
Abstract
Background
Understanding the contribution of outdoor-resting Anopheles mosquitoes to residual malaria transmission is important in terms of scaling up vector control towards malaria elimination in South Africa. The aim of this project was to assess the potential role of Anopheles parensis and other Anopheles species in residual malaria transmission, using sentinel surveillance sites in the uMkhanyakude District of northern KwaZulu-Natal Province.
Methods
Monthly vector surveillance was conducted at the sentinel sites from January 2017 to May 2018. Outdoor-placed clay pot resting traps were used to collect male and female adult Anopheles mosquitoes. All Anopheles gambiae complex and Anopheles funestus group specimens collected were identified to species and all females were screened for Plasmodium falciparum circumsporozoite protein (CSP) by enzyme-linked immunosorbent assay (ELISA). Samples showing infectivity for P. falciparum were further verified by a nested PCR and subsequent DNA sequence analysis.
Results
From a sample of 491 anophelines, Anopheles arabiensis (n = 228) and An. parensis (n = 194) were the most abundant. Other species collected included Anopheles merus (n =11), Anopheles quadriannulatus (n = 10), Anopheles leesoni (n = 29), Anopheles rivulorum (n =18), and Anopheles vaneedeni (n =1). Of the 317 female specimens screened for P. falciparum CSP, one Anopheles arabiensis and one An. parensis showed positive by ELISA and Plasmodium nested PCR. For the An. parensis specimen, confirmation of its species identity was based on sequence analysis of the ITS2 region, and the presence of P. falciparum DNA was further confirmed by sequence analysis.
Conclusions
Anopheles parensis is a potential vector of malaria in South Africa although its contribution to transmission is likely to be minimal at best owing to its strong zoophilic tendency. By contrast, An. arabiensis is a major vector that is primarily responsible for the bulk of residual malaria transmission in South Africa. As all recently collected sporozoite-positive Anopheles mosquitoes were found in outdoor-placed resting traps, it is necessary to introduce interventions that can be used to control outdoor-resting vector populations while maintaining the efficacy of South Africa’s indoor house spraying operations.
Similar content being viewed by others

Background
Malaria in South Africa is endemic in the low altitude northeastern border regions of KwaZulu-Natal, Mpumalanga and Limpopo provinces, and transmission follows a seasonal trend with peaks during the rainy season of November to April. Almost all infections are Plasmodium falciparum [1]. Malaria vector control operations in the affected provinces primarily utilize insecticide-based indoor residual spraying (IRS) with supplementary larviciding in select districts where incidence is highest. These interventions have reduced overall incidence to a point where elimination is a feasible prospect, leading to the development of an elimination strategy [2]. There was however an unexpected and substantial increase in cases in 2017 throughout the southern African region [3]. This increase coupled with ongoing residual transmission, despite control operations in South Africa, has necessitated intensified vector surveillance activities designed to better understand the entomological drivers of transmission [4].
The primary vectors of malaria in South Africa are Anopheles funestus and Anopheles arabiensis [5]. The former species is highly anthropophilic and endophilic, making it especially susceptible to control by IRS. As a result of South Africa’s IRS operations, the incidence of An. funestus has been reduced to a point where it is almost entirely undetectable using a range of surveillance methods [6]. Anopheles arabiensis on the other hand shows variable feeding and resting habits, and will readily rest outdoors making it less amenable to control by IRS. This species has recently been directly incriminated in malaria transmission in South Africa [7] and is considered to be the primary contributor to residual incidence. Added to this list is Anopheles merus, which has been directly implicated in transmission in southern Mozambique [8] and which occurs in South Africa in all malarious provinces [9, 10]. Anopheles vaneedeni, an outdoor resting member of the An. funestus group [11], has recently been incriminated as a secondary vector of malaria in South Africa and a likely contributor to residual transmission [12].
Members of the An. funestus group that have been incriminated in malaria transmission in other African countries are Anopheles rivulorum, Anopheles leesoni and Anopheles parensis (tentative) in Tanzania [13,14,15], Anopheles rivulorum in Kenya [16], and Anopheles longipalpis in Kenya [17]. Anopheles parensis periodically appears as a potential vector. This species is predominantly zoophilic and rarely takes blood from humans even though some populations have shown a strong inclination to rest indoors [18, 19]. A single positive An. parensis (1 out of 4 specimens sampled) using nested PCR was detected in Bagamoya, Tanzania [14]. Samples of An. parensis from Uganda gave a P. falciparum infection rate of 4.2% (n = 94) using a Taqman assay but this was not confirmed by nested PCR, suggesting the detection of false positives [20]. Tentative evidence of malaria infectivity by this species was obtained from northern KwaZulu-Natal Province in South Africa based on a sample of 149 specimens of which 20 (13.4%) showed positive using the standard ELISA assay [19] for P. falciparum circumsporozoite protein. Follow-up PCR analysis however did not reveal any positives and it was subsequently shown that all ELISA positive specimens had taken blood from non-human sources [19]. The status of An. parensis as a malaria vector, therefore, remains to be confirmed.
Understanding the contribution of outdoor-resting anophelines to residual malaria transmission is important in terms of scaling up vector control toward malaria elimination in South Africa. The aim of this project was therefore to assess the potential role of An. parensis and other Anopheles species in residual malaria transmission using sentinel sites in the uMkhanyakude District of northern KwaZulu-Natal Province, which currently experiences very low malaria incidence and is the closest of all of South Africa’s malaria-endemic districts to achieving elimination status.
Methods
Entomological surveillance
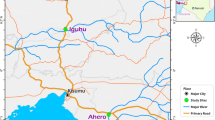
Monthly entomological surveillance was conducted for just over 1 year—January 2017 to May 2018—at sentinel sites in the rural village of Mamfene, in the uMkhanyakude District of KwaZulu-Natal Province, South Africa (Fig. 1). Traditional clay pots (Fig. 2) were deployed outside households (with the home-owners’ consent) to serve as resting traps from which live Anopheles mosquitoes were collected in the cool early hours of each morning (between 06:00 and 08:00). Five households were recruited for this purpose and two pots were deployed in each (n = 10 pots). All pots were cleared of mosquitoes for 3 days each month during the surveillance period, except for June 2017 and February 2018. There were no collections during those two months owing to logistical constraints.

a Location of the field site at Mamfene (yellow pin circled in red), KwaZulu-Natal Province, South Africa. Map data © 2018 AfriGIS (Pty) Ltd, Google. b Spatial distribution of 10 clay pots (blue pins) deployed as outdoor-placed resting traps at the field site in KwaZulu-Natal Province. Image © 2018 DigitalGlobe, Google Earth © Google 2018

An example of the locally-sourced clay pots that were used as outdoor-placed resting traps for Anopheles mosquito collections in KwaZulu-Natal Province, South Africa
Molecular identification
All specimens were identified by external morphology using dichotomous keys [21]. Those identified as Anopheles gambiae complex or An. funestus group were dry-preserved on silica gel and transported to the Vector Control Reference Laboratory of the National Institute for Communicable Diseases in Johannesburg for further processing. Each specimen was identified to species using the appropriate polymerase chain reaction (PCR) protocol for either An. gambiae complex [22] or An. funestus group [23, 24].
Plasmodium infectivity testing
Every female anopheline mosquito was screened for the presence of P. falciparum circumsporozoite protein (CSP) by means of an enzyme-linked immunosorbent assay (ELISA) [25, 26]. The ELISA homogenates of all positive specimens were then boiled at 100 °C for 10 min followed by a repeat ELISA to confirm or refute the presence of CSP [27]. Those that remained ELISA-positive were verified by a nested Plasmodium PCR assay [28]. The internal transcribed spacer 2 (ITS2) region of an anopheline female showing positive for a P. falciparum infection was amplified by ITS2 PCR assay [23] to be used for DNA sequence analysis to verify the species identification.
DNA sequence analysis
Sequencing analysis of the ITS2 and the nested Plasmodium PCR products was used to confirm the anopheline species identity and infection with P. falciparum. Purification and sequencing of the PCR products was performed by Macrogen. Subsequently, the chromatograms of the sequences were manually edited by BioEdit version 7.2.5 [39]. The Emboss Needle pairwise sequence alignment tool (https://www.ebi.ac.uk/Tools/psa/emboss_needle/nucleotide.html) was used to compare the analysed An. parensis ITS2 sequence with the established An. parensis ITS2 reference sequences (GenBank accession number JN994144.1) [29]. The unknown Plasmodium sequence was compared with the reference P. falciparum Pf8 18S ribosomal RNA gene, partial sequence (GenBank accession number: KC428742.1) [30].
Results
Entomological surveillance
Adult Anopheles mosquitoes were collected from outdoor-resting traps deployed around households in Mamfene, northern KwaZulu-Natal Province (Figs. 1 and 2), during the period January 2017 to May 2018. A total of 491 male and female specimens were identified as either An. gambiae complex or An. funestus group using morphological features [21]. These were subsequently identified to species using the standard PCR assays for each group [22,23,24]. Anopheles arabiensis, a member of the An. gambiae complex, was the most abundant (n = 228). Other members of the An. gambiae complex included An. merus (n = 11) and An. quadriannulatus (n = 10) (Fig. 3). Within the An. funestus group, An. parensis was most abundant (n = 194) followed by An. leesoni (n = 29), An. rivulorum (n = 18) and An. vaneedeni (n = 1) (Fig. 3). There was an overall preponderance of females across all species collected (F = 240.58, df = 1, P < 0.004); 163 females and 86 males of the An. gambiae complex, and 154 females and 88 males of the An. funestus group (Fig. 3). The monthly and seasonal distribution of collected anophelines showed a significant increase in mosquito numbers in March 2017 (Fig. 4.). This increase in relative mosquito population density is likely attributable to high rainfall experienced in Mamfene from February to March 2017.

Anopheles mosquitoes by species group and gender collected using outdoor-placed clay pots deployed as resting traps at a field site at Mamfene, KwaZulu-Natal Province, South Africa, from January 2017 to May 2018

Monthly sampling of male and female Anopheles by species group/complex in Mamfene, KwaZulu-Natal Province, South Africa, from January 2017 to May 2018. Mosquitoes were collected over a 3-day period each month. No mosquitoes were collected for June 2017 and February 2018 due to logistical constraints
Vector incrimination
All female mosquitoes were screened for P. falciparum CSP by a two-step ELISA [25,26,27] and were subsequently validated by nested PCR and DNA sequence analysis where necessary. The majority of positive samples from the first ELISA assay did not appear positive on the second that included a boiling step. Of the 163 female An. gambiae complex specimens screened for CSP, one An. arabiensis (collected in October 2017) showed positive by ELISA and Plasmodium nested PCR [28]. As this An. arabiensis population has been implicated in malaria transmission before [7], no further DNA sequence analysis was considered necessary. Of the 154 female An. funestus group specimens screened, one female identified as An. parensis (also collected in October 2017) showed positive for P. falciparum CSP by ELISA and nested Plasmodium PCR. These data indicate a P. falciparum infectivity rate of 0.67% (n = 149) for An. arabiensis and 0.83% (n = 120) for An. parensis at Mamfene.
Validation of Anopheles species identity and the presence of a Plasmodium infection
The species identity of the infective An. parensis female was confirmed after standard PCR [23] ITS2 sequences showed 99% homology between this specimen and that for a reference sequence of this species in GenBank (accession number: JN994144.1) [25]. The P. falciparum infection in this An. parensis specimen was confirmed by 95% sequence homology of the nested Plasmodium PCR product with the P. falciparum isolate Pf8 18S ribosomal RNA gene, partial sequence (GenBank accession number: KC428742.1) [30].
Discussion
The morphological and molecular taxonomy of the An. funestus group is comparatively complex [11], making identification of member species potentially problematic. The need for morphological identification prior to molecular analysis has been highlighted [31], and is especially critical in terms of follow-on vector incrimination. This is because Anopheles species misidentification can easily occur when entirely reliant on nested PCR assays, and sequence information can be misleading in terms of the critical thresholds required for conspecific homology between unknown samples and banked sequences linked to voucher specimens. In addition to the complexities of Anopheles species identification, establishing Plasmodium infectivity (as distinct from infection) also requires careful use of methodologies and data interpretation. It is for this reason that the two-step ELISA assays for CSP detection were conducted prior to molecular verification for the An. parensis samples. Given that An. parensis has not been unambiguously incriminated in malaria transmission before, it was considered necessary to ensure that all morphological, PCR, ELISA and sequence data were sufficiently aligned to confirm Plasmodium infectivity, and that all data were either double or triple-checked for quality assurance. A similar approach was recently conducted for the incrimination of An. vaneedeni (also a member of the An. funestus group) as a secondary malaria vector in South Africa [12].
It should be noted that the single ELISA technique used to detect Plasmodium sporozoites can lack sensitivity and specificity. It is however more likely to give a false positive than a false negative result, especially in zoophilic species such as An. parensis [19, 27]. The false positive ELISA result that can occur in zoophilic Anopheles species is due to a cross-reacting heat-unstable antigen [27]. Heating the ELISA lysate, as was done in the experiments described here, reduces the possibility of obtaining false positives. Additionally, the use of nested PCR for the detection of Plasmodium sporozoites is a highly sensitive method [28] that further eliminates the chance of false positives. Although Taqman assays [32, 33] are also effective for detecting Plasmodium in anophelines, they can be prohibitively expensive. Owing to South Africa’s comparatively low malaria incidence and the general rarity of Plasmodium infective mosquitoes, the two-step ELISA method followed by nested PCR is the most cost-effective and ideal method for detecting and confirming Plasmodium infectivity.
It is reasonable to infer that the single An. parensis female that presented with P. falciparum CSP was infective for malaria. This at least shows that this species is a potential malaria vector. There are however other considerations that need to be taken into account in terms of its contribution to malaria transmission. Available data show that An. parensis seldom takes blood from humans, being more inclined to feed on livestock animals and rest outdoors [34, 35], as has previously been demonstrated for the northern KwaZulu-Natal population [19]. This reduces its potential contribution to malaria transmission substantially, but not necessarily to zero. This is reinforced by earlier records that describe a high malaria incidence setting on the Kenyan coast in which An. parensis was abundant but no sporozoite positive specimens were found (based on salivary gland dissections) despite the sympatric An. funestus population showing a sporozoite positive index of 7% [35]. Anopheles parensis has nevertheless been recorded as an exophilic feeder on humans [35] with a comparatively high human blood index in some ecosystems [34, 36], and can be found resting both indoors and outdoors [37].
Anopheles arabiensis is a major malaria vector throughout its distribution including northern KwaZulu-Natal [7, 38]. It is worth noting that both the infective An. arabiensis and An. parensis specimens were collected in October 2017. Although October is generally a low-incidence month in South Africa, 2017 was an unusual year in that unexpectedly high numbers of cases were recorded from May onwards (unpublished National Department of Health statistics). Incidence in KwaZulu-Natal was nevertheless low compared to other endemic provinces which precludes associating these sporozoite-positive specimens with any case or cluster of cases. Also important to note is that both of these specimens were caught resting outdoors, highlighting the importance of addressing residual transmission in South Africa by targeting outdoor-resting vectors in addition to those targeted indoors by the provincial IRS programmes. Intensive provincial larval source management programmes, including winter larviciding, and community outreach programmes designed to educate on personal protection measures and treatment-seeking, are currently under development to address this issue.
Conclusions
It is concluded that An. parensis is a potential vector of malaria but that its contribution to transmission in South Africa’s endemic provinces is likely to be minimal at best owing to its strong zoophilic tendency [19]. By contrast, An. arabiensis is a major vector throughout its distribution and is primarily responsible for the bulk of residual malaria transmission in South Africa. As all recently collected sporozoite-positive Anopheles mosquitoes were found in outdoor-placed resting traps in northern KwaZulu-Natal Province [7, 12], it is necessary to introduce additional interventions, intensive larval source management in particular, that can be used to control outdoor-resting vector populations while maintaining the efficacy of South Africa’s IRS operations for the ongoing control of indoor-resting vectors.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request. All data presented in this study is stored in the malaria vector database of the Vector Control Reference Laboratory, National Institute for Communicable Diseases, and is available on request.
Abbreviations
- CSP:
-
circumsporozoite protein
- DNA:
-
deoxyribonucleic acid
- ELISA:
-
enzyme-linked immunosorbent assay
- IRS:
-
indoor residual spraying
- ITS2:
-
internal transcribed spacer 2
- PCR:
-
polymerase chain reaction
- RNA:
-
ribonucleic acid
References
Maharaj R, Raman J, Morris N, Moonasar D, Durrheim DN, Seocharan I, et al. Epidemiology of malaria in South Africa: from control to elimination. S Afr Med J. 2013;103(10 Suppl 2):779–83.
Raman J, Morris N, Frean J, Brooke B, Blumberg L, Kruger P, et al. Reviewing South Africa’s malaria elimination strategy (2012–2018): progress, challenges and priorities. Malar J. 2016;15:438.
World Health Organization. World malaria report. Geneva: World Health Organization; 2018.
Christian R, Dahan-Moss Y, Braack L, Munhenga G, Kaiser M, Lobb L, et al. Malaria vector surveillance report, South Africa, January–December, 2017. NICD Bulletin. 2018;6:1.
Brooke BD, Koekemoer L, Kruger P, Urbach J, Misiani E, Coetzee M. Malaria vector control in South Africa. S Afr Med J. 2013;103:784–8.
Coetzee M, Hunt RH, Wilkerson R, Della Torre A, Coulibaly MB, Besanksky NJ. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa. 2013;3619:246.
Dandalo LC, Brooke BD, Munhenga G, Lobb LN, Zikhali J, Ngxongo SP, et al. Population dynamics and Plasmodium falciparum (Haemosporida: Plasmodiidae) infectivity rates for the malaria vector Anopheles arabiensis (Diptera: Culicidae) at Mamfene, KwaZulu-Natal. J Med Entomol. 2017;54(6):1758–66.
Cuamba N, Mendis C. The role of Anopheles merus in malaria transmission in an area of southern Mozambique. J Vector Borne Dis. 2009;46:157–9.
Mbokazi F, Coetzee M, Brooke B, Govere J, Reid A, Owiti P, et al. Changing distribution and abundance of the malaria vector Anopheles merus in Mpumalanga Province, South Africa. Public Health Action. 2018;8(Suppl 1):S39–43.
Coetzee M, Craig M, Le Sueur D. Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex. Parasitol Today. 2000;16:74–7.
Coetzee M, Koekemoer LL. Molecular systematics and insecticide resistance in the major African malaria vector Anopheles funestus. Annu Rev Entomol. 2013;58:393–412.
Burke A, Dandalo L, Munhenga G, Dahan-Moss Y, Mbokazi F, Ngxongo S, et al. A new malaria vector mosquito in South Africa. Sci Rep. 2017;7:43779.
Wilkes TJ, Matola YG, Charlwood JD. Anopheles rivulorum, a vector of human malaria in Africa. Med Vet Entomol. 1996;10:108–10.
Temu EA, Minjas JN, Tuno N, Kawada H, Takagi M. Identification of four members of the Anopheles funestus (Diptera: Culicidae) group and their role in Plasmodium falciparum transmission in Bagamoyo coastal Tanzania. Acta Trop. 2007;102:119–25.
Kyalo D, Amratia P, Mundia CW, Mbogo CM, Coetzee M, Snow RW. A geo-coded inventory of anophelines in the Afrotropical Region south of the Sahara: 1898–2016. Wellcome Open Res. 2017;2:57.
Kawada H, Dida GO, Sonye G, Njenga SM, Mwandawiro C, Minakawa N. Reconsideration of Anopheles rivulorum as a vector of Plasmodium falciparum in western Kenya: some evidence from biting time, blood preference, sporozoite positive rate, and pyrethroid resistance. Parasit Vectors. 2012;5:230.
Ogola EO, Fillinger U, Ondiba IM, Villinger J, Masiga DK, Torto B, et al. Insights into malaria transmission among Anopheles funestus mosquitoes, Kenya. Parasit Vectors. 2018;11:577.
Kamau L, Koekemoer LL, Hunt RH, Coetzee M. Anopheles parensis: the main member of the Anopheles funestus species group found resting inside human dwellings in Mwea area of central Kenya toward the end of the rainy season. J Am Mosq Control Assoc. 2003;19:130–3.
Mouatcho JC, Hargreaves K, Koekemoer LL, Brooke BD, Oliver SV, Hunt RH, et al. Indoor collections of the Anopheles funestus group (Diptera: Culicidae) in sprayed houses in northern KwaZulu-Natal, South Africa. Malar J. 2007;6:30.
Mulamba JC, Hargreaves K, Koekemoer LL, Brooke BD, Oliver SV, Hunt RH, et al. Contrasting Plasmodium infection rates and insecticide susceptibility profiles between the sympatric sibling species Anopheles parensis and Anopheles funestus s.s.: a potential challenge for malaria vector control in Uganda. Parasit Vectors. 2014;7:71.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa South of the Sahara (Afrotropical region). Johannesburg: South African Institute for Medical Research; 1987.
Scott JA, Brogdon WG, Collins FH. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993;49:520–9.
Koekemoer LL, Kamau L, Hunt RH, Coetzee M. A cocktail polymerase chain reaction assay to identify members of the Anopheles funestus (diptera: culicidae) group. Am J Trop Med Hyg. 2002;6:804–11.
Cohuet A, Simard F, Toto JC, Kengne P, Coetzee M, Fontenille D. Species identification within the Anopheles funestus group of malaria vectors in Cameroon and evidence for a new species. Am J Trop Med Hyg. 2003;69:200–5.
Wirtz RA, Zavala F, Charoenvit Y, Campbell GH, Burkot TR, Schneider I, et al. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull World Health Organ. 1987;65:39–45.
Burkot TR, Williams JL, Schneider I. Identification of Plasmodium falciparum-infected mosquitoes by a double antibody enzyme-linked immunosorbent assay. Am J Trop Med Hyg. 1984;33:783–8.
Durnez L, Van Bortel W, Denis L, Roelants P, Veracx A, Trung HD, et al. False positive circumsporozoite protein ELISA: a challenge for the estimation of the entomological inoculation rate of malaria and for vector incrimination. Malar J. 2011;10:195.
Snounou G, Viriyakosol S, Zhu XP, Jarra W, Pinheiro L, Rosario VE, et al. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol. 1993;6:315–20.
Norris LC, Norris DE. Phylogeny of anopheline (Diptera: Culicidae) species in southern Africa, based on nuclear and mitochondrial genes. J Vector Ecol. 2015;40:16–27.
Ngassa Mbenda HG, Das A. Molecular evidence of Plasmodium vivax mono and mixed malaria parasite infections in Duffy-negative native Cameroonians. PLoS One. 2014;9:e103262.
Erlank E, Koekemoer LL, Coetzee M. The importance of morphological identification of African anopheline mosquitoes (Diptera: Culicidae) for malaria control programmes. Malar J. 2018;17:43.
Bass C, Nikou D, Blagborough AM, Vontas J, Sinden RE, Martin S, Williamson MS, et al. PCR-based detection of Plasmodium in Anopheles mosquitoes: a comparison of a new high-throughput assay with existing methods. Malar J. 2008;7:177.
Kefi M, Mavridis K, Simões ML, Dimopoulos G, Siden-Kiamos I, Vontas J. New rapid one-step PCR diagnostic assay for Plasmodium falciparum infective mosquitoes. Sci Rep. 2018;8:1462.
Muturi EJ, Kamau L, Jacob BG, Muriu S, Mbogo CM, Shililu J, et al. Spatial distribution, blood feeding pattern, and role of Anopheles funestus complex in malaria transmission in central Kenya. Parasitol Res. 2009;105:1041–6.
Gillies MT, De Meillon B. The Anophelinae of Africa south of the Sahara (Ethiopian zoogeographical region). Johannesburg: South African Institute for Medical Research; 1968.
Kweka EJ, Mahande AM, Nkya WM, Assenga C, Lyatuu EE, Nyale E, et al. Vector species composition and malaria infectivity rates in Mkuzi, Muheza District, north-eastern Tanzania. Tanzan J Health Res. 2008;10:46–9.
Kamau L, Munyekenye GO, Koekemoer LL, Hunt RH, Coetzee M. A survey of the Anopheles funestus (Diptera: Culicidae) group of mosquitoes from 10 sites in Kenya with special emphasis on population genetic structure based on chromosomal inversion karyotypes. J Med Entomol. 2003;40:664–71.
Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, et al. A global map of dominant malaria vectors. Parasit Vectors. 2012;5:69.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–8.
Acknowledgements
Dr. Givemore Munhenga, Dr. Maria Kaiser, Mrs. Leanne Lobb, Mr. Oliver Wood, Dr. Power Tshikae, Mr. Jabulani Zhikali and Mr. Dumisani Dlamini are thanked for their support of and assistance with the collection of specimens.
Funding
These activities were sponsored by the National Institute for Communicable Diseases, a CDC/GDD (Global Diseases Detection programme) grant (U19GH000622-01 MAL01), the DST/NRF South African Research Chair Initiative to MC and LK, the International Atomic Energy Agency (Research Contract No. 17904; 19099 and SAF 5013/5014), the Industrial Development Corporation and the South African Nuclear Energy Corporation (NECSA) through its Nuclear Technologies in Medicine Biosciences Initiative (NTeMBI)—a national platform funded by the Department of Science and Technology.
Author information
Authors and Affiliations
Contributions
AB collected and analysed the data, conducted the laboratory analysis of material, and assisted with writing the manuscript. YD-M interpreted the DNA sequence analysis results and assisted with writing the manuscript. BB, FD, MC, BQ and LK conceived the study, assisted with data interpretation and the drafting of the manuscript. BB interpreted the data and produced the final version of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
An ethics approval waiver was obtained from the animal ethics screening committee of the University of the Witwatersrand, Johannesburg, South Africa. Reference: A Burke Waiver (2) 24-02-2017-O. Informed consent was obtained from all home-owners affected by the surveillance portion of this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Burke, A., Dahan-Moss, Y., Duncan, F. et al. Anopheles parensis contributes to residual malaria transmission in South Africa. Malar J 18, 257 (2019). https://doi.org/10.1186/s12936-019-2889-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-019-2889-5