
Abstract
Considering the pivotal role played by erosive organisms in the marine habitat and the scanty knowledge of this phenomenon in the Mediterranean Sea, the present study aimed to identify the pioneer excavating organisms occurring in the first stages of bioerosion, providing the first estimation of their rate of erosion along the North Adriatic Sea. Bioerosion activity was investigated by deploying sets of limestone panels for a period of 2 years. Moreover, micro-computed tomography was used for the first time to study polychaete erosion, confirming it is a very useful tool for the description of erosion processes. Blocks were placed in the touristic harbor Marina Dorica (Ancona) in July 2017 and periodically collected, the first time after a 6-month period, then bimonthly. Two species of the polychaete genus Polydora were the only boring organisms that settled on the experimental substrates. Erosion rate was estimated by measuring the holes produced by both worm species, and the eroded volumes obtained by resin casts of the holes. Polydora appeared after 6 months of exposure significantly changing its density rates (holes·100 cm−2·month−1) during the exposure, being comparable with densities reported from tropical areas. The erosion rate on the contrary is lower, reaching values of 0.053 g·cm−2·year−1.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Destruction of carbonate substrates due to organism activities, or bioerosion (Neumann, 1966), is a common process in several marine ecosystems from temperate (Cerrano et al., 2001a; Wisshak et al., 2005; Schönberg & Wisshak, 2014), to tropical (Hutchings et al., 1992, 2013; Hutchings & Peyrot-Clausade, 2002; Pari et al., 2002; Tribollet et al., 2002; Hutchings, 2008) and even polar regions (Cerrano et al., 2001b; Meyer et al., 2021). Boring organisms participate to a wide range of regulation and maintenance ecosystem services: in fact, they enhance habitat complexity offering refuge to several taxa and contribute to the carbonate cycle, with the erosion of coastal areas and the production of fine sediments (Naylor et al., 2012; Davidson et al., 2018). Moreover, considering that their boring activity can be influenced by a variety of environmental factors (e.g., nutrients, salinity, temperature, pH), in the context of the current climate crisis, the study of the bioerosion processes became a timely topic (Chazottes et al., 2008; Schönberg & Wisshak, 2014; Schönberg et al., 2017).
Schönberg & Wisshak (2014) reported that only the 11% of the research on bioerosion were conducted in the Mediterranean Sea; since then, only 7 papers have been found on SCOPUS (https://www.scopus.com) using “bioerosion,” “boring species,” and “Mediterranean” as search terms. In particular, studies about the rates of bioerosion in Mediterranean basin are even scantier (Calcinai et al., 2008, 2011; Färber et al., 2015, 2016) and exclusively about sponges and polychaetes.
Several organisms belonging to different taxa are involved in calcareous-boring activities; they can be microborers as cyanobacteria (Tribollet et al., 2011; Pica et al., 2016) and fungi (Gleason et al., 2017), or macroborers as sponges (Bavestrello et al., 1991; Calcinai et al., 2008, 2011), molluscs (Ćurin et al., 2014; Peharda et al., 2015), echinoderms (Belaústegui et al., 2017), and polychaetes (Martin & Britayev, 1998; Hutchings, 2008). Experimental studies for the evaluation of bioerosion rates are principally conducted using experimental blocks made of dead coral substrate (Tribollet et al., 2002; Hutchings, 2008), limestone panels (Wisshak et al., 2005; Casoli et al., 2019), shells (Calcinai et al., 2007), and wood (Reish et al., 2018). These experimental blocks also provide essential information about successional stages of the boring communities. In the first place, the local pattern of currents influences the larval dispersion (Tribollet et al., 2002) affecting the recruitment of boring organisms. The following successional stages, mainly facilitated by the borers, lead to the achievement of mature boring communities (Hutchings, 2008). Polychaetes have been reported as the pioneer-dominant borers on newly available submerged substrates and considered among major bioeroders along with sponges and molluscs (Hutchings et al., 1992, 2011). Nevertheless, studies about values of bioerosion of polychaetes are not abundant. Numerous researches assessed this topic in coral reefs ecosystems, obtaining a great variation among their estimates: Tribollet and Golubic (2005) (0.010–0.130 kg CaCO3·m−2·year−1) together with Yeung et al. (2021) (0.013–0.162 kg CaCO3·m−2·year−1) presented the lowest values, followed by Kiene and Hutchings (1994) (0.104 kg·m−2 ·year−1), Hutchings (2008) (0.356 kg·m−2·year−1), Chazottes et al. (2002) (0.050–0.400 kg CaCO3 m−2 ·year−1), and Davies and Hutchings (1983) which presented the highest value in the literature (1.800 kg·m−2·year−1). Generally, this group present lower values than sponges, considered as the principal erosive organisms in later succession stages (Pari et al., 2002). Only recently the colonization dynamics of boring polychaetes has been studied in the Mediterranean Sea (Tyrrhenian Sea), using experimental substrates (Casoli et al., 2019; Gravina et al., 2019), but still no data are available regarding their erosion rates.
Several families of polychaetes are involved in bioerosion processes, such as Eunicidae, Cirratulidae, Sabellidae, and Spionidae. These worms exhibit a variety of feeding preferences but are in general considered as filter and surface deposit feeders. Boring polychaetes use both chemical secretions and mechanical excavation for dissolving the rocks and making their holes (Hutchings, 2008); some sabellids, for example, although limited to a few species, have developed glandular areas along the ventral part of the body, while in some polydorids (Spionidae) the erosion of the carbonate substrate is possible thanks to the modified chaetae present on the 5th chaetiger, in addition to the production of chemicals compounds (Zottoli & Carricket, 1974; Sato-Okoshi & Okoshi, 1993; Hutchings, 2011). Polydorid is one of the most studied boring taxa and this is principally due to the serious damages these polychaetes inflict on oysters and abalone shells in aquaculture systems (Sato-Okoshi et al., 2008; Walker, 2011). These polychaetes are easily recognizable on the substrate for their typical 8-shaped holes, due to the two closed openings they produce divided by a thin mucous layer, allowing palps to move in the surrounding ground looking for food particles. The longitudinal section of the hole, where the worm lives in, is U-shaped, and each 8-shaped hole corresponds to a single boring individual. Not all the genera belonging to this taxon have been recognized as boring (Sato-Okoshi, 1999); sometimes, it’s not even easy to attribute the species to the correct taxon because of the similar morphological characteristics shared by a group of species, thus making identification a difficult task (see for example Simon & Sato-Okoshi, 2015; Bertasi, 2016; Çinar & Dagli, 2021), such as the case of the species complex Polydora ciliata (Johnston, 1838) (Mustaquim, 1986, 1988; Manchenko & Radashevsky, 1998).
Micro-computed tomography (micro-CT) has been extensively applied for studying internal and external anatomical features of several taxa such as corals (Morales Pinzón et al., 2014; Enochs et al., 2016; Urushihara et al., 2016), sponges (Heim & Nickel, 2010), or polychaetes (Faulwetter et al., 2013; Parapar et al., 2017, 2019, 2021), and in terms of analysis of bioerosion traces produced by micro- and macroborers (Beuck et al., 2007). Regarding annelids, micro-CT has been implemented to visualize, from a three-dimensional perspective, bioturbation in polychaetes (Pennafirme et al., 2019) and the boring pattern produced by the Siboglinid Osedax (Higgs et al., 2010). To the best of our knowledge, this is the first time micro-CT is used to visualize the development of boring polychaete cavities.
Considering the role that bioerosion plays in the marine habitat and the scanty knowledge of this phenomenon in the Mediterranean Sea, this study aims to assess on experimental limestone panels: (i) the pioneer, bioerosive organisms occurring in the first stages of bioerosion along the North Adriatic Sea and (ii) the annual rate of bioerosion during this first colonization phase. A diagram summarizing the different techniques implemented along the study was produced and added as Supplementary Material (Online Resource 1).
Methods
Study site, experimental design, and sample treatment
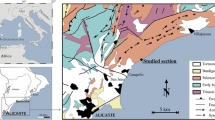
Marina Dorica (43°36.65ʹ N; 13°28.91ʹ E) is a touristic harbor in the northern-west coast of the Ancona Promontory (Adriatic Sea, Italy) (Fig. 1). In this site, five sets of three experimental blocks (15 in total) were placed under a wharf (W3) at a depth of 1 m. These blocks were made of Pietra Leccese (Margiotta, 2006), a common calcareous formation along Italian coasts consisting in a compact biomicrite made of planktonic foraminifera with its origin in the Miocene (Bossio et al., 2002).

Map of the study site, located in the touristic harbor of Marina Dorica (Ancona, Italy)
Blocks were positioned between the poles using a line and kept separated by plastic tubes 20 cm long. The wharf is in proximity of the canal port where the water exchange, with the open sea, is higher with respect to the inner port. The calcareous blocks were submerged from July 2017 to July 2019, and the first set of three blocks was collected in January’18 after 6 months from the positioning. Later, the other sets were collected bimonthly, up to July’18; the last three blocks were collected after 24 months of immersion (in total), in July 2019. When removed from the structure, the blocks were separately placed in plastic bags with sea water and transferred to the laboratory; at this point, all the encrusting organisms were carefully removed from the blocks and the blocks were kept in aquaria, under a continuous sea-water circulation. The cleaning procedure did not affect the integrity of boring worms and keeping them alive in aquaria allowed to precisely localize their holes. Magnesium chloride (7%) was added to the water to relax polychaetes and to facilitate their extraction from the substrate; right after the worms were extracted, forcing them out of their holes with the help of a Pasteur pipette, fixed in alcohol solution (95%) and identified. For scanning electron microscopy (SEM) analyses, several specimens were primary fixed in ethanol 20%, then dehydrated through a graded ethanol series (50%, 70%, 90%, 100%), and dried with Hexamethyldisilazane 98% until complete evaporation following Shively & Miller (2009). Finally, they were attached on stubs, sputtered with gold-platinum and observed under SEM Philips XL20.
Density and erosion parameters assessment
For each block, the 8-shaped Polydora’s holes were counted, calculating their average density (holes·100 cm−2). The two openings, creating an 8-shaped sign on the rock surface, approximately have equal diameter (d) (Fig. 2a); for this reason, only one diameter (of the single opening) per each hole was measured under stereomicroscope; the surface of each opening (OS) (Fig. 2a) was considered circular in shape and calculated with the formula OS = (d/2)2π. The value of the surface erosion produced by a single worm (8-shaped hole) was obtained doubling the OS values.

Three methodologies used to extract polydorid’s morphometrics. a Model used as a proxy for cavity volume; b Resin cast; c 3D reconstruction of Polydora’s cavities after micro-CT processing. (d diameter, h height, OS surface of a single opening)
To assess the volume of carbonate material removed by the polychaetes, a polyurethane resin (PU800) (www.antichitabelsito.it) was injected into the holes of the blocks, with a surgical syringe. Then, the blocks were dissolved in acid chloride (37%) and the hardened resin casts were removed. For this purpose, the blocks of March, May, and July 2018 were used. The resin casts reproduced worm excavations (Fig. 2b), whose regular form allowed us to approximate its shape to two paired cylinders (Fig. 2a) to calculate the total volume (TV) excavated by a single worm; we applied the formula TV = hm2OS, where 2OS is the average value of the excavated area (i.e., the double openings; see above) calculated for each period of sampling, and h stands for the height of the cast; the value hm was obtained by calculating the average height of 15 casts for each period (Fig. 2a, b). The total volume excavated in July 2018 was then used to estimate the erosion rate of polychaetes as mm3 of excavated substrate per mm−2 of substrate, per year−1. To evaluate the weight (g) of the substrate eroded by polychaetes per surface and year−1, the volumes removed by these after one year of exposure (July 2018), were multiplied by the specific gravity of the “Leccese” rock (2.577 g·cm-3) and by the specific gravity of the micritic limestone of the maiolica formation that mainly characterizes the Conero substratum (2.5 g·cm−3) (Calcinai et al., 2011).
In order to compare the density of excavating worms (= holes) on the blocks between the different times of exposure, it was standardized by dividing its density values by their time of exposure.
Parallel to the approach made with resin casts, Polydora samples from January and July 2018 were micro-CT-scanned at Centro di Ricerca e Servizio di Microscopia delle Nanostrutture (CISMiN) at the Polytechnic University of Marche, using a Skyscan 1174. During scanning, the sample was rotated in 0.20° angular increments reaching a total of 180° rotation, acquiring an X-ray absorption radiograph was digitally recorded at each rotation step. The X-ray source was set at 50 kV and 800 µA and an aluminum filter was placed between the sample and the camera in order to maximize the energy transmission through the sample. Scans were performed at a maximum pixel size of 25.25 µm. Initial processing of the projection images was performed with cone-beam reconstruction software (Nrecon v.1.7.3.1., Skyscan), resulting in a series of axial cross-sections. Post-processing of the cross-sections was conducted using the Amira software edition from the Zuse Institute Berlin, ZIB Amira 5 version 2019.1 (Stalling et al., 2005), where, in the first place, a Gaussian smoothing filter using a 3 × 3 × 3 voxel kernel was applied to reduce image noise. Then, bioerosive pores were differentiated from the rocky substrate by the following: (i) cross-sections segmentation by an interactive thresholding, defining the solid substrate; (ii) then, ambient occlusion module was applied to the binarized cross-sections in order to allow erosion pores to be visualized separately, as digital ‘casts’ in a greyscale depending on the occlusion of the same; and (iii) a final interactive thresholding was applied again to allow the software to perform volume calculations. Once single cavities were individualized on the 3D reconstructions, dimensions such as diameter, length, and volume values were extracted for each reconstructed erosion channel (Fig. 2c). Two sampling periods were assessed by the tomographic approach in order to test the technique, January and July 2018.
Statistical analysis
Given the proximity among the three blocks extracted at each time period, it was not possible to consider them as independent replicates. This is the reason why a repeated-measures ANOVA test was conducted to compare groups of samples exposed to different periods of immersion. The data were previously checked for homogeneity of variance using the Levene’s test, and whenever data did not meet the assumptions for parametric analyses, they were logarithmically transformed. Moreover, when the assumptions for parametric analyses were not met, even after logarithmical transformation, data were compared by the equivalent non-parametric analysis, the Friedman test.
All statistical analyses were performed using PAST 4 (Hammer et al., 2001) and graphical outputs with R software (R Core Team, 2020).
Results
Boring organism, density, and erosion traces
Boring species recorded into the experimental blocks belonged to two species: Polydora ciliata complex (Annelida: Spionidae) (Walker, 2011) and Polydora sp.. The P. ciliata complex specimens had the typical characters of the species (Fig. 3), but since the first description of the species refers to tubicolous non-boring forms on mud sediments (Johnston, 1838), for the boring sibling species of this complex, only genetic analyses can distinguish among different taxa (Manchenko & Radashevsky, 1993, 1998). Polydora sp. is a form showing some of the characters of the P. ciliata complex, but differs in having (i) dark bar pigmentation of the palps, (ii) the pigmentation present on the prostomium and dorsal side of the 3rd–4th segments, and (iii) the stout special chaetae of the 5th segment without a spur or secondary tooth. In fact, the pigmentation on both palps and prostomium, as well as in the anterior segments, closely resembles that of Polydora haswelli Blake & Kudenov, (1978) (see figure in Sato-Okoshi & Abe, 2013); however, the shape of the stout modified chaetae of the 5th segment differs from P. haswelli, in lacking any clear lateral accessory flange (Radashevsky et al., 2006). The form present in our samples resembles the worms collected by Boscolo and Giovanardi (2002) in the external coloration; these worms were found as borers infesting the bivalve Ruditapes philippinarum (A. Adams & Reeve, 1850) (cited as Tapes philippinarum) shells in the northern Adriatic and attributed by the Authors to P. ciliata. Therefore, considering that P. haswelli is actually not reported for the Mediterranean Sea, our specimens deserve further taxonomic analyses, possibly matched with molecular ones, since the 18S gene sequencing is already available for some Japanese specimens of P. haswelli (Sato-Okoshi & Abe, 2013).

SEM pictures of a specimen of Polydora ciliata complex, showing the main taxonomic characters. a–c Dorsal and lateral views of the prostomium and first few segments showing lack of notochaetae at the 1st segment, and the start of neuropodial uncini and branchiae at the 7th segment. d Modified stout chaetae of the 5th segments with a typical spur (sometimes a small tooth). e–f posterior part of the body showing the lack of both special chaetae and branchiae in the last segments, and the plate-like pygidium
However, from an ecological and functional point of view, both species found in our samples seem to exert the same role as boring pattern and activity, and therefore, their effects can be considered as they were a single taxonomic entity. Bearing this in mind, signs of bioerosion made by Polydora spp. have been present since the first sampling in January 2018, where after six months of immersion, we could already count a total of 80 specimens in all three blocks. Moreover, their density changed along the periods of immersion, reaching the highest value in July 2018, after 1 year of submersion (n = 290 in three blocks with a density of 32.8 ± 15.8 holes·cm−2).
Considering exposure time, the density of excavating worms (= holes) was comprised between 1.51 ± 0.51 worms 100 cm−2·month−1 in January 2018 (after 6 months of exposure) and 2.74 ± 1.32 worms 100 cm−2·month−1 in July 2018, after 12 months (Fig. 4). Repeated-measures ANOVA test revealed significant differences in relation to the periods of immersion (P < 0.05), specially between July 2018 and July 2019 (Tukey’s test P = 0.01296).

Density rate of the excavating worms (holes) (average ± SD) along the period of exposure
The average size of diameters of a single opening is shown in Fig. 5. The smallest value of 0.70 ± 0.19 mm was recorded in May’18 (after 10 months of immersion) while the higher value was 0.81 ± 0.13 mm in July’19, after 24 months of immersion. The Friedman test showed no significant differences (P > 0.05) between diameters among periods of immersion.

Average diameter (± SD) of Polydora’s hole (single opening) along the period of exposure
The heights of the resin casts were evaluated for holes produced in March, May, and July 2018. They are between 18.4 ± 4.5 mm in May and 19.2 ± 4.1 mm in March. Repeated-measures ANOVA test revealed no significant differences (P > 0.05) in the heights of the holes, reproduced by the casts.
Erosion rates assessment
The eroded volumes were ranged from 6.4 mm3·cm−2 of substrate (March’18, after 6 months of exposure) to 21 mm3·cm−2 of substrate (July’18, after a year). Assuming that substrate erosion is constant over time and considering the periods of blocks’ immersion, the eroded volume rates varied between 9.788 mm3·cm−2·year−1 (March ‘18) and 21.086 mm3·cm−2·year−1 (July’18). The rates obtained using the full-year period eroded volume was considered as a more complete estimate, possibly accounting for seasonal variations. In this context, erosion rate was calculated by multiplying the yearly eroded volume by the specific gravity of the Leccese rock, obtaining a rate of 0.0543 g·cm−2·year−1. If we consider the specific gravity of the Conero rock present along Ancona Promontory, the erosion rates on the Conero area would be around 0.053 g·cm−2·year−1.
Micro-CT characterization
Micro-CT was applied for two of the experimental time steps: January ‘18, where two subsamples were scanned resolving three fully reconstructed holes, presenting an averaged diameter of 0.71 ± 0.09 mm and a length of 8.32 ± 3.08 mm; while for July ‘18, a single subsample resolved three holes, with an averaged diameter of 0.78 ± 0.14 mm and an average length of 9.39 ± 3.29 mm. The 3D reconstructions obtained from the scanning allowed also to calculate the volume for each of the holes, obtaining averaged eroded volumes of 7.52 ± 4.73 mm3 for January ‘18 and 8.51 ± 5.52 mm3 for July ‘18.
Discussion
The aim of this study was to describe the first phases of bioerosion processes along the North Adriatic Sea coast and provide the first estimation of the relative bioerosion rates. The North-Italian Adriatic coast is characterized almost entirely by sandy beaches while the Conero Promontory, where Ancona is located, is a massive calcareous promontory colonized by both epilithic and endolithic organisms as sponges and bivalves (Calcinai et al., 2009). Up until today, only sponge bioerosion has been studied on this peculiar ecosystem (Calcinai et al., 2011). At Mediterranean level, knowledge regarding polychaete erosion rates is still absent. For some borers, as sponges (Cliona adriatica Calcinai et al., 2011) and bivalves (Rocellaria dubia (Pennant, 1777)), it has been possible to estimate the erosion with non-destructive methods, allowing to calculate the volume of the internal cavities by mean of its external openings (Schiaparelli et al., 2005; Calcinai et al., 2011). For polydorids, only destructive methods, such as cracking the substrates, have been used (Simon & Sato-Okoshi, 2015; Radashevsky et al., 2017); considering that heights of the excavations did not vary significantly with the time of exposure, we suggest to estimate the volume eroded by Polydora spp. directly by measuring the diameter of the openings, considering the erosion cavity as a double cylinder, and avoiding destructive approach; moreover, we also propose, for the first time, micro-CT three-dimensional reconstructions as a non-destructive method for evaluating polydorids’ erosion cavities. The erosion pattern in Polydora seems different in respect of other taxa as the bivalve R. dubia (see Schiaparelli et al., 2005), the date mussel (Lithophaga lithophaga, (Linnaeus, 1758)), or the piddock, (Pholas dactylus Linnaeus, 1758) where both the erosion cavities and the superficial openings enlarge following the mollusks’ growth.
This study shows that, after 6 months, the polydorid Polydora spp. appeared into the blocks, and no other excavating organisms were detected during the following 2 years of experiment. No evidence was recorded for boring sponges, bivalves, or echinoderms, widely considered as the principal taxa involved in boring processes both in tropical and temperate seas (Schönberg & Wisshak, 2014). These organisms are known to appear in late successional stages of the boring community (after 2 years, e.g., Chazottes et al., 1995), after worms that, in the coral reefs, are always reported as pioneers (Hutchings & Peyrot–Clausade, 2002; Hutchings, 2008; Hutchings et al., 2013). In this study, blocks were immersed only for 2 years but, although in the short term, this experiment highlights that also in the Mediterranean Sea, the first succession phase in the boring community mirrors that of the tropical areas (see, e.g., Hutchings et al., 1992; Tribollet et al., 2002).
The density of Polydora spp. significantly changed with the time of exposure, as reported for other bioeroders in studies conducted in the tropical areas (Chazotte et al., 1995; Hutchings et al., 2002; Hutchings et al., 2013). Recently, Casoli et al. (2019) registered the presence of only 10 specimens of Polydora ciliata and of another excavating worm (Dodecaceria concharum Örsted, 1843), in experimental blocks in the Tyrrhenian Sea. In contrast, our experimental blocks presented a high abundance of Polydora spp. specimens, reaching its maximum densities after one year. This sequence coincides with the results of Casoli et al. (2019) and Gravina et al. (2019) that found Polydora exclusively present in the first period, in blocks immersed for 1 year, substituted later by D. concharum, the only worm present in 2- and 3-year-old blocks. Consequently, species of the genus Polydora may be considered as pioneer species characterizing the first period of succession also in the Adriatic Sea.
In terms of eroded volumes, the higher values recorded in July 2018 are probably due to the settlement of a new cohort of Polydora spp., that, as reported by Blake (1969), it is characterized by one or two reproductive events in a single season, usually in late spring. In fact, during the observation in vivo, done on the samples of May 2018, several mature specimens were recorded.
Regarding the density, per unit of volume, of the boring polychaetes, our data varied between 10 ± 4.5 and 36.25 ± 23.3 worms·100 cm−3. Values comparable to those reported in coral reefs by Hutchings and Peyrot-Clausade (2002) which assessed abundances of some coral-boring polydorids in the French Polynesia, obtaining densities from 0.5 ± 0.8 to 36.3 ± 33.7 ind·100 cm−3; higher values as those reported by Hutchings et al. (2013) for Polydora spp. (303.0 ± 72.75 ind·100 cm−3). On the other hand, bioerosion rates, here assessed, are much lower than those reported in tropical areas (see data in the introduction). This is not surprising, as it is known that the bioerosion rate in the tropics exceeds the ones occurring in temperate and cold waters (Wisshak, 2006). Nevertheless, the comparison between present data and those from literature allows only some very general considerations; in literature, the erosion rates are often calculated without discriminating among the taxa, i.e., grouping polychaetes and sipunculans as “worms,” and being this group mainly clustered together with bivalves, as macroborers. Moreover, differences observed in terms of density and erosion rates may be due to the different lithologies used during the experiments; in fact, it has been demonstrated that the bioerosion rate is higher in porous substrates (corals) in respect of denser material (Leccese rock) (Calcinai et al., 2007).
Thanks to the micro-CT analysis, a clear three-dimensional view of the erosion patterns produced by Polydora spp. was obtained, showing a unified “8-shaped hole” which occasionally gets separated into two single funnels along its development. Measurements taken from the digital casts obtained similar values that the ones extracted from resin casts; yet the tomography did not only enabled us to extract measures, but also to observe the position and orientation of the cavities into the substrate, as well as to explore bioerosion pattern as a whole, otherwise impossible to obtain by more invasive approaches (e.g., Online Resource 2, where two worms appeared to have avoided perforating each other cavities by slightly changing direction during their erosion activity). Nevertheless, even if micro-CT has proved its value on the assessment of erosive patterns, its application on high-density substrates can cause artifacts during the scanning, thus limiting the maximum volume of sample to be analyzed. A posterior subsampling would make the technique to lose its non-invasive character, risking to damage the cavities inside the sample (as observed on Online Resource 2, where the cut on the substrate damaged two cavities). Moreover, operational costs for a high number of samples can be considerable, for which the coupling of more cost-effective techniques, such as resin casts, should be considered.
Conclusions
This is the first study evaluating bioerosion rates produced by polychaetes, organisms which have a dominant role in the bioerosion processes affecting calcareous substrates, both natural and artificial, in the Mediterranean Sea. The study has shown how the spionid polychaetes Polydora spp. trigger the earlier stages of bioerosion in the North Adriatic Sea, and that it is possible to estimate the etched volume with non-destructive methods. In the sea, the bioerosion affects any kind of calcareous substrate, playing a relevant role in several marine ecosystem dynamics. Bio-erosive successions in temperate climates are poorly studied. Moreover, in the near future, due to the increasing of organic pollution in coastal waters together with a general warming, an increment of the bioeroders’ activity is expected. Long-term experiments using substrates having different lithologies, at different depths and considering waters with different contents of organic matter, are needed to provide a wider and more precise picture of the bioerosion trends of the Mediterranean Sea.
Data availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
Change history
21 July 2022
Missing Open Access funding information has been added in the Funding Note.
References
Bavestrello, G., B. Calcinai & M. Sarà, 1991. Delectona ciconiae sp. nov. (Porifera, Demospongiae) boring in the scleraxis of Corallium rubrum. Journal of the Marine Biological Association of the United Kingdom 76: 867–873.
Belaústegui, Z., F. Muñiz, J. H. Nebelsick, R. Domènech & J. Martinell, 2017. Echinoderm ichnology: bioturbation, bioerosion and related processes. Journal of Paleontology 91: 643–661.
Bertasi, F., 2016. The occurrence of the alien species Polydora cornuta Bosc, 1802 (Polychaeta: Spionidae) in North Adriatic lagoons: an overlooked presence. Italian Journal of Zoology 83(1): 77–88.
Beuck, L., A. Vertino, E. Stepina, M. Karolczak & O. Pfannkuche, 2007. Skeletal response of Lophelia pertusa (Scleractinia) to bioeroding sponge infestation visualised with micro-computed tomography. Facies 53: 157–176.
Blake, J. A., 1969. Reproduction and larval development of Polydora from Northern New England (Polychaeta: Spionidae). Ophelia 7: 1–63.
Blake, J. A. & J. D. Kudenov, 1978. The Spionidae (Polychaeta) from southeastern Australia and adjacent areas with a revision of the genera. Memoirs of the National Museum of Victoria 39: 171–280.
Boscolo, R. & O. Giovanardi, 2002. Polydora ciliata shell infestation in Tapes philippinarum Manila clam held out of the substrate in the Adriatic Sea, Italy. Journal of Invertebrate Pathology 79: 197–198.
Bossio, A., R. Mazzei, B. Monteforti & G. Salvatorini, 2002. Note illustrative della carta geologica dell’area di Lèuca (con Appendice biostratigrafica a cura di Foresi L.M., Mazzei R., Salvatorini G.). Atti Della Società Toscana Di Scienze Naturali, Memorie Serie A 107: 97–163.
Calcinai, B., F. Azzini, G. Bavestrello, L. Gaggero & C. Cerrano, 2007. Excavating rates and boring pattern of Cliona albimarginata (Porifera: Clionaidae) in different substrata. In Beresi, M. S. (ed), Porifera Research: Biodiversity, Innovation & Sustainability Museu Nacional, Rio de Janeiro: 203–210.
Calcinai, B., C. Cerrano, N. Iwasaki & G. Bavestrello, 2008. Sponges boring into precious corals: an overview with description of a new species of Alectona (Demospongiae, Alectonidae) and a worldwide identification key for the genus. Marine Ecology 29: 273–279.
Calcinai, C., G. Bavestrello, B. Betti, M. Bo, C. Cerrano, C. G. Di Camillo, M. Martinelli, S. Puce & S. Tazioli, 2009. Storie vitali peculiari del benthos di substrato duro nei SIC marini marchigiani. Biologia Marina Mediterranea 16: 42–45.
Calcinai, C., G. Bavestrello, G. Cuttone & C. Cerrano, 2011. Excavating sponges from the Adriatic Sea: description of Cliona adriatica sp. nov. (Demospongiae: Clionaidae) and estimation of its boring activity. Journal of the Marine Biological Association of the United Kingdom 91: 339–346.
Casoli, E., S. Ricci, F. Antonelli, C. S. Perasso, G. Ardizzone & M. F. Gravina, 2019. Colonization dynamic on experimental limestone substrata: the role of encrusting epilithics favouring boring polychaetes. Hydrobiologia 842: 101–112.
Cerrano, C., G. Bavestrello, C. N. Bianchi, B. Calcinai, R. Cattaneo-Vietti, C. Morri & M. Sarà, 2001a. The role of sponge bioerosion in Mediterranean coralligeneous accretion. In Di Castri, F. & H. A. Mooney (eds), Mediterranean Ecosystems Springer, New York: 235–240.
Cerrano, C., G. Bavestrello, B. Calcinai, R. Cattaneo-Vietti, M. Chiantore, M. Guidetti & A. Sarà, 2001b. Bioerosive processes in Antarctic Sea. Polar Biology 24: 790–792.
Chazottes, V., T. Le Campion-Alsumard & M. Peyrot-Claused, 1995. Bioerosion rates on coral reef: interaction between macroborers, microborers and grazer (Moorea, French Polynesia). Palaeogeography, Palaeoclimatology, Palaeoecology 113: 189–198.
Chazottes, V., T. Le Campion-Alsumard, M. Peyrot-Clausade & P. Cuet, 2002. The effects of eutrophication-related alterations to coral reef communities on agents and rates of bioerosion (Reunion Island, Indian Ocean). Coral Reefs 21: 375–390.
Chazottes, V., J. J. Reijmer & E. Cordier, 2008. Sediment characteristics in reef areas influenced by eutrophication-related alterations of benthic communities and bioerosion processes. Marine Geology 250: 114–127.
Çinar, M. E. & E. Dagli, 2021. Bioeroding (boring) polychaete species (Annelida: Polychaeta) from the Aegean Sea (eastern Mediterranean). Journal of the Marine Biological Association of the United Kingdom 101: 1–10.
Ćurin, M., M. Peharda, B. Calcinai & S. Golubić, 2014. Incidence of damaging endolith infestation of the edible mytilid bivalve Modiolus barbatus. Marine Biology Research 10: 179–189.
Davidson, T. M., A. H. Altieri, G. M. Ruiz & M. E. Torchin, 2018. Bioerosion in a changing world: a conceptual framework. Ecology Letters 21: 422–438.
Davies, P. J. & P. A. Hutchings, 1983. Initial colonization, erosion and accretion of coral substrate. Coral Reefs 2(1): 27–35.
Enochs, I. C., D. P. Manzello, H. H. Wirshing, R. Carlton & J. Serafy, 2016. Micro-CT analysis of the Caribbean octocoral Eunicea flexuosa subjected to elevated pCO2. ICES Journal of Marine Science 73: 910–991.
Färber, C., M. Wisshak, I. Pyko, N. Bellou & A. Freiwald, 2015. Effects of water depth, seasonal exposure, and substrate orientation on microbial bioerosion in the Ionian Sea (Eastern Mediterranean). PLoS ONE 10: e0126495.
Färber, G., J. Titschack, C. Schönberg, K. Ehrig, K. Boos, D. Baum, B. Illerhaus, U. Asgaard, R. G. Bromley, A. Freiwald & M. Wisshak, 2016. Long-term macrobioerosion in the Mediterranean Sea assessed by micro-computed tomography. Biogeosciences 13: 3461–3474.
Faulwetter, S., A. Vasileiadou, M. Kouratoras, T. Dailianis & C. Arvanitidis, 2013. Micro-computed tomography: introducing new dimensions to taxonomy. ZooKeys 263: 1–45.
Gleason, F. H., M. G. Gadd, J. I. Pitt & A. W. D. Larkum, 2017. The roles of endolithic fungi in bioerosion and disease in marine ecosystems I. General concepts. Mycology. 8: 205–215.
Gravina, M. F., F. Antonelli, C. Sacco-Perasso, A. Cesaretti, E. Casoli & S. Ricci, 2019. The role of polychaetes in bioerosion of submerged mosaic floors in the Underwater Archaeological Park of Baiae, Naples, Italy. Facies 65(2): 19.
Hammer, Ø., D. A. T. Harper & P. D. Ryan, 2001. Past: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 4–9.
Heim, I. & M. Nickel, 2010. Description and molecular phylogeny of Tethya leysae sp. nov. (Porifera, Demospongiae, Hadromerida) from the Canadian Northeast Pacific with remarks on the use of microtomography in sponge taxonomy. Zootaxa. 2422: 1–21.
Higgs, N. D., A. G. Glover, T. G. Dahlgren & C. T. S. Little, 2010. Using computed-tomography to document borings by Osedax mucofloris in whale bone. Cahiers de Biologie Marine 5: 401–405.
Hutchings, P. A., 2008. Role of polychaetes in bioerosion of coral substrate. In Tapanila, L. & M. Wisshak (eds), Current Developments in Bioerosion Springer, Dordrecht: 249–264.
Hutchings, P. A., 2011. Bioerosion. In Hopley, D. (ed), Encyclopedia of Modern Coral Reefs: Structure, Form and Process Springer, Dordrecht: 139–156.
Hutchings, P. A. & M. Peyrot-Clausade, 2002. The distribution and abundance of boring species of polychaetes and sipunculans in coral substrates in French Polynesia. Journal of Experimental Marine Biology & Ecology 269: 101–121.
Hutchings, P. A., W. E. Kiene, R. B. Cunningham & C. Donnelly, 1992. Experimental Investigation of bioerosion at Lizard Island, Great Barrier Reef. Part 1. Patterns in the distribution and extent of non-colonial, boring communities. Coral Reefs 11: 23–31.
Hutchings, P., M. Peyrot-Clausade & A. Stuken, 2013. Internal macrobioerosion on five species of Acropora following the 1998 bleaching event: Implications for the long-term impact of bleaching on the Great Barrier Reef. Pacific Conservation Biology 19: 409–417.
Kiene, W. E. & P. A. Hutchings, 1994. Bioerosion experiments at Lizard Island, Great Barrier Reef. Coral Reefs 13: 91–98.
Manchenko, G. P. & V. I. Radashevsky, 1993. Genetic differences between two sibling species of the Polydora ciliata complex (Polychaeta: Spionidae). Biochemical Systematics and Ecology 21: 543–548.
Manchenko, G. P. & V. I. Radashevsky, 1998. Genetic evidence for two sibling species within Polydora cf. ciliata (Polychaeta: Spionidae) from the Sea of Japan. Marine Biology 131: 489–495.
Margiotta, S., 2006. Bio-cronostratigrafia a foraminiferi planctonici dei sedimenti miocenici nell’area di Strudà (Lecce, Puglia). Geologica Romana 39: 1–14.
Martin, D. & T. A. Britayev, 1998. Symbiotic polychaetes: review of known species. Oceanography & Marine Biology and annual Review 36: 217–340.
Meyer, N., M. Wisshak & A. Freiwald, 2021. Bioerosion ichnodiversity in barnacles from the Ross Sea, Antarctica. Polar Biology 44(4): 667–682.
Morales Pinzón, A., M. Orkisz, C. M. Rodríguez Useche, J. S. Torres González, S. Teillaud, J. A. Sánchez & M. Hernández Hoyos, 2014. A semi-automatic method to extract canal pathways in 3D micro-CT images of octocorals. PLoS ONE. 9: e85557.
Mustaquim, J., 1986. Morphological variation in Polydora ciliata complex (Polychaeta: Annelida). Zoological Journal of the Linnean Society 86: 75–88.
Mustaquim, J., 1988. Electrophoretic variation of isozymes in Polydora ciliata complex (Polychaeta Spionidae) Comparative Biochemistry & Physiology 91: 197–205.
Naylor, L. A., M. A. Coombes & H. A. Viles, 2012. Reconceptualising the role of organisms in the erosion of rock coasts: a new model. Geomorphology 157: 17–30.
Neumann, A. C., 1966. Observations on coastal erosion in Bermuda and measurements of the boring rate of the sponge, Cliona lampa. Limnology & Oceanography 11: 92–108.
Parapar, J., M. Candás, X. Cunha-Veira & J. Moreira, 2017. Exploring annelid anatomy using micro-computed tomography: a taxonomic approach. Zoologischer Anzeiger 270: 19–42.
Parapar, J., C. Caramelo, M. Candás, X. Cunha-Veira & J. Moreira, 2019. An integrative approach to the anatomy of Syllis gracilis Grube, 1840 (Annelida) using micro-computed X-ray tomography. PeerJ 7: e7251.
Parapar, J., M. Al-Kandari, M. Cands & J. Moreira, 2021. A new species of Polyophthalmus (Annelida, Opheliidae) from the Arabian Gulf, with an insight on internal anatomy and diversity of the genus. Zootaxa 5052(4): 501–528.
Pari, N., M. Peyrot-Clausade & P. A. Hutchings, 2002. Bioerosion of experimental substrates on high islands and atoll lagoons (French Polynesia) during 5 years of exposure. Journal of Experimental Marine Biology & Ecology 276: 109–127.
Peharda, M., B. Calcinai, S. Puljas, S. Golubic, J. Arapov & J. Thèbault, 2015. Endoliths in Lithophaga lithophaga shells – variation in intensity of infection and species occurrence. Marine Environmental Research 108: 91–99.
Pennafirme, S., A. S. Machado, A. C. Machado, R. T. Lopes, I. C. B. Lima & M. A. C. Crapez, 2019. Monitoring bioturbation by a small marine polychaete using microcomputed tomography. Micron 121: 77–83.
Pica, D., A. Tribollet, S. Golubic, M. Bo, C. G. Di Camillo, B. Bavestrello & S. Puce, 2016. Microboring organisms in living stylasterid corals (Cnidaria, Hydrozoa). Marine Biology Research 12: 573–582.
R Core Team, 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Radashevsky, V. I., P. C. Lana & R. C. Nalesso, 2006. Morphology and biology of Polydora species (Polychaeta: Spionidae) boring into oyster shells in South America, with the description of a new species. Zootaxa 1353: 1–37.
Radashevsky, V. I., J.-W. Choi & M. C. Gambi, 2017. Morphology and biology of Polydora hoplura Claparède, 1868 (Annelida: Spionidae). Zootaxa 4282: 543–555.
Reish, D. J., T. V. Gerlinger & R. R. Ware, 2018. Comparison of the polychaetous annelids populations on suspended test panels in Los Angeles Harbor in 1950–1951 with the populations in 2013–2014. Bulletin, Southern California Academy of Sciences 117: 82–90.
Sato-Okoshi, W., 1999. Polydorid species (Polychaeta: Spionidae) in Japan, with descriptions of morphology, ecology and burrow structure. 1. Boring species. Journal of the Marine Biological Association of the United Kingdom 79: 831–848.
Sato-Okoshi, W. & K. Okoshi, 1993. Microstructure of scallop and oyster shells infested with boring Polydora. Nippon Suisan Gakkaishi 59: 1243–1247.
Sato-Okoshi, W. & H. Abe, 2013. Morphology and molecular analysis of the 18S rRNA gene of oyster shell borers, Polydora species (Polychaeta: Spionidae), from Japan and Australia. Journal of the Marine Biological Association of the United Kingdom 93: 1279–1286.
Sato-Okoshi, W., K. Okoshi & J. Shaw, 2008. Polydorid species (Polychaeta: Spionidae) in south-western Australian waters with special reference to Polydora uncinata and Boccardia knoxi. Journal of the Marine Biological Association of the United Kingdom 88: 491–501.
Schiaparelli, S., G. Franci, G. Albertelli & R. Cattaneo-Vietti, 2005. A non-destructive method to evaluate population structure and bioerosion activity of the boring bivalve Gastrochaena dubia. Journal of Coastal Research 21: 383–386.
Schönberg, C. H. L. & M. Wisshak, 2014. Marine bioerosion. In Goffredo, S. & Z. Dubinsky (eds), The Mediterranean Sea: its History and Present Challenges. Springer, Dordrecht.
Schönberg, C. H. L., J. K. H. Fang, M. Carreiro-Silva, A. Tribollet & M. Wisshak, 2017. Bioerosion: the other ocean acidification problem: Contribution to the Themed Issue: ‘Ocean Acidification.’ ICES Journal of Marine Science 74: 895–925.
Shively, S. & W. R. Miller, 2009. The use of HMDS (hexamethyldisilazane) to replace critical point drying (CPD) in the Preparation of Tardigrades for SEM (Scanning Electron Microscope) Imaging. Transactions of the Kansas Academy of Science 112: 198–200.
Simon, C. A. & W. Sato-Okoshi, 2015. Polydorid polychaetes on farmed molluscs: distribution, spread and factors contributing to their success. Aquaculture Environment Interactions 7: 147–166.
Stalling, D., M. Westerhof & H. C. Hege, 2005. Amira: a highly interactive system for visual data analysis. In Hansen, C. D. & C. R. Johnson (eds), The Visualization Handbook Elsevier, Amsterdam: 749–767.
Tribollet, A. & S. Golubic, 2005. Cross-shelf differences in the pattern and pace of bioerosion of experimental carbonate substrates exposed for 3 years on the northern Great Barrier Reef, Australia. Coral Reefs 24(3): 422–434.
Tribollet, A., G. Decherf, P. A. Hutchings & M. Peyrot-Clausade, 2002. Large-scale spatial variability in bioerosion of experimental coral substrates on the Great Barrier Reef (Australia): importance of microborers. Coral Reefs 21: 424–432.
Tribollet, A., S. Golubic, G. Radtke & J. Reitner, 2011. On microbiocorrosion. In Reitner, J., N.-V. Quéric & G. Arp (eds), Advances in Stromatolite Geobiology. Lecture Notes in Earth Sciences Springer, Berlin: 265–276.
Urushihara, Y., H. Hasegawa & N. Iwasaki, 2016. X-ray micro-CT observation of the apical skeleton of Japanese white coral Corallium konojoi. Journal of Experimental Marine Biology and Ecology 475: 124–128.
Walker, L. M., 2011. A review of the current status of the Polydora - complex (Polychaeta: Spionidae) in Australia and a checklist of recorded species. Zootaxa 2751: 40–62.
Wisshak, M., 2006. High-Latitude Bioerosion: The Kosterfjord Experiment - Lecture Notes in Earth Sciences, Springer, Berlin:
Wisshak, M., M. Gektidis, A. Freiwald & C. Lundälv, 2005. Bioerosion along a bathymetric gradient in a cold-temperate setting (Kosterfjord, SW Sweden): an experimental study. Facies 51: 93–117.
Yeung, Y. H., J. Y. Xie, Y. Zhao, H. Y. Yu, C. Chen, W. W. Lu & J. W. Qiu, 2021. Rapid external erosion of coral substrate in subtropical Hong Kong waters. Marine Pollution Bulletin 169: 112495.
Zottoli, K. A. & M. R. Carriker, 1974. Burrow morphology, tube formation, and microarchitecture of shell dissolution by the spionid polychaete Polydora websteri. Marine Biology 27: 307–316.
Acknowledgements
The authors would like to thank Dr. Fabrizio Torsani and Dr. Daniela Pica for assistance during scientific diving. Thanks are due to Dr. Gravina M.F. and Dr. Radashevsky V.I., for their valuable advice regarding polydorid species’ taxonomy, and to Dr. Maria Laura Gatto, for her assistance during the Micro-CT processing. The authors are also thankful to the Reviewers for their precious comments.
Funding
Open access funding provided by Marche Polytechnic University within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
TPM contributed to methodology, software, visualization, data curation, writing-reviewing, and editing. LP contributed to conceptualization, data curation, writing, methodology, and original draft preparation. CC contributed to conceptualization, supervision, writing-reviewing, and editing. BC contributed to conceptualization, project administration, supervision, methodology, original draft preparation, writing-reviewing, and editing. MCG contributed to taxonomic consideration and writing-reviewing. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Handling Editor: Iacopo Bertocci
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file1 (AVI 145621 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pulido Mantas, T., Pola, L., Cerrano, C. et al. Bioerosion features of boring polydorid polychaetes in the North Adriatic Sea. Hydrobiologia 849, 1969–1980 (2022). https://doi.org/10.1007/s10750-022-04839-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-022-04839-3