
Summary
Three different protein prenyltransferases (farnesyltransferase and geranylgeranyltransferases I and II) catalyze the attachment of prenyl lipid anchors 15 or 20 carbons long to the carboxyl termini of a variety of eukaryotic proteins. Farnesyltransferase and geranylgeranyltransferase I both recognize a 'Ca1a2X' motif on their protein substrates; geranylgeranyltransferase II recognizes a different, non-CaaX motif. Each enzyme has two subunits. The genes encoding CaaX protein prenyltransferases are considerably longer than those encoding non-CaaX subunits, as a result of longer introns. Alternative splice forms are predicted to occur, but the extent to which each splice form is translated and the functions of the different resulting isoforms remain to be established. Farnesyltransferase-inhibitor drugs have been developed as anti-cancer agents and may also be able to treat several other diseases. The effects of these inhibitors are complicated, however, by the overlapping substrate specificities of geranylgeranyltransferase I and farnesyltransferase.
Similar content being viewed by others
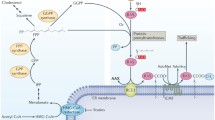

Gene organization and evolutionary history
Gene organization
Protein prenylation is the posttranslational attachment of either a farnesyl group or a geranylgeranyl group via a thioether linkage (-C-S-C-) to a cysteine at or near the carboxyl terminus of the protein. Farnesyl and geranylgeranyl groups are polyisoprenes, unsaturated hydrocarbons with a multiple of five carbons; the chain is 15 carbons long in the farnesyl moiety and 20 carbons long in the geranylgeranyl moiety (see Table 1). There are three different protein prenyltransferases in humans: farnesyltransferase (FT) and geranylgeranyltransferase 1 (GGT1) share the same motif (the CaaX box) around the cysteine in their substrates, and are thus called CaaX prenyltransferases, whereas geranylgeranyltransferase 2 (GGT2, also called Rab geranylgeranyltransferase) recognizes a different motif and is thus called a non-CaaX prenyltransferase [1]. Each protein consists of two subunits, α and β; the α subunit of FT and GGT1 is encoded by the same gene, FNTA (see Table 1).
The genomic organization of the human genes that encode protein prenyltransferases is shown in Figure 1. It is interesting to note that the genes encoding both subunits of non-CaaX prenyltransferases are much smaller (about 6-9 kilobases, kb) than those of the CaaX prenyltransferases (about 30-76 kb). The number of exons in the two types of genes are similar; the differences in gene size therefore result from differing intron lengths.

Gene structures and chromosomal locations of human protein prenyltransferase subunit genes. The chromosome diagrams and the locations of the genes on the cytogenetic map are according to the NCBI MapViewer [67]. The sizes of the genes are indicated but are not drawn to scale. (a) Genes encoding CaaX protein prenyltransferases are relatively long; (b) genes encoding non-CaaX protein prenyltransferases are much shorter.
Automatic comparisons of data from expressed sequence tags (ESTs) with genes (for example using the program Acembly, for which the results are available from the NCBI AceView server [2]) shows that all the human protein prenyltransferase genes have multiple alternative splice variants. The extent of translation of the various predicted transcripts and the structures and functions of the resulting proteins remain to be established experimentally; some of the predicted transcripts may be derived from missplicing rather than being real splice variants.
Evolutionary history
Protein prenyltransferases are currently known only in eukaryotes, but they are widespread, being found in vertebrates, insects, nematodes, plants, fungi and protozoa, including several parasites. The chromosomal locations and number of exons from protein prenyltransferase genes in the major eukaryotic model organisms are shown in Table 2. The α and β subunits have different folds, so are unlikely to have arisen from a common ancestor. Molecular phylogenetic analysis shows that orthologous proteins in different species are more closely related to each other than to their paralogs in the same species; the relationships between the β subunits are not fully clear. As well as the known genes, processed pseudogenes resulting from retrotransposition events have been found in the human and mouse genomes [3, 4].
The α subunits of protein prenyltransferases consist of tetratricopeptide repeats and are part of the tetratricopeptide repeat superfamily [5], which also includes functionally diverse proteins involved in transcription, co-chaperoning, protein transport, cell-cycle control and phosphorylation. Although evolution of repeat proteins is difficult to analyze and interpret, Zhang and Grishin [6] have deduced convincingly that the FNTA and RABGGTA genes originated from a common ancestor that already contained multiple tetratricopeptide repeats rather than having independently amplified the number of motifs as the families diverged over time.
The evolutionary history of the β subunits of protein prenyltransferases has received much less attention so far than that of the α subunits [7]. They can, however, be shown using standard sequence analysis tools such as PSI-Blast [8] or HMMer [9] to be part of a superfamily of prenyltransferases. Whereas the β subunits of protein prenyltransferases attach lipid anchors to proteins, other members of the superfamily catalyze different reactions involving polyisoprenes, such as the cyclization of polyisoprene derivatives during the synthesis of hopanoids in bacteria [10], cycloartenol in plants [11], ergosterol in fungi [12] and lanosterol in vertebrates [13, 14]; these products are then generally processed further to produce essential substances such as cholesterol, steroid hormones or vitamin D and their equivalents. A merged PFAM [15] domain (PF00432) has been created containing the β subunits of protein prenyltransferases, as well as squalene-hopene cyclases and lanosterol and cycloartenol synthases. Intriguingly, structural superposition of the conserved (α-α)6 barrel forming one half of bacterial squalene-hopene cyclase (the enzyme that synthesizes hopanoids) [16] with the β subunit of rat FT [17] reveals correspondence not only of their secondary structural elements but also of the parts of their active or binding sites (Figure 2). This would make sense if the different enzymes have retained common reaction mechanisms - or at least common substrate-binding characteristics - during evolution. Indeed, all of the members of the prenyltransferase superfamily bind substrates containing isoprenyl units, although they bind different numbers and variations of such units and the underlying processing and binding mechanisms seem to have diverged widely over time.

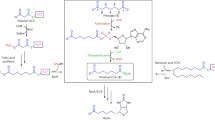
The structural relationship between protein prenyltransferase β subunits and squalene-hopene cyclases. (a) The complete structure of squalene-hopene cyclase from the bacterium Alicyclobacillus acidocaldarius (Protein Data Bank (PDB) identifier 3SQC [68]). (b) Structural superposition of the conserved (α-α)6 barrels of squalene-hopene cyclase (blue) and the rat FT β subunit (cyan; see Figure 3 for the full structure of the FT). Blue spheres, functionally important residues in the half of the squalene-hopene cyclase that is shown (the sphere indicates the van der Waals radius); pink spheres, farnesyl-pyrophosphate and Zn2+ bound to the β subunit of FT; pink residues in stick representation, the Ca1a2X peptide of the substrate protein. (c) Alignment of the two proteins on the basis of the structural superposition. Secondary structural elements are colored as in (b); black residues with colored background represent α helix and white residues with colored background represent β sheet. Conserved residues and similar residues are shown below the alignment; 1, polar; 2, small; 3, aromatic; 4, hydrophobic. The lower-case letters represent residues within 3SQC that were not easily superimposed on the 1D8D structure (that is, gaps in 1D8D). The molecular representations were created using VMD [69].
Characteristic structural features
The CaaX prenyltransferases FT and GGT1 and the non-CaaX prenyltransferase GGT2 share the same heterodimeric structure [1] (Figure 3). In the α subunits of both types of protein prenyltransferases, seven tetratricopeptide repeats are formed by pairs of helices (helices 2 to 15) that are stabilized by conserved intercalating residues. The α subunits of GGT2 in mammals and plants also have an immunoglobulin-like domain between the fifth and sixth tetratricopeptide repeat, as well as leucine-rich repeats at the carboxyl terminus. The functions of these additional domains in GGT2 are as yet undefined, but they are apparently not directly involved in the interaction with substrates and Rab escort proteins (see below) [18, 19]. The tetratricopeptide repeats of the α subunit form a right-handed superhelix, which embraces the (α-α)6 barrel of the β subunit [20]. The β subunits include most of the substrate- and lipid-binding pockets [20] and their tight association with the respective α subunits is required for proper function [21]. Compared with FT, GGT2 has a larger hydrophobic pocket in the β subunit to accommodate the longer lipid [22].

It is difficult to estimate the effect of alternative splicing on the structure of protein prenyltransferases. We would expect that the integrity of the structure of the β subunits would be more sensitive to non-terminal truncations than are the α subunits, because the modular structure of the tetratricopeptide repeat motifs in the α subunits would allow truncations and additions without severe consequences for the hydrophobic packing of the structure.
Enzyme mechanism
CaaX prenyltransferases recognize the carboxy-terminal Ca1a2X motif (see Table 1) [23, 24] of substrate proteins, usually after binding farnesyl-pyrophosphate [25] or geranylgeranylpyrophosphate [26]. The lipid anchors are then transferred by a catalytic mechanism that depends on formation of a complex between a Zn2+ cation and the cysteine of the Ca1a2X motif [20]. High concentrations of Mg2+ are required for optimal enzymatic activity of FT [27], though this is apparently not the case for GGT1 [28]. The Zn2+ is suggested to be required for the proper conformation of the substrate peptide [17]. The major conformational change in the transfer step seems to be a rotation of the prenylpyrophosphate in the binding pocket and not of parts of the enzyme itself. A detailed picture of the reaction pathway that involves electrophilic and nucleophilic mechanisms is given by a series of structures representing the different states [28] as well as by kinetic measurements [29].
In contrast to FT and GGT1, GGT2 does not require a very specific carboxy-terminal motif [30] apart from the availability of several cysteines close to the carboxyl terminus that are often arranged -CC, -CXC, -CCX, -CCXX, -CCXXX or, in a few cases, with only a single cysteine as in -CXXX. If the motif consists of two cysteines in close proximity, two geranylgeranyl moieties are usually added.
GGT2 recognizes the structural features of a complex of the substrate with an escort protein (Rab escort protein (REP), previously known as component A) and then scans the carboxyl terminus for prenylatable cysteines [30]. The catalytic mechanism of lipid transfer from geranylgeranylpyrophosphate to the protein substrate also requires Zn2+, and the following model has been presented for how the double geranylgeranylation could take place on the basis of insights from the reaction pathway of FT [28]. After attachment of the first prenyl group, the lipid chain is translocated over the enzyme surface into another hydrophobic groove upon binding of the second geranylgeranylpyrophosphate. Finally, binding of a third geranylgeranylpyrophosphate releases the whole complex of the now doubly geranylgeranylated substrate with its escort protein; the escort protein is also involved in the transport of the substrate to the target membrane [31, 32].
Localization and function
The results of systematic oligonucleotide microarray experiments catalogued in the human gene-expression index database [33, 34] show that protein prenyltransferases are expressed in a variety of tissues. Both CaaX prenyltransferases seem to be active (that is, their α and β subunits are coexpressed) in a range of tissues, and the non-CaaX prenyltransferase subunits are also expressed in several more tissues. It should be noted, however, that many of the expression levels listed [33, 34] are close to the detection threshold and could therefore result from cross-hybridization between close homologs rather than true expression. Interestingly, the α subunits also appear to be expressed in tissues that lack expression of the corresponding β subunits; this suggests that single subunits, or isoforms of them, might have additional, prenylation-independent functions in the cell.
As the α subunit of CaaX prenyltransferases is shared between FT and GGT1, its expression must be higher than that of each β subunit if it is to form 1:1 complexes with the β subunits of both enzymes. These higher expression levels seem to be transcriptionally regulated by different promoters; in order to produce recombinant CaaX prenyltransferases in the laboratory, it is thus necessary to downregulate expression of the α subunits when coexpressing with β subunits [35].
Lipid anchors are common posttranslational modifications that can direct the subcellular localization of proteins. Other lipid modifications, such as myristoylation [36–38], palmitoylation [39, 40] and glycosylphosphatidylinositol (GPI) anchors [41, 42], are mainly important for attachment of the protein to membranes, but lipid modification by protein prenyltransferases seems to have a more complex role: the farnesyl and geranylgeranyl moieties attached to the substrates are directly involved in protein-protein interactions as well as in protein-membrane interactions [43]. The importance of protein prenyltransferases is illustrated by the involvement of their substrates in critical cellular pathways and diseases [44].
Substrates and functions of CaaX prenyltransferases
Typical substrates that are farnesylated by FT include many members of the Ras superfamily of small GTPases (H-Ras, K-Ras, N-Ras, Ras2, Rap2, RhoB (which is also geranylgeranylated), RhoE, Rheb, TC10, and TC21), as well as the nuclear lamina proteins lamin A and B, the kinetochore proteins CENP-E and CENP-F, fungal mating factors, cGMP phosphodiesterase α, γ subunit variants of G proteins, DnaJ heat-shock protein homologs, rhodopsin kinase, the peroxisomal membrane proteins Pex19 and PxF and paralemmin (a neural protein suggested to be involved in membrane dynamics). GGT1 preferentially geranylgeranylates some of the other small GTPases (such as Rac1, Rac2, RalA, Rap1A, Rap1B, RhoA, and RhoB (which is also farnesylated, as noted above), RhoC, Cdc42, Rab8 (which is also geranylgeranylated by GGT2), Rab11, and Rab13, as well as some γ-subunit variants of G proteins, cGMP phosphodiesterase β and the plant calmodulin CaM53. Typically, prenylation by CaaX protein prenyltransferases is accompanied by further posttranslational processing, most often involving cleavage of the carboxy-terminal tripeptide (-a1a2X) followed by carboxymethylation of the carboxyl terminus [45–47]. Palmitoylation is another modification that sometimes takes place after prenylation [48].
Because several prenylated substrates are involved in diseases, inhibition of protein prenyltransferases has great potential for medical applications. A boom in the field was triggered by the finding that inhibition of FT in mice that have tumors derived from H-Ras-transformed cells leads to tumor regression, while the inhibitor has no adverse effect on the organism [49]. This led to successful completion of clinical phase I trials of farnesyl transferase inhibitors (FTIs), but in phase II trials the efficacy of the inhibitors towards a broad spectrum of different cancer cells (such as K-Ras-transformed cells) was far below the high expectations that arose from the phase I trials. Surprisingly, however, beneficial effects were found for other, non-neoplastic diseases; for example, diabetic retinopathy and macular degeneration [50]. The unexpected physiological effects of FT inhibition are partly due to a striking cross-specificity between the two CaaX prenyltransferases: both FT and GGT1 can use either farnesyl-pyrophosphate or GGPP to a certain extent to transfer lipids to several of each others' substrates as well as their own [51, 52], and several substrates can be either farnesylated or geranylgeranylated. The substrates probably compete in vivo for the enzymes loaded with the preferred polyprenylpyrophosphate, and the type of modification that is added depends on the relative affinity of the substrates for the enzymes.
In cancer cells, FTIs are known to affect growth in soft agar (anchorage-independent growth), cell-cycle progression at the G1-S phase and the G2-M phase checkpoints, the morphology of the cytoskeleton (formation of actin stress fibers) and induction of apoptosis. The substrate proteins that have been suggested to be involved in these effects include H-Ras [49], Rheb [53], CENP-E [54] and RhoB [55]; FTI effects are reviewed in more detail elsewhere [56]. The effects of FT inhibition in cells transformed with H-Ras differ from those on cells transformed with K-Ras. This difference has been attributed to the ability of GGT1 to alternatively prenylate some proteins, through the cross-specificity mentioned above, including K-Ras but not H-Ras (see Figure 4). RhoB can be both farnesylated and geranylgeranylated; Prendergast and colleagues [57] have suggested that the different levels of farnesylated and geranylgeranylated RhoB in FTI-treated cells compared with untreated cells could contribute to the observed effects of FTIs.

Differential prenylation of substrates of CaaX protein prenyltransferases (a) under normal conditions and (b) in the presence of a FT inhibitor. Substrates are shown in the middle of each panel, with the two enzymes above and below. 'Vital-F' and 'Vital-GG' represent all the proteins for which farnesylation (Vital-F) or geranylgeranylation (Vital-GG) are essential; RhoB-GG represents geranylgeranylated RhoB and RhoB-F represents farnesylated RhoB. Thick arrow, normal prenylation activity; thin arrow, reduced prenylation activity through cross-specificity of the non-preferred enzyme; arrows with crosses, blocked prenylation activity during inhibition of FT; brackets, substituting prenylation activity by GGT1 during inhibition of FT.
Substrates and functions of non-CaaX prenyltransferases
The main substrates for prenylation by GGT2 are the Rab family of proteins, the largest group of small GTPases in the Ras superfamily. There are at least 60 different Rabs in humans [58]. They interact with the Rab escort protein REP, which is required for the prenylation of Rabs by GGT2 [30], and are involved in the docking of transport vesicles to their specific target membranes [59].
As with CaaX protein prenyltransferases, deficiencies in prenylation by non-CaaX protein prenyltransferases are relevant to diseases [59, 60]. A mutation inactivating a start codon of the major transcript of the α subunit of GGT2 is one of the many mutations involved in the recessively inherited Hermansky-Pudlak syndrome and related disorders [61, 62], in which platelet synthesis, platelet organelle function and pigmentation are affected. X-linked choroideremia (CHM) results in retinal degeneration, with symptoms starting from night blindness in young people and progressing over decades until vision is completely lost [63]. It is caused by loss-of-function mutations in the CHM gene, which encodes Rab escort protein 1 (REP1) [64]. Loss-of function mutations in the Rab27a gene cause Griscelli syndrome, whose symptoms are similar to Hermansky-Pudlak syndrome and other diseases associated with insufficient Rab prenylation [60].
Frontiers
There are several issues that merit further study in the regulation of protein prenyltransferases. Firstly, it is not clear how the concomitant transcription of the two subunits from two different chromosomes is regulated or where and how the subunits meet to build up functional prenyltransferases. Secondly, given that there are multiple splice variants, it is likely that additional variants of subunits will be found to have distinct functions or regulatory roles; an example is a variant of the FT/GGT1 α subunit that has been reported to be directly involved in signaling by transforming growth factor β and activin [65]. Interpretation of results in areas ranging from molecular biology to clinical trials must take into account possible isoforms with varying functions or altered interactions to avoid erroneous conclusions.
A third issue is the striking differences in gene size and intron length between the two types of protein prenyltransferases. One of several possible factors that could have caused this is a difference in evolutionary selection pressures. Whereas FT and GGT1 partly compensate each other functionally, there is no counterpart for GGT2. Furthermore, formation of a complex between the substrate and an escort protein is necessary for recognition by GGT2 and the conservation of additional binding sites at the surface is therefore required. Also, the severity of the effect when the prenylation of different substrates is abolished may vary. Finally, the size of the genomic region containing the gene might alter its accessibility to the transcription machinery and the time needed to complete transcription, so gene size may affect or be affected by expression levels. The implications of these factors for the exact evolutionary history of the protein prenyltransferase genes (such as the relative ages of the subunits and the order of duplication events) remain to be established.
Finally, more research is also needed on the effects of FTIs. After the rush to develop inhibitors, basic research is now needed as well as clinical trials in order to improve the understanding of the basic processes involved [66]. For example, it cannot be ruled out that some effects of FTIs are not a direct consequence of inhibiting prenylation but are instead due to cross-reactivity with proteins from completely different pathways. It is tempting to speculate that one of the proteins that are evolutionarily related to the protein prenyltransferases (such as other prenyltransferases) could be affected by FTIs; the selectivity of existing FTIs, which do not inhibit even the much more closely related GGTs, makes this scenario most unlikely, however. The next task is to identify clearly the proteins whose altered prenylation causes the observed effects of FT inhibition. Given the multiplicity and heterogeneity of these effects, it is clear that they cannot be attributed to one single farnesylated protein that lacks a lipid modification because of FT inhibition; rather, alterations in the function of several proteins probably cause the observed effects, with variations depending on the cell type, disease and organism. Further research may eventually lead to FTIs being used successfully to treat cancers and other diseases.
References
Casey PJ, Seabra MC: Protein prenyltransferases. J Biol Chem. 1996, 271: 5289-5292. 10.1074/jbc.271.10.5289. A short minireview describing protein prenyltransferases, written before the structure was known.
AceView. Alignment of expressed sequence tags and mRNAs to the human genome, showing alternative splice forms., [http://www.humangenes.org]
Andres DA, Milatovich A, Ozcelik T, Wenzlau JM, Brown MS, Goldstein JL, Francke U: cDNA cloning of the two subunits of human CAAX farnesyltransferase and chromosomal mapping of FNTA and FNTB loci and related sequences. Genomics. 1993, 18: 105-112. 10.1006/geno.1993.1432. Cloning of human FNTA and FNTB; 'related sequences' refers to processed pseudogenes.
Dhawan P, Yang E, Kumar A, Mehta KD: Genetic complexity of the human geranylgeranyltransferase I beta-subunit gene: a multigene family of pseudogenes derived from mis-spliced transcripts. Gene. 1998, 210: 9-15. 10.1016/S0378-1119(98)00042-0. The authors suggest that there are 13 GGT1B pseudogenes, but these seem to correspond to only two in the human genome.
Blatch GL, Lassle M: The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. BioEssays. 1999, 21: 932-939. 10.1002/(SICI)1521-1878(199911)21:11<932::AID-BIES5>3.3.CO;2-E. Review describing the family of TPR-containing proteins.
Zhang H, Grishin NV: The alpha-subunit of protein prenyltransferases is a member of the tetratricopeptide repeat family. Protein Sci. 1999, 8: 1658-1667. Evolutionary history and relationships of the FT α subunit.
Liang PH, Ko TP, Wang AH: Structure, mechanism and function of prenyltransferases. Eur J Biochem. 2002, 269: 3339-3354. 10.1046/j.1432-1033.2002.03014.x. A review giving a quick overview of isoprenylpyrophosphate synthases, isoprenyl cyclases and protein prenyltransferases that share processing polyisoprene derivatives in some way with partly overlapping structures and reaction mechanisms.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ: Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997, 25: 3389-3402. 10.1093/nar/25.17.3389. The standard reference for the sequence similarity search tools of the BLAST suite.
Eddy SR: Profile hidden Markov models. Bioinformatics. 1998, 14: 755-763. 10.1093/bioinformatics/14.9.755. A sensitive alternative method to BLAST for sequence similarity searches.
Wendt KU, Poralla K, Schulz GE: Structure and function of a squalene cyclase. Science. 1997, 277: 1811-1815. 10.1126/science.277.5333.1811. The first crystal structure of a squalene cyclase.
Nes WD, Venkatramesh M: Enzymology of phytosterol transformations. Crit Rev Biochem Mol Biol. 1999, 34: 81-93. A review on plant hormone synthesis, including the cycloartenol synthases, which are homologous to the beta subunit of protein prenyltranserases.
Sturley SL: Conservation of eukaryotic sterol homeostasis: new insights from studies in budding yeast. Biochim Biophys Acta. 2000, 1529: 155-163. 10.1016/S1388-1981(00)00145-1. A review including discussion of ergosterol synthesis in fungi.
Janowski BA, Willy PJ, Devi TR, Falck JR, Mangelsdorf DJ: An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature. 1996, 383: 728-731. 10.1038/383728a0. A description of intermediate steps in steroid hormone and cholesterol synthesis.
Kim JH, Lee JN, Paik YK: Cholesterol biosynthesis from lanosterol. A concerted role for Sp1 and NF-Y-binding sites for sterol-mediated regulation of rat 7- dehydrocholesterol reductase gene expression. J Biol Chem. 2001, 276: 18153-18160. 10.1074/jbc.M101661200. Analysis of the cholesterol biosynthesis pathway, starting from lanosterol.
Bateman A, Birney E, Durbin R, Eddy SR, Howe KL, Sonnhammer EL: The Pfam protein families database. Nucleic Acids Res. 2000, 28: 263-266. 10.1093/nar/28.1.263. A protein families database automatically generated by clustering proteins into families according to similarity identified with hidden Markov models.
Wendt KU, Lenhart A, Schulz GE: The structure of the membrane protein squalene-hopene cyclase at 2.0 Å resolution. J Mol Biol. 1999, 286: 175-187. 10.1006/jmbi.1998.2470. High-resolution structure of squalene-hopene cyclase plus discussion of catalytic mechanism and membrane interaction surfaces.
Long SB, Casey PJ, Beese LS: The basis for K-Ras4B binding specificity to protein farnesyltransferase revealed by 2 Å resolution ternary complex structures. Structure Fold Des. 2000, 8: 209-222. 10.1016/S0969-2126(00)00096-4. The first crystal structure of FT in complex with farnesyl-pyrophosphate.
Dursina B, Thoma NH, Sidorovitch V, Niculae A, Iakovenko A, Rak A, Albert S, Ceacareanu AC, Kolling R, Herrmann C, et al: Interaction of yeast Rab geranylgeranyl transferase with its protein and lipid substrates. Biochemistry. 2002, 41: 6805-6816. 10.1021/bi016067w. Shows that immunoglobulin-like and leucine-rich repeat domains are not required for non-CaaX prenylation.
Pylypenko O, Rak A, Reents R, Niculae A, Sidorovitch V, Cioaca MD, Bessolitsyna E, Thoma NH, Waldmann H, Schlichting I, et al: Structure of rab escort protein-1 in complex with rab geranylgeranyltransferase. Mol Cell. 2003, 11: 483-494. Structure of the complex of GGT2 with REP1.
Park HW, Boduluri SR, Moomaw JF, Casey PJ, Beese LS: Crystal structure of protein farnesyltransferase at 2.25 angstrom resolution. Science. 1997, 275: 1800-1804. 10.1126/science.275.5307.1800. The first crystal structure of protein FT.
Andres DA, Goldstein JL, Ho YK, Brown MS: Mutational analysis of alpha-subunit of protein farnesyltransferase. Evidence for a catalytic role. J Biol Chem. 1993, 268: 1383-1390. The first evidence for residues of the α subunit being involved in catalysis.
Zhang H, Seabra MC, Deisenhofer J: Crystal structure of Rab geranylgeranyltransferase at 2.0 Å resolution. Structure Fold Des. 2000, 8: 241-251. 10.1016/S0969-2126(00)00102-7. The first crystal structure of GGT2.
Moores SL, Schaber MD, Mosser SD, Rands E, O'Hara MB, Garsky VM, Marshall MS, Pompliano DL, Gibbs JB: Sequence dependence of protein isoprenylation. J Biol Chem. 1991, 266: 14603-14610. Detailed analysis of the amino-acid composition of the motifs recognized by protein prenyltransferases.
Caplin BE, Hettich LA, Marshall MS: Substrate characterization of the Saccharomyces cerevisiae protein farnesyltransferase and type-I protein geranylgeranyltransferase. Biochim Biophys Acta. 1994, 1205: 39-48. 10.1016/0167-4838(94)90089-2. Characterization of substrate motifs in yeast and the cross-specificity between FT and GGT1.
Reiss Y, Goldstein JL, Seabra MC, Casey PJ, Brown MS: Inhibition of purified p21ras farnesyl:protein transferase by Cys-AAX tetrapeptides. Cell. 1990, 62: 81-88. An early biochemical analysis of FT including farnesyl-pyrophosphate binding and inhibition with CaaX tetrapeptides.
Yokoyama K, McGeady P, Gelb MH: Mammalian protein geranylgeranyltransferase-I: substrate specificity, kinetic mechanism, metal requirements, and affinity labeling. Biochemistry. 1995, 34: 1344-1354. Complete biochemical characterization of mammalian GGT1.
Saderholm MJ, Hightower KE, Fierke CA: Role of metals in the reaction catalyzed by protein farnesyltransferase. Biochemistry. 2000, 39: 12398-12405. 10.1021/bi0011781. The metal requirements of FT catalysis.
Long SB, Casey PJ, Beese LS: Reaction path of protein farnesyltransferase at atomic resolution. Nature. 2002, 419: 645-650. 10.1038/nature00986. Completion of a series of structures resembling intermediate steps of the reaction pathway of FT. Discussion of implications for all three protein prenyltransferases.
Furfine ES, Leban JJ, Landavazo A, Moomaw JF, Casey PJ: Protein farnesyltransferase: kinetics of farnesyl pyrophosphate binding and product release. Biochemistry. 1995, 34: 6857-6862. Kinetic analysis of the reaction pathway of FT.
Alexandrov K, Simon I, Yurchenko V, Iakovenko A, Rostkova E, Scheidig AJ, Goody RS: Characterization of the ternary complex between Rab7, REP-1 and Rab geranylgeranyl transferase. Eur J Biochem. 1999, 265: 160-170. 10.1046/j.1432-1327.1999.00699.x. GGT2 prenylation requires formation of complex of GGT2 with substrate and escort protein.
Alexandrov K, Horiuchi H, Steele-Mortimer O, Seabra MC, Zerial M: Rab escort protein-1 is a multifunctional protein that accompanies newly prenylated rab proteins to their target membranes. EMBO J. 1994, 13: 5262-5273. Rab escort protein is not only required for the prenylation reaction but also accompanies Rab substrates to the membrane. This article also presents its homology to the GDP dissociation inhibitor.
Benito-Moreno RM, Miaczynska M, Bauer BE, Schweyen RJ, Ragnini A: Mrs6p, the yeast homologue of the mammalian choroideraemia protein: immunological evidence for its function as the Ypt1p Rab escort protein. Curr Genet. 1994, 27: 23-25. Characterization of the yeast Rab/Ypt escort protein.
Haverty PM, Weng Z, Best NL, Auerbach KR, Hsiao LL, Jensen RV, Gullans SR: HugeIndex: a database with visualization tools for high-density oligonucleotide array data from normal human tissues. Nucleic Acids Res. 2002, 30: 214-217. 10.1093/nar/30.1.214. Systematic mRNA expression analysis of human tissues.
HuGE Index. A database of systematic mRNA expression analyses of human tissues., [http://hugeindex.org/]
Tsao KL, Waugh DS: Balancing the production of two recombinant proteins in Escherichia coli by manipulating plasmid copy number: high-level expression of heterodimeric Ras farnesyltransferase.. Protein Expr Purif. 1997, 11: 233-240. 10.1006/prep.1997.0794. The α subunit has to be downregulated when coexpressed with the β subunit in order to yield high levels of recombinant FT.
Gordon JI, Duronio RJ, Rudnick DA, Adams SP, Gokel GW: Protein N-myristoylation. J Biol Chem. 1991, 266: 8647-8650. Compact review on protein N-myristoylation.
Maurer-Stroh S, Eisenhaber B, Eisenhaber F: N-terminal N-myristoylation of proteins: refinement of the sequence motif and its taxon-specific differences. J Mol Biol. 2002, 317: 523-540. 10.1006/jmbi.2002.5425. A detailed update of the sequence motif for N-myristoylation as well as enzyme-substrate interactions.
Maurer-Stroh S, Eisenhaber B, Eisenhaber F: N-terminal N-myristoylation of proteins: prediction of substrate proteins from amino acid sequence. J Mol Biol. 2002, 317: 541-557. 10.1006/jmbi.2002.5426. Large-scale prediction unveils a series of new target proteins for protein N-myristoylation.
Morello JP, Bouvier M: Palmitoylation: a post-translational modification that regulates signalling from G-protein coupled receptors. Biochem Cell Biol. 1996, 74: 449-457. A review on the importance of palmitoylation in G-protein signaling.
Resh MD: Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim Biophys Acta. 1999, 1451: 1-16. 10.1016/S0167-4889(99)00075-0. A short review on myristoyl and palmitoyl anchors for membrane targeting.
Chatterjee S, Mayor S: The GPI-anchor and protein sorting. Cell Mol Life Sci. 2001, 58: 1969-1987. A review on glycosylphosphatidylinositol lipid anchors.
Eisenhaber B, Bork P, Eisenhaber F: Post-translational GPI lipid anchor modification of proteins in kingdoms of life: analysis of protein sequence data from complete genomes. Protein Eng. 2001, 14: 17-25. 10.1093/protein/14.1.17. Full genome analyses of GPI-anchor-containing proteins.
Sinensky M: Functional aspects of polyisoprenoid protein substituents: roles in protein-protein interaction and trafficking. Biochim Biophys Acta. 2000, 1529: 203-209. 10.1016/S1388-1981(00)00149-9. Role of farnesyl and geranylgeranyl anchors in protein-protein interactions.
Sinensky M: Recent advances in the study of prenylated proteins. Biochim Biophys Acta. 2000, 1484: 93-106. 10.1016/S1388-1981(00)00009-3. Review on enzymology and function of protein prenylation.
Trueblood CE, Boyartchuk VL, Picologlou EA, Rozema D, Poulter CD, Rine J: The CaaX proteases, Afc1p and Rce1p, have overlapping but distinct substrate specificities. Mol Cell Biol. 2000, 20: 4381-4392. 10.1128/MCB.20.12.4381-4392.2000. Characterization of endoproteases cleaving the carboxy-terminal tripeptide after prenylation of cysteine in the CaaX motif.
Pei J, Grishin NV: Type II CAAX prenyl endopeptidases belong to a novel superfamily of putative membrane-bound metalloproteases. Trends Biochem Sci. 2001, 26: 275-277. 10.1016/S0968-0004(01)01813-8. A description of a protein superfamily that includes CaaX prenyl endoproteases.
Bergo MO, Leung GK, Ambroziak P, Otto JC, Casey PJ, Gomes AQ, Seabra MC, Young SG: Isoprenylcysteine carboxyl methyltransferase deficiency in mice. J Biol Chem. 2001, 276: 5841-5845. 10.1074/jbc.C000831200. A description of the methyltransferase that carboxymethylates cysteines at the carboxyl terminus of prenylated proteins.
Hancock JF, Magee AI, Childs JE, Marshall CJ: All ras proteins are polyisoprenylated but only some are palmitoylated. Cell. 1989, 57: 1167-1177. Palmitoylation of Ras proteins sometimes takes place after prenylation.
Kohl NE, Omer CA, Conner MW, Anthony NJ, Davide JP, deSolms SJ, Giuliani EA, Gomez RP, Graham SL, Hamilton K, et al: Inhibition of farnesyltransferase induces regression of mammary and salivary carcinomas in ras transgenic mice. Nat Med. 1995, 1: 792-797. FT inhibition causes tumor regression in H-Ras transforming mice with minimal side-effects on the organism.
Prendergast GC, Rane N: Farnesyltransferase inhibitors: mechanism and applications. Expert Opin Investig Drugs. 2001, 10: 2105-2116. Detailed review on FTI, including their effects on non-neoplastic diseases.
Armstrong SA, Hannah VC, Goldstein JL, Brown MS: CAAX geranylgeranyl transferase transfers farnesyl as efficiently as geranylgeranyl to RhoB. J Biol Chem. 1995, 270: 7864-7868. 10.1074/jbc.270.43.25879. Reports that GGT1 can farnesylate RhoB as well as geranylgeranylating it, a surprising finding.
Yokoyama K, Zimmerman K, Scholten J, Gelb MH: Differential prenyl pyrophosphate binding to mammalian protein geranylgeranyltransferase-I and protein farnesyltransferase and its consequence on the specificity of protein prenylation. J Biol Chem. 1997, 272: 3944-3952. 10.1074/jbc.272.1.503. Detailed analysis of prenyl pyrophosphate cross-specificity of FT and GGT1.
Tamanoi F, Kato-Stankiewicz J, Jiang C, Machado I, Thapar N: Farnesylated proteins and cell cycle progression. J Cell Biochem suppl. 2001, Suppl 37: 64-70. 10.1002/jcb.10067. The involvement of farnesylated proteins in the cell cycle in the light of the effects of FT inhibition.
Ashar HR, James L, Gray K, Carr D, Black S, Armstrong L, Bishop WR, Kirschmeier P: Farnesyl transferase inhibitors block the farnesylation of CENP-E and CENP-F and alter the association of CENP-E with the microtubules. J Biol Chem. 2000, 275: 30451-30457. 10.1074/jbc.M003469200. A lack of prenylation of CENP-E affects microtubule-centromere interaction at the G2/M phase checkpoint.
Lebowitz PF, Casey PJ, Prendergast GC, Thissen JA: Farnesyltransferase inhibitors alter the prenylation and growth-stimulating function of RhoB. J Biol Chem. 1997, 272: 15591-15594. 10.1074/jbc.272.25.15591. Altering the prenylation status of RhoB alters its functions.
Tamanoi F, Gau CL, Jiang C, Edamatsu H, Kato-Stankiewicz J: Protein farnesylation in mammalian cells: effects of farnesyltransferase inhibitors on cancer cells. Cell Mol Life Sci. 2001, 58: 1636-1649. A review on cellular processes affected by FT inhibition in mammalian cancers.
Prendergast GC: Actin' up: RhoB in cancer and apoptosis. Nat Rev Cancer. 2001, 1: 162-168. 10.1038/35101096. A short review of RhoB function, including a hypothesis on the effects of FTIs.
Pereira-Leal JB, Seabra MC: Evolution of the Rab family of small GTP-binding proteins. J Mol Biol. 2001, 313: 889-901. 10.1006/jmbi.2001.5072. An extensive analysis of Rab proteins in eukaryotic genomes.
Seabra MC, Mules EH, Hume AN: Rab GTPases, intracellular traffic and disease. Trends Mol Med. 2002, 8: 23-30. 10.1016/S1471-4914(01)02227-4. An update of Rab functions and localizations, plus their involvement in disease.
Pereira-Leal JB, Hume AN, Seabra MC: Prenylation of Rab GTPases: molecular mechanisms and involvement in genetic disease. FEBS Lett. 2001, 498: 197-200. 10.1016/S0014-5793(01)02483-8. A minireview of Rab prenylation deficiency and disease.
Huizing M, Anikster Y, Gahl WA: Hermansky-Pudlak syndrome and related disorders of organelle formation. Traffic. 2000, 1: 823-835. 10.1034/j.1600-0854.2000.011103.x. A review on Hermansky-Pudlak syndrome and related disorders.
Detter JC, Zhang Q, Mules EH, Novak EK, Mishra VS, Li W, McMurtrie EB, Tchernev VT, Wallace MR, Seabra MC, et al: Rab geranylgeranyl transferase alpha mutation in the gunmetal mouse reduces Rab prenylation and platelet synthesis. Proc Natl Acad Sci USA. 2000, 97: 4144-4149. 10.1073/pnas.080517697. Genetic and molecular basis for Rab prenylation deficiency in Hermansky-Pudlak syndrome.
Seabra MC: New insights into the pathogenesis of choroideremia: a tale of two REPs. Ophthalmic Genet. 1996, 17: 43-46. The mechanism of choroideremia pathology involves an inability of Rab escort protein 2 to fully substitute for function of Rab escort protein 1.
van den Hurk JA, Schwartz M, van Bokhoven H, van de Pol TJ, Bogerd L, Pinckers AJ, Bleeker-Wagemakers EM, Pawlowitzki IH, Ruther K, Ropers HH, et al: Molecular basis of choroideremia (CHM): mutations involving the Rab escort protein-1 (REP-1) gene. Hum Mutat. 1997, 9: 110-117. 10.1002/(SICI)1098-1004(1997)9:2<110::AID-HUMU2>3.3.CO;2-9. A loss-of-function mutation in the CHM gene that codes for Rab escort protein 1 results in choroideremia.
Wang T, Danielson PD, Li BY, Shah PC, Kim SD, Donahoe PK: The p21(RAS) farnesyltransferase alpha subunit in TGF-beta and activin signaling. Science. 1996, 271: 1120-1122. An isoform of FT α subunit that has a non-classical function.
Cox AD, Der CJ: Farnesyltransferase inhibitors: promises and realities. Curr Opin Pharmacol. 2002, 2: 388-393. 10.1016/S1471-4892(02)00181-9. A review on the pitfalls of FT inhibition.
Homo sapiens genome view. Graphical display of features of the human genome sequence., [http://www.ncbi.nlm.nih.gov/mapview/map_search.cgi?]
Protein Data Bank: structure explorer - 3SQC. Information on the three-dimensional structure of bacterial squalene-hopene cyclase., [http://www.rcsb.org/pdb/cgi/explore.cgi?pdbId=3SQC]
Visual Molecular Dynamics. Free software for displaying, animating, and analyzing large biomolecular systems., [http://www.ks.uiuc.edu/Research/vmd/]
Protein Data Bank: structure explorer - 1D8D. Information on the three-dimensional structure of rat FT., [http://www.rcsb.org/pdb/cgi/explore.cgi?pdbId=1D8D]
Acknowledgements
The authors are grateful for useful comments from Maria Novatchkova and Alexander Schleiffer as well as for continuous support from Boehringer Ingelheim. This project has been partly funded by the Fonds zur Förderung der wissenschaftlichen Forschung Österreichs (FWF grant P15037) and by the Austrian National Bank (OeNB).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Maurer-Stroh, S., Washietl, S. & Eisenhaber, F. Protein prenyltransferases. Genome Biol 4, 212 (2003). https://doi.org/10.1186/gb-2003-4-4-212
Published:
DOI: https://doi.org/10.1186/gb-2003-4-4-212