migration on SDS-PAGE and enzyme-linked immunosorbent assays have shown that Tir binds to the type III ATPase EscN via CesT [46] which allows for its subsequent secretion through the T3SS [47]. Similarly, crystallography studies have shown that SycN–YscB form a heterodimeric chaperone that permits the secretion of YopN in Yersinia pestis [48]. Moreover, LcrH plays a role in controlling the levels of secretion of Yop and YopD [49, 50].
An important family of chaperones is the SycE family in the T3SS of members of the Yersinia genus [51, 52]. This family includes Syct, a chaperone that harbours a binding site containing residues 52–103 of YopT and is required for the secretion of YopT in Yersinia enterocolitica [53]. Three additional chaperones have been described for the Yop of Y. enterocolitica, SycE, SycH, and SycO, acting as chaperones for YopE, YopH, and YopO, respectively [54].
In Salmonella sp., the multicargo T3SS chaperone SrcA interacts with the ATPase, SPI-2-encoded effector SsaN, and SteD, an adaptor that drives the ubiquitination and degradation of MHCII [55]. Mutation experiments have further shown that SrcA is required for the secretion of both of these effector proteins [44, 55]. Meanwhile, another chaperone, SseA is known to act as a chaperone allowing for the translocation of SseB and SseD [44]. In Salmonella enterica serovar Typhimurium, mutations in sseA has been shown to prevent the assembly of the SPI-2 T3SS and to result in loss of virulence and impaired intracellular replication [56].
Shigella flexneri Spa15 chaperone is unusual because it has multiple substrates: it has been shown to enhance the stability of IpaA, IpgB1, OspC3, and OspB [57]. In addition, several homologs of this chaperone have been described, including the SycB that acts as a chaperone for YspB in Y. enterocolitica [58].
The PscG–PscF complex in Pseudomonas aeruginosa is an example of class III that is required for the assembly of a stable needle for the T3SS. Consequently, mutations in these chaperones have been shown to result in an impairment of the T3SS functions and reduction of its cytotoxic effects on macrophages [59].
2.3 Roles of the T3SS
2.3.1 Effects of the T3SS on host cell signalling
Cell physiology is regulated by interconnected networks and intercellular cell signals. A common strategy by microbial pathogens is to interfere with these signals by suppressing or mimicking cellular messengers, notably using effector proteins secreted directly into the cell cytoplasm through the T3SS. Among the common targets of the T3SS are the cytoskeleton, endosomal trafficking, particularly those involved in phagocytosis, mitogen-activated protein (MAP) kinase, and nuclear factor-κB (NF-κB) pathways as well as the innate immunity [60, 61].
The MAP kinase pathway is one of the main cell signals controlling proliferation progression, stress, and inflammation response and plays a vital role in immunity of the host to bacterial pathogens [62]. Two pathways have been described, termed “canonical” and “non-canonical”, that both involve stimulation of membrane receptors of the EGF receptor family, ultimately leading to the cascade activation of transcription factors [62, 63]. Enteropathogenic bacteria in particular interfere with a variety of host cell mechanisms through the mitogen-activated protein kinase (MAPK) and NF-κB pathways [61]. In Yersinia, screening of host genes using RNA interference allowed to identify 19 NF-κB-regulated genes [64]. These genes included the heat shock protein H 1 (HSPH1) and encompass a variety of functions, including the NF-κB, the MAPK and the ERK signalling pathways, ion channel activity, and even cell growth regulation. Normally, these genes are upregulated by NF-κB, although this does not occur in the presence of wild-type Y. enterocolitica. However, curing this bacterium from the virulence plasmid pYV (that carries the bacterium T3SS) restores expression of these genes in infected cells, leading the authors to conclude that they were the target of Y. enterocolitica T3SS [64].
Moreover, research on the T3SS effector NleB1 from enteropathogenic E. coli showed that the effector NleB1 binds to the death domain of the Fas-associated protein [65]. This binding prevents the formation of the canonical death-inducing signalling complex (DISC) and the proteolytic activation of caspase-8 in order to avert apoptosis [65]. Similarly, another T3SS protein has been described as preventing cleavage of caspase-8, caspase-3, and the receptor-interacting serine/threonine protein kinase 1 (RIPK1) as well as binding caspase-4, -8, and -9 to inhibit their activity [66].
2.3.2 Cytotoxic effects
The cellular cytoskeleton is a network of actin and microtubules that act together to give shape and rigidity to eukaryotic cells while also playing a role in cellular trafficking, cellular uptake and cell movement. A major aspect of the cytoskeleton is the Rho family that constitutes a subfamily of the broader Ras super-family. Rho are small GTPases modulating actin organization, cell cycle progression and gene expression and playing a role in the organization of guanine exchange factors (GEF) and GTPase activating proteins (GAP) to change the inactive GDP- bound to active GTP- bound [67].
Members of the Rho family are a major target for effector proteins of the T3SS, either for inhibition or activation of these networks. For example, in Yersinia spp., translocation of the recruiting effector YopD through the cell membrane allows for the formation of channels through which YopE is delivered into the cell cytoplasm [32]. YopE GTPase inactivates members of the Rho family, including RhoA, Rac1, but not Cdc42 which is not considered an in vivo target for YopE [68]. This inactivation leads to disruption of the actin cytoskeleton [68, 69] and elicits a proinflammatory signalling response culminating in cytotoxicity [70]. However YopE is also recognised as a danger signal by macrophages and it has been shown that presence of YopE increased macrophage mediated killing of Yersinia bacteria [71]. Another effector of Yersinia spp., YopT, is a cysteine protease that induces apoptosis of macrophages through the inhibition of the MAPK and NFκB pathways [72], disturbing the F-actin structure of the cytoskeleton and resulting in a rounding of affected cells [73]. Interestingly, YopT competes with YopE for the same pool of Rac1, and the two effector proteins have antagonistic properties [71]. Similarly, YopK/YopQ (the name varies between homolog in different species of Yersinia) plays a role in the control of Yop translocation across the host cells [74]. In Y. pseudotuberculosis, YopQ has been associated with cytotoxic effects on polymorphonuclear leucocytes, although the precise mechanisms of actions remain unclear [75]. Similarly, YopJ effector has an inhibitory function for MAPK and NF-κB [76]. This effector prevents phosphorylation of serine and threonine residues MAPKK6 and interrupts the pathway [76, 77], reducing the production of the anti-apoptotic regulator Bcl-2 and leading to apoptosis of infected macrophages [78]. Moreover, YopE interferes on the signalling by acetylation of MEK2, MEK6 and, IKK as well as promoted apoptosis in infected macrophages [72, 78].
Salmonella enterica also induces apoptosis in macrophages through an unknown NF-κB-independent mechanism that relies on signalling through the kepro-inflammatory activator caspase 1 [61, 63, 79, 80]. Ectopic expression of the SPI-2 T3SS effector, SseL, did not support a role in the down-regulation of the host immune response and in particular the NF-kB pathway [81].
2.3.3 Other means of circumventing the immune response
In addition to causing apoptosis, bacterial pathogens can protect themselves from phagocytosis and the immune system by interfering with the host cells in other ways. For example, YopE and YopT can hinder the maturation of prointerleukin-1β in macrophages [82]. Meanwhile, EspF and EspJ of E. coli inhibit the PI3K and Src signalling required for phagocytosis [61].
Moreover, in Citrobacter rodentium and other Attaching/Effacing (A/E) pathogens like E. coli, the metalloprotease effector NleC cleaves the transcription factor p65, a transcription factor involved in the transcription of NF-κB in colon epithelial cells isolated from the intestine of mice. The resulting p651–38 fragment then interferes with p65/ RPS3 interactions to hinder the transcription of NF-κB and NF-κB signalling [83]. Likewise, the effector SpvB secreted by the Salmonella pathogenicity island-2 (SPI-2) T3SS of Salmonella enterica serovar Typhimurium has been shown to promote the expression of the E3 ligase KEAP1 [84]. KEAP1 acts as a repressor for IKKb which is in turn a promoter of NF-kB, therefore this effector proteins allows the bacterium to supress the NF-kB-dependent signalling pathway associated with the immune response [84].
Two distinct T3SS have been described in members of the Salmonella genus, located on the SPI-1 and SPI-2 and correspondingly named T3SS1 and T3SS2 [85, 86]. Both of these T3SS are required for bacterial virulence: one of the pathways of intracellular infection in Salmonella spp. involves the formation of salmonella containing vacuoles (SCV) in which the bacterium can multiply overtime [87]. Salmonella spp. requires both T3SS1 and T3SS2 to survive within these SCV and effector proteins interfere with the maturation of the vacuoles [87], although the precise mechanisms involved remain unclear. Effectors from T3SS1 are generally detected earlier in the infection process and it has been suggested that they are more involved with the earlier stages of the vacuole maturation while effectors from T3SS2 are more involved with later stages [87]. Finally, experiments using mutants of Acidovorax avenae identified an effector protein (A. avenae K1 suppression factor 1) that was associated with supressing the cellular response following recognition of flagellin associated molecular patterns [88].
2.3.4 Interference with the cytoskeleton to promote internalisation and cell invasion
Finally, a major role of the T3SS is in tissue colonisation to assist with bacterial attachment and to provide a mechanism of intracellular invasion through the trigger mechanism. Tir molecules are effectors which exist in two distinct forms, secreted by enteropathogenic and enterohaemmorhagic E. coli, respectively [89]. Both forms of Tir stimulate actin assembly through Nck-independent pathways leading to the recruitment of F-actin at the site of contact and the formation of pedestal structures to which the bacteria can then attach tightly [89].
A large class of effector proteins, best known in Shigella spp. and Salmonella spp., are characterised by their Trp-X-X-X-Glu motif [90]. These proteins are known to activate the Rho GTPases RhoA, Cdc42, and Rac1, resulting in the reconfiguration of the cytoskeleton and the formation of stress fibres, filopodia, and ruffling of the host cell membrane, leading to the intracellular uptake of the bacteria [90,91,92]. Similarly, the effector proteins SopE, SopE2, and SigD of Salmonella can activate actin in a Arp2/3-dependent manner [93], resulting in membrane ruffling that facilitates bacterial invasion [94]. Following internalisation, the activation of Cdc42 and Rac is reversed by SptP, another effector protein while IpgD destabilizes tight junctions, facilitating the formation of SVC [93,94,95].
3 T3SS in aquatic pathogens
As mentioned, many T3SS-harbouring bacterial pathogens have been associated with diseases in fish and other aquatic animals. However, despite this importance, much remains to be learned about these virulence factors. A review of what is currently known is presented in the following pages.
3.1 Edwardsiella spp.
The Edwardsiella genus represents a small but growing group of facultative anaerobic enteric bacteria and includes pathogens causing disorders in both fish and mammals. Mutation studies performed on 7 genes from the T3SS of E. tarda resulted in an increase in the bacterium LD50 (at least tenfold) as well as a decreased survival and growth of the bacterium in fish phagocytes following silencing of the T3SS [96]. The first effector of T3SS discovered in E. tarda was EseG whose secretion is dependent on the activity of the EscB chaperone [97]. EseG shares partial homology with the effectors SseG and SseF of Salmonella spp. and like these two proteins was found to interact with the cytoskeletons of the host cells, suggesting that it too might be involved in promoting internalisation of bacteria into fish cells [97]. In addition, the inner rod protein of the T3SS, alongside the flagellin molecule, induces pyroptosis activating the death process in host cells [98]. Pyroptosis starts with the activation of caspase-1 via the inflammasome complex, resulting in the secretion of IL-1β, IL-18 and TNF-α and concluding in cell death [98]. The protein EsaE is a membrane bound protein required for the secretion of T3SS effectors and its deletion has been shown to result in the attenuation of the bacterium [99]. Liu et al., furthermore demonstrated that the expression of E. tarda T3SS was regulated through a ternary complex involving EsaB, Esa, and EsaM [100]. Notably, they found that the activity of EsaL was pH dependent with EsaL suppressing secretion at pH 7.2 but promoting it under acidic conditions (pH 5.5) [100].
E. ictaluri is best known as a pathogen of catfish and sequencing of E. ictaluri plasmids pEI1 and pEI2, revealed remarkable similarities with the T3SS of E. tarda as well as SlrP, SspH1, and SspH2 in Salmonella typhimurium and IpaH of S. flexneri. pEI2 has also limited similarities to Spa15 of S. flexneri 5 and InvB of Shigella sonnei and S. typhimurium [101]. Signature-tagged mutagenesis have shown that these regions were necessary for the ability of E. ictaluri to cause infections [102, 103]. Both acidification and subsequent neutralization of the Edwardsiella containing vacuoles (ECV) are required to trigger the translocation of the E. ictaluri effectors [103], a feature that appears unique among bacterial pathogens. In contrast, transcription and assembly of the related Salmonella SPI-2 T3SS only requires acidification of the SCV [104]. The EseN effector plays an important role in E. ictaluri invasion, where it activates extracellular replication in catfish head kidney by dephosphorylation in vivo in the head kidney of infected fish, and ex vivo in macrophages derived from the head kidney [105]. EseN can also shut down signal-regulated kinases 1 and 2 (ERK1/2) early in the infectious process [80, 105]. Furthermore, there is evidence of reduced numbers of bacteria in infected tissue and in mortality in channel catfish (Ictalurus punctatus) infected with bacteria with a deleted eseN gene bacteria compared to wild-type infected tissue [105]. Finally, low pH and low phosphate triggers the expression of the T3SS effectors EseB, EseC, and EseD expression as well as the PEI1and PEI2 encoded, ESeH and ESeI [106].
Edwardsiella piscicida, has been described more recently and has been associated with diseases in aquatic animals, e.g. catfish and zebrafish [107]. It possesses both a T3SS and a T6SS, which are responsible to transport 12 effectors with synergistic activity [108]. In-frame deletion mutants of the T3SS structural genes EpDssaV, EpDesaM, EpDyscR in E. piscicida have resulted in loss of virulence in the bacterium and further analysis found that the mutants had potential efficacy as vaccines in catfish fingerlings [109]. The T3SS E. piscicida triggers TNF-α and IL-1β, synchronously, and activates inflammatory cytokines contributing to spreading the infection in host cells [110]. More recently, transcriptomic analysis using the regulator EsrB have allowed to identify 6 novel effector proteins that were co-expressed alongside proteins of the T3SS, although the exact role of these effectors remains to be elucidated [111].
3.2 Vibrio spp.
Vibrio species include a wide array of pathogenic bacteria found in both marine and terrestrial animals and these bacteria are considered endemic in water reservoirs throughout the world and are uncommon in the fact that many members of this genus commonly harbour two chromosomes, of varying size [112]. Most members of the genus Vibrio only harbour one T3SS [113]. However, sequencing has shown that several pathogenic strains of Vibrio parahaemolyticus harbour two distinct T3SS gene clusters encoding two different apparatus known as T3SS1, homolog to the one shared by other Vibrio spp., and T3SS2, carried on the main and secondary chromosome, respectively [113].
The T3SS1 showed an organisation comparable to that of the T3SS of Yersinia spp. [114]. Mutant strains of V. parahaemolyticus deficient in either T3SS1 or T3SS2 activity have been generated by Park et al. [114] and infection studies using these mutants have shown that T3SS1 was involved in the cytotoxic activity of the bacterium [114]. Activity of the T3SS1 on fibroblasts has shown an impact on the expression of 398 host genes [115]. Multiple T3SS1 effector genes have been identified [113], these include the effectors VopQ, VopS, VPA0450 and VopR (VP1683). VopQ has been linked to cell autophagy and toxicity via JNK-pathways leading to the expression of IL-8, caspase-1, IL-1β, and IL-18 and the inactivation of Cdc42, culminating in cellular cell death [116,117,118]. Moreover, the fic domain of VopS promotes cytoskeleton destruction via RhoA, Rac1 and Cdc42 (Rho GTPases) AMPylation [116, 119]. VopS and VopQ are homologous proteins, collaborating to promote cell autophagy concomitantly [119,120,121]. Moreover, VPA0450 was reported to disrupt the bacterial cell membrane by interfering with the cytoskeleton, resulting in membrane blebbing [122]. In addition, the regulators EsxA, EsxC, ExsD and ExsE have been shown to be necessary for the expression of the T3SS1. ExsA acts as a positive transcriptional regulator and plays a regulatory role for T3SS1 and shows DNA binding motifs linked to multiple T3SS1 genetic operons and promotes the expression of the T3SS1. ExsD binds ExsA to block expression of the T3SS1 genes while Exsc binds ExsD to allow for their expression [123]. Expression of the T3SS is regulated through the regulatory protein ExsA and several environmental factors like temperature above 30 °C or elevated presence of magnesium in the medium have been correlated with an increased activity [124]. T3SS1 is further involved in suppressing inflammatory responses through the inactivation of Cdc42 which otherwise acts as a stimulator to activate inflammasomes, NLRP3 and NLRC4 in host cells [117]. In V. alginolyticus, the effector proteins Val1686 and Val1680 include a Fic-domain, and have been shown to induce apoptosis, cell rounding and cell lysis through Val1686-dependent Rho GTPase inhibition [125]. However blocking the caspase 3 apoptotic pathway does not prevent cell rounding, membrane pore forming or cell death, suggesting that this T3SS can act through other cellular pathways beside caspase 3 [126]. Conversely, the T3SS1 was also shown to upregulate several pathways involved in cell-survival, a feature that the authors suggested might help the bacterium reduce the immune response by masking its cytopathic activity [115].
Somewhat less information is available regarding the rarer T3SS2. Screening of 155 environmental isolates (defined by the absence of the thermostable direct haemolysin (TDH) and TDH-related haemolysin) for genes coding for 2 elements of the T3SS2 (vscC2 and vopP), detected these genes in 10 or 8 isolates, respectively [127]. This suggested that this virulence factor might be uncommon in environmental isolates. Similarly, screening of environmental isolates from grouper and milkfish suggested that presence of T3SS2-associated genes was also variable, ranging from 14 to 100% of the isolates, depending on the gene [128]. When present, the T3SS2 has been linked to enterotoxicity [129, 130]. Notably, the T3SS2 is necessary for the secretion of the effector protein VopT which is a homolog of the Pseudomonas exoenzyme T (ExoT) [130]. Expression of VopT has been shown to inhibit growth in yeast culture while mutant of V. parahaemolyticus deficient in VopT displayed reduced cytotoxic activity against Caco-2 cells [130]. However, compared to the T3SS1, the T3SS2 appears to only be active against a limited number of cell lines [131]. Finally, Calder et al. have described the role of the T3SS2 in the formation of biofilms [132].
3.3 Aeromonas spp.
Aeromonas salmonicida is another major pathogen in aquaculture and the causative agent of furunculosis [132, 133]. The T3SS of A. salmonicida is likely its most important virulence factor [134] but was only relatively recently reported, its discovery probably hindered by the fact that T3SS are lost when the bacterium is cultivated above 20 °C [135,136,137], due to the T3SS gene cluster being locating in a thermosensitive region [138]. Among the most important effector proteins secreted by the T3SS of A. salmonicida are AopH, Ati2, AexT, AopP, AopO, AopN and ExsE [134], that have been associated with cytoskeletal collapse and immune system response impairment [139], with aopO being upregulated during the infectious process [140]. Moreover, AcrV and AopB are homologues in LcrV and YopB in Yersinia which stimulate IL-10 production in the cell, consequently supressing the immune response [141].
More recently, experimental infections of Oncorhynchus mykiss by wild-type A. salmonicida harbouring a fully functional T3SS, a mutated T3SS ascV (ΔascV), or a strain in which the T3SS had been completely lost confirmed the central role of the T3SS in the establishment of disease, with ΔascV being attenuated and the T3SS-less strain being totally avirulent [142]. Moreover, the authors also performed RT-qPCR on the anterior kidneys of infected fish showing a down-regulation of several immune genes associated with T3SS activity. Notably, expression of the interleukin 2 (a cytokine regulating proliferation of T-cells) and interferon gamma (a cytokine produced by T-cells) as well as that of the markers CD4 and CD8 (both expressed by different sub-populations of T-cells) were very strongly downregulated in fish infected with the wild-type and ΔascV strains but not in the strain without T3SS [142]. These results suggested that the T3SS of A. salmonicida had a strong immunosuppressive effect, particularly targeting different populations of T-cells, even if the precise effectors involved remain to be elucidated [142].
The effector AexT has been linked to cell cytotoxicity. It is considered homologous to P. aeruginosa bifunctional toxins exoenzyme S (ExoS) and exoenzyme T (ExoT), having a GAP function, activating members of the Rho family and resulting in the depolymerisation of ADP-ribosyltransferase actin and cell rounding [143]. Studies in mice have shown that deletion of aexU gene from the genome of A. salmonicida resulted in loss of virulence [144]. Furthermore, immunization of mice with recombinant AexU protected them from subsequent lethal challenge dose by the wild-type bacteria.
Notably, presence of T3SS genes is less systematic in A. hydrophila, the other major fish pathogen in this genus [133], as only some of the strains of the bacterium harbour all the genes for a functional T3SS [145]. However, sequencing of the gene cluster of A. hydrophila AH-3 using primers derived from sequences in A. salmonicida revealed the presence of 35 T3SS genes, 20 of these genes were homologous to genes on A. salmonicida and at least half of the remaining 15 proteins appeared novel to Aeromonas [19]. Silencing of the T3SS by deletion of the ascV gene resulted in loss of virulence in both rainbow trout and mice, showing the role of the T3SS in the disease process [19]. This decreased virulence of the mutants was confirmed in dwarf gourami (Trichogaster lalius) and was associated with reduced cytotoxicity and increased phagocytosis, which could highlight some mechanisms of action of this T3SS [146]. Epidemiologically, screening for the presence of several genes associated with the T3SS showed that the strains harbouring such genes were more likely to be associated with outbreaks of disease [145]. Similarly, screening of Aeromonas spp. from human patients has also shown that Aeromonas caviae, generally regarded as less virulent, were less likely to harbour a T3SS than other members of the genus [147]. The regulatory mechanisms of this T3SS in A. hydrophila have been investigated and it was shown that it was expressed in response to several environmental factors, including calcium depletion, high magnesium concentration, and high temperature [148]. The latter makes an interesting contrast with A. salmonicida where the T3SS is lost at high temperature. As previously stated, several of the effector proteins secreted by the T3SS of A. hydrophila are homologous or present similarities with those of A. salmonicida. For example, the first part of the aexT gene is identical in A. hydrophila and A. salmonicida [149], while the second half is different. The same study identified the protein ADP-ribosyltransferase activity and showed that mutation of aexT resulted in a slight reduction of their virulence both in vitro and in vivo in both three-spot gourami (Trichogaster trichopterus) or mice [149]. Similarly, aexU was found to have similarities with aexT of A. salmonicida [144, 150]. In addition, this protein also demonstrated GAP activity resulting in the disruption of actin filaments, inhibiting cytokine secretion and resulting in apoptosis in HeLa cells [151]. Like for A. salmonicida, deletion of the gene resulted in increased phagocytosis and decreased virulence of the bacterium [144, 150, 151]. Despite these advances, much is still to be learned about the T3SS of A. hydrophila. Notably, the precise targets and mechanisms of actions of the known T3SS effectors remain to be determined. Moreover, because of the variability in T3SS between isolates and the limited numbers of strains that have been investigated, it is likely that a significant number of effector proteins remains to be identified.
Finally, Matys et al. have recently reviewed the effector proteins secreted by bacteria of the Aeromonas genus, and this article should be of great interest to readers [152].
3.4 Flavobacterium spp.
The genus Flavobacterium encompasses many species and is ubiquitous in aquatic and soil environments. Among these species, several are well known as opportunistic or true pathogens. In particular Flavobacterium psychrophilum which is a major problem in particular in fish hatcheries, and Flavobacterium columnare, which is mainly known as a pathogen in warm water fish, notably in I. punctatus [153]. Even within a species, it is known that different strains can vary wildly in term of their virulence [154, 155]. For example, in F. columnare, examination of the bacterial genome has allowed to subdivide members of the species in several subgroups termed genomovars [156, 157], and these genetic groups appear to have at least some correlation with the virulence of the isolates [155]. T3SS are not unknown within this genus: examination of the genomes from two strains belonging to two different genomovar revealed the presence of a partial T3SS on the genome of F. columnare ATCC 49512, considered avirulent in catfish, and a complete T3SS on the genome of F. columnare 94-081, belonging to the genomovar II and considered highly virulent [158]. Similar examination performed on 4 strains belonging to genomovar I with varying degrees of virulence identified T3SS on all genomes [159] and suggested that differences in virulence were likely related to differences in chemotaxis and bacterial adhesion rather than the T3SS. Overall, the T3SS of Flavobacterium spp. does not appear to have garnered much attention. In the future, it might be of interest to systematically investigate the prevalence of this virulence factor within both clinical and environmental isolates of Flavobacterium spp. as well as clarify the role that it might play in the establishment of disease by deletion mutation or other silencing of the genes involved.
3.5 Yersinia ruckeri
Yersinia ruckeri is a major pathogen, particularly well-known in salmonid fish, and associated with generalized bacteraemia and septicaemia [160]. The presence of a T3SS in Y. ruckeri was reported by Gunasena et al. in 2003 [161]. Subsequent sequencing of the complete Yersinia ruckeri SC09 genome has allowed to further confirm the components of a ysa T3SS [162]. This T3SS is unusual because of its chromosomal location and it is carried on an operon with a moderate (ranging from 60 to 37%) identity to four genes of Y. enterocolitica [161]: YsaV (protein ID: CNI46870.1), YsaK (protein ID: CNI46839.1), YsaN (protein ID: CNI46802.1) and CDS19 (protein ID: NZ_KN150747.1). Furthermore, it displays similarities in gene sequence, arrangement and gene content with that of Y. enterocolitica biotype 1B and SPI-1 [163]. However, beside the sequencing of these genes, no information is currently available regarding the function of this T3SS and of their role in the virulence, internalization, replication and invasion mechanisms [162, 164]. This is particularly surprising considering the importance of Y. ruckeri as a fish pathogen and the amount of research that have otherwise focused on this pathogen.
3.6 Pseudomonas spp.
Pseudomonas spp. are gram-negative bacteria belonging to the γ-proteobacteria [165] that can display a high level of resistance for antibiotics [166] and infect a wide range of animals including human and fish [167, 168]. For example, P. aeruginosa is a normal part of the fish microbiota but may behave as an opportunistic pathogen in immunocompromised fish, resulting in ulcers and hemorrhagic septicemia [168]. The T3SS of P. aeruginosa is similar in structure with that of other gram-negative bacteria like Salmonella, Shigella and Yersinia spp. [169] and it has been theorized that this T3SS might have evolved to kill environmental amoeba as a defense mechanism against predation [170]. Four effector proteins (ExoT, ExoS, ExoU and ExoY) have been described in P. aeruginosa. Intriguingly, ExoS and ExoU are, for unknown reason, rarely secreted together in the same strain and have been associated with apoptosis and rapid cell lysis, respectively [166]. Furthermore, deletion mutations have shown that ExoU plays a role in damaging the lung epithelium in mice [171]. The other two effector proteins include ExoT and ExoY. ExoT has been linked to apoptosis in cultured cells following disruption of the mitochondrial membrane and leaking of cytochrome C into the cytosol [172, 173]. Furthermore, this effector also interferes with the actin cytoskeleton in order to inhibit phagocytosis, as well as slow wound healing by preventing epithelial cell migration in order to facilitate bacterial colonization [174]. The last known effector protein is ExoY which promotes secretion of the cyclic pyrimidine nucleotides cGMP and cUMP which play an important regulatory role in apoptosis and ion channel regulation, as well as smooth muscle control, including in the blood vessels [175].
While most of this work has been conducted on mammalian models, and in particular a mouse model, the proteins targeted by P. aeruginosa are well conserved and it is plausible that the T3SS has similar effects on fish cells. Moreover, these mechanisms of action, notably induction of apoptosis, escape from the immune system, and epithelial damage and delayed healing are consistent with the clinical signs associated with P. aeruginosa infections in fish [168].
4 New potential avenues for research
While T3SS appear commonplace among aquatic bacterial pathogens, they are not universal in every species. Therefore, it would be of interest to systematically screen and sequence both clinical and environmental bacterial isolates to identify the presence of T3SS. This is particularly the case for members of the genus Aeromonas and Flavobacterium in which the presence of T3SS is only sporadic. Moreover, comparing the prevalence of these virulence factors between clinical and environmental isolates would provide us with some understanding of their role in disease.
In addition, sequencing of the T3SS regions would allow to identify some of the effector proteins based on homology with previously characterized proteins in other species. This approach could be complemented by mutation experiments targeting structural element of the secretion apparatus followed by a comparison of the mutant secretome to that of the wild type isolate. Effector proteins are the most variable part of the T3SS, including between isolates of the same species, therefore there are likely many such proteins that remain to be identified and this approach has the advantage not to be reliant on sequence homology with already known effector proteins.
Moreover, the mutants could then be tested for infection and virulence, both in vivo and in vitro, to assess their role in the infection process. This approach would be relevant for bacterial fish pathogen in which the presence of a T3SS is known but its role in the establishment of disease remains unclear, like in Y. ruckeri or Flavobacterium spp. The specific mechanisms through which the expression of these T3SS is regulated is also an area of interest as is the target and mechanisms of actions of individual effector proteins. The later could be investigated by transfecting cells with plasmids expressing the gene of interest. The effect of this transfection on the cells physiology could then be investigated [176]. Molecules belonging to the T3SS would also represent interesting targets for the development of prophylaxis or therapeutic treatments. For example, the T3SS of P. aeruginosa has been targeted and several small molecules have been identified with inhibitory properties on the T3SS [166] as well as antibodies targeting the protein PcrV [177]. Similarly, additional vaccine attempts have been conducted targeting the homologous LcrV on Y. pestis [177] and AopO, a T3SS effector of A. salmonicida, has proven immunogenic in O. mykiss [140]. Targeting virulence mechanisms rather than the pathogen itself is advantageous not only because it offers new targets but also because it is generally accepted that it exerts a lower selective pressure on the pathogen, leading to a delayed emergence of resistance mechanisms [178]. Currently, most of these research efforts are being conducted on human pathogens; however, it is highly plausible that many of these therapeutants will be equally effective on fish pathogens. Finally, interference with the T3SS could also be performed by targeting its regulatory mechanisms. In particular, some of T3SS are known to be regulated through quorum sensing and strategies that target quorum sensing molecules, such as quorum quenching probiotics, have already been identified in fish health management [179, 180].
5 Conclusions
T3SS constitute a way for bacterial pathogens to manipulate the physiology of the host cells. They represent a powerful and versatile tool, and some of the most important virulence factors of gram-negative bacteria. Several examples show that it is also the case for multiple bacterial fish pathogens. However, comparatively less is known about their exact repertoire of secreted effector proteins as well as the targets and precise mode of action of these effectors. This is for example the case of otherwise well-known pathogens such as Y. ruckeri or members of the genus Vibrio or Aeromonas. In this context, much research effort, as detailed above, is still required to improve our understanding of the role of the T3SS in aquatic bacterial pathogens, how common they are and how they contribute to specific diseases as well as clarifying the factors governing their expression. Moreover, molecules belonging to these T3SS might also represent new targets for the development of vaccines or even new therapeutic treatments.
Availability of data and materials
The data supporting the conclusions of this article is included within the article.
Abbreviations
- A/E pathogens:
-
Attaching/effacing pathogens
- ATPase:
-
Adenosine triphosphatase
- Bp:
-
Base pair
- CD4:
-
Cluster of differentiation 4
- CD8:
-
Cluster of differentiation 8
- Cdc42:
-
Cell division control protein 42 homolog
- DISC:
-
Death-inducing signalling complex
- cGMP:
-
Cyclic guanosine monophosphate
- cUMP:
-
Cyclic uridine monophosphate
- ECV:
-
Edwardsiella Containing vacuoles
- ERK:
-
Extracellular signal-regulated kinases
- ExoS:
-
Exoenzyme S
- ExoT:
-
Exoenzyme T
- Fic domain:
-
Filamentation induced by c-AMP domain
- GAP:
-
GTPase- activating protein
- GEF:
-
Guanine exchange factors
- GTPase:
-
Nucleotide guanosine triphosphatase
- HSPH1:
-
Heat Shock Protein H 1
- IL-1β:
-
Interleukin 1 β
- IL-2:
-
Interleukin 2
- IL-8:
-
Interleukin 8
- IL-10:
-
Interleukin 10
- IL-18:
-
Interleukin 18
- IKKb:
-
Inhibitor of Nuclear Factor Kappa B Kinase Subunit Beta
- JNK pathway:
-
C-Jun N-terminal kinase pathway
- kDa:
-
Kilodalton
- Keap1:
-
Kelch ECH associating protein 1
- Map:
-
Mitochondrial associated protein
- MAPK:
-
Mitogen-activated protein kinases
- M ring:
-
Membrane ring
- NF-κB:
-
Nuclear factor-κB
- RAC1:
-
Ras-related C3 botulinum toxin substrate 1
- RAS:
-
Rat sarcoma viral oncogene homolog
- RIPK1:
-
Receptor-interacting serine/threonine protein kinase 1
- Rho:
-
Ras homolog
- SCV:
-
Salmonella containing vacuoles
- TDH:
-
Thermostable direct haemolysin
- Tir:
-
Translocated intimin receptor
- TLR:
-
Toll-like receptor
- TNF-α:
-
Tumour necrosis factor α
- T3SS:
-
Type III secretion system
- Yop:
-
Yersinia outer protein
- Ysc:
-
Yop secretion
References
FAO (2016) The state of world fisheries and aquaculture. Romes
FAO (2018) The State of World Fisheries and Aquaculture Meeting the Sustainable Development Goals. Italy, Rome
Little DC, Newton RW, Beveridge MCM (2016) Aquaculture: a rapidly growing and significant source of sustainable food? Status, transitions and potential. Proc Nutr Soc 75:274–286. https://doi.org/10.1017/S0029665116000665
World Bank, The World Bank (2013) FISH TO 2030: Prospects for Fisheries and Aquaculture
Adams A, Aoki T, Franck CJ (2005) Recent Technological Advancements on Aquatic Animal Health and Their Contributions Toward Reducing Disease Risks - a Review. In: Bondad-Reantaso MG, Mohan CN, Crumlish M, Subasinghe RP (eds) Diseases in Aquaculture VI, Proceedings of the sixth symposium on diseases in Asian Aquaculture. Fish Health Section, Asian Fisheries Society, Manila, Philippines, Colombo, pp 71–88
Leung TLF, Bates AE (2013) More rapid and severe disease outbreaks for aquaculture at the tropics: implications for food security. J Appl Ecol 50:215–222. https://doi.org/10.1111/1365-2644.12017
Silhavy TJ, Kahne D, Walker S (2010) The bacterial cell envelope. Cold Spring Harb Perspect Biol 2:a000414. https://doi.org/10.1101/cshperspect.a000414
Miller SI, Salama NR (2018) The gram-negative bacterial periplasm: Size matters. PLoS Biol 16:e2004935. https://doi.org/10.1371/journal.pbio.2004935
Green ER, Mecsas J (2016) Bacterial Secretion Systems: An Overview. Virulence Mechanisms of Bacterial Pathogens. ASM Press, Washington, DC, USA, pp 213–239
Costa TRD, Felisberto-Rodrigues C, Meir A, Prevost MS, Redzej A, Trokter M, Waksman G (2015) Secretion systems in Gram-negative bacteria: structural and mechanistic insights. Nat Rev Microbiol 13:343–359. https://doi.org/10.1038/nrmicro3456
Lauber F, Deme JC, Lea SM, Berks BC (2018) Type 9 secretion system structures reveal a new protein transport mechanism. Nature 564:77–82. https://doi.org/10.1038/s41586-018-0693-y
Warne B, Harkins CP, Harris SR, Vatsiou A, Stanley-Wall N, Parkhill J, Peacock SJ, Palmer T, Holden MTG (2016) The Ess/Type VII secretion system of Staphylococcus aureus shows unexpected genetic diversity. BMC Genomics 17:222. https://doi.org/10.1186/s12864-016-2426-7
Moest TP, Méresse S (2013) Salmonella T3SSs: successful mission of the secret(ion) agents. Curr Opin Microbiol 16:38–44. https://doi.org/10.1016/j.mib.2012.11.006
Pallen MJ, Beatson SA, Bailey CM (2005) Bioinformatics, genomics and evolution of non-flagellar type-III secretion systems: a Darwinian perpective. FEMS Microbiol Rev 29:201–229. https://doi.org/10.1016/j.femsre.2005.01.001
Diepold A, Armitage JP (2015) Type III secretion systems: the bacterial flagellum and the injectisome. Philos Trans R Soc Lond B Biol Sci 370:20150020. https://doi.org/10.1098/rstb.2015.0020
Troisfontaines P, Cornelis GR (2005) Type III Secretion: more systems than you think. Physiology 20:326–339. https://doi.org/10.1152/physiol.00011.2005
Abrusci P, McDowell MA, Lea SM, Johnson S (2014) Building a secreting nanomachine: a structural overview of the T3SS. Curr Opin Struct Biol 25:111–117. https://doi.org/10.1016/j.sbi.2013.11.001
Notti RQ, Stebbins CE (2016) The Structure and Function of Type III Secretion Systems. Virulence Mechanisms of Bacterial Pathogens. ASM Press, Washington, DC, USA, pp 241–264
Vilches S, Urgell C, Merino S, Chacón MR, Soler L, Castro-Escarpulli G, Figueras MJ, Tomás JM (2004) Complete type III secretion system of a mesophilic Aeromonas hydrophila strain. Appl Environ Microbiol 70:6914–6919. https://doi.org/10.1128/AEM.70.11.6914-6919.2004
Wagner S, Grin I, Malmsheimer S, Singh N, Torres-Vargas CE, Westerhausen S (2018) Bacterial type III secretion systems: a complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol Lett 365:fny201. https://doi.org/10.1093/femsle/fny201
Tsai C-L, Burkinshaw BJ, Strynadka NCJ, Tainer JA (2015) The Salmonella type III secretion system virulence effector forms a new hexameric chaperone assembly for export of effector/chaperone complexes. J Bacteriol 197:672–675. https://doi.org/10.1128/JB.02524-14
Lara-Tejero M, Kato J, Wagner S, Liu X, Galan JE (2011) A sorting platform determines the order of protein secretion in bacterial type III systems. Science 331:1188–1191. https://doi.org/10.1126/science.1201476
Diepold A, Wiesand U, Cornelis GR (2011) The assembly of the export apparatus (YscR, S, T, U, V) of the Yersinia type III secretion apparatus occurs independently of other structural components and involves the formation of an YscV oligomer. Mol Microbiol 82:502–514. https://doi.org/10.1111/j.1365-2958.2011.07830.x
Diepold A, Kudryashev M, Delalez NJ, Berry RM, Armitage JP (2015) Composition, formation, and regulation of the cytosolic C-ring, a dynamic component of the type III secretion injectisome. PLoS Biol 13:e1002039. https://doi.org/10.1371/journal.pbio.1002039
Izoré T, Job V, Dessen A (2011) Biogenesis, regulation, and targeting of the type III secretion system. Structure 19:603–612. https://doi.org/10.1016/j.str.2011.03.015
Yang X, Guo Y, Luo J, Pu X, Li M (2013) Effective identification of Gram-negative bacterial type III secreted effectors using position-specific residue conservation profiles. PLoS One 8:e84439. https://doi.org/10.1371/journal.pone.0084439
Spreter T, Yip CK, Sanowar S, André I, Kimbrough TG, Vuckovic M, Pfuetzner RA, Deng W, Yu AC, Finlay BB, Baker D, Miller SI, Strynadka NCJ (2009) A conserved structural motif mediates formation of the periplasmic rings in the type III secretion system. Nat Struct Mol Biol 16:468–476. https://doi.org/10.1038/nsmb.1603
Diepold A, Amstutz M, Abel S, Sorg I, Jenal U, Cornelis GR (2010) Deciphering the assembly of the Yersinia type III secretion injectisome. EMBO J 29:1928–1940. https://doi.org/10.1038/emboj.2010.84
Gamez AM (2013) Structural analysis of a forkhead-associated domain from the type III secretion system protein YscD. University of California, San Diego
Kowal J, Chami M, Ringler P, Müller SA, Kudryashev M, Castaño-Díez D, Amstutz M, Cornelis GR, Stahlberg H, Engel A (2013) Structure of the dodecameric Yersinia enterocolitica secretin YscC and its trypsin-resistant core. Structure 21:2152–2161. https://doi.org/10.1016/j.str.2013.09.012
Burghout P, Beckers F, de Wit E, van Boxtel R, Cornelis GR, Tommassen J, Koster M (2004) Role of the pilot protein YscW in the biogenesis of the YscC secretin in Yersinia enterocolitica. J Bacteriol 186:5366–5375. https://doi.org/10.1128/JB.186.16.5366-5375.2004
Montagner C, Arquint C, Cornelis GR (2011) Translocators YopB and YopD from Yersinia enterocolitica form a multimeric integral membrane complex in eukaryotic cell membranes. J Bacteriol 193:6923–6928. https://doi.org/10.1128/JB.05555-11
Habenstein B, El Mammeri N, Tolchard J, Lamon G, Tawani A, Berbon M, Loquet A (2019) Structures of Type III Secretion System Needle Filaments. In: springer nature. pp 109–131
Daniel C, Dewitte A, Poiret S, Marceau M, Simonet M, Marceau L, Descombes G, Boutillier D, Bennaceur N, Bontemps-Gallo S, Lemaître N, Sebbane F (2019) Polymorphism in the Yersinia LcrV antigen enables immune escape from the protection conferred by an LcrV-Secreting Lactococcus Lactis in a Pseudotuberculosis mouse model. Front Immunol 10:1830. https://doi.org/10.3389/fimmu.2019.01830
Wei T, Gong J, Qu G, Wang M (2019) Interactions between Yersinia pestis V- antigen (LcrV) and human Toll-like receptor 2 (TLR2) in a modelled protein complex and potential mechanistic insights. BMC Immunol 20:48
Costa TRD, Edqvist PJ, Bröms JE, Åhlund MK, Forsberg Å, Francis MS (2010) YopD self-assembly and binding to LcrV facilitate type III secretion activity by Yersinia pseudotuberculosis. J Biol Chem 285:25269–25284. https://doi.org/10.1074/jbc.M110.144311
Goure J, Broz P, Attree O, Cornelis GR, Attree I (2005) Protective anti-V antibodies inhibit Pseudomonas and Yersinia translocon assembly within host membranes. J Infect Dis 192:218–225. https://doi.org/10.1086/430932
Coleman MA, Cappuccio JA, Blanchette CD, Gao T, Arroyo ES, Hinz AK, Bourguet FA, Segelke B, Hoeprich PD, Huser T, Laurence TA, Motin VL, Chromy BA (2016) Expression and association of the Yersinia pestis translocon proteins, YopB and YopD, are facilitated by nanolipoprotein particles. PLoS One 11:e0150166. https://doi.org/10.1371/journal.pone.0150166
Sheahan K-L, Isberg RR (2015) Identification of mammalian proteins that collaborate with type III secretion system function: involvement of a chemokine receptor in supporting translocon activity. MBio 6:e02023-e2114. https://doi.org/10.1128/mBio.02023-14
Burkinshaw BJ, Strynadka NCJ (2014) Assembly and structure of the T3SS. Biochim Biophys Acta 1843:1649–1663. https://doi.org/10.1016/j.bbamcr.2014.01.035
Parsot C, Hamiaux C, Page A-L (2003) The various and varying roles of specific chaperones in type III secretion systems. Curr Opin Microbiol 6:7–14. https://doi.org/10.1016/s1369-5274(02)00002-4
Cooper CA, Zhang K, Andres SN, Fang Y, Kaniuk NA, Hannemann M, Brumell JH, Foster LJ, Junop MS, Coombes BK (2010) Structural and biochemical characterization of SrcA, a multi-cargo type III secretion chaperone in Salmonella required for pathogenic association with a host. PLoS Pathog 6:e1000751. https://doi.org/10.1371/journal.ppat.1000751
Ellis RJ (2006) Molecular chaperones: assisting assembly in addition to folding. Trends Biochem Sci 31:395–401. https://doi.org/10.1016/j.tibs.2006.05.001
Allison SE, Tuinema BR, Everson ES, Sugiman-Marangos S, Zhang K, Junop MS, Coombes BK (2014) Identification of the docking site between a type III secretion system ATPase and a chaperone for effector cargo. J Biol Chem 289:23734–23744. https://doi.org/10.1074/jbc.M114.578476
Page A-L, Sansonetti P, Parsot C (2002) Spa15 of Shigella flexneri, a third type of chaperone in the type III secretion pathway. Mol Microbiol 43:1533–1542. https://doi.org/10.1046/j.1365-2958.2002.02835.x
Gauthier A, Finlay BB (2003) Translocated intimin receptor and its chaperone interact with ATPase of the type III secretion apparatus of enteropathogenic Escherichia coli. J Bacteriol 185:6747–6755. https://doi.org/10.1128/JB.185.23.6747-6755.2003
Thomas NA, Deng W, Puente JL, Frey EA, Yip CK, Strynadka NCJ, Finlay BB (2005) CesT is a multi-effector chaperone and recruitment factor required for the efficient type III secretion of both LEE- and non-LEE-encoded effectors of enteropathogenic Escherichia coli. Mol Microbiol 57:1762–1779. https://doi.org/10.1111/j.1365-2958.2005.04802.x
Schubot FD, Jackson MW, Penrose KJ, Cherry S, Tropea JE, Plano GV, Waugh DS (2005) Three-dimensional structure of a macromolecular assembly that regulates type III secretion in Yersinia pestis. J Mol Biol 346:1147–1161. https://doi.org/10.1016/j.jmb.2004.12.036
Edqvist PJ, Aili M, Liu J, Francis MS (2007) Minimal YopB and YopD translocator secretion by Yersinia is sufficient for Yop-effector delivery into target cells. Microbes Infect 9:224–233. https://doi.org/10.1016/j.micinf.2006.11.010
Singh SK, Boyle AL, Main ERG (2013) LcrH, a Class II chaperone from the type three secretion system, has a highly flexible native structure. J Biol Chem 288:4048–4055. https://doi.org/10.1074/jbc.M112.395889
Hu X, Lee MS, Wallqvist A (2009) Interaction of the disordered Yersinia effector protein YopE with its cognate chaperone SycE. Biochemistry 48:11158–11160. https://doi.org/10.1021/bi9017347
Locher M, Lehnert B, Krauss K, Heesemann J, Groll M, Wilharm G (2005) Crystal structure of the Yersinia enterocolitica type III secretion chaperone SycT. J Biol Chem 280:31149–31155. https://doi.org/10.1074/jbc.M500603200
Büttner CR, Cornelis GR, Heinz DW, Niemann HH (2005) Crystal structure of Yersinia enterocolitica type III secretion chaperone SycT. Protein Sci 14:1993–2002. https://doi.org/10.1110/ps.051474605
Letzelter M, Sorg I, Mota LJ, Meyer S, Stalder J, Feldman M, Kuhn M, Callebaut I, Cornelis GR (2006) The discovery of SycO highlights a new function for type III secretion effector chaperones. EMBO J 25:3223–3233. https://doi.org/10.1038/sj.emboj.7601202
Godlee C, Cerny O, Durkin CH, Holden DW (2019) SrcA is a chaperone for the Salmonella SPI-2 type three secretion system effector SteD. Microbiology 165:15–25. https://doi.org/10.1099/mic.0.000732
Ruiz-Albert J, Mundy R, Yu X-J, Beuzón CR, Holden DW (2003) SseA is a chaperone for the SseB and SseD translocon components of the Salmonella pathogenicity-island-2-encoded type III secretion system. Microbiology 149:1103–1111. https://doi.org/10.1099/mic.0.26190-0
Page A-L, Parsot C (2002) Chaperones of the type III secretion pathway: jacks of all trades. Mol Microbiol 46:1–11. https://doi.org/10.1046/j.1365-2958.2002.03138.x
Foultier B, Troisfontaines P, Vertommen D, Rider M, Parsot C, Cornelis GR (2003) Identification of substrates and chaperone from the Yersinia enterocolitica 1B Ysa Type III secretion system. Infect Immun 71:242–253. https://doi.org/10.1128/IAI.71.1.242-253.2003
Quinaud M, Plé S, Job V, Contreras-Martel C, Simorre JP, Attree I, Dessen A (2007) Structure of the heterotrimeric complex that regulates type III secretion needle formation. Proc Natl Acad Sci USA 104:7803–7808. https://doi.org/10.1073/pnas.0610098104
Raymond B, Young JC, Pallett M, Endres RG, Clements A, Frankel G (2013) Subversion of trafficking, apoptosis, and innate immunity by type III secretion system effectors. Trends Microbiol 21:430–441. https://doi.org/10.1016/j.tim.2013.06.008
Pinaud L, Sansonetti PJ, Phalipon A (2018) Host cell targeting by enteropathogenic bacteria T3SS effectors. Trends Microbiol 26:266–283. https://doi.org/10.1016/j.tim.2018.01.010
Robert J (2015) MAP Kinase Pathway. Textbook of Cell Signalling in Cancer. Springer International Publishing, Cham, pp 27–41
Shan L, He P, Sheen J (2007) Intercepting host MAPK signaling cascades by bacterial type III effectors. Cell Host Microbe 1:167–174. https://doi.org/10.1016/j.chom.2007.04.008
Micheva-Viteva SN, Shou Y, Nowak-Lovato KL, Rector KD, Hong-Geller E (2013) c-KIT signaling is targeted by pathogenic Yersinia to suppress the host immune response. BMC Microbiol 13:249. https://doi.org/10.1186/1471-2180-13-249
Pearson JS, Giogha C, Ong SY, Kennedy CL, Kelly M, Robinson KS (2013) A type III effector antagonizes death receptor signalling during bacterial gut infection. Nature 501:247–251. https://doi.org/10.1038/nature12524
Pollock GL, Oates CVL, Giogha C, Wong Fok Lung T, Ong SY, Pearson JS, Hartland EL (2017) Distinct roles of the antiapoptotic effectors NleB and NleF from enteropathogenic Escherichia coli. Infect Immun 85:e01071-e1116. https://doi.org/10.1128/IAI.01071-16
Ridley AJ (2001) Rho GTPases and cell migration. J Cell Sci 114:2713–2722. https://doi.org/10.1242/jcs.114.15.2713
Aili M, Isaksson EL, Hallberg B, Wolf-Watz H, Rosqvist R (2006) Functional analysis of the YopE GTPase-activating protein (GAP) activity of Yersinia pseudotuberculosis. Cell Microbiol 8:1020–1033. https://doi.org/10.1111/j.1462-5822.2005.00684.x
Rosqvist R, Forsberg Å, Wolf-Watz H (1991) Microinjection of the Yersinia YopE cytotoxin in mammalian cells induces actin microfilament disruption. Biochem Soc Trans 19:1131–1132. https://doi.org/10.1042/bst0191131
Mejía E, Bliska JB, Viboud GI (2008) Yersinia controls type III effector delivery into host cells by modulating Rho activity. PLoS Pathog 4:e3. https://doi.org/10.1371/journal.ppat.0040003
Wang X, Parashar K, Sitaram A, Bliska JB (2014) The GAP activity of type III effector YopE triggers killing of Yersinia in macrophages. PLoS Pathog 10:e1004346. https://doi.org/10.1371/journal.ppat.1004346
Schmidt G (2011) Yersinia enterocolitica outer protein T (YopT). Eur J Cell Biol 90:955–958. https://doi.org/10.1016/j.ejcb.2010.12.005
Iriarte M, Cornelis GR (1998) YopT, a new Yersinia Yop effector protein, affects the cytoskeleton of host cells. Mol Microbiol 29:915–929. https://doi.org/10.1046/j.1365-2958.1998.00992.x
Tan Y, Liu W, Zhang Q, Cao S, Zhao H, Wang T, Qi Z, Han Y, Song Y, Wang X, Yang R, Du Z (2017) Yersinia pestis YopK inhibits bacterial adhesion to host cells by binding to the extracellular matrix adaptor protein matrilin-2. Infect Immun 85:e01069-e1116. https://doi.org/10.1128/IAI.01069-16
Thorslund SE, Ermert D, Fahlgren A, Erttmann SF, Nilsson K, Hosseinzadeh A, Urban CF, Fällman M (2013) Role of YopK in Yersinia pseudotuberculosis resistance against polymorphonuclear leukocyte defense. Infect Immun 81:11–22. https://doi.org/10.1128/IAI.00650-12
Mukherjee S (2006) Yersinia YopJ acetylates and inhibits kinase activation by blocking phosphorylation. Science 312:1211–1214. https://doi.org/10.1126/science.1126867
Vossenkämper A, Marchès O, Fairclough PD, Warnes G, Stagg AJ, Lindsay JO, Evans PC, Luong LA, Croft NM, Naik S, Frankel G, MacDonald TT (2010) Inhibition of NF-κB signaling in human dendritic cells by the enteropathogenic Escherichia coli effector protein NleE. J Immunol 185:4118–4127. https://doi.org/10.4049/jimmunol.1000500
Pandey AK, Sodhi A (2009) Recombinant YopJ induces apoptosis in murine peritoneal macrophages in vitro: involvement of mitochondrial death pathway. Int Immunol 21:1239–1249. https://doi.org/10.1093/intimm/dxp086
El-Aouar Filho RA, Nicolas A, De Paula Castro TL, Deplanche M, De Carvalho Azevedo VA, Goossens PL, Taieb F, Lina G, Le Loir Y, Berkova N (2017) Heterogeneous family of cyclomodulins: smart weapons that allow bacteria to hijack the eukaryotic cell cycle and promote infections. Front Cell Infect Microbiol 7:208. https://doi.org/10.3389/fcimb.2017.00208
Bhavsar AP, Guttman JA, Finlay BB (2007) Manipulation of host-cell pathways by bacterial pathogens. Nature 449:827–834. https://doi.org/10.1038/nature06247
Mesquita FS, Holden DW, Rolhion N (2013) Lack of Effect of the Salmonella Deubiquitinase SseL on the NF-κB Pathway. PLoS One 8:e53064. https://doi.org/10.1371/journal.pone.0053064
Schotte P, Denecker G, Van Den Broeke A, Vandenabeele P, Cornelis GR, Beyaert R (2004) Targeting Rac1 by the Yersinia effector protein YopE inhibits caspase-1-mediated maturation and release of interleukin-1beta. J Biol Chem 279:25134–25142. https://doi.org/10.1074/jbc.M401245200
Hodgson A, Wier EM, Fu K, Sun X, Yu H, Zheng W, Sham HP, Johnson K, Bailey S, Vallance BA, Wan F (2015) Metalloprotease NleC suppresses host NF-κB/inflammatory responses by cleaving p65 and interfering with the p65/RPS3 interaction. PLoS Pathog 11:e1004705. https://doi.org/10.1371/journal.ppat.1004705
Yang S, Deng Q, Sun L, Zhu Y, Dong K, Wu S, Huang R, Li Y (2021) Salmonella effector SpvB inhibits NF-κB activity via KEAP1-mediated downregulation of IKKβ. Front Cell Infect Microbiol 11:641412. https://doi.org/10.3389/fcimb.2021.641412
Ramos-Morales F (2012) Impact of Salmonella enterica Type III Secretion System Effectors on the Eukaryotic Host Cell. ISRN Cell Biol 2012:787934. https://doi.org/10.5402/2012/787934
Shea JE, Hensel M, Gleeson C, Holden DW (1996) Identification of a virulence locus encoding a second type III secretion system in Salmonella typhimurium. Proc Natl Acad Sci U S A 93:2593–2597. https://doi.org/10.1073/pnas.93.6.2593
Steele-Mortimer O (2008) The Salmonella-containing vacuole: moving with the times. Curr Opin Microbiol 11:38–45. https://doi.org/10.1016/j.mib.2008.01.002
Kawaguchi T, Nakamura M, Hirai H, Furukawa T, Kondo M, Che F-S (2021) AKSF1 isolated from the rice-virulent strain Acidovorax avenae K1 is a novel effector that suppresses PAMP-triggered immunity in rice. Mol Plant Microbe Interact 34:186–197. https://doi.org/10.1094/MPMI-10-20-0271-R
Brady MJ, Campellone KG, Ghildiyal M, Leong JM (2007) Enterohaemorrhagic and enteropathogenic Escherichia coli Tir proteins trigger a common Nck-independent actin assembly pathway. Cell Microbiol 9:2242–2253. https://doi.org/10.1111/j.1462-5822.2007.00954.x
Bulgin R, Raymond B, Garnett JA, Frankel G, Crepin VF, Berger CN, Arbeloa A (2010) Bacterial guanine nucleotide exchange factors SopE-like and WxxxE effectors. Infect Immun 78:1417–1425. https://doi.org/10.1128/IAI.01250-09
Killackey SA, Sorbara MT, Girardin SE (2016) Cellular aspects of Shigella pathogenesis: focus on the manipulation of host cell processes. Front Cell Infect Microbiol 6:38. https://doi.org/10.3389/fcimb.2016.00038
Schroeder GN, Hilbi H (2008) Molecular pathogenesis of Shigella spp.: controlling host cell signaling, invasion, and death by type III secretion. Clin Microbiol Rev 21:134–156. https://doi.org/10.1128/CMR.00032-07
Hänisch J, Kölm R, Wozniczka M, Bumann D, Rottner K, Stradal TEB (2011) Activation of a RhoA/myosin II-dependent but Arp2/3 complex-independent pathway facilitates Salmonella invasion. Cell Host Microbe 9:273–285. https://doi.org/10.1016/j.chom.2011.03.009
Terebiznik MR, Vieira OV, Marcus SL, Slade A, Yip CM, Trimble WS, Meyer T, Finlay BB, Grinstein S (2002) Elimination of host cell PtdIns(4,5)P(2) by bacterial SigD promotes membrane fission during invasion by Salmonella. Nat Cell Biol 4:766–773. https://doi.org/10.1038/ncb854
Mason D, Mallo GV, Terebiznik MR, Payrastre B, Finlay BB, Brumell JH, Rameh L, Grinstein S (2007) Alteration of epithelial structure and function associated with PtdIns(4,5)P2 degradation by a bacterial phosphatase. J Gen Physiol 129:267–283. https://doi.org/10.1085/jgp.200609656
Tan YP (2005) Role of type III secretion in Edwardsiella tarda virulence. Microbiology 151:2301–2313. https://doi.org/10.1099/mic.0.28005-0
Xie H-X, Yu HB, Zheng J, Nie P, Foster LJ, Mok YK, Finlay BB, Leung KY (2010) EseG, an effector of the type III secretion system of Edwardsiella tarda, triggers microtubule destabilization. Infect Immun 78:5011–5021. https://doi.org/10.1128/IAI.00152-10
Xie H-X, Lu J-F, Rolhion N, Holden DW, Nie P, Zhou Y, Yu X-J (2014) Edwardsiella tarda-Induced cytotoxicity depends on its type III secretion system and flagellin. Infect Immun 82:3436–3445. https://doi.org/10.1128/IAI.01065-13
Zhou Y, Liu LY, He TT, Laghari ZA, Nie P, Gao Q, Xie H-X (2016) Edwardsiella tarda EsaE (Orf19 protein) is required for the secretion of type III substrates, and pathogenesis in fish. Vet Microbiol 190:12–18. https://doi.org/10.1016/j.vetmic.2016.05.003
Liu LY, Nie P, Yu HB, Xie H-X (2017) Regulation of Type III secretion of translocon and effector proteins by the EsaB/EsaL/EsaM complex in Edwardsiella tarda. Infect Immun 85:e00322-e417. https://doi.org/10.1128/IAI.00322-17
Fernandez DH, Pittman-Cooley L, Thune RL (2001) Sequencing and analysis of the Edwardsiella ictaluri plasmids. Plasmid 45:52. https://doi.org/10.1006/plas.2000.1499
Thune RL, Fernandez DL, Benoit JL, Kelly-Smith M, Rogge ML, Booth NJ, Landry CA, Bologna RA (2007) Signature-tagged mutagenesis of Edwardsiella ictaluri identifies virulence-related genes, including a Salmonella pathogenicity island 2 class of type III secretion systems. Appl Environ Microbiol 73:7934–7946
Baumgartner WA, Dubytska LP, Rogge ML, Mottram PJ, Thune RL (2014) Modulation of vacuolar pH is required for replication of Edwardsiella ictaluri in channel catfish macrophages. Infect Immun 82:2329–2336. https://doi.org/10.1128/IAI.01616-13
Yu X-J, McGourty K, Liu M, Unsworth KE, Holden DW (2010) pH sensing by intracellular Salmonella induces effector translocation. Science 328:1040–1043. https://doi.org/10.1126/science.1189000
Dubytska LP, Thune R (2018) Edwardsiella ictaluri type III secretion system (T3SS) effector EseN is a phosphothreonine lyase that inactivates ERK1/2. Dis Aquat Organ 130:117–129. https://doi.org/10.3354/dao03255
Rogge ML, Thune RL (2011) Regulation of the Edwardsiella ictaluri type III secretion system by pH and phosphate concentration through EsrA, EsrB, and EsrC. Appl Environ Microbiol 77:4293–4302. https://doi.org/10.1128/AEM.00195-11
Zhou P, Han X, Ye X, Zheng F, Yan T, Xie Q, Zhang Y-A, Curtiss R, Zhou Y (2020) Phenotype, virulence and immunogenicity of Edwardsiella piscicida cyclic AMP receptor protein (Crp) mutants in catfish host. Microorganisms 8:517. https://doi.org/10.3390/microorganisms8040517
Leung KY, Wang Q, Yang Z, Siame BA (2019) Edwardsiella piscicida : a versatile emerging pathogen of fish. Virulence 10:555–567. https://doi.org/10.1080/21505594.2019.1621648
Edrees A, Abdelhamed H, Nho SW, Park SB, Karsi A, Austin FW, Essa M, Pechan T, Lawrence ML (2018) Construction and evaluation of type III secretion system mutants of the catfish pathogen Edwardsiella piscicida. J Fish Dis 41:805–816. https://doi.org/10.1111/jfd.12784
Hu T, Chen R, Zhang L, Wang Z, Yang D, Zhang Y, Liu X, Liu Q (2019) Balanced role of T3SS and T6SS in contribution to the full virulence of Edwardsiella piscicida. Fish Shellfish Immunol 93:871–878. https://doi.org/10.1016/j.fsi.2019.08.014
Liu Y, Zhao L, Yang M, Yin K, Zhou X, Leung KY, Liu Q, Zhang Y, Wang Q (2017) Transcriptomic dissection of the horizontally acquired response regulator EsrB reveals its global regulatory roles in the physiological adaptation and activation of T3SS and the cognate effector repertoire in Edwardsiella piscicida during infection. Virulence 8:1355–1377. https://doi.org/10.1080/21505594.2017.1323157
Okada K, Iida T, Kita-Tsukamoto K, Honda T (2005) Vibrios commonly possess two chromosomes. J Bacteriol 187:752–757. https://doi.org/10.1128/JB.187.2.752-757.2005
Miller KA, Tomberlin KF, Dziejman M (2019) Vibrio variations on a type three theme. Curr Opin Microbiol 47:66–73. https://doi.org/10.1016/j.mib.2018.12.001
Park K-S, Ono T, Rokuda M, Jang M-H, Okada K, Iida T, Honda T (2004) Functional characterization of two type III secretion systems of Vibrio parahaemolyticus. Infect Immun 72:6659–6665. https://doi.org/10.1128/IAI.72.11.6659-6665.2004
De Nisco NJ, Kanchwala M, Li P, Fernandez J, Xing C, Orth K (2017) The cytotoxic type 3 secretion system 1 of Vibrio rewires host gene expression to subvert cell death and activate cell survival pathways. Sci Signal 10:aa14501. https://doi.org/10.1126/scisignal.aal4501
Luong P, Kinch LN, Brautigam CA, Grishin NV, Tomchick DR, Orth K (2010) Kinetic and structural insights into the mechanism of AMPylation by VopS fic domain. J Biol Chem 285:20155–20163. https://doi.org/10.1074/jbc.M110.114884
Higa N, Toma C, Koizumi Y, Nakasone N, Nohara T, Masumoto J, Kodama T, Iida T, Suzuki T (2013) Vibrio parahaemolyticus effector proteins suppress inflammasome activation by interfering with host autophagy signaling. PLoS Pathog 9:e1003142. https://doi.org/10.1371/journal.ppat.1003142
Matlawska-Wasowska K, Finn R, Mustel A, O’Byrne CP, Baird AW, Coffey ET, Boyd A (2010) The Vibrio parahaemolyticus Type III Secretion Systems manipulate host cell MAPK for critical steps in pathogenesis. BMC Microbiol 10:329. https://doi.org/10.1186/1471-2180-10-329
Yarbrough ML, Li Y, Kinch LN, Grishin NV, Ball HL, Orth K (2009) AMPylation of Rho GTPases by Vibrio VopS disrupts effector binding and downstream signaling. Science 323:269–272. https://doi.org/10.1126/science.1166382
Osorio CR (2018) T3SS effectors in Vibrios: homology in sequence, diversity in biological functions? Virulence 9:721–723. https://doi.org/10.1080/21505594.2018.1435965
Sreelatha A, Bennett TL, Zheng H, Jiang Q-X, Orth K, Starai VJ (2013) Vibrio effector protein, VopQ, forms a lysosomal gated channel that disrupts host ion homeostasis and autophagic flux. Proc Natl Acad Sci USA 110:11559–11564. https://doi.org/10.1073/pnas.1307032110
Broberg CA, Zhang L, Gonzalez H, Laskowski-Arce MA, Orth K (2010) A Vibrio effector protein is an inositol phosphatase and disrupts host cell membrane integrity. Science 329:1660–1662. https://doi.org/10.1126/science.1192850
Zhou X, Konkel ME, Call DR (2010) Regulation of type III secretion system 1 gene expression in Vibrio parahaemolyticus is dependent on interactions between ExsA, ExsC, and ExsD. Virulence 1:260–272. https://doi.org/10.4161/viru.1.4.12318
Liu AC, Thomas NA (2015) Transcriptional profiling of Vibrio parahaemolyticus exsA reveals a complex activation network for type III secretion. Front Microbiol 6:1089. https://doi.org/10.3389/fmicb.2015.01089
Zhao Z, Liu J, Deng Y, Huang W, Ren C, Call DR, Hu C (2018) The Vibrio alginolyticus T3SS effectors, Val1686 and Val1680, induce cell rounding, apoptosis and lysis of fish epithelial cells. Virulence 9:318–330. https://doi.org/10.1080/21505594.2017.1414134
Zhao Z, Chen C, Hu C-Q, Ren C-H, Zhao J-J, Zhang L-P, Jiang X, Luo P, Wang Q-B (2010) The type III secretion system of Vibrio alginolyticus induces rapid apoptosis, cell rounding and osmotic lysis of fish cells. Microbiology 156:2864–2872. https://doi.org/10.1099/mic.0.040626-0
Caburlotto G, Gennari M, Ghidini V, Tafi M, Lleo MM (2009) Presence of T3SS2 and other virulence-related genes in tdh -negative Vibrio parahaemolyticus environmental strains isolated from marine samples in the area of the Venetian Lagoon, Italy. FEMS Microbiol Ecol 70:506–514. https://doi.org/10.1111/j.1574-6941.2009.00764.x
Tey YH, Jong KJ, Fen SY, Wong HC (2015) Genetic variation in Vibrio parahaemolyticus isolated from the aquacultural environments. Lett Appl Microbiol 60:321–327. https://doi.org/10.1111/lam.12372
Li L, Meng H, Gu D, Li Y, Jia M (2019) Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol Res 222:43–51. https://doi.org/10.1016/j.micres.2019.03.003
Kodama T, Rokuda M, Park KS, Cantarelli VV, Matsuda S, Iida T, Honda T (2007) Identification and characterization of VopT, a novel ADP-ribosyltransferase effector protein secreted via the Vibrio parahaemolyticus type III secretion system 2. Cell Microbiol 9:2598–2609. https://doi.org/10.1111/j.1462-5822.2007.00980.x
Hiyoshi H, Kodama T, Iida T, Honda T (2010) Contribution of Vibrio parahaemolyticus virulence factors to cytotoxicity, enterotoxicity, and lethality in mice. Infect Immun 78:1772–1780. https://doi.org/10.1128/IAI.01051-09
Menanteau-Ledouble S, Kumar G, Saleh M, El-Matbouli M (2016) Aeromonas salmonicida: updates on an old acquaintance. Dis Aquat Organ 120:49–68. https://doi.org/10.3354/dao03006
Cipriano RC, Austin B (2011) Furunculosis and other aeromonad diseases. In: Woo PT, Bruno DW (eds) Fish diseases and disorders. Volume 3: viral, bacterial and fungal infections, 2nd ed. CABI, Wallingford, pp 424–483
Vanden Bergh P, Heller M, Braga-Lagache S, Frey J (2013) The Aeromonas salmonicida subsp. salmonicida exoproteome: determination of the complete repertoire of Type-Three Secretion System effectors and identification of other virulence factors. Proteome Sci 11:42. https://doi.org/10.1186/1477-5956-11-42
Frey J, Origgi FC (2016) Type III secretion system of Aeromonas salmonicida undermining the host’s immune response. Front Mar Sci 3:130. https://doi.org/10.3389/fmars.2016.00130
Burr SE, Stuber K, Wahli T, Frey J (2002) Evidence for a Type III Secretion System in Aeromonas salmonicida subsp. salmonicida. J Bacteriol 184:5966–5970. https://doi.org/10.1128/jb.184.21.5966-5970.2002
Daher RK, Filion G, Tan SG, Dallaire-Dufresne S, Paquet VE, Charette SJ (2011) Alteration of virulence factors and rearrangement of pAsa5 plasmid caused by the growth of Aeromonas salmonicida in stressful conditions. Vet Microbiol 152:353–360. https://doi.org/10.1016/j.vetmic.2011.04.034
Vincent AT, Bernatchez A, Frey J, Charette SJ (2019) A mesophilic Aeromonas salmonicida strain isolated from an unsuspected host, the migratory bird pied avocet. Microorganisms 7:592. https://doi.org/10.3390/microorganisms7120592
Vanden Bergh P, Frey J (2014) Aeromonas salmonicida subsp. salmonicida in the light of its type-three secretion system. Microb Biotechnol 7:381–400. https://doi.org/10.1111/1751-7915.12091
Menanteau-Ledouble S, El-Matbouli M (2016) Antigens of Aeromonas salmonicida subsp. salmonicida specifically induced in vivo in Oncorhynchus mykiss. J Fish Dis 39:1015–1019. https://doi.org/10.1111/jfd.12430
Vanden Bergh P, Burr SE, Benedicenti O, von Siebenthal B, Frey J, Wahli T (2013) Antigens of the type-three secretion system of Aeromonas salmonicida subsp. salmonicida prevent protective immunity in rainbow trout. Vaccine 31:5256–5261. https://doi.org/10.1016/j.vaccine.2013.08.057
Origgi FC, Benedicenti O, Segner H, Sattler U, Wahli T, Frey J (2017) Aeromonas salmonicida type III secretion system-effectors-mediated immune suppression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol 60:334–345. https://doi.org/10.1016/j.fsi.2016.12.006
Fehr D, Burr SE, Gibert M, D’Alayer J, Frey J, Popoff MR (2007) Aeromonas exoenzyme T of Aeromonas salmonicida is a bifunctional protein that targets the host cytoskeleton. J Biol Chem 282:28843–28852. https://doi.org/10.1074/jbc.M704797200
Sha J, Wang SF, Suarez G, Sierra JC, Fadl AA, Erova TE, Foltz SM, Khajanchi BK, Silver A, Graf J, Schein CH, Chopra AK (2007) Further characterization of a type III secretion system (T3SS) and of a new effector protein from a clinical isolate of Aeromonas hydrophila–part I. Microb Pathog 43:127–146. https://doi.org/10.1016/j.micpath.2007.05.002
Carvalho-Castro GA, Lopes CO, Leal CAG, Cardoso PG, Leite RC, Figueiredo HCP (2010) Detection of type III secretion system genes in Aeromonas hydrophila and their relationship with virulence in Nile tilapia. Vet Microbiol 144:371–376. https://doi.org/10.1016/j.vetmic.2010.01.021
Yu HB, Rao PSS, Lee HC, Vilches S, Merino S, Tomas JM, Leung KY (2004) A type III secretion system is required for Aeromonas hydrophila AH-1 pathogenesis. Infect Immun 72:1248–1256. https://doi.org/10.1128/IAI.72.3.1248
Chacón MR, Soler L, Groisman EA, Guarro J, Figueras MJ (2004) Type III secretion system genes in clinical Aeromonas isolates. J Clin Microbiol 42:1285–1287. https://doi.org/10.1128/JCM.42.3.1285-1287.2004
Vilches S, Jimenez N, Tomás JM, Merino S (2009) Aeromonas hydrophila AH-3 type III secretion system expression and regulatory network. Appl Environ Microbiol 75:6382–6392. https://doi.org/10.1128/AEM.00222-09
Vilches S, Wilhelms M, Yu HB, Leung KY, Tomás JM, Merino S (2008) Aeromonas hydrophila AH-3 AexT is an ADP-ribosylating toxin secreted through the type III secretion system. Microb Pathog 44:1–12. https://doi.org/10.1016/j.micpath.2007.06.004
Sierra JC, Suarez G, Sha J, Foltz SM, Popov VL, Galindo CL, Garner HR, Chopra AK (2007) Biological characterization of a new type III secretion system effector from a clinical isolate of Aeromonas hydrophila—Part II. Microb Pathog 43:147–160. https://doi.org/10.1016/j.micpath.2007.05.003
Sierra JC, Suarez G, Sha J, Baze WB, Foltz SM, Chopra AK (2010) Unraveling the mechanism of action of a new type III secretion system effector AexU from Aeromonas hydrophila. Microb Pathog 49:122–134. https://doi.org/10.1016/j.micpath.2010.05.011
Matys J, Turska-Szewczuk A, Sroka-Bartnicka A (2020) Role of bacterial secretion systems and effector proteins - insights into Aeromonas pathogenicity mechanisms. Acta Biochim Pol 67:283–293. https://doi.org/10.18388/abp.2020_5410
Starliper CE, Schill WB, Schill, (2011) Flavobacterial Diseases: Columnaris Disease, Cold-water Disease and Bacterial Gill Disease. In: Woo PTK, Bruno DW (eds) Fish Diseases and Disorders, 2nd edn. CAB International Publishing, New York, pp 606–631
Soto E (2007) Genetic and virulence diversity of Flavobacterium columnare. Mississippi State
Soto E, Mauel MJ, Karsi A, Lawrence ML (2008) Genetic and virulence characterization of Flavobacterium columnare from channel catfish (Ictalurus punctatus). J Appl Microbiol 104:1302–1310. https://doi.org/10.1111/j.1365-2672.2007.03632.x
LaFrentz BR, Waldbieser GC, Welch TJ, Shoemaker CA (2014) Intragenomic heterogeneity in the 16S rRNA genes of Flavobacterium columnare and standard protocol for genomovar assignment. J Fish Dis 37:657–669. https://doi.org/10.1111/jfd.12166
Triyanto WH (1999) Genotypic diversity of strains of Flavobacterium columnare from diseased fishes. Fish Pathol 34:65–71. https://doi.org/10.3147/jsfp.34.65
Kumru S, Tekedar HC, Gulsoy N, Waldbieser GC, Lawrence ML, Karsi A (2017) Comparative analysis of the Flavobacterium columnare Genomovar I and II Genomes. Front Microbiol 8:1375. https://doi.org/10.3389/fmicb.2017.01375
Declercq AM, Tilleman L, Gansemans Y, De Witte C, Haesebrouck F, Van Nieuwerburgh F, Smet A, Decostere A (2021) Comparative genomics of Flavobacterium columnare unveils novel insights in virulence and antimicrobial resistance mechanisms. Vet Res 52:18. https://doi.org/10.1186/s13567-021-00899-w
Kumar G, Menanteau-Ledouble S, Saleh M, El-Matbouli M (2015) Yersinia ruckeri, the causative agent of enteric redmouth disease in fish. Vet Res 46:103. https://doi.org/10.1186/s13567-015-0238-4
Gunasena DK, Komrower JR, Macintyre S (2003) The Fish Pathogen Yersinia ruckeri Possesses a TTS System. In: Skurnik M, Bengoechea J, Granfors K (eds) The Genus Yersinia. Kluwer Academic Publishers, Boston, pp 105–107
Liu T, Wang K-Y, Wang J, Chen D-F, Huang X-L, Ouyang P, Geng Y, He Y, Zhou Y, Min J (2016) Genome sequence of the fish pathogen Yersinia ruckeri SC09 provides insights into niche adaptation and pathogenic mechanism. Int J Mol Sci 17:557. https://doi.org/10.3390/ijms17040557
Wrobel A, Leo JC, Linke D (2019) Overcoming fish defences: the virulence factors of Yersinia ruckeri. Genes (Basel) 10:700. https://doi.org/10.3390/genes10090700
Menanteau-Ledouble S, Lawrence ML, El-Matbouli M (2018) Invasion and replication of Yersinia ruckeri in fish cell cultures. BMC Vet Res 14:81. https://doi.org/10.1186/s12917-018-1408-1
Chevalier S, Bouffartigues E, Bodilis J, Maillot O, Lesouhaitier O, Feuilloley MGJ, Orange N, Dufour A, Cornelis P (2017) Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol Rev 41:698–722
Anantharajah A, Mingeot-Leclercq M-PP, Van Bambeke F (2016) Targeting the type three secretion system in Pseudomonas aeruginosa. Trends Pharmacol Sci 37:734–749. https://doi.org/10.1016/j.tips.2016.05.011
Khairnar K, Raut MP, Chandekar RH, Sanmukh SG, Paunikar WN (2013) Novel bacteriophage therapy for controlling metallo-beta-lactamase producing Pseudomonas aeruginosa infection in Catfish. BMC Vet Res 9:264. https://doi.org/10.1186/1746-6148-9-264
Algammal AM, Mabrok M, Sivaramasamy E, Youssef FM, Atwa MH, El-Kholy AW, Hetta HF, Hozzein WN (2020) Emerging MDR-Pseudomonas aeruginosa in fish commonly harbor oprL and toxA virulence genes and blaTEM, blaCTX-M, and tetA antibiotic-resistance genes. Sci Rep 10:15961. https://doi.org/10.1038/s41598-020-72264-4
Lombardi C, Tolchard J, Bouillot S, Signor L, Gebus C, Liebl D, Fenel D, Teulon JM, Brock J, Habenstein B, Pellequer JL, Faudry E, Loquet A, Attrée I, Dessen A, Job V (2019) Structural and functional characterization of the type three secretion system (T3SS) needle of Pseudomonas aeruginosa. Front Microbiol 10:573. https://doi.org/10.3389/fmicb.2019.00573
Matz C, Moreno AM, Alhede M, Manefield M, Hauser AR, Givskov M, Kjelleberg S (2008) Pseudomonas aeruginosa uses type III secretion system to kill biofilm-associated amoebae. ISME J 2:843–852. https://doi.org/10.1038/ismej.2008.47
Finck-Barbançon V, Goranson J, Zhu L, Sawa T, Wiener-Kronish JP, Fleiszig SM, Wu C, Mende-Mueller L, Frank DW (1997) ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury. Mol Microbiol 25:547–557. https://doi.org/10.1046/j.1365-2958.1997.4891851.x
Wood SJ, Goldufsky JW, Bello D, Masood S, Shafikhani SH (2015) Pseudomonas aeruginosa ExoT induces mitochondrial apoptosis in target host cells in a manner that depends on its GTPase-activating protein (GAP) domain activity. J Biol Chem 290:29063–29073. https://doi.org/10.1074/jbc.M115.689950
Kaminski A, Gupta KH, Goldufsky JW, Lee HW, Gupta V, Shafikhani SH (2018) Pseudomonas aeruginosa ExoS induces intrinsic apoptosis in target host cells in a manner that is dependent on its GAP domain activity. Sci Rep 8:14047. https://doi.org/10.1038/s41598-018-32491-2
Garrity-Ryan L, Shafikhani S, Balachandran P, Nguyen L, Oza J, Jakobsen T, Sargent J, Fang X, Cordwell S, Matthay MA, Engel JN (2004) The ADP ribosyltransferase domain of Pseudomonas aeruginosa ExoT contributes to its biological activities. Infect Immun 72:546–558. https://doi.org/10.1128/IAI.72.1.546-558.2004
Beckert U, Wolter S, Hartwig C, Bähre H, Kaever V, Ladant D, Frank DW, Seifert R (2014) ExoY from Pseudomonas aeruginosa is a nucleotidyl cyclase with preference for cGMP and cUMP formation. Biochem Biophys Res Commun 450:870–874. https://doi.org/10.1016/j.bbrc.2014.06.088
Fehr D, Casanova C, Liverman A, Blazkova H, Orth K, Dobbelaere D, Frey J, Burr SE (2006) AopP, a type III effector protein of Aeromonas salmonicida, inhibits the NF-κB signalling pathway. Microbiology 152:2809–2818. https://doi.org/10.1099/mic.0.28889-0
Fasciano AC, Shaban L, Mecsas J (2019) Promises and challenges of the type three secretion system injectisome as an antivirulence target. EcoSal Plus. https://doi.org/10.1128/ecosalplus.ESP-0032-2018
Fleitas Martínez O, Cardoso MH, Ribeiro SM, Franco OL (2019) Recent advances in anti-virulence therapeutic strategies with a focus on dismantling bacterial membrane microdomains, toxin neutralization, quorum-sensing interference and biofilm inhibition. Front Cell Infect Microbiol 9:74. https://doi.org/10.3389/fcimb.2019.00074
Ghanei-Motlagh R, Mohammadian T, Gharibi D, Khosravi M, Mahmoudi E, Zarea M, El-Matbouli M, Menanteau-Ledouble S (2021) Quorum quenching probiotics modulated digestive enzymes activity, growth performance, gut microflora, haemato-biochemical parameters and resistance against Vibrio harveyi in Asian seabass (Lates calcarifer). Aquaculture 531:735874. https://doi.org/10.1016/j.aquaculture.2020.735874
Ghanei-Motlagh R, Gharibi D, Mohammadian T, Khosravi M, Mahmoudi E, Zarea M, Menanteau-Ledouble S, El-Matbouli M (2021) Feed supplementation with quorum quenching probiotics with anti-virulence potential improved innate immune responses, antioxidant capacity and disease resistance in Asian seabass (Lates calcarifer). Aquaculture 535:736345. https://doi.org/10.1016/j.aquaculture.2021.736345
Gazi AD, Sarris PF, Fadouloglou VE, Charova SN, Mathioudakis N, Panopoulos NJ, Kokkinidis M (2012) Phylogenetic analysis of a gene cluster encoding an additional, rhizobial-like type III secretion system that is narrowly distributed among Pseudomonas syringae strains. BMC Microbiol 12:188. https://doi.org/10.1186/1471-2180-12-188
Lilic M, Vujanac M, Stebbins CE (2006) A common structural motif in the binding of virulence factors to bacterial secretion chaperones. Mol Cell 21:653–664. https://doi.org/10.1016/j.molcel.2006.01.026
Funding
Open Access funding for this article was provided by the University of Veterinary Medicine Vienna (Vetmeduni Vienna).
Author information
Authors and Affiliations
Contributions
HRE and SML performed the literature review and wrote the manuscript. SML and MEL conceived and designed the manuscript. MEL supervised the writing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Proteins belonging to the seven main T3SS families classified based on their homology. The table is reproduced with permission from Gazi et al. [181].
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Rahmatelahi, H., El-Matbouli, M. & Menanteau-Ledouble, S. Delivering the pain: an overview of the type III secretion system with special consideration for aquatic pathogens. Vet Res 52, 146 (2021). https://doi.org/10.1186/s13567-021-01015-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-021-01015-8