
Abstract
Background
Aeromonas salmonicida subsp. salmonicida, the etiologic agent of furunculosis, is a major pathogen of fisheries worldwide. Several virulence factors have been described, but the type-three secretion system (T3SS) is recognized as having a major effect on virulence by injecting effectors directly into fish cells. In this study we used high-throughput proteomics to display the differences between in vitro secretome of A. salmonicida wild-type (wt, hypervirulent, JF2267) and T3SS-deficient (isogenic ΔascV, extremely low-virulent, JF2747) strains in exponential and stationary phases of growth.
Results
Results confirmed the secretion of effectors AopH, AexT, AopP and AopO via T3SS, and for the first time demonstrated the impact of T3SS in secretion of Ati2, AopN and ExsE that are known as effectors in other pathogens. Translocators, needle subunits, Ati1, and AscX were also secreted in supernatants (SNs) dependent on T3SS. AopH, Ati2, AexT, AopB and AopD were in the top seven most abundant excreted proteins. EF-G, EF-Tu, DnaK, HtpG, PNPase, PepN and MdeA were moderately secreted in wt SNs and predicted to be putative T3 effectors by bioinformatics. Pta and ASA_P5G088 were increased in wt SNs and T3-associated in other bacteria. Ten conserved cytoplasmic proteins were more abundant in wt SNs than in the ΔascV mutant, but without any clear association to a secretion system. T1-secreted proteins were predominantly found in wt SNs: OmpAI, OmpK40, DegQ, insulinase ASA_0716, hypothetical ASA_0852 and ASA_3619. Presence of T3SS components in pellets was clearly decreased by ascV deletion, while no impact was observed on T1- and T2SS. Our results demonstrated that the ΔascV mutant strain excreted well-described (VapA, AerA, AerB, GCAT, Pla1, PlaC, TagA, Ahe2, GbpA and enolase) and yet uncharacterized potential toxins, adhesins and enzymes as much as or even more than the wt strain. Other putative important virulence factors were not detected.
Conclusions
We demonstrated the whole in vitro secretome and T3SS repertoire of hypervirulent A. salmonicida. Several toxins, adhesins and enzymes that are not part of the T3SS secretome were secreted to a higher extent in the extremely low-virulent ΔascV mutant. All together, our results show the high importance of an intact T3SS to initiate the furunculosis and offer new information about the pathogenesis.
Similar content being viewed by others


Background
Aeromonas salmonicida subsp. salmonicida, a gram-negative bacterium, is the etiologic agent of furunculosis, a frequent and major pathogen of fisheries worldwide which is generating significant economic losses related to deficits in zootechnical profits and the intensive use of antibiotics [1]. To date, several virulence factors have been characterized for A. salmonicida: the type three secretion system (T3SS) encoded on a large plasmid and described for the first time in the Aeromonas genus in our laboratory ten years ago [2, 3]; the surface layer protein VapA [4]; a type I pilus [5]; three type IV pilus systems [6, 7]; superoxide dismutases [8] and some extracellular proteins including serine protease (AspA) [9], glycerophospholipid:cholesterol acyltransferase (GCAT or SatA) [9, 10] and several hemolysins (aerolysins) [11]. Other putative virulence factors were identified without experimental evidence [12]. However, the T3SS is the only one recognized as having a major effect on virulence, as independent studies have shown that isogenic mutant strains for T3SS structural proteins are non-virulent both in vitro and in vivo [2, 13–16]. T3SS is also called “injectisome” because it enables the secretion and simultaneous injection of effector proteins produced in the prokaryotic cytoplasm across the bacterial envelope and then, through a needle and a translocon complex, into the target eukaryotic cells across their membrane [17]. Once injected in the eukaryotic cytosol, effector proteins are able to modulate cell signalling pathways, or alternatively disrupt the dynamics of the cytoskeleton, thereby modulating host cell biology for the benefit of the pathogen [17].
Currently, four different virulent effectors have been investigated for the A. salmonicida T3SS, and only two have been studied in detail: the bifunctional toxin AexT, which possesses a GTPase-activating domain acting on small monomeric GTPases of the Rho family and an ADP-ribosylating domain, which ADP-ribosylates both muscular and non-muscular actin [18, 19]; AopP, which inhibits the NF-κ B signaling pathway by preventing translocation of NF-k B into the nucleus of the target cells [20]. AopO, which is related to Yersinia YopO/YpkA [14] and AopH with similarity to Yersinia YopH [14], represent two further potential effectors that have been characterized in less detail. AexT, AopO and AopH toxins do not seem to be solely responsible for Aeromonas virulence because individual knock-out mutations of these genes [14] or a triple-effector knock-out mutant [21] keep a virulent phenotype or show only delayed virulence, such as in the case of ΔaexT mutants [14, 19]. Given that A. salmonicida mutants that are defective for T3SS fully lose their pathogenicity, we hypothesize that other important cytotoxic proteins might be injected by these Aeromonas nanosyringes into the fish cell cytoplasm.
The aim of this work was to use high-throughput proteomics to display the secretome of A. salmonicida subsp. salmonicida wild-type (wt, hypervirulent) and an isogenic T3SS-deficient mutant (ΔascV, extremely low-virulent) during the exponential-growth phase (GP) and the stationary phase (SP). In this article, which is the second part of the work, authors characterized the whole in vitro repertoire of T3SS effectors and new virulence factors of A. salmonicida. In the first part, “The Aeromonas salmonicida subsp. salmonicida exoproteome: global analysis, moonlighting proteins and putative antigens for vaccination against furunculosis”, the same authors focused on the general analysis of proteomics data, the presence of cytoplasmic proteins with putative moonlighting activities in supernatants and the identification of putative antigens for fish vaccination against furunculosis.
Results and discussion
A. salmonicida T3SS and comparison to other appendages
A. salmonicida subsp. salmonicida wt strain was previously shown to cause 80% - 100% mortality in rainbow trout at 500 cfu inoculated intraperitoneally, while the ΔascV deletion mutant derived thereof was shown to be non-virulent causing 0% mortality under the same conditions [15, 22]. In order to further show the strong attenuation due to the ΔascV deletion mutation, rainbow trout kept under the same conditions were challenged intraperitoneally with 108 cfu, an infectious dose which is not representative of what happens in natural infection. These fish showed only a slight mortality of 20% after 14 days post infection showing the high degree of attenuation obtained with the ΔascV mutation. We assume that the residual mortality observed in this experiment is solely due to the excessive load of bacteria applied.
We identified a total of 2136 A. salmonicida proteins with PMSS and LFQ values among the different experimental conditions (see Methods for explanations and the first part of this work for raw data) for 1861 and 2070 proteins respectively. These values correspond to a semi-quantitative abundance estimate of protein species present in SDS-PAGE gels and were used as a surrogate for the amount of secreted proteins in concentrated SNs and the amount of produced proteins in whole pellets.
In our MS analysis we identified 45 proteins of the A. salmonicida T3SS. The effectors should only be secreted or detected in higher quantity in wt SNs (in GP and SP) in comparison to the ΔascV mutant (Table 1). Our results confirmed the secretion of the well-described AopH, AexT, AopP and AopO effectors (Figure 1, Table 1). Moreover, we demonstrated the secretion of additional T3SS effectors for the first time. Ati2 (ASA_P5G045), an inositol polyphosphate 5-phosphatase already described as a putative T3SS effector [12], was strongly secreted in wt SNs (as much as AexT, 20 times more than in ΔascV mutant SN). Ati2 is homologous to the Vibrio parahaemolyticus T3SS effector VPA0450 and Photorhabdus luminescens Plu4615 (87% identity over 495 amino acids [aa]). This effector disrupts cytoskeletal binding sites on the inner surface of host membranes, causes plasma membrane blebbing and probably contributes to cell death by facilitating lysis [23]. Our data showed that Ati1 (ASA_P5G046), the chaperone of Ati2, was also secreted in wt SNs by the T3SS, whereas all other T3SS chaperones (SycE, SycH, SycO, AscY, Acr1, Acr2, AscB, AcrG, AscG, AscE, AscO and AcrR) were only present in pellets and were never secreted (Figure 2) suggesting that Ati1 might be injected with Ati2 into fish cells. AopN (ASA_P5G075) was secreted by the T3SS in wt SNs, but to a lower extent than the previous effectors. AopN homologues in other bacteria are T3SS effectors which play a role in virulence and can have a dual role: controlling the secretion of translocator proteins inside bacteria and suppressing immunity when T3 translocated inside host cells [24–26].

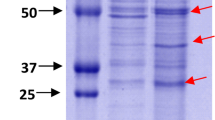
SDS gel electrophoresis of A. salmonicida subsp. salmonicida proteins from GP wt and Δ ascV mutant SNs stained with Coomassie blue. SDS gel electrophoresis of proteins from supernatants (SN) of wild-type (wt) and ΔascV mutant (mut) strains in exponential (GP) phase of growth. Proteins corresponding to the most abundant bands are indicated. The molecular weights (kDa) of the Protein Ladder are shown on both sides of the figure.

Ratios of different T3SS components in A. salmonicida wt versus mutant strain. The plot represents the logarithm base two of the ratios [wt/mut] PMSS values for each protein identified in pellet (X-axis) and SNs (Y-axis). For T3SS components, exponential growth phase (GP) values are represented by circles and stationary phase (SP) values by squares. Red, the T3SS effectors; dark green, T3SS chaperones; white, translocators; black, needle components; orange, OM secretin ring; light green, IM export ring; dark blue, C ring ATPase; light blue, transcriptional regulators. Values of other proteins in GP and SP are shown with dark blue triangles and yellow squares respectively. The hatched line represents the threshold that we used to identify proteins 4-fold more present in wt SNs.
AopH, Ati2 and AexT were the most secreted A. salmonicida proteins in wt SNs (GP or SP) (Figure 1, Additional files 1 and 2). When we calculated the ratio of [SN/pellet] quantities for each effector, we observed that AopP, AopH, AexT and Ati2 showed a high proportion in concentrated SNs, whereas this proportion was weak for AopO and AopN. This suggests that the in vitro secretion of AopO and AopN in wt SNs was significantly less efficient than AopP, AopH, AexT and Ati2.
We observed that AscX (ASA_P5G072) and ExsE (ASA_P5G062) were T3 secreted in wt SNs (Table 1). The same observation was made for YscX in Yersinia pestis [27]. YscX does not seem to be a T3SS effector, but it plays a role with its chaperone (YscY) and YscV in the export of needle components (YscF and YscI) [28]. In Pseudomonas aeruginosa, it was shown that the T3 secretion in extracellular medium and the T3 translocation into host cell of ExsE was required for transcriptional induction of the T3SS [29–31]. It is not known whether ExsE plays a role within the host cell.
Our proteomic analysis logically detected all translocon components (AopB, AopD, AcrV and AcrH) in A. salmonicida wt secretome (Figures 1 and 2, Table 1). AopB and AopD were among the top ten most abundant secreted proteins (Additional file 2). As expected, the elements of the T3SS needle (AscF, AscP, AscH and AscI) were also oversecreted in wt SNs and T3SS proteins of the OM ring (AscJ, AscD, AscC and ExsB), the inner membrane export apparatus (AscV, AscR, AscT and AscU) and the C ring/ATPase (AscL, AscK, AscN and AscQ) were only detected in pellets (Figure 2 and Additional files 1, 2 and 3).
Our study did not detect T3SS effectors AopX (homologuous to V. parahaemolyticus VopR [VP1683], P. luminescens plu4750) and ASA_0010 (homologuous to V. parahaemolyticus VopS [VP1686]) [32], suggesting that the mutations present in these genes in the genome of A. salmonicida A449 [12] and also in our wt strain prevent their production. However, the chaperone of VopS effector (ASA_0011) was detected, but only to a weak level in GP in the wt pellet.
From these results we concluded that our MS analysis localized 100% of T3SS components that are structurally linked to the bacteria and associated to pellets (cytoplasmic chaperones, OM, IM and C rings proteins) or T3 secreted and associated to SNs (effectors, translocon and needle elements) with effectiveness and accuracy. These results also support the idea that highly conserved cytoplasmic proteins unexpectedly present in A. salmonicida SNs and detailed in the first part of this work were not due to cell lysis.
The quantity of T3SS proteins was systematically lower in SP pellets (wt or mutant), and significantly lower in mutant pellets in comparison to wt (Additional files 1 and 3 for individual T3SS components), suggesting that the T3SS production was at a maximum when bacteria were in the phase of active multiplication and that the ΔascV knock-out mutation induced a strong down-regulation of the expression of many T3SS genes. AopD, AopB, AopH, AscV, Ati2, AcrV, AopO and AexT were the most abundant T3SS proteins present in the GP wt pellet (Additional file 3) and the difference in quantity (PMSS value) observed between the pellets of the wt and the ΔascV mutant in GP was confirmed by western blotting for AopD, AcrV and AexT (Additional file 4).
This underexpression of T3SS genes from different operons argues that the ascV deletion modulates the transcription regulation of several T3SS components and is not due to a polar effect. Strickingly, weak amounts of T3SS effectors/translocators were found in ΔascV mutant SNs (AopH, AexT, AopD, Ati2, AopP, AopN, AopB and ExsE by order of decreasing importance), but clearly to a lower extent than in wt SNs (Figure 3A and Additional files 1 and 2). As for the wt strain, the presence of these T3SS elements in mutant SNs was unlikely to be due to bacterial lysis given that (referred to the first part of this work for details): (i) ~90% of predicted cytoplasmic proteins in mutant pellets were never detected in SNs, (ii) GroEL, a marker of cell lysis, was among the most abundant proteins present in mutant pellets but was absent from SNs, and (iii) EF-Tu amount in mutant SNs decreased from GP to SP. The presence of T3SS effectors/translocators in mutant SNs was also unlikely to be due to a contamination between wt and mutant samples because, for example, the [wt/mutant] PMSS ratios of these T3SS-secreted components were 10-fold higher for AopP to 110-fold higher for AopB in GP SNs of wt when compared to ΔascV and were therefore not proportional. Burr and collaborators [2] did not detect AexT secretion in the ΔascV mutant SN, but they used unconcentrated SNs. Our samples were ~200 times more concentrated in this study. When we used total sum of PMSS values to calculate [intra bacterial effectors or translocators/T3SS structural components] we found that proportions were similar in wt and mutant strains (Figure 3B) assuming that, even if ascV was deleted, A. salmonicida kept the same proportion between the intracellular stock of effectors/translocators and the other T3SS structural components. As already mentioned, the proportion of [extra bacterial effectors or translocators/T3SS structural components] showed that the T3 secretion capacity was strongly impaired for the mutant strain during GP and SP, but this difference with the wt strain was weaker during the SP (Figures 3). This could mean that small amounts of effectors and translocators accumulated progressively in the mutant SNs along growth phases.

Proportion of T3SS effectors and translocators. The total PMSS value of T3SS effectors or translocators present in wt and mutant SNs during GP and SP are shown in diagram (A). Quantities were strongly decreased in mutant SNs but low levels of T3SS effectors/translocators were detected and increased during SP. Diagram (B), represents the proportion of [extra- or intrabacterial effectors or translocators/T3SS structural components] calculated with the total PMSS value of each category. These results showed that despite the ascV deletion, A. salmonicida kept the same proportion between intrabacterial effectors/translocators and the other components of the T3SS while strong differences were observed between wt and mutant for the proportion of extrabacterial effectors/translocators.
The mutant strain might continue to release these T3SS components in SNs, either from the resting structural T3SS components or by an alternative secretion pathway. Recent publications argue that the T3SS arose from an exaptation of the flagellum, i.e. the recruitment of part of the flagellum structure for the evolution of the new protein delivery function [33, 34] and, the secretion of T3SS effectors through flagella in the extracellular medium has been described in other bacteria [35, 36]. The secretion of effectors/translocators by this process is unlikely in A. salmonicida given that functional lateral and polar flagella were not detected (Figure 4), thus confirming the results of studies showing that operons coding for A. salmonicida flagella contain several mutations [12]. However, we could imagine that FlhA (ASA_1351, polar flagella) and/or LfhA (ASA_0352, lateral flagella), showing respectively 56% and 55% of similarity with AscV might partially supply the function of this T3SS component. Such possible interactions between FlhA and the T3SS have been described in Chlamydia pneumonia [37]. While no mutations are predicted in these genes in A. salmonicida their expression was not detected in our pellets, but we cannot exclude that they were expressed below the detection limit of our system as our proteomic analysis did not cover the total proteome (59% of proteins common to all Aeromonas sp. were identified). Another possibility is that two mechanisms of effectors/translocators secretion operate in parallel along phases of growth, the first would be actively dependent on intact T3SSs while the second, clearly less efficient, would explain the progressive accumulation of effectors/translocators in the extracellular medium of the mutant strain. The alternative secretion of T3SS effectors through classical/unclassical pathways has never been described contrary to major constituents of the OM ring which are transported to the periplasm by the Sec-dependent secretion pathway [38]. Furthermore, the presence of T3SS effectors in the periplasma [39] and OMVs [40] has rarely been described. Another possibility might be the formation of double-bilayer OMVs (diameter from 100 to 250 nm) containing cytosolic components, as recently described in Shewanella [41], but GroEL would have been detected in SNs. Another study showed that in the absence of the host cell, at least YopH (homologous to AopH), YopE (AexT) and YopB/YopD translocators were excreted homogeneously at the Yersinia surface without physical association with the injectisome [42]. While the T3-dependent secretion of effectors in SNs is well-characterized, it is not known if these OM-associated effectors are excreted at the bacterial surface through a T3SS-dependent pathway. As a result, further investigations are necessary to clarify this point.

Protein abundance of A. salmonicida appendages. The diagram represents total PMSS values of A. salmonicida secretion systems (T1- to T6SS), flagella (lateral and polar), pili (T1 pilus [Fim], T4 pili [Tap, Flp and Msh] and fimbriae (MatB). Only the structural components were taken into account for the T3SS. Logically, the strongest difference in protein amount was observed for the T3SS. No differences were observed in pellets for T1- (103 ABC-transporters) and T2SS. In SNs, the strong difference between wt and mutant strains for T1SS values is due to the higher amount of VapA in mutant SNs.
When we compared the total LFQ values of A. salmonicida secretion systems (T1- to T6SS), flagella (lateral and polar), pili (T1 pilus [Fim], T4 pili [Tap, Flp and Msh] and fimbriae (MatB), it was clear that the T3SS was the most expressed system by A. salmonicida (Figure 4). T1- and T2SS were expressed just as much in wt and mutant pellets, showing that their expression and function was not impaired by the knock-out mutation in ascV. All of the other systems (flagella, pili, fimbriae, T4-, T5- and T6SS) were either not expressed at all or were expressed to a lower level, suggesting that they could be impaired by mutations similar to the ones observed in the reference A449 strain [12].
Other putative virulence factors oversecreted in A. salmonicida wt SNs
We combined several thresholds to identify additional putative A. salmonicida T3SS effectors and T3-independent virulence factors. We targeted wt secreted proteins with PMSS values over 25, a PMSS or LFQ intensity 4-fold increased in the wt SN, and a PEP value inferior to 10-8 or equal to zero (Figure 2 and Table 2). We then performed bioinformatics analyses to predict whether a peptide signal for Sec-, Tat- or T3 dependent secretion was present in the N-terminal part of secreted proteins.
From 466 proteins detected in SNs, only 26 proteins were more abundant in wt than in mutant SNs, while their presence was approximatively similar in pellets. Among the first targeted proteins, seven were surprisingly designated by bioinformatics as T3 effectors (EF-G, EF-Tu, DnaK, HtpG, PNPase, MdeA, PepN and OpdA), and two proteins without a predicted motif for T3 secretion were shown to have homologues that are T3 secreted in other bacteria (Pta and ASA_P5G088) (Table 2). These proteins were secreted to a clear lesser extent than previously described T3SS effectors, and these results should therefore be interpreted with caution and need further investigations in order to confirm that they are (T3-) secreted. Strikingly, homologues of these proteins are present in eukaryotic cells, where they play fundamental roles and sometimes alternative (moonlighting) functions (EF-1α for EF-Tu [43], HSP70 and HSP90 for DnaK and HtpG [44–46], eukaryotic aminopeptidases and thimet oligopeptidase for PepN and OpdA [47–49]). For example, these molecular chaperones play a role in the virulence of other pathogens and are considered as new targets for therapy [50, 51]. It is tempting to assume that EF-G, EF-Tu, DnaK, HtpG, PepN and OpdA might be injected by A. salmonicida into host cells in order to interfere with these functions.
Polynucleotide phosphorylase PNPase has pleiotropic roles in bacteria such as degrading mRNA (degradosome) and mediating post-transcriptional regulation [52]. However, it was shown that PNPase was required for the optimal functioning of Yersinia T3SS and enhanced the ability of the bacterium to withstand the killing activities of murine macrophages [53]. In Salmonella enterica and Dickeya dadantii, PNPase downregulated the transcription of T3SS genes [54, 55].
Although they did not have the N-terminal motif for T3-secretion, the phosphate acetyl transferase (Pta, ASA_3402) and the putative α/β hydrolase ASA_P5G088 of A. salmonicida were targeted by our screening as putative T3SS effectors. In Salmonella, a Pta mutant showed that this enzyme was associated to virulence [56], and a recent study demonstrated that E. coli Pta (E2348C_2437, 83% similarity with A. salmonicida Pta) might be secreted by the T3SS [57]. Interestingly, homologues of ASA_P5G088 in V. parahaemolyticus (35% and 38% of similarity with VP1677 and VP1678) were T3 secreted [58].
Ten cytoplasmic proteins were more abundant in wt vs ΔascV mutant SNs, did not have any predicted signal for a secretion system and were not characterized as T3SS effectors in other bacteria (Table 2). TypA (or BipA) is a GTPase that was associated to virulence [59, 60] through regulation of the T3SS [61, 62]. Interestingly, even though the TypA N-terminal part does not contain a predicted signal for T3 secretion, it shares three conserved motifs with the N-terminal part of EF-G and EF-Tu. Unclearly, ribosomal protein 30S S1, 30S S6, 50S L24 and L3, IleS, LeuS, Tkt, AcnB, and WecB were more abundant in wt SNs. All of these components were discovered to be associated to the A. salmonicida surfacome and in the secretome of other bacteria (refer to the Additional file 8 of the Part 1 of this work, “The Aeromonas salmonicida subsp. salmonicida exoproteome: global analysis, moonlighting proteins and putative antigens for vaccination against furunculosis” for details). AcnB and WecB have homologous proteins that have been associated to the virulence in other bacteria (Additional file 5).
Six proteins with a predicted T1 peptide signal were systematically found either in higher amounts or only in wt SNs compared to the ΔascV mutant (Table 2). That was the case for OmpAI (ASA_1267) and OmpK40 (ASA_1544), which were linked to virulence in Aeromonas and other bacteria (Additional file 5). The presence of these OM proteins in SNs was not an artefact given that OmpAII (ASA_1266) was produced just as much in pellets as OmpAI but was never detected in SNs (Additional file 1). The periplasmic trypsin-like serine protease DegQ (ASA_0330), the insulinase ASA_0716 (zinc-dependent peptidase M16), the putative OM lipoprotein ASA_0852, and the putative ABC-type Fe3 + -hydroxamate transport system component ASA_3619 were also increased in wt SNs, and such proteins have also been related to virulence in other bacteria (Additional file 5). Interestingly, A. hydrophila homologues of ASA_0852 and ASA_3619 were found in all toxic extracellular product fractions of the bacterium [63].
Analysis of previously-described and newly detected putative virulence factors
Besides the T3SS, other virulence factors of A. salmonicida have been characterized (or predicted) in the literature, and certain conserved proteins are homologous to virulent toxins, adhesins and enzymes identified in other bacteria (Table 3, Additional file 5). We identified the tetragonal surface virulence array protein VapA, aerolysin AerA, hemolysin AerB, esterase SatA, extracellular phospholipase PlaA1, phospholipase PlaC, the metalloprotease/mucinase, serine protease Ahe2, chitin/ N-acetylglucosamine-binding protein (ASA_0604), extracellular nuclease (ASA_1199), enolase (ASA_3475), and outer membrane endopeptidase PepO. Our results showed that all these toxins and enzymes were secreted as much as or more as in the extremely low-virulent ΔascV mutant (Figure 5, Table 3 and Additional file 2) and they highlighted that an intact T3SS is primordial to initiate the disease. This observation is supported by studies demonstrating that the deletion of T3SS genes completely abolishes the virulence [2, 3, 13–16, 24, 64].

Correlation of protein contents between wt and T3SS-mutant. Each plot represents the PMSS (A and B) or LFQ (C and D) values for each protein identified in wt (X-axis) and/or mutant (Y-axis) strains, in supernatants (SNs, A and C) and pellets (B and D). Values of exponential growth phase (GP) are dark blue triangles and stationary phase (SP) values are yellow squares. The global distribution of wt vs mutant protein values was linear in all conditions, but with a larger repartition in SNs than in pellets, thereby indicating differences in protein secretion between wt and ΔascV strains. Red values = T3SS proteins; orange = VapA, AerA, AerB, Ahe2, SatA and Asx; light blue = TagA, microbial collagenase, extracellular desoxyribonuclease ASA_1199, PlaA1, PlaC, Amy1, CdxA, ChiB, Chi2 and AmyA.
Our proteomic study also characterized, the secretion in SNs of other putative virulent toxins, adhesins and enzymes conserved among Aeromonas sp. for the first time (Table 3, Additional file 5), in decreasing order of quantity in SNs (Additional file 2): the large RTX (repeats in toxin) exoprotein Asx (ASA_0826), a microbial collagenase (ASA_3723), an unknown protein ASA_2541 that could be co-expressed/secreted with Ahe2, the nuclease NucH (ASA_2206), a leucine aminopeptidase (ASA_3073), two large unknown proteins with a Ig-like domain (homologues to pRA1_0073 in IncA/C plasmids and ASA_P4G163), chitinases CdxA, Chi2 (ASA_2142) and ChiB (ASA_3320), the solute receptor TAXI (TRAP-associated extracytoplasmic immunogenic) of a TRAP transporter (ASA_3982), the immune inhibitor A metalloprotease PrtV (ASA_0849), the metalloprotease LasA, a LysM domain-containing protein (ASA_1027), the hypothetical GlyGly-CTERM protein ASA_1998, the micrococcal nuclease (SNase-like) ASA_P4G031, the azurin, and the Type I pilus subunit FimD. All of these proteins were as much as or more secreted in mutant SNs (Table 3 and Additional file 2), highlighting once again that an intact T3SS is primordial to initiate the disease. The putative hemolysin ASA_1523 was only detected in pellets and in higher quantity in the mutant strain.
In the genome of A. salmonicida A449, Zonular Occludens Toxins (Zot, ASA_2003 and ASA_2015), elastase AhpB and toxic extracellular endopeptidase AsaP1 genes are impaired by deletions and insertion elements. According to these observations, we did not detect any polypeptides for these proteins in our MS experiments, suggesting that they would be also disrupted in our A. salmonicida strain. Furthermore, the insecticidal cytolytic delta-endotoxin (ASA_2128), putative RTX toxins (ASA_0127, ASA_1674 and ASA_1675), a secreted metalloprotease (ASA_1723) and the pullulanase PulA were not identified, and their expression might be induced in the host. Finally, 15 prophage proteins were identified in pellets (12% of prophage genes detected in the genome of the reference strain A449) and only one (ASA_2013) was detected in SNs, but without any significant differences between the wt and mutant strains.
Conclusions
The comparison by high-throughput proteomics of A. salmonicida secretomes from wt and T3SS-deficient strains is a powerful method that gave us the opportunity (i) to characterize the full in vitro repertoire of T3SS effectors represented mainly by AopH, Ati2, AexT, AopP, AopO, AopN and ExsE, (ii) to identify new putative virulence factors that are secreted in the extracellular medium or might be translocated into the host cell by the T3SS or alternative mechanisms, and (iii) to confirm that A. salmonicida secreted toxins, adhesins and enzymes that have been described until now and are additionally found in this study are secreted to a higher extent in the extremely low-virulent ΔascV mutant.
Our results also clearly show that the deletion of one gene (ascV in this study) can induce the down-regulation of several other genes (only associated to the T3SS in our study), not necessary transcriptionally linked in the same operon. To respect the molecular Koch’s postulates, we can conclude from this study that each work investigating phenotypic characters by site-directed mutagenesis should ideally be completed by a larger analysis studying the impact of the mutation over the global gene expression.
Due to the fact that we studied in vitro secretomes, T3SS effectors that we have found might be considered as the first line of weapons that A. salmonicida uses to invade fish and initiate the disease. Inside the salmonid, bacteria might induce the expression of genes specific to the A. salmonicida species and present in genomic islands (such as the cluster of genes [ASA_1049 to ASA_1052] homologous to the Vibrio Seventh pandemic Island-I [VSP-I]) that might be necessary to survive in new environments [65]. Interestingly, T3SS effectors predicted by bioinformatics are two times more abundant in genomic regions specific to A. salmonicida (15% of specific genes) than in genetic regions common to all Aeromonas species (8% of common genes). Further proteomics studies will be necessary in order to confirm the in vivo A. salmonicida secretome.
Methods
Cell culture and preparation of bacterial supernatants and pellets for LC-MS/MS
Aeromonas salmonicida wt and ΔascV mutant strains used in this study were characterized in a previous work [15]. To get A. salmonicida wt cultures into a maximum T3SS activation state, we used JF2267 strain which was freshly reisolated from an experimentally infected dead fish (JF5054). This re-isolated strain was highly virulent, since intraperitoneal inoculation of only 500 cfu per fish was sufficient to induce 70 to 80% of mortality in challenge assays [22]. The ΔascV mutant strain JF2747 is considered to have extremely low-virulence because 105 cfu/fish induced no mortality [15], and 108 cfu/fish induced a weak mortality of only 20%.
To precipitate and concentrate proteins from the supernatant of wt and ΔascV A. salmonicida, 50 ml of TSB medium were inoculated with 109 bacteria and cultivated at 18°C under shaking (160 rpm) in the presence of protease inhibitors (Complete, Roche Diagnostics). The bacterial growth was stopped during the exponential phase of growth (DO600 = ~1.5) and the stationary phase (DO600 >2.0). Supernatants were separated from bacterial pellets by centrifugation (6.000 × g, 10 min, 4°C) and filtration through a 0.22 μM Acrodisc filter (low protein binding, PALL Life Sciences). The bacterial pellets were resuspended in 10 ml of PBS, and 250 μL of these solutions were mixed with 250 μL of SDS loading buffer and heated at 100°C for 5 min. To precipitate proteins from supernatants, 12.5 ml of 100% ice-cold trichloroacetic acid were added to the solutions (20% final concentration), then immediately vortexed and incubated overnight on ice. Supernatants were removed and brown protein pellets were suspended and washed several times by centrifugation in ice-cold 100% acetone in 2 ml Eppendorf tubes (low binding protein). Finally, the pellets were dried, diluted in 250 μL of SDS loading buffer (~200 times concentration) and heated at 100°C for 5 min. Proteins were separated in non-adjacent wells (to avoid well to well contamination) on 15% acrylamide SDS-PAGE gels and stained with Coomassie. One run for each of the eight biological conditions (wt vs mutant, GP vs SP and SN vs pellet) was completely sliced from the stacking gel to the buffer front in 20 to 25 bands, and each band was cut into small (~1 mm3) cubes for protein in-gel digestion and MS analysis, as described elsewhere [66, 67]. Peptide sequencing was made on a LTQ Orbitrap XL mass spectrometer (ThermoFisher Scientific, Bremen; Germany) equipped with a Rheos Allegro nano flow system with AFM flow splitting (Flux Instruments, Reinach; Switzerland) and a nano electrospray ion source operated at a voltage of 1.7 kV. Peptide separation was performed on a Magic C18 column (5 μm, 100 Å, 0.075 × 70 mm) using a flow rate of ~400 nL/min and a linear gradient of 5 to 40% acetonitrile in water/0.1% (v/v) formic acid during 60 min.
The mass spectrometry proteomics data were deposited to the ProteomeXchange Consortium (http://proteomecentral.proteomexchange.org) via the PRIDE partner repository [68] with the dataset identifier PXD000429 and DOI 10.6019/PXD000429.
LC-MS/MS data interpretation
LC-MS/MS data interpretation was made against the current UniProtKB database release (2012_06) of all known A. salmonicida protein sequences. Two methods of relative protein quantification were used. The peptide-matching score summation (PMSS) is a label-free technique that assumes ideal scoring for proteins as the summative of the identification scores of their constituent peptides freed upon digestion. A higher score represents a more abundant protein [69]. The EasyProt search algorithm [70] was used for this, as described in [67]. The obtained mass spectrometric raw data were also analyzed with MaxQuant, version 1.2.2.5 [71], and its label-free quantitation (LFQ) algorithms [72] allowed quantitative comparisons. MaxQuant settings were as follows: Accepted false discovery rates at peptide, modified peptide and protein level were set at 1% using the reversed sequence database. Carbamidomethylation on Cys was set as a fixed modification. Oxidation of Met, acetylation on protein N-terminus, and phosphorylation on Set/Thr/Tyr were set as variable modifications with a precursor mass tolerance of 6 ppm in the main search, while only oxidation and acetylation with a mass accuracy of 20 ppm was used in the first search. Trypsin cleavage specificity was set at full with a maximum 2 missed cleavages and the allowance of up to three modifications per peptide of length between 6–25 amino acids. Fragment spectra were filtered to the 6 most intense peaks per 100 Da mass windows and searched with a mass tolerance of 0.5 Da. Protein identifications were accepted with at least 2 razor and unique peptide identifications. For label free quantification (LFQ), at least 2 unmodified or acetylated protein N-terminal peptides were required, and matching within a 2 minute time frame between samples was allowed. Only proteins with significant increased PMSS and LFQ values in GP and SP of wt vs mutant SNs were developed in the text.
Bioinformatics analysis
Detection of signal sequences for secretion was carried out using the SignalP 4.1 server (http://www.cbs.dtu.dk/services/SignalP/) [73], TatP 1.0 server (http://www.cbs.dtu.dk/services/TatP/) [74] and the T3SS effector prediction softwares from Modlab® (http://gecco.org.chemie.uni-frankfurt.de/T3SS_prediction/T3SS_prediction.html) [75] and EffectiveT3 (http://www.effectors.org/) [76]. The list of A. salmonicida ABC transporters was provided by ABCdb (https://www-abcdb.biotoul.fr/) [77] and prohage genes by PHAST (http://phast.wishartlab.com/index.html) [78].
Abbreviations
- APC:
-
Antigen presenting cell
- CTL:
-
Cytotoxic T lymphocytes
- GP:
-
Exponential phase of growth
- LFQ:
-
Label-free quantitation
- PMSS:
-
Peptide-matching score summation
- SN:
-
Supernatant
- SP:
-
Stationary phase of growth
- T3SS:
-
Type-three secretion system
- wt:
-
wild-type.
References
Janda JM, Abbott SL: The genus Aeromonas : taxonomy, pathogenicity, and infection. Clin Microbiol Rev 2010, 23: 35–73. 10.1128/CMR.00039-09
Burr SE, Stuber K, Wahli T, Frey J: Evidence for a type III secretion system in Aeromonas salmonicida subsp. salmonicida . J Bacteriol 2002, 184: 5966–5970. 10.1128/JB.184.21.5966-5970.2002
Stuber K, Burr SE, Braun M, Wahli T, Frey J: Type III secretion genes in Aeromonas salmonicida subsp salmonicida are located on a large thermolabile virulence plasmid. J Clin Microbiol 2003, 41: 3854–3856. 10.1128/JCM.41.8.3854-3856.2003
Noonan B, Trust TJ: The synthesis, secretion and role in virulence of the paracrystalline surface protein layers of Aeromonas salmonicida and A. hydrophila . FEMS Microbiol Let 1997, 154: 1–7. 10.1111/j.1574-6968.1997.tb12616.x
Dacanay A, Boyd JM, Fast MD, Knickle LC, Reith ME: Aeromonas salmonicida Type I pilus system contributes to host colonization but not invasion. Dis Aq Org 2010, 88: 199–206.
Masada CL, LaPatra SE, Morton AW, Strom MS: An Aeromonas salmonicida type IV pilin is required for virulence in rainbow trout Oncorhynchus mykiss . Dis Aq Org 2002, 51: 13–25.
Boyd JM, Dacanay A, Knickle LC, Touhami A, Brown LL, Jericho MH, Johnson SC, Reith M: Contribution of type IV pili to the virulence of Aeromonas salmonicida subsp salmonicida in Atlantic salmon ( Salmo salar l.). Infect Immun 2008, 76: 1445–1455. 10.1128/IAI.01019-07
Dacanay A, Johnson SC, Bjornsdottir R, Ebanks RO, Ross NW, Reith M, Singh RK, Hiu J, Brown LL: Molecular characterization and quantitative analysis of superoxide dismutases in virulent and avirulent strains of Aeromonas salmonicida subsp salmonicida . J Bact 2003, 185: 4336–4344. 10.1128/JB.185.15.4336-4344.2003
Vipond R, Bricknell IR, Durant E, Bowden TJ, Ellis AE, Smith M, MacIntyre S: Defined deletion mutants demonstrate that the major secreted toxins are not essential for the virulence of Aeromonas salmonicida . Infect Immun 1998, 66: 1990–1998.
Lee KK, Ellis AE: Glycerophospholipid-cholesterol acyltransferase complexed with lipopolysaccharide (Lps) is a major lethal exotoxin and cytolysin of Aeromonas salmonicida - Lps stabilizes and enhances toxicity of the enzyme. J Bact 1990, 172: 5382–5393.
Hirono I, Aoki T: Cloning and characterization of 3 hemolysin genes from Aeromonas salmonicida . Microb Pathogenesis 1993, 15: 269–282. 10.1006/mpat.1993.1077
Reith ME, Singh RK, Curtis B, Boyd JM, Bouevitch A, Kimball J, Munholland J, Murphy C, Sarty D, Williams J, et al.: The genome of Aeromonas salmonicida subsp salmonicida A449: insights into the evolution of a fish pathogen. BMC Genomics 2008, 9: 1–15. 10.1186/1471-2164-9-1
Burr SE, Wahli T, Segner H, Pugovkin D, Frey J: Association of type III secretion genes with virulence of Aeromonas salmonicida subsp salmonicida . Dis Aq Org 2003, 57: 167–171.
Dacanay A, Knickle L, Solanky KS, Boyd JM, Walter JA, Brown LL, Johnson SC, Reith M: Contribution of the type III secretion system (TTSS) to virulence of Aeromonas salmonicida subsp salmonicida . Microbiology - SGM 2006, 152: 1847–1856. 10.1099/mic.0.28768-0
Burr SE, Pugovkin D, Wahli T, Segner H, Frey J: Attenuated virulence of an Aeromonas salmonicida subsp salmonicida type III secretion mutant in a rainbow trout model. Microbiology - SGM 2005, 151: 2111–2118. 10.1099/mic.0.27926-0
Daher RK, Filion G, Tan SGE, Dallaire-Dufresne S, Paquet VE, Charette SJ: Alteration of virulence factors and rearrangement of pAsa5 plasmid caused by the growth of Aeromonas salmonicida in stressful conditions. Vet Microbiol 2011, 152: 353–360. 10.1016/j.vetmic.2011.04.034
Marlovits TC, Stebbins CE: Type III secretion systems shape up as they ship out. Curr Opin Microbiol 2010, 13: 47–52. 10.1016/j.mib.2009.11.001
Burr SE, Stuber K, Frey J: The ADP-ribosylating toxin, AexT, from Aeromonas salmonicida subsp salmonicida is translocated via a type III secretion pathway. J Bact 2003, 185: 6583–6591. 10.1128/JB.185.22.6583-6591.2003
Fehr D, Burr SE, Gibert M, d'Alayer J, Frey J, Popoff MR: Aeromonas exoenzyme T of Aeromonas salmonicida is a bifunctional protein that targets the host cytoskeleton. J Biol Chem 2007, 282: 28843–28852. 10.1074/jbc.M704797200
Fehr D, Casanova C, Liverman A, Blazkova H, Orth K, Dobbelaere D, Frey J, Burr SE: AopP, a type III effector protein of Aeromonas salmonicida , inhibits the NF- kappa B signalling pathway. Microbiology 2006, 152: 2809–2818. 10.1099/mic.0.28889-0
Fast MD, Tse B, Boyd JM, Johnson SC: Mutations in the Aeromonas salmonicida subsp salmonicida type III secretion system affect Atlantic salmon leucocyte activation and downstream immune responses. Fish Shellfish Immun 2009, 27: 721–728. 10.1016/j.fsi.2009.09.009
Vanden Bergh P, Burr SE, Benedicenti O, von Siebenthal B, Frey J, Wahli T: Antigens of the type-three secretion system of A. salmonicida subsp. salmonicida prevent protective immunity in rainbow trout. Vaccine in press
Broberg CA, Zhang LL, Gonzalez H, Laskowski-Arce MA, Orth K: A Vibrio effector protein is an inositol phosphatase and disrupts host cell membrane integrity. Science 2010, 329: 1660–1662. 10.1126/science.1192850
Crabill E, Karpisek A, Alfano JR: The Pseudomonas syringae HrpJ protein controls the secretion of type III translocator proteins and has a virulence role inside plant cells. Mol Microbiol 2012, 85: 225–238. 10.1111/j.1365-2958.2012.08097.x
Archuleta TL, Du YQ, English CA, Lory S, Lesser C, Ohi MD, Ohi R, Spiller BW: The Chlamydia effector chlamydial outer protein N (CopN) sequesters tubulin and prevents microtubule assembly. J Biol Chem 2011, 286: 33992–33998. 10.1074/jbc.M111.258426
Nagamatsu K, Kuwae A, Konaka T, Nagai S, Yoshida S, Eguchi M, Watanabe M, Mimuro H, Koyasu S, Abe A: Bordetella evades the host immune system by inducing IL-10 through a type III effector, BopN. J Exp Med 2009, 206: 3073–3088. 10.1084/jem.20090494
Day JB, Plano GV: The Yersinia pestis YscY protein directly binds YscX, a secreted component of the type III secretion machinery. J Bact 2000, 182: 1834–1843. 10.1128/JB.182.7.1834-1843.2000
Diepold A, Wiesand U, Amstutz M, Cornelis GR: Assembly of the Yersinia injectisome: the missing pieces. Mol Microbiol 2012, 85: 878–892. 10.1111/j.1365-2958.2012.08146.x
Urbanowski ML, Lykken GL, Yahr TL: A secreted regulatory protein couples transcription to the secretory activity of the Pseudomonas aeruginosa type III secretion system. Proc Natl Acad Sci USA 2005, 102: 9930–9935. 10.1073/pnas.0504405102
Rietsch A, Vallet-Gely I, Dove SL, Mekalanos JJ: ExsE, a secreted regulator of type III secretion genes in Pseudomonas aeruginosa . Proc Natl Acad Sci USA 2005, 102: 8006–8011. 10.1073/pnas.0503005102
Urbanowski ML, Brutinel ED, Yahr TL: Translocation of ExsE into Chinese hamster ovary cells is required for transcriptional induction of the Pseudomonas aeruginosa type III secretion system. Infect Immun 2007, 75: 4432–4439. 10.1128/IAI.00664-07
Broberg CA, Calder TJ, Orth K: Vibrio parahaemolyticus cell biology and pathogenicity determinants. Microbes Infect 2011, 13: 992–1001. 10.1016/j.micinf.2011.06.013
Abby SS, Rocha EPC: The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet 2012, 8: 1–15.
Erhardt M, Namba K, Hughes KT: Bacterial nanomachines: the flagellum and type III injectisome. Cold Spring Harb Perspect Biol 2010, 2: 1–23.
Young GM, Schmiel DH, Miller VL: A new pathway for the secretion of virulence factors by bacteria: The flagellar export apparatus functions as a protein-secretion system. Proc Natl Acad Sci USA 1999, 96: 6456–6461. 10.1073/pnas.96.11.6456
Ehrbar K, Winnen B, Hardt WD: The chaperone binding domain of SopE inhibits transport via flagellar and SPI-1 TTSS in the absence of InvB. Mol Microbiol 2006, 59: 248–264. 10.1111/j.1365-2958.2005.04931.x
Stone CB, Bulir DC, Gilchrist JD, Toor RK, Mahony JB: Interactions between flagellar and type III secretion proteins in Chlamydia pneumoniae . BMC Microbiol 2010, 10: 1–12. 10.1186/1471-2180-10-1
Schroeder GN, Hilbi H: Molecular pathogenesis of Shigella spp.: controlling host cell signaling, invasion, and death by type III secretion. Clin Microbiol Rev 2008, 21: 134–156. 10.1128/CMR.00032-07
Nilles ML, Williams AW, Skrzypek E, Straley SC: Yersinia pestis LcrV forms a stable complex with LcrG and may have a secretion-related regulatory role in the low-Ca2+ response. J Bact 1997, 179: 1307–1316.
Chowdhury C, Jagannadham MV: Virulence factors are released in association with outer membrane vesicles of Pseudomonas syringae pv. tomato T1 during normal growth. Biochim Biophys Acta-Proteins and Proteomics 2013, 1834: 231–239. 10.1016/j.bbapap.2012.09.015
Perez-Cruz C, Carrion O, Delgado L, Martinez G, Lopez-Iglesias C, Mercade E: New type of outer membrane vesicle produced by the Gram-negative bacterium Shewanella vesiculosa M7T: implications for DNA content. Appl Environ Microbiol 2013, 79: 1874–1881. 10.1128/AEM.03657-12
Akopyan K, Edgren T, Wang-Edgren H, Rosqvist R, Fahlgren A, Wolf-Watz H, Fallman M: Translocation of surface-localized effectors in type III secretion. Proc Natl Acad Sci USA 2011, 108: 1639–1644. 10.1073/pnas.1013888108
Gonen H, Smith CE, Siegel NR, Kahana C, Merrick WC, Chakraburtty K, Schwartz AL, Ciechanover A: Protein synthesis elongation-factor Ef-1- alpha is essential for ubiquitin-dependent degradation of certain N- alpha -acetylated proteins and may be substituted for by the bacterial elongation-factor Ef-Tu. Proc Natl Acad Sci USA 1994, 91: 7648–7652. 10.1073/pnas.91.16.7648
Tsan MF, Gao B: Heat shock proteins and immune system. J Leukoc Biol 2009, 85: 905–910. 10.1189/jlb.0109005
Roberts RJ, Agius C, Saliba C, Bossier P, Sung YY: Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: a review. J Fish Dis 2010, 33: 789–801. 10.1111/j.1365-2761.2010.01183.x
Tamura Y, Torigoe T, Kukita K, Saito K, Okuya K, Kutomi G, Hirata K, Sato N: Heat-shock proteins as endogenous ligands building a bridge between innate and adaptive immunity. Immunotherapy 2012, 4: 841–852. 10.2217/imt.12.75
Rock KL, York IA, Goldberg AL: Post-proteasomal antigen processing for major histocompatibility complex class I presentation. Nat Immunol 2004, 5: 670–677. 10.1038/ni1089
Kim E, Kwak H, Ahn K: Cytosolic aminopeptidases influence MHC class I-mediated antigen presentation in an allele-dependent manner. J Immunol 2009, 183: 7379–7387. 10.4049/jimmunol.0901489
Kessler JH, Khan S, Seifert U, Le Gall S, Chow KM, Paschen A, Bres-Vloemans SA, de Ru A, van Montfoort N, Franken KLMC, et al.: Antigen processing by nardilysin and thimet oligopeptidase generates cytotoxic T cell epitopes. Nat Immunol 2011, 12: 45–67. 10.1038/ni.1974
Neckers L, Tatu U: Molecular chaperones in pathogen virulence: emerging new targets for therapy. Cell Host Microbe 2008, 4: 519–527. 10.1016/j.chom.2008.10.011
Kulzer S, Charnaud S, Dagan T, Riedel J, Mandal P, Pesce ER, Blatch GL, Crabb BS, Gilson PR, Przyborski JM: Plasmodium falciparum -encoded exported hsp70/hsp40 chaperone/co-chaperone complexes within the host erythrocyte. Cell Microbiol 2012, 14: 1784–1795. 10.1111/j.1462-5822.2012.01840.x
Carpousis AJ: The RNA degradosome of Escherichia coli : an mRNA-degrading machine assembled on RNase E. Annu Rev Microbiol 2007, 61: 71–87. 10.1146/annurev.micro.61.080706.093440
Rosenzweig JA, Schesser K: Polynucleotide phosphorylase and the T3SS. Adv Exp Med Biol 2007, 603: 217–224. 10.1007/978-0-387-72124-8_19
Ygberg SE, Clements MO, Rytkonen A, Thompson A, Holden DW, Hinton JCD, Rhen M: Polynucleotide phosphorylase negatively controls spv virulence gene expression in Salmonella enterica . Infect Immun 2006, 74: 1243–1254. 10.1128/IAI.74.2.1243-1254.2006
Zeng QA, Ibekwe AM, Biddle E, Yang CH: Regulatory mechanisms of exoribonuclease PNPase and regulatory small RNA on T3SS of Dickeya dadantii . Mol Plant Microbe Interact 2010, 23: 1345–1355. 10.1094/MPMI-03-10-0063
Kim YR, Brinsmade SR, Yang Z, Escalante-Semerena J, Fierer J: Mutation of phosphotransacetylase but not isocitrate lyase reduces the virulence of Salmonella entetica serovar typhimurium in mice. Infect Immun 2006, 74: 2498–2502. 10.1128/IAI.74.4.2498-2502.2006
Deng WY, Yu HB, de Hoog CL, Stoynov N, Li YL, Foster LJ, Finlay BB: Quantitative proteomic analysis of type III secretome of enteropathogenic Escherichia coli reveals an expanded effector repertoire for attaching/effacing bacterial pathogens. Mol Cell Proteomics 2012, 11: 692–709. 10.1074/mcp.M111.013672
Zhou XH, Nydam SD, Christensen JE, Konkel ME, Orfe L, Friel P, Call DR: Identification of potential type-III secretion proteins via heterologous expression of Vibrio parahaemolyticus DNA. Appl Environ Microbiol 2012, 78: 3492–3494. 10.1128/AEM.07977-11
Chaudhuri RR, Peters SE, Pleasance SJ, Northen H, Willers C, Paterson GK, Cone DB, Allen AG, Owen PJ, Shalom G, et al.: Comprehensive identification of Salmonella enterica serovar typhimurium genes required for infection of BALB/c mice. PLoS Pathog 2009, 5: 1–13.
Sabbagh SC, Lepage C, McClelland M, Daigle F: Selection of Salmonella enterica serovar typhi genes involved during interaction with human macrophages by screening of a transposon mutant library. PloS One 2012, 7: 1–13.
Grant AJ, Farris M, Alefounder P, Williams PH, Woodward MJ, O'Connor CD: Co-ordination of pathogenicity island expression by the BipA GTPase in enteropathogenic Escherichia coli (EPEC). Mol Microbiol 2003, 48: 507–521. 10.1046/j.1365-2958.2003.t01-1-03447.x
Neidig A, Yeung AT, Rosay T, Tettmann B, Strempel N, Rueger M, Lesouhaitier O, Overhage J: TypA is involved in virulence, antimicrobial resistance and biofilm formation in Pseudomonas aeruginosa . BMC Microbiol 2013, 13: 77. 10.1186/1471-2180-13-77
Pridgeon JW, Klesius PH, Song L, Zhang D, Kojima K, Mobley JA: Identification, virulence, and mass spectrometry of toxic ECP fractions of West Alabama isolates of Aeromonas hydrophila obtained from a 2010 disease outbreak. Vet Microbiol 2013, 164: 336–343. 10.1016/j.vetmic.2013.02.020
Tanaka KH, Dallaire-Dufresne S, Daher RK, Frenette M, Charette SJ: An insertion sequence-dependent plasmid rearrangement in Aeromonas salmonicida causes the loss of the type three secretion system. Plos One 2012, 7: 1–8.
Studer N, Frey J, Vanden Bergh P: Clustering subspecies of Aeromonas salmonicida using IS630 typing. BMC Microbiol 2013, 13: 36. 10.1186/1471-2180-13-36
Gunasekera K, Wuthrich D, Braga-Lagache S, Heller M, Ochsenreiter T: Proteome remodelling during development from blood to insect-form Trypanosoma brucei quantified by SILAC and mass spectrometry. BMC Genomics 2012, 13: 556. 10.1186/1471-2164-13-556
Al Kaabi A, Traupe T, Stutz M, Buchs N, Heller M: Cause or effect of arteriogenesis: compositional alterations of microparticles from CAD patients undergoing external counterpulsation therapy. PloS One 2012, 7: 1–10.
Vizcaino JA, Cote RG, Csordas A, Dianes JA, Fabregat A, Foster JM, Griss J, Alpi E, Birim M, Contell J, et al.: The Proteomics Identifications (PRIDE) database and associated tools: status in 2013. Nucleic Acids Res 2013, 41: D1063-D1069. 10.1093/nar/gks1262
Stalder D, Haeberli A, Heller M: Evaluation of reproducibility of protein identification results after multidimensional human serum protein separation. Proteomics 2008, 8: 414–424. 10.1002/pmic.200700527
Gluck F, Hoogland C, Antinori P, Robin X, Nikitin F, Zufferey A, Pasquarello C, Fetaud V, Dayon L, Muller M, et al.: EasyProt - An easy-to-use graphical platform for proteomics data analysis. J Proteomics 2013, 79: 146–160.
Cox J, Mann M: MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol 2008, 26: 1367–1372. 10.1038/nbt.1511
Luber CA, Cox J, Lauterbach H, Fancke B, Selbach M, Tschopp J, Akira S, Wiegand M, Hochrein H, O'Keeffe M, et al.: Quantitative proteomics reveals subset-specific viral recognition in dendritic cells. Immunity 2010, 32: 279–289. 10.1016/j.immuni.2010.01.013
Petersen TN, Brunak S, von Heijne G, Nielsen H: SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods 2011, 8: 785–786. 10.1038/nmeth.1701
Bendtsen JD, Nielsen H, Widdick D, Palmer T, Brunak S: Prediction of twin-arginine signal peptides. BMC Bioinformatics 2005, 6: 167. 10.1186/1471-2105-6-167
Lower M, Schneider G: Prediction of type III secretion signals in genomes of gram-negative bacteria. PloS One 2009, 4: 1–9. 10.1371/journal.pone.0005361
Jehl MA, Arnold R, Rattei T: Effective - a database of predicted secreted bacterial proteins. Nucleic Acids Res 2011, 39: D591-D595. 10.1093/nar/gkq1154
Fichant G, Basse MJ, Quentin Y: ABCdb: an online resource for ABC transporter repertories from sequenced archaeal and bacterial genomes. FEMS Microbiol Let 2006, 256: 333–339. 10.1111/j.1574-6968.2006.00139.x
Zhou Y, Liang Y, Lynch KH, Dennis JJ, Wishart DS: PHAST: a fast phage search tool. Nucleic Acids Res 2011, 39: W347-W352. 10.1093/nar/gkr485
Acknowledgements
This research was funded by the Swiss National Science Foundation grant no. 31003A-135808. We are especially grateful to Dr. Thomas Whali (Centre for Fish and Wildlife Health, Vetsuisse Faculty, University of Bern, Switzerland) for his rapid and thorough help with in vivo challenges. The authors are especially grateful to Dr Cedric Simillion (Department of Clinical Research, University of Bern, Switzerland) for his invaluable technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors have declared that no competing interests exist.
Authors’ contributions
PVB conceived of the study, carried out the experiments, analyzed data from MS, performed bioinformatic analyses and drafted the manuscript. SB-L and MH performed MS experiments and interpretation of MS data. JF helped to draft the manuscript. Authors read and approved the final manuscript.
Electronic supplementary material
12953_2013_453_MOESM1_ESM.xlsx
Additional file 1: Table: PMSS, LFQ values, ratios, PEP values, subcellular localization, secretion system signals for each protein identified in SNs and pellets of wt and mutant strains in GP and SP. Column B: Proteins names. Red = T3SS components; dark red = other virulence factors (toxins, enzymes and adhesins); light red = putative secondary virulence factors; yellow = proteins specific of JF2267 or B526; mauve = multidrud resistance-associated proteins; orange = ABC transporters; light green = proteins associated to flagella, pili, T4SS; dark blue = phage proteins; light blue = cytoplasmic moonlighting proteins present in SNs; grey = T5SS; light pink T6SS, pink: transposases. Column E: A449 Loci. Grey = genes conserved among Aeromonas sp.; white = genes shared with at least one other Aeromonas species; green = genes specific of A. salmonicida; yellow = genes specific of A. salmonicida JF2267 and B526; pink = transposases. (XLSX 917 KB)
12953_2013_453_MOESM2_ESM.pdf
Additional file 2: Figure: most abundant proteins in A. salmonicida supernatants of wt and mutant strains in GP and SP. The diagram represents the most abundant proteins secreted by A. salmonicida (in decreasing order of PMSS values in wt SN during GP). Below the name of the protein circles represent T3SS components (red), other virulence factors (toxins, enzymes and adhesins) (pink), putative secondary virulence factors (yellow) and cytoplasmic proteins with putative moonlighting activity (green). (PDF 131 KB)
12953_2013_453_MOESM3_ESM.pdf
Additional file 3: Figure: most abundant proteins in A. salmonicida pellets of wt and mutant strains in GP and SP. The diagram represents the most abundant proteins detected in A. salmonicida pellets (in decreasing order of PMSS values in wt pellet during GP). Below the name of the protein circles represent T3SS components (red), other virulence factors (toxins, enzymes and adhesins) (pink), putative secondary virulence factors (yellow) and secreted cytoplasmic proteins with putative moonlighting activity (green). (PDF 178 KB)
12953_2013_453_MOESM4_ESM.pdf
Additional file 4: Figure: confirmation by western blotting of the difference in quantity observed between the pellets of the wt and the ΔascV mutant in the GP for AopD, AcrV and AexT. (PDF 3 MB)
12953_2013_453_MOESM5_ESM.doc
Additional file 5: Table: A. salmonicida secreted proteins that have homologues in other bacteria with a putative role in virulence. (DOC 270 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Vanden Bergh, P., Heller, M., Braga-Lagache, S. et al. The Aeromonas salmonicida subsp. salmonicida exoproteome: determination of the complete repertoire of Type-Three Secretion System effectors and identification of other virulence factors. Proteome Sci 11, 42 (2013). https://doi.org/10.1186/1477-5956-11-42
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1477-5956-11-42