
Abstract
Clinical, but not experimental evidence has suggested that air pollution particulate matter (PM) aggravates myocardial infarction (MI). Here, we aimed to describe mechanisms and consequences of PM exposure in an experimental model of MI. C57BL/6J mice were challenged with a PM surrogate (Residual Oil Fly Ash, ROFA) by intranasal installation before MI was induced by permanent ligation of the left anterior descending coronary artery. Histological analysis of the myocardium 7 days after MI demonstrated an increase in infarct area and enhanced inflammatory cell recruitment in ROFA-exposed mice. Mechanistically, ROFA exposure increased the levels of the circulating pro-inflammatory cytokines TNF-α, IL-6, and MCP-1, activated myeloid and endothelial cells, and enhanced leukocyte recruitment to the peritoneal cavity and the vascular endothelium. Notably, these effects on endothelial cells and circulating leukocytes could be reversed by neutralizing anti-TNF-α treatment. We identified alveolar macrophages as the primary source of elevated cytokine production after PM exposure. Accordingly, in vivo depletion of alveolar macrophages by intranasal clodronate attenuated inflammation and cell recruitment to infarcted tissue of ROFA-exposed mice. Taken together, our data demonstrate that exposure to environmental PM induces the release of inflammatory cytokines from alveolar macrophages which directly worsens the course of MI in mice. These findings uncover a novel link between air pollution PM exposure and inflammatory pathways, highlighting the importance of environmental factors in cardiovascular disease.
Similar content being viewed by others

Introduction
Cardiovascular disease represents the leading cause of mortality worldwide [26]. Chronic and acute inflammation triggered by traditional and non-traditional risk factors has been identified as the driving force behind cardiovascular pathologies including atherosclerosis and myocardial infarction (MI) [14, 31, 49]. In fact, environmental factors, such as air pollution, strongly influence initiation and outcome of cardiovascular disease [32]. Accordingly, numerous epidemiological studies identified exposure to environmental particulate matter (PM)—one of the main components of what is widely referred to as ‘air pollution’—as a major cause of increased mortality from MI [6]. Particularly, an acute exposure to PM elevates the risk of MI within a few hours [39]. Following MI, increased risk of death, progression to heart failure, and subsequent MI have been associated with PM exposure in humans [52]. Moreover, daily changes in airborne PM levels correlate with increased hospitalization due to MI, heart failure, arrhythmias, and stroke [22]. In cumulative analyses, up to 80 % of the increase in PM-associated mortality was caused by cardiovascular complications, while respiratory complications accounted for only 20 % of total mortality, indicating that cardiovascular, but not respiratory disease is the main complication of air pollution [4]. In line with these findings, exposure to PM accelerated experimental atherosclerosis in hyperlipidemic rabbits [51] and worsened cardiac function in previous animal studies [28].
Based on the observation that PM inhalation increases the levels of circulating pro-inflammatory cytokines, inflammation may be the cause of amplified cardiovascular mortality in PM-exposed individuals [29, 40]. However, neither the underlying mechanisms of the proposed inflammatory response, nor the direct participation of PM in experimental MI has been tested so far. To interrogate a potential direct functional link between PM exposure and MI, we have hypothesized that PM—by its surrogate Residual Oil Fly Ash (ROFA)—directly modulates traits of experimental MI in mice.
Methods
An expanded Methods section is available in the online-only Data Supplement.
Particulate matter
ROFA particles were collected from Boston Edison Co., Mystic Power Plant, Mystic, CT, US, burning low-sulfur residual oil (No. 6 fuel oil) and were kindly provided by Dr. J. Godleski (Harvard School of Public Health, Harvard University, Boston, MA, US). ROFA samples were extensively characterized regarding their chemical composition and particle size. Vanadium, nickel, and iron are the predominant metals present as water-soluble sulfates in ROFA. Their particle mean aerodynamic diameter is 2.06 ± 1.57 μm [8, 38]. ROFA containing suspensions were freshly prepared by suspending particles in sterile saline solution at 5 mg/mL, followed by incubation in an ultrasonic water bath for 5 min before use.
Animal protocols
Male 8-week-old wild type or lymphocyte-free Rag1-deficient (Rag1−/−) mice on a C57BL/6J background (Jackson Laboratories) were exposed to a suspension of ROFA particles in saline (1 mg/kg body weight) or the same volume of sterile saline solution (control group) by intranasal (i.n.) instillation as previously described [42]. MI was induced by permanent ligation of the left anterior descending coronary artery (LAD) [20]. All experimental protocols were approved by the Animal Ethics Committee of the University of Freiburg, Germany. Every procedure was carried out in accordance with institutional guidelines.
Histology
Seven days after MI, hearts were excised, embedded in Tissue-Tek O.C.T. compound (Sakura Finetek), frozen, and sectioned into 6 μm slices. Infarct area was demarked by Masson’s Trichrome staining. The anti-CD11b antibody clone M1/70 (BD Biosciences) was used as a pan-myeloid cell marker in immunohistochemistry.
Flow cytometry
Cell suspensions were stained for flow cytometry as previously described [47] and acquired on a Canto II cytometer (BD Biosciences). Antibodies were used as indicated. For intracellular cytokine staining, leukocytes were fixed and permeabilized by the Cytofix/Cytoperm kit (BD Biosciences) and incubated with fluorochrome-conjugated anti-TNF-α, anti-IL-6, and anti-MCP-1 antibodies (eBioscience). Data were analyzed with FlowJo (TreeStar). To quantify leukocytes resident in the heart, infarcted myocardial tissue was excised, weighted, digested, and the obtained cell suspensions were analyzed as previously described [20]. Specifically, cardiac monocytes were identified as CD45+, Lin− (Lin = CD3, CD19, CD49b, Ly6G, NK1.1), CD11b+, F4/80low, MHCIIlow, CD11clow, CD68low, CD115+ cells, subdivided into Ly6Chigh and Ly6Clow subsets; neutrophils were identified as CD45+, Lin+, CD11b+, MHCIIlow, CD11clow, SSChigh, Ly6Cint cells; cardiac macrophages were identified as CD45+, Lin−, CD11b+, F4/80high (MHCII+, CD68high) cells. Alveolar macrophages were identified in bronchoalveolar lavage (BAL) samples as CD45+, CD11blow, Siglec-F+, CD11c+, CD64+, F4/80int cells (Supplemental Fig. 2).
Intravital microscopy
Three hours after the exposure to ROFA particles, mice were anesthetized and leukocytes were fluorescently labeled by a retro-orbital injection of rhodamine. A loop of ileum was exteriorized and intravital microscopy was performed on mesenteric veins as previously described [47]. Rolling leukocyte flux was defined as the number of leukocytes moving at a lower velocity than erythrocytes. Adherent leukocytes were defined as cells that remained stationary for at least 30 s. Injection of 200 ng murine TNF-α i.p. served as positive control.
Sterile peritonitis
One hour after ROFA exposure, C57BL/6J mice received an i.p. injection of 2 ml 4 % thioglycollate broth. After 4 h, a peritoneal lavage was performed and peritoneal exudate cells (PECs) were quantified in a hemocytometer after red cell lysis.
Cytokine levels
A cytometric bead array (CBA assay, BD Biosciences) was used to quantify cytokine levels in plasma, BAL, and cell culture supernatants according to manufacturer’s protocol.
Endothelium activation markers
Plasma levels of soluble ICAM-1 (sICAM-1) and VCAM-1 (sVCAM-1) were measured by ELISA (R&D Systems). Alternatively, ICAM-1 and VCAM-1 expression was quantified on in vitro cultured mouse endothelial cells by immunohistochemistry as previously described [43].
Conditioned plasma
Mouse endothelial cells from non-exposed C57BL/6J mice were isolated and cultured as previously described [47]. Neutrophils, monocytes, and macrophages were isolated by magnetic bead separation. Cells were incubated with plasma (1 % v/v) from saline- or ROFA-exposed mice, or with ROFA particles (1 μg/mL). As indicated, incubation was performed in the presence of a blocking anti-TNF-α antibody (10 μg/mL). After 24 h, cell culture supernatants were analyzed for cytokines, and cellular expression of adhesion molecules was analyzed by immunohistochemistry and flow cytometry as previously described [48].
Macrophage depletion
Alveolar macrophages were depleted from C57BL/6J mice before ROFA exposure by an i.n. instillation of 50 μl dichloromethylene bisphosphonate (clodronate) liposomes (5 mg/mL) as previously described [53]. PBS-loaded liposomes were used as control. Alternatively, 200 μl clodronate liposomes were injected i.v. for systemic depletion of macrophages.
Statistical analysis
Data are presented as mean ± SEM. Unpaired Student’s t test was used to analyze differences between two groups. ANOVA followed by Student–Newman–Keuls post hoc test was performed to evaluate differences between more than two groups. Statistical significance was considered at p < 0.05.
Results
Exposure to ROFA particles aggravates myocardial infarction
To test whether exposure to PM modulates MI, 8-week-old C57BL/6J mice were challenged with ROFA particles (1 mg/kg body weight) or an equal volume of sterile saline solution by i.n. instillation. After 1 day, MI was induced by permanent ligation of the left anterior descending coronary artery (LAD). Mice were subsequently instilled with either ROFA or saline solution on a daily basis. On day 7, hearts were excised and cross sections of the myocardium were stained with Masson’s Trichrome to depict infarction area (Fig. 1a). We detected a significant increase in infarction size by up to 67 ± 13 % (Fig. 1b) and decrease in collagen density (Fig. 1c) in ROFA-treated mice compared with respective control. These data indicate an adverse cardiac remodeling in ROFA-exposed mice. We also identified increased CD11b+ myeloid cell accumulation in hearts of ROFA-treated mice by immunohistochemistry (Fig. 1d). We observed no differences in overall survival (data not shown) or MI-associated arrhythmias in ROFA-exposed mice (Supplemental Fig. 1), but a slight increase in the duration of ventricular complexes 3 days after MI in this group (Supplemental Table 1). The latter is consistent with previous observations that ROFA exposure enhances the vulnerability to cardiac arrhythmias [46]. To further characterize the dynamics of myeloid cell infiltration to cardiac tissue, leukocyte populations in the heart were quantified by flow cytometry on days 0, 3, and 7 after LAD ligation (Fig. 1e). Following MI, ROFA-exposed mice showed enhanced accumulation of neutrophils, inflammatory Ly6Chigh monocytes, and macrophages (Fig. 1f). Particularly, macrophage numbers increased by 125 ± 31 % in ROFA-exposed mice at day 7 (Fig. 1f). These results demonstrate that exposure to ROFA particles increases MI size and myocardial inflammatory cell accumulation, in particular that of macrophages.

Exposure to ROFA particles aggravates MI. C57BL/6J mice were exposed to ROFA particles (1 mg/kg body weight) or an equal volume of sterile saline solution by i.n. instillation. MI was induced by permanent ligation of the left anterior descending coronary artery (LAD). After 7 days, infarct area was demarked and quantified by Masson’s Trichrome staining in cross sections of heart tissue at increasing distance from the site of coronary artery ligation (a, b). At 2 mm from the site of LAD ligation, collagen density was evaluated within the infarcted tissue (c), and myeloid cell infiltration was assessed by immunohistochemistry for the pan-myeloid marker CD11b (d). Myeloid cell infiltration was characterized by flow cytometry of digested hearts (e). Leukocyte populations of infarcted tissue from saline- and ROFA-exposed mice were quantified at the indicated time points after LAD ligation (f). Data are presented as mean ± SEM of at least seven mice per group. *p < 0.05
ROFA particles promote trafficking of inflammatory leukocytes by enhancing leukocyte activation
To directly test whether an acute exposure to ROFA particles enhances inflammatory cell recruitment, a main pathomechanism in MI, we tested two in vivo models. First, we treated C57BL/6J mice by an i.n. instillation of ROFA particles (1 mg/kg body weight), saline solution, or an i.p. injection of murine TNF-α (200 ng), and monitored leukocyte recruitment in mesenteric venules by intravital microscopy (Fig. 2a). Interestingly, we observed a significant increase in the number of rolling and adhering leukocytes in ROFA-exposed mice (Fig. 2a, b, Supplemental Videos I–III). Accordingly, average rolling velocity was decreased after ROFA exposure (Fig. 2c). TNF-α i.p. treatment was used as a positive control and increased leukocyte rolling and adhesion, as well as decreased mean rolling velocity (Fig. 2c). Secondly, we tested whether ROFA exposure is capable of modulating leukocyte migration in vivo. Therefore, C57BL/6J mice were treated with ROFA as described above, and the number of cells accumulating in the peritoneal cavity was quantified. We did not observe relevant changes after ROFA exposure alone (Fig. 2d). However, after induction of sterile peritonitis by thioglycollate, simultaneous exposure to ROFA increased the number of peritoneal exudate cells (PECs) by 74 ± 12 % compared to saline (Fig. 2d), indicating that a ROFA exposure can boost, but not initiate leukocyte migration. To explore a potential mechanism, we assessed leukocyte activation by quantifying the abundance of the activation epitope CBRM1/5 on the leukocyte integrin CD11b/CD18 (Mac-1), an important mediator of leukocyte rolling and adhesion [10]. Interestingly, ROFA treatment significantly increased CD11b activation on circulating neutrophils and inflammatory Ly6Chigh monocytes (Fig. 2e). Moreover, we observed a significant increase in neutrophil- and Ly6Chigh monocyte-platelet aggregates in this group (Fig. 2f). Notably, plasma levels of the soluble fraction of the endothelial adhesion molecules ICAM-1 and VCAM-1 increased (Fig. 2g), suggesting that a ROFA exposure also induces endothelial cell activation. Taken together, these findings indicate that the exposure to ROFA particles induces inflammatory leukocyte recruitment, possibly by the up-regulation of adhesion molecules on endothelial and circulating myeloid cells.

A single instillation of ROFA particles promotes adhesion and migration of pro-inflammatory leukocytes. C57BL/6J mice were treated with either saline, a suspension of ROFA particles in saline (1 mg/kg body weight), or murine TNF-α (200 ng i.p.) as positive control. After 3 h, leukocyte recruitment in mesenteric venules was assessed in intravital microscopy. Leukocytes were stained by rhodamine (a) and leukocyte rolling, adhesion (b), and cumulative frequency of the rolling velocity (c) were quantified. The inlay in c represents leukocyte mean rolling velocity. To evaluate whether ROFA treatment primes leukocytes to migrate, the number of cells residing in the peritoneal cavity (PECs) was quantified in saline- or ROFA-exposed mice. Leukocyte migration was forced by inducing sterile peritonitis by an i.p. injection of 4 % thioglycollate broth (d). To assess leukocyte activation, expression of the CD11b activation epitope (CRBM1/5, e) and formation of leukocyte-platelet aggregates (f) were quantified on myeloid cells by flow cytometry. Plasma markers of endothelial activation (g) were quantified by ELISA. Data are presented as mean ± SEM of at least 10 mice per group
Activation of leukocytes and endothelial cells following ROFA exposure can be reversed by blockade of pro-inflammatory cytokines
It has been suggested that exposure to PM increases the levels of circulating pro-inflammatory cytokines, but it is not known whether these cytokines are required for inflammatory leukocyte recruitment after PM inhalation [5]. Therefore, we hypothesized that cytokines circulating in the blood activate leukocytes and endothelial cells in our model. We first measured TNF-α, IL-6, and MCP-1 in the plasma of exposed mice. Consistent with previous reports, we detected elevated cytokine levels, such as TNF-α (an increase up to 327 ± 100 %) and IL-6 (an increase up to 1686 ± 469 %) compared with samples from control mice (Fig. 3a). Next, we tested the impact of cell culture media supplemented with conditioned plasma from saline- or ROFA-exposed mice (1 % v/v) to ex vivo cultures of isolated myeloid and endothelial cells. Supplementation of plasma from mice exposed to ROFA increased activation of leukocytes and endothelial cells in vitro, as assessed by CD11b activation and expression of endothelial adhesion factors (Fig. 3b, c). Interestingly, a neutralizing anti-TNF-α antibody reversed these effects. Similar results were obtained with a blocking anti-IL-6 antibody (data not shown). These findings indicate that pro-inflammatory cytokines circulating in the plasma after ROFA exposure increase activation of leukocytes and endothelial cells.

Activation of leukocytes and endothelial cells by ROFA is reversed by blockade of the pro-inflammatory cytokine TNF-α. C57BL/6J mice were exposed to saline or a suspension containing ROFA particles (1 mg/kg body weight) by i.n. instillation, and plasma levels of pro-inflammatory cytokines were quantified by a cytometric bead array (a). Isolated myeloid and endothelial cells were incubated in vitro with conditioned plasma (1 % v/v) from saline- or ROFA-exposed mice in the presence or absence of a blocking anti-TNF-α antibody (10 μg/ml). After 24 h, the abundance of the CD11b activation epitope CBRM1/5 was quantified by flow cytometry in myeloid cells (b). On endothelial cells, expression of ICAM-1 and VCAM-1 was determined by flow cytometry (c). Data are presented as mean ± SEM from at least nine mice per group (a) and from at least three independent experiments (b, c)
ROFA particles do not directly activate endothelial or circulating myeloid cells
Nano-scale particles and soluble PM constituents can break through the respiratory epithelia and exert direct effects in systemic circulation within minutes to hours [33, 45]. To address whether ROFA particles circulating in the blood directly induce a cellular response, we challenged endothelial and circulating myeloid cells, such as neutrophils and monocytes, with a concentration of ROFA particles higher than expected after inhalation (1 μg/mL). Interestingly, exposure to ROFA particles had no effect on the expression of ICAM-1, VCAM-1, P-Selectin, or E-Selectin in cultured endothelial cells, as shown by immunohistochemistry and flow cytometry (Fig. 4a, b), while the addition of TNF-α strongly up-regulated these markers. Also, neutrophils and monocytes did not significantly increase TNF-α secretion after incubation with ROFA particles (Fig. 4c). These findings suggest that ROFA particles cannot directly activate endothelial or circulating myeloid cells.

Direct exposure to ROFA particles does not activate endothelial or circulating myeloid cells. Endothelial cells were incubated in vitro with saline, ROFA particles (1 μg/mL), or TNF-α (10 ng/mL) as positive control. Expression of the adhesion molecules ICAM-1, VCAM-1, P-Selectin, and E-Selectin were quantified after 24 h by immunohistochemistry (a) as well as by flow cytometry (b). Isolated myeloid cells were incubated in vitro with ROFA particles (1 μg/mL), and TNF-α levels were measured in cell culture supernatants (c). IgG Isotype in panel a indicates the staining with an unspecific FITC- or TRITC-coupled isotype antibody. Data are presented as mean ± SEM of at least three independent experiments per group. ns indicates not significant
Exposure to ROFA particles induces the release of pro-inflammatory cytokines by alveolar macrophages
PM inhalation can trigger the accumulation of macrophages in the lung and the release of inflammatory cytokines [3, 17]. Therefore, we hypothesized that circulating pro-inflammatory cytokines detectable in blood may originate from alveolar macrophages. To give proof to this concept, we isolated macrophages from a splenocyte cell suspension by magnetic bead separation and incubated the obtained cells in RPMI media supplemented with either saline or ROFA particles. Macrophages exposed to ROFA showed increased CD11b activation (Fig. 5a, b) and augmented secretion of pro-inflammatory cytokines (Fig. 5c). Next, we confirmed that ROFA exposure was associated with an increase in total leukocyte count in bronchoalveolar lavage (BAL) as previously reported [17] (Fig. 5d). ROFA instillation strongly increased pro-inflammatory cytokine levels in BAL, including TNF-α and IL-6 (Fig. 5e). To track down the cellular origin of these cytokines, we performed intracellular cytokine staining in distinct leukocyte subpopulations by flow cytometry, and identified alveolar macrophages (CD45+, CD11blow, Siglec-F+, CD11c+, CD64+, F4/80int, Supplemental Fig. 2) as the primary source of these cytokines (Fig. 5f, g). Other cells, such as T- or B-lymphocytes, did not show relevant cytokine production (data not shown). Notably, Toll-like receptor (TLR) 4 expression in alveolar macrophages, but not in circulating myeloid cells, was increased (Supplemental Fig. 3), suggesting a functional role of TLR pathways in macrophage activation following ROFA exposure. These results indicate that alveolar macrophages are the predominant source of pro-inflammatory cytokines in the lung after exposure to environmental PM.

Exposure to ROFA particles induces the release of pro-inflammatory cytokines by alveolar macrophages. Macrophages isolated from the spleen were cultured in the presence of ROFA particles (1 μg/mL) or medium alone (RPMI). Cell activation was assessed by expression of the CD11b activation epitope CBRM1/5 (a, b). The concentration of pro-inflammatory cytokines was determined in cell culture supernatants (c). To quantify cytokine secretion in vivo, C57BL/6J mice were exposed to saline solution or ROFA particles (1 mg/kg body weight) by i.n. instillation. A bronchoalveolar lavage (BAL) was performed after 3 h and total cell numbers (d) and the concentration of pro-inflammatory cytokines were quantified (e). The BAL cell suspension was analyzed by flow cytometry. Alveolar macrophages were identified as CD45+, CD11blow, Siglec-F+, CD11c+, CD64+, F4/80int cells (Supplemental Fig. 2), and intracellular cytokine staining was performed for TNF-α and IL-6 (f, g). Data are presented as mean ± SEM of at least three independent experiments (a–c) or at least six animals per group (d–g)
Specific depletion of alveolar macrophages prevents ROFA-mediated inflammation
We hypothesized that ROFA exposure induces an inflammatory response driven by alveolar macrophages thereby impairing MI healing. To proof this concept in a loss-of-function approach, alveolar macrophages were depleted in our model by clodronate liposomes. Clodronate is a strongly hydrophilic bisphosphonate that—when delivered in liposomes—induces apoptosis in macrophages after internalization [44]. In a first approach, we injected clodronate liposomes i.v. to systemically deplete macrophages. PBS-loaded liposomes were used as control. After 24 h, i.v. clodronate pretreatment abolished ROFA-induced macrophage recruitment to the lung and reduced local and systemic cytokine levels in this group (Supplemental Fig. 4), indicating that macrophages are a relevant source of circulating pro-inflammatory cytokines. To specifically deplete alveolar macrophages, we delivered clodronate liposomes by i.n. instillation. This strategy was highly effective in depleting macrophages from the lung, but not from other locations (Supplemental Fig. 5). Alveolar macrophages were suppressed up to 5 days after a single dose of clodronate (Supplemental Fig. 6). Specific depletion of alveolar macrophages was effective in preventing both local and systemic cytokine release following ROFA exposure (Supplemental Fig. 7). To rule out that some of the effects induced by ROFA are at least partially mediated by the adaptive immune system, such as by lymphocytes [2, 41], we exposed lymphocyte-free Rag-1−/− mice to ROFA particles. Notably, the cellular and inflammatory response resembled that of wild type mice (Supplemental Fig. 8), indicating that lymphocytes do not play a significant role in the acute inflammatory response initiated by ROFA exposure.
Finally, to confirm that alveolar macrophages ultimately determine the inflammatory response in infarcted cardiac tissue after ROFA exposure, we tested a combined model of alveolar macrophage depletion, ROFA exposure, and MI (Fig. 6a). Clodronate depletion effectively reduced alveolar macrophages 7 days after MI (Fig. 6b) and protected from ROFA-mediated increase in BAL and systemic TNF-α levels (Fig. 6c, d). In accord with the finding that blood Ly6Chigh monocyte activation correlated with plasma TNF-α levels in ROFA-exposed mice (Supplemental Fig. 9), we observed lowered expression of the CD11b activation epitope in circulating inflammatory Ly6Chigh monocytes in clodronate-pretreated ROFA-exposed mice (Fig. 6e). As a result, lack of alveolar macrophages prevented from macrophage accumulation in infarcted tissue after ROFA exposure (Fig. 6f, g). Moreover, depletion of alveolar macrophages was also effective in reducing the ROFA-mediated increase in leukocyte rolling and adhesion in intravital microscopy (Fig. 6h, i, Supplemental videos IV–VI), suggesting that enhanced inflammatory cell recruitment is the cause of increased macrophage accumulation in infarcted tissue of ROFA-exposed mice.

Specific depletion of alveolar macrophages prevents ROFA-mediated inflammation. To test their simultaneous effects, depletion of alveolar macrophages, ROFA exposure, and MI were performed gradually (a). After 7 days, numbers of alveolar macrophages in the BAL were determined by flow cytometry (b). TNF-α levels in BAL (c) and plasma (d) were quantified after 7 days, or at the indicated time points. Expression of the CD11b activation marker on inflammatory Ly6Chigh blood monocytes was expressed as MFI (e). Myeloid cell subsets in infarcted hearts, including macrophages, were quantified after 7 days (f, g). Rolling and adhering cells in mesenteric venules were quantified in intravital microscopy after depletion of alveolar macrophages and subsequent ROFA exposure (h, i). Data are presented as mean ± SEM of at least seven mice pre group
Discussion
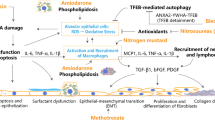
Exposure to environmental PM correlates with cardiovascular disease and mortality from MI [1, 3, 5]. However, it is not known whether and by which mechanism PM influence MI. In this study, we present the novel finding that the exposure to ROFA particles aggravates experimental MI by boosting inflammatory cell recruitment to the myocardium. Mechanistically, four major findings support our model (Fig. 7): (1) an acute ROFA exposure induced sustained activation of macrophages, but not of lymphocytes, resident in the lung. (2) Activated alveolar macrophages secreted pro-inflammatory cytokines, such as TNF-α and IL-6, into the circulation. (3) This increase in systemic cytokines promoted expression of myeloid and endothelial adhesion factors, and pre-disposed for enhanced leukocyte rolling, adhesion, and transmigration. Notably, anti-cytokine therapy reversed these effects. (4) Experimental depletion of alveolar macrophages reduced inflammatory cytokine release and protected from inflammatory cell recruitment induced by ROFA in vivo. Accordingly, attenuation of myeloid cell recruitment into the myocardium has been reported to improve MI healing [31].

Proposed model of PM-induced aggravation of myeloid cell recruitment during MI. Following PM inhalation, alveolar macrophages are directly activated by PM and orchestrate lung inflammation. The release of pro-inflammatory mediators, such as TNF-α and IL-6, into the blood stream results in increased expression of adhesion factors in myeloid cells and the endothelium. Consequently, leukocyte migration to sites of pre-established inflammation is boosted. Within the infarcted myocardium, monocytes and macrophages accumulate, ultimately leading to impaired tissue remodeling and increased infarction area
Numerous studies have previously suggested different mechanisms. Firstly, a direct uptake of PM by phagocytes, such as by macrophages, has been demonstrated [21]. Secondly, it is thought that certain PM constituents may induce an oxidative stress response [50]. Airborne PM is comprised of a heterogeneous mixture of solid and liquid particles suspended in air, varying in size, chemical composition and sources of origin [5]. Anthropogenic emissions are the main contributors to environmental PM burden and mainly consist of motor vehicle emissions and fossil fuel combustion during power generation and industrial processes [32]. ROFA particles often present an aerodynamic diameter below 2.5 μm (PM2.5), a size that penetrates the lung deeper and has been shown to be more closely associated with PM adverse health effects rather than coarser particles (PM10) [5]. ROFA particles are especially rich in soluble transition metals, that can enhance the inflammatory response triggered by PM, through generation of reactive oxygen species (ROS) [7]. In our study, we tested PM rich in water-soluble transition metals [8, 36, 38]. Some deleterious effects observed in local lung injury appear to be caused by reactive oxygen species which may potentially be overcome by over-expression of superoxide dismutase [17, 18, 27, 28]. An interesting link is provided by the observation that increased oxidative damage after PM exposure leads to the generation of oxidized lipid species, such as 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine (oxPAPC) in the bronchoalveolar lavage fluid. The later can be engulfed by macrophages mediated by pattern-recognition receptors, such as Toll-like receptors (TLRs), and potentiate inflammatory signaling [24]. In accord, TLR4-deficient mice are protected from PM-associated inflammation [24] and MI [37]. In our study, TLR4 expression was enhanced in alveolar macrophages after ROFA exposure, but not in circulating leukocytes, raising the possibility that TLR4 may indeed mediate some inflammatory pathways initiated by ROFA. However, we also observed that ROFA particles alone induced pro-inflammatory cytokine expression in isolated alveolar macrophages, indicating that such pathways are not required for, but rather synergistically support the inflammatory response after ROFA exposure.
The concept that PM exposure induces leukocyte recruitment by promoting both rolling and firm adhesion has been reported earlier. In particular, Nurkiewicz et al. have shown that rats exposed to ROFA exhibited increased leukocyte deposition in the spinotrapezius muscle [34]. This effect was discussed to be caused by deposition of myeloperoxidase in the arterial wall [35]. Moreover, we previously showed that ROFA instillation activates intravascular neutrophils [28], as also confirmed in this study. However, it has not been elucidated whether leukocytes are activated per se or whether this activation is secondary to circulating mediators of inflammation. By testing conditioned plasma from ROFA-exposed mice on in vitro cultured leukocytes and endothelial cells, we showed that cytokines are needed to induce cell activation, while ROFA particles alone are not sufficient to fully activate cell types participating in leukocyte recruitment. In vitro, circulating TNF-α levels in ROFA-exposed mice caused ICAM-1 and VCAM-1 expression in endothelial cells, as well as CD11b (Mac-1) integrin activation in myeloid cells, a prerequisite for leukocyte adhesion to ICAM-1 [9, 13]. This is particularly interesting since TNF-α is capable of inducing endothelial activation [23]. Given that nano-scale particles and soluble PM constituents have been found in systemic circulation after PM inhalation [33, 45], a slight effect of ROFA particles on endothelial and circulating leukocytes cannot be completely ruled out. As proof-of-concept, we have demonstrated that therapeutic inhibition of the prototypic pro-inflammatory cytokine TNF-α reverses the effects of PM. In accord, it has been recently shown that TNF-α blockade is also effective to prevent PM-associated worsening in cardiac energetic and contractile function [30]. Our findings do not exclude the participation of alternative pro-inflammatory mediators. In particular, Fijimaki et al. have demonstrated that IL-6-deficient mice failed to accumulate leukocytes in the lung after the exposure to diesel exhaust particles [12], proposing that IL-6 also participates in lung inflammation in this setting.
In our study, alveolar macrophages are the source of TNF-α and IL-6 after ROFA exposure. These data confirm previous studies showing that macrophage activation occurs in vitro and in vivo after exposure to PM [21]. It has already been demonstrated that cytokine levels detectable in BAL rise after PM inhalation [11, 17]. However, the cellular origin of these cytokines has not been tracked down until now. Another important finding from our study is that circulating TNF-α levels rise following PM exposure, even in the context of already elevated TNF-α plasma levels after MI. Notably, lymphocytes can promote acute and chronic inflammation by cytokine secretion and providing chemotactic stimuli for myeloid cells, particularly in the setting of MI [55]. To further elucidate the relative participation of lymphocytes and macrophages, Rag1−/− mice lacking mature lymphocytes and clodronate-pretreated mice lacking alveolar macrophages were used as experimental models. Interestingly, local and systemic inflammation in Rag1−/− mice resembled that of wild type mice, indicating that T- and B-lymphocytes do not play a relevant role in this context. On the contrary, we observed that some cytokine levels were even higher in Rag1−/− than in wild type mice, suggesting a protective role of some lymphocyte subsets, as previously suggested for T-regulatory cells in lung inflammation [54]. Specific depletion of alveolar macrophages reduced local and systemic inflammation after ROFA exposure, indicating that alveolar macrophages are the source of circulating pro-inflammatory cytokines in our model. Of note, we have not observed that i.n. clodronate affects myeloid cell survival at other locations than the lung. This is particularly important, since MI is driven by the mobilization of monocytes from secondary lymphoid organs and the bone marrow to the heart, where they rapidly differentiate into macrophages within a couple of days [31]. Accordingly, lack of alveolar macrophages prevented excessive inflammatory leukocyte accumulation in infarcted tissue of ROFA-exposed mice. In this context, it is also possible that ROFA exposure may trigger the release of monocytes from the bone marrow of lymphoid organs, although we have not observed elevated numbers of circulating myeloid cells following ROFA exposure. On the other hand, other effector functions of alveolar macrophages apart from cytokine release may also contribute to the observed effects. These questions will have to be answered in future studies.
Despite ROFA is an accepted model particle for air pollution PM [16], a potential limitation of our study is that an acute exposure to ROFA may only partially reflect environmental exposures as present in urban areas of industrialized countries, in particular when PM is present at lower concentrations. However, acute exposure to oil fly ashes [15, 19, 25] in humans has been reported to induce a comparable degree of lung injury and inflammation as observed in the present study.
In conclusion, the presented findings unravel some of the key mechanisms by which PM worsens the outcome following MI, directly linking air pollution PM with the activation of pathologic inflammatory pathways. After all, our data highlight the importance of environmental factors in cardiovascular disease.
References
Analitis A, Katsouyanni K, Dimakopoulou K, Samoli E, Nikoloulopoulos AK, Petasakis Y, Touloumi G, Schwartz J, Anderson HR, Cambra K, Forastiere F, Zmirou D, Vonk JM, Clancy L, Kriz B, Bobvos J, Pekkanen J (2006) Short-term effects of ambient particles on cardiovascular and respiratory mortality. Epidemiology 17:230–233. doi:10.1097/01.ede.0000199439.57655.6b
Arantes-Costa FM, Grund LZ, Martins MA, Lima C (2014) Airborne pollutant ROFA enhances the allergic airway inflammation through direct modulation of dendritic cells in an uptake-dependent mechanism. Int Immunopharmacol 22:9–20. doi:10.1016/j.intimp.2014.06.020
Brook RD (2008) Cardiovascular effects of air pollution. Clin Sci (Lond) 115:175–187. doi:10.1042/CS20070444
Brook RD, Franklin B, Cascio W, Hong Y, Howard G, Lipsett M, Luepker R, Mittleman M, Samet J, Smith SC Jr, Tager I, Expert Panel on P, Prevention Science of the American Heart A (2004) Air pollution and cardiovascular disease: a statement for healthcare professionals from the Expert Panel on Population and Prevention Science of the American Heart Association. Circulation 109:2655–2671. doi:10.1161/01.CIR.0000128587.30041.C8
Brook RD, Rajagopalan S, Pope CA 3rd, Brook JR, Bhatnagar A, Diez-Roux AV, Holguin F, Hong Y, Luepker RV, Mittleman MA, Peters A, Siscovick D, Smith SC Jr, Whitsel L, Kaufman JD, American Heart Association Council on E, Prevention CotKiCD, Council on Nutrition PA, Metabolism (2010) Particulate matter air pollution and cardiovascular disease: an update to the scientific statement from the American Heart Association. Circulation 121:2331–2378. doi:10.1161/CIR.0b013e3181dbece1
Brunekreef B, Holgate ST (2002) Air pollution and health. Lancet 360:1233–1242. doi:10.1016/S0140-6736(02)11274-8
Chen LC, Lippmann M (2009) Effects of metals within ambient air particulate matter (PM) on human health. Inhal Toxicol 21:1–31. doi:10.1080/08958370802105405
Chen Y, Shah N, Huggins FE, Huffman GP (2004) Investigation of the microcharacteristics of PM2.5 in residual oil fly ash by analytical transmission electron microscopy. Environ Sci Technol 38:6553–6560. doi:10.1021/es049872h
Diamond MS, Springer TA (1993) A subpopulation of Mac-1 (CD11b/CD18) molecules mediates neutrophil adhesion to ICAM-1 and fibrinogen. J Cell Biol 120:545–556
Dunne JL, Collins RG, Beaudet AL, Ballantyne CM, Ley K (2003) Mac-1, but not LFA-1, uses intercellular adhesion molecule-1 to mediate slow leukocyte rolling in TNF-alpha-induced inflammation. J Immunol 171:6105–6111. doi:10.4049/jimmunol.171.11.6105
Farraj AK, Haykal-Coates N, Winsett DW, Hazari MS, Carll AP, Rowan WH, Ledbetter AD, Cascio WE, Costa DL (2009) Increased non-conducted P-wave arrhythmias after a single oil fly ash inhalation exposure in hypertensive rats. Environ Health Perspect 117:709–715. doi:10.1289/ehp.0800129
Fujimaki H, Kurokawa Y, Yamamoto S, Satoh M (2006) Distinct requirements for interleukin-6 in airway inflammation induced by diesel exhaust in mice. Immunopharmacol Immunotoxicol 28:703–714. doi:10.1080/08923970601067433
Galkina E, Ley K (2007) Vascular adhesion molecules in atherosclerosis. Arterioscler Thromb Vasc Biol 27:2292–2301. doi:10.1161/ATVBAHA.107.149179
Gerdes N, Zirlik A (2011) Co-stimulatory molecules in and beyond co-stimulation—tipping the balance in atherosclerosis? Thromb Haemost 106:804–813. doi:10.1160/TH11-09-0605
Ghio AJ, Gilbey JG, Roggli VL, Richards JH, McGee JK, Carson JL, Devlin RB, Cascio WE (2001) Diffuse alveolar damage after exposure to an oil fly ash. Am J Respir Crit Care Med 164:1514–1518
Ghio AJ, Silbajoris R, Carson JL, Samet JM (2002) Biologic effects of oil fly ash. Environ Health Perspect 110:89–94
Ghio AJ, Suliman HB, Carter JD, Abushamaa AM, Folz RJ (2002) Overexpression of extracellular superoxide dismutase decreases lung injury after exposure to oil fly ash. Am J Physiol Lung Cell Mol Physiol 283:L211–L218. doi:10.1152/ajplung.00409.2001
Gurgueira SA, Lawrence J, Coull B, Murthy GG, Gonzalez-Flecha B (2002) Rapid increases in the steady-state concentration of reactive oxygen species in the lungs and heart after particulate air pollution inhalation. Environ Health Perspect 110:749–755
Hauser R, Elreedy S, Hoppin JA, Christiani DC (1995) Airway obstruction in boilermakers exposed to fuel oil ash. A prospective investigation. Am J Respir Crit Care Med 152:1478–1484
Hilgendorf I, Gerhardt LM, Tan TC, Winter C, Holderried TA, Chousterman BG, Iwamoto Y, Liao R, Zirlik A, Scherer-Crosbie M, Hedrick CC, Libby P, Nahrendorf M, Weissleder R, Swirski FK (2014) Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ Res 114:1611–1622. doi:10.1161/CIRCRESAHA.114.303204
Hiraiwa K, van Eeden SF (2013) Contribution of lung macrophages to the inflammatory responses induced by exposure to air pollutants. Mediat Inflamm 2013:619523. doi:10.1155/2013/619523
Hoek G, Brunekreef B, Fischer P, van Wijnen J (2001) The association between air pollution and heart failure, arrhythmia, embolism, thrombosis, and other cardiovascular causes of death in a time series study. Epidemiology 12:355–357
Iademarco MF, Barks JL, Dean DC (1995) Regulation of vascular cell adhesion molecule-1 expression by IL-4 and TNF-alpha in cultured endothelial cells. J Clin Invest 95:264–271. doi:10.1172/JCI117650
Kampfrath T, Maiseyeu A, Ying Z, Shah Z, Deiuliis JA, Xu X, Kherada N, Brook RD, Reddy KM, Padture NP, Parthasarathy S, Chen LC, Moffatt-Bruce S, Sun Q, Morawietz H, Rajagopalan S (2011) Chronic fine particulate matter exposure induces systemic vascular dysfunction via NADPH oxidase and TLR4 pathways. Circ Res 108:716–726. doi:10.1161/CIRCRESAHA.110.237560
Kim JY, Wand MP, Hauser R, Mukherjee S, Herrick RF, Christiani DC (2003) Association of expired nitric oxide with occupational particulate exposure. Environ Health Perspect 111:676–680
Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, Alvarado M, Anderson HR, Anderson LM, Andrews KG, Atkinson C, Baddour LM, Barker-Collo S, Bartels DH, Bell ML, Benjamin EJ, Bennett D, Bhalla K, Bikbov B, Bin Abdulhak A, Birbeck G, Blyth F, Bolliger I, Boufous S, Bucello C, Burch M, Burney P, Carapetis J, Chen H, Chou D, Chugh SS, Coffeng LE, Colan SD, Colquhoun S, Colson KE, Condon J, Connor MD, Cooper LT, Corriere M, Cortinovis M, de Vaccaro KC, Couser W, Cowie BC, Criqui MH, Cross M, Dabhadkar KC, Dahodwala N, De Leo D, Degenhardt L, Delossantos A, Denenberg J, Des Jarlais DC, Dharmaratne SD, Dorsey ER, Driscoll T, Duber H, Ebel B, Erwin PJ, Espindola P, Ezzati M, Feigin V, Flaxman AD, Forouzanfar MH, Fowkes FG, Franklin R, Fransen M, Freeman MK, Gabriel SE, Gakidou E, Gaspari F, Gillum RF, Gonzalez-Medina D, Halasa YA, Haring D, Harrison JE, Havmoeller R, Hay RJ, Hoen B, Hotez PJ, Hoy D, Jacobsen KH, James SL, Jasrasaria R, Jayaraman S, Johns N, Karthikeyan G, Kassebaum N, Keren A, Khoo JP, Knowlton LM, Kobusingye O, Koranteng A, Krishnamurthi R, Lipnick M, Lipshultz SE, Ohno SL et al (2012) Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380:2095–2128. doi:10.1016/S0140-6736(12)61728-0
Magnani ND, Marchini T, Tasat DR, Alvarez S, Evelson PA (2011) Lung oxidative metabolism after exposure to ambient particles. Biochem Biophys Res Commun 412:667–672. doi:10.1016/j.bbrc.2011.08.021
Marchini T, Magnani N, D’Annunzio V, Tasat D, Gelpi RJ, Alvarez S, Evelson P (2013) Impaired cardiac mitochondrial function and contractile reserve following an acute exposure to environmental particulate matter. Biochim Biophys Acta 1830:2545–2552. doi:10.1016/j.bbagen.2012.11.012
Marchini T, Magnani ND, Paz ML, Vanasco V, Tasat D, Gonzalez Maglio DH, Alvarez S, Evelson PA (2014) Time course of systemic oxidative stress and inflammatory response induced by an acute exposure to Residual Oil Fly Ash. Toxicol Appl Pharmacol 274:274–282. doi:10.1016/j.taap.2013.11.013
Marchini T, D’Annunzio V, Paz ML, Cáceres L, Garcés M, Perez V, Tasat D, Vanasco V, Magnani N, Gonzalez Maglio D, Gelpi RJ, Alvarez S, Evelson P (2015) Selective TNF-α targeting with infliximab attenuates impaired oxygen metabolism and contractile function induced by an acute exposure to air particulate matter. Am J Physiol Heart Circ Physiol 309:H1621–H1628. doi:10.1152/ajpheart.00359.2015
Nahrendorf M, Pittet MJ, Swirski FK (2010) Monocytes: protagonists of infarct inflammation and repair after myocardial infarction. Circulation 121:2437–2445. doi:10.1161/CIRCULATIONAHA.109.916346
Nel A (2005) Atmosphere. Air pollution-related illness: effects of particles. Science 308:804–806. doi:10.1126/science.1108752
Nemmar A, Hoet PH, Vanquickenborne B, Dinsdale D, Thomeer M, Hoylaerts MF, Vanbilloen H, Mortelmans L, Nemery B (2002) Passage of inhaled particles into the blood circulation in humans. Circulation 105:411–414. doi:10.1161/hc0402.104118
Nurkiewicz TR, Porter DW, Barger M, Castranova V, Boegehold MA (2004) Particulate matter exposure impairs systemic microvascular endothelium-dependent dilation. Environ Health Perspect 112:1299–1306
Nurkiewicz TR, Porter DW, Barger M, Millecchia L, Rao KM, Marvar PJ, Hubbs AF, Castranova V, Boegehold MA (2006) Systemic microvascular dysfunction and inflammation after pulmonary particulate matter exposure. Environ Health Perspect 114:412–419
Ostachuk A, Evelson P, Martin S, Dawidowski L, Sebastian Yakisich J, Tasat DR (2008) Age-related lung cell response to urban Buenos Aires air particle soluble fraction. Environ Res 107:170–177. doi:10.1016/j.envres.2008.01.007
Oyama J, Blais C Jr, Liu X, Pu M, Kobzik L, Kelly RA, Bourcier T (2004) Reduced myocardial ischemia-reperfusion injury in toll-like receptor 4-deficient mice. Circulation 109:784–789. doi:10.1161/01.CIR.0000112575.66565.84
Pattanaik S, Huggins FE, Huffman GP (2012) Chemical speciation of Fe and Ni in residual oil fly ash fine particulate matter using X-ray absorption spectroscopy. Environ Sci Technol 46:12927–12935. doi:10.1021/es301080s
Pope CA 3rd, Muhlestein JB, May HT, Renlund DG, Anderson JL, Horne BD (2006) Ischemic heart disease events triggered by short-term exposure to fine particulate air pollution. Circulation 114:2443–2448. doi:10.1161/CIRCULATIONAHA.106.636977
Riediker M (2007) Cardiovascular effects of fine particulate matter components in highway patrol officers. Inhal Toxicol 19:99–105. doi:10.1080/08958370701495238
Roberts JR, Young SH, Castranova V, Antonini JM (2009) The soluble nickel component of residual oil fly ash alters pulmonary host defense in rats. J Immunotoxicol 6:49–61. doi:10.1080/15476910802630379
Southam DS, Dolovich M, O’Byrne PM, Inman MD (2002) Distribution of intranasal instillations in mice: effects of volume, time, body position, and anesthesia. Am J Physiol Lung Cell Mol Physiol 282:L833–L839. doi:10.1152/ajplung.00173.2001
Stachon P, Peikert A, Michel NA, Hergeth S, Marchini T, Wolf D, Dufner B, Hoppe N, Ayata CK, Grimm M, Cicko S, Schulte L, Reinohl J, von zur Muhlen C, Bode C, Idzko M, Zirlik A (2014) P2Y6 deficiency limits vascular inflammation and atherosclerosis in mice. Arterioscler Thromb Vasc Biol 34:2237–2245. doi:10.1161/ATVBAHA.114.303585
van Rooijen N, Hendrikx E (2010) Liposomes for specific depletion of macrophages from organs and tissues. Methods Mol Biol 605:189–203. doi:10.1007/978-1-60327-360-2_13
Wallenborn JG, McGee JK, Schladweiler MC, Ledbetter AD, Kodavanti UP (2007) Systemic translocation of particulate matter-associated metals following a single intratracheal instillation in rats. Toxicol Sci 98:231–239. doi:10.1093/toxsci/kfm088
Wellenius GA, Saldiva PH, Batalha JR, Krishna Murthy GG, Coull BA, Verrier RL, Godleski JJ (2002) Electrocardiographic changes during exposure to residual oil fly ash (ROFA) particles in a rat model of myocardial infarction. Toxicol Sci 66:327–335. doi:10.1093/toxsci/66.2.327
Wolf D, Hohmann JD, Wiedemann A, Bledzka K, Blankenbach H, Marchini T, Gutte K, Zeschky K, Bassler N, Hoppe N, Rodriguez AO, Herr N, Hilgendorf I, Stachon P, Willecke F, Duerschmied D, von zur Muhlen C, Soloviev DA, Zhang L, Bode C, Plow EF, Libby P, Peter K, Zirlik A (2011) Binding of CD40L to Mac-1′s I-domain involves the EQLKKSKTL motif and mediates leukocyte recruitment and atherosclerosis—but does not affect immunity and thrombosis in mice. Circ Res 109:1269–1279. doi:10.1161/CIRCRESAHA.111.247684
Wolf D, Jehle F, Anto Michel N, Bukosza EN, Rivera J, Chen YC, Hoppe N, Dufner B, Ortiz Rodriguez A, Colberg C, Nieto L, Rupprecht B, Wiedemann A, Schulte L, Peikert A, Bassler N, Lozhkin A, Hergeth SP, Stachon P, Hilgendorf I, Willecke F, von Zur Muhlen C, von Elverfeldt D, Binder CJ, Aichele P, Varo N, Febbraio MA, Libby P, Bode C, Peter K, Zirlik A (2014) Co-inhibitory suppression of T cell activation by CD40 protects from obesity and adipose tissue inflammation in mice. Circulation 129:2414–2425. doi:10.1161/CIRCULATIONAHA.113.008055
Wolf D, Stachon P, Bode C, Zirlik A (2014) Inflammatory mechanisms in atherosclerosis. Hamostaseologie 34:63–71. doi:10.5482/HAMO-13-09-0050
Wu S, Deng F, Wei H, Huang J, Wang H, Shima M, Wang X, Qin Y, Zheng C, Hao Y, Guo X (2012) Chemical constituents of ambient particulate air pollution and biomarkers of inflammation, coagulation and homocysteine in healthy adults: a prospective panel study. Part Fibre Toxicol 9:49. doi:10.1186/1743-8977-9-49
Yatera K, Hsieh J, Hogg JC, Tranfield E, Suzuki H, Shih CH, Behzad AR, Vincent R, van Eeden SF (2008) Particulate matter air pollution exposure promotes recruitment of monocytes into atherosclerotic plaques. Am J Physiol Heart Circ Physiol 294:H944–H953. doi:10.1152/ajpheart.00406.2007
Zanobetti A, Schwartz J (2007) Particulate air pollution, progression, and survival after myocardial infarction. Environ Health Perspect 115:769–775. doi:10.1289/ehp.9201
Zasłona Z, Przybranowski S, Wilke C, van Rooijen N, Teitz-Tennenbaum S, Osterholzer JJ, Wilkinson JE, Moore BB, Peters-Golden M (2014) Resident alveolar macrophages suppress, whereas recruited monocytes promote, allergic lung inflammation in murine models of asthma. J Immunol 193:4245–4253. doi:10.4049/jimmunol.1400580
Zhang WC, Wang YG, Zhu ZF, Wu FQ, Peng YD, Chen ZY, Yang JH, Wu JJ, Lian YT, He MA, Wu TC, Cheng LX (2014) Regulatory T cells protect fine particulate matter-induced inflammatory responses in human umbilical vein endothelial cells. Mediat Inflamm 2014:869148. doi:10.1155/2014/869148
Zouggari Y, Ait-Oufella H, Bonnin P, Simon T, Sage AP, Guerin C, Vilar J, Caligiuri G, Tsiantoulas D, Laurans L, Dumeau E, Kotti S, Bruneval P, Charo IF, Binder CJ, Danchin N, Tedgui A, Tedder TF, Silvestre JS, Mallat Z (2013) B lymphocytes trigger monocyte mobilization and impair heart function after acute myocardial infarction. Nat Med 19:1273–1280. doi:10.1038/nm.3284
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding sources
This study was supported by research grants from the Deutsche Forschungsgemeinschaft (DFGZI743/3-1 and 3-2) and from the Else-Kröner-Fresenius Stiftung (EKFSP30/10 and A43/10) to Dr. Zirlik. Dr. Marchini was funded by a fellowship from the German Academic Exchange Service (DAAD) and from the Argentinean National Scientific and Technical Research Council (CONICET).
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
T. Marchini and D. Wolf contributed equally to this work.
P. Evelson and A. Zirlik share senior authorship.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Marchini, T., Wolf, D., Michel, N.A. et al. Acute exposure to air pollution particulate matter aggravates experimental myocardial infarction in mice by potentiating cytokine secretion from lung macrophages. Basic Res Cardiol 111, 44 (2016). https://doi.org/10.1007/s00395-016-0562-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00395-016-0562-5