
Abstract
Milk consumption is a hallmark of western diet. According to common believes, milk consumption has beneficial effects for human health. Pasteurization of cow’s milk protects thermolabile vitamins and other organic compounds including bioactive and bioavailable exosomes and extracellular vesicles in the range of 40–120 nm, which are pivotal mediators of cell communication via systemic transfer of specific micro-ribonucleic acids, mRNAs and regulatory proteins such as transforming growth factor-β. There is compelling evidence that human and bovine milk exosomes play a crucial role for adequate metabolic and immunological programming of the newborn infant at the beginning of extrauterine life. Milk exosomes assist in executing an anabolic, growth-promoting and immunological program confined to the postnatal period in all mammals. However, epidemiological and translational evidence presented in this review indicates that continuous exposure of humans to exosomes of pasteurized milk may confer a substantial risk for the development of chronic diseases of civilization including obesity, type 2 diabetes mellitus, osteoporosis, common cancers (prostate, breast, liver, B-cells) as well as Parkinson’s disease. Exosomes of pasteurized milk may represent new pathogens that should not reach the human food chain.
Similar content being viewed by others

Introduction
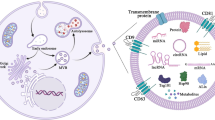
Exosomes (40–120 nm) are members of a larger spectrum of extracellular vesicles (EVs) of up to 1000 nm that mediate cell-to-cell communication and cell function [1,2,3,4]. Milk exosomes and milk microvesicles (MVs) are released from mammary gland epithelial cells (MECs) of all mammals including humans and dairy cows [5,6,7,8,9]. Exosomes are formed from inward budding of endosomes resulting in membrane-surrounded multivesicular bodies (MVBs), which are secreted by fusion of the MVBs with the cell membrane. MVs are released directly by budding of the plasma membrane like milk fat globules (MFGs). Both pathways are highly regulated and appear to be conserved amongst different species [10]. In 2013, Melnik et al. [11] postulated that “milk is not just food” but a genetic transfection system activating mechanistic target of rapamycin complex 1 (mTORC1) signaling and microRNA (miR) transfer for postnatal growth. Today, compelling evidence confirms this functional hypothesis of milk signaling. Milk’s exosomal miRs serve as a biomolecular software for maternal-neonatal communication which is important for epigenetic gene regulation that is required for developmental processes of the newborn infant [12]. Abundantly present miRs in milk-derived EVs including miR-148a are highly conserved between mammals [13]. Various exosome-specific proteins, lipids, mRNAs, circular RNAs, non-coding miRs and regulatory proteins such as transforming growth factor-β (TGF-β) are crucial signaling components delivered by milk exosomes [5, 6, 14, 15]. Evidence has been provided that breast milk exosomes and their miR cargo play a key role for the appropriate maturation of the intestine, development of the gut microbiome and programming of the intestinal mucosa-associated lymphatic tissue (MALT) as well as thymic T cell differentiation [16,17,18,19,20,21,22,23,24,25,26]. The deficiency of milk exosomes in artificial formulas increases the risk for inappropriate metabolic and immunological programming of the newborn infant [8, 9, 18, 19], a major determinant for the development of diseases of civilization in later life such as allergic diseases and obesity [18, 19]. Under physiological conditions, the transfer of milk-derived exosomes and their miR-mediated impact on epigenetic regulation is restricted to the period of maternal lactation in all mammals, except Neolithic humans, who are exposed to dairy milk exosomes after the nursing period for several decades. Since the 1950s, when widely available refrigeration technology allowed the distribution of pasteurized milk and milk products, bioactive bovine milk exosomes entered the human food chain in a large scale (Fig. 1). It is the intention of this review article to provide epidemiological and translational evidence that dairy milk-derived exosomes and their cargo contribute to the pathogenesis of common diseases of civilization and should thus be regarded as critical pathogens, that have to be eliminated from the human food chain.

Transfer of dairy milk exosomes to the human milk consumer. Genetic dairy cow selection enhances mammary epithelial cell miR-148a expression, a crucial epigenetic mechanism enhancing milk yield that potentially also increases milk exosome miR-148a content. Persistent pregnancy of dairy cows further promotes estrogen-stimulated expression of miR-148a and miR-21. Milk exosomes also contain miR-155 and transforming growth factor-β (TGF-β), which promotes the expression of miR-155. Pasteurization has no significant effect on milk exosome integrity and exosomal miR bioavailability. Large scale pasteurization and cooling technology promoted the persistent entry of dairy milk exosomes and their miRs into the human food chain
Dairy milk exosomes and their miR cargo are bioavailable for the milk consumer
Reinhardt et al. [27] characterized the proteome of bovine milk exosomes and reported a greatly reduced presence of MFG membrane (MFGM) proteins in the fraction of cow milk exosomes, which suggests that milk exosome secretion pathways originate from Golgi and differ from that of MFGs, which resemble holocrine secretion of lipid droplets directly from the endoplasmic reticulum (ER). Bovine milk exosomes (50–100 nm) isolated by ultracentrifugation from the 100,000×g pellet from the milk of mid-lactation Holstein cows are enriched in tumor susceptibility gene-101 (TSG101), a protein component of the vesicular trafficking process and depleted in MFGM proteins such as lactaderin/MGFE8 [26]. Benmoussa et al. [28] confirmed that cow milk exosomes of the 100,000×g pellet fraction are positive for the exosome markers TSG101, apoptosis-linked gene 2-interacting protein X (ALIX), heat shock protein 70 (HSP70) and contain bovine miR-223 and miR-125b. A large quantity of bovine milk miR-223 and miR-125b resisted digestion under simulated gastrointestinal tract conditions, which supports their bioaccessibility [28]. Recently, a subset of milk MVs (100 nm in diameter) with proteins commonly found in MFGM has been characterized that sediments at low speed ultracentrifugation (35,000×g) and contains and protects the bulk of milk miRs from degradation [29, 30]. At present, there is a lack of information on the potential systemic biological effects and trafficking characteristics of this 35 K subset of milk EVs to the milk consumer.
This review focuses on milk exosomes of the 100,000×g fraction (100 K). It is generally appreciated that exosomes participate in cell-to-cell communication and gene regulation, facilitated by the transfer of miRs, proteins and lipids from donor to recipient cells. Bovine milk exosomes contain nearly 400 miRs and selected proteins [31,32,33,34] that resist the harsh conditions in the gastrointestinal tract [14, 28, 32], are taken up via receptor-mediated endocytosis by intestinal epithelial cells [35], vascular endothelial cells [36], and reach distant tissues across species boundaries after oral administration [35,36,37,38,39]. The most sophisticated and extensively controlled study of Manca et al. [39] recently demonstrated that bovine milk exosomes derived from commercial pasteurized skim milk reached the systemic circulation of mice and distributed widely among murine tissues. A variety of different tracers used in their study suggests that milk exosomes and their miR cargo accumulate in the brain, an important finding, which is consistent with proven exosomal delivery of Cre-recombinase to the brain [40]. Bovine miRs were analyzed by RNase H2-dependent PCR (rhPCR) in plasma collected from 11 healthy volunteers before and 6 h after consumption of 1.0 L of commercial 1%-fat cow’s milk. This method (rhPCR) is able to distinguish between bovine and human miRs with small variations in the nucleotide sequence. Notably, plasma concentrations of Bos taurus (bta)-miR-21-5p and bta-miR-30a-5p were > 100% higher 6 h after milk consumption than before milk intake, a finding confirming the bioavailability of dairy milk exosomes in humans [41]. The majority of dairy milk miRs including miR-148a, miR-21, miR-29b and miR-155 survive pasteurization and refrigerated storage but are significantly reduced after boiling or ultra-heat treatment (UHT) [32, 42,43,44,45]. Baier et al. [46] demonstrated the bioavailability of milk-borne miRs in humans using commercial milk (1% fat) that contained 148 ± 42 pmol/L of miR-29b. In a dose-dependent manner, human volunteers absorbed considerable amounts of miR-29b from cow milk resulting in a plasma peak of miR-29b at about 4 h to 6 h postprandial associated with an intracellular increase of miR-29b in peripheral blood mononuclear cells (PBMC) [46]. Furthermore, it has been demonstrated that bovine milk exosomes are taken up by human macrophages [47].
Recent evidence underlines that bacterial fermentation of milk decreases the size, protein- and miR content of milk exosomes [48]. It has been demonstrated that milk-derived exosomes are taken up by Escherichia coli K-12 MG1655 and Lactobacillus plantarum WCFS promoting bacterial growth [49]. In contrast to pasteurization (78 °C), boiling (100 °C), and ultra-heat treatment (130 °C) of milk decreased the levels of milk miRs [43, 44].
Taken together, there is compelling evidence that dairy milk exosomes of pasteurized commercial milk reach the systemic circulation and tissues of the human milk consumer. Whereas human breast milk-derived exosomes are of critical importance for infant health and appropriate development, programming and tissue maturation [5,6,7, 11, 16,17,18,19,20,21,22,23,24,25,26], continued exposure of humans to dairy milk-derived exosomes after the nursing period may exert adverse effects on human health.
Allergic diseases
Exosomes and exosomal miR signaling play a key role during postnatal programming and tissue maturation of the infant [5,6,7,8,9, 50]. Breastfeeding has a protective effect on the prevention of allergic rhinitis, allergic asthma and atopic dermatitis [51]. Breastfeeding in contrast to commercial artificial formula feeding is regarded as the most efficient primary prevention of allergic asthma in childhood [52,53,54,55]. Allergy-prone and allergic individuals exhibit reduced numbers and function of regulatory T cells (Tregs) [56]. Forkhead box P3 (FoxP3) is the master transcription factor of Tregs and controls Treg differentiation and maintenance of Treg-mediated immune tolerance [57,58,59]. Tooley et al. [60] demonstrated that maternal rat milk, but not formula, prevented β-lactoglobulin-induced allergy in rat pups. Thus, maternal milk in contrast to formula contains an ingredient conferring an allergy-preventive effect. Notably, Admyre et al. [16] showed that the addition of human breast milk exosomes to PBMCs increased the number of FoxP3+CD4+CD25+ Tregs in a dose-dependent manner. FOXP3 gene expression is controlled by epigenetic mechanisms as well as TGF-β [61,62,63]. The Treg-specific demethylation region (TSDR) is a critical region of the FOXP3 promoter, which controls FoxP3 expression. TSDR methylation reduces FoxP3 expression, whereas TSDR demethylation promotes FoxP3 expression, respectively [61, 62]. In allergic individuals, an increased TSDR methylation has been observed [64, 65]. In contrast, tolerance induction and cessation of allergy was associated with TSDR FOXP3 demethylation [65, 66]. Both DNA methyltransferase 1 (DNMT1) and DNMT3B are associated with the FOXP3 locus in CD4+T cells [67]. miR-148a directly targets DNMT1, whereas miR-21 indirectly downregulates DNMT1 expression by targeting an important autoimmune gene, Ras guanyl nucleotide-releasing protein 1 (RASGRP1), which mediates the Ras-MAPK pathway upstream of DNMT1 [68]. miR-29b is another miR species that negatively regulates DNMT1 expression [69,70,71]. Importantly, miR-148a, miR-21, miR-29b and miR-155 are cargos of human and bovine milk exosomes [17, 23, 35, 45, 48, 72]. Golan-Gerstl et al. [44] demonstrated that incubation of breast milk exosomes with intestinal cells increased their miR-148a content resulting in decreased expression of DNMT1. Importantly, miR levels in dairy milk were not significantly affected by pasteurization [43,44,45].
Consumption of raw cow milk during early infancy exhibited a preventive effect on the development of allergic diseases and increased the number of FoxP3 + Tregs [73,74,75]. The existence of a postnatal window for milk-induced Treg maturation has been proposed [76]. We hypothesized that thymic maturation of Tregs is mediated by milk-derived exosomes [18, 19]. miR-155 plays a critical role in the maturation of thymic Tregs [18, 19]. miR-155 targets suppressor of cytokine signaling 1 (SOCS1), a critical inhibitor of signal transducer and activator of transcription 5 (STAT5), which promotes the expression of FoxP3 [77]. Higher amounts of SOCS1 protein suppress IL-12 and IFNγ signaling inhibiting Th1 cell differentiation, while promoting Th2 cell induction [78, 79]. TGF-β as well promotes thymic Treg (tTreg) cell development by inducing FoxP3 expression repressing T cell clonal deletion and peripheral Treg cell differentiation [63, 80]. Notably, bovine milk exosomes contain both miR-155 and TGF-β [14, 32, 45], a fact, that further supports milk exosome-driven maturation of tTregs [18, 19]. It is likely that milk exosomes or exosome-derived molecules, which reach distant tissue including the brain may also accumulate in the thymus [39], an organ exhibiting extensive exosome traffic [81,82,83]. Milk exosomes may thus support thymic epithelial cell-derived exosomes in tTreg differentiation and maturation, a potential augmenting mechanism which may explain allergy prevention early in life by consumption of raw cow’s milk, during a period when the infant’s thymus is still functional operative (Fig. 2).

Milk exosomes and allergy prevention. Human breast milk and dairy milk exosomes transfer miR-148a and miR-29b, which both suppress DNA methyltransferase 1 (DNMT1). DNMT1 controls the methylation status of the Treg-specific demethylation region (TSDR) on the FOXP3 promoter. DNMT1 suppression (TSDR hypomethylation) increases FoxP3 expression. Milk exosome-derived miR-155 inhibits suppressor of cytokine signaling 1 (SOCS1), a negative regulator of the JAK-STAT pathway that increases the expression of signal transducer and activator of transcription 5 (STAT5) promoting FoxP3 expression. Milk exosome-derived transforming growth factor-β (TGF-β) enhances SMAD5 signaling that further increases FoxP3 expression, especially in the thymus. Milk exosomes thus promote the induction of FoxP3, the master transcription factor of regulatory T cells (Tregs), the potential mechanism preventing allergy development by breast feeding or raw farm milk consumption during early infancy
In addition, milk exosomes may have a direct effect on MALT homeostasis. TGF-β, a component of milk exosomes [14], has been shown to induce miR-155 in both freshly isolated and lamina propria T cell lymphoblasts [84]. miR-155 targets IL-2 inducible T-cell kinase (ITK) and decreases ITK and IL-2 mRNA suggesting a TGF-β-dependent function for miR-155 in modulating cytokine and T-cell immune responses in the gut [84]. Intriguingly, the concentration of TGF-β1 in colostrum samples from mothers of infants with IgE-mediated cow’s milk allergy (CMA) was significantly lower than from mothers of infants with non-IgE-mediated CMA [85] pointing to an important role of TGF-β/miR-155 signaling in intestinal immune homeostasis.
In contrast to breastfeeding, artificial infant formula powder contains no bioactive exosomes and only minor amounts of exosomal TGF-β and miRs (for instance < 10% of miR-148a) compared to raw cow’s milk [32, 86]. This may be a reasonable explanation for the superiority of breastfeeding in allergy prevention compared to formula feeding [18, 19]. These data strongly indicate that milk exosomes are of critical importance for the maturation of the immune system during the postnatal period and early infancy.
Fetal macrosomia
The Developmental Origins of Health and Disease (DOHaD) hypothesis underlines the impact of prenatal and postnatal epigenetic factors in the transmission of obesity and cardiovascular diseases [87, 88]. Accelerated fetal growth and increased birth weight are well-known risk factors for the development of obesity and T2DM [89,90,91]. Disturbances of the intrauterine milieu can induce lifelong deviations of metabolic programming [92]. Exosomes have been identified as key players for fetal-maternal communication and vice versa [93]. As milk exosomes and their cargo products are able to overcome tissue barriers including the intestinal and blood–brain barrier and are distributed in various tissues [39], it is conceivable that they may also reach the placenta of women consuming pasteurized milk. Worldwide gynecological societies such as the American College of Obstetricians and Gynecologists recommend increased milk and dairy consumption during pregnancy as a rich source of calcium [94]. During 1996–2002, the Danish National Birth Cohort collected data on midpregnancy diet of 50,117 mother-infant-pairs and ascertained birth outcomes [95]. This study demonstrated that increased milk consumption during pregnancy was associated with an increase in placental and birth weight [95]. Maternal milk consumption, fetal growth, and the risks of neonatal complications have been investigated in the Generation R Study in Rotterdam including 3405 mothers [96]. Maternal milk consumption of > 3 glasses/day was associated with greater fetal weight gain in the third trimester of pregnancy, which led to an 88 g higher birth weight than that with milk intake of none to 1 glass/day. In addition, head circumference tended to be 2.3 cm larger when mothers consumed > 3 glasses/day. This association appeared to be limited only to milk, whereas protein intake from non-dairy food or cheese was not associated with an increase in birth weight [96]. A systematic review of all studies and case reports supported the conclusion that only milk consumption but not the intake of fermented milk/milk products increased birth weight [97, 98]. Thus, some compounds of unfermented milk not related to milk proteins, accelerate fetal growth.
Two independent studies confirmed increased expression of miR-21 in placenta tissue of infants born with macrosomia (birth weight > 4000 g) [99, 100]. Milk miR-21, a signature miR of commercial dairy milk and cargo of bovine milk exosomes [71, 72, 80], most likely reaches the placenta and increases placental and trophoblast growth. miR-21 plays important roles in growth of trophoblastic cell lines [101] and increases PI3K-AKT- and mTORC1 signaling by suppression of various key inhibitory checkpoints such as PTEN [11]. Stimulation of trophoblast mTORC1 activity enhances the transfer of branched-chain amino acids (BCAAs) to the fetus, a constellation that may promote BCAA-mTORC1-driven fetal macrosomia [102,103,104,105,106,107] (Fig. 3). About 5–10% of cases exhibiting Beckwith-Wiedemann syndrome (BWS), an overgrowth syndrome characterized by macrosomia, macroglossia, and abdominal wall defects, are caused by loss-of-function mutations of cyclin-dependent kinase inhibitor 1C (CDKN1C, p57kip2) [108,109,110,111,112]. CDKN1C is the cyclin-dependent kinase inhibitor of G1 cyclin complexes that functions as a negative regulator of cellular growth and proliferation [113]. Notably, miR-21 is one of several miRs that directly target CDKN1C [113], a further epigenetic mechanism linking dairy milk exosome intake during pregnancy to fetal macrosomia.

Dairy milk exosomes and fetal macrosomia. Milk exosome-derived miR-21 may increase placental miR-21 content promoting mTORC1 signaling via inhibition of phosphatase and tensin homolog (PTEN) and other regulatory checkpoints. Increased mTORC1-mediated placental growth enhances the nutrient transfer to the fetus. In the trophoblast, upregulated mTORC1 increases the expression of L-type amino acid transporters (LAT) and glucose transporter 1 (GLUT1), thus overstimulating the diaplacental flux of branched-chain amino acids (BCAAs) and glucose to the fetus promoting fetal overgrow (macrosomia). miR-21 also targets CDKN1C, a critical checkpoint for fetal growth mutated in Beckwith-Wiedemann syndrome
Adipogenesis and obesity
The worldwide epidemic of obesity is a growing health problem, associated with increased risk of chronic diseases especially type 2 diabetes mellitus (T2DM). Young mice who had long-term ad libitum access to commercial whole cow’s milk in comparison to mice that received low fat milk or controls that had no access to dairy milk exhibited increased body weight and epididymal fat mass [114]. There is increasing interest in the role of exosomes and exosome-transferred miRs in the regulation of mesenchymal stem cell (MSC)-derived adipogenesis [115,116,117,118,119,120]. miRs regulate adipogenic lineage commitment in MSCs and hence govern fat cell numbers [115]. MSCs arise from a variety of tissues, including bone marrow and adipose tissue and, accordingly, have the potential to differentiate into multiple cell types, including osteoblasts and adipocytes [117]. An inverse relationship exists in adipogenic and osteogenic lineage commitment and differentiation, such that signaling pathways induce adipogenesis at the expense of osteogenesis and vice versa [117]. Peroxisome proliferator-activated receptor γ (PPARγ) is known to function as a master transcriptional regulator of adipocyte differentiation, but inhibits osteoblast differentiation [117]. In contrast, inducers of osteogenic differentiation, such as bone morphogenetic protein (BMP) and wingless-type MMTV integration site family members (Wnt), inhibit the function of PPARγ transactivation during MSC differentiation towards adipocytes [120]. Notably, MSCs differentiated on osteoblast extracellular matrix (ECM) with adipogenic exosomes showed expression of adipogenic lineage genes, while MSCs differentiated on adipocyte ECM with osteoblast exosomes showed osteogenic lineage genes [118]. These findings indicate that exosomes might override ECM-mediated instructive signals during lineage specification of MSCs [118]. Accumulating evidence indicates that miRs act as switches for MSCs to differentiate into either osteogenic or adipogenic lineages [120]. Based on these observations, it is conceivable that dairy milk-delivered exosomes and their miR cargo as well may interfere with MSC-derived adipogenesis and osteogenesis.
miR-148a, a component of milk exosomes, is increased in adipose tissues from obese individuals and mice fed a high-fat diet (HFD) [121]. miR-148a suppresses its target gene Wnt1, an endogenous inhibitor of adipogenesis. Ectopic expression of miR-148a accelerated differentiation and partially rescued Wnt1-mediated inhibition of adipogenesis, whereas knockdown of miR-148a inhibited adipogenesis [121, 122]. In addition, miR-148a has been shown to silence Wnt10b, a further endogenous inhibitor of adipogenesis during 3T3-L1 cell differentiation [123]. A further study demonstrated that increased expression of miR-148a via suppression of DNMT1 enhanced adipocyte differentiation [124]. In the absence of DNMT1, adipocyte-specific gene expression and lipid accumulation occurred precociously [124]. Yang et al. [125] recently demonstrated that DNA methylation biphasically regulates 3T3-L1 preadipocyte differentiation [125]. Inhibition of DNA methylation at late stage of preadipocyte differentiation promoted lipogenesis and adipocyte phenotype in 3T3-L1 cells. This is likely mediated by induction of sterol regulatory element-binding transcription factor 1c (SREBF1c), whose promoter activity is upregulated by DNA demethylation during adipogenesis [125]. Persisting transfer of milk exosomal miR-148a may thus enhance SREBF1c-mediated lipid accumulation in adipocytes (Fig. 4). Remarkably, the MIR148A gene has been identified as an obesity risk gene in humans exhibiting single nucleotide polymorphisms which enhance miR-148a expression [126,127,128].

Dairy milk exosomes and adipogenesis. miR-21 induces the differentiation of mesenchymal stem cells (MSCs) towards adipocytes via activation of peroxisome proliferator-activated receptor PPARγ (PPARγ). miR-148a directly targets and suppresses the inhibitors of adipogenesis Wingless 1 (WNT1) and WNT10B increasing the expression of PPARγ and CCAAT/enhancer binding protein α (C/EBPα). miR-148a-mediated suppression of DNMT1 via promoter hypomethylation increases the expression of fat mass and obesity-associated gene (FTO), PPARγ and sterol regulatory element binding-transcription factor 1 (SREBF1). The mRNA demethylase FTO removes a m6A mark on RUNX1T1 mRNA generating its short splice variant RUNX1T1-S, which relieves RUNX1T1-mediated inhibition of C/EBPβ. Activated C/EBPβ activates the key adipogenic transcription factors C/EBPα and PPARγ. miR-148a targets PRKAA1, the catalytic α-unit of AMP-activated protein kinase (AMPK), the key negative regulator of mechanistic target of rapamycin complex 1 (mTORC1). mTORC1 activation enhances the expression of PPARγ and SREBF1, key lipogenic transcription factors. In addition, miR-148a targets salt-inducible kinase 1 (SIK1), and thereby relieves its inhibitory action on SREBF1. Milk exosome-derived miR-148a is thus an adipogenesis promoting factor that operates at pivotal regulatory checkpoints enhancing the risk of obesity
miR-21, another signature miR of bovine milk exosomes, is involved in adipocyte differentiation [129,130,131,132]. Kim et al. [129] showed that miR-21 governs human adipose tissue-derived MSC differentiation towards adipocytes. Furthermore, a correlation between miR-21 level and adipocyte numbers in the epididymal fat of mice fed a HFD has been observed [129]. Mei et al. [130] reported that overexpression of miR-21 in MSCs elevated the expression level of the differentiation-associated gene PPARγ, whereas miR-21 knockdown reduced PPARγ expression. miR-21 modulated ERK-MAPK activity by repressing Sprouty 2 (SPRY2), a known regulator of the receptor tyrosine kinase signaling pathway, that controls the magnitude of ERK-MAPK signaling during MSC differentiation [130]. Kang et al. [131] confirmed that miR-21 promotes adipocyte differentiation. It has recently been demonstrated that miR-21 expression was twofold greater in adipose tissue of patients with T2DM [132].
miR-29b, another abundant exosome-derived miR of cow’s milk, is also involved in adipogenesis [133]. During normal adipogenic differentiation of adipose tissue-derived stromal cells, upregulation of miR-29b promoted adipogenesis. Remarkably, miR-29 family members enhance lactation performance in dairy cow MECs via suppression of DNMT3A and DNMT3B. In contrast, inhibition of miR-29 s caused global DNA hypermethylation and increased the methylation levels of promoters of lactation-related genes, including casein αs1 (CSN1S1), E74-like factor 5 (ElF5), PPARγ, SREBF1, and glucose transporter 1 (GLUT1) and thereby reduced the secretion of lactoprotein, triacylglycerols and lactose by dairy cow MECs [133]. Thus, promoter demethylation of lipidogenic genes via miR-mediated DNMT suppression enhances both adipogenesis and lactation.
Overexpression of miR-155 in mice has been shown to reduce brown adipose tissue (BAT) mass [134]. Thus, milk exosome-derived miR-155 may attenuate BAT differentiation and thermogenesis via BAT, an unfavorable condition promoting lipid and energy storage in white adipose tissue (WAT) further promoting obesity.
Hyperphagia
Whole cow’s milk consumption in young mice not only increased body weight but also caloric intake [114]. The suppression of satiety signals during the period of lactation may be an intrinsic mechanism of milk signaling to enhance anabolism during the postnatal growth phase. As milk exosomes and their cargo products pass the blood–brain barrier and reach the brain [39, 135, 136], they may interfere with hypothalamic control centers of satiety feedback regulation. It has recently been shown that hypothalamic stem cells control ageing speed partly through exosomal miRs [137]. The brain-gut-axis is an interdependent system affecting neural functions and controlling eating behavior [138]. One of the hormones sending satiety signals to the hypothalamus is cholecystokinin (CCK), which is secreted from intestinal mucosa cells when the duodenum is filled with food [138]. CCK binds and signals via CCK1 receptor (CCK1R) and CCK2R. CCK2R knock out mice developed obesity that was associated with hyperphagia [139]. Suppression of feeding and concomitantly increased expression of hypothalamic proopiomelanocortin after intracerebroventricular injection of gastrin into control mice demonstrates that hypothalamic CCK2Rs mediate inhibition of food intake [139]. CCK2R deletion was associated with increased body weight and hypothalamic neuropeptide Y (NPY) content, which explains the increased food intake in CCK2R knockout mice [140]. Notably, the gene expressing CCK2R (CCKBR) is a direct target gene of miR-148a [141]. Thus, miR-148a of milk exosomes via suppression of satiety signals may maintain the state of a “hungry brain”, advantageous for postnatal growth but critical for long-term energy balance in adults (Fig. 5).

Milk exosomes and hyperphagia. Milk, a feeding and signaling system promoting postnatal anabolism and growth, most likely interferes with satiety control in the hypothalamus, which is possible as milk exosomes accumulate in the brain. Cholecystokinin (CCK) is released by duodenal I-cell during intestinal nutrient abundance. CKK is an important hormone that induces satiety signals in the hypothalamus via binding to CCK receptor 2 (CCKR2). CCKR2 is a direct target of miR-148a. It is thus conceivable that milk exosomes maintain a “hungry brain” to increase milk intake during the breastfeeding period. Persistent milk exosome intake by consumption of pasteurized cow’s milk may maintain this hyperphagic state, a further mechanism promoting obesity
Type 2 diabetes mellitus
T2DM is an increasing epidemic in developed countries and is closely linked to obesity (diabesity). Most epidemiological studies and systematic reviews promote the view that milk and dairy products are good for metabolic health and may reduce the risk of T2DM [142,143,144,145,146,147]. Dairy product intake may be inversely associated with risk of T2DM, but the evidence is inconclusive for total dairy products and sparse for individual types of dairy products. There are only few epidemiological studies that compare the risk of milk versus fermented milk/products. This, however, is of critical importance because fermentation of milk negatively affects the bioactivity of milk exosomes and their miR cargo [48]. The largest study that investigated the association of T2DM with intake of milk versus fermented milk products is the European Prospective Investigation into Cancer and Nutrition (n = 340,234) [148]. Whereas the consumption of fermented milk and fermented milk products confirmed an inverse relation to T2DM risk, the intake of non-fermented milk showed an increased risk for T2DM [148]. Furthermore, the Physicians’ Health Study (n = 21,660) demonstrated a significant increase in T2DM risk in relation to the consumption of whole milk [149]. Data presented from the Framingham Heart Study Offspring Cohort demonstrated a nonlinear correlation between milk consumption and prediabetes (defined by fasting glucose plasma levels ≥ 100 to < 126 mg/dl). More than 5 servings of milk per week significantly increased the risk of prediabetes [150]. None of these studies considered thermal milk processing (pasteurization versus UHT), which is of crucial importance for the bioavailability and function of exosomes in commercial milk products.
Milk protein provides abundant essential BCAAs including leucine, which activates mTORC1 [151], a key driver of anabolism, growth and insulin secretion [152]. Elevated BCAA plasma levels correlate with an increased risk of insulin resistance and T2DM [153,154,155,156,157,158,159].
Cow milk exosomes provide miR-29b, an abundant miR of dairy milk that survives pasteurization and exhibits the same nucleotide sequence as human miR-29b [42]. Consumption of pasteurized cow’s milk by healthy volunteers increased plasma levels of miR-29b including intracellular miR-29b levels in PBMCs [46]. Notably, diabetes researchers regard the miR-29s as a diabetogenic miR family [160,161,162,163,164]. Intriguingly, miR-29b controls the expression of branched chain α-ketoacid dehydrogenase (BCKD) complex in the cell via targeting the BCKD core protein dihydrolipoamide branched-chain acyltransferase (DBT) [165]. In accordance to a recent study, early-onset and classical forms of T2DM showed impaired expression of BCKD genes involved in muscle BCAA catabolism [166]. miR-29b-mediated inhibition of BCKD activity decreases BCAA catabolism, a meaningful metabolic switch for the newborn mammal. The essential BCAAs are required for the synthesis of many functional and structural proteins [167]. Therefore, BCAAs should not be wasted for purposes of energy generation during postnatal growth [154]. In addition, BCAAs play a key role as activators of mTORC1, which orchestrates cell growth and anabolism [168,169,170,171]. In cells with impaired leucine catabolism, mTORC1 signaling towards phosphorylation of ribosomal protein S6 kinase 1 (S6K1) was significantly increased [172].
Insulinotropic amino acids, especially leucine and glutamine, are amino acids highly enriched in milk proteins that are capable to increase insulin secretion [173,174,175]. Leucine supplementation in mice stimulated insulin secretion of pancreatic islets, which was associated with an activation of the PI3K/AKT/mTORC1 pathway [174]. Insulin has growth-promoting effects and binds to insulin- and insulin-like growth factor 1 (IGF-1) receptors. Insulin regulates appetite, body temperature, white fat mass, and glucose metabolism. Importantly, insulin signaling modulates neurotransmitter activity, neuronal function and synaptogenesis, critical events during the postnatal period [176]. Inhibition of BCKD by exosomal miR-29b may serve to increase β-cells BCAA levels further promoting mTORC1-mediated insulin secretion during the postnatal growth phase. However, this is a critical regulatory switch enhancing endoplasmic reticulum (ER) stress and β-cell apoptosis in the long run [151]. In fact, chronic exposure to leucine in vitro has been shown to induce β-cell dysfunction in INS-1E cells and mouse islets [177].
Milk miR-29b-mediated increases in BCAA levels and BCAA-driven mTORC1 activation in peripheral tissues explain insulin resistance by S6K1-mediated inhibitory phosphorylation of insulin receptor substrate 1 (IRS-1), a key checkpoint of insulin signaling [178,179,180,181] (Fig. 6). SPARC (secreted protein acidic and rich in cysteine, also known as osteonectin or BM-40) may represent an important link between obesity and T2DM [182]. Overexpression of SPARC in cultured β-cells resulted in a 2.4-fold increase in insulin secretion in high glucose conditions [183]. Reduced SPARC expression was demonstrated in primary islets from subjects with diabetes compared with controls [183]. It has been demonstrated that SPARC is a direct target of miR-29b [184, 185]. Importantly, overexpression of miR-29s reduced glucose uptake and GLUT4 levels [185].

Dairy milk exosomes and type 2 diabetes mellitus. In peripheral tissues (muscle and adipose tissue), miR-29b promotes insulin resistance via inhibiting the core protein dihydrolipoamide branched-chain acyltransferase (DBT) of branched-chain alpha-ketoacid dehydrogenase (BCKD), the rate limiting enzyme of branched-chain amino acid (BCAA) degradation. Resulting increases in intracellular BCAA levels enhance mTORC1-S6K1 activity. Overstimulated S6K1 phosphorylates and inhibits insulin-receptor substrate 1 (IRS-1) thereby decreasing insulin signaling and glucose transporter 1 (GLUT1) translocation to the cell membrane. In addition, miR-29b-mediated suppression of secreted protein acidic and rich in cysteine (SPARC) reduces GLUT4 activity. miR-29b-mediated attenuation of BCAA degradation increases BCAA-mTORC1-mediated insulin synthesis. miR-29b-mediated suppression of SPARC reduces insulin secretion. In addition, miR-148a-mediated suppression of V-Maf musculoaponeurotic fibrosarcoma oncogene homolog B (MAFB) in the β-cell reduces glucose-dependent insulin secretion. Continued miR-29b-mediated overstimulation of insulin synthesis and miR148a- and miR-29b-mediated impairment of insulin secretion enhances endoplasmic reticulum (ER) stress promoting β-cell apoptosis, the hallmark of type 2 diabetes mellitus
The Maf basic leucine-zipper-containing transcription factor MAFB is required for the generation of functional β-cell populations by directly activating insulin gene transcription and key regulators of β-cell differentiation and function [186]. Importantly, MAFB increases the expression of MAFA, which is important to maintain β-cell function in adults [187]. Notably, MAFB is a direct target of miR-148a [188]. Suppressed expression of MAFB in murine and human β-cells has been associated with decreased glucose-dependent insulin secretion [189].
Thus, persistent milk exosome miR-29b-BCAA-mediated overstimulation of insulin synthesis and BCAA-mTORC1-dependent insulin resistance identify dairy milk exosomes as potential promoters of T2DM. Both, miR-29b-mediated suppression of SPARC and miR-148a-mediated suppression of MAFB impair insulin secretion, a potential mechanism enhancing ER stress and β-cell apoptosis.
Atherosclerosis, cardiovascular and overall mortality
Atherosclerosis and cardiovascular disease are the major causes of death in industrialized countries. Two large Swedish cohorts, one with 61,433 women and one with 45,339 men determined the association between milk consumption and time to mortality [190]. For every glass of milk, which in Sweden is primarily pasteurized milk, the adjusted hazard ratio of all-cause mortality was 1.15 (1.13–1.17) in women and 1.03 (1.01–1.04) in men, respectively. A recent study from Northern Sweden including 103,256 adult participants reported that high consumers of nonfermented milk (≥ 2.5 times/day) had a 32% increased hazard (HR: 1.32; 95% CI 1.18, 1.48) for all-cause mortality compared with that of subjects who consumed milk ≤ 1 time/week [191]. In contrast, fermented milk intake and cheese intake were negatively associated with mortality [191]. A systematic review and updated dose–response meta-analysis of prospective cohort studies partially funded by dairy associations reported a 7% lower risk of stroke with an increment of 200 g milk daily [192]. The association of milk with total stroke was nonlinear, with the strongest inverse association around 125 g/day. For milk intake in the range of 125–750 g/day the inverse association remained significant, but was attenuated. Based on the same studies, whole milk intake was significantly associated with a higher risk of stroke per 200 g/day with no heterogeneity. In contrast, total fermented dairy intake (200 g/day) was associated with a 9% lower risk of stroke [192]. The Prospective Urban Rural Epidemiology (PURE) study investigators reported no increase in cardiovascular mortality by dairy intake including milk consumption in low-income and middle-income countries. Milk intake > 1 serving vs no intake was associated with lower risk of cardiovascular mortality [193]. The category > 1 serving vs no intake is not suitable to identify a dose-relationship between milk intake and mortality risk. Yogurt was associated with lower risk of cardiovascular mortality [193]. Again, all these questionnaire-based studies did not provide information on the type of thermal processing (pasteurized vs UHT) of milk.
Whereas whole milk consumption appears to be associated with an increased risk of mortality, fermented milk and fermented milk products are not. The presence of milk-derived bioactive exosomes and their miR content may play a key role explaining this discrepancy. Non-coding RNAs and miRs are in the recent focus of lipid and atherosclerosis research [194, 195]. During atherosclerosis, the gradual accumulation of lipids into the subendothelial space of damaged arteries results in several lipid modification processes followed by macrophage uptake in the arterial wall. Cholesterol accumulation within monocyte-derived macrophages and their transformation into foam cells make up the characteristic fatty streaks observed in the early stages of atherosclerosis [196, 197]. Notably, milk-derived exosomes and their miR content are taken up by human macrophages [47]. Exosome-derived miRs are regarded as potential biomarkers of atherosclerosis [198, 199]. It has recently been demonstrated that miR-148a promotes the differentiation of monocytes into macrophages and induces M1 but inhibits M2 polarization [200]. Macrophages overexpressing miR-148a exhibited enhanced ability to engulf and kill bacteria, which was mediated by excessive production of reactive oxygen species (ROS). Furthermore, PTEN has been detected as a direct target gene of miR-148a in macrophages. Macrophages overexpressing miR-148a via upregulation of AKT signaling increased the production of ROS and pro-inflammatory cytokines through upregulation of NF-κB signaling [200]. Peritoneal macrophages of organic dust-exposed mice which were fed a milk exosome-enriched diet exhibited an M1 shift compared to an M2 phenotype in mice fed a milk exosome-deficient diet [201]. In macrophages of mice which received a diet enriched in milk exosomes, interleukin 6 (IL-6), TNF, and IL-12/23 were significantly elevated [201]. Remarkably, a dose-dependent correlation between milk consumption and plasma levels of IL-6 has been reported in humans [190].
miR-148a directly targets the expression of low density-lipoprotein (LDL) receptors (LDLR) [202, 203], the pivotal regulators of cholesterol homeostasis and hepatic LDL clearance [204]. In addition, miR-148a directly targets ATP-binding cassette transporter 1 (ABCA1) [202], the key player for reverse cholesterol transport [205, 206]. For the growing infant, milk exosome-derived miR-148a via suppression of LDL-mediated hepatic cholesterol uptake and impairment of reverse cholesterol transport from peripheral tissues may serve to provide sufficient amounts of cholesterol for growth of distant tissues as well as steroid hormone biosynthesis. The persistence of this lipid metabolic switch into adulthood may however exert atherogenic effects. ABCA1 expression is induced during differentiation of human monocytes into macrophages in vitro [205]. In macrophages, both ABCA1 mRNA and protein expression are upregulated in the presence of acetylated low-density lipoprotein (AcLDL) [205, 206]. Milk exosome-derived miR-148a via targeting ABCA1 may thus attenuate macrophage cholesterol efflux promoting foam cell formation (Fig. 7). In fact, histopathologic examination of ABCA1(−/−) mice at ages 7, 12 and 18 months demonstrated a striking accumulation of lipid-laden macrophages [207]. It has recently been demonstrated that when miR-148a/152 was overexpressed, DNMT1 expression was suppressed, whereas the expression of adipose differentiation-related protein (ADRP) was enhanced, and the contents of total cholesterol (TC) and cholesteryl ester (CE) were increased in cultured macrophage foam cells [208]. Conversely, downregulation of miR-148a/152 led to elevated DNMT1 expression, reduced ADRP expression, and lowered contents of TC and CE [208]. Antisense miR-148a administration has recently been proposed as a new treatment option of atherogenic dyslipidemia [203].

Dairy milk exosomes and atherogenesis. Milk exosomes are taken up by monocytes and macrophages. miR-148a stimulates the differentiation of monocytes to macrophages, especially of macrophages of the pro-inflammatory M1 type. miR-148a-mediated suppression of low density-lipoprotein (LDL) receptor (LDLR) expression increases circulating LDLs that after ageing-dependent chemical modifications are scavenged by macrophages. miR-148a-mediated suppression of ATP binding cassette transporter 1 (ABCA1) attenuates reverse cholesterol transport and thus further promotes lipid accumulation in macrophages. miR-148a-mediated suppression of DNA methyltransferase 1 (DNMT1) enhances the expression of adipose differentiation-related protein (ADRP) further promoting foam cell formation
Several findings connect miRs to cardiovascular pathology. Neointimal formation is a common pathological phenotype in diverse cardiovascular diseases such as atherosclerosis and coronary heart disease. miR-21 has been related to vascular neointimal lesion formation, whereas downregulation of overexpressed miR-21 decreased neointima formation in rat carotid artery after angioplasty [209]. Upregulated miR-21 in endothelial cells suppressed apoptosis and increased eNOS phosphorylation and nitric oxide production [210]. In all these settings, miR-21 upregulation inhibited apoptosis and induced proliferation of vascular smooth muscle cells, contributing to the formation of neointima thickening in vivo [211]. Importantly, it has been demonstrated that milk exosomes are taken up by human vascular endothelial cells via endocytosis [36]. It is thus conceivable that systemic uptake of milk exosomes exerts adverse miR-21-mediated effects on vascular homeostasis.
Taken together, persistent dietary exposure to exosomal miR-148a and miR-21 derived from pasteurized, unfermented milk could exert atherogenic activities that may increase cardiovascular morbidity and mortality.
Bone remodeling, osteoporosis and fracture risk
It is common belief that consumption of cow’s milk, an abundant source of calcium, promotes bone strength and bone health, a major reason to recommend higher milk intake during pregnancy, infancy, adolescence and adulthood. However, greater milk consumption during teenage years was not associated with a lower risk of hip fracture in older adults [212]. Michaëlsson et al. [190] in Sweden reported that for every glass of milk in women no reduction was observed in fracture risk with higher milk consumption for any fracture or for hip fracture. However, high milk intake was associated with higher fracture incidence in Swedish women [190]. In contrast, among US men and women, 1 glass of milk per day was associated with an 8% lower risk of hip fracture [213]. Thus, the role of dairy foods and the quantity of milk intake for hip fracture still remains controversial. Recent evidence indicates that fermented milk in comparison to non-fermented milk exerts a protective effect on hip fracture rates and bone mineral density [214, 215]. Biver et al. [216] prospectively followed a cohort of 65-year-old healthy Swiss women and showed that age-related Ct bone loss was attenuated at non-bearing bone sites in fermented dairy product consumers, but not in milk consumers, independently of total energy, calcium, and protein intakes. According to a recent study, there was insufficient evidence to deduce the association between milk consumption and risk of hip fracture, which was however reduced by yogurt and cheese consumption [217].
Until today, no epidemiological study considered the heat processing (pasteurization vs UHT) of milk and did not report on the presence or absence of bioactive milk exosomes and their miR cargo [190, 212,213,214,215,216,217,218,219,220,221,222,223,224,225]. The presence of bioavailable milk exosomes is however of utmost importance to understand the differences in the biological function of pasteurized versus UHT milk on bone homeostasis.
Bone structure and homeostasis is controlled by MSCs. In the bone marrow, multipotent MSCs undergo differentiation into various anchorage-dependent cell types, including osteoblasts and adipocytes. At the cellular level, the MSC pool in the bone marrow niche shows a biased differentiation towards adipogenesis at the cost of osteogenesis [226]. This differentiation shift leads to decreased bone formation, contributing to the etiology of osteoporosis [226]. Since the identification of the v-MAF oncogene in an avian tumor virus, the MAF protein family has grown rapidly, forming a unique subclass of basic-leucine zipper transcription (bZIP) factors. MAF family members appear to play important roles in the regulation of MSC differentiation [227]. Nishikawa et al. [228] demonstrated that decreased expression of MAF in mouse MSCs, which regulated MSC bifurcation into osteoblasts and adipocytes by cooperating with the osteogenic transcription factor RUNX2 and inhibiting the expression of the adipogenic transcription factor PPARγ, impaired osteogenesis [228]. The crucial role of MAF in both osteogenesis and adipogenesis was underscored by in vivo observations of delayed bone formation in perinatal MAF(−/−) mice, and accelerated formation of fatty marrow associated with bone loss in aged MAF(±) mice.
MAF and MAFB are direct target genes of miR-148a [188, TargetScanHuman7.2]. Long-term exposure to milk exosome-derived miR-148a may thus favor adipogenesis in the bone on the expense of osteogenesis. Bone remodeling is a life-long process to maintain bone homeostasis. Its imbalance causes bone porosity and increases the risk of fracture. The balance is controlled by bone-forming osteoblasts and bone-resorbing osteoclasts interacting with blood-vessel-forming endothelial cells [229]. There is compelling evidence that exosomes and their miR cargo play a crucial role in bone remodeling [230,231,232,233,234,235,236]. Kelch et al. [234] recently reported that miR-148a and miR-21 are significantly upregulated in serum and osteoclasts of patients with osteoporosis. In accordance, increased levels of miR-148a and miR-21 have been detected in sera of type 1 diabetes patients (T1DM) versus non-diabetic subjects. In patients with T1DM, who exhibit reduced bone mineral density (BMD) associated with an increased risk of fractures, miR-148a expression showed an inverse correlation with BMD [237]. Remember that milk exosomal miRs have been shown to increase in PBMCs of milk consumers [46]. In addition, the uptake of dairy milk exosomes and their miRs by human macrophages has also been demonstrated [47]. Peripheral blood monocytes (PBMs) are an important source of osteoclast precursors and cytokines produced by PBMs have profound effects on osteoclast differentiation, activation, and apoptosis [238]. Receptor activator of nuclear factor kappaB ligand (RANKL) induces osteoclast formation from hematopoietic cells via regulation of various transcription factors. MAFB negatively regulates RANKL-induced osteoclast differentiation [239]. Intriguingly, miR-148a targets MAFB, a critical inhibitor of RANKL, thereby promoting the differentiation of monocytes to pre-osteoclasts [188]. miR-148a was reported to be dramatically upregulated during M-CSF + RANKL-induced osteoclastogenesis of CD14+ PBMCs [188]. miR-21, another exosomal signature miR of dairy milk, suppresses programmed cell death 4 (PDCD4) [240, 241], a critical inhibitor of c-Fos, which is important for the differentiation of pre-osteoclasts to osteoclasts [234]. Intriguingly, Oliveira et al. [232], demonstrated that dairy milk exosomes (100,000×g fraction) promoted osteoclast differentiation associated with an increased expression of c-Fos (Fig. 8).

Milk exosome and bone homeostasis. MiRs play a key role in the regulation of bone remodeling executed by bone-resorbing osteoclasts and bone-forming osteoblasts. Blood monocytes are a primary source of osteoclast precursor cells. Upregulation of receptor activator of nuclear factor B ligand (RANKL), V-Fos FBJ murine osteosarcoma viral oncogene homolog (c-Fos) and transforming growth factor-β (TGF-β) promote osteoclastogenesis. miR-148a via targeting V-maf musculoaponeurotic fibrosarcoma oncogene homolog B (MAFB) increases RANKL expression. miR-21 via targeting programmed cell death 4 (PDCD4) increases c-Fos activity. Notably, miR-148a, miR-21 and TGF-β are provided by dairy milk exosomes. Addition of commercial milk-derived exosomes to bone marrow-derived osteoclast precursor cells increased osteoclast formation. Overexpression of miR-148a triggers mesenchymal stem cells (MCS) to differentiate into adipocytes and attenuates osteoblast differentiation. Persistent intake of dairy milk exosomes may thus disturb the delicate balance of bone remodeling favoring osteoclastogenesis over osteoblastogenesis, a critical mechanism promoting osteoporosis and fracture risk
Oral administration of dairy milk-derived exosomes to female DBA/1 J mice during 7 weeks did not alter the tibia trabecular bone area but increased the number of osteocytes [242]. The highest dose of milk exosomes markedly increased the woven bone tissue. The exposure of MSCs to bovine milk exosomes during 21 days resulted in less mineralization but higher cell proliferation. Interestingly, milk exosomes reduced the collagen production, but enhanced the expression of genes characteristic for immature osteoblasts [242]. A kinetic study showed that milk exosomes upregulated many osteogenic genes within the first 4 days. However, the production of type I collagen and expression of its genes (COL1A1 and COL1A2) were markedly reduced at days 21 and 28. At day 28, milk exosomes again lead to higher proliferation, but mineralization was significantly increased. This was associated with increased expression of sclerostin, a marker for osteocytes, and reduced SPARC (osteonectin), which is associated to bone matrix formation and bone mineralization [242].
Earlier studies suggested that TGF-β increases osteoclast formation via action on osteoclast precursors [243]. TGF-β is a component of dairy milk exosomes [14] and has been shown to promote the differentiation of blood monocytes into osteoclasts [240]. Addition of TGF-β and dexamethasone to peripheral blood (PB) monocytes led to higher number of nuclei in multinuclear cells and increased expression of tartrate resistant acid phosphatase (TRACP) 5a and 5b, CR and NFATc1 in PB-derived osteoclasts depicting the higher osteoclastogenic potential and responsiveness to TGF-β and dexamethasone in PB monocytes [244].
There is further evidence that miR-148a inversely regulates adipocyte and osteoblast differentiation [233]. Supplementing miR-148a activity inhibited cell growth and induced stromal ST2 cells to differentiate into mature adipocytes. By contrast, supplementation of miR-148a blunted osteoblast differentiation. Lysine-specific demethylase 6b (Kdm6b), a recently identified regulator of osteoblast differentiation, was shown to be a direct target of miR-148a. Overexpression of Kdm6b attenuated miR-148a-mediated stimulation of adipogenic differentiation. Thus, miR-148a reciprocally regulates adipocyte and osteoblast differentiation through directly targeting Kdm6b [233].
Collectively, accumulating translational evidence supports the view that persistent uptake of pasteurized dairy milk and their bioactive exosomal miRs after the skeletal growth period activate osteoclastogenesis and impairs osteoblastogenesis, an unfavorable deviation for adults disturbing the appropriate balance for bone remodeling and explaining the association of obesity, diabetes mellitus and osteoporosis [245] (Fig. 8).
Parkinson’s disease
Epidemiological evidence supports a correlation between milk intake and risk of Parkinson’s disease (PD) [246,247,248,249,250,251]. A large meta-analysis reported a linear dose–response relationship for milk consumption and PD [249]. PD risk increased by 17% for every 200 g/day increment in milk intake [249]. Analyses were based on data from 2 large prospective cohort studies, the Nurses’ Health Study (n = 80,736) and the Health Professionals Follow-up Study (n = 48,610) confirmed an increased risk of PD associated with consumption of skim and low-fat milk [251]. Notably, there is no increased PD risk for fermented milk products such as yogurt [248, 249]. Neuron density in substantia nigra was lowest in nonsmoking decedents who consumed high amounts of milk (> 473 ml/day) [250]. After removing cases of PD and dementia with Lewy bodies, adjusted neuron density in all but the dorsomedial quadrant was 41.5% lower for milk intake > 473 ml/day versus intake that was less [250]. Thus, milk intake, but not fermented milk appears to exert neurodegenerative effects in PD.
Growing evidence indicates that exosomes are prominent mediators of neurodegenerative diseases. Exosomes of PD patients contain neurodegenerative disease-associated proteins such α-synuclein (α-syn) and facilitate their spread to the extracellular environment [252,253,254,255]. There is increasing evidence that exosome lipids promote α-syn aggregation [256]. Aggregation of exogenous α-syn was accelerated by exosomes irrespective of whether they were derived from control cells or cells overexpressing α-syn suggesting that the lipids in exosomes were sufficient for the catalytic effect to arise [252, 256]. As milk exosomes have been detected to cross the blood–brain barrier and accumulate in the brain [39], it is conceivable that dairy milk exosomes may promote α-syn aggregation and spreading.
There is recent interest in the regulatory role of exosomal miRs in the pathogenesis of PD [257], which according to recent concepts is related to neuroinflammation [258]. Prajapati et al. [259] demonstrated that TNFα is a potential regulator of miRs which may regulate mitochondrial functions and neuronal cell death, having important implication in pathogenesis of PD. TNFα induced the expression of miR-155 [259]. Recently, Thome et al. [260] found significant upregulation of miR-155 in an in vivo model of PD produced by adeno-associated-virus-mediated expression of α-syn. Using a mouse with a complete deletion of miR-155, they found that a loss of miR-155 reduced proinflammatory responses to α-syn and blocked α-syn-induced neurodegeneration. In primary microglia from miR-155(−/−) mice, they observed a markedly reduced inflammatory response to α-syn fibrils. Treatment of these microglia with a synthetic mimic of miR-155 restored the inflammatory response to α-syn fibrils. These results suggest that miR-155 plays a central role in the inflammatory response to α-syn in the brain and in α-syn-related neurodegeneration [260]. Importantly, miR-155 is one of the major immune regulatory miRs in cow’s milk that most likely invades into the brain [39].
Methylation of human α-syn gene SNCA intron 1 decreased its gene expression, while inhibition of DNA methylation activated SNCA expression. Methylation of SNCA intron 1 was reduced in DNA from sporadic PD patients’ substantia nigra, putamen, and cortex [261]. In fact, CpG demethylation in the promoter region of SNCA enhances α-syn expression and affects the pathogenesis of PD [262]. It has been shown that α-syn sequesters DNMT1 from the nucleus, which might be a novel mechanism for epigenetic alterations in Lewy body diseases [263]. Milk exosome-derived miR-148a may be another epigenetic mechanism, which via targeting DNMT1 may increase α-syn expression promoting PD pathogenesis [8] (Fig. 9).

Dairy milk exosomes and pathogenesis of Parkinson’s disease. Milk exosomes preferentially accumulate in the brain. Milk exosome-derived suppression of DNA methyltransferase 1 (DNMT1) reduces SNCA promoter methylation resulting in increased expression of α-synuclein. α-Synuclein promotes nuclear extrusion of DNMT1. Aggregates of α-synuclein induce neuroinflammation and increase tumor necrosis factor-α (TNF-α)-mediated upregulation of miR-155, which further enhances neuroinflammation. Milk exosomes via binding of α-synuclein to exosome membrane lipids may promote the spread of neurotoxic α-synuclein in the brain. Suppression of miR-155 attenuated α-synuclein-induced neuroinflammation in models of Parkinson’s disease
Collectively, milk exosomes and their miRs, which accumulate in the brain after milk consumption [39], might be the critical promoters involved in the initiation and progression of PD in humans consuming pasteurized cow’s milk.
Colorectal cancer
Two large meta-analyses came to the conclusion that milk consumption but not the consumption of fermented milk products has a protective effect on the development of colorectal cancer (CRC) [264, 265]. A recent systematic review and meta-analysis of cohort studies confirmed that an increase of 200 g/day of milk intake was associated with a decreased risk of CRC [266]. It has been shown that bacterial fermentation attacks the integrity of cow milk exosomes associated with a reduction of miR recovery [48, 49]. The protein content and size of bovine milk exosomes was significantly reduced in fermented cow’s milk associated with a substantial loss of miRs (miR-29b, miR-21) compared to unfermented raw milk [48]. Malignant epithelial cells of CRC exhibit a reduced expression of miR-148a [267,268,269,270], which increases the expression of DNMT1 that functions as a tumor promoter in CRC [271,272,273]. Intriguingly, Golan-Gerstl et al. [44] demonstrated that the incubation of CRC cells (Lim 1215) with human breastmilk exosomes increased the cellular content of miR-148a. In exosome incubation experiments with normal intestinal cells (CRL 1831), a significant decrease in DNMT1 was observed [44]. It is thus possible that milk exosome-mediated uptake of bovine miR-148a, which is identical with human miR-148a (mirbase.org), targets DNMT1 and thereby attenuates the action of this critical promoter of CRC initiation and progression. Increased expression of Rho-associated coiled coil-containing protein kinase 1 (ROCK1) also plays a key role in CRC pathogenesis [274, 275], which as well is a direct target of miR-148a [276].
Chronic inflammation triggers cellular events that can promote malignant transformation of cells and carcinogenesis. Chronic intestinal inflammation is a well-known stimulus of CRC cancerogenesis [277]. Several inflammatory mediators, especially TNFα, IL-6 and IL-10 have been shown to participate in both the initiation and progression of cancer including colitis-associated CRC [277]. miR-148a has a substantial impact on immune regulation and cancerogenesis [278]. miR-148a inhibits the production of cytokines including TNFα, IL-6, IL-12 and impairs innate response and antigen presentation of Toll-like receptor (TLR)-triggered dendritic cells by targeting calcium/calmodulin-dependent protein kinase IIα (CaMKIIα) [279]. Furthermore, TGF-β and miR-155, both components of milk exosomes, suppressed intestinal T cells and had a protective effect on the development of colitis [84].
Thus, mounting epidemiological and translational evidence indicates that milk exosomes via transfer of miR-148a and miR-155 may have a preventive effect on CRC cancerogenesis (Fig. 10).

Dairy milk exosomes and colorectal cancer prevention. DNA methyltransferase 1 (DNMT1) and Rho-associated coiled coil-containing protein kinase 1 (ROCK1) are overexpressed in colorectal cancer (CRC) cells. DNMT1 is regarded as a tumor promoter in CRC. Milk exosome uptake by CRC intestinal epithelial cells (IEC) increases intracellular levels of miR-148a, which suppresses DNMT1 and ROCK1, a potential mechanism explaining CRC prevention by consumption of pasteurized milk
Prostate cancer
Among European men, prostate cancer (PCa) is the most common cancer and the third leading cancer cause of death [280]. In 2018, there were an estimated 3.91 million new cases of cancer including 450,000 cases of PCa [280]. In the Physicians’ Health Study (n = 21,660) an association between whole milk intake and PCa-specific mortality among U.S. male physicians has been demonstrated [149]. A large meta-analysis of 11 population-based cohort studies involving 778,929 individuals reported a linear dose–response relationship between increase of whole milk intake and increase of PCa mortality risk [281]. A recent study confirmed that in comparison to men who consumed < 1 servings/day of whole milk, those who drank ≥ 3 servings/day had an increased hazard of PCa mortality [282]. In 996 African American and 1064 European American men diagnosed with PCa, a higher whole milk intake was associated with higher odds of high-aggressive PCa [283]. Pettersson et al. [284] showed that men with the highest versus lowest intake of whole milk were at an increased risk of PCa progression. A prospective study among 1334 men with non-metastatic PCa in the Cancer of the Prostate Strategic Urologic Research Endeavor (CaPSURE™) reported that whole milk consumption after PCa diagnosis was associated with increased risk of recurrence, particularly among very overweight or obese men [285]. Milk fat contains branched-chain fatty acids, whose metabolism is disturbed in PCa patients. A central role for fatty acid oxidation in supplying energy to the PCa cell is supported by the observation that the peroxisomal enzyme α-methylacyl-CoA racemase (AMACR), which facilitates the transformation of branched-chain fatty acids to a form suitable for β-oxidation, is highly overexpressed in PCa compared with normal prostate [286,287,288]. Branched-chain fatty acids in milk and dairy products markedly enhance AMACR expression in PCa cells in vitro [289].
In contrast to whole milk, total dairy and fermented milk products did not correlate with PCa risk [281,282,283,284,285]. Milk fat and calcium are obviously not the primary causative nutritional factors in whole milk for PCa initiation and progression, as milk fat is abundant in cheese and other fermented milk products. In contrast to fermented milk products, pasteurized milk transfers milk exosomes to the consumer.
Cancer cells communicate closely with the cells in their microenvironment, and this communication promotes malignancy via abnormal growth, invasion, drug resistance and metastasis. Increasing evidence illustrates that exosomes derived from tumor cells trigger tumor initiation, tumor cell growth and progression, metastasis, and drug resistance [290,291,292,293,294,295,296,297]. Exosome release by PCa cells modify the tumor microenviroment and play a key role in PCa initiation and progression [298,299,300,301,302,303].
miR-21, a signature miR of dairy milk, is overexpressed in PCa cells [304, 305], in blood serum [304, 306], PBMCs [307], serum exosomes [308] well as urinary exosomes of PCa patients [309, 310]. miR-21 is regarded as an oncomir, that inhibits pivotal tumor suppressor genes such as PTEN, p57kip2 (CDKN1C), PDCD4, MARCKS and others [240, 311,312,313,314]. Loss of function of the PTEN tumor suppressor, upregulating the phosphoinositide 3-kinase (PI3K)-AKT signaling network, is recognized as one of the most common driving events in PCa development [311]. Overexpression of miR-21 has been associated with chemo resistance and PCa progression [315, 316]. In accordance, an increase in miR-21 helps PCa cells to overcome androgen deprivation [317].
Epithelial–mesenchymal transition (EMT) plays a pivotal role in the conversion from benign to malignant phenotypes. There is accumulating evidence that exosomes via miR transfer prepare the pre-metastatic niche [318]. TGF-β signaling plays a further key role in EMT-mediated cancer progression [319]. PCa-derived exosomes, which in accordance with milk exosomes contain TGF-β [14], dominantly dictated a program of MSC differentiation generating myofibroblasts with functional properties consistent with cancer promotion [320]. Remarkably, it has been shown that human breast milk exosomes could promote EMT via TGF-β2 [15]. Commercial milk exosomes via transfer of TGF-β may further augment TGF-β-mediated EMT. Recent evidence has been provided that inhibition of DNMT1 induces EMT and the cancer stem cell (CSC) phenotype facilitating tumorigenesis in PCa cells [321]. Notably, miR-148a, miR-21 and miR-29b, major miRs of dairy milk exosomes, synergistically attenuate DNMT1 expression [68,69,70,71]. Furthermore, DNMT1 negatively controls the activity of androgen receptor signaling, which plays a key role in PCa pathogenesis [322].
miR-155 is upregulated in PCa tissues and cell lines and promotes cell proliferation by targeting annexin 7 [323]. SOCS1 functions as a tumor suppressor in PCa and its expression is reduced in PCa tissue [324]. miR-221-mediated suppression of SOCS1 enhanced cell proliferation and metastasis through in PCa [325]. Importantly, miR-155 as well targets SOCS1 [77]. miR-155 is an important exosomal immune regulatory miR of human and bovine milk [17, 45, 84]. miR-30d is another suppressor of SOCS1 [326]. miR-30d is overexpressed in PCa tissue and is inversely related to SOCS1 expression [326]. Remarkably, miR-30d is another signature miR of commercial cow’s milk [84].
Milk consumption has been linked with increased expression of fat mass and obesity-associated gene (FTO) via miR-148a-mediated suppression of DNMT1 [327]. Epidemiology studies show that FTO SNPs (including rs9939609, rs17817449, rs8050136, rs1477196, rs6499640, rs16953002, rs11075995, and rs1121980) are associated with increased FTO expression, overweight/obesity and increased risk of various types of cancers, including PCa [328]. FTO has recently been shown to increase the expression of C/EBPα and C/EBPβ [329, 330], which are upregulated in PCa tissue [331,332,333]. Remarkably, a C/EBP binding motif has been identified in the AMACR promotor [334]. Milk exosome-derived miR-148a via epigenetic enhancement of FTO-C/EBP-signaling may enhance AMACR expression in PCa allowing malignant cells to utilize branched-chain fatty acids as an alternative energy source for PCa growth and metastasis.
Bernichtein et al. [335] failed to observe any proliferative effects of “whole cow’s milk” in two mouse models of benign prostatic hyperplasia (probasin-Prl mice, Pb-Prl) or pre-cancerous PIN lesions (KIMAP mice). They reported decreased levels of the cell proliferation marker Ki-67. Notably, these investigators did not use “whole milk” as claimed on their paper’s title but instead used powdered milk re-suspended in water [335]. There is good reason to assume that cow milk powder in analogy to infant formula misses bioactive exosomal miRs [86]. In contrast, Tate et al. [336] observed a 30% increase in proliferation of LNCaP cells in culture after addition of commercial cow’s milk, which contains bioactive milk exosomes. The incidence of lactose intolerance, a natural protection for milk and milk exosome consumption, in PCa patients was significantly less than that in the control group [337].
Dairy milk exosomes via transfer of oncogenic miRs and TGF-β may promote growth and PCa progression in consumers of pasteurized whole milk but not fermented milk or milk protein preparations (Fig. 11).

Dairy milk exosomes and prostate tumorigenesis. Milk exosome-derived miR-148a, miR-29b and miR-21 suppress DNA methyltransferase 1 (DNMT1), a critical step in prostate cancer (PCa) epithelial–mesenchymal transition (EMT) and cancer stem cell (CSC) formation. miR-148a-mediated suppression of DNMT1 enhances the expression of fat mass and obesity-associated gene (FTO), which increases the expression of CCAAT enhancer element binding protein-β (C/EBPβ), a potential mechanism increasing the expression of α-methylacyl-CoA racemase (AMACR). DNMT1 is also a negative regulator of androgen receptor (AR) signaling. Milk exosomes provide miR-155 and transforming growth factor-β (TGFβ), which further induces miR-155, which inhibits suppressor of cytokine signaling 1 (SOCS1), a pivotal inhibitor of EMT. Milk-derived exosomal miR-21 increases the pool of a key oncogenic miR, which suppresses key checkpoint regulators of cell cycle progression and apoptotic signaling including phosphatase and tensin homolog (PTEN), cyclin-dependent kinase inhibitor 1C (CDKN1C, p57kip2), programmed cell death 4 (PDCD4), myristolylated alanine-rich protein kinase C substrate (MARCKS) and others
Breast cancer
Breast cancer (BC) is the most common cancer in women in industrialized countries. In Europe, 523,000 cases of female BC have been estimated in 2018 [280]. A prospective study of 25,892 Norwegian women reported that consumers of 750 ml or more of full-fat milk daily had a relative risk of 2.91 compared with those who consumed < 150 ml [338]. Wang et al. [339] surveyed risk factors for BC in women (n = 122,058) residing in urban and rural areas of eastern China. Among women residing in rural areas, obesity and a high intake of milk were identified as risk factors for BC. A case–control study in Mexico (97 BC patients, 104 controls) reported that high milk consumption increased BC risk by 7.2 times, whereas the consumption of meat was not significantly associated with BC risk [340]. According to a case–control study (n = 333) in Uruguay, high intakes of whole milk was associated with significant increased risk of BC, whereas fermented milk products were associated with significant decreased risk [341]. In a large Swedish cohort (n = 22,788), people with lactose intolerance, characterized by low consumption of milk, had decreased risks of BC [342]. In contrast, an older pooled analysis of cohort studies (n = 351,041) [343] found no significant associations between intake of dairy products and risk of BC. Notably, at present no epidemiological study clearly compared the effect of whole milk versus fermented milk products and there are missing data on the type of heat processing of milk in all epidemiological studies.
Although, epidemiological correlations for whole milk consumption and BC are less established than those for whole milk intake and PCa, tumor-derived exosomes as well play a key role in tumor initiation and progression in BC [344,345,346,347,348,349]. The widespread post-transcriptional regulatory role of miRs is of recent interest in estrogen receptor (ER)-positive BC, comprising about 65%–70% of BCs [350]. Estrogen/ERα activation can modulate miR expression, which may contribute to ER+ breast carcinogenesis [350]. Estradiol (E2) treatment of BC MCF7 cells doubled the expression levels of miR-148a and miR-21 [351]. An ER binding site has been demonstrated on the MIR21 gene [352]. E2 induced miR-148a in MCF-7 and MDA-MB-231 cells [353]. miR-21 is overexpressed in BC compared with normal breast tissue and has been associated with advanced stage, lymph node positivity, and reduced survival time [354, 355]. miR-21 is a major miR component of exosomes released by cancer-associated fibroblasts and cancer-associated adipocytes [356, 357], which promote tumor progression [358, 359]. In BC patients, increased miR-21 in the systemic circulation exists either freely or in exosomes [360]. Notably, in postmenopausal women 6 weeks of tamoxifen treatment decreased miR-21 levels suggesting that this miR may be important for BC tumorigenesis [361]. Circulating levels of miR-21 are significantly higher in plasma samples of BC patients, when compared healthy controls [360, 361]. miR-21 is even regarded as a marker of BC exosomes [362] and was found to be selectively enriched in human BC exosomes in the plasma of patients with BC [362,363,364]. A further increase of exosomal miR-21 via milk intake is apparently not suitable for patients with BC, neither the transfer of milk-derived exosomal miR-148a. In BC cell line MCF7, a miR-148a mimic increased estrogen receptor-α (ERα) expression, whereas a miR-148a inhibitor decreased ERα expression [365]. It has been shown that miR-148a regulates ERα expression through DNMT1-mediated DNA methylation in BC cells [366]. In contrast, it has been reported that miR-148a targets B-cell lymphoma 2 (BCL-2), which is frequently upregulated in BC [367]. Nuclear receptor NR4A1 (Nur77) promotes BC invasion and metastasis by activating TGF-β signaling [368, 369]. NR4A1 binding induces a BCL-2 conformational change that exposes its BH3 domain, resulting in conversion of BCL-2 from an anti-apoptotic to a pro-apoptotic protein [370].
BRCA1, a well-known tumor suppressor, abrogates the repression of miR-155, a bona fide oncomir [371]. miR-155 is overexpressed in BC tissue and accelerates the growth of tumor cell lines in vivo and induces tamoxifen resistance [371, 372]. In BC cells, FoxP3 induced miR-155 through transcriptional repression of BRCA1. Notably, miR-155 is known to induce FoxP3 expression [373]. For patients with early stage or localized BC, there were high levels of miR-155 in both plasma and blood cells [373]. Santos et al. [374] recently reported that exosomes enriched in miR-155 added to BC cells induced chemo resistance and promoted EMT. Ectopic expression of miR-155 significantly promoted the proliferation of BC cells, the formation of soft agar foci in vitro, and the development of tumors in nude mice [374]. In BC cells, RNA interference silencing of SOCS1 recapitulates the oncogenic effects of miR-155, whereas restoration of SOCS1 expression attenuated the tumor-promoting function of miR-155, suggesting that miR-155 exerts its oncogenic role by negatively regulating SOCS1 [375]. Thus, exosomal transfer of miR-155, a miR component of colostrum and mature cow’s milk [86, 376], may promote BC tumorigenesis.
TGF-β signaling features a growth inhibitory effect at an early stage but aggressive oncogenic activity at the advanced malignant state [377,378,379]. Recent efforts in BC therapy are directed against growth factor pathway including TGF-β signaling in BC [379]. Notably, TGF-β1 has been shown to promote the expression of miR-155 [380]. TGF-β2 is significantly upregulated in breast milk exosomes during weaning/early involution. Breast milk exosomes containing high levels of TGF-β2 induced changes in both benign and malignant breast epithelial cells, consistent with the development and progression of BC, suggesting a role for high TGF-β2-expressing breast milk exosomes in influencing BC risk [14]. BC exosomes contain TGF-β2, which suppresses T cell proliferation, a critical pathway used by BC cells to escape immune surveillance [381]. In addition, exosomes from BC cells via TGF-β upregulation converted adipose tissue-derived MSCs into myofibroblast-like cells [382].
Collectively, BC-derived exosomes and dairy milk-derived exosomes both contain and transfer miR-21, miR-155 and TGFβ2, which may exert synergistic effects in breast cancerogenesis (Fig. 12).

Dairy milk exosomes and breast cancerogenesis. Milk exosomes via transfer of miR-148a and miR-148a-mediated suppression of DNA methyltransferase 1 (DNMT1) enhances the expression of estrogen receptor-α (ERα). ERα promotes des expression of miR-21, which targets critical genes involved in PI3K-AKT signaling and cell cycle control. Exosome-derived transforming growth factor-β (TGF-β) induces the expression of miR-155. miR-155 enhances the expression of FoxP3, a critical inhibitor of the tumor suppressor breast cancer 1 gene (BRCA1). Downregulation of BRCA1 further enhances the expression of miR-155, which is a pivotal inhibitor of suppressor of cytokine signaling 1 (SOCS1) finally promoting epithelial–mesenchymal transition (EMT). Dairy milk exosomes thus contribute to BC tumorigenesis via enhancing key oncogenic components involved in the pathogenesis of BC
Hepatocellular carcinoma
In the European Prospective Investigation into Cancer and Nutrition cohort including 477,206 participants showed a significant positive hepatocellular carcinoma (HCC) risk for the consumption of milk and cheese but not yogurt [383]. Increased expression of miR-148a has been reported in hepatitis B virus-induced HCC resulting from HBx antigen-induced upregulation of von Willebrand factor C and EGF domain-containig protein [384]. Hepatitis C virus-induced upregulation of miR-155 has been shown to promote hepatocarcinogenesis [385]. miR-21 expression was significantly upregulated in HCC tissues relative to nontumor livers [386]. Exosome-associated miR-21 is markedly elevated in serum of patients with HCC [387]. It has been reported that miR-155 is linked to the recurrence and prognosis of HCC following liver transplantation [381]. In the HCC cell line Huh-7, miR-155 is overexpressed and exhibited altered levels of expression of certain cellular adhesion molecules related to EMT [381]. Furthermore, TGF-β1 upregulated the expression of miR-155 in HCC cells in vitro, which led to the conclusion that increased levels of miR-155 in HCC cells, possibly due to stimulation by TGF-β1, accelerate EMT in the liver. Notably, the liver is a major target of bovine milk exosomes [39]. Recent evidence indicates that miR-155 suppresses p53-induced nuclear protein 1 (TP53INP1), a critical step that is involved in liver cancer stem cell acquisition and self-renewal [388]. TP53INP1 is a p53-inducible gene that regulates p53-dependent apoptosis, downregulates the expression of SPARC and is repressed by miR-155 [389,390,391]. Recent findings indicate that loss of SOCS1-dependent control over EMT may contribute to MET-mediated migration, invasion and metastatic growth of HCC [392].
Translational evidence indicates that milk-derived exosomes via transfer of onocogenic miR-148a, miR-21, miR-155 and TGF-β may promote the development of HCC (Fig. 13).

Dairy milk exosomes and hepatocellular carcinoma. After oral administration milk exosomes accumulate preferentially in the liver and may increase the hepatocellular levels of miR-148a, miR-21 and miR-155, which are upregulated in hepatocellular carcinoma (HCC). In hepatitis B virus (HCB)-associated HCC overexpression of miR-148a may be further increased by milk exosome-derived miR-148a. Milk exosome-derived transforming growth factor-β (TGF-β) may further increase the expression of miR-155, which downregulates suppressor of cytokine signaling 1 (SOCS1), thereby enhancing epithelial–mesenchymal transition (EMT). MiR-155-mediated suppression of p53-induced nuclear protein 1 (TP53INP1) promotes cancer stem cell (CSC) proliferation in the liver. Milk exosomes may thus increase the risk for HCC. Thus, milk exosomes may augment the tumorigenic effects of hepatitis B (HCB) and hepatitis C virus (HCC)-induced upregulation of miR-148a and miR-155, respectively
Diffuse large B-cell lymphoma
A recent meta-analysis investigated the relation between dairy product consumption and Non-Hodgkin lymphoma (NHL). Milk has been identified to increase the risk of diffuse large B-cell lymphoma (DLBCL) [393]. The dose–response analysis suggested that the risk of NHL increased by 6% for each 200 g/day increment of milk consumption but not yogurt [393].
DLBCL have 10- to 30-fold higher copy numbers of miR-155 than do normal circulating B cells [394]. Epstein Barr virus (EBV)-positive DLBCL is an entity included in the 2016 WHO classification of lymphoid neoplasms [395]. It has been demonstrated that the expression of miR-155 is induced by EBV [396]. EBV acts on noninfected macrophages in the tumor through exosome secretion and thereby augments lymphoma development [397]. Plasma miR-155 expression was significantly upregulated in DLBCL patients compared to healthy individuals [398]. DLBCL cases with an elevated level of miR-155 had shorter overall survival than those with a lower miR-155 expression [398]. Intriguingly, SOCS1, the target of miR-155 and miR-30d, is frequently mutated in patients with DLBCL [399,400,401]. One-fourth of DLBCL and follicular lymphomas carried SOCS1 mutations, which were preferentially targeted to SHM hotspot motifs and frequently inactivating mutations [401]. Furthermore, increased serum expression levels of miR-21 have been detected in patients with DLBCL associated with negative prognostic outcome [402]. Exosomal transfer of milk-dervived miR-155 and miR-21 to circulating B-cells may initiate or promote DLBCL progression (Fig. 14).

Dairy milk exosomes and tumorigenesis of diffuse large B-cell lymphoma. miR-155 levels are significantly upregulated in diffuse large B-cell lymphoma (DLBCL). Epstein Barr Virus (EBV) infection in DLBCL induces miR-155 expression, which attenuates the expression of suppressor of cytokine signaling 1 (SOCS1). Loss-of-function mutations of SOCS1 (SOCS1*) have also been found in DLBCL further reducing SOCS1. Milk exosomes via transfer of miR-155 and miR-155-inducing transforming growth factor-β (TGF-β) may further promote tumorigenesis of DLBCL
MIRNA148A and dairy cow lactation performance
Epigenetic regulation of bovine MECs plays a crucial role in the biosynthesis of milk lipid and protein components. miR-148a targets key mediators involved in triacylglycerol and cholesterol homeostasis such as ABCA1, LDLR and CPT1A [403]. All members of the miR-148/152 family (miR-148a, miR-148b, and miR-152) share identical seed sequences [278, 404]. DNMT1 is a direct target of both miR-148a and miR-152 [68, 405]. The expression of miR-152 significantly increased during lactation in MECs of dairy cows producing high quality milk compared to lower miR-152 levels in cows producing low quality milk [406]. Forced expression of miR-152 in dairy cow MECs resulted in a marked reduction of DNMT1 at both the mRNA and protein levels [406]. In goat MECs, miR-148a induced milk triacylglycerol synthesis [407]. miR-148a expression can regulate PPARA and promoted triacylglycerol (TAG) synthesis while the knockdown of miR-148a impaired TAG synthesis in goat MEC. In addition, miR-148a cooperates with miR-17-5p to regulate fatty acid metabolism by repressing PPARGC1A and PPARA in goat MECs. Lactogenic hormones such as prolactin induce cellular and extracellular miR-148a expression in bovine MECs [408]. miR-148a belongs to the most abundantly expressed miRs of bovine milk since it accounts for more than 10% of the read counts in each stage of dairy cow lactation [409]. Directional selection of miR regulatory variants was important in the domestication and subsequent selection that gave rise to modern taurine cattle. The MIR148A gene has been identified as a candidate of domestication genes of modern cattle [410]. Co-expression network and pathway analyses identified bovine MIR148A as a major determinant enhancing milk yield [411]. Exaggerated miR-148a expression resulting in decreased DNMT1 expression is critical epigenetic change that induces lactation performance. Furthermore, persistent pregnancy of cows via increased E2 production may enhance E2-mediated miR-148a- and miR-21 expression in bovine MECs [333,334,335], thereby increasing the exosomal content of miR-148a and miR-21 (Fig. 15). In fact, increased miR-21 levels have been detected in skim milk of pregnant versus cyclic cows [354]. Thus, genetic selection of high performance dairy cows with enhanced miR-148a expression and pregnancy-dependent E2 production may be associated with an enrichment of miR-148a and miR-21 in milk exosomes enhancing the exposure of the human consumer of pasteurized milk to oncogenic miRs.

Hypothesized increase in dairy milk exosomal miR content by genetic selection and persistent pregnancy of dairy cows. MIR148A is a domestication gene of dairy cows increasing milk yield. Estrogens, which are upregulated in pregnant dairy cows, stimulated the expression of miR-148a and miR-21, which attenuate the expression of DNA methyltransferase 1 (DNMT1). Various lipogenesis-inducing genes such as sterol regulatory element binding factor 1 (SREBF1), peroxisome proliferator-activated receptor-γ (PPARG) and estrogen receptor-α (ERα). Procedures that increase dairy cow lactation performance are associated with an upregulation of lactation-promoting miRs, which may enter the human food chain in higher amounts
Conclusion
Milk exosomes execute an evolutionary program under control of the lactation genome. They assist in the regulation of growth, tissue maturation, metabolic and immunological programming of the newborn infant. Obviously, this ancient exosome system has developed for maternal-neonatal communication and operates exclusively during the postnatal period of mammals except Neolithic humans who are persistently exposed to this system. The transfer of exosome-protected bovine milk miRs to human consumers has been considered to be one of the most important miR-based inter-species epigenetic communication related to the pathogenesis of human diseases [7,8,9,10,11,12, 412]. The restriction of milk exosomes to the postnatal period has been secured by physiological lactose intolerance that appears after weaning in all mammals except lactase (LCT)-mutated humans that could persistently abuse this postnatal epigenetic doping system. Unfortunately, the beneficial growth promoting and tissue-supporting effects of milk exosomes during early infancy may turn into obesigenic, diabetogenic, osteoclastogenic, cancerogenic and neurodegenerative signaling in adulthood. Dairy milk-derived exosomes should thus be regarded as potential pathogens that have spread unnoticeably into the human food chain. Technical progress with the introduction of pasteurization and refrigeration technology selected and preserved bovine milk exosomes, because the thermic conditions of pasteurization are not sufficient to eliminate bioactive milk exosomes [42,43,44]. Furthermore, pasteurization reduces exosome-degrading lactobacteria. Bacterial fermentation of milk attacks milk exosome proteins, reduces their size and miR content [48, 49].
As the milk exosome system is an archaic and highly conserved signaling system of all mammals [44], miRs exhibit a high degree of sequence homology [13, 44]. Remarkably, seed sequences of human and bovine miR-148a, miR-21, miR-29b, and miR-155 are identical (mirbase.org). Manca et al. [39] provided compelling evidence that bovine milk exosomes of pasteurized commercial milk reach the systemic circulation and distribute in tissues of different species. Efforts of dairy research to increase lactation performance and milk yield of dairy cows may have increased bovine miR-148a and miR-21 expression and their transfer to human milk consumer via milk exosomes [9].
Translational evidence supports the view that dairy milk exosomes are potential pathogens for human health (Table 1). This view is in accordance with epidemiological evidence showing adverse health effects for unfermented milk but no adverse or even beneficial health effects for fermented milk and milk products. Based on translational evidence, we conclude that milk exosomes should not reach the human food chain. Pasteurization of milk is an inappropriate method to prevent the spread of milk exosomes to the human milk consumer. In this regard, UHT is much more effective [43]. Other choices under recent investigation are microwave treatment or ultra-sonication of milk exosomes [39, 413].
Due to their low antigenicity, excellent bioavailability in many tissues and easy crossing of tissue boundaries such as the intestinal and blood–brain barrier, pharmacology became highly interested in bovine milk exosomes as therapeutic delivery systems of small interfering RNAs, drugs and phytochemicals [38, 414]. However, these new milk-exosome-based therapeutic options are a double-edged sword, because milk exosomes function as a Trojan horse not only transferring the new compound of interest but also the intrinsic exosome cargo such as oncogenic miRs and EMT-promoting TGF-β. Furthermore, it is of critical concern that exosomes may transfer viral RNA or DNA [415]. Novel replication-competent circular DNA molecules with potential proliferative activity have been detected in commercial milk of dairy cows [416,417,418,419,420], that via milk exosome transfer may reach distant tissue including the brain. Exosomes and their cargo are also involved in the spread of neurotoxic proteins such as α-syn, amyloid-β and prions [392, 393, 421,422,423,424,425]. Therefore, dairy milk-derived exosomes, although representing an easily accessible and abundant source of exosomes, are apparently not suitable for the treatment of human diseases. Milk exosomes meet the definition of bioactive food compounds and have an impact on human metabolism and gene regulation [426]. Before employing milk exosomes as drug delivery systems or supplements of infant formula, their unique intrinsic roles in the transmission of exosomal miRs and their potential ability to spread viral or neurotoxic pathogens require much more attention and most careful studies before introducing milk exosomes as carrier systems for the treatment of human diseases. Dairy milk exosomes should be regarded as potential new pathogens promoting western diseases.
Abbreviations
- ABCA1:
-
ATP-binding cassette, subfamily A, member 1
- ADRP:
-
adipose differentiation-related protein
- ALIX:
-
apoptosis-linked gene 2-interacting protein X
- α-syn:
-
alpha-synuclein
- AMACR:
-
alpha-methylacyl-CoA racemase
- BAT:
-
brown adipose tissue
- BC:
-
breast cancer
- BCAA:
-
branched-chain amino acid
- BCKD:
-
branched-chain alpha-ketoacid dehydrogenase
- BMD:
-
bone mineral density
- BMP:
-
bone morphogenetic protein
- BRCA1:
-
breast cancer 1 gene
- CaMKIIα:
-
calcium/calmodulin-dependent protein kinase II alpha
- CCK:
-
cholecystokinin
- CCK1R:
-
CCK receptor 1 = CCK A receptor
- CCK2R:
-
CCK receptor 2 = CCK B receptor
- CD63:
-
melanoma-associated antigen MLA1
- CD81:
-
target of antiproliferative antibody (TAPA1)
- CDKN1C:
-
cyclin-dependent kinase inhibitor 1C (p57kip2)
- CE:
-
cholesteryl ester
- C/EBP:
-
CCAAT enhancer element binding protein
- c-Fos:
-
V-Fos FBJ murine osteosarcoma viral oncogene homolog
- CRC:
-
colorectal cancer
- CSN1S1:
-
casein alpha s1
- DBT:
-
dihydrolipoamide branched-chain acyltransferase
- DLBCL:
-
diffuse large B-cell lymphoma
- DNMT:
-
DNA methyltransferase
- DOHaD:
-
developmental origins of health and disease
- EBV:
-
Epstein Barr virus
- ECM:
-
extracellular matrix
- ElF5:
-
E74-like factor 5
- EMT:
-
epithelial–mesenchymal transition
- ER:
-
estrogen receptor
- ERK:
-
mitogen-activated protein kinase 4
- EV:
-
extracellular vesicle
- FoxP3:
-
forkhead box P3 (scurfin)
- GLUT:
-
glucose transporter
- hASC:
-
human adipose tissue-derived mesenchymal stem cell
- HCC:
-
hepatocellular carcinoma
- HSP70:
-
heat shock protein 70
- HFD:
-
high-fat diet
- IL:
-
interleukin
- IFN:
-
interferon
- IRS-1:
-
insulin receptor substrate 1
- ITK:
-
IL-2 inducible T-cell kinase
- Kdm6b:
-
lysine-specific demethylase 6b
- LDL:
-
low density-lipoprotein
- LDLR:
-
low density lipoptrotein receptor
- MAFB:
-
V-Maf musculoaponeurotic fibrosarcoma oncogene homolog B
- MAPK:
-
mitogen-activated protein kinase 1
- MARCKS:
-
myristolylated alanine-rich protein kinase C substrate
- MEC:
-
mammary epithelial cell
- MFG:
-
milk fat globule
- miR:
-
micro ribonucleic acid
- MFGM:
-
milk fat globule membrane
- MSC:
-
mesenchymal stem cell
- mTORC1:
-
mechanistic target of rapamycin complex 1
- MV:
-
microvesicles
- MVB:
-
multivesicular body
- NPY:
-
neuropeptide Y
- NHL:
-
non-Hodgkin lymphoma
- PBM:
-
peripheral blood monocytes
- PBMC:
-
peripheral blood mononuclear cell
- PD:
-
Parkinson’s disease
- PCa:
-
prostate cancer
- PPAR:
-
peroxisome proliferator-activated receptor
- PPARGC1A:
-
peroxisome proliferator-activated receptor-gamma, coactivator 1, alpha
- RANKL:
-
receptor activator of nuclear factor κB ligand
- RASGRP1:
-
ras guanyl nucleotide-releasing protein 1
- rhPCR:
-
RNase H2-dependent polymerase chain reaction
- ROCK1:
-
Rho-associated coiled coil-containing protein kinase 1
- ROS:
-
reactive oxygen species
- SOCS:
-
suppressor of cytokine signaling
- SPARC:
-
secreted protein acidic and rich in cysteine
- SPRY2:
-
sprouty 2
- SREBF1:
-
sterol regulatory element-binding transcription factor 1
- STAT:
-
signal transducer and activator of transcription
- TAG:
-
triacylglycerol
- TC:
-
total cholesterol
- T2DM:
-
type 2 diabetes mellitus
- TGF-β:
-
transforming growth factor-beta
- TNF:
-
tumor necrosis factor
- TLR:
-
toll-like receptor
- Treg:
-
regulatory T cell
- TSDR:
-
treg-specific demethylation region
- TSG101:
-
susceptibility gene-101
- UHT:
-
ultra-heat treatment
- UTR:
-
untranslated region
- WAT:
-
white adipose tissue
- Wnt:
-
wingless-type MMTV integration site family members
References
Yáñez-Mó M, Siljander PR, Andreu Z, Zavec AB, Borràs FE, Buzas EI, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066.
Iraci N, Leonardi T, Gessler F, Vega B, Pluchino S. Focus on extracellular vesicles: physiological role and signalling properties of extracellular membrane vesicles. Int J Mol Sci. 2016;17:171.
Abels ER, Breakefield XO. Introduction to extracellular vesicles: biogenesis, RNA cargo selection, content, release, and uptake. Cell Mol Neurobiol. 2016;36:301–12.
Zhang J, Li S, Li L, Li M, Guo C, Yao J, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinform. 2015;13:17–24.
Alsaweed M, Hartmann PE, Geddes DT, Kakulas F. MicroRNAs in breastmilk and the lactating breast: potential immunoprotectors and developmental regulators for the infant and the mother. Int J Environ Res Public Health. 2015;12:13981–4020.
Cui J, Zhou B, Ross SA, Zempleni J. Nutrition, microRNAs, and human health. Adv Nutr. 2017;8:105–12.
Zempleni J, Aguilar-Lozano A, Sadri M, Sukreet S, Manca S, Wu D, et al. Biological activities of extracellular vesicles and their cargos from bovine and human milk in humans and implications for infants. J Nutr. 2017;147:3–10.
Melnik BC, Schmitz G. Milk’s role as an epigenetic regulator in health and disease. Diseases. 2017;5(1):E12.
Melnik BC, Schmitz G. MicroRNAs: milk’s epigenetic regulators. Best Pract Res Clin Endocrinol Metab. 2017;31:427–42.
Lawson C, Kovacs D, Finding E, Ulfelder E, Luis-Fuentes V. Extracellular vesicles: evolutionarily conserved mediators of intercellular communication. Yale J Biol Med. 2017;90:481–91.
Melnik BC, John SM, Schmitz G. Milk is not just food but most likely a genetic transfection system activating mTORC1 signaling for postnatal growth. Nutr J. 2013;12:103.
Melnik BC, Kakulas F, Geddes DT, Hartmann PE, John SM, Carrera-Bastos P, et al. Milk miRNAs: simple nutrients or systemic functional regulators? Nutr Metab (Lond). 2016;13:42.
van Herwijnen MJC, Driedonks TAP, Snoek BL, Kroon AMT, Kleinjan M, Jorritsma R, et al. Abundantly present miRNAs in milk-derived extracellular vesicles are conserved between mammals. Front Nutr. 2018;5:81.
Pieters BC, Arntz OJ, Bennink MB, Broeren MG, van Caam AP, Koenders MI, et al. Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-β. PLoS ONE. 2015;10:e0121123.
Qin W, Tsukasaki Y, Dasgupta S, Mukhopadhyay N, Ikebe M, Sauter ER. Exosomes in human breast milk promote EMT. Clin Cancer Res. 2016;22:4517–24.
Admyre C, Johansson SM, Qazi KR, Filén JJ, Lahesmaa R, Norman M, et al. Exosomes with immune modulatory features are present in human breast milk. J Immunol. 2007;179:1969–78.
Zhou Q, Li M, Wang X, Li Q, Wang T, Zhu Q, et al. Immune-related microRNAs are abundant in breast milk exosomes. Int J Biol Sci. 2012;8:118–23.
Melnik BC, John SM, Schmitz G. Milk: an exosomal microRNA transmitter promoting thymic regulatory T cell maturation preventing the development of atopy? J Transl Med. 2014;12:43.
Melnik BC, John SM, Carrera-Bastos P, Schmitz G. Milk: a postnatal imprinting system stabilizing FoxP3 expression and regulatory T cell differentiation. Clin Transl Allergy. 2016;6:18.
de la Torre Gomez C, Goreham RV, Bech Serra JJ, Nann T, Kussmann M. “Exosomics”- A review of biophysics, biology and biochemistry of exosomes with a focus on human breast milk. Front Genet. 2018;9:92.
Liao Y, Du X, Li J, Lönnerdal B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol Nutr Food Res. 2017;61:11.
Sitarik AR, Bobbitt KR, Havstad SL, Fujimura KE, Levin AM, Zoratti EM, et al. Breast milk transforming growth factor β is associated with neonatal gut microbial composition. J Pediatr Gastroenterol Nutr. 2017;65:e60–7.
Kahn S, Liao Y, Du X, Xu W, Li J, Lönnerdal B. Exosomal microRNAs in milk from mothers delivering preterm infants survive in vitro digestion and are taken up by human intestinal cells. Mol Nutr Food Res. 2018;62:e1701050.
Martin C, Patel M, Williams S, Arora H, Sims B. Human breast milk-derived exosomes attenuate cell death in intestinal epithelial cells. Innate Immun. 2018;24:278–84.
Le Doare K, Holder B, Bassett A, Pannaraj PS. Mother’s milk: a purposeful contribution to the development of the infant microbiota and immunity. Front Immunol. 2018;9:361.
Sakaguchi K, Koyanagi A, Kamachi F, Harauma A, Chiba A, Hisata K, et al. Breast- feeding regulates immune system development via transforming growth factor-β in mice pups. Pediatr Int. 2018;60:224–31.
Reinhardt TA, Lippolis JD, Nonnecke BJ, Sacco RE. Bovine milk exosome proteome. J Proteomics. 2012;75:1486–92.
Benmoussa A, Lee CH, Laffont B, Savard P, Laugier J, Boilard E, et al. Commercial dairy cow microRNAs resist digestion under simulated gastrointestinal tract conditions. J Nutr. 2016;146:2206–15.
Benmoussa A, Ly S, Shan ST, Laugier J, Boilard E, Gilbert C, et al. A subset of extracellular vesicles carries the bulk of microRNAs in commercial dairy cow’s milk. J Extracell Vesicles. 2017;6:1401897.
Benmoussa A, Gotti C, Bourassa S, Gilbert C, Provost P. Identification of protein markers for extracellular vesicle (EV) subsets in cow’s milk. J Proteom. 2018;3919(18):30322. https://doi.org/10.1016/j.jprot.2018.08.010 (Epub ahead of print).
Hata T, Murakami K, Nakatani H, Yamamoto Y, Matsuda T, Aoki N. Isolation of bovine milk-derived microvesicles carrying mRNAs and microRNAs. Biochem Biophys Res Commun. 2010;396:528–33.
Izumi H, Kosaka N, Shimizu T, Sekine K, Ochiya T, Takase M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J Dairy Sci. 2012;95:4831–41.
Koh YQ, Peiris HN, Vaswani K, Meier S, Burke CR, Macdonald KA, et al. Characterization of exosomes from body fluids of dairy cows. J Anim Sci. 2017;95:3893–904.
Cai M, He H, Jia X, Chen S, Wang J, Shi Y, et al. Genome-wide microRNA profiling of bovine milk-derived exosomes infected with Staphylococcus aureus. Cell Stress Chaperones. 2018;23:663–72.
Wolf T, Baier SR, Zempleni J. The intestinal transport of bovine milk exosomes is mediated by endocytosis in human colon carcinoma Caco-2 cells and rat small intestinal IEC-6 cells. J Nutr. 2015;145:2201–6.
Kusuma RJ, Manca S, Friemel T, Sukreet S, Nguyen C, Zempleni J. Human vascular endothelial cells transport foreign exosomes from cow’s milk by endocytosis. Am J Physiol Cell Physiol. 2016;310:C800–7.
Arntz OJ, Pieters BC, Oliveira MC, Broeren MG, Bennink MB, de Vries M, et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol Nutr Food Res. 2015;59:1701–12.
Munagala R, Aqil F, Jeyabalan J, Gupta RC. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016;371:48–61.
Manca S, Upadhyaya B, Mutai E, Desaulniers AT, Cederberg RA, White BR, et al. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci Rep. 2018;8:11321.
Sterzenbach U, Putz U, Low LH, Silke J, Tan SS, Howitt J. Engineered exosomes as vehicles for biologically active proteins. Mol Ther. 2017;25:1269–78.
Wang L, Sadri M, Giraud D, Zempleni J. RNase H2-dependent polymerase chain reaction and elimination of confounders in sample collection, storage, and analysis strengthen evidence that microRNAs in bovine milk are bioavailable in humans. J Nutr. 2018;148:153–9.
Howard KM, Jati Kusuma R, Baier SR, Friemel T, Markham L, Vanamala J, et al. Loss of miRNAs during processing and storage of cow’s (Bos taurus) milk. J Agric Food Chem. 2015;63:588–92.
Kirchner B, Pfaffl MW, Dumpler J, von Mutius E, Ege MJ. microRNA in native and processed cow’s milk and its implication for the farm milk effect on asthma. J Allergy Clin Immunol. 2016;137:1893–5.
Golan-Gerstl R, Elbaum Shiff Y, Moshayoff V, Schecter D, Leshkowitz D, Reif S. Characterization and biological function of milk-derived miRNAs. Mol Nutr Food Res. 2017;61(10):1700009.
Oh S, Park MR, Son SJ, Kim Y. Comparison of total RNA isolation methods for analysis of immune-related microRNAs in market milks. Korean J Food Sci Anim Resour. 2015;35:459–65.
Baier SR, Nguyen C, Xie F, Wood JR, Zempleni J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J Nutr. 2014;144:1495–500.
Izumi H, Tsuda M, Sato Y, Kosaka N, Ochiya T, Iwamoto H, et al. Bovine milk exosomes contain microRNA and mRNA and are taken up by human macrophages. J Dairy Sci. 2015;98:2920–33.
Yu S, Zhao Z, Sun L, Li P. Fermentation results in quantitative changes in milk- derived exosomes and different effects on cell growth and survival. J Agric Food Chem. 2017;65:1220–8.
Yu S, Zhao Z, Xu X, Li M, Li P. Characterization of three different types of extracellular vesicles and their impact on bacterial growth. Food Chem. 2019;272:372–8.
Melnik BC, Kakulas F. Milk exosomes and microRNAs: potential epigenetic regulators. In: Preedy VR, Patel VB, editors. Handbook of nutrition, diet, and epigenetics. Berlin: Springer; 2017.
Ip S, Chung M, Raman G, Chew P, Magula N, DeVine D, et al. Breastfeeding and maternal and infant health outcomes in developed countries. Evid Rep Technol Assess (Full Rep). 2007;153:1–186.
Bener A, Ehlayel MS, Alsowaidi S, Sabbah A. Role of breast feeding in primary prevention of asthma and allergic diseases in a traditional society. Eur Ann Allergy Clin Immunol. 2007;39:337–43.
Oddy WH. Breastfeeding, childhood asthma, and allergic disease. Ann Nutr Metab. 2017;70(Suppl 2):26–36.
Azad MB, Vehling L, Lu Z, Dai D, Subbarao P, Becker AB, et al. Breastfeeding, maternal asthma and wheezing in the first year of life: a longitudinal birth cohort study. Eur Respir J. 2017;49:1602019.
Klopp A, Vehling L, Becker AB, Subbarao P, Mandhane PJ, Turvey SE, et al. Modes of infant feeding and the risk of childhood asthma: a prospective birth cohort study. J Pediatr. 2017;190:192–9.
Palomares O, Akdis M, Martín-Fontecha M, Akdis CA. Mechanisms of immune regulation in allergic diseases: the role of regulatory T and B cells. Immunol Rev. 2017;278:219–36.
Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003;299:1057–61.
Pellerin L, Jenks JA, Bégin P, Bacchetta R, Nadeau KC. Regulatory T cells and their roles in immune dysregulation and allergy. Immunol Res. 2014;58:358–68.
Li X, Zheng Y. Regulatory T cell identity: formation and maintenance. Trends Immunol. 2015;36:344–53.
Tooley KL, El-Merhibi A, Cummins AG, Grose RH, Lymn KA, DeNichilo M, et al. Maternal milk, but not formula, regulates the immune response to beta-lactoglobulin in allergy-prone rat pups. J Nutr. 2009;139:2145–51.
Polansky JK, Kretschmer K, Freyer J, Floess S, Garbe A, Baron U, et al. DNA methylation controls Foxp3 gene expression. Eur J Immunol. 2008;38:1654–63.
Huehn J, Beyer M. Epigenetic and transcriptional control of Foxp3+ regulatory T cells. Semin Immunol. 2015;27:10–8.
Chen W, Konkel JE. Development of thymic Foxp3(+) regulatory T cells: tGF-β matters. Eur J Immunol. 2015;45:958–65.
Hinz D, Bauer M, Röder S, Olek S, Huehn J, Sack U, et al. Cord blood Tregs with stable FOXP3 expression are influenced by prenatal environment and associated with atopic dermatitis at the age of one year. Allergy. 2012;67:380–9.
Paparo L, Nocerino R, Cosenza L, Aitoro R, D’Argenio V, Del Monaco V, et al. Epigenetic features of FoxP3 in children with cow’s milk allergy. Clin Epigenetics. 2016;8:86.
Wang M, Yang V, Davidson EJ, Joetham A, Takeda K, O’Connor BP, et al. Forkhead box protein 3 demethylation is associated with tolerance induction in peanut-induced. J Allergy Clin Immunol. 2018;141:659–70.
Lal G, Bromberg JS. Epigenetic mechanisms of regulation of Foxp3 expression. Blood. 2009;114:3727–35.
Pan W, Zhu S, Yuan M, Cui H, Wang L, Luo X, et al. MicroRNA-21 and microRNA-148a contribute to DNA hypomethylation in lupus CD4+ T cells by directly and indirectly targeting DNA methyltransferase 1. J Immunol. 2010;184:6773–81.
Garzon R, Liu S, Fabbri M, Liu Z, Heaphy CE, Callegari E, et al. MicroRNA-29b induces global DNA hypomethylation and tumor suppressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood. 2009;113:6411–8.
Qin H, Zhu X, Liang J, Wu J, Yang Y, Wang S, et al. MicroRNA-29b contributes to DNA hypomethylation of CD4+ T cells in systemic lupus erythematosus by indirectly targeting DNA methyltransferase 1. J Dermatol Sci. 2013;69:61–7.
Yang Y, Jin Z, Dong R, Zheng C, Huang Y, Zheng Y, et al. MicroRNA-29b/142-5p contribute to the pathogenesis of biliary atresia by regulating the IFN-γ gene. Cell Death Dis. 2018;9:545.
Baddela VS, Nayan V, Rani P, Onteru SK, Singh D. Physicochemical biomolecular insights into buffalo milk-derived nanovesicles. Appl Biochem Biotechnol. 2016;178:544–57.
Waser M, Michels KB, Bieli C, Flöistrup H, Pershagen G, von Mutius E, et al. Inverse association of farm milk consumption with asthma and allergy in rural and suburban populations across Europe. Clin Exp Allergy. 2007;37:661–70.
Loss G, Apprich S, Waser M, Kneifel W, Genuneit J, Büchele G, et al. The protective effect of farm milk consumption on childhood asthma and atopy: the GABRIELA study. J Allergy Clin Immunol. 2011;128:766–73.
Lluis A, Depner M, Gaugler B, Saas P, Casaca VI, Raedler D, et al. Increased regulatory T-cell numbers are associated with farm milk exposure and lower atopic sensitization and asthma in childhood. J Allergy Clin Immunol. 2014;133:551–9.
Schröder PC, Illi S, Casaca VI, Lluis A, Böck A, Roduit C, et al. A switch in regulatory T cells through farm exposure during immune maturation in childhood. Allergy. 2017;72:604–15.
Lu LF, Thai TH, Calado DP, Chaudhry A, Kubo M, Tanaka K, et al. Foxp3-dependent microRNA155 confers competitive fitness to regulatory T cells by targeting SOCS1 protein. Immunity. 2009;30:80–91.
Yoshimura A, Naka T, Kubo M. SOCS proteins, cytokine signalling and immune regulation. Nat Rev Immunol. 2007;7:454–65.
Harada M, Nakashima K, Hirota T, Shimizu M, Doi S, Fujita K, et al. Functional polymorphism in the suppressor of cytokine signaling 1 gene associated with adult asthma. Am J Respir Cell Mol Biol. 2007;36:491–6.
Liu M, Li S, Li MO. TGF-β control of adaptive immune tolerance: a break from Treg cells. BioEssays. 2018;40(11):e1800063.
Wang GJ, Liu Y, Qin A, Shah SV, Deng ZB, Xiang X, et al. Thymus exosomes-like particles induce regulatory T cells. J Immunol. 2008;181:5242–8.
Skogberg G, Telemo E, Ekwall O. Exosomes in the thymus: antigen transfer and vesicles. Front Immunol. 2015;6:366.
Lundberg V, Berglund M, Skogberg G, Lindgren S, Lundqvist C, Gudmundsdottir J, et al. Thymic exosomes promote the final maturation of thymocytes. Sci Rep. 2016;6:36479.
Das LM, Torres-Castillo MD, Gill T, Levine AD. TGF-β conditions intestinal T cells to express increased levels of miR-155, associated with down-regulation of IL-2 and itk mRNA. Mucosal Immunol. 2013;6:167–76.
Saarinen KM, Vaarala O, Klemetti P, Savilahti E. Transforming growth factor-beta1 in mothers’ colostrum and immune responses to cows’ milk proteins in infants with cows’ milk allergy. J Allergy Clin Immunol. 1999;104:1093–8.
Chen X, Gao C, Li H, Huang L, Sun Q, Dong Y, et al. Identification and characterization of microRNAs in raw milk during different periods of lactation, commercial fluid, and powdered milk products. Cell Res. 2010;20:1128–37.
Oestreich AK, Moley KH. Developmental and transmittable origins of obesity-associated health disorders. Trends Genet. 2017;33:399–407.
Soubry A. Epigenetics as a driver of developmental origins of health and disease: did we forget the fathers? Bioessays. 2018;40(1):1700113.
Lawlor DA, Smith GD, O’Callaghan M, Alati R, Mamun AA, Williams GM, et al. Epidemiologic evidence for the fetal overnutrition hypothesis: findings from the mater-university study of pregnancy and its outcomes. Am J Epidemiol. 2007;165:418–24.
Davey Smith G, Steer C, Leary S, Ness A. Is there an intrauterine influence on obesity? Evidence from parent child associations in the Avon Longitudinal Study of Parents and Children (ALSPAC). Arch Dis Child. 2007;92:876–80.
Catalano PM, Farrell K, Thomas A, Huston-Presley L, Mencin P, de Mouzon SH, et al. Perinatal risk factors for childhood obesity and metabolic dysregulation. Am J Clin Nutr. 2009;90:1303–13.
Tanvig M. Offspring body size and metabolic profile—effects of lifestyle intervention in obese pregnant women. Dan Med J. 2014;61:B4893.
Adam S, Elfeky O, Kinhal V, Dutta S, Lai A, Jayabalan N, et al. Review: fetal–maternal communication via extracellular vesicles—implications for complications of pregnancies. Placenta. 2017;54:83–8.
The American College of Obstetricians and Gynecologist: Nurtrtion during pregnancy. https://www.acog.org/Patients/FAQs/Nutrition-During-Pregnancy.
Olsen SF, Halldorsson T, Willett WC, Knudsen VK, Gillman MW, Mikkelsen TB, et al. Milk consumption during pregnancy is associated with increased infant size at birth: prospective cohort study. Am J Clin Nutr. 2007;86:1104–10.
Heppe DH, van Dam RM, Willemsen SP, den Breeijen H, Raat H, Hofman A, et al. Maternal milk consumption, fetal growth, and the risks of neonatal complications: the Generation R Study. Am J Clin Nutr. 2011;94:501–9.
Melnik BC, John SM, Schmitz G. Milk consumption during pregnancy increases birth weight, a risk factor for the development of diseases of civilization. J Transl Med. 2015;13:13.
Melnik BC. The potential impact of maternal milk consumption during pregnancy on mTORC1-driven fetal growth. In: Bhattacharya N, Stubblefield PG, editors. Human fetal growth and development. First and second trimester. Berlin: Springer; 2016. p. 235–58.
Jiang H, Wu W, Zhang M, Li J, Peng Y, Miao TT, et al. Aberrant upregulation of miR-21 in placental tissues of macrosomia. J Perinatol. 2014;34:658–63.
Zhang JT, Cai QY, Ji SS, Zhang HX, Wang YH, Yan HT, et al. Decreased miR-143 and increased miR-21 placental expression levels are associated with macrosomia. Mol Med Rep. 2016;13:3273–80.
Chaiwangyen W, Ospina-Prieto S, Photini SM, Schleussner E, Markert UR, Morales- Prieto DM. Dissimilar microRNA-21 functions and targets in trophoblastic cell lines of different origin. Int J Biochem Cell Biol. 2015;68:187–96.
Lager S, Powell TL. Regulation of nutrient transport across the placenta. J Pregnancy. 2012;2012:179827.
Larqué E, Ruiz-Palacios M, Koletzko B. Placental regulation of fetal nutrient supply. Curr Opin Clin Nutr Metab Care. 2013;16:292–7.
Roos S, Jansson N, Palmberg I, Säljö K, Powell TL, Jansson T. Mammalian target of rapamycin in the human placenta regulates leucine transport and is down-regulated in restricted fetal growth. J Physiol. 2007;582:449–59.
Jansson T, Aye IL, Goberdhan DC. The emerging role of mTORC1 signaling in placental nutrient-sensing. Placenta. 2012;33(Suppl 2):e23–9.
Roos S, Lagerlöf O, Wennergren M, Powell TL, Jansson T. Regulation of amino acid transporters by glucose and growth factors in cultured primary human trophoblast cells is mediated by mTOR signaling. Am J Physiol Cell Physiol. 2009;297:C723–31.
Jansson N, Rosario FJ, Gaccioli F, Lager S, Jones HN, Roos S, et al. Activation of placental mTOR signaling and amino acid transporters in obese women giving birth to large babies. J Clin Endocrinol Metab. 2013;98:105–13.
Sood R, Zehnder JL, Druzin ML, Brown PO. Gene expression patterns in human placenta. Proc Natl Acad Sci USA. 2006;103:5478–83.
Weksberg R, Shuman C, Beckwith JB. Beckwith–Wiedemann syndrome. Eur J Hum Genet. 2010;18:8–14.
Romanelli V, Belinchón A, Benito-Sanz S, Martínez-Glez V, Gracia-Bouthelier R, Heath KE, et al. CDKN1C (p57(Kip2)) analysis in Beckwith–Wiedemann syndrome (BWS) patients: genotype-phenotype correlations, novel mutations, and polymorphisms. Am J Med Genet A. 2010;152A:1390–7.
Brioude F, Netchine I, Praz F, Le Jule M, Calmel C, Lacombe D, et al. Mutations of the imprinted CDKN1C gene as a cause of the overgrowth Beckwith-Wiedemann syndrome: clinical spectrum and functional characterization. Hum Mutat. 2015;36:894–902.
Mussa A, Russo S, de Crescenzo A, Freschi A, Calzari L, Maitz S, et al. Fetal growth patterns in Beckwith–Wiedemann syndrome. Clin Genet. 2016;90:21–7.
Stampone E, Caldarelli I, Zullo A, Bencivenga D, Mancini FP, Della Ragione F, et al. Genetic and epigenetic control of CDKN1C expression: importance in cell commitment and differentiation, tissue homeostasis and human diseases. Int J Mol Sci. 2018;19(4):E1055.
Bar Yamin H, Barnea M, Genzer Y, Chapnik N, Froy O. Long-term commercial cow’s milk consumption and its effects on metabolic parameters associated with obesity in young mice. Mol Nutr Food Res. 2014;58:1061–8.
McGregor RA, Choi MS. microRNAs in the regulation of adipogenesis and obesity. Curr Mol Med. 2011;11:304–16.
Zhang Y, Yu M, Tian W. Physiological and pathological impact of exosomes of adipose tissue. Cell Prolif. 2016;49:3–13.
Yuan Z, Li Q, Luo S, Liu Z, Luo D, Zhang B, et al. PPARγ and Wnt signaling in adipogenic and osteogenic differentiation of mesenchymal stem cells. Curr Stem Cell Res Ther. 2016;11:216–25.
Narayanan K, Kumar S, Padmanabhan P, Gulyas B, Wan ACA, Rajendran VM. Lineage-specific exosomes could override extracellular matrix mediated human mesenchymal stem cell differentiation. Biomaterials. 2018;182:312–22.
Pardo F, Villalobos-Labra R, Sobrevia B, Toledo F, Sobrevia L. Extracellular vesicles in obesity and diabetes mellitus. Mol Aspects Med. 2018;60:81–91.
Kang H, Hata A. The role of microRNAs in cell fate determination of mesenchymal stem cells: balancing adipogenesis and osteogenesis. BMB Rep. 2015;48:319–23.
Shi C, Zhang M, Tong M, Yang L, Pang L, Chen L, et al. miR-148a is associated with obesity and modulates adipocyte differentiation of mesenchymal stem cells through Wnt signaling. Sci Rep. 2015;5:9930.
Shi C, Pang L, Ji C, Wang J, Lin N, Chen J, et al. Obesity-associated miR148a is regulated by cytokines and adipokines via a transcriptional mechanism. Mol Med Rep. 2016;14:5707–12.
Cho YM, Kim TM, Hun Kim D, Hee Kim D, Jeong SW, Kwon OJ. miR-148a is a downstream effector of X-box-binding protein 1 that silences Wnt10b during adipogenesis of 3T3-L1 cells. Exp Mol Med. 2016;48:e226.
Londoño Gentile T, Lu C, Lodato PM, Tse S, Olejniczak SH, Witze ES, et al. DNMT1 is regulated by ATP-citrate lyase and maintains methylation patterns during adipocyte differentiation. Mol Cell Biol. 2013;33:3864–78.
Yang X, Wu R, Shan W, Yu L, Xue B, Shi H. DNA methylation biphasically regulates 3T3-L1 preadipocyte differentiation. Mol Endocrinol. 2016;30:677–87.
Schwenk RW, Vogel H, Schürmann A. Genetic and epigenetic control of metabolic health. Mol Metab. 2013;2:337–47.
Monda KL, Chen GK, Taylor KC, Palmer C, Edwards TL, Lange LA, et al. A meta-analysis identifies new loci associated with body mass index in individuals of African ancestry. Nat Genet. 2013;45:690–6.
Voisin S, Almén MS, Zheleznyakova GY, Lundberg L, Zarei S, Castillo S, et al. Many obesity-associated SNPs strongly associate with DNA methylation changes at proximal promoters and enhancers. Genome Med. 2015;7:103.
Kim YJ, Hwang SH, Cho HH, Shin KK, Bae YC, Jung JS. MicroRNA 21 regulates the proliferation of human adipose tissue-derived mesenchymal stem cells and high-fat diet-induced obesity alters microRNA 21 expression in white adipose tissues. J Cell Physiol. 2012;227:183–93.
Mei Y, Bian C, Li J, Du Z, Zhou H, Yang Z, et al. miR-21 modulates the ERK-MAPK signaling pathway by regulating SPRY2 expression during human mesenchymal stem cell differentiation. J Cell Biochem. 2013;114:1374–84.
Kang M, Yan LM, Zhang WY, Li YM, Tang AZ, Ou HS. Role of microRNA-21 in regulating 3T3-L1 adipocyte differentiation and adiponectin expression. Mol Biol Rep. 2013;40:5027–34.
Guglielmi V, D’Adamo M, Menghini R, Cardellini M, Gentileschi P, Federici M, et al. MicroRNA 21 is up-regulated in adipose tissue of obese diabetic subjects. Nutr Healthy Aging. 2017;4:141–5.
Bian Y, Lei Y, Wang C, Wang J, Wang L, Liu L, et al. Epigenetic regulation of miR-29s affects the lactation activity of dairy cow mammary epithelial cells. J Cell Physiol. 2015;230:2152–63.
Chen Y, Siegel F, Kipschull S, Haas B, Fröhlich H, Meister G, et al. miR-155 regulates differentiation of brown and beige adipocytes via a bistable circuit. Nat Commun. 2013;4:1769.
Yang T, Martin P, Fogarty B, Brown A, Schurman K, Phipps R, et al. Exosome delivered anticancer drugs across the blood–brain barrier for brain cancer therapy in Danio rerio. Pharm Res. 2015;32:2003–14.
Matsumoto J, Stewart T, Banks WA, Zhang J. The transport mechanism of extracellular vesicles at the blood–brain barrier. Curr Pharm Des. 2017;23:6206–14.
Zhang Y, Kim MS, Jia B, Yan J, Zuniga-Hertz JP, Han C, et al. Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature. 2017;548:52–7.
Zanchi D, Depoorter A, Egloff L, Haller S, Mählmann L, Lang UE, et al. The impact of gut hormones on the neural circuit of appetite and satiety: a systematic review. Neurosci Biobehav Rev. 2017;80:457–75.
Clerc P, Coll Constans MG, Lulka H, Broussaud S, Guigné C, Leung-Theung-Long S, et al. Involvement of cholecystokinin 2 receptor in food intake regulation: hyperphagia and increased fat deposition in cholecystokinin 2 receptor-deficient mice. Endocrinology. 2007;148:1039–49.
Chen H, Kent S, Morris MJ. Is the CCK2 receptor essential for normal regulation of body weight and adiposity? Eur J Neurosci. 2006;24:1427–33.
Yu B, Lv X, Su L, Li J, Yu Y, Gu Q, et al. MiR-148a functions as a tumor suppressor by targeting CCK-BR via inactivating STAT3 and Akt in human gastric cancer. PLoS ONE. 2016;11:e0158961.
Pasin G, Comerford KB. Dairy foods and dairy proteins in the management of type 2 diabetes: a systematic review of the clinical evidence. Adv Nutr. 2015;6:245–59.
Cândido FG, Ton WT, Alfenas Rde C. Dairy products consumption versus type 2 diabetes prevention and treatment; a review of recent findings from human studies. Nutr Hosp. 2013;28:1384–95.
Tong X, Dong JY, Wu ZW, Li W, Qin LQ. Dairy consumption and risk of type 2 diabetes mellitus: a meta-analysis of cohort studies. Eur J Clin Nutr. 2011;65:1027–31.
Aune D, Norat T, Romundstad P, Vatten LJ. Dairy products and the risk of type 2 diabetes: a systematic review and dose-response meta-analysis of cohort studies. Am J Clin Nutr. 2013;98:1066–83.
Díaz-López A, Bulló M, Martínez-González MA, Corella D, Estruch R, Fitó M, et al. Dairy product consumption and risk of type 2 diabetes in an elderly Spanish Mediterranean population at high cardiovascular risk. Eur J Nutr. 2016;55:349–60.
Guasch-Ferré M, Becerra-Tomás N, Ruiz-Canela M, Corella D, Schröder H, Estruch R, et al. Total and subtypes of dietary fat intake and risk of type 2 diabetes mellitus in the Prevención con Dieta Mediterránea (PREDIMED) study. Am J Clin Nutr. 2017;105:723–35.
Sluijs I, Forouhi NG, Beulens JW, van der Schouw YT, Agnoli C, Arriola L, et al. The amount and type of dairy product intake and incident type 2 diabetes: results from the EPIC-InterAct Study. Am J Clin Nutr. 2012;96:382–90.
Song Y, Chavarro JE, Cao Y, Qiu W, Mucci L, Sesso HD, et al. Whole milk intake is associated with prostate cancer-specific mortality among U.S. male physicians. J Nutr. 2013;143:189–96.
Hruby A, Ma J, Rogers G, Meigs JB, Jacques PF. Associations of dairy intake with incident prediabetes or diabetes in middle-aged adults vary by both dairy type and glycemic status. J Nutr. 2017;147:1764–75.
Melnik BC. Milk—a nutrient system of mammalian evolution promoting mTORC1-dependent translation. Int J Mol Sci. 2015;16:17048–87.
Melnik BC. The pathogenic role of persistent milk signaling in mTORC1- and milk-microRNA-driven type 2 diabetes mellitus. Curr Diabetes Rev. 2015;11:46–62.
McCormack SE, Shaham O, McCarthy MA, Deik AA, Wang TJ, Gerszten RE, et al. Circulating branched-chain amino acid concentrations are associated with obesity and future insulin resistance in children and adolescents. Pediatr Obes. 2013;8:52–61.
Yoon MS. The emerging role of branched-chain amino acids in insulin resistance and metabolism. Nutrients. 2016;8:E405.
Ruiz-Canela M, Guasch-Ferré M, Toledo E, Clish CB, Razquin C, Liang L, et al. Plasma branched chain/aromatic amino acids, enriched Mediterranean diet and risk of type 2 diabetes: case-cohort study within the PREDIMED Trial. Diabetologia. 2018;61:1560–71.
Andersson-Hall U, Gustavsson C, Pedersen A, Malmodin D, Joelsson L, Holmäng A. Higher concentrations of BCAAs and 3-HIB are associated with insulin resistance in the transition from gestational diabetes to type 2 diabetes. J Diabetes Res. 2018;2018:4207067.
Bloomgarden Z. Diabetes and branched-chain amino acids: what is the link? J Diabetes. 2018;10:350–2.
Arany Z, Neinast M. Branched chain amino acids in metabolic disease. Curr Diab Rep. 2018;18:76.
Yadao DR, MacKenzie S, Bergdahl A. Reducing branched-chain amino acid intake to reverse metabolic complications in obesity and type 2 diabetes. J Physiol. 2018;596:3455–6.
Arnold N, Koppula PR, Gul R, Luck C, Pulakat L. Regulation of cardiac expression of the diabetic marker microRNA miR-29. PLoS ONE. 2014;9:e103284.
Chakraborty C, Doss CG, Bandyopadhyay S, Agoramoorthy G. Influence of miRNA in insulin signaling pathway and insulin resistance: micro-molecules with a major role in type-2 diabetes. Wiley Interdiscip Rev RNA. 2014;5:697–712.
Ślusarz A, Pulakat L. The two faces of miR-29. J Cardiovasc Med (Hagerstown). 2015;16:480–90.
Wu B, Miller D. Involvement of microRNAs in diabetes and its complications. Methods Mol Biol. 2017;1617:225–39.
Massart J, Sjögren RJO, Lundell LS, Mudry JM, Franck N, O’Gorman DJ, et al. Altered miR-29 expression in type 2 diabetes influences glucose and lipid metabolism in skeletal muscle. Diabetes. 2017;66:1807–18.
Mersey BD, Jin P, Danner DJ. Human microRNA (miR29b) expression controls the amount of branched chain alpha-ketoacid dehydrogenase complex in a cell. Hum Mol Genet. 2005;14:3371–7.
Hernández-Alvarez MI, Díaz-Ramos A, Berdasco M, Cobb J, Planet E, Cooper D, et al. Early-onset and classical forms of type 2 diabetes show impaired expression of genes involved in muscle branched-chain amino acids metabolism. Sci Rep. 2017;7:13850.
Brosnan JT, Brosnan ME. Branched-chain amino acids: enzyme and substrate regulation. J Nutr. 2006;136(1 Suppl):207S–2011S.
Dodd KM, Tee AR. Leucine and mTORC1: a complex relationship. Am J Physiol Endocrinol Metab. 2012;302:E1329–42.
Bar-Peled L, Sabatini DM. Regulation of mTORC1 by amino acids. Trends Cell Biol. 2014;24:400–6.
Duan Y, Li F, Tan K, Liu H, Li Y, Liu Y, et al. Key mediators of intracellular amino acids signaling to mTORC1 activation. Amino Acids. 2015;47:857–67.
Jewell JL, Kim YC, Russell RC, Yu FX, Park HW, Plouffe SW, et al. Metabolism. Differential regulation of mTORC1 by leucine and glutamine. Science. 2015;347:194–8.
Schriever SC, Deutsch MJ, Adamski J, Roscher AA, Ensenauer R. Cellular signaling of amino acids towards mTORC1 activation in impaired human leucine catabolism. J Nutr Biochem. 2013;24:824–31.
Yang J, Chi Y, Burkhardt BR, Guan Y, Wolf BA. Leucine metabolism in regulation of insulin secretion from pancreatic beta cells. Nutr Rev. 2010;68:270–9.
de Oliveira CA, Latorraca MQ, de Mello MA, Carneiro EM. Mechanisms of insulin secretion in malnutrition: modulation by amino acids in rodent models. Amino Acids. 2011;40:1027–34.
Tricò D, Frascerra S, Baldi S, Mengozzi A, Nesti L, Mari A, et al. The insulinotropic effect of a high-protein nutrient preload is mediated by the increase of plasma amino acids in type 2 diabetes. Eur J Nutr. 2018. https://doi.org/10.1007/s00394-018-1778-y.
Kleinridders A, Ferris HA, Cai W, Kahn CR. Insulin action in brain regulates systemic metabolism and brain function. Diabetes. 2014;63:2232–43.
Liu Z, Jeppesen PB, Gregersen S, Larsen LB, Hermansen K. Chronic exposure to leucine in vitro induces β-cell dysfunction in INS-1E cells and mouse islets. J Endocrinol. 2012;215:79–88.
Um SH, D’Alessio D, Thomas G. Nutrient overload, insulin resistance, and ribosomal protein S6 kinase 1, S6K1. Cell Metab. 2006;3:393–402.
Tremblay F, Brûlé S, Hee Um S, Li Y, Masuda K, et al. Identification of IRS-1 Ser- 1101 as a target of S6K1 in nutrient- and obesity-induced insulin resistance. Proc Natl Acad Sci USA. 2007;104:14056–61.
Melnik BC. Leucine signaling in the pathogenesis of type 2 diabetes and obesity. World J Diabetes. 2012;3:38–53.
Catalán V, Gómez-Ambrosi J, Rodríguez A, Ramírez B, Andrada P, Rotellar F, et al. Expression of S6K1 in human visceral adipose tissue is upregulated in obesity and related to insulin resistance and inflammation. Acta Diabetol. 2015;52:257–66.
Kos K, Wilding JP. SPARC: a key player in the pathologies associated with obesity and diabetes. Nat Rev Endocrinol. 2010;6:225–35.
Harries LW, McCulloch LJ, Holley JE, Rawling TJ, Welters HJ, Kos K. A role for SPARC in the moderation of human insulin secretion. PLoS ONE. 2013;8:e68253.
Wang C, Gao C, Zhuang JL, Ding C, Wang Y. A combined approach identifies three mRNAs that are down-regulated by microRNA-29b and promote invasion ability in the breast cancer cell line MCF-7. J Cancer Res Clin Oncol. 2012;138:2127–36.
Song H, Ding L, Zhang S, Wang W. MiR-29 family members interact with SPARC to regulate glucose metabolism. Biochem Biophys Res Commun. 2018;497:667–74.
Artner I, Blanchi B, Raum JC, Guo M, Kaneko T, Cordes S, et al. MafB is required for islet beta cell maturation. Proc Natl Acad Sci USA. 2007;104:3853–8.
Artner I, Hang Y, Mazur M, Yamamoto T, Guo M, Lindner J, et al. MafA and MafB regulate genes critical to beta-cells in a unique temporal manner. Diabetes. 2010;59:2530–9.
Cheng P, Chen C, He HB, Hu R, Zhou HD, Xie H, et al. miR-148a regulates osteoclastogenesis by targeting V-maf musculoaponeurotic fibrosarcoma oncogene homolog B. J Bone Miner Res. 2013;28:1180–90.
Dai C, Kayton NS, Shostak A, Poffenberger G, Cyphert HA, Aramandla R, et al. Stress-impaired transcription factor expression and insulin secretion in transplanted human islets. J Clin Invest. 2016;126:1857–70.
Michaëlsson K, Wolk A, Langenskiöld S, Basu S, Warensjö Lemming E, Melhus H, et al. Milk intake and risk of mortality and fractures in women and men: cohort studies. BMJ. 2014;349:g6015.
Tognon G, Nilsson LM, Shungin D, Lissner L, Jansson JH, Renström F, et al. Nonfermented milk and other dairy products: associations with all-cause mortality. Am J Clin Nutr. 2017;105:1502–11.
de Goede J, Soedamah-Muthu SS, Pan A, Gijsbers L, Geleijnse JM. Dairy consumption and risk of stroke: a systematic review and updated dose-response meta-analysis of prospective cohort studies. J Am Heart Assoc. 2016;5(5):e002787.
Dehghan M, Mente A, Rangarajan S, Sheridan P, Mohan V, Iqbal R, et al. Association of dairy intake with cardiovascular disease and mortality in 21 countries from five continents (PURE): a prospective cohort study. Lancet. 2018;392:2288–97.
Singh AK, Aryal B, Zhang X, Fan Y, Price NL, Suárez Y, et al. Posttranscriptional regulation of lipid metabolism by non-coding RNAs and RNA binding proteins. Semin Cell Dev Biol. 2018;81:129–40.
Aryal B, Suárez Y. Non-coding RNA regulation of endothelial and macrophage functions during atherosclerosis. Vascul Pharmacol. 2018;1891(17):30360–9. https://doi.org/10.1016/j.vph.2018.03.001 (Epub ahead of print).
Schmitz G, Grandl M. Lipid homeostasis in macrophages—implications for atherosclerosis. Rev Physiol Biochem Pharmacol. 2008;160:93–125.
Maguire EM, Pearce SWA, Xiao Q. Foam cell formation: a new target for fighting atherosclerosis and cardiovascular disease. Vascul Pharmacol. 2018;1891(18):30108. https://doi.org/10.1016/j.vph.2018.08.002 (Epub ahead of print).
Zhang Y, Hu YW, Zheng L, Wang Q. Characteristics and roles of exosomes in cardiovascular disease. DNA Cell Biol. 2017;36:202–11.
Lu M, Yuan S, Li S, Li L, Liu M, Wan S. The exosome-derived biomarker in atherosclerosis and its clinical application. J Cardiovasc Transl Res. 2018. https://doi.org/10.1007/s12265-018-9796-y (Epub ahead of print).
Huang F, Zhao JL, Wang L, Gao CC, Liang SQ, An DJ, et al. miR-148a-3p mediates Notch signaling to promote the differentiation and M1 activation of macrophages. Front Immunol. 2017;8:1327.
Nordgren TM, Heires AJ, Zempleni J, Swanson BJ, Wichman C, Romberger DJ. Bovine milk-derived extracellular vesicles enhance inflammation and promote M1 polarization following agricultural dust exposure in mice. J Nutr Biochem. 2018;64:110–20.
Goedeke L, Rotllan N, Canfrán-Duque A, Aranda JF, Ramírez CM, Araldi E, et al. MicroRNA-148a regulates LDL receptor and ABCA1 expression to control circulating lipoprotein levels. Nat Med. 2015;21:1280–9.
Rotllan N, Price N, Pati P, Goedeke L, Fernández-Hernando C. microRNAs in lipoprotein metabolism and cardiometabolic disorders. Atherosclerosis. 2016;246:352–60.
Goldstein JL, Brown MS. The LDL receptor. Arterioscler Thromb Vasc Biol. 2009;29:431–8.
Langmann T, Klucken J, Reil M, Liebisch G, Luciani MF, Chimini G, et al. Molecular cloning of the human ATP-binding cassette transporter 1 (hABC1): evidence for sterol-dependent regulation in macrophages. Biochem Biophys Res Commun. 1999;257:29–33.
Schmitz G, Kaminski WE, Orsó E. ABC transporters in cellular lipid trafficking. Curr Opin Lipidol. 2000;11:493–501.
McNeish J, Aiello RJ, Guyot D, Turi T, Gabel C, Aldinger C, et al. High density lipoprotein deficiency and foam cell accumulation in mice with targeted disruption of ATP-binding cassette transporter-1. Proc Natl Acad Sci USA. 2000;97:4245–50.
Yang A, Sun Y, Gao Y, Yang S, Mao C, Ding N, et al. Reciprocal regulation between miR-148a/152 and DNA methyltransferase 1 is associated with hyperhomocysteinemia-accelerated atherosclerosis. DNA Cell Biol. 2017;36:462–74.
Ji R, Cheng Y, Yue J, Yang J, Liu X, Chen H, et al. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of MicroRNA in vascular neointimal lesion formation. Circ Res. 2007;100:1579–88.
Weber M, Baker MB, Moore JP, Searles CD. MiR-21 is induced in endothelial cells by shear stress and modulates apoptosis and eNOS activity. Biochem Biophys Res Commun. 2010;393:643–8.
Dellago H, Preschitz-Kammerhofer B, Terlecki-Zaniewicz L, Schreiner C, Fortschegger K, Chang MW, et al. High levels of oncomiR-21 contribute to the senescence-induced growth arrest in normal human cells and its knock-down increases the replicative lifespan. Aging Cell. 2013;12:446–58.
Feskanich D, Bischoff-Ferrari HA, Frazier AL, Willett WC. Milk consumption during teenage years and risk of hip fractures in older adults. JAMA Pediatr. 2014;168:54–60.
Feskanich D, Meyer HE, Fung TT, Bischoff-Ferrari HA, Willett WC. Milk and other dairy foods and risk of hip fracture in men and women. Osteoporos Int. 2018;29:385–96.
Michaëlsson K, Wolk A, Lemming EW, Melhus H, Byberg L. Intake of milk or fermented milk combined with fruit and vegetable consumption in relation to hip fracture rates: a cohort study of swedish women. J Bone Miner Res. 2018;33:449–57.
Tu MY, Chen HL, Tung YT, Kao CC, Hu FC, Chen CM. Short-term effects of kefir-fermented milk consumption on bone mineral density and bone metabolism in a randomized clinical trial of osteoporotic patients. PLoS ONE. 2015;10:e0144231.
Biver E, Durosier-Izart C, Merminod F, Chevalley T, van Rietbergen B, Ferrari SL, et al. Fermented dairy products consumption is associated with attenuated cortical bone loss independently of total calcium, protein, and energy intakes in healthy postmenopausal women. Osteoporos Int. 2018;29:1771–82.
Bian S, Hu J, Zhang K, Wang Y, Yu M, Ma J. Dairy product consumption and risk of hip fracture: a systematic review and meta-analysis. BMC Public Health. 2018;18:165.
Feskanich D, Willett WC, Stampfer MJ, Colditz GA. Milk, dietary calcium, and bone fractures in women: a 12-year prospective study. Am J Public Health. 1997;87:992–7.
Feskanich D, Willett WC, Colditz GA. Calcium, vitamin D, milk consumption, and hip fractures: a prospective study among postmenopausal women. Am J Clin Nutr. 2003;77:504–11.
Bischoff-Ferrari HA, Dawson-Hughes B, Baron JA, Kanis JA, Orav EJ, Staehelin HB, et al. Milk intake and risk of hip fracture in men and women: a meta-analysis of prospective cohort studies. J Bone Miner Res. 2011;26:833–9.
Sahni S, Tucker KL, Kiel DP, Quach L, Casey VA, Hannan MT. Milk and yogurt consumption are linked with higher bone mineral density but not with hip fracture: the Framingham Offspring Study. Arch Osteoporos. 2013;8:119.
Sahni S, Mangano KM, Tucker KL, Kiel DP, Casey VA, Hannan MT. Protective association of milk intake on the risk of hip fracture: results from the Framingham Original Cohort. J Bone Miner Res. 2014;29:1756–62.
Vogel KA, Martin BR, McCabe LD, Peacock M, Warden SJ, McCabe GP, et al. The effect of dairy intake on bone mass and body composition in early pubertal girls and boys: a randomized controlled trial. Am J Clin Nutr. 2017;105:1214–29.
Fardellone P, Séjourné A, Blain H, Cortet B, Thomas T. GRIO Scientific Committee: osteoporosis: is milk a kindness or a curse? Joint Bone Spine. 2017;84:275–81.
Bergholdt HKM, Larsen MK, Varbo A, Nordestgaard BG, Ellervik C. Lactase persistence, milk intake, hip fracture and bone mineral density: a study of 97 811 Danish individuals and a meta-analysis. J Intern Med. 2018. https://doi.org/10.1111/joim.12753 (Epub ahead of print).
Infante A, Rodríguez CI. Osteogenesis and aging: lessons from mesenchymal stem cells. Stem Cell Res Ther. 2018;9:244.
Blank V, Andrews NC. The Maf transcription factors: regulators of differentiation. Trends Biochem Sci. 1997;22:437–41.
Nishikawa K, Nakashima T, Takeda S, Isogai M, Hamada M, Kimura A, et al. Maf promotes osteoblast differentiation in mice by mediating the age-related switch in mesenchymal cell differentiation. J Clin Invest. 2010;120:3455–65.
Behera J, Tyagi N. Exosomes: mediators of bone diseases, protection, and therapeutics potential. Oncoscience. 2018;5:181–95.
van Wijnen AJ, van de Peppel J, van Leeuwen JP, Lian JB, Stein GS, Westendorf JJ, et al. MicroRNA functions in osteogenesis and dysfunctions in osteoporosis. Curr Osteoporos Rep. 2013;11:72–82.
Liu M, Sun Y, Zhang Q. Emerging role of extracellular vesicles in bone remodeling. J Dent Res. 2018;97:859–68.
Oliveira MC, Di Ceglie I, Arntz OJ, van den Berg WB, van den Hoogen FH, Ferreira AV, et al. Milk-derived nanoparticle fraction promotes the formation of small osteoclasts but reduces bone resorption. J Cell Physiol. 2017;232:225–33.
Tian L, Zheng F, Li Z, Wang H, Yuan H, Zhang X, et al. miR-148a-3p regulates adipocyte and osteoblast differentiation by targeting lysine-specific demethylase 6b. Gene. 2017;627:32–9.
Kelch S, Balmayor ER, Seeliger C, Vester H, Kirschke JS, van Griensven M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci Rep. 2017;7:15861.
Liu J, Li D, Wu X, Dang L, Lu A, Zhang G. Bone-derived exosomes. Curr Opin Pharmacol. 2017;34:64–9.
Xie Y, Chen Y, Zhang L, Ge W, Tang P. The roles of bone-derived exosomes and exosomal microRNAs in regulating bone remodeling. J Cell Mol Med. 2017;21:1033–41.
Grieco GE, Cataldo D, Ceccarelli E, Nigi L, Catalano G, Brusco N, et al. Serum levels of miR-148a and miR-21-5p are increased in type 1 diabetic patients and correlated with markers of bone strength and metabolism. Noncoding RNA. 2018;4(4):E37.
Zhou Y, Deng HW, Shen H. Circulating monocytes: an appropriate model for bone- related study. Osteoporos Int. 2015;26:2561–72.
Kim K, Kim JH, Lee J, Jin HM, Kook H, Kim KK, et al. MafB negatively regulates RANKL-mediated osteoclast differentiation. Blood. 2007;109:3253–9.
Buscaglia LE, Li Y. Apoptosis and the target genes of microRNA-21. Chin J Cancer. 2011;30:371–80.
Liao J, Liu R, Shi YJ, Yin LH, Pu YP. Exosome-shuttling microRNA-21 promotes cell migration and invasion-targeting PDCD4 in esophageal cancer. Int J Oncol. 2016;48:2567–79.
Oliveira MC, Arntz OJ, Blaney Davidson EN, van Lent PL, Koenders MI, van der Kraan PM, et al. Milk extracellular vesicles accelerate osteoblastogenesis but impair bone matrix formation. J Nutr Biochem. 2016;30:74–84.
Quinn JM, Itoh K, Udagawa N, Hausler K, Yasuda H, Shima N, et al. Transforming growth factor beta affects osteoclast differentiation via direct and indirect actions. J Bone Miner Res. 2001;16:1787–94.
Kylmäoja E, Nakamura M, Turunen S, Patlaka C, Andersson G, Lehenkari P, et al. Peripheral blood monocytes show increased osteoclast differentiation potential compared to bone marrow monocytes. Heliyon. 2018;4:e00780.
Neglia C, Argentiero A, Chitano G, Agnello N, Ciccarese R, Vigilanza A, et al. Diabetes and obesity as independent risk factors for osteoporosis: updated results from the ROIS/EMEROS registry in a population of five thousand post-menopausal women living in a region characterized by heavy environmental pressure. Int J Environ Res Public Health. 2016;13:E1067.
Chen H, Zhang SM, Hernán MA, Willett WC, Ascherio A. Diet and Parkinson’s disease: a potential role of dairy products in men. Ann Neurol. 2002;52:793–801.
Park M, Ross GW, Petrovitch H, White LR, Masaki KH, Nelson JS, et al. Consumption of milk and calcium in midlife and the future risk of Parkinson disease. Neurology. 2005;64:1047–51.
Kyrozis A, Ghika A, Stathopoulos P, Vassilopoulos D, Trichopoulos D, Trichopoulou A. Dietary and lifestyle variables in relation to incidence of Parkinson’s disease in Greece. Eur J Epidemiol. 2013;28:67–77.
Jiang W, Ju C, Jiang H, Zhang D. Dairy foods intake and risk of Parkinson’s disease: a dose-response meta-analysis of prospective cohort studies. Eur J Epidemiol. 2014;29:613–9.
Abbott RD, Ross GW, Petrovitch H, Masaki KH, Launer LJ, Nelson JS, et al. Midlife milk consumption and substantia nigra neuron density at death. Neurology. 2016;86:512–9.
Hughes KC, Gao X, Kim IY, Wang M, Weisskopf MG, Schwarzschild MA, et al. Intake of dairy foods and risk of Parkinson disease. Neurology. 2017;89:46–52.
Properzi F, Ferroni E, Poleggi A, Vinci R. The regulation of exosome function in the CNS: implications for neurodegeneration. Swiss Med Wkly. 2015;145:w14204.
Vella LJ, Hill AF, Cheng L. Focus on extracellular vesicles: exosomes and their role in protein trafficking and biomarker potential in Alzheimer’s and Parkinson’s disease. Int J Mol Sci. 2016;17:173.
Chistiakov DA, Chistiakov AA. α-Synuclein-carrying extracellular vesicles in Parkinson’s disease: deadly transmitters. Acta Neurol Belg. 2017;117:43–51.
Tofaris GK. A critical assessment of exosomes in the pathogenesis and stratification of Parkinson’s disease. J Parkinsons Dis. 2017;7:569–76.
Grey M, Dunning CJ, Gaspar R, Grey C, Brundin P, Sparr E, et al. Acceleration of α- synuclein aggregation by exosomes. J Biol Chem. 2015;290:2969–82.
Leggio L, Vivarelli S, L’Episcopo F, Tirolo C, Caniglia S, Testa N, et al. microRNAs in Parkinson’s disease: from pathogenesis to novel diagnostic and therapeutic approaches. Int J Mol Sci. 2017;18:E2698.
Calabrese V, Santoro A, Monti D, Crupi R, Di Paola R, Latteri S, et al. Aging and Parkinson’s disease: inflammaging, neuroinflammation and biological remodeling as key factors in pathogenesis. Free Radic Biol Med. 2018;115:80–91.
Prajapati P, Sripada L, Singh K, Bhatelia K, Singh R, Singh R. TNF-α regulates miRNA targeting mitochondrial complex-I and induces cell death in dopaminergic cells. Biochim Biophys Acta. 2015;1852:451–61.
Thome AD, Harms AS, Volpicelli-Daley LA, Standaert DG. microRNA-155 regulates alpha-synuclein-induced inflammatory responses in models of Parkinson disease. J Neurosci. 2016;36:2383–90.
Jowaed A, Schmitt I, Kaut O, Wüllner U. Methylation regulates alpha-synuclein expression and is decreased in Parkinson’s disease patients’ brains. J Neurosci. 2010;30:6355–63559.
Matsumoto L, Takuma H, Tamaoka A, Kurisaki H, Date H, Tsuji S, et al. CpG demethylation enhances alpha-synuclein expression and affects the pathogenesis of Parkinson’s disease. PLoS ONE. 2010;5:e15522.
Desplats P, Spencer B, Coffee E, Patel P, Michael S, Patrick C, Adame A, et al. Alpha-synuclein sequesters Dnmt1 from the nucleus: a novel mechanism for epigenetic alterations in Lewy body diseases. J Biol Chem. 2011;286:9031–7.
Aune D, Lau R, Chan DS, Vieira R, Greenwood DC, Kampman E, et al. Dairy products and colorectal cancer risk: a systematic review and meta-analysis of cohort studies. Ann Oncol. 2012;23:37–45.
Ralston RA, Truby H, Palermo CE, Walker KZ. Colorectal cancer and nonfermented milk, solid cheese, and fermented milk consumption: a systematic review and meta- analysis of prospective studies. Crit Rev Food Sci Nutr. 2014;54:1167–79.
Vieira AR, Abar L, Chan DSM, Vingeliene S, Polemiti E, Stevens C, et al. Foods and beverages and colorectal cancer risk: a systematic review and meta-analysis of cohort studies, an update of the evidence of the WCRF-AICR Continuous Update Project. Ann Oncol. 2017;28:1788–802.
Takahashi M, Cuatrecasas M, Balaguer F, Hur K, Toiyama Y, Castells A, et al. The clinical significance of MiR-148a as a predictive biomarker in patients with advanced colorectal cancer. PLoS ONE. 2012;7:e46684.
Tsai HL, Yang IP, Huang CW, Ma CJ, Kuo CH, Lu CY, et al. Clinical significance of microRNA-148a in patients with early relapse of stage II stage and III colorectal cancer after curative resection. Transl Res. 2013;162:258–68.
Baltruskeviciene E, Schveigert D, Stankevicius V, Mickys U, Zvirblis T, Bublevic J, et al. Down-regulation of miRNA-148a and miRNA-625-3p in colorectal cancer is associated with tumor budding. BMC Cancer. 2017;17:607.
Zhu Y, Gu L, Li Y, Lin X, Shen H, Cui K, Chen L, et al. miR-148a inhibits colitis and colitis-associated tumorigenesis in mice. Cell Death Differ. 2017;24:2199–209.
Foran E, Garrity-Park MM, Mureau C, Newell J, Smyrk TC, Limburg PJ, et al. Upregulation of DNA methyltransferase-mediated gene silencing, anchorage-independent growth, and migration of colon cancer cells by interleukin-6. Mol Cancer Res. 2010;8:471–81.
Huang C, Liu H, Gong XL, Wu L, Wen B. Expression of DNA methyltransferases and target microRNAs in human tissue samples related to sporadic colorectal cancer. Oncol Rep. 2016;36:2705–14.
Guo Y, Wang M, Jia X, Zhu H, Zhi Y, Yuan L. Wnt signaling pathway upregulates DNMT1 to trigger NHERF1 promoter hypermethylation in colon cancer. Oncol Rep. 2018;40:1165–73.
Zucchini C, Martinelli M, De Sanctis P, Rodia MT, Mattei G, Ugolini G, et al. Possible gender-related modulation by the ROCK1 gene in colorectal cancer susceptibility. Pathobiology. 2015;82:252–348.
Zhu QD, Zhou QQ, Dong L, Huang Z, Wu F, Deng X. MiR-199a-5p inhibits the growth and metastasis of colorectal cancer cells by targeting ROCK1. Technol Cancer Res Treat. 2018;17:1533034618775509.
Zheng B, Liang L, Wang C, Huang S, Cao X, Zha R, et al. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancer. Clin Cancer Res. 2011;17:7574–83.
Landskron G, De la Fuente M, Thuwajit P, Thuwajit C, Hermoso MA. Chronic inflammation and cytokines in the tumor microenvironment. J Immunol Res. 2014;2014:149185.
Friedrich M, Pracht K, Mashreghi MF, Jäck HM, Radbruch A, Seliger B. The role of the miR-148/-152 family in physiology and disease. Eur J Immunol. 2017;47:2026–38.
Liu X, Zhan Z, Xu L, Ma F, Li D, Guo Z, et al. MicroRNA-148/152 impair innate response and antigen presentation of TLR-triggered dendritic cells by targeting CaMKIIα. J Immunol. 2010;185:7244–51.
Ferlay J, Colombet M, Soerjomataram I, Dyba T, Randi G, Bettio M, et al. Cancer incidence and mortality patterns in Europe: estimates for 40 countries and 25 major cancers in 2018. Eur J Cancer. 2018;103:356–87.
Lu W, Chen H, Niu Y, Wu H, Xia D, Wu Y. Dairy products intake and cancer mortality risk: a meta-analysis of 11 population-based cohort studies. Nutr J. 2016;15:91.
Downer MK, Batista JL, Mucci LA, Stampfer MJ, Epstein MM, Håkansson N, et al. Dairy intake in relation to prostate cancer survival. Int J Cancer. 2017;140:2060–9.
Steck SE, Omofuma OO, Su LJ, Maise AA, Woloszynska-Read A, Johnson CS, et al. Calcium, magnesium, and whole-milk intakes and high-aggressive prostate cancer in the North Carolina-Louisiana Prostate Cancer Project (PCaP). Am J Clin Nutr. 2018;107:799–807.
Pettersson A, Kasperzyk JL, Kenfield SA, Richman EL, Chan JM, Willett WC, et al. Milk and dairy consumption among men with prostate cancer and risk of metastases and prostate cancer death. Cancer Epidemiol Biomarkers Prev. 2012;21:428–36.
Tat D, Kenfield SA, Cowan JE, Broering JM, Carroll PR, Van Blarigan EL, et al. Milk and other dairy foods in relation to prostate cancer recurrence: data from the cancer of the prostate strategic urologic research endeavor (CaPSURE™). Prostate. 2018;78:32–9.
Kuefer R, Varambally S, Zhou M, Lucas PC, Loeffler M, Wolter H, et al. Alpha-Methylacyl-CoA racemase: expression levels of this novel cancer biomarker depend on tumor differentiation. Am J Pathol. 2002;161:841–8.
Zadra G, Photopoulos C, Loda M. The fat side of prostate cancer. Biochim Biophys Acta. 2013;1831:1518–32.
Wu X, Daniels G, Lee P, Monaco ME. Lipid metabolism in prostate cancer. Am J Clin Exp Urol. 2014;2:111–20.
Mobley JA, Leav I, Zielie P, Wotkowitz C, Evans J, Lam YW, et al. Branched fatty acids in dairy and beef products markedly enhance alpha-methylacyl-CoA racemase expression in prostate cancer cells in vitro. Cancer Epidemiol Biomarkers Prev. 2003;12:775–83.
Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev. 2013;32:623–42.
Kahlert C, Kalluri R. Exosomes in tumor microenvironment influence cancer progression and metastasis. J Mol Med (Berl). 2013;91:431–7.
Kosaka N, Yoshioka Y, Tominaga N, Hagiwara K, Katsuda T, Ochiya T. Dark side of the exosome: the role of the exosome in cancer metastasis and targeting the exosome as a strategy for cancer therapy. Future Oncol. 2014;10:671–81.
Yu S, Cao H, Shen B, Feng J. Tumor-derived exosomes in cancer progression and treatment failure. Oncotarget. 2015;6:37151–68.
Zhang X, Yuan X, Shi H, Wu L, Qian H, Xu W. Exosomes in cancer: small particle, big player. J Hematol Oncol. 2015;8:83.
Suchorska WM, Lach MS. The role of exosomes in tumor progression and metastasis (Review). Oncol Rep. 2016;35:1237–44.
Guo W, Gao Y, Li N, Shao F, Wang C, Wang P, et al. Exosomes: new players in cancer (Review). Oncol Rep. 2017;38:665–75.
Abak A, Abhari A, Rahimzadeh S. Exosomes in cancer: small vesicular transporters for cancer progression and metastasis, biomarkers in cancer therapeutics. PeerJ. 2018;6:e4763.
Soekmadji C, Russell PJ, Nelson CC. Exosomes in prostate cancer: putting together the pieces of a puzzle. Cancers (Basel). 2013;5:1522–44.
Hessvik NP, Sandvig K, Llorente A. Exosomal miRNAs as biomarkers for prostate cancer. Front Genet. 2013;4:36.
Liu CM, Hsieh CL, Shen CN, Lin CC, Shigemura K, Sung SY. Exosomes from the tumor microenvironment as reciprocal regulators that enhance prostate cancer progression. Int J Urol. 2016;23:734–44.
Frediani JN, Fabbri M. Essential role of miRNAs in orchestrating the biology of the tumor microenvironment. Mol Cancer. 2016;15:42.
Pan J, Ding M, Xu K, Yang C, Mao LJ. Exosomes in diagnosis and therapy of prostate cancer. Oncotarget. 2017;8:97693–700.
Vlaeminck-Guillem. Extracellular vesicles in prostate cancer carcinogenesis, diagnosis, and management. Front Oncol. 2018;8:222.
Zedan AH, Hansen TF, Assenholt J, Pleckaitis M, Madsen JS, Osther PJS. microRNA expression in tumour tissue and plasma in patients with newly diagnosed metastatic prostate cancer. Tumour Biol. 2018;40:1010428318775864.
Kumar B, Rosenberg AZ, Choi SM, Fox-Talbot K, De Marzo AM, Nonn L, et al. Cell-type specific expression of oncogenic and tumor suppressive microRNAs in the human prostate and prostate cancer. Sci Rep. 2018;8:7189.
Kotb S, Mosharafa A, Essawi M, Hassan H, Meshref A, Morsy A. Circulating miRNAs 21 and 221 as biomarkers for early diagnosis of prostate cancer. Tumour Biol. 2014;35:12613–7.
Yang B, Liu Z, Ning H, Zhang K, Pan D, Ding K, et al. MicroRNA-21 in peripheral blood mononuclear cells as a novel biomarker in the diagnosis and prognosis of prostate cancer. Cancer Biomark. 2016;17:223–30.
Malla B, Aebersold DM, Dal Pra A. Protocol for serum exosomal miRNAs analysis in prostate cancer patients treated with radiotherapy. J Transl Med. 2018;16:223.
Foj L, Ferrer F, Serra M, Arévalo A, Gavagnach M, Giménez N, et al. Exosomal and non-exosomal urinary miRNAs in prostate cancer detection and prognosis. Prostate. 2017;77:573–83.
Ghorbanmehr N, Gharbi S, Korsching E, Tavallaei M, Einollahi B, Mowla SJ. miR- 21-5p, miR-141-3p, and miR-205-5p levels in urine-promising biomarkers for the identification of prostate and bladder cancer. Prostate. 2018. https://doi.org/10.1002/pros.23714 (Epub ahead of print).
Wise HM, Hermida MA, Leslie NR. Prostate cancer, PI3K, PTEN and prognosis. Clin Sci (Lond). 2017;131:197–210.
Mishra S, Lin CL, Huang TH, Bouamar H, Sun LZ. MicroRNA-21 inhibits p57Kip2 expression in prostate cancer. Mol Cancer. 2014;13:212.
Lu Z, Liu M, Stribinskis V, Klinge CM, Ramos KS, Colburn NH, et al. MicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 gene. Oncogene. 2008;27:4373–9.
Li T, Li D, Sha J, Sun P, Huang Y. MicroRNA-21 directly targets MARCKS and promotes apoptosis resistance and invasion in prostate cancer cells. Biochem Biophys Res Commun. 2009;383:280–5.
Shi GH, Ye DW, Yao XD, Zhang SL, Dai B, Zhang HL, et al. Involvement of microRNA-21 in mediating chemo-resistance to docetaxel in androgen-independent prostate cancer PC3 cells. Acta Pharmacol Sin. 2010;31:867–73.
Guan Y, Wu Y, Liu Y, Ni J, Nong S. Association of microRNA-21 expression with clinicopathological characteristics and the risk of progression in advanced prostate cancer patients receiving androgen deprivation therapy. Prostate. 2016;76:986–93.
Ribas J, Ni X, Haffner M, Wentzel EA, Salmasi AH, Chowdhury WH, et al. miR-21: an androgen receptor-regulated microRNA that promotes hormone-dependent and hormone-independent prostate cancer growth. Cancer Res. 2009;69:7165–9.
Banyard J, Bielenberg DR. The role of EMT and MET in cancer dissemination. Connect Tissue Res. 2015;56:403–13.
Katsuno Y, Lamouille S, Derynck R. TGF-β signaling and epithelial–mesenchymal transition in cancer progression. Curr Opin Oncol. 2013;25:76–84.
Chowdhury R, Webber JP, Gurney M, Mason MD, Tabi Z, Clayton A. Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro- invasive myofibroblasts. Oncotarget. 2015;6:715–31.
Lee E, Wang J, Yumoto K, Jung Y, Cackowski FC, Decker AM, et al. DNMT1 regulates epithelial–mesenchymal transition and cancer stem cells, which promotes prostate cancer metastasis. Neoplasia. 2016;18:553–66.
Valdez CD, Kunju L, Daignault S, Wojno KJ, Day ML. The E2F1/DNMT1 axis is associated with the development of AR negative castration resistant prostate cancer. Prostate. 2013;73:1776–85.
Cai ZK, Chen Q, Chen YB, Gu M, Zheng DC, Zhou J, et al. microRNA-155 promotes the proliferation of prostate cancer cells by targeting annexin 7. Mol Med Rep. 2015;11:533–8.
Chevrier M, Bobbala D, Villalobos-Hernandez A, Khan MG, Ramanathan S, Saucier C, et al. Expression of SOCS1 and the downstream targets of its putative tumor suppressor functions in prostate cancer. BMC Cancer. 2017;17:157.
Shao N, Ma G, Zhang J, Zhu W. miR-221-5p enhances cell proliferation and metastasis through post-transcriptional regulation of SOCS1 in human prostate cancer. BMC Urol. 2018;18:14.
Kobayashi N, Uemura H, Nagahama K, Okudela K, Furuya M, Ino Y, et al. Identification of miR-30d as a novel prognostic maker of prostate cancer. Oncotarget. 2012;3:1455–71.
Melnik BC. Milk: an epigenetic amplifier of FTO-mediated transcription? Implications for western diseases. J Transl Med. 2015;13:385.
Deng X, Su R, Stanford S, Chen J. Critical enzymatic functions of FTO in obesity and cancer. Front Endocrinol (Lausanne). 2018;9:396.
Su R, Dong L, Li C, Nachtergaele S, Wunderlich M, Qing Y, et al. R-2HG exhibits anti-tumor activity by targeting FTO/m6A/MYC/CEBPA signaling. Cell. 2018;172:90–105.
Martin Carli JF, LeDuc CA, Zhang Y, Stratigopoulos G, Leibel RL. FTO mediates cell-autonomous effects on adipogenesis and adipocyte lipid content by regulating gene expression via 6 mA DNA modifications. J Lipid Res. 2018;59:1446–60.
Yin H, Lowery M, Glass J. In prostate cancer C/EBPalpha promotes cell growth by the loss of interactions with CDK2, CDK4, and E2F and by activation of AKT. Prostate. 2009;69:1001–16.
Zhang J, Gonit M, Salazar MD, Shatnawi A, Shemshedini L, Trumbly R, et al. C/EBPalpha redirects androgen receptor signaling through a unique bimodal interaction. Oncogene. 2010;29:723–38.
Kim MH, Fields J. Translationally regulated C/EBP beta isoform expression upregulates metastatic genes in hormone-independent prostate cancer cells. Prostate. 2008;68:1362–71.
Zha S, Isaacs WB. A nonclassic CCAAT enhancer element binding protein binding site contributes to alpha-methylacyl-CoA racemase expression in prostate cancer. Mol Cancer Res. 2005;3:110–8.
Bernichtein S, Pigat N, Capiod T, Boutillon F, Verkarre V, Camparo P, et al. High milk consumption does not affect prostate tumor progression in two mouse models of benign and neoplastic lesions. PLoS ONE. 2015;10:e0125423.
Tate PL, Bibb R, Larcom LL. Milk stimulates growth of prostate cancer cells in culture. Nutr Cancer. 2011;63:1361–6.
Agarwal MM, Rana SV, Mandal AK, Malhotra S, Khandelwal N, Kumar S, et al. Lactose intolerance in prostate cancer patients: incidence and associated factors. Scand J Gastroenterol. 2008;43:270–6.
Gaard M, Tretli S, Løken EB. Dietary fat and the risk of breast cancer: a prospective study of 25,892 Norwegian women. Int J Cancer. 1995;63:13–7.
Wang F, Yu L, Wang F, Liu L, Guo M, Gao D, et al. Risk factors for breast cancer in women residing in urban and rural areas of eastern China. J Int Med Res. 2015;43:774–89.
Galván-Salazar HR, Arreola-Cruz A, Madrigal-Pérez D, Soriano-Hernández AD, Guzman-Esquivel J, Montes-Galindo DA, et al. Association of milk and meat consumption with the development of breast cancer in a western Mexican population. Breast Care (Basel). 2015;10:393–6.
Ronco AL, De Stéfani E, Dáttoli R. Dairy foods and risk of breast cancer: a case–control study in Montevideo, Uruguay. Eur J Cancer Prev. 2002;11:457–63.
Ji J, Sundquist J, Sundquist K. Lactose intolerance and risk of lung, breast and ovarian cancers: aetiological clues from a population-based study in Sweden. Br J Cancer. 2015;112:149–52.
Missmer SA, Smith-Warner SA, Spiegelman D, Yaun SS, Adami HO, Beeson WL, et al. Meat and dairy food consumption and breast cancer: a pooled analysis of cohort studies. Int J Epidemiol. 2002;31:78–85.
Villagrasa A, Álvarez PJ, Osuna A, Garrido JM, Aránega A, Rodríguez-Serrano F. Exosomes derived from breast cancer cells, small Trojan horses? J Mammary Gland Biol Neoplasia. 2014;19:303–13.
Yu DD, Wu Y, Shen HY, Lv MM, Chen WX, Zhang XH, et al. Exosomes in development, metastasis and drug resistance of breast cancer. Cancer Sci. 2015;106:959–64.
Lowry MC, Gallagher WM, O’Driscoll L. The role of exosomes in breast cancer. Clin Chem. 2015;61:1457–65.
Jia Y, Chen Y, Wang Q, Jayasinghe U, Luo X, Wei Q, et al. Exosome: emerging biomarker in breast cancer. Oncotarget. 2017;8:41717–33.
He Y, Deng F, Yang S, Wang D, Chen X, Zhong S, et al. Exosomal microRNA: a novel biomarker for breast cancer. Biomark Med. 2018;12:177–88.
Hesari A, Golrokh Moghadam SA, Siasi A, Rahmani M, Behboodi N, Rastgar- Moghadam A, et al. Tumor-derived exosomes: potential biomarker or therapeutic target in breast cancer? J Cell Biochem. 2018;119:4236–40.
Howard EW, Yang X. microRNA regulation in estrogen receptor-positive breast cancer and endocrine therapy. Biol Proced Online. 2018;20:17.
Masuda M, Miki Y, Hata S, Takagi K, Sakurai M, Ono K, et al. An induction of microRNA, miR-7 through estrogen treatment in breast carcinoma. J Transl Med. 2012;10(Suppl 1):S2.
Bhat-Nakshatri P, Wang G, Collins NR, Thomson MJ, Geistlinger TR, Carroll JS, et al. Estradiol-regulated microRNAs control estradiol response in breast cancer cells. Nucleic Acids Res. 2009;37:4850–61.
Tao S, He H, Chen Q, Yue W. GPER mediated estradiol reduces miR-148a to promote HLA-G expression in breast cancer. Biochem Biophys Res Commun. 2014;451:74–8.
Schanzenbach CI, Kirchner B, Ulbrich SE, Pfaffl MW. Can milk cell or skim milk miRNAs be used as biomarkers for early pregnancy detection in cattle? PLoS ONE. 2017;12:e0172220.
Corcoran C, Friel AM, Duffy MJ, Crown J, O’Driscoll L. Intracellular and extracellular microRNAs in breast cancer. Clin Chem. 2011;57:18–32.
Donnarumma E, Fiore D, Nappa M, Roscigno G, Adamo A, Iaboni M, et al. Cancer-associated fibroblasts release exosomal microRNAs that dictate an aggressive phenotype in breast cancer. Oncotarget. 2017;8:19592–608.
Au Yeung CL, Co NN, Tsuruga T, Yeung TL, Kwan SY, Leung CS, et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016;7:11150.
Bussard KM, Mutkus L, Stumpf K, Gomez-Manzano C, Marini FC. Tumor-associated stromal cells as key contributors to the tumor microenvironment. Breast Cancer Res. 2016;18:84.
Bayraktar R, Van Roosbroeck K, Calin GA. Cell-to-cell communication: microRNAs as hormones. Mol Oncol. 2017;11:1673–86.
Abrahamsson A, Dabrosin C. Tissue specific expression of extracellular microRNA in human breast cancers and normal human breast tissue in vivo. Oncotarget. 2015;6:22959–69.
Kumar S, Keerthana R, Pazhanimuthu A, Perumal P. Overexpression of circulating miRNA-21 and miRNA-146a in plasma samples of breast cancer patients. Indian J Biochem Biophys. 2013;50:210–4.
Bahrami A, Aledavood A, Anvari K, Hassanian SM, Maftouh M, Yaghobzade A, et al. The prognostic and therapeutic application of microRNAs in breast cancer: tissue and circulating microRNAs. J Cell Physiol. 2018;233:774–86.
Guzman N, Agarwal K, Asthagiri D, Yu L, Saji M, Ringel MD, et al. Breast cancer- specific miR signature unique to extracellular vesicles includes “microRNA-like” tRNA fragments. Mol Cancer Res. 2015;13:891–901.
Wang M, Ji S, Shao G, Zhang J, Zhao K, Wang Z, et al. Effect of exosome biomarkers for diagnosis and prognosis of breast cancer patients. Clin Transl Oncol. 2018;20:906–11.
Hannafon BN, Trigoso YD, Calloway CL, Zhao YD, Lum DH, Welm AL, et al. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016;18:90.
Xu Y, Chao L, Wang J, Sun Y. miRNA-148a regulates the expression of the estrogen receptor through DNMT1-mediated DNA methylation in breast cancer cells. Oncol Lett. 2017;14:4736–40.
Li Q, Ren P, Shi P, Chen Y, Xiang F, Zhang L, et al. MicroRNA-148a promotes apoptosis and suppresses growth of breast cancer cells by targeting B-cell lymphoma 2. Anticancer Drugs. 2017;28:588–95.
Zhou F, Drabsch Y, Dekker TJ, de Vinuesa AG, Li Y, Hawinkels LJ, et al. Nuclear receptor NR4A1 promotes breast cancer invasion and metastasis by activating TGF-β signalling. Nat Commun. 2014;5:3388.
Hedrick E, Safe S. Transforming growth factor β/NR4A1-inducible breast cancer cell migration and epithelial-to-mesenchymal transition is p38α (mitogen-activated protein kinase 14) dependent. Mol Cell Biol. 2017;37(18):e00306–17.
Lin B, Kolluri SK, Lin F, Liu W, Han YH, Cao X, et al. Conversion of Bcl-2 from protector to killer by interaction with nuclear orphan receptor Nur77/TR3. Cell. 2004;116:527–40.
Chang S, Wang RH, Akagi K, Kim KA, Martin BK, Cavallone L, et al. Tumor suppressor BRCA1 epigenetically controls oncogenic microRNA-155. Nat Med. 2011;17:1275–82.
Shen R, Wang Y, Wang CX, Yin M, Liu HL, Chen JP, et al. MiRNA-155 mediates TAM resistance by modulating SOCS6-STAT3 signalling pathway in breast cancer. Am J Transl Res. 2015;7:2115–26.
Gao S, Wang Y, Wang M, Li Z, Zhao Z, Wang RX, et al. MicroRNA-155, induced by FOXP3 through transcriptional repression of BRCA1, is associated with tumor initiation in human breast cancer. Oncotarget. 2017;8:41451–64.
Santos JC, Lima NDS, Sarian LO, Matheu A, Ribeiro ML, Derchain SFM. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci Rep. 2018;8:829.
Jiang S, Zhang HW, Lu MH, He XH, Li Y, Gu H, et al. MicroRNA-155 functions as an OncomiR in breast cancer by targeting the suppressor of cytokine signaling 1 gene. Cancer Res. 2010;70:3119–27.
Sun Q, Chen X, Yu J, Zen K, Zhang CY, Li L. Immune modulatory function of abundant immune-related microRNAs in microvesicles from bovine colostrum. Protein Cell. 2013;4:197–210.
Zu X, Zhang Q, Cao R, Liu J, Zhong J, Wen G, et al. Transforming growth factor-β signaling in tumor initiation, progression and therapy in breast cancer: an update. Cell Tissue Res. 2012;347:73–84.
Alexe G, Reiss M. Transforming growth factor-beta signaling: emerging stem cell target in metastatic breast cancer? Breast Cancer Res Treat. 2009;115:453–95.
Du J, Yu Y, Zhan J, Zhang H. Targeted therapies against growth factor signaling in breast cancer. Adv Exp Med Biol. 2017;1026:125–46.
Li DP, Fan J, Wu YJ, Xie YF, Zha JM, Zhou XM. MiR-155 up-regulated by TGF-β promotes epithelial–mesenchymal transition, invasion and metastasis of human hepatocellular carcinoma cells in vitro. Am J Transl Res. 2017;9:2956–65.
Rong L, Li R, Li S, Luo R. Immunosuppression of breast cancer cells mediated by transforming growth factor-β in exosomes from cancer cells. Oncol Lett. 2016;11:500–4.
Cho JA, Park H, Lim EH, Lee KW. Exosomes from breast cancer cells can convert adipose tissue-derived mesenchymal stem cells into myofibroblast-like cells. Int J Oncol. 2012;40:130–8.
Duarte-Salles T, Fedirko V, Stepien M, Trichopoulou A, Bamia C, Lagiou P, et al. Dairy products and risk of hepatocellular carcinoma: the European Prospective Investigation into Cancer and Nutrition. Int J Cancer. 2014;135:1662–72.
Yuan K, Lian Z, Sun B, Clayton MM, Ng IO, Feitelson MA. Role of miR-148a in hepatitis B associated hepatocellular carcinoma. PLoS ONE. 2012;7:e35331.
Zhang Y, Wei W, Cheng N, Wang K, Li B, Jiang X, et al. Hepatitis C virus-induced up-regulation of microRNA-155 promotes hepatocarcinogenesis by activating Wnt signaling. Hepatology. 2012;56:1631–40.
Yoon JS, Kim G, Lee YR, Park SY, Tak WY, Kweon YO, et al. Clinical significance of microRNA-21 expression in disease progression of patients with hepatocellular carcinoma. Biomark Med. 2018. https://doi.org/10.2217/bmm-2018-0096 (Epub ahead of print).
Pu C, Huang H, Wang Z, Zou W, Lv Y, Zhou Z, et al. Extracellular vesicle-associated mir-21 and mir-144 are markedly elevated in serum of patients with hepatocellular carcinoma. Front Physiol. 2018;9:930.
Liu F, Kong X, Lv L, Gao J. MiR-155 targets TP53INP1 to regulate liver cancer stem cell acquisition and self-renewal. FEBS Lett. 2015;589:500–6.
Okamura S, Arakawa H, Tanaka T, Nakanishi H, Ng CC, Taya Y, et al. p53DINP1, a p53-inducible gene, regulates p53-dependent apoptosis. Mol Cell. 2001;8:85–94.
Seux M, Peuget S, Montero MP, Siret C, Rigot V, Clerc P, et al. TP53INP1 decreases pancreatic cancer cell migration by regulating SPARC expression. Oncogene. 2011;30:3049–61.
Gironella M, Seux M, Xie MJ, Cano C, Tomasini R, Gommeaux J, et al. Tumor protein 53-induced nuclear protein 1 expression is repressed by miR-155, and its restoration inhibits pancreatic tumor development. Proc Natl Acad Sci USA. 2007;104:16170–5.
Gui Y, Khan MGM, Bobbala D, Dubois C, Ramanathan S, Saucier C, et al. Attenuation of MET-mediated migration and invasion in hepatocellular carcinoma cells by SOCS1. World J Gastroenterol. 2017;23:6639–49.
Wang J, Li X, Zhang D. Dairy product consumption and risk of non-Hodgkin lymphoma: a meta-analysis. Nutrients. 2016;8:120.
Eis PS, Tam W, Sun L, Chadburn A, Li Z, Gomez MF, et al. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc Natl Acad Sci USA. 2005;102:3627–32.
Castillo JJ, Beltran BE, Miranda RN, Young KH, Chavez JC, Sotomayor EM. EBV-positive diffuse large B-cell lymphoma, not otherwise specified: 2018 update on diagnosis, risk-stratification and management. Am J Hematol. 2018;93:953–62.
Yin Q, McBride J, Fewell C, Lacey M, Wang X, Lin Z, et al. MicroRNA-155 is an Epstein–Barr virus-induced gene that modulates Epstein–Barr virus-regulated gene expression pathways. J Virol. 2008;82:5295–306.
Higuchi H, Yamakawa N, Imadome KI, Yahata T, Kotaki R, Ogata J, et al. Role of exosomes as a proinflammatory mediator in the development of EBV-associated lymphoma. Blood. 2018;131:2552–67.
Ahmadvand M, Eskandari M, Pashaiefar H, Yaghmaie M, Manoochehrabadi S, Khakpour G, et al. Over expression of circulating miR-155 predicts prognosis in diffuse large B-cell lymphoma. Leuk Res. 2018;70:45–8.
Mottok A, Renné C, Seifert M, Oppermann E, Bechstein W, Hansmann ML, et al. Inactivating SOCS1 mutations are caused by aberrant somatic hypermutation and restricted to a subset of B-cell lymphoma entities. Blood. 2009;114:4503–6.
Schif B, Lennerz JK, Kohler CW, Bentink S, Kreuz M, Melzner I, et al. SOCS1 mutation subtypes predict divergent outcomes in diffuse large B-cell lymphoma (DLBCL) patients. Oncotarget. 2013;4:35–47.
Schuhmacher B, Bein J, Rausch T, Benes V, Tousseyn T, Vornanen M, et al. JUNB, DUSP2, SGK1, SOCS1 and CREBBP are frequently mutated in T-cell/histiocyte rich large B-cell lymphoma. Haematologica. 2018. https://doi.org/10.3324/haematol.2018.203224 (Epub ahead of print).
Li J, Fu R, Yang L, Tu W. miR-21 expression predicts prognosis in diffuse large B-cell lymphoma. Int J Clin Exp Pathol. 2015;8:15019–24.
Wagschal A, Najafi-Shoushtari SH, Wang L, Goedeke L, Sinha S, deLemos AS, et al. Genome-wide identification of microRNAs regulating cholesterol and triglyceride homeostasis. Nat Med. 2015;21:1290–7.
Chen Y, Song YX, Wang ZN. The microRNA-148/152 family: multi-faceted players. Mol Cancer. 2013;12:43.
Ji W, Yang L, Yuan J, Yang L, Zhang M, Qi D, et al. MicroRNA-152 targets DNA methyltransferase 1 in NiS-transformed cells via a feedback mechanism. Carcinogenesis. 2013;34:446–53.
Wang J, Bian Y, Wang Z, Li D, Wang C, Li Q, et al. MicroRNA-152 regulates DNA methyltransferase 1 and is involved in the development and lactation of mammary glands in dairy cows. PLoS ONE. 2014;9:e101358.
Chen Z, Luo J, Sun S, Cao D, Shi H, Loor JJ. miR-148a and miR-17-5p synergistically regulate milk TAG synthesis via PPARGC1A and PPARA in goat mammary epithelial cells. RNA Biol. 2017;14:326–38.
Muroya S, Hagi T, Kimura A, Aso H, Matsuzaki M, Nomura M. Lactogenic hormones alter cellular and extracellular microRNA expression in bovine mammary epithelial cell culture. J Anim Sci Biotechnol. 2016;7:8.
Do DN, Li R, Dudemaine PL, Ibeagha-Awemu EM. MicroRNA roles in signalling during lactation: an insight from differential expression, time course and pathway analyses of deep sequence data. Sci Rep. 2017;7:44605.
Braud M, Magee DA, Park SD, Sonstegard TS, Waters SM, MacHugh DE, et al. Genome-wide microRNA binding site variation between extinct wild aurochs and modern cattle identifies candidate microRNA-regulated domestication genes. Front Genet. 2017;8:3.
Do DN, Dudemaine PL, Li R, Ibeagha-Awemu EM. Co-expression network and pathway analyses reveal important modules of miRNAs regulating milk yield and component traits. Int J Mol Sci. 2017;18:E1560.
Perge P, Nagy Z, Decmann Á, Igaz I, Igaz P. Potential relevance of microRNAs in inter-species epigenetic communication, and implications for disease pathogenesis. RNA Biol. 2017;14:391–401.
Zhao Z, Yu S, Xu M, Li P. Effects of microwave on extracellular vesicles and microRNA in milk. J Dairy Sci. 2018;101:2932–40.
Betker JL, Angle BM, Graner MW, Anchordoquy TJ. The potential of exosomes from cow milk for oral delivery. J Pharm Sci. 2018;3549(18):30726. https://doi.org/10.1016/j.xphs.2018.11.022 (Epub ahead of print).
van Dongen HM, Masoumi N, Witwer KW, Pegtel DM. Extracellular vesicles exploit viral entry routes for cargo delivery. Microbiol Mol Biol Rev. 2016;80:369–86.
Whitley C, Gunst K, Müller H, Funk M, Zur Hausen H, de Villiers EM. Novel replication-competent circular DNA molecules from healthy cattle serum and milk and multiple sclerosis-affected human brain tissue. Genome Announc. 2014;2(4):e00849–914.
Zur Hausen H, de Villiers EM. Dairy cattle serum and milk factors contributing to the risk of colon and breast cancers. Int J Cancer. 2015;137:959–67.
Zur Hausen H, Bund T, de Villiers EM. Infectious agents in bovine red meat and milk and their potential role in cancer and other chronic diseases. Curr Top Microbiol Immunol. 2017;407:83–116.
Falida K, Eilebrecht S, Gunst K, Zur Hausen H, de Villiers EM. Isolation of two virus- like circular DNAs from commercially available milk samples. Genome Announc. 2017;5(17):e00266–317.
Eilebrecht S, Hotz-Wagenblatt A, Sarachaga V, Burk A, Falida K, Chakraborty D, et al. Expression and replication of virus-like circular DNA in human cells. Sci Rep. 2018;8:2851.
Vella LJ, Sharples RA, Lawson VA, Masters CL, Cappai R, Hill AF. Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J Pathol. 2007;211:582–90.
Vella LJ, Greenwood DL, Cappai R, Scheerlinck JP, Hill AF. Enrichment of prion protein in exosomes derived from ovine cerebral spinal fluid. Vet Immunol Immunopathol. 2008;124:385–93.
Bellingham SA, Guo BB, Coleman BM, Hill AF. Exosomes: vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front Physiol. 2012;3:124.
Sardar Sinha M, Ansell-Schultz A, Civitelli L, Hildesjö C, Larsson M, Lannfelt L, et al. Alzheimer’s disease pathology propagation by exosomes containing toxic amyloid-beta oligomers. Acta Neuropathol. 2018;136:41–56.
Cheng L, Zhao W, Hill AF. Exosomes and their role in the intercellular trafficking of normal and disease associated prion proteins. Mol Aspects Med. 2018;60:62–8.
Zempleni J, Sukreet S, Zhou F, Wu D, Mutai E. Milk-derived exosomes and metabolic regulation. Annu Rev Anim Biosci. 2018. https://doi.org/10.1146/annurev-animal-020518-115300 (Epub ahead of print).
Authors’ contributions
BCM performed translational research and wrote the manuscript. GS analyzed the data on exosome microRNA biology and improved the manuscript. Both authors read and approved the final manuscript.
Acknowledgements
The authors thank Dr. Evelyn Osró for providing scientific literature for this manuscript.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data are provided by scientific peer-reviewed publications that are accessible by PubMed.
Consent for publication
Both authors provide their consent for publication.
Ethics approval and consent to participate
Does not apply to this publication.
Funding
The authors declare that this paper is not funded.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Melnik, B.C., Schmitz, G. Exosomes of pasteurized milk: potential pathogens of Western diseases. J Transl Med 17, 3 (2019). https://doi.org/10.1186/s12967-018-1760-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-018-1760-8