
Abstract
Background
Root-to-shoot translocation of zinc (Zn) and cadmium (Cd) depends on the concentrations of both metals in the medium. A previous study on tobacco (Nicotiana tabacum) pointed to the contribution of NtZIP1, NtZIP2, NtZIP4 and NtIRT1-like in the regulation of this phenomenon. To learn more, Zn and Cd accumulation, root/shoot distribution and the expression of ZIP genes were investigated in the apical, middle and basal root parts.
Results
We show that Zn/Cd status-dependent root-shoot distribution of both metals was related to distinct metal accumulation in root parts. At low Zn and Cd in the medium, the apical part contained the highest metal level; at higher concentrations, the middle and basal parts were the major sink for excess metal. The above were accompanied by root part-specific expression pattern modifications of ZIPs (NtZIP1-like, NtZIP2, NtZIP4A/B, NtZIP5A/B, NtZIP5-like, NtZIP8, NtZIP11, NtIRT1, and NtIRT1-like) that fell into four categories with respect to the root part. Furthermore, for lower Zn/Cd concentrations changes were noted for NtZIP5A/B and NtZIP5-like only, but at higher Zn and Cd levels for NtZIP1-like, NtZIP5-like, NtZIP8, NtZIP11, NtIRT1, and NtIRT1-like. NtZIP1, here renamed to NtZIP5B, was cloned and characterized. We found that it was a zinc deficiency-inducible transporter involved in zinc and cadmium uptake from the soil solution primarily by the middle root part.
Conclusions
We conclude that regulation of the longitudinal distribution of Zn and Cd is highly specific, and that the apical, middle and basal root parts play distinct roles in Zn/Cd status-dependent control of metal translocation efficiency to shoots, including the stimulation of Zn translocation to shoots in the presence of Cd. These results provide new insight into the root part-specific unique role of NtZIP5B and other ZIP genes in the longitudinal distribution of zinc and cadmium and their contribution to the regulation of root-to-shoot translocation.
Similar content being viewed by others


Background
Tobacco is known to accumulate metals (including Zn and Cd) in aerial parts more efficiently than many other species [1, 2]. It has, therefore, been frequently used for phytoextraction purposes [3, 4]. On the other hand, these characteristics put smokers at risk of being exposed to cadmium accumulated in shoots. Thus, in-depth knowledge of the molecular mechanisms governing the rate of translocation of metals to shoots might generate future biotechnology-based modifications in Zn/Cd accumulation in the aerial parts of tobacco plants.
The regulation pathways of Zn homeostasis are not uniquely specific for this metal. Conversely, its regulation via common pathways is closely related to the homeostasis of other metals, as well as some toxic metals including Cd. This has been termed metal cross-homeostasis. It is based on one metal transport gene being regulated by more than one metal, and on metal transport proteins or chelating compounds having diverse substrate specificities [5,6,7]. There are strong competitive interactions between Zn and Cd. At the whole plant level these relationships are manifested, among others, by changes in Zn and Cd uptake and translocation to shoots in a manner dependent on the concentration of both metals in the medium [8, 9]. For example, excess Zn affects the uptake and root/shoot distribution of Cd. In A. halleri or T. caerulescens (current name, Noccaea caerulescens), Cd transfer to shoots was competitively inhibited by Zn [10, 11]. The presence of Cd in the medium/soil also perturbs the uptake and translocation of Zn. Reduction of Zn transfer to shoots in the presence of Cd was reported, e.g., in N. rustica and T. caerulescence [12, 13], however, no changes were observed in tumbleweed or in A. thaliana [14, 15].
Despite their common occurrence, the mechanisms underlying this phenomenon have been only fragmentarily depicted. To learn more about the regulation of Zn/Cd supply-dependent root-to-shoot translocation of these metals in tobacco, the aim of this study was to examine the possible contribution of ZIP genes (encoding proteins belonging to the family of ZRT\IRT-related proteins) to the root/shoot distribution of both metals under different metal concentrations in the medium. The focus on ZIP genes results from SSH (Suppression Subtractive Hybridization)-based identification of tobacco NtZIP1, NtZIP2, NtZIP4 and IRT1-like as genes having different expression under combinations of low to high Zn/Cd concentrations in the medium. This accompanied changes in the root/shoot partitioning of both metals [9]. Proteins from the ZIP family mediate the transport of a range of metals, including Zn and Cd, to the cytoplasm either from the extracellular space or from internal stores [16]. In Arabidopsis, AtZIP1 (localized to the tonoplast) is involved in Zn and Mn transport, and Zn- and Fe-deficiency upregulated its expression [17]. The ability to transport Cd was detected, e.g., for MtZIP1 [18] and OsZIP1 [19] but not for the new tobacco NtZIP1-like and NtZIP11 genes encoding a Zn- (but not Cd- or Fe-) uptake protein [20, 21]. AtZIP2 from A. thaliana and MtZIP2 from Medicago truncatula are Zn uptake proteins localized to the plasma membrane, although AtZIP2 is also able to transport Mn, Cu, and potentially Cd [17, 22,23,24]. For comparison, the ability to transport Zn and Cd has also been shown for NtZIP4 [25], MnZIP4 (from Morus notabilis) [26], AtIRT1 [27, 28], and PsIRT1 (from Pisum sativum) [29].
In this study, we focused on root-specific processes associated with changes in Zn and Cd root/shoot partitioning and in the expression pattern of ZIP genes that depended on combinations of low and high concentrations of both metals in the medium. For the first time, analysis was performed not on whole roots, but on root parts (apical, middle, basal). Here, nine ZIP genes (NtZIP1-like, NtZIP2, NtZIP4A/B, NtZIP5A/B, NtZIP5-like, NtZIP8, NtZIP11, NtIRT1, and NtIRT1-like) were included in the analysis. Furthermore, as the result of detailed analysis NtZIP1 was renamed NtZIP5, and two copies, NtZIP5A and NtZIP5B, were identified. NtZIP5B was cloned and functionally characterized. Our results provide new insights into the role of three root parts in Zn/Cd status-dependent accumulation of Zn and Cd and into the expression of ZIP genes associated with distinct Zn and Cd root/shoot partition patterns.
Results
Zn/Cd supply-dependent root/shoot metal distribution
The aim of this study was to determine to what extent the levels of Zn and Cd in the medium affect the root-to-shoot translocation of both metals. It was shown that in plants exposed to a range of Zn (0; 1; 5; 10; 50 μM) and Cd (0; 0.1; 0.25; 1; 4 μM) concentrations for 17 days, Zn/Cd accumulation and translocation efficiency depended on the pairwise combination of both metals (Figs. 1-2; Additional file 1).

Cadmium concentration in plants grown under various Zn and Cd concentrations. 3.5-week old plants grown in the quarter-strength Knop’s medium were exposed for 17 days to the control medium supplemented with pairwise combinations of Zn (0; 1; 5; 10; 50 μM) and Cd (0; 0.25; 1; 4 μM) concentrations. Concentrations of metals were determined in the shoots (a), roots (b) and whole plants (c). The Translocation Factor (TF) was determined as the ratio of shoot/root concentrations of Cd (d). Concentrations of Cd were not measured for plants grown at the combination 4 μM Cd + 50 μM Zn since it was too toxic. Inserts in the (a, b, c) represent parts of the graph with decreased scale at the Y axis. Values correspond to arithmetic means ±SD (n = 10). Significance of differences between all values were evaluated by Student’s t-test (P ≤ 0.05), and it has been determined that each pair of values is significantly different if the SD bars do not overlap. Significance has not been marked above the bars with the letters or arrows to keep the clarity of the graph

Zinc concentration in plants grown under various Zn and Cd concentrations. 3.5-week old plants grown in the quarter-strength Knop’s medium were exposed for 17 days to the control medium supplemented with pairwise combinations of Zn (0; 1; 5; 10; 50 μM) and Cd (0; 0.25; 1; 4 μM) concentrations. Concentrations of metals were determined in the shoots (a), roots (b) and whole plants (c). The Translocation Factor (TF) was determined as the ratio of shoot/root concentrations of Zn (d). Concentrations of Zn were not measured for plants grown at the combination 4 μM Cd + 50 μM Zn since it was too toxic. Inserts in the (a, b, c) represent parts of the graph with decreased scale at the Y axis. Values correspond to arithmetic means ±SD (n = 10). Significance of differences between all values were evaluated by Student’s t-test (P ≤ 0.05), and it has been determined that each pair of values is significantly different if the SD bars do not overlap. Significance has not been marked above the bars with the letters or arrows to keep the clarity of the graph
As the Cd concentration in the medium increased (regardless of the accompanying Zn level), the enhanced concentration of Cd in plants (Fig. 1a-c) was accompanied by decreased efficiency of root-to-shoot translocation (Fig. 1d). Concentrations of Cd in the shoots of plants grown at each tested Cd level were modified by accompanying Zn concentrations (Fig. 1a, b). Interestingly, at 0 Zn/4 μM Cd, the concentration of Cd in the shoots was 3- to 4-times lower compared with variants containing Zn (Fig. 1a); this did not result from a decreased translocation rate (Fig. 1d), but likely from lower Cd uptake in the absence of Zn under Zn deficiency (Fig. 1c). Moreover, the presence of 50 μM Zn reduced Cd translocation (Fig. 1d), and the accompanying high level of Cd in roots (Fig. 1b) indicated enhanced retention of Cd in this organ.
The presence of Cd also modified Zn shoot/root distribution (Fig. 2a-d). The pattern, however, was not uniform. In the presence of high (4 μM) Cd, Zn translocation to shoots was reduced (relative to the medium without Cd), however, only for plants exposed to 5–50 μM Zn. Notably, the opposite effect, an increase in Zn translocation in the presence of 0.25 and 1 μM Cd was observed in plants grown at low/medium Zn concentrations (0 to 1 μM) (Fig. 2d).
The effects of long-term exposure of zinc and cadmium on Cd-dependent stimulation of Zn translocation to shoots
Next, the study focused on Zn/Cd combinations at which Cd-dependent stimulation of Zn root-to-shoot translocation was detected (0 Zn + 0.25 μM Cd; 1 μM Zn + 0.25 μM Cd; 1 μM Zn + 1 μM Cd vs medium without Cd). The efficiency of retention of a metal in roots is one of the critical factors in controlling the rate of its root-to-shoot transfer. Depending on the combination of Zn/Cd concentrations in the medium, different root parts dominated in the accumulation of Zn and Cd. Under Zn deficiency the distribution of Zn among the apical, middle and basal root parts of plants grown without Cd and in the presence of 0.25 μM Cd was similar, with the highest Zn concentration found in the apical parts (Fig. 3a1). The Zn distribution changed at higher Zn and Cd status, and at 1 μM Zn + 1 μM Cd the middle and basal parts became the major root storage sites for both Zn and Cd (Fig. 3a2, b2). At lower Zn/Cd concentrations in the medium (0 Zn + 0.25 μM Cd), accumulation of Cd remained at the same level in three root parts (Fig. 3b1). These results point to the importance of the apical parts in Zn absorption under Zn deficiency, while at higher metal concentrations the basal part contributes more efficiently to metal accumulation.

Zinc and cadmium concentration in the apical, middle and basal parts of roots from plants grown under various Zn and Cd concentrations. 3.5-week old plants grown in the quarter-strength Knop’s medium were exposed to the control medium supplemented with pairwise combinations of Zn (0; 1 μM) and Cd (0; 0.25; 1 μM) concentrations for 17 days. Concentrations of metals were measured in the apical parts (2.5 cm) from the main and lateral roots; basal (proximal) parts (1/4 of the total length of the main roots); middle part (remaining parts of the main roots). The basal and the middle parts of the roots were collected from the main root only (adventitious roots were excised). Zinc concentration (a): at Zn-deficiency (a1), at 1 μM Zn (a2). Cadmium concentration (b): at Zn-deficiency (b1), at 1 μM Zn (b2). Values correspond to arithmetic means ±SD (n = 10). Significance of differences between all values were evaluated by Student’s t-test (P ≤ 0.05), and it has been determined that each pair of values is significantly different if the SD bars do not overlap. Significance has not been marked above the bars with the letters or arrows to keep the clarity of the graph
The expression of nine ZIP genes was determined under the above conditions. Differences among the apical, middle, and basal root parts allowed categorization of ZIP genes into four groups (Fig. 4 a-d). The first contains ZIPs with the highest expression in the apical part (NtZIP2, NtZIP5A/B, NtIRT1, NtIRT1-like) (Fig. 4a1-a5). The second comprises NtZIP1-like and NtZIP8 expressed preferentially in the basal root part (Fig. 4b1-b2). The third is represented by NtZIP4A/B with equal expression in the three root parts (Fig. 4c1-c2). The remaining NtZIP5-like and NtZIP11 fall into the fourth group characterized by intermediate expression that changed in different ways in the root parts under the tested combinations of Zn/Cd concentrations (Fig. 4d1-d2).

Expression of tobacco ZIP genes in the apical, middle and basal parts of roots from plants grown under various Zn and Cd concentrations. 3.5-week old plants grown in the quarter-strength Knop’s medium were exposed to the control medium supplemented with pairwise combinations of Zn (0; 1 μM) and Cd (0; 0.25; 1 μM) for 17 days. ZIP genes preferentially expressed in the apical root part (a): NtZIP2 (a1), NtZIP5A (a2), NtZIP5B (a3), NtIRT1 (a4), NtIRT1-like (a5); in the basal root part (b): NtZIP1-like (b1), NtZIP8 (b2); equally expressed in tested three root parts (c): NtZIP4A (c1), NtZIP4B (c2); mixed expression pattern for tested three root parts (d): NtZIP5-like (d1), NtZIP11 (d2). Gene expression was normalized to the PP2A level. Values correspond to arithmetic means ±SD (n = 3); those with the ratio greater than 2 are considered significantly different
Furthermore, the transcript level of each tested ZIP gene in the apical, middle and basal root part depended on the combinations of Zn/Cd in the medium, although to different extents (Fig. 4). Expression was modified by Zn availability (all ZIPs, except NtZIP2 and NtZIP11, were upregulated by Zn-deficiency), and by the Zn/Cd concentration ratios. Here we investigated the root part-specific expression of ZIPs that accompanied Cd-dependent stimulation of Zn root-to-shoot translocation detected at different combinations of Zn to Cd concentrations: 0.25 μM Cd + 0 Zn, 0.25 μM Cd + 1 μM Zn, 1 μM Cd + 1 μM Zn (Fig. 2d). At each of these three combinations, different ZIP expression patterns were found. Adding 0.25 μM Cd to the Zn-deficient medium decreased expression of NtZIP5A/B in all three root parts (Fig. 4a2-a3,) NtIRT1 in the apical (Fig. 4a4) and NtZIP5-like in the middle part (Fig. 4d1), but its presence in the medium containing 1 μM Zn resulted in lower expression of NtZIP2 in the basal root part only (Fig. 4a1). Furthermore, at the combination of 1 μM Cd + 1 μM Zn (relative to 0 μM Cd + 1 μM Zn) increased transcript levels were noted either in all three root parts (NtZIP11; Fig. 4d2), in the apical and middle parts (NtZIP8, Fig. 4b2), in the basal and middle parts (NtZIP1-like; Fig. 4b1), or in the apical part only (NtZIP5-like, NtIRT1-like, NtIRT1, NtZIP5A) (Fig. 4d1, a5, a4, a2, respectively).
The detected root part-specific changes indicate a distinct function of each part in mineral nutrition at low/sufficient Zn in the medium and in the presence/absence of toxic Cd, in which the ZIPs under study play specific roles.
Short-term exposure to Cd: expression and accumulation study
The detected increased transcript level of several ZIP genes upon 17-day exposure to 1 μM Cd (Fig. 4) suggested induction by Cd. Here, short-term treatment (3 h, 1 day, 3 days) with 1 μM Cd + 1 μM Zn (Figs. 5, 6) indicated Cd-induced increase in the expression of NtZIP5B (apical part) and NtZIP1-like (all root parts - Fig. 5c, f, and leaves - Fig. 6a). The opposite effect, downregulation of NtIRT1 and NtIRT1-like was found in the middle root part (Fig. 5d, e). To complement the study, the Zn and Cd root/shoot distribution was determined (Fig. 7). It was shown that Zn translocation was already enhanced after 3-day exposure to 1 μM Cd + 1 μM Zn (as compared with medium without Cd).

Expression of ZIP genes in the apical, middle and basal parts of roots from plants after short-term exposure to Cd. 5.5-week old plants grown in the quarter-strength Knop’s medium were exposed to the control medium containing 1 μM Zn with (1 μM Cd) or without Cd for 3 h, 1 day, or 3 days. Expression of NtZIP2A (a), NtZIP5A (b), NtZIP5B (c), NtIRT1 (d), NtIRT1-like (e), NtZIP1-like (f), NtZIP8 (g), NtZIP4A (h), NtZIP4B (i), NtZIP5-like (j), NtZIP11 (k). Gene expression was normalized to the PP2A level. Values correspond to arithmetic means ±SD (n = 3); those with the ratio greater than 2 are considered significantly different

Expression of ZIP genes in the leaves of tobacco plants after short-term exposure to Cd. 5.5-week old plants grown in the quarter-strength Knop’s medium were exposed to the control medium containing 1 μM Zn with (1 μM Cd) or without Cd for 3 h, 1 day, or 3 days. Expression of NtZIP1-like (a), NtZIP4A (b), NtZIP4B (c), NtZIP5-like (d), NtZIP8 (e), NtZIP11 (f). Expression of NtZIP2, NtZIP5A, NtZIP5B, NtIRT1, NtIRT1-like were not detected in leaves. Gene expression was normalized to the PP2A level. Values correspond to arithmetic means ±SD (n = 3); those with the ratio greater than 2 are considered significantly different

Zinc and cadmium concentration in the shoots and roots of tobacco plants after 3-day exposure to Cd. 5.5-week old plants grown in the quarter-strength Knop’s medium were exposed to the control medium containing 1 μM Zn with (1 μM Cd) or without Cd for 3 days. Zn concentration in the shoots (a), and roots (b). Cd concentrations in the shoots (d) and roots (e). The translocation factor (TF) was determined as the ratio of shoot/root concentrations of Zn (c), and Cd (f). Values correspond to means ±SD (n = 10). Significance of differences between all values were evaluated by Student’s t-test (P ≤ 0.05), and it has been determined that each pair of values is significantly different if the SD bars do not overlap. Significance has not been marked above the bars with the letters or arrows to keep the clarity of the graph
Isolation of NtZIP5B and bioinformatics analysis
The above experiments indicated that Cd-dependent stimulation of Zn translocation to shoots was accompanied by downregulation of NtZIP5B specifically during conditions of zinc deficiency (Fig. 4a3). To learn more about the physiological function of this gene, it was cloned and characterized. Tobacco is an allotetraploid species [30], and numerous genes are present in two copies. Accordingly, here we found two copies of NtZIP5: NtZIP5A and NtZIP5B (Additional file 2). Detailed bioinformatics analysis showed that NtZIP5A has a nucleotide sequence identical with NtZIP1 previously identified by Sano et al. [31], and here we provided data based on which NtZIP1 was renamed NtZIP5A.
Our study showed that the sequence of NtZIP1 (acc. no AB 505626) is identical with the nucleotide sequence NM_001325745.1 described as Nicotiana tabacum zinc transporter 5-like (LOC107803903) mRNA and also shares the highest homology (95%) with the predicted N. tabacum zinc transporter 5-like (LOC107774100) mRNA (XM_016593570) (Additional file 2). The nucleotide sequences NM_001325745.1 and XM_016593570, and corresponding amino acid (aa) sequences were further used as a query for Blast searches for tobacco genes/proteins with the highest homology. As shown in Additional files 2a and 2b, the identified genes/proteins represented other ZIP5-like and ZIP5 from tobacco and other species, which share 100 to 83% identity. These results together with the data from the phylogenetic analysis (Fig. 8; Additional file 3) were the basis for changing the name of sequence A/B 505626.1/NM_001325745.1 from NtZIP1 to NtZIP5A, and sequence XM_016593570.1 was named NtZIP5B.

Phylogenetic analysis of ZIP transporters from selected nine species. Unrooted phylogenetic tree for the ZIP proteins was constructed based on amino acid sequences identified in the NCBI database, using MEGA 7.0 software. The length of branches are proportional to the degree of divergence. Numbers in the figure represent bootstrap values (1000 replicates). The accession numbers are as follows: Nicotiana tabacum NtZIP5A (NP_001312674.1), NtZIP5B (XP_016449056.1), NtZIP1-like (XM_016652513), Capsicum annuum CaZIP5 (PHT75648.1), Medicago truncatula MtZIP5 (XP_013461166.1), MtZIP1 (XP_013464193.1), Morus notabilis MnZIP5 (XP_010093125.1), Vitis vinifera VvZIP1 (XP_002264603.2), Arabidopsis thaliana AtZIP1 (NP_187881.1), AtZIP2 (NP_200760.1), AtZIP3 (NP_180786.1) AtZIP4 (NP_001320672.1), AtZIP5 (NP_172022.1), AtZIP6 (NP_180569.1), AtZIP7 (NP_178488.1), AtZIP9 (NP_001329099.1), AtZIP10 (NP_174411.2), AtZIP11 (NP_564703.1), AtZIP12 (NP_201022.1), AtIRT1 (NP_567590.3), AtIRT2 (NP_193703.2), AtIRT3 (NP_001321605.1), Vigna radiata VrZIP1 (XP_014501490.1), Hordeum vulgare HvZIP5 (ACN93833.1), Oryza sativa OsZIP1 (XP_015633357.1), OsZIP5 (XP_015637510.1)
The open reading frame (ORF) of NtZIP5A/NtZIP5B consists of 1017/1029 bp with 96.46% identity and encodes a predicted protein of 339/343 aa with 98.23% identity. Three exons were identified within the genomic sequences (Additional file 2e; Additional file 3). The phylogenetic relationships are demonstrated on a phylogenetic tree (Fig. 8) and in a Table showing sequence identity at the nucleotide and amino acid level (Additional file 3). It was shown that ZIP5 proteins from different plant species formed two distinct clades. First, both NtZIP5A and NtZIP5B cluster together with CaZIP5, MnZIP5 and MtZIP5 and show 83.63 to 69.62% identity to them. Furthermore, AtZIP5 and AtZIP3 reside on the closest sub-branch. The second ZIP5 clade includes Monocotyledonous HvZIP5 and OsZP5, which displayed the lowest identity (56.13 to 54.13%). Similarly, NtZIP5A/B show low identity (54.03 to 62.54%) to ZIP1 proteins (NtZIP1-like, AtZIP1 and VvZIP1), which reside on the closest sub-branch.
The aa sequences of NtZIP5A/B are aligned with ZIP5 and ZIP1 proteins identified in the phylogenetic tree (Figs. 8, 9). In agreement with the structure of ZIP family members from other plants, eight transmembrane domains (TMD), a very short C-terminal tail, and a hydrophilic region between TM domains III/IV were identified. TM III and IV of NtZIP5A/B proteins contain potential metal-binding histidine residues. The length of this region in NtZIP5A/B is 51/55 aa, respectively, which falls within the range of aa numbers typical for plant ZIPs (Additional file 2e). Moreover, amino acids within the TM-IV domain of NtZIP5A/B match the proposed ZIP signature domain [32]. However, within the variable region between the III/IV TMDs, NtZIP5A has three histidine residues, whereas NtZIP5B has five (Fig. 9).

Amino acid alignment of predicted ZIP5 and ZIP1 proteins from different species. Sequences were aligned using Clustal Omega https://www.ebi.ac.uk/Tools/msa/clustalo/. The prediction of membrane-spanning regions was performed using Phobius programme (http://phobius.sbc.su.se/), and indicated as lines above the sequences, and numbered TM I–VIII respectively. Sequences of the transmembrane domains (TM) are marked with light grey; identical amino acids with dark grey; histidine in black. Dashes indicate gaps
In silico analysis of the promoter regions of NtZIP5A and NtZIP5B (1797 bp and 1726 bp), identified several cis-acting regulatory elements regulated by metals and other abiotic stresses having a high degree of similarity with respect to location and types of elements (Additional file 4). In both copies, in the same location we found ZDRE elements (zinc deficiency-related elements), IDE2 (iron deficiency-responsive element 2), ABRE and ABRE4 (abscisic acid-responsive), MYB-like sequence (MYB binding region from A. thaliana), Box 4 and GT1 (abiotic stresses such as wounding and pathogen response elements), STRE (Stress Response Element) for heat, osmotic, low pH and nutrient starvation, and CAAT-box (common cis-acting element in promoter and enhancer regions in Pisum sativum), whereas light-responsive G-box was detected in a different location [33,34,35,36,37]. Additional copies of Box-4 and STRE were identified in NtZIP5A, while IDE2, ABRE, and GT-1 in NtZIP5B. Furthermore, four distinct cis-acting elements were found exclusively in the promoter of NtZIP5A: Box-S and W-box (wounding and pathogen response elements), P-box (giberelin response), and TCT (for light response). Alignments of the promoter regions of NtZIP5A and NtZIP5B from three different cultivars (K326, Basma Xanthi, and K326) showed identity of sequences (Additional file 4b-d).
Bioinformatics analysis to determine the subcellular localization of NtZIP5B was performed with the use of the ProtComp programme [38, 39]. As shown in Additional file 5, the NtZIP5B protein was predicted to be localized at the plasma membrane.
Functional analysis of NtZIP5B in yeast
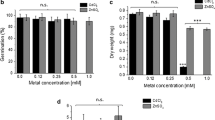
Increased sensitivity to Cd of the wild-type yeast DY1457 expressing pAG426-NtZIP5B as compared with the wild-type expressing pAG426 (Fig. 10a) indicated involvement of NtZIP5B in Cd uptake. Expression of pAG426-NtZIP5B in the zrt1zrt2Δ mutant line improved growth on minimal medium with increasing concentrations of EGTA (Fig. 10b). Moreover, it led to enhanced sensitivity to high Zn (Fig. 10c). Both results suggest that NtZIP5B mediates Zn uptake.

Complementation by NtZIP5B cDNA of yeast mutants defective in metal uptake on selective media. Yeast cells: DY1457, Δzrt1zrt2 (defective in Zn uptake) were transformed with the empty vector pAG426 as a control (e.v.), or with the vector carrying NtZIP5B with the stop codon pAG426-ZIP5B (NtZIP5B). Yeast cultures were adjusted to an OD600 of 0.2, and 3 μl of serial dilutions (from left to right in each panel) was spotted on SC-URA medium containing 2% (w/v) galactose solidified with 2% agar, supplemented with CdCl2 (a); EGTA (b), or ZnCl2 (c). The plates were incubated for 3–6 d at 30 °C. The images are representative of three independent experiments taken after 3 days of growth
Tissue-specific expression of NtZIP5B
Analysis of NtZIP5Bprom-GUS transgenic tobacco (seven homozygous T2 lines) showed no expression under control conditions (Fig. 11a1-a2), induction was noted at Zn deficiency (g. 11b1, b2).

GUS staining pattern of transgenic plants expressing NtZIP5Bprom::GUS. GUS expression in 4-week-old transgenic and wild-type seedlings grown at control conditions (a) and in the Zn-deficient (−Zn) medium (b) for 4 days. Whole roots (a1, b1); whole leaves (a2, b2); Basal root part (b1a) and the cross section (b1-b); Root segment from the middle part with the protruding lateral root (b1-c) and the cross section (b1-d); root apex (b1-e) and cross sections at different positions as indicated by arrows (b1-f, b1-g, b1-h); the apical root part (b1-i) and the cross sections from the position indicated by arrow (b1-j); Cross section through the leaf blade (b3) and two chosen areas at higher magnification (b3-a, b3-b). Magnification bars in different colours represent indicated length
GUS activity was not detected in the basal root part (Fig. 11b1-a, b1-b). High levels were found in all tissues of the root tips (except the quiescent center and the root cap) and in newly formed lateral roots (Fig. 11b1-c, b1-d, b1-e, b1-f, b1-g, b1-h). The intensity of staining increased in the epidermis at a distance of approximately 4 mm from the root tip and further up in the more mature region where it was localized primarily in the epidermis (Fig. 11b1-i, b1-j). These results indicate the contribution of NtZIP5B to the uptake of minerals from the medium by epidermal cells, also from the apoplast by cells comprising internal tissues. This function is specific to certain root parts only, primarily to the mature segments of the apical root parts.
Interestingly, in the leaves GUS activity was low and unevenly distributed (Fig. 11b2). Such distribution of NtZIP5B expression sites was confirmed by performing histochemical staining directly on sections from fresh leaves (Fig. 11b3). Blue staining was seen in the groups of mesophyll cells (Fig. 11b3-a, b3-b). Thus visually indistinguishable mesophyll cells have differing abilities to take up Zn.
Discussion
The apical, middle and basal root parts are distinct constituents of the root with specific contributions to Zn and Cd sequestration and Cd-dependent stimulation of Zn translocation to shoots
Our research showed that in tobacco under both Zn-deficiency and at 1 μM Zn in the medium, Zn translocation to shoots increased in the presence of Cd. This effect was lost at higher Zn concentrations (Fig. 2d). Competitive interactions between Zn and Cd modify the root-to-shoot translocation efficiency of both metals [9, 40,41,42], however the underlying mechanisms remain unknown. Two basic processes are decisive in the regulation of metal root/shoot partitioning; (i) sequestration in root cells, which determines the availability of the metal for xylem loading, (ii) efficiency of loading into xylem [43,44,45,46,47,48]. The majority of current research has been conducted on whole roots composed longitudinally of zones that are at different developmental stages (meristematic, differentiation, primary and secondary structures). It is not known if each zone plays distinct role in the regulation of metal root-to-shoot distribution.
In this study, the Zn and Cd accumulation in three root parts was compared among combinations of Zn/Cd concentrations representing contrasting values of the Zn Translocation Factor. Thus, the experimental variant with Cd (where the stimulation of Zn translocation occurred) was compared with the reference combination without Cd (0 μM Zn + 0.25 μM Cd vs 0 μM Zn + 0 μM Cd); (1 μM Zn + 0.25 μM Cd vs 1 μM Zn + 0 μM Cd); (1 μM Zn + 1 μM Cd vs 1 μM Zn + 0 μM Cd). We showed that the capacity of the apical, middle and basal root parts to accumulate Zn and Cd is different and depends on the concentrations of both metals in the soil solution (Fig. 3). The detected Zn/Cd status-dependent root part-specific distribution of Zn and Cd implicates distinct roles of the apical, middle and basal root parts in retention of both metals, consequently in the regulation of their translocation to shoots. Under Zn deficiency, the overall Zn concentration in roots was low (Fig. 2b), thus the distribution pattern characterized by the highest level of the metal in the apical part (Fig. 3a1) likely reflects the necessity of providing Zn to differentiating cells. When the Zn and Cd concentrations increased, the middle and basal parts of the root took over the major role of a sink for excess metal (Fig. 3a2, b2), which might protect the young apical zone from their toxicity. It is not known whether under higher metal status redistribution of metals is involved, or if uptake from the medium directly by the middle part (or even the basal part) is stimulated. The mechanisms remain unknown, since to date, neither the metal accumulation capacity of root parts, nor expression patterns specific for them have been taken into account. Only a few studies have demonstrated the longitudinal variation of metal influx. For example, Laporte et al. [49] showed that the efficiency of the longitudinal translocation of 109Cd applied locally was low, and the absorption of 109Cd at the tips of first-order sunflower roots was 2.9-fold that of the basal region. The root part-specific retention of Zn and Cd and less effective redistribution of 109Cd than 65Zn was also reported by Page and Feller [50]. They suggested the contribution of the xylem-to-phloem transfer of 65Zn and 109Cd taken up by roots in the regulation of the longitudinal distribution of both metals in roots.
Of importance was the discovery that Zn/Cd status-dependent distribution of both metals between the root parts (Figs. 1-3) was accompanied by specific changes in ZIPs expression patterns, which fell into four categories (Fig. 4). The first category included NtZIP2, NtZIP5A/B, NtIRT1, NtIRT1-like with the highest expression in the apical part; the second, those with the highest expression in the basal part (NtZIP1-like and NtZIP8); the third was represented by NtZIP4A/B with uniform expression in three root parts; and the fourth, by genes whose expression was not specific for any part of the root (NtZIP5-like and NtZIP11). These observations expose the complexity of the regulation of metal transfer along the whole root and within its tissues in response to changing metal status, and indicate the existence of signals specific to the apical, middle, and basal root parts that are generated upon a particular combination of Zn and Cd concentrations. Regulation of longitudinal distribution requires numerous genes involved in the uptake of metals, their retention in the particular root parts or effective redistribution, and in their transfer to the shoot.
Seven ZIPs were Zn-deficiency inducible, and upregulation was restricted to the apical root part only for NtIRT1-like, to the apical and middle part for NtIRT1, to the middle and basal part for NtZIP8, and to all root parts for NtZIP1-like, NtZIP5A/B, NtZIP5-like, NtZIP4A/B (Fig. 4). Furthermore, we detected upregulation by 17-day exposure to 1 μM Cd of NtIRT1, NtZIP1-like, and NtZIP11 in all three root parts, NtZIP5A/B and NtZIP5-like in apical parts, and NtZIP8 in apical and middle parts (Fig. 4). Knowing that enhanced transcript levels upon long-term treatment do not necessarily indicate upregulation by Cd itself, but might result from secondary changes in metal homeostasis [51,52,53], short-term exposure (3 h or 1 day) was applied. Detected upregulation of NtZIP5B in the apical root part, and NtZIP1-like in all root parts, and downregulation of NtIRT1 and NtIRT1-like in the middle root part (Fig. 5) suggest root-part-specific regulation by Cd. For comparison, Yoshihara et al. [52] showed upregulation of NtIRT1 in whole tobacco roots, and Barabasz et al. [25], downregulation of NtZIP4A/B upon 3-day exposure to 4 μM Cd in leaves, but not in roots.
From the results here described we conclude that the regulation of the longitudinal distribution of Zn and Cd is highly specific, and that the apical, middle and basal root parts play a specific role in accumulation and, likely, in the control of the efficiency of metal translocation to shoots, including the detected stimulation of Zn translocation to shoots in the presence of Cd (Fig. 2). Here, based on distinct expression of tested ZIP genes in root parts between combinations of medium containing Cd (at which Zn translocation was enhanced) and the reference medium without Cd, ZIP transporters were identified as candidates to be players involved in stimulation of Zn translocation (Fig. 4). However, as shown at the graphic summary (Additional file 6) it turned out that for each combination of Zn/Cd concentrations, different ZIP genes with modified expression were identified: (i) NtZIP5-like and NtZIP5A/B at 0 μM Zn + 0.25 μM Cd; (ii) NtZIP2 at 1 μM Zn + 0.25 μM Cd, (iii) NtZIP11, NtZIP8, NtZIP1-like, NtIRT1, NtIRT1-like, and NtZIP5-like at 1 μM Zn + 1 μM Cd. Thus, the same phenomenon (stimulation of Zn translocation in the presence of Cd) detected at distinct combinations of Zn and Cd concentrations (0 μM Zn + 0.25 μM Cd; 1 μM Zn + 0.25 μM Cd; 1 μM Zn + 1 μM Cd) was accompanied by modified expression of different ZIP genes in a root part-specific manner (Additional file 6). This indicates that distinct molecular mechanisms or regulatory processes that contribute to Cd-dependent stimulation of Zn translocation to shoots may be involved at different Zn/Cd concentrations in the medium. This is likely linked to different metal statuses in the tested combinations (including exposure to low and severe stress resulting from the presence of Cd) that determine the expression pattern of sets of genes involved in different defense mechanisms [7, 54, 55]. To fully understand the underlying mechanisms, further research is needed to identify all regulatory components of the metal cross-homeostasis network.
NtZIP5A/B –a novel Zn deficiency-inducible transporter involved in Zn and Cd uptake
NtZIP1 (AB505626 / NM_001325745.1) was first cloned by Sano et al. [31] from tobacco BY-2 cells. However, he focused primarily on NtNRAMP1, and only an initial study on NtZIP1 was performed. In our study, we renamed the gene NtZIP1 as NtZIP5A based on detailed bioinformatics analysis and cloning. First, sequences with the highest homology to NM_001325745.1 (identified in the NCBI data base) all belonged to ZIP5 genes (Additional file 2c-e). Second, phylogenetic analysis of predicted proteins showed that tobacco NtZIP5A/NtZIP5B formed a distinct clade with dicot CaZIP5, MnZIP5, MtZIP5, AtZIP5 and AtZIP3 (Fig. 8), while two monocot HvZIP5 and OsZIP5 formed a separate branch. These suggest functional divergence between the Dicotyledonous and Monocotyledonous ZIP5 transporters. Moreover, alignments of predicted amino acids of NtZIP5A/NtZIPB with other ZIP proteins (Fig. 9) also showed high sequence conservation with typical ZIP structures (eight transmembrane domains, a short C-terminal tail, a variable region between TM3-TM4 containing histidine residues) [32, 56, 57].
Furthermore, a yeast growth assay using the zrt1zrt2 mutant line and wild type expressing NtZIP5B suggested that Zn and Cd are substrates (Fig. 10). Together with in silico bioinformatics analysis suggesting localization of NtZIP5B in the plasma membrane (Additional file 5), these results point to this protein’s role in Zn and Cd uptake. For comparison, HvZIP5, OsZIP5, MtZIP5, and ZmZIP5 have also been shown to be Zn uptake proteins, moreover, HvZIP5, OsZIP5 and ZmZIP5 have been localized to the plasma membrane [58,59,60,61,62].
In the tobacco genome, most genes are present in two copies originating from one of the two ancestors, N. tomentosiformis or N. sylvestris [30]. Here, two NtZIP5 homologous genes, NtZIP5A and NtZIP5B, were identified (Additional file 2), which is similar to the presence of NtMTP1a and NtMTP1b, or NtHMA4α and NtHMA4β [63, 64]. In our study, the comparable expression patterns between NtZIP5A and NtZIP5B found in the apical, middle, and basal parts of roots (Fig. 4a1-a5), and absence of expression in leaves (legend to Fig. 6), suggested their similar functions in the regulation of metal homeostasis. Analysis of the cis-acting elements within the promoter region of new NtZIP5A/B genes showed the same number and localization of ZDRE elements responsible for upregulation under Zn-deficiency [36] and two IDE2s (iron-deficiency-responsive element 2) [33, 35]. Thus, both NtZIP5A and NtZIP5B likely play a similar role in the regulation of metal homeostasis. Numerous other regulatory sequences also displayed identity, with the exception, however, of four elements that were detected only within the NtZIP5A promoter, which are regulators of responses to wounding and pathogens (Box S and W-box), light (TCT motif), and giberellin (P-box) (Additional file 4). Hence compared with NtZIP5B, one might speculate that NtZIP5A could be involved more broadly in signaling pathways induced by multiple factors. Interestingly, the presence of sequences responsible for responses to various abiotic and biotic stresses and to abscisic acid (ABA) points likely not only to the existence of mechanisms by which metal transporters are regulated by numerous factors, but also to the importance of Zn homeostasis in a plant’s response to them. These could be a manifestation of cross-homeostasis not only between different metals, but also between abiotic and biotic stresses.
In addition, as shown by GUS expression analysis, NtZIP5B primarily provides root tissues with Zn under Zn-deficiency conditions (Fig. 11b1). Strong expression detected within the apical meristem across all tissues (Fig. 11b1-e, b1-f, b1-g) and further up in the epidermis (Fig. 11b1-i, b1-j) points to its function in the uptake of minerals from the medium. NtZIP5B is the new ZIP protein for which involvement in the absorption of Zn directly from the soil solution has been indicated. This function was first ascribed to the IRT1 protein [27, 28, 65], also to IRT3 [66], and recently to NtZIP4B, which provided Zn for the whole root up to its basal part, except for the very apical meristem [25]. NtZIP5B provides metals also to palisade parenchyma cells, but interestingly, not across the entire leaf blade (Fig. 11b2, b3), indicating the distinct ability of mesophyll cells to store Zn, as suggested by Siemianowski et al. [67].
Conclusions
The presented results show that the apical, middle and basal root parts played distinct roles in the sequestration of Zn and Cd that depended on the reciprocal combinations of the concentrations of both metals. The detected regulation of metal distribution within root parts could be regarded as a mechanism protecting young meristematic cells within the root apex. Moreover, it might contribute to the regulation of Zn/Cd root-to-shoot distribution, likely by formation of a metal pool available for translocation to shoots. Next, the identified categories of the root part-specific expression pattern of tobacco ZIP genes that depended on the Zn/Cd status provide a first glimpse into the complexity of the regulatory cross-talk between Zn and Cd. This is a key element in determining metal distribution among root parts and the contribution of underlying processes in the regulation of metal root-to-shoot translocation, including the stimulation of Zn translocation in the presence of Cd and associated changes in the expression level of ZIP genes. We were the first to show the possible involvement of ZIP genes in this process, although to get a complete picture it is necessary not only to fully understand their physiological role, but also to identify other genes acting in concert.
Furthermore, we conclude that the newly cloned tobacco NtZIP5B is a Zn-deficiency inducible transporter involved in absorption of Zn and Cd from the soil solution. It performs uptake functions in the root epidermis, but also provides Zn to internal root tissues such as xylem parenchyma, and in leaves to groups of palisade parenchyma cells. The irregular expression of NtZIP5 within the palisade parenchyma indicates that these cells have a distinct capacity to take up metals. Further study is necessary to reveal the identity of these cells.
Methods
Plant material, growth conditions and treatments
The experiments were performed on tobacco (Nicotiana tabacum var. Xanthi) plants. Seeds were obtained from the stock of the Institute of Biochemistry and Biophysics PAS, Warszawa, Poland in 2002, since then they were propagated in the greenhouse of the University of Warsaw for our experiments. Plants were cultivated in a controlled environment chamber as previously described [25].
Surface-sterilized seeds in 8% sodium hypochlorite (w/v) were grown for the first three weeks on vertically positioned Petri dishes containing quarter-strength Knop’s medium, 2% sucrose (w/v) and 1% agar (w/v). Afterwards plants were transferred to aeriated hydroponic control medium (quarter-strength Knop’s; composition is given in Barabasz et al. [68]). They were grown in 2 L pots (5 plants per pot) for a period depending on the experiment (details below). The nutrient solution was renewed every 3 days.
Plant samples were collected at the end of each experiment. For expression analysis, plant material was frozen in liquid nitrogen and stored at − 80 °C. The experiments were done in three independent biological replicates. For each experimental variant, plant material was collected and pooled from 10 to 30 plants in total.
For determination of Zn and Cd concentrations, roots were washed in ice-cold 5 mM CaCl2 according to Barabasz et al. [68]. Plant samples were dried at 50 °C and stored at room temperature (RT) until analysis.
Long-term experiments
Three-week-old seedlings were transferred from the agar-solidified medium to the hydroponic control medium and grown for three days, then exposed for 17 days to the control medium supplemented with different Zn (as ZnSO4) and Cd (as CdCl2) concentrations.
To determine Zn and Cd root/shoot partitioning, plants were exposed to pairwise combinations of Zn (0; 1; 5; 10; 50 μM) and Cd (0; 0.25; 1; 4 μM) concentrations, then whole roots and shoots were collected.
To determine Zn and Cd concentrations and the expression of tobacco ZIP genes in the apical (distal), middle, and basal (proximal) parts of the roots, plants were exposed to Cd (0; 0.25; 1 μM) in the presence of 1 μM Zn or without Zn, then the following root parts were collected: (i) apical (distal) parts (2.5 cm) from the main and lateral roots (containing meristematic tissues, differentiation zone and young primary tissues); (ii) basal (proximal) parts equal to 1/4 of the total length of the main roots (containing primary tissues and within the central cylinder, secondary conductive tissues); (iii) middle parts making up the remaining parts of the main roots (containing primary tissues). The basal and the middle parts of the roots were collected from the main root only (adventitious roots were excised). Taking into consideration the changes of the root branching pattern in plants exposed to a set of Zn and Cd concentrations in the medium, to be able to compare samples collected from plants grown at different medium compositions, the basal and the middle parts of the roots were collected from the main root only.
Short-term experiments
Three-week-old seedlings were transferred from the agar-solidified medium to hydroponic control medium for 2.5 weeks, then were exposed to pairwise combinations of Zn (0; 1 μM) and Cd (0; 1 μM) for 3 h, 1 day, or 3 days. Leaves (the 2nd and 3rd counting from the base), and roots (collected as described above) were frozen for expression analysis. Moreover, after 3-day Cd, treatment roots and shoots were collected for determination of Zn and Cd concentrations.
Transcript analysis
The quantitative real-time PCR (qRT-PCR) based analysis was performed as described by Papierniak et al. [20]. In brief, the relative quantities of each transcript in the samples was calculated based on the comparative ΔCt (threshold cycle) method [69]. As an internal control tobacco NtPP2A (protein phosphatase 2A; AJ007496) was used, and its stability in the range of applied metal concentrations are given in Additional file 7. Primer sequences are listed in Additional file 8. For each sample, reactions were set up in three repetitions to calculate means. Studies were conducted on a minimum of three biological replicates.
Determination of metal concentrations
Mineralization and determination of Zn and Cd concentrations by Atomic Absorption Spectrophotometry (AAS) were performed according to Barabasz et al. [53]. To compare the efficiency of Zn and Cd root-to-shoot translocation between plants grown at the applied medium compositions, the Translocation Factor (TF) was determined as the ratio of shoot/root Zn and Cd concentrations [9].
Cloning of NtZIP5B and generation of constructs
In this study, the full length open reading frame (ORF) of NtZIP5B was amplified by PCR using Phusion HF polymerase (Thermo Scientific) with the cDNA transcribed from total RNA. The primer sequences are given in Additional file 2c, d, and in Additional file 8. Full length NtZIP5B containing the STOP codon was cloned into a vector pENTR™/D-TOPOR (Invitrogen), and the presence of a correct insert in the plasmid pENTR/D-TOPO-NtZIP5B was checked by sequencing (Genomed, Poland).
For yeast complementation assays the vector pAG426-NtZIP5B was generated by LR recombination between the pENTR/D-TOPO-NtZIP5B and the destination vector pAG426GAL-cccB-EGFP.
To determine the tissue-specific expression of NtZIP5B, the promoter sequence was cloned with the use of pENTR Directional TOPO Cloning Kits (Invitrogen). The promoter sequence of 1726 bp upstream of the start codon was amplified with the use of primers specific to the genomic sequence AWOK01252628.1, and cloned to the vector pENTR/D-TOPO. The CACC sequence was added to the forward primers (Additional file 8). One Shot TOP10 E. coli was transformed with the construct pENTR/D::ZIP5Bprom and the insert was sequenced. The NtZIP5B promoter was cloned in translational fusion with the uidA gene (GUS gene) in the destination binary vector, pMDC163, by LR recombination and the sequence of the insert in the construct pMDC163::NtZIP5Bprom::GUS was checked again.
NtZIP5 sequence analysis
The nucleotide cDNA sequences of NtZIP5A and NtZIP5B were translated to protein sequences with the use of the ExPASy translate tool (http://web.expasy.org/translate/). Similarity searches were performed using the BLAST algorithm. Multiple sequence alignment of the NtZIP5A/B and ZIP proteins from chosen plant species was performed with CLustalW [70]. The phylogenetic tree was constructed with MEGA 7.0 software [71] using the maximum likelihood method with 1000 bootstrap replicates. Potential transmembrane domains were identified with Phobius software [72]. To identify Nicotiana tabacum ZIP sequences in the NCBI database, BLASTn (Basic Local Alignment Search Tool) searches were performed based on homology to already known A. thaliana sequences. The subcellular localizations of NtZIP4B protein was predicted with the use of the ProtComp v. 9.0 online; http://www.softberry.com/berry.phtml?topic=protcomppl&group=programs&subgroup=proloc.
Yeast growth assay
Two Saccharomyces cerevisiae strains were used in the experiments: (i) wild-type DY1457 (MATa, ade1 can1 his3 leu2 trp1 ura3), (ii) mutant ZHY3 –zrt1zrt2 (DY1457 + zrt1::LEU2, zrt2::HIS3) defective in high- and low-affinity zinc uptake. The prepared construct pAG426-NtZIP5B and the empty vector pAG426GAL were used for yeast transformation using the lithium acetate protocol [73].
The zrt1zrt2Δ strain defective in high- and low-affinity Zn uptake was used to determine if NtZIP5B mediates Zn uptake. Yeast were grown as described by Barabasz et al. [25]. Briefly, the wild type (WT) DY1457 was transformed with the empty vector pAG426GAL, and the mutant zrt1zrt2 with pAG426-NtZIP5B. Next, they were cultivated on the SC-URA medium (containing 0.2 mM Zn) with galactose. Liquid cultures were spotted onto agar-solidified plates containing SC-URA with GAL supplemented with 0.5 to 2 mM EGTA (ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid) or with 0.1 to 2 mM Zn concentrations.
To check whether NtZIP5B transports Cd, the WT strain DY1457 was transformed with the empty vector pAG426GAL, and with the vector pAG426-NtZIP5B. The growth of yeast cultivated on an agar-solidified SC-URA with GAL medium containing 5 to 50 μM Cd (CdCl2) was monitored.
Tissue-specific expression of NtZIP5B prom::GUS
The pMDC163::NtZIP5Bprom::GUS construct was used for tobacco leaf disc transformation using an Agrobacterium-mediated method as previously described [68]. Transformants were selected for hygromycin resistance. The T2 homozygous lines were selected from T1 plant lines with a segregation ratio of 3:1 (tolerant:sensitive).
Three-week-old transformants (seven lines) and the wild type were transferred from the agar-solidified medium to hydroponic control medium for 3 days, then exposed to the medium without Zn (and in parallel to the control one) for 4 days. Whole seedlings were used for GUS histochemical staining. Representative individuals from representative lines were photographed. To visualize the expression of NtZIP5B at the tissue/cell level, stained root fragments were embedded in 3% agarose, 150 μ sections were cut on a Vibratome (Leica VT1000S, Heidelberg, Germany) and used for microscopic analysis (OPTA-TECH microscope). In addition, leaf cross-sections made from fresh unstained leaves on a Vibratome were subjected to histochemical staining. GUS activity was determined as described by Barabasz et al. [25].
Statistical analysis
Evaluation of statistical significance was performed at the 0.05 probability level using Student’s t-test. Experiments were performed with at least three independent replicates.
Availability of data and materials
Datasets supporting conclusions of this article are included within the article and its additional files.
Abbreviations
- Aa:
-
Amino acid
- AAS:
-
Atomic Absorption Spectrophotometry
- ABA:
-
Abscisic acid
- ABRE:
-
Abscisic acid-responsive element
- BLAST:
-
Basic Local Alignment Search Tool
- Cd:
-
Cadmium
- Cu:
-
Cooper
- EGTA:
-
Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid
- GAL:
-
Galactose
- GUS:
-
β-glucuronidase
- IDE:
-
Iron deficiency-responsive element
- IRT:
-
Iron regulated transporter
- Mn:
-
Manganese
- MYB-like:
-
Myeloblastosis-like
- ORF:
-
Open reading frame
- RT:
-
Room temperature
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- SSH:
-
Suppression Subtractive Hybridization
- STRE:
-
Stress Response Element
- TMD:
-
Transmembrane domain
- WT:
-
Wild type
- ZDRE:
-
Zinc deficiency-related element
- ZIP:
-
ZRT/IRT related Proteins
- Zn:
-
Zinc
- ZRT:
-
Zinc related transporter
References
Lugon-Moulin N, Zhang M, Gadani F, Rossi L, Koller D, Krauss M, et al. Critical review of the science and options for reducing cadmium in tobacco (Nicotiana tabacum L.) and other plants. Adv. Agron. 2004;83:111–80.
Lozano-Rodriguez E, Hernandez L, Bonay P, Carpena-Ruiz R. Distribution of cadmium in shoot and root tissues of maize and pea plants: physiological disturbances. J Exp Bot. 1997;48:123–8.
Vangronsveld J, Herzig R, Weyens N, Boulet J, Adriaensen K, Ruttens A, et al. Phytoremediation of contaminated soils and groundwater: lessons from the field. Environ Sci Pollut Res. 2009;16:765–94.
Herzig R, Nehnevajova E, Pfistner C, Schwitzguebel J-P, Ricci A, Keller C. Feasibility of labile Zn phytoextraction using enhanced tobacco and sunflower: results of five- and one-year field-scale experiments in Switzerland. Int J Phytoremed. 2014;16:735–54.
Grotz N, Guerinot ML. Molecular aspects of cu, Fe and Zn homeostasis in plants. Biochim Biophys Acta. 1763;2006:595–608.
Broadley MR, White PJ, Hammond JP, Zelko I, Lux A. Zinc in plants. New Phytol. 2007;173:677–702.
Sinclair SA, Krämer U. The zinc homeostasis network of land plants. Biochim Biophys Acta. 1823;2012:1553–67.
Krupa Z, Siedlecka A, Skórzyńska-Polit E, Maksymiec W. Heavy-metal interactions with plant nutrients. In: MNV P, Strzałka K, editors. Physiology and biochemistry of metal toxicity and tolerance in plants. Dordrecht: Kluwer Academic Publishers; 2002. p. 287–301.
Barabasz A, Klimecka M, Kendziorek M, Weremczuk A, Ruszczyńska A, Bulska E, et al. The ratio of Zn to cd supply as a determinant of metal-homeostasis gene expression in tobacco and its modulation by overexpressing the metal exporter AtHMA4. J Exp Bot. 2016;67:6201–14.
Ueno D, Iwashita T, Zhao F-J, Ma JF. Characterization of cd translocation and identification of the cd form in xylem sap of the cd-hyperaccumulator Arabidopsis halleri. Plant Cell Phys. 2008;49:540–8.
Küpper H, Kochian LV. Transcriptional regulation of metal transport genes and mineral nutrition during acclimatization to cadmium and zinc in the cd/Zn hyperaccumulator, Thlaspi caerulescens (Ganges population). New Phytol. 2010;185:114–29.
Bovet L, Rossi L, Lugon-Moulin L. Cadmium partitioning and gene expression studies in Nicotiana tabacum and Nicotiana rustica. Physiol. Plantarum. 2006;128:466–75.
Assunção AGL, Bleeker P, ten Bookum WM, Vooijs R, Schat H. Intraspecific variation of metal preference patterns for hyperaccumulation in Thlaspi caerulescens: evidence from binary metal exposures. Plant Soil. 2008;303:289–99.
de la Rosa G, Peralta-Videa JR, Montes M, Parsons JG, Cano-Aguilera I, Gardea-Torresday JL. Cadmium uptake and translocation in tumbleweed (Salsola kali), a potential cd-hyperaccumulator desert plant species: ICP/OES and XAS studies. Chemosphere. 2004;55:1159–68.
Cun P, Sarrobert C, Richaud P, Chevalier A, Soreau P, Auroy P, et al. Modulation of Zn/cd P1B2-ATPase activities in Arabidopsis impacts differently on Zn and cd contents in shoots and seeds. Metallomics. 2014;6:2109–16.
Palmer CM, Guerinot ML. Facing the challenges of cu, Fe and Zn homeostasis in plants. Nature Chem Biol. 2009;5:333–40.
Milner MJ, Seamon J, Craft E, Kochian LV. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J Exp Bot. 2013;64:369–81.
López-Millán A-F, Ellis DR, Grusak MA. Identification and characterization of several new members of the ZIP family of metal ion transporters in Medicago truncatula. Plant Mol Biol. 2004;54:583–96.
Ramesh SA, Shin R, Eide DJ, Schachtman DP. Differential metal selectivity and gene expression of two zinc transporters from rice. Plant Physiol. 2003;133:126–34.
Papierniak A, Kozak K, Kendziorek M, Barabasz A, Palusińska M, Tiuryn J, et al. Contribution of NtZIP1-like to the regulation of Zn homeostasis Frontiers. Plant Sci. 2018;9:185.
Kozak K, Papierniak A, Barabasz A, Kendziorek M, Palusińska M, et al. 2019. NtZIP11, a new Zn transporter specifically upregulated in tobacco leaves by toxic Zn level. Env. Exp. bot. 2019;157:69-78.
Grotz N, Fox T, Connolly E, Park W, Guerinot ML, Eide D. Identification of a family of zinc transporter genes from Arabidopsis that respond to zinc deficiency. Proc Natl Acad Sci U S A. 1998;95:7220–4.
Wintz H, Fox T, Wu YY, Feng V, Chen W, Chang HS, et al. Expression Profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J Biol Chem. 2003;278:47644–53.
Burleigh SH, Kristensen BK, Bechmann IE. A plasma membrane zinc transporter from Medicago truncatula is up-regulated in roots by Zn fertilization, yet down-regulated by arbuscular mycorrhizal colonization. Plant Mol Biol. 2004;52:1077–88.
Barabasz A, Palusińska M, Papierniak A, Kendziorek M, Kozak K, Williams LE, et al. Functional analysis of NtZIP4B and Zn status-dependent expression pattern of tobacco ZIP genes. Frontiers Plant Sci. 2019;9:1984.
Fan W, Liu C, Cao B, Qin M, Long D, Xiang Z, Zhao A. Genome-wide identification and characterization of four gene families putatively involved in cadmium uptake, Translocation and Sequestration in Mulberry. Front Plant Sci. 2018;9:879.
Eide D, Broderius M, Fett J, Guerinot ML. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc Natl Acad Sci U S A. 1996;93:5624–8.
Rogers EE, Eide DJ, Guerinot ML. Altered selectivity in an Arabidopsis metal transporter. Proc Natl Acad Sci U S A. 2000;97:12356–60.
Cohen CK, Garvin DF, Kochian LV. Kinetic properties of a micronutrient transporter from Pisum sativum indicate a primary function in Fe uptake from the soil. Planta. 2004;218:784–92.
Kenton A, Parokonny AS, Gleba YY, Bennett MD. Characterization of the Nicotiana tabacum L. genome by molecular cytogenetics. Mol. Gen. Genomics. 1993;240:159–69.
Sano T, Yoshihara T, Handa K, Sato MH, Nagata T, Hasezawa S. Metal ion homeostasis mediated by Nramp transporters in plant cells - focused on increased resistance to iron and cadmium ion. In: Weigert R, editor. Crosstalk and integration of membrane trafficking pathways. Rijeka, Shanghai: INTECH; 2012. p. 214–28.
Eng BH, Guerinot D, Eide MH, Saier J. Sequence analyses and phylogenetic characterization of the ZIP family of metal ion transport proteins. J Membrane Biol. 1998;166:1–7.
Kobayashi T, Nakayama Y, Itai RN, Nakanishi H, Yoshihara T, Mori S, et al. Identification of novel cis-acting elements, IDE1 and IDE2, of the barley IDS2 gene promoter conferring iron-deficiency-inducible, root-specific expression in heterogeneous tobacco plants. Plant J. 2003;36:780–93.
Zhang W, Ruan J, Ho T, You Y, Yu T, Quatrano RS. Cis-regulatory element based targeted gene finding: genome-wide identification of abscisic acid- and abiotic stress-responsive genes in Arabidopsis thaliana. Bioinformatics. 2005;21:3074–81.
Ogo Y, Kobayashi T, Itai RN, Nakanishi H, Kakei Y, Takahashi M, et al. A novel NAC transcription factor, IDEF2, that recognizes the iron deficiency-responsive element 2 regulates the genes involved in iron homeostasis in plant. J Biol Chem. 2008;283:13407–17.
Assunção AGL, Herrero E, Lin Y-F, Lin Y-F, Huettel B, Talukdar S, et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc Natl Acad Sci U S A. 2010;107:10296–301.
Ambawat S, Sharma P, Yadav NR, Yadav RC. 2013. MYB transcription factor genes as regulators for plant responses: an overview. Physiol Mol Biol Plants. 2013;19:307–21.
Yu CS, Chen YC, Lu CH, Hwang JK. Prediction of protein subcellular localization. Proteins. 2006;64:643–51.
Fu X-Z, Zhou X, Xing F, Ling L-L, Chun, CP, Cao L, et al. Genome-wide identification, cloning and functional analysis of the Zinc/Iron-regulated transporter-like protein (ZIP) gene family in Trifoliate Orange (Poncirus trifoliata L. Raf.). Frontiers Plant Sci. 2017;8:588.
Hart JJ, Welch RM, Norvell WA, Kochian LV. Transport interactions between cadmium and zinc in roots of bread and durum wheat seedlings. Physiol Plantarum. 2002;116:73–8.
Cataldo DA, Garland TR, Wildung RE. Cadmium uptake kinetics in intact soybean plants. Plant Physiol. 1983;3:844–8.
Pence NS, Larsen PB, Ebbs SD, Letham DLD, Lasat MM, Garvin DF, et al. The molecular physiology of heavy metal transport in the Zn/cd hyperaccumulator Thlaspi caerulescens. Proc Natl Acad Sci U S A. 2000;97:4956–60.
Lasat MM, Baker AJM, Kochian LV. Altered zinc compartmentation in the root symplasm and stimulated Zn2+ absorption into the leaf as mechanisms involved in zinc hyperaccumulation in Thlaspi caerulescens. Plant Physiol. 1998;118:875–83.
Hanikenne M, Talke IN, Haydon MJ, Lanz C, Nolte A, Motte P, et al. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature. 2008;453:391–5.
Papoyan A, Piñeros M, Kochian LV. Plant Cd2+ and Zn2+ status effects on root and shoot heavy metal accumulation in Thlaspi caerulescens. New Phytol. 2007;175:51–8.
Xing JP, Jiang RE, Ueno D, Ma JF, Schat H, McGrath SP, et al. Variation in root-to-shoot translocation of cadmium and zinc among different accessions of the hyperaccumulators Thlaspi caerulescens and Thlaspi praecox. New Phytol. 2008;178:315–25.
Mori S, Uraguchi S, Ishikawa S, Arao T. Xylem loading process is a critical factor in determining cd accumulation in the shoots of Solanum melongena and Solanum torvum. Env Exp Bot. 2009;67:127–32.
Clemens S, Ma JF. Toxic heavy metal and metalloid accumulation in crop plants and foods. Ann Rev Plant Biol. 2016;67:489–512.
Laporte MA, Denaix L, Pagès L, Sterckeman T, Flénet F, Dauguet S, et al. Longitudinal variation in cadmium influx in intact first order lateral roots of sunflower (Helianthus annuus. L). Plant Soil. 2013;372:581–95.
Page V, Feller U. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat plants. Annals Bot. 2005;96:425–34.
Fodor F, Gáspár L, Morales F, Goforcena Y, Lucena JJ, Cseh E, et al. Effects of two iron sources on iron and cadmium allocation in poplar (Populus alba) plants exposed to cadmium. Tree Physiol. 2005;25:1173–80.
Yoshihara T, Hodoshima H, Miyano Y, Shoji K, Shimada H, Goto F. Cadmium inducible Fe deficiency responses observed from macro and molecular views in tobacco pants. Plant Cell Rep. 2006;25:365–73.
Barabasz A, Wilkowska A, Ruszczyńska A, Bulska E, Hanikenne M, Czarny M, et al. Metal response of transgenic tomato plants expressing P1B-ATPase. Physiol. Plantarum. 2012;145:315–31.
Kacperska A. Sensor types in signal transduction pathways in plant cells responding to abiotic stressors: do they depend on stress intensity? Physiol. Plantarum. 2004;122:159–68.
Ricachenevsky FK, Menguer PK, Sperotto RA, Fett JP. Got to hide your Zn away: molecular control of Zn accumulation and biotechnological applications. Plant Sci. 2015;236:1–17.
Guerinot ML. The ZIP family of metal transporters. Biochim Biophys Acta. 2000;1465:190–8.
Nishida S, Mizuno T, Obata H. Involvement of histidine-rich domain of ZIP family transporter TjZNT1 in metal ion specificity. Plant Physiol Bioch. 2008;46:601–6.
Pedas P, Schjoerring JK, Husted S. Identification and characterization of zinc-starvation-induced ZIP transporters from barley roots. Plant Physiol Biochem. 2009;47:377–83.
Lee S, Jeong HJ, Kim SA, Lee J, Guerinot ML, An G. OsZIP5 is a plasma membrane zinc transporter in rice. Plant Mol Biol. 2010;73:507–17.
Stephens BW, Cook DR, Grusak MA. Characterization of zinc transport by divalent metal transporters of the ZIP family from the model legume Medicago truncatula. Biometals. 2011;24:51–8.
Li S, Zhou X, Huang Y, Zhu L, Zhang S, Zhao Y, et al. Identification and characterization of the zinc-regulated transporters, iron-regulated transporter-like protein (ZIP) gene family in maize. BMC Plant Biol. 2013;13:114.
Tiong J, McDonald G, Genc Y, Shirley N, Langridge P, Chuang CY. Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytol. 2015;207:1–13.
Shingu Y, Kudo T, Ohsato S, Kimura M, Ono Y, Yamaguchi I, et al. Characterization of genes encoding metal tolerance proteins isolated from Nicotiana glauca and Nicotiana tabacum. Bioch Biophys Res Communic. 2005;331:675–80.
Hermand V, Julio E, deBorne FD, Punshon T, Ricachenevsky FK, Bellec A, et al. Inactivation of two newly identified tobacco heavy metal ATPases leads to reduced Zn and cd accumulation in shoots and reduced pollen germination. Metallomics. 2014;6:1427–40.
Eckhardt U, Marques AM, Buckhout TJ. Two iron-regulated cation transporters from tomato complement metal uptake-deficient yeast mutants. Plant Mol Biol. 2001;45:437–48.
Lin Y-F, Liang H-M, Yang S-Y, Boch A, Clemens S, Chen C-C, et al. Arabidopsis IRT3 is a zinc-regulated and plasma membrane localized zinc/iron transporter. New Phytol. 2009;182:392–404.
Siemianowski O, Barabasz A, Weremczuk A, Ruszczyńska A, Bulska E, Williams LE, et al. Development of Zn-related necrosis in tobacco is enhanced by expressing AtHMA4 and depends on the apoplastic Zn levels. Plant Cell Env. 2013;36:1093–104.
Barabasz A, Krämer U, Hanikenne M, Rudzka J, Antosiewicz DM. Metal accumulation in tobacco expressing Arabidopsis halleri metal hyperaccumulation gene depends on external supply. J Exp Bot. 2010;61:3057–67.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 2001;25:402–8.
Thompson JD, Higgins DG, Gibson TJ. Clustal-W – improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl Acids Res. 1994;22:4673–80.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30:2725–9.
Käll L, Krogh A, Sonnhammer ELL. A combined Transmembrane topology and signal peptide prediction method. J Mol Biol. 2004;338:1027–36.
Gietz DR, Schiestl RH. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nature Prot. 2007;3:31–3.
Acknowledgements
Not applicable.
Funding
This work was financially supported by National Science Center, Poland (Grant OPUS 8 No. 2014/15/B/NZ9/02303 to DMA). DMA wrote the grant proposal which has been accepted for financing by National Science Center, Poland.
Author information
Authors and Affiliations
Contributions
MP carried out all experiments, performed data analysis; AB contributed to the study concept, expression analysis and AAS based measurements, supervised the majority of experiments; AP, KK and KM contributed to hydroponic experiments; DMA designed the study concept, coordinated the research and supervised experiments, performed data analysis and wrote the manuscript. All authors critically revised the manuscript and approved the final version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1.
Dry weight of plant samples used to measure Zn and Cd concentrations
Additional file 2
Nucleotide and amino acid sequences of NtZIP5A/B and other selected ZIP5 from other plant species
Additional file 3
Sequence identity between the NtZIP5A, NtZIP5B and chosen ZIP5 and ZIP1 nucleotide sequences (a) and predicted proteins (b) from selected species
Additional file 4.
Supplementary information on cloning of NtZIP5B promoter (primers, promoter sequences)
Additional file 5.
Bioinformatics analysis of NtZIP5B subcellular localization
Additional file 6.
Graphical presentation of root part-specific changes in ZIPs expression
Additional file 7.
Stability of PP2A
Additional file 8.
Primer sequences used for expression analysis, cloning of NtZIP5B and promoter isolation
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Palusińska, M., Barabasz, A., Kozak, K. et al. Zn/Cd status-dependent accumulation of Zn and Cd in root parts in tobacco is accompanied by specific expression of ZIP genes. BMC Plant Biol 20, 37 (2020). https://doi.org/10.1186/s12870-020-2255-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-020-2255-3