
Abstract
Background
Lignocellulosic biomass is a promising renewable feedstock for biofuel production. Acetate is one of the major inhibitors liberated from hemicelluloses during hydrolysis. An understanding of the toxic effects of acetate on the fermentation microorganism and the efficient utilization of mixed sugars of glucose and xylose in the presence of hydrolysate inhibitors is crucial for economic biofuel production.
Results
A new microarray was designed including both coding sequences and intergenic regions to investigate the acetate stress responses of Zymomonas mobilis 8b when using single carbon sources of glucose or xylose, or mixed sugars of both glucose and xylose. With the supplementation of exogenous acetate, 8b can utilize all the glucose with a similar ethanol yield, although the growth, final biomass, and ethanol production rate were reduced. However, xylose utilization was inhibited in both media containing xylose or a mixed sugar of glucose and xylose, although the performance of 8b was better in mixed sugar than xylose-only media. The presence of acetate caused genes related to biosynthesis, the flagellar system, and glycolysis to be downregulated, and genes related to stress responses and energy metabolism to be upregulated. Unexpectedly, xylose seems to pose more stress on 8b, recruiting more genes for xylose utilization, than does acetate. Several gene candidates based on transcriptome results were selected for genetic manipulation, and a TonB-dependent receptor knockout mutant was confirmed to have a slight advantage regarding acetate tolerance.
Conclusions
Our results indicate Z. mobilis utilized a different mechanism for xylose utilization, with an even more severe impact on Z. mobilis than that caused by acetate treatment. Our study also suggests redox imbalance caused by stressful conditions may trigger a metabolic reaction leading to the accumulation of toxic intermediates such as xylitol, but Z. mobilis manages its carbon and energy metabolism through the control of individual reactions to mitigate the stressful conditions. We have thus provided extensive transcriptomic datasets and gained insights into the molecular responses of Z. mobilis to the inhibitor acetate when grown in different sugar sources, which will facilitate future metabolic modeling studies and strain improvement efforts for better xylose utilization and acetate tolerance.
Similar content being viewed by others


Background
Lignocellulosic biomass is considered to be a renewable and sustainable resource to address global challenges on environmental protection, energy security, and economic development, and cellulosic ethanol production has made significant progress at the pilot and demonstration scales. However, the toxic compounds generated during the deconstruction processes of pretreatment and enzymatic saccharification to release fermentable sugars such as glucose and xylose inhibit the microbial catalyst performance during fermentation to ethanol. These inhibitors include weak acids (such as acetic acid), aldehydes (for example, furfural), and lignin degradation products (such as vanillin) [1]. Acetic acid, liberated from hemicelluloses during biomass deconstruction, is one of the more dominant inhibitors due to its high concentration in lignocellulosic hydrolysates and its toxic effect on proton gradient homeostasis as a weak acid [2],[3]. The development of robust microbial catalysts capable of maintaining high productivity in the presence of acetate and other inhibitors is crucial for commercialization of biochemical conversion processes for biofuel production, and numerous efforts are being devoted to meeting this goal [3].
Although yeast remains a major microbial biocatalyst for ethanol production, other microorganisms such as E. coli and Zymomonas mobilis have also received significant attention. Z. mobilis, a Gram-negative facultative anaerobic ethanologenic bacterium, has excellent industrial characteristics such as unique anaerobic use of the Entner-Doudoroff (ED) pathway that results in a high ethanol yield per mole of glucose consumed, high specific productivity, high ethanol titers, and notable ethanol tolerance [4]-[9]. In addition, the availability of genome sequence for multiple cultivars [10]-[14], operon prediction tools [15], metabolic modeling results [16]-[19], and strain engineering methods [20]-[25] accelerates the research progress in Z. mobilis. However, wild-type Z. mobilis can only utilize glucose, fructose, and sucrose as carbon sources, and cannot utilize pentoses like xylose, which is the second most abundant sugar in pretreated and saccharified biomass slurries. An engineered Z. mobilis strain 8b was constructed expressing heterologous genes of talB, tktA, and xylAB for xylose utilization as well as truncating the endogenous lactate dehydrogenase gene ldh for improved flux to ethanol [23]. Z. mobilis 8b is more sensitive to acetate when grown in xylose. The IC50 value (chemical concentration inhibiting 50% cell growth) of acetate when Z. mobilis 8b is grown in xylose is 50 mM, compared to the value of 210 mM when glucose is used as the carbon source [1]. The concentration of acetate in a typical hydrolysate prepared from pretreated corn stover at 20% solids loading is about 82 mM, which will completely inhibit cell growth on xylose.
Despite advances in engineering strains of Z. mobilis for pentose utilization [23],[26]-[28], co-utilization of glucose and pentoses remains problematic, especially in the presence of inhibitory compounds such as acetate and furfural, and more work will be needed to achieve the high overall ethanol yields required for a commercial process [29]-[33]. Furthermore, despite recent systems biology studies undertaken to unravel the inhibitor tolerance mechanism of Z. mobilis for end-product ethanol [15],[34], the single inhibitor acetate [35],[36] or furfural [37], as well as the comprehensive hydrolysate toxic compounds [24], no transcriptomic study has yet been conducted focusing on the effect and interaction of pretreatment inhibitors and carbon source. This work addresses that knowledge gap and will provide insights on the development of a robust Z. mobilis with high pentose utilization efficiency for economic biofuel (both ethanol and advanced hydrocarbon) production.
Results
Physiological response to acetate and different sugars
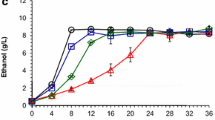
As we noted in a previous report for Z. mobilis ZM4 [35], the presence of acetate negatively affected cellular growth and ethanol production in Z. mobilis fermentations. This is especially true with xylose as the carbon source (Figure 1). The maximum biomass of Z. mobilis in terms of OD600nm value in different media (RMG, RMX, or RMGX) without exogenous acetate supplementation was lower than that in media with acetate supplementation, delaying the time to reach the stationary phase and maximal ethanol titers (Figure 1). Correspondingly, the growth rates (h-1) in cultures grown in the absence of acetate (RMG or RMX) were at least 50% higher than the growth rates in acetate-supplemented cultures (RMGAc or RMXAc): 0.43 ± 0.01 and 0.093 ± 0.003 in RMG and RMX compared to 0.28 ± 0.021 and 0.056 ± 0.002 in RMGAc or RMXAc, respectively. The sugar source had a dramatic impact on cellular growth; the growth rate of Z. mobilis 8b dropped sharply from 0.43 when using glucose to 0.093 when xylose was used as the sole carbon source. The supplementation of glucose restored the cellular growth, and the growth rates of Z. mobilis 8b in mixed sugars of glucose and xylose (RMGX and RMGXAc) were same as those in RMG and RMGAc (Figure 1).

Cell growth, substrate sugar utilization, and ethanol production in RMG8 (A), RMX8 (B), and RMG4X4 (C) media with and without the supplementation of NH 4 OAc.
Sugar utilization and ethanol production were monitored by HPLC (Figure 1). The elimination of lactate production is consistent with the lactate dehydrogenase gene deletion in the 8b background [23]. Corresponding to the decreased growth, Z. mobilis 8b consumed sugar more slowly with acetate supplementation. Ethanol production corresponded to the substrate (glucose or xylose) consumption in all conditions studied in this work; the ethanol was produced gradually as the sugar was consumed and reached its maximum when the glucose or xylose was used up (Figure 1).
Transcriptomic profiling of Z. mobilis in response to acetate using different carbon sources
New array design and microarray experiment
To reflect the improved genome annotation [38], we designed a new NimbleGen high-density 12-plex microarray to investigate all genetic features including the gene coding region (CDS) and intergenic regions. The new array contains 146-K features for both expression and tiling arrays covering both the genome and five plasmids as well as heterologous genes (talB, tktA, araBAD, and xylAB) engineered into Z. mobilis for xylose and arabinose metabolism [26],[27].
Samples were taken at exponential phase for the single sugar (RMG or RMX) fermentation experiment, and at different post-inoculation time points of exponential, transition, and stationary phases for mixed sugar (RMGX) fermentation (Figure 1). The time point between the end of the exponential phase and the beginning of the stationary phase was selected as the transition phase. Gene expression profiles were analyzed using a NimbleGen high-density expression array as described previously [35]. In addition to conditions using a sole carbon source of glucose or xylose only, the acetate effect on 8b during mixed sugar RMGX fermentation was also investigated to better model a real production situation using pretreated and saccharified biomass slurries containing both glucose and xylose. In total, 36 arrays were used (Additional file 1: Table S1), and two types of analyses were performed to investigate the effect of acetate on 8b under a variety of conditions: 1) a comparison of the effects of acetate with media (RMG, RMX, or RMGX) and growth phase (log, transition, or stationary) held constant; 2) a comparison of the effects of growth phase (log, transition, or stationary) within different media (RMG, RMX, or RMGX) with or without acetate supplementation.
Array data quality and qRT-PCR confirmation
The quality of data was checked with good correlation (Additional file 2: Figure S1) before the statistical analyses. The microarray data were also assessed using several approaches, and the quality was found to be high. The correlation coefficients for microarray data from biological replicates were good, with r > 0.9 for each comparison (Additional file 2: Figure S1A, B). In addition, the microarray data of biological replicates were also grouped together closely based on hierarchical clustering and principal components analyses (Additional file 2: Figure S1C, D), and the data can be grouped or separated as individual time points or by treatment.
Moreover, our newly designed microarray contains features of engineered genes such as talB, tktA, araBAD, and xylAB. Z. mobilis 8b contains talB, tktA, and xylAB but not araBAD. The array data are consistent with our strain background with the talB, tktA, and xylAB genes expressed but not araBAD (Additional file 2: Figure S2). Because our array datasets are derived from a high-density tiling array containing multiple probes for each gene, we may obtain information of post-transcriptional regulation through the transcriptomic study. Our results showed that the probe intensities for these engineered genes in different gene locations were different and the probes adjacent to the 5-prime region of these genes had higher expression intensity than those in the C-terminal region (Additional file 2: Figure S2).
Furthermore, gene expression profiles in terms of probe intensity can be overlaid across the whole genome to identify whether there are expression differences in the genome. In this study, most chromosomal regions were expressed evenly under different conditions. However, some regions had relatively higher or lower abundance than the average probe intensity under different conditions (for example, single sugar of RMG and RMX with and without the supplementation of exogenous acetate at exponential phase), which can help visualize the genome regions with different expression patterns and give an overview of gene expression profiles under different conditions (Additional file 2: Figure S3). On the contrary, only small sporadic regions in the plasmids had an expression intensity higher than the background signal and most areas had no expression (Additional file 2: Figure S3). Of the five plasmids reported in Z. mobilis[38], sequencing reads were only mapped to portions of three of them, calling for further study to investigate the plasmid profile and the plasmid stability of Z. mobilis after metabolic engineering and the role of plasmids in Z. mobilis physiology and metabolic networks.
Finally, ten genes from arrays with different expression values were picked for qPCR using a Bio-Rad CFX96 instrument and the Roche qPCR kit. Genes with significant array P-values in each comparison were selected, and the correlation between array and qPCR was plotted. The results indicate that our array had good quality with relatively high correlations between qRT-PCR and array results as in our previous reports [15],[35],[36],[39], although the correlation between RMGAc and RMG is relatively weak with a squared correlation coefficient value of 0.79 (Additional file 2: Figure S4).
Transcriptomic analysis of the effect of acetate and carbon source
Overview of acetate and sugar effect
Analysis of variance (ANOVA) analyses using JMP Genomics (SAS Inc., Cary, NC) were carried out using sugar and acetate treatment as variables, and the results indicated that acetate had a more dramatic impact on the transcriptome of 8b in RMG than that of cells grown in RMX, and that the difference between carbon sources is more dramatic than the effect caused by acetate treatment (Additional file 2: Figure S5; Table 1; Additional file 3: Table S2). The overview of significantly differentially expressed genes is shown in Additional file 2: Figure S5. The number of differentially expressed genes in response to acetate or carbon source is summarized in Table 1, and the detailed gene expression information is listed in Additional file 3: Table S2 (S2A for the single sugar experiment and S2B for the mixed sugar array study). With acetate treatment in RMG, only 148 genetic features (including both genes and intergenic region features) were differentially expressed, with 30 genes upregulated and 43 downregulated (Table 1, Additional file 3: Table S2-2). When xylose was used as the sole carbon source, 369 genetic features were differentially expressed compared to that using glucose as the sole carbon source including 113 genes upregulated and 128 genes downregulated (Table 1, Additional file 3: Table S2-4). The RMGX cultures grew much better than the RMX cultures (Figure 1) and, correspondingly, fewer genetic features were differentially expressed in the mixed sugar condition in the exponential phase, with only 28 genes upregulated and 17 downregulated compared to samples taken from the RMG medium (Table 1, Additional file 3: Table S2-17).
In addition, when acetate was supplemented into the medium, fewer genetic features were significantly differentially expressed when xylose was used as the sole carbon source than when glucose was used (76 versus 148) (Table 1, Additional file 3: Table S2-2, S2-3). This result further indicates that the utilization of the non-native sugar xylose in the absence of glucose presents a stressful environment for the cells, and the addition of acetate into RMX only causes an incremental transcriptional change since common stress-responsive genes were already expressed in the stressful xylose environment. Alternatively, cells could face dramatic perturbation when shifting from the favorable growth condition using glucose to other stressful conditions (such as xylose utilization or acetate resistance), and more transcriptional regulation is required for cells to adapt to the stress.
Time-course microarray investigation of the mixed sugar fermentation
In addition to the comparisons of the Z. mobilis 8b transcriptional profiles in RMG, RMX, or RMGX in the absence and presence of acetate, we also investigated the effect of acetate on the gene expression profiles of 8b at the different phases, log, transition, and stationary, in mixed sugar fermentation (Additional file 2: Figure S5, Additional file 3: Table S2-8 to S2-16). In the exponential phase, the effect of mixed sugars on gene expression in the presence of acetate is more like that of glucose than that of xylose with more differentially expressed genetic features (154 features, Table 1, Additional file 3: Table S2-11). In addition, there were more common differentially expressed genetic features shared between mixed sugar and glucose than between mixed sugar and xylose (Figure 2). This suggests that the supplementation of glucose into xylose can alleviate the stress caused by xylose and favor cellular metabolism and growth, which is consistent with the growth, sugar utilization, and ethanol production profiles (Figure 1, Additional file 4: Table S3).

Venn diagrams of common genetic features shared by shock stress responses in RMG8, RMX8, and RMG4X4. A: the acetate upregulated features with at least two-fold increase and shared with those upregulated in single sugar of RMG (G_Ac_UP) or RMX (X_Ac_UP), and mixed sugar of RMGX (GX_Ac_UP); B: the acetate downregulated features with at least two-fold decrease and shared with those downregulated in single sugar of RMG (G_Ac_D) or RMX (X_Ac_D), and mixed sugar of RMGX (GX_Ac_D); C: genetic features upregulated in xylose RMX (X/G_UP) or mixed sugar of RMGX (GX/G_UP) compared to that in RMG with at least two-fold increase without acetate supplementation, X/G_Ac_UP and GX/G_Ac_UP for those with acetate supplementation; D: genetic features downregulated in xylose RMX (X/G_D) or mixed sugar of RMGX (GX/G_D) compared to that in RMG with at least two-fold increase without acetate supplementation, X/G_Ac_D and GX/G_Ac_D for those with acetate supplementation.
The subsets of expressed genes with at least two-fold changes were used for their potential interactions using the STRING 9.05 precomputed database [40], and the results indicated that acetate upregulated genes in mixed sugar fermentation had better interactions than those of the downregulated ones (Additional file 2: Figure S6). Similar to the individual sugar fermentation, the upregulated genes in the mixed sugar fermentation also included genes encoding cysteine synthase (Additional file 2: Figure S6A). The downregulated ones are also similar to those in individual sugar fermentation, which are involved in biosynthesis (for example, ribosomal proteins), flagellar biosynthesis, and glycolysis (Additional file 2: Figure S6B).
However, we need to point out that these genes may be related to growth phase and may not be acetate specific. For example, dramatic differences were detected when 8b entered the transition phase from the log phase with a large number of genes highly differentially expressed (Additional file 2: Figure S5I, K, M, Additional file 3: Table S2-8, S2-14). The difference between the transition and stationary phases (Additional file 2: Figure S5J, L, N, Additional file 3: Table S2-9, S2-15) was less dramatic than that of the log to transition period. We believe that some genes identified in this study are mainly growth phase dependent rather than acetate or xylose related. This is consistent with our previous studies comparing the impact of stressors (such as oxygen and ethanol) at various stages of the growth phase [15],[39]. Caution is needed to identify and interpret stressor-related gene candidates using microarray data from one single time point.
Transcriptomic study of genes responsive to acetate resistance
We begin our transcriptomic profiling of 8b in response to acetate in RMG, RMX, and RMGX by identifying the common genetic features and those specific to each condition (Figure 2). Briefly, 30 chromosomal genes were upregulated by acetate in RMG (Table 1, Additional file 3: Table S2-2). Six of these are involved in sulfur metabolism including five genes involved in cysteine synthesis (CysC, N, S, K < ZMO003, 4, 7, 0748>, and ZMO0006) and an ABC transport system SsuACB (ZMO1261-3) involved in sulfonate import. The 43 genes downregulated by acetate in RMG were involved in cell motility (for flagellar assembly) and protein synthesis (ribosome component proteins). In the RMX medium, acetate upregulated 18 chromosomal genes (involved in chemotaxis and signal transduction) and downregulated 26 chromosomal genes (involved in biosynthesis and transport) (Table 1, Additional file 3: Table S2-3). However, in the mixed sugar medium RMGX, 60 chromosomal genes were found to be upregulated by acetate, mainly involved in oxidation-reduction reactions and amino acid metabolism. Acetate caused 44 chromosomal genes to be downregulated in RMGX, mainly coding for hypothetical proteins (Table 1, Additional file 3: Table S2-11).
Genes shared under different conditions were investigated, and ZMO1682 is the only upregulated gene responsive to acetate supplementation in all three growth conditions. This gene encodes L-aspartate beta-decarboxylase involved in amino acid metabolism (Figure 2A, Additional file 3: Table S2-2, S2-3, S2-11). Another gene upregulated by acetate in both RMG and RMX but not in RMGX is ZMO1181, which encodes N-formylglutamate amidohydrolase. Only one other gene, ZMO1885 (ncr) encoding NADH-flavin oxidoreductase, is upregulated by acetate in RMX and RMGX. In contrast, ten genes are upregulated by acetate in both RMG and RMGX. Three of these are involved in sulfur metabolism: cysteine synthase CysD, I, J (ZMO0005, 8, 9) and individual genes are involved in stress response: Fe-S oxidoreductase (ZMO0022), aldo/keto reductase (Dkg, ZMO1344), and catalase (ZMO0918). Other upregulated genes encode the TonB-dependent receptor (ZMO0031), glucan biosynthesis protein D (ZMO0905), L-sorbose reductase (ZMO1449), and a potential alanine/aspartate transporter (ZMO1681) (Figure 2A, Additional file 3: Table S2-2, S2-3, S2-11).
The two genes downregulated by acetate in RMG, RMX, and RMGX are ZMO0128 and ZMO1522, which are annotated as TonB-dependent receptors (Figure 2B, Additional file 3: Table S2-2, S2-3, S2-11). Nine additional genes were downregulated by acetate in RMG and RMGX. With the exception of ZMO0127 (encoding a nuclease S1), ZMO0130 (phoC, encoding an acid phosphatase precursor), ZMO1020 (encoding a decarboxylase 2), and ZMO0412 (encoding a potential multiple antibiotic transporter), the other five genes (ZMO0095, 0110, 0131, 0912, and 1521) encode hypothetical proteins.
Clearly from the Venn diagrams (Figure 2), the genes responsive to acetate in RMX, RMG, or RMGX alone far exceed those shared in two or more media. These results indicate that acetate has little further effect than the stress of growth in xylose compared to growth in glucose or glucose plus xylose. The slightly altered pattern of genes differentially expressed between RMG and RMGX suggests that xylose does exert subtle effects in RMGX (Figure 2, Additional file 3: Table S2-2, S2-11).
Transcriptomic study of genes responsive to xylose utilization
To investigate the effect of the non-native sugar xylose on 8b gene expression, the transcriptomic profile of 8b in RMG was compared to those of cells grown in RMX (Additional file 3: Table S2-4) or RMGX (Additional file 3: Table S2-17). Only one common gene was identified to be upregulated, ZMO1268, a hypothetical protein containing a truncated MarR domain with very low similarity (Evalue = 2.31e-03), no matter whether the exogenous acetate was present or not.
Since the growth and gene expression profiles in RMGX were similar to those in RMG (Figure 2, Table 1), we focused our subsequent analysis of differentially expressed genes on results from cells grown in RMG and RMX. The utilization of non-native xylose apparently poses a significant metabolic burden to the cells, with 113 genes upregulated and 128 downregulated (Table 1, Additional file 3: Table S2-4), and the gene functions of these differentially expressed genes have much in common with acetate-responsive genes as discussed above as well as with other stress (ethanol or furfural) responsive genes as described in previous reports [34],[41]. In the presence of acetate, only 40 genes were upregulated and 35 genes downregulated in RMX compared to RMG (Table 1, Additional file 3: Table S2-5). This indicates that multiple stressors elicit the same basic response.
Indeed, 7 upregulated genes and 16 downregulated ones are common between the acetate-responsive genes in RMG (Additional file 3: Table S2-2) and the xylose-responsive genes without acetate supplementation (RMX versus RMG, Additional file 3: Table S2-4). These common genes may serve as genetic targets for improving microbial tolerance to different stressors. However, we also point out that, despite the commonality, there are many more transcriptional responses unique to the individual stressors. Growth in xylose led to upregulation of 113 genes and downregulation of 128 genes compared to growth in glucose, while only 23 genes were acetate upregulated and 27 were acetate downregulated (Figure 2, Table 1, Additional file 3: Table S2-2, S2-4).
Genetics confirmation
As noted above, our transcriptomic analysis provided targets for strain improvement to increase acetate tolerance. We constructed chromosomal knockin mutant strains to overexpress the genes that were upregulated to acetate as well as knockout mutants of the upregulated and downregulated genes. We hypothesized that the overexpression of the upregulated genes as well as deletion of the downregulated genes could confer growth advantage in the presence of acetate, while the deletion of the upregulated genes would pose a negative effect. Unfortunately, our knockin strategy was unsuccessful in generating stable mutants, perhaps due to the choice of a constitutive strong promoter in our constructs (data not shown). However, we were successful in constructing three knockout mutants (8b-KO1181, 8b-KO1682, and 8b-KO0128, named for the cognate gene). These mutant strains grew similarly as their parental strain 8b in RMG without exogenous acetate supplementation (data not shown). 8b-KO1181 and 8b-KO1682 were indistinguishable from the control strain, while 8b-KO0128 had a slight advantage in acetate resistance for cultures grown in RMG plus acetate (Additional file 2: Figure S7). A BLAST search of the Z. mobilis genome indicated that ZMO1682 and ZMO1181 are homologues of ZMO0342 (aatA) and ZMO1395, respectively, and these homologous gene products may compensate for the effect of the deletion. Similarly, the existence of multiple homologues of ZMO0128 in Z. mobilis (Additional file 2: Figure S8) may explain why the knockout of ZMO0128 only conferred a slight advantage in acetate tolerance. Further directed knockout mutants to eliminate other homologous genes may boost the effect of acetate tolerance.
Discussion
The exogenous acetate stress or non-native xylose utilization shifted redox balance, leading to toxic intermediate xylitol formation and glycolysis inhibition
Glycolysis and fermentation provide not only the necessary metabolic intermediates but also energy forms of ATP and redox NAD(P)H for anabolism, and the enzymes involved in glycolysis are among the most abundant proteins in Z. mobilis[15]. The stressful conditions trigger energy conservation and inhibit central metabolism with genes related to stress response being upregulated [15],[39]. In addition, the expression levels for genes encoding the glycolytic enzymes were also affected by the stressful condition. For example, our previous study suggests that the transcriptional levels of glycolytic enzymes (such as Gpm, Eno, and Pyk) were reduced in the aerobic culture compared to that of the anaerobic culture. In addition, glycolysis intermediates were accumulated which may be due to the reduced glycolysis efficiency [39]. A similar energy conservation phenomenon under stressful conditions has also been observed for E. coli[42],[43].
To investigate the impact of acetate supplementation and xylose utilization on glycolysis and fermentation, the metabolic pathway including xylose metabolism, glycolysis, and fermentation in the Z. mobilis 8b strain was reconstructed, and transcriptomic data were mapped into the pathway for xylose and acetate responsive genes under different conditions (Figure 3). When glucose was used as the only carbon source, the efficient Z. mobilis fermentation pathway enzymes Pdc (ZMO1360) and AdhB (ZMO1596) allowed fast ethanol production with minimal xylitol and acetate produced as byproducts (Figure 3, Additional file 4: Table S3). However, when xylose was the only carbon source, the acetate and xylitol levels were much higher than those in the RMG cultures (Additional file 4: Table S3). Comparing cells grown in RMX versus RMG, the ssdA (ZMO1754) gene encoding aldehyde dehydrogenase, which catalyzes the conversion of acetaldehyde to acetate, was upregulated eight-fold, and the adhA (ZMO1236) gene, encoding an iron-containing group III alcohol dehydrogenase proposed to oxidize ethanol into acetaldehyde [44],[45], was also upregulated about four-fold. Similar results were seen when exogenous acetate was added into the medium, which suggests that the acetate accumulation may cause a buildup of the NADH reducing equivalents, leading to redox imbalance (Figure 3).

Metabolic pathway of xylose degradation, glycolysis, and fermentation in Z. mobilis 8b strain. The box adjacent to the enzyme contains gene expression information from Additional file 3: Table S2-1, Table S2-6. The top row is for sugar effect: xylose-responsive genes compared to glucose in log phase without or with the supplementation of exogenous acetate from left to right column respectively, and the middle row is for acetate effect in log phase when exogenous acetate was added into single sugar of glucose or xylose respectively, from left to right column (Additional file 3: Table S2-1). The bottom row is for acetate effect in mixed sugar in log, transition, or stationary phase respectively, from left to right column (Additional file 3: Table S2-6). Absolute values greater than 1.5-fold (number in the box is log2-based value) were used, with red-filled box indicating upregulation and blue-filled box downregulation.
Excessive amounts of NADH could then drive the reaction of xylose conversion towards the accumulation of xylitol instead of the production of D-xylulose to regenerate NAD for redox balance (Figure 3). As evidence of this hypothesis, there was a high correlation (R2 = 0.99) between the xylose consumed and the xylitol accumulated (Additional file 4: Table S3). In addition, ZMO0976, a gene proposed to convert xylose to xylitol [46], was also upregulated consistent with a need to balance the excessive NADH accumulated, which is detrimental to the cells. The knockout of this gene has been proved to help Z. mobilis resist acetate toxicity [29]. Furthermore, ZMO1984, a paralogue to ZMO0976, was also upregulated similarly in response to xylose, which may further drive the reaction toward xylitol production (Figure 3). Although the reaction from xylose to xylitol could mitigate the problem of redox imbalance, xylitol is toxic to the cells and causes growth inhibition. In addition, the gfo gene encoded by ZMO0689 was reported to carry out the reaction from xylose to xylitol as well [47], and it was downregulated in RMX compared to RMG. Taken together, these observations indicate that the conversion of xylose to xylitol is more complicated than previously thought, as this reaction is involved in multiple responses to metabolic stressors, all leading to a toxic dead-end product of xylose (Figure 3).
Glf (ZMO0366) has been proposed to serve as a xylose transporter in addition to its primary role as a glucose transporter [37]. A major facilitator superfamily (MFS) family sugar transporter ZMO0293 (EC: 1.3.1.74) is homologous to Glf with a high similarity score of 191. ZMO0293 encodes a symporter with 12 transmembrane domains, which also has high similarity to xylose and arabinose symporters of E. coli (for example, XylE). ZMO0293 was downregulated when using xylose (RMX) compared to that in RMG in the absence or presence of exogenous acetate. In addition, when xylose was the sole carbon source, ZMO0293 was downregulated further in the presence of acetate (Figure 3). Furthermore, ZMO0293 was downregulated in late phases (transition or stationary) compared to the log phase with or without the acetate supplementation. This is consistent with its xylose downregulation phenomenon, since glucose was completely consumed with xylose present when reaching the transition or stationary phases (Figure 1, Additional file 3: Table S2-1, S2-6, Additional file 4: Table S3). We therefore propose that ZMO0293 may be the major transporter involved in xylose importation, and the downregulation of ZMO0293 in the presence of xylose may explain the poor xylose utilization by Z. mobilis and could be a genetic target to improve xylose utilization in Z. mobilis (for example, by changing the promoter region to release the transcriptional repression in the presence of xylose).
As described above, acetate significantly inhibits growth in RMX medium. About half of the xylose was still remaining in the media at 148 h post inoculation and the ethanol concentration was only about half that in RMX without acetate supplementation (Figure 3, Additional file 4: Table S3). The addition of acetate into RMX caused the downregulation of ZMO0293, which could reduce xylose uptake in Z. mobilis. At the same time, the Pfl gene (ZMO1570) was upregulated, which could drive more pyruvate into the acetate production, causing NADH buildup and more xylitol accumulation. For example, when about half of the xylose (38 g) was consumed, the xylitol concentration was 2.38 g/L in the absence of acetate compared to 3.37 g/L with the supplementation of acetate (Additional file 4: Table S3). Similarly, when the RMGX medium was used, xylitol was higher in the presence of acetate than in its absence (Figure 1, Additional file 4: Table S3). This result indicates that the stress from exogenous acetate can cause a toxic chain reaction by driving carbon flux toward acetate production with excessive NADH accumulation, shifting from complete metabolism of xylose via xylulose to xylitol generation, inhibiting growth and ethanol production (Figure 1).
Alternative energy metabolism pathways to overcome the energy shortage due to glycolysis inhibition
Previous studies have indicated that, besides energy conservation, microorganisms might bypass the central glycolysis pathway and utilize alternative pathways (such as the nitrogen metabolism in Clostridium thermocellum ATCC27405 [48]) for carbon metabolism and energy regeneration to survive the harsh conditions. Additionally, sulfur assimilation was suggested to contribute to Z. mobilis ethanol stress response and hydrolysate tolerance [15],[24]. In this study, several sulfur assimilation-related genes were also upregulated in RMG medium in the presence of acetate. For example, cysI, J, K, N were upregulated, and genes related to glutathione synthesis needed for assembly of Fe-S clusters were also upregulated in a response to acetate. Other general stress response genes were also upregulated in response to acetate in this study including genes encoding proteases (Clp), chaperone proteins (GroEL), and nucleases, which will help repair the damaged proteins and DNA to ensure accurate cellular function. Furthermore, metabolism of these protein and DNA degradation products may also help resolve any energy imbalances.
Moreover, ZMO1181 was upregulated in both RMG and RMX media in the presence of acetate. ZMO1181 is a single gene transcription unit (TU) encoding N-formylglutamate amidohydrolase, which catalyzes the hydrolysis reaction to release amino acids such as alanine from bacterial cell-wall glycopeptides as a substrate for alternative carbon metabolism. All of these results suggest that Z. mobilis may bypass the inhibition on central metabolism by external stressful conditions and utilize alternative carbon and energy metabolism pathways to provide energy and metabolic precursors to keep redox balanced and the cell growing under stressful conditions (Figure 3). Additional aspects of energy conservation and generation will be addressed in the following section.
Amino acid decarboxylase for energy generation
Amino acid decarboxylation has been suggested to help microorganisms resist acid stress. For example, aspartate-alanine antiporters and histidine decarboxylation enzymes help proton motive force (PMF) generation and ATP formation in Lactobacillus sp. [49],[50]. In addition, four genes involved in amino acid decarboxylation were reported to be associated with acid tolerance of L. acidophilus[51]. The consistent upregulation of ZMO1682 in Z. mobilis 8b in response to acetate was independent of sugar source of glucose, xylose, or the mixed sugar of RMGX, indicating its critical role in resistance to the weak acid of acetate. ZMO1682 encodes an aspartate aminotransferase and forms an operon with ZMO1681, which encodes a putative aspartate-alanine antiporter. This operon is coexpressed under ethanol and salt conditions [15] as well as the acetate condition in this study. This operon likely responds to the stress such as xylose utilization and acetate supplementation in the early stage, and the upregulation of this operon may help Z. mobilis generate PMF and form ATP for acetate tolerance. During acetate stress conditions, genes encoding proteases and chaperone proteins were upregulated (Additional file 3: Table S2-2, S2-3, S2-11), which may help repair and at the same time degrade these damaged proteins to release the amino acids as substrates for this decarboxylation reaction and resist the acetate stress.
The dilemma of sugar transport on substrate importation and energy conservation
Glf (ZMO0366, EC: 1.3.1.74) was regarded as the major sugar transporter and reported to be responsive for the transport of non-native sugars such as xylose [37]. In this study, glf was only downregulated from the log phase to transition and stationary phases in RMGX in the presence of exogenous acetate. Its expression was similar in RMG or RMX with or without acetate supplementation at log phase (Additional file 3: Table S2). However, Z. mobilis growth in RMG or RMX was dramatically different (Figure 1). Besides the impact of toxic xylitol accumulation and redox imbalance as discussed above, other genetic elements may also be responsive for these physiology differences of Z. mobilis.
As discussed briefly above, the expression of XylE homologous gene ZMO0293 was growth-phase dependent in the mixed sugar fermentation and downregulated in late growth phases when glucose was consumed. The expression of ZMO0293 was also downregulated by xylose (Additional file 3: Table S2-4, S2-5). This is different from its E. coli XylE symporter, which is repressed by glucose but induced by xylose. The expression of ZMO0293 was also downregulated by acetate in RMX medium (Additional file 3: Table S2-3), which indicates that the existence of acetate in the media may reduce xylose transport, thus interfering with carbon metabolism. Conceivably, the importation of both sugar and proton by this kind of MFS family symporter would change the PMF gradient and further acidify the cell, especially in the presence of the weak acid acetate or in stationary phase when metabolism decreases and the cells are less capable of pumping protons out for energy regeneration. Although these gene expression patterns and cell physiology differences indicate the involvement of sugar transport in stress response, further work is needed to systematically investigate the transport kinetics and substrate specificity of Glf and ZMO0293 under similar growth conditions to those used for this study. Upregulation of another transporter ZMO1452 (AraJ) with low similarity to Glf from log to transition phase (Additional file 3: Table S2-14) and its downregulation from transition to stationary (Additional file 3: Table S2-15) as well as the downregulation in the presence of acetate in stationary phase (Additional file 3: Table S2-6) demonstrate the importance and complexity of sugar transport in adapting to different environmental conditions.
TonB-dependent transporters and energy conservation
TonB-dependent receptors are a family of beta-barrel proteins in the Gram-negative bacteria outer membrane, which can transport substrates of iron chelators (siderophores), oligosaccharides, and polypeptides against the concentration gradient with two other components of a periplasma-facing TonB protein and a plasma membrane localized machinery (ExbBD). In E. coli, the TonB protein interacts with outer membrane receptor proteins that carry out high-affinity binding and energy-dependent uptake of specific substrates into the periplasmic space [2]. In this study, two genes (ZMO0128 and ZMO1522) annotated as TonB-dependent receptors were consistently downregulated under different conditions. Potentially, ZMO0128 is a paralogue to four other Z. mobilis genes annotated as TonB-dependent receptors (ZMO0789, ZMO1463, ZMO1568, and ZMO1694) with a close relationship (Additional file 2: Figure S8). ZMO1522 is not similar to ZMO0128 and distinct from other genes annotated as TonB-dependent receptors such as ZMO0171, ZMO0979, ZMO1040, and ZMO1260. Six other TonB-dependent receptors are more similar to ZMO0128 than to ZMO1522 (Additional file 2: Figure S8). The downregulation of these two TonB-dependent receptors may therefore reduce energy needs for substrate uptake and reserve the energy for stress responses. The ZMO0128 knockout mutant did have a slight advantage over the wild-type strain for acetate tolerance (Additional file 2: Figure S7). However, further investigation is needed to identify the substrates that these TonB-dependent receptors are binding to, and the relationship between the substrate uptake repression and energy conservation.
Conclusions
The physiological responses and transcriptomic profiles of an engineered xylose-utilizing Z. mobilis strain 8b to acetate stress using glucose or xylose individually or in mixed sugar fermentations were investigated. The non-native sugar xylose poses a stress for 8b, and the supplementation of acetate impedes the cell growth of 8b, resulting in slow substrate utilization and ethanol production. The transcriptomic profiles indicate that the molecular responses of 8b to xylose and acetate are dynamic and complicated. The utilization of the non-native sugar, xylose, elicits a more dramatic effect on transcriptional regulation than the inhibitor, acetate. Our study also suggests that stressful conditions such as growth in xylose in the presence and absence of acetate cause a redox imbalance. As the cells adjust metabolism to address redox imbalance, carbon flux may shift towards the accumulation of a toxic intermediate such as xylitol. In addition, Z. mobilis appears to manage its energy balance through the reduction of energy-consuming reactions such as expression of genes encoding efflux pumps and substrate transporters as well as energy regeneration through alternative carbon and energy pathways such as amino acid decarboxylation. Several target gene candidates based on the transcriptomic results were selected for genetic manipulation, and a TonB-dependent receptor knockout mutant was shown to convey acetate tolerance. Our study has provided insights into the molecular responses of the model ethanologenic bacterium Z. mobilis to the inhibitor acetate in different sugar sources, which will help further strain metabolic engineering efforts for better xylose utilization and acetate tolerance (for example, deletion of ZMO0293 for improved xylose transportation and utilization of the substrate channeling technique to direct the substrate xylose into the pentose phosphate pathway instead of toxic intermediate xylitol accumulation).
Materials and methods
Bacterial strain and growth conditions
Z. mobilis 8b was revived from frozen glycerol stocks for about 6 to 8 h in 10 mL RM (10 g/L yeast extract, 2 g/L KH2PO4) + 2% glucose at 33°C prior to inoculating the seed cultures overnight in RM + 5% glucose (RMG5) using shake flasks filled to 80% capacity at 33°C at 120 rpm. When the glucose concentration reached approximately 20 to 40 g/L, the cells were spun down at 3,840 × g for 10 min at room temperature (RT) and resuspended in either RMG (8% glucose), RMX (8% xylose), or RMGX (4% glucose, 4% xylose) at a ten-fold concentration and used as inocula for fermentation studies.
Controlled batch fermentations
Fermentations were performed in BioStat-Q Plus fermentors (Sartorius Stedim Biotech, France) at 300-mL working volumes using Z. mobilis 8b. RMG, RMX, or RMGX media were used for fermentation. The fermentors were inoculated at an OD600nm unit of 0.1 approximately. The fermentation was conducted at 30°C with an agitation rate of 150 rpm. The pH control set point was maintained at 6.0 by automatic titration with 2 N KOH. Samples were harvested during fermentation at different time points for transcriptomic and HPLC analyses as described previously [39]. The time point between the end of the exponential phase and the beginning of the stationary phase was selected as the transition phase. Specifically for the mixed sugar fermentation with multiple sampling time points, the exponential, transition, and stationary time points without acetate supplementation are 7, 12, and 36.5 h post inoculation, whereas the exponential, transition, and stationary time points with acetate supplementation are 11.5, 21, and 59 h post inoculation.
Bioscreen C high-throughput toxicity assay
The high-throughput Bioscreen C assay was carried out as reported previously [1],[52]. Briefly, Z. mobilis cells were revived from overnight culture with OD600nm adjusted to 1.5, and 10 μL seed culture was added into 290 μL media with an initial OD600nm of about 0.05. Growth was then monitored using a Bioscreen C instrument (GrowthCurves, Piscataway, NJ, USA) with three technical replicates. The experiments were repeated at least two times.
Extracellular metabolite analysis with HPLC
HPLC analysis was used for the measurements of the extracellular metabolite concentration of glucose, xylose, acetate, ethanol, xylitol, glycerol, and lactate in 0.2 μm-filtered samples taken at different time points during fermentation. Soluble fermentation products were identified by comparison with retention times and peak areas of corresponding standards as described previously [15],[35],[39],[53].
RNA extraction and ds-cDNA synthesis
Total RNA was isolated from the cell pellet resuspended in TRIzol reagent (Invitrogen, Carlsbad, CA) as described previously [39]. Each total RNA preparation was treated with RNase-free DNase I (Ambion, Austin, TX) to digest residual genomic DNA and subsequently purified using the RNeasy Mini Kit (Qiagen, Valencia, CA). Total cellular RNA was quantified with a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE) and RNA quality was assessed with an Agilent Bioanalyzer (Agilent, Santa Clara, CA). Purified RNA of high quality was used as the template to generate ds-cDNA using Invitrogen ds-cDNA synthesis kit (Invitrogen, CA).
Microarray sample labeling, hybridization, scan, and statistical analysis of array data
Ds-cDNA was labeled, hybridized, washed, and scanned following the NimbleGen protocols. Hybridizations were conducted using a 12-bay hybridization station (BioMicro Systems Inc., Salt Lake City, UT), the arrays were dried using a Maui wash system (BioMicro Systems Inc.) and scanned with a SureScan high-resolution DNA microarray scanner (Agilent Technologies, CA), and the images were quantified using NimbleScan software (Roche NimbleGen, IN). Statistical analyses were done with JMP Genomics 5.1 software (SAS Institute, Cary, NC) as described previously [35],[39]. Briefly, raw data of probe fluorescence intensity were log2 transformed and imported into JMP Genomics for analysis. A distribution analysis and data correlation analysis were conducted as a quality control step. The overlaid kernel density estimates derived from the distribution analysis allowed the visualization of sources of variation based on strain and treatment, as well as variation attributed to technical factors, such as array. The data were subsequently normalized using the LOESS Normalization algorithm within JMP Genomics. An analysis of variance (ANOVA) was performed to determine differential expression levels between conditions and time points using the false discovery rate (FDR) testing method (P < 0.05). The significantly differentially expressed gene lists for different comparisons were generated and outputted as Excel sheets. Microarray studies have been submitted to NCBI GEO database with the accession number GSE57553. The interactions among differentially expressed genes were investigated using the STRING 9.05 precomputed database [40] available at http://string.embl.de/.
Chromosomal knockout deletion constructs
The Gibson Assembly kit (New England BioLabs, Ipswich, MA) was used to assemble upstream and downstream fragments (about 1.2 kb each) of the target deletion gene with the spectinomycin gene aadA in the middle. The assembled product was used as a template for PCR amplification. The purified PCR product was directly electroporated into the Z. mobilis. Transformants appearing on RMG plates with the supplementation of spectinomycin were screened using colony PCR. Colonies with correct PCR product sizes were selected as deletion candidates after sequencing confirmation using the Sanger sequencing service from GenScript (Piscataway, NJ, USA).
Authors’ contributions
MZ conceived and led the project. SY, MAF, YCC, and MZ designed the experiment. SY, MAF, and YCC carried out the fermentation. MAF generated the HPLC data. SY did the RNA extraction, qPCR, and knockout mutant construction. SDB’s lab did the microarray. SY and MZ analyzed the data. SY wrote the manuscript. PTP conducted extensive review, and MAF, SDB, and MZ provided comments for revision. All authors read and approved the final manuscript.
Additional files
Abbreviations
- CDS:
-
gene coding region
- ED:
-
Entner-Doudoroff pathway
- HPLC:
-
high-performance liquid chromatography
- IC50:
-
chemical concentration inhibiting 50% cell growth
- MFS:
-
major facilitator superfamily transporter
- PMF:
-
proton motive force
- TU:
-
transcription unit
References
Franden MA, Pilath HM, Mohagheghi A, Pienkos PT, Zhang M: Inhibition of growth of Zymomonas mobilis by model compounds found in lignocellulosic hydrolysates. Biotechnol Biofuels. 2013, 6: 99-10.1186/1754-6834-6-99.
Zaldivar J, Ingram LO: Effect of organic acids on the growth and fermentation of ethanologenic Escherichia coli LY01. Biotechnol Bioeng. 1999, 66: 203-210. 10.1002/(SICI)1097-0290(1999)66:4<203::AID-BIT1>3.0.CO;2-#.
Mills TY, Sandoval NR, Gill RT: Cellulosic hydrolysate toxicity and tolerance mechanisms in Escherichia coli. Biotechnol Biofuels. 2009, 2: 26-10.1186/1754-6834-2-26.
Dien BS, Cotta MA, Jeffries TW: Bacteria engineered for fuel ethanol production: current status. Appl Microbiol Biotechnol. 2003, 63: 258-266. 10.1007/s00253-003-1444-y.
Gunasekaran P, Raj KC: Ethanol fermentation technology - Zymomonas mobilis. Curr Sci. 1999, 77: 56-68.
Panesar PS, Marwaha SS, Kennedy JF: Zymomonas mobilis: an alternative ethanol producer. J Chem Technol Biotechnol. 2006, 81: 623-635. 10.1002/jctb.1448.
Rogers PL, Goodman AE, Heyes RH: Zymomonas ethanol fermentations. Microbiol Sci. 1984, 1: 133-136.
Rogers PL, Jeon YJ, Lee KJ, Lawford HG: Zymomonas mobilis for fuel ethanol and higher value products. Adv Biochem Eng Biotechnol. 2007, 108: 263-288.
Swings J, De Ley J: The biology of Zymomonas mobilis. Bacteriol Rev. 1977, 41: 1-46.
Zhao N, Bai Y, Zhao XQ, Yang ZY, Bai FW: Draft genome sequence of the flocculating Zymomonas mobilis strain ZM401 (ATCC 31822). J Bacteriol. 2012, 194: 7008-7009. 10.1128/JB.01947-12.
Desiniotis A, Kouvelis VN, Davenport K, Bruce D, Detter C, Tapia R, Han C, Goodwin LA, Woyke T, Kyrpides NC, Typas MA, Pappas KM: Complete genome sequence of the ethanol-producing Zymomonas mobilis subsp. mobilis centrotype ATCC 29191. J Bacteriol. 2012, 194: 5966-5967. 10.1128/JB.01398-12.
Kouvelis VN, Davenport KW, Brettin TS, Bruce D, Detter C, Han CS, Nolan M, Tapia R, Damoulaki A, Kyrpides NC, Typas MA, Pappas KM: Genome sequence of the ethanol-producing Zymomonas mobilis subsp. pomaceae lectotype strain ATCC 29192. J Bacteriol. 2011, 193: 5049-5050. 10.1128/JB.05273-11.
Kouvelis VN, Saunders E, Brettin TS, Bruce D, Detter C, Han C, Typas MA, Pappas KM: Complete genome sequence of the ethanol producer Zymomonas mobilis NCIMB 11163. J Bacteriol. 2009, 191: 7140-7141. 10.1128/JB.01084-09.
Seo JS, Chong H, Park HS, Yoon KO, Jung C, Kim JJ, Hong JH, Kim H, Kim JH, Kil JI, Park CJ, Oh HM, Lee JS, Jin SJ, Um HW, Lee HJ, Oh SJ, Kim JY, Kang HL, Lee SY, Lee KJ, Kang HS: The genome sequence of the ethanologenic bacterium Zymomonas mobilis ZM4. Nat Biotechnol. 2005, 23: 63-68. 10.1038/nbt1045.
Yang S, Pan C, Tschaplinski TJ, Hurst GB, Engle NL, Zhou W, Dam PA, Xu Y, Rodriguez M, Dice L, Johnson CM, Davison BH, Brown SD: Systems biology analysis of Zymomonas mobilis ZM4 ethanol stress responses. PLoS One. 2013, 8: e68886-10.1371/journal.pone.0068886.
Lee KY, Park JM, Kim TY, Yun H, Lee SY: The genome-scale metabolic network analysis of Zymomonas mobilis ZM4 explains physiological features and suggests ethanol and succinic acid production strategies. Microb Cell Fact. 2010, 9: 94-10.1186/1475-2859-9-94.
Widiastuti H, Kim JY, Selvarasu S, Karimi IA, Kim H, Seo JS, Lee DY: Genome-scale modeling and in silico analysis of ethanologenic bacteria Zymomonas mobilis. Biotechnol Bioeng. 2011, 108: 655-665. 10.1002/bit.22965.
Pentjuss A, Odzina I, Kostromins A, Fell DA, Stalidzans E, Kalnenieks U: Biotechnological potential of respiring Zymomonas mobilis: a stoichiometric analysis of its central metabolism. J Biotechnol. 2013, 165: 1-10. 10.1016/j.jbiotec.2013.02.014.
Altintas MM, Eddy CK, Zhang M, McMillan JD, Kompala DS: Kinetic modeling to optimize pentose fermentation in Zymomonas mobilis. Biotechnol Bioeng. 2006, 94: 273-295. 10.1002/bit.20843.
Kerr AL, Jeon YJ, Svenson CJ, Rogers PL, Neilan BA: DNA restriction-modification systems in the ethanologen, Zymomonas mobilis ZM4. Appl Microbiol Biotechnol. 2011, 89: 761-769. 10.1007/s00253-010-2936-1.
Jia X, Wei N, Wang T, Wang H: Use of an EZ-Tn5-based random mutagenesis system to create a Zymomonas mobilis with significant tolerance to heat stress and malnutrition. J Ind Microbiol Biotechnol. 2013, 40: 811-822. 10.1007/s10295-013-1287-1.
Zhang X, Wang T, Zhou W, Jia X, Wang H: Use of a Tn5-based transposon system to create a cost-effective Zymomonas mobilis for ethanol production from lignocelluloses. Microb Cell Fact. 2013, 12: 41-10.1186/1475-2859-12-41.
Zhang M, Chou YC, Howe W, Eddy C, Evans K, Mohagheghi A: Zymomonas pentose-sugar fermenting strains and uses thereof.US Patent 2007, US7223575.,
Skerker JM, Leon D, Price MN, Mar JS, Tarjan DR, Wetmore KM, Deutschbauer AM, Baumohl JK, Bauer S, Ibáñez AB, Mitchell VD, Wu CH, Hu P, Hazen T, Arkin AP: Dissecting a complex chemical stress: chemogenomic profiling of plant hydrolysates. Mol Syst Biol. 2013, 9: 674-10.1038/msb.2013.30.
Yang S, Keller M, Brown S: Genomics on pretreatment inhibitor tolerance of Zymomonas mobilis. Microbial Stress Tolerance for Biofuels. Edited by: Liu ZL. 2012, Springer, Berlin Heidelberg,
Zhang M, Eddy C, Deanda K, Finkelstein M, Picataggio S: Metabolic engineering of a pentose metabolism pathway in ethanologenic Zymomonas mobilis. Science. 1995, 267: 240-243. 10.1126/science.267.5195.240.
Deanda K, Zhang M, Eddy C, Picataggio S: Development of an arabinose-fermenting Zymomonas mobilis strain by metabolic pathway engineering. Appl Environ Microbiol. 1996, 62: 4465-4470.
Ma Y, Dong H, Zou S, Hong J, Zhang M: Comparison of glucose/xylose co-fermentation by recombinant Zymomonas mobilis under different genetic and environmental conditions. Biotechnol Lett. 2012, 34: 1297-1304. 10.1007/s10529-012-0897-4.
Agrawal M, Wang Y, Chen RR: Engineering efficient xylose metabolism into an acetic acid-tolerant Zymomonas mobilis strain by introducing adaptation-induced mutations. Biotechnol Lett. 2012, 34: 1825-1832. 10.1007/s10529-012-0970-z.
Gao Q, Zhang M, McMillan JD, Kompala DS: Characterization of heterologous and native enzyme activity profiles in metabolically engineered Zymomonas mobilis strains during batch fermentation of glucose and xylose mixtures. Appl Biochem Biotechnol. 2002, 98-100: 341-355. 10.1385/ABAB:98-100:1-9:341.
Jeon YJ, Svenson CJ, Rogers PL: Over-expression of xylulokinase in a xylose-metabolising recombinant strain of Zymomonas mobilis. FEMS Microbiol Lett. 2005, 244: 85-92. 10.1016/j.femsle.2005.01.025.
Mohagheghi A, Evans K, Chou YC, Zhang M: Cofermentation of glucose, xylose, and arabinose by genomic DNA-integrated xylose/arabinose fermenting strain of Zymomonas mobilis AX101. Appl Biochem Biotechnol. 2002, 98-100: 885-898. 10.1385/ABAB:98-100:1-9:885.
Mohagheghi A, Evans K, Finkelstein M, Zhang M: Cofermentation of glucose, xylose, and arabinose by mixed cultures of two genetically engineered Zymomonas mobilis strains. Appl Biochem Biotechnol. 1998, 70-72: 285-299. 10.1007/BF02920145.
He MX, Wu B, Shui ZX, Hu QC, Wang WG, Tan FR, Tang XY, Zhu QL, Pan K, Li Q, Su XH: Transcriptome profiling of Zymomonas mobilis under ethanol stress. Biotechnol Biofuels. 2012, 5: 75-10.1186/1754-6834-5-75.
Yang S, Land ML, Klingeman DM, Pelletier DA, Lu T-YS, Martin SL, Guo H-B, Smith JC, Brown SD: Paradigm for industrial strain improvement identifies sodium acetate tolerance loci in Zymomonas mobilis and Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 2010, 107: 10395-10400. 10.1073/pnas.0914506107.
Yang S, Pan C, Hurst GB, Dice L, Davison BH, Brown SD: Elucidation of Zymomonas mobilis physiology and stress responses by quantitative proteomics and transcriptomics. Front Microbiol. 2014, 5: 246-
Ren C, Chen T, Zhang J, Liang L, Lin Z: An evolved xylose transporter from Zymomonas mobilis enhances sugar transport in Escherichia coli. Microb Cell Fact. 2009, 8: 66-10.1186/1475-2859-8-66.
Yang S, Pappas KM, Hauser LJ, Land ML, Chen G-L, Hurst GB, Pan C, Kouvelis V, Typas M, Pelletier DA, Klingeman DM, Chang Y-J, Samatova NF, Brown SD: Improved genome annotation for Zymomonas mobilis. Nat Biotechnol. 2009, 27: 893-894. 10.1038/nbt1009-893.
Yang S, Tschaplinski TJ, Engle NL, Carroll SL, Martin SL, Davison BH, Palumbo AV, Rodriguez M, Brown SD: Transcriptomic and metabolomic profiling of Zymomonas mobilis during aerobic and anaerobic fermentations. BMC Genomics. 2009, 10: 34-10.1186/1471-2164-10-34.
Jensen LJ, Kuhn M, Stark M, Chaffron S, Creevey C, Muller J, Doerks T, Julien P, Roth A, Simonovic M, Bork P, von Mering C: STRING 8 - a global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009, 37: D412-D416. 10.1093/nar/gkn760.
He MX, Wu B, Shui ZX, Hu QC, Wang WG, Tan FR, Tang XY, Zhu QL, Pan K, Li Q, Su XH: Transcriptome profiling of Zymomonas mobilis under furfural stress. Appl Microbiol Biotechnol. 2012, 95: 189-199. 10.1007/s00253-012-4155-4.
Weber H, Polen T, Heuveling J, Wendisch VF, Hengge R: Genome-wide analysis of the general stress response network in Escherichia coli: sigmaS-dependent genes, promoters, and sigma factor selectivity. J Bacteriol. 2005, 187: 1591-1603. 10.1128/JB.187.5.1591-1603.2005.
Jozefczuk S, Klie S, Catchpole G, Szymanski J, Cuadros-Inostroza A, Steinhauser D, Selbig J, Willmitzer L: Metabolomic and transcriptomic stress response of Escherichia coli. Mol Syst Biol. 2010, 6: 364-10.1038/msb.2010.18.
Kalnenieks U, Galinina N, Toma MM, Marjutina U: Ethanol cycle in an ethanologenic bacterium. FEBS Lett. 2002, 522: 6-8. 10.1016/S0014-5793(02)02923-X.
Kalnenieks U, Galinina N, Toma MM, Pickford JL, Rutkis R, Poole RK: Respiratory behaviour of a Zymomonas mobilis adhB::kan(r) mutant supports the hypothesis of two alcohol dehydrogenase isoenzymes catalysing opposite reactions. FEBS Lett. 2006, 580: 5084-5088. 10.1016/j.febslet.2006.08.034.
Agrawal M, Chen RR: Discovery and characterization of a xylose reductase from Zymomonas mobilis ZM4. Biotechnol Lett. 2011, 33: 2127-2133. 10.1007/s10529-011-0677-6.
Zhang X, Chen G, Liu W: Reduction of xylose to xylitol catalyzed by glucose-fructose oxidoreductase from Zymomonas mobilis. FEMS Microbiol Lett. 2009, 293: 214-219. 10.1111/j.1574-6968.2009.01529.x.
Yang S, Giannone RJ, Dice L, Yang ZK, Engle NL, Tschaplinski TJ, Hettich RL, Brown SD: Clostridium thermocellum ATCC27405 transcriptomic, metabolomic and proteomic profiles after ethanol stress. BMC Genomics. 2012, 13: 336-10.1186/1471-2164-13-336.
Abe K, Ohnishi F, Yagi K, Nakajima T, Higuchi T, Sano M, Machida M, Sarker RI, Maloney PC: Plasmid-encoded asp operon confers a proton motive metabolic cycle catalyzed by an aspartate-alanine exchange reaction. J Bacteriol. 2002, 184: 2906-2913. 10.1128/JB.184.11.2906-2913.2002.
Molenaar D, Bosscher JS, Ten Brink B, Driessen AJ, Konings WN: Generation of a proton motive force by histidine decarboxylation and electrogenic histidine/histamine antiport in Lactobacillus buchneri. J Bacteriol. 1993, 175: 2864-2870.
Azcarate-Peril MA, Altermann E, Hoover-Fitzula RL, Cano RJ, Klaenhammer TR: Identification and inactivation of genetic loci involved with Lactobacillus acidophilus acid tolerance. Appl Environ Microbiol. 2004, 70: 5315-5322. 10.1128/AEM.70.9.5315-5322.2004.
Franden MA, Pienkos PT, Zhang M: Development of a high-throughput method to evaluate the impact of inhibitory compounds from lignocellulosic hydrolysates on the growth of Zymomonas mobilis. J Biotechnol. 2009, 144: 259-267. 10.1016/j.jbiotec.2009.08.006.
Mohagheghi A, Linger J, Smith H, Yang S, Dowe N, Pienkos PT: Improving xylose utilization by recombinant Zymomonas mobilis strain 8b through adaptation using 2-deoxyglucose. Biotechnol Biofuels. 2014, 7: 19-10.1186/1754-6834-7-19.
Acknowledgments
Funding for this work was provided by the DOE Bioenergy Technologies Office (BETO). Research work conducted in SDB’s laboratory is sponsored in part by the Laboratory Directed Research and Development Program of Oak Ridge National Laboratory (ORNL) and leveraged resources supported by the BioEnergy Science Center (BESC). BESC is a Department of Energy Bioenergy Research Center supported by the Office of Biological and Environmental Research in the Department of Energy Office of Science. Oak Ridge National Laboratory is managed by UT-Battelle, LLC, for the DOE under contract DE-AC05-00OR22725. The authors wish to acknowledge Dawn M. Klingeman and Courtney Johnson at ORNL for their technical help with the RNA extraction and microarray, as well as Ali Mohagheghi at the National Renewable Energy Laboratory for his assistance with the fermentation.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Electronic supplementary material
13068_2014_140_MOESM1_ESM.docx
Additional file 1: Table S1.: Information for array data used for acetate stress response transcriptomic profiling. F: fermentor number. (DOCX 137 KB)
13068_2014_140_MOESM2_ESM.docx
Additional file 2: Figure S1.: The quality control analyses of microarray data by JMP Genomics. Figure S2. Example of the recombinant gene expression for Z. mobilis in RMG at exponential phase for each probe with two biological replicates listed together for each gene. Figure S3. Transcriptomic profiles of Z. mobilis gDNA and plasmid genes in single sugar of RMG8 or RMX8 with and without the supplementation of exogenous ammonium acetate based on tiling array data. Figure S4. Correlations between qRT-PCR and microarray. Figure S5. The ANOVA modeling of acetate shock response microarray data using JMP Genomics. Figure S6. Interactions among acetate upregulated genes and downregulated genes for Z. mobilis 8b grown in mixed sugar of RMG4X4 with at least twofold significant increase (data from Table S2-7) using the STRING precomputed protein-interaction database. Figure S7. Bioscreen C result of wild-type 8b and ZMO0128 knockout mutant 8b-KO0128 grown in RMG8 with the supplementation of ammonium acetate (15 g/L). Figure S8. Relationship among TonB-dependent receptors in Z. mobilis. (DOCX 5 MB)
13068_2014_140_MOESM3_ESM.zip
Additional file 3: Table S2.: Lists of significantly differentially expressed genes for single sugar experiment (S2A including 5 Excel sheets from S2-1 to S2-5) and for mixed sugar array study (S2B including 13 Excel sheets from S2-6 to S2-18). (ZIP 20 MB)
13068_2014_140_MOESM4_ESM.docx
Additional file 4: Table S3.: HPLC data of glucose, xylose, xylitol, glycerol, acetic acid, and ethanol (g/L) when 8b grown in RMG8, RMX8, and RMG4X4 media with and without the supplementation of NH4OAc at different time points. (DOCX 159 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Yang, S., Franden, M.A., Brown, S.D. et al. Insights into acetate toxicity in Zymomonas mobilis8b using different substrates. Biotechnol Biofuels 7, 140 (2014). https://doi.org/10.1186/s13068-014-0140-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-014-0140-8