
Abstract
Background
Vascular endothelial growth factor A (VEGF-A) plays an integral role in angiogenesis by contributing to growth, development, and metastasis of solid tumors. Recently, a single-nucleotide polymorphism +936C/T located in the VEGF-A 3′ untranslated region (UTR) facilitated the susceptibility of colorectal cancer. The association between VEGF-A gene polymorphism +936C/T and colorectal cancer risk has been widely studied in the last decade, but presently, the results furnished remain enigmatic. Hence, the study aimed to investigate the association between VEGF-A +936C/T miRNA binding site polymorphism and the risk of developing colorectal cancer.
Methods
This meta-analysis included 13 published case-control studies covering 3465 cases (colorectal cancer) and 3476 healthy controls. Publication bias was examined by means of Begg’s funnel plots and Egger’s regression tests. The quality of the studies included was evaluated using Newcastle-Ottawa scale. Subgroup analyses were performed in accordance to the various ethnicities of the study subjects and the study quality.
Results
From the data obtained, it is implied that VEGF-A +936C/T polymorphism did not correlate with elevated colorectal cancer risk in all genetic models. But the results acquired from the subgroup analysis in over dominant model (CT vs. CC + TT: OR = 1.5047, 95% CI = 1.19–1.90) suggest that VEGF-A +936C/T polymorphism leads to the raise in the risk of developing CRC among the East Asian population. No association was observed in Caucasian and South Asian population.
Conclusions
Our results indicate that VEGF-A +936C/T polymorphism is not a risk factor for developing CRC in Caucasian and South Asian population. However, the East Asian population was related to an increased risk of developing colorectal cancer due to the presence of the minor allele.
Similar content being viewed by others


Background
Colorectal cancer (CRC) is currently the third most widespread cancer type and the foremost cause of cancer mortality in both men and women worldwide [1, 2]. It has been assessed that 1.4 million individuals are suffering from CRC every year, 65% of whom are from developed nations; this ailment causes about 700,000 deaths every year, and 3.5 million people continue to live with CRC [3]. It is a multifaceted disease which occurs due to the influence of numerous reasons such as environmental factors and genetic variations [4, 5], which are the root cause to considerably influence the risk of CRC [6, 7]. Angiogenesis is the process of production of new blood vessels that play a vital role in cancer development and metastases. The vascular endothelial growth factor A (VEGF-A), a heparin-binding glycoprotein, contributes to mitogenic, angiogenic, and vascular permeability activities explicit for endothelial cells. VEGF-A gene is located at chromosome 6 and comprises of eight exons [8]. VEGF-A gene belongs to the VEGF family; it encompasses four VEGF amino acid isomeric residues VEGF165, VEGF189, VEGF121, and VEGF206. This gene is extremely polymorphic, its promoter, 5′ and 3′ untranslated regions (UTRs), has a diverse range of single-nucleotide polymorphism SNPs [9]. MicroRNAs (miRNAs) are a group of single-stranded noncoding RNA molecules which regulate gene expression by binding to cognate sequences of the 3′ UTR regions of mRNAs. Their binding leads to reduction in protein translation and an increase in mRNA degradation [10]. SNPs present in the 3′ UTR targeted by miRNAs can either eliminate existing binding sites or produce illegitimate binding sites or can affect miRNA:mRNA interactions and target the mRNA expression [11]. This process results in the regulation and expression of target genes.
There are more than 15 VEGF SNPs that have been reported in diverse types of cancers [12, 13]. VEGF SNPs such as +936C > T, −2578C > A, +405C > G, -634G>C, −460C > T, and −1154G > A have been extensively studied [14]. SNPs located in the 3′ UTR region of the VEGF gene were found to be correlated with variations in VEGF protein production [15]. Among the many polymorphisms, +936C/T polymorphism located at miR-199a binding site in the VEGF-A 3′ UTR region has been demonstrated to perform a functional role. This polymorphism has been significantly associated with different types of cancer like oral, breast, colorectal cancer [16, 17], and other diseases with a recognized angiogenic basis [18].
MiR-199 is an imperative vertebrate-specific miRNA, which is associated with a wide variety of cellular and developmental mechanisms like tumor growth and progression [19]. It has been reported that MiR-199a downregulates and also suppresses tumor progression in prostate adenocarcinoma [20], chondrosarcoma [21], hepatocellular carcinoma [22], and ovarian cancer [23]. In recent times, it has been testified that miR-199a is unusually expressed in CRC [24,25,26,27]. MiR-199a plays an important role in repressing the migration and invasion of CRC cells in hypoxia-inducible factor 1-alpha/vascular endothelial growth factor (HIF-1α/VEGF) pathway and targets the discoidin domain receptor 1 [28, 29]. Shweiki et al. reported that VEGF-A production was controlled by numerous stimuli, among which hypoxia is the most significant one [30]. In hypoxic conditions, HIF-1α translocates into the nucleus and heterodimerizes with HIF-1β by binding to hypoxia-responsive elements on several genes, such as VEGF-A [31]. The underlying mechanism of miR-199a in regulation, development, and progression of CRC still remains indistinct as miR-199a has numerous target genes [32], which may be involved in tumor initiation and progression of CRC. Therefore, SNPs situated in the miRNA binding sites may affect the expression of miRNA target genes and contribute to the susceptivity of humans to develop diseases [33,34,35,36]. Thereby, it is hypothesized that SNPs present in the potential miRNA binding sites of VEGF-A gene may influence the susceptibility and progression of colorectal cancer.
Ongoing studies have demonstrated that SNPs situated in the VEGF-A gene may contribute to the development of colorectal cancer. Hence, it is commonly speculated that due to the varied ethnicities and inadequate sample size, the results of these studies remain inconclusive. Thus, this updated meta-analysis includes all eligible case-control studies which were implemented to inspect whether VEGF-A miRNA binding site polymorphism +936C/T was associated with the risk of developing colorectal cancer.
Methods
Literature search criteria
A systematic literature search investigating the association of VEGF-A gene polymorphism and colorectal cancer risk was performed utilizing PubMed (www.ncbi.nlm.nih.gov/pubmed), ScienceDirect (www.sciencedirect.com), and Google Scholar (www.scholar.google.com) databases to find relevant publications up to June 2020. The systematic search was conducted adopting different combinations of relevant keywords: colorectal neoplasm, polymorphism, and genetic vascular endothelial growth factor. Furthermore, the reference lists of original studies were examined manually for additional literature. All the appropriate studies, abstracts, and titles were checked cautiously to prevent duplication of datasets.
Inclusion and exclusion criteria
Studies that were included in the present meta-analysis had to comply the following norms: (1) only case-control studies, (2) studies which focused on +936C/T polymorphism and colorectal cancer risk, (3) adequate data about allele or genotype frequencies containing +936C/T genotypes (CC, CT, and TT) which could be expressed as odds ratio (OR) and corresponding 95% confidence interval (95% CI), and (4) only studies on human subjects with full content in English. Accordingly, the following exclusion criteria were also used: (1) duplication of retrieved information; (2) studies that have only case population details, reviews, abstracts, and editorials that were published in journals; (3) studies with inadequate genotype data; (4) pharmacogenetic studies, pharmacokinetic studies, interim analysis, and case reports were excluded; and (5) if a study fails to satisfy Hardy–Weinberg equilibrium (HWE).
Data extraction and quality assessment
Data was independently and separately collected and checked for discrepancies by two investigators (KSS and ZSS). The studies included in the meta-analysis are listed in Table 1. The data collected for the study comprised the name of the first author, year of publication, population, ethnicity, genotyping method, sample size of case and control groups, genotype distributions in case and control groups, HWE, and p-values for controls.
Statistical analysis
Pooled odds ratio (OR) and corresponding 95% confidence interval limits were calculated for evaluating the strength of the associations between the VEGF-A +936C/T polymorphism with susceptibility to colorectal cancer. The pooled odds ratios were calculated by fixed effects model or random effects model, according to the heterogeneity level. The amount of heterogeneity was identified by performing the Cochran’s Q-test and the I2 test. The amount of heterogeneity was calculated according to the following scale: 75% ≥ I2 = very severe heterogeneity, 50 ≥ I2 < 75% = severe heterogeneity, 25 ≥ I2 < 50% = moderate heterogeneity, and I2 < 25% = very less heterogeneity; high-resolution forest plots were prepared to portray both OR and 95% CI limits. To assess publication bias, we examined funnel plots. As there was minimal inter-study heterogeneity, sensitivity analysis was not performed. All included studies were tested for genotypic distribution of the VEGF-A +936C/T polymorphism in the control group with the HWE principle using the chi-square goodness-of-fit test. p < 0.05 was considered as statistically significant. Subgroup analysis was performed to check the association between VEGF-A +936C/T polymorphism and the ethnicity of the study subjects. The analysis was conducted using a gratuitous web tool MetaGenyo [37], a web application framework for RStudio. Backend computations were carried out in R using available packages and custom scripts.
Quality assessment of the studies
The quality of all the studies included in the meta-analysis was assessed by Newcastle-Ottawa scale (NOS) [38]. NOS is a star system which allows semiquantitative evaluation of nonrandomized study quality; it comprises of eight items which are categorized into three major components such as selection, comparability and exposure (case-control studies), or outcome (cohort studies). The scale ranges from zero to nine stars; the number of stars represented the highest methodological quality. The study is considered to be of good quality if the total score is above 5. Subgroup analysis was performed according to the quality of the study and the total score obtained.
Results
Meta-analysis
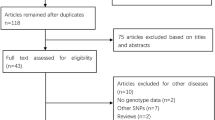
The process implemented for retrieval and selection of papers in this meta-analysis is shown in Fig. 1. The baseline characteristics of the included studies are briefed in Table 1. A total of 553 studies were retrieved after a comprehensive search in the electronic databases available, out of which 535 studies were omitted after reading the abstract as they did not match the study criteria. Of the remaining 18 studies, 2 studies did not satisfy HWE, and 3 studies had inadequate genotype data; eventually, 13 studies were included in the study [39,40,41,42,43,44,45,46,47,48,49,50,51]. Out of these seven belonged to Caucasian population, five belonged to East Asians, and one South Asian. The quality of studies included was assessed, and points were given on a scale of five to nine (Table 2). Thirteen studies were of good quality (Table 2).

A schematic, to set forth the steps taken for the meta-analysis study
Association between VEGF-A +936C/T polymorphism and colorectal cancer risk
Thirteen studies compared the association of VEGF-A +936C/T with colorectal cancer patients including a sample size of n = 3465 and healthy controls having a sample size of n = 3476. Fixed-effects model was used; pooled OR investigations and the results were represented as forest plots (Fig. 2 a and b). With respect to the OR and 95% CI, no significant association was observed between VEGF-A +936C/T polymorphism and colorectal cancer (Fig. 2 a and b; Table 3) in the allelic (C vs. T: OR = 0.99; 95% CI = 0.91–1.09); recessive (CC vs. CT+TT: OR =1.02; 95% CI = 0.9202–1.1352); dominant (CC + CT vs. TT: OR = 0.80; 95% CI = 0.58–1.10); overdominant (CT vs. CC + TT: OR = 0.95; 95% CI = 0.85–1.06); and the CC vs. CT : (OR = 1.04; 95% CI = 0.93–1.16) genetic models. However, the subgroup analysis revealed significant results in the overdominant model (CT vs. CC + TT: OR = 1.5047, 95% CI = 1.19–1.90) in East Asians (Table 4). In Caucasian population, no association was observed in the overdominant model (CT vs. CC + TT: OR = 0.8843, 95% CI = 0.74–1.04). The funnel plot of studies evaluating the role of +936C/T appeared symmetric (Fig. 3), suggesting the absence of publication bias (Egger’s test p-value = 0.8515). The subgroup analysis for the study quality revealed significant results in the CC vs. CT model (OR = 1.82, 95% CI = 1.07–3.12). The forest plot for the recessive model showed heterogeneity of I2 =66%, and CC vs. CT model showed I2 = 63% (Fig. 2 a and b).

a Association between VEGF-A +936C/T polymorphism and colorectal cancer risk in CC vs. CT + TT model presented as a forest plot. b Association between VEGF-A +936C/T polymorphism and colorectal cancer risk in CC vs. CT model presented as a forest plot

Association between VEGF-A +936C/T polymorphism and colorectal cancer risk presented as a funnel plot
Discussion
VEGF is an imperative regulator of tumor angiogenesis, associated with the development and progression of multiple cancers [52]. This gene portrays a pivotal role and acts as an important prognostic factor in a variety of tumors, including CRC. Numerous studies related to the risk and diagnosis of breast cancer [53] and non-small cell lung cancer [54] have demonstrated their impact on VEGF SNPs. Several studies have also revealed that VEGF protein production during colorectal carcinogenesis is correlated with polymorphisms situated in the 5′ and 3′ UTR of the VEGF gene and their promoter region [13, 55]. However, very limited studies have reported about the association of VEGF-A 3′ UTR miRNA SNPs with the susceptibility to CRC.
MiR-199a is an intronic miRNA discovered in 2003 and associated with development of various diseases [56]. It has been reported that MiR-199a is a possible inhibitor of HIF-1α/VEGF pathway. MiR-199a targets the 3′ UTR of HIF-1α and HIF-2α and leads to the decrease in hypoxia-increased HIF levels. HIF-1α is an important transcription factor, which plays a crucial role in CRC development and progression [57]. Overexpression of VEGF gene in CRC cells is observed often; this gene portrays a vital role in angiogenesis and cell proliferation which makes it a potential target for cancer therapy. The mechanism involved in VEGF-A 3′ UTR binding site polymorphism regulating the development and progression of CRC remains scanty. Currently, there are very few studies related to miR199a significance in angiogenesis and colorectal cancer. Hence, we wanted to elucidate the contribution of VEGF-A 3′ UTR binding site polymorphism to colorectal cancer by performing a meta-analysis.
The 3′ UTR of the VEGF gene has demonstrated to enhance the stability of mRNA and also leads to the hypoxic induction of the VEGF gene [18, 58, 59]. Recently, genes designated as Hu family have been ascertained, and their products have shown to bind to the AU-rich element of 3′ UTR of numerous genes, including the VEGF mRNA [18, 60]. It is proposed that the proteins of the Hu family change the VEGF mRNA conformation so that the mRNA is not affected by RNase. SNPs in the 3′ UTR have shown to be related with the deregulation of the affected genes [61]. Therefore, the SNPs in the 3′ UTR may modify the mRNA conformational integrity, bringing about genetic variation of VEGF gene expression. The VEGF +936 C/T is a significant functional polymorphism that has demonstrated to alter the susceptibility of various diseases such as cancer [62, 63]. Chen et al. stated that the total viability of non-small cell lung carcinoma patients and their response to chemotherapy was afflicted of VEGF +936C/T gene polymorphism [55]. Zhang et al. reported that VEGF +936C/T was significantly correlated with glioma susceptibility and may act as a genetic marker [64]. VEGF +936C/T serves as a marker for disease aggression, relapse, and an important factor for the poor prognosis of epithelial ovarian cancer [65].
Most of the studies reported lack of association between CRC and +936C/T in Caucasian population [47, 51, 66]. These results were consistent with studies recently published in the Caucasian population [40, 41]. Our meta-analysis showed concordance to these results since no association was observed in Caucasian population (Table 4). However, the +936T allele increases the risk of CRC in East Asian population such as Korean and Chinese populations [47, 49]. Similarly, Jang et al. reported that the +936T allele was associated with an increased susceptibility to CRC [44, 67]. These results were concordant to the results obtained in our meta-analysis (Table 4).
Jeon et al. study dealing with the association amidst VEGF 3′ UTR polymorphisms and CRC susceptibility in Koreans stated that VEGF 1451C > T and 1725G > A could render to CRC susceptibility [68]. Also, metastasis and angiogenesis are eminently connected to VEGF expression in solid tumors. VEGF +936-T allele leads to reduced plasma VEGF levels in young healthy population of Caucasians [69]; on the other hand, the underlying mechanism remains to be elusive. Two interpretations were put forth; (1) +936 C/T mutation induces the loss of a potential binding site for AP-4, which is a transcription factor enhancing expression of several viral and cellular genes by binding to specific enhancer sites [70, 71]. (2) This variant may hinder the binding of hypoxia-induced protein to the 3′ UTR of VEGF-A mRNA, which may lead to a significantly diminished half-life of the mRNA.
Subsequently, the polymorphisms in VEGF-A gene 3′ UTR can alter the implied binding sites of transcription factors, which stimulate impaired proteins and malformations. This malformation could illustrate why the +936-T allele carriers have a lower risk of developing cancer like breast cancer, small cell lung cancer, and oral squamous cell carcinoma [72, 73]. Lack of association was observed in VEGF-A +936 C/T polymorphism and CRC in Han Chinese in Sichuan province subsequent to Bonferroni correction [39]. Studies in large numbers are requisite to directly check for miRNA binding activity to VEGF-A 3′ UTR polymorphisms and to regulate the mechanism by which these polymorphisms might have an effect on cellular proliferation and cancer advancement. These studies might have a great clinical impact for all diseases associated with abnormal angiogenesis and hypoxic conditions.
Majority of studies reported a lack of association between CRC and VEGF-A +936C/T polymorphism. Lately, few case-control studies have been carried out to examine the association between VEGF gene polymorphisms and CRC susceptibility [39,40,41]. However, the possible effect of VEGF-A +936C/T polymorphisms on VEGF-A production as well as tumor development and progression in CRC still remains ambiguous. Hence, the purpose of this study was to investigate the effect of VEGF-A +936C/T polymorphisms on susceptibility to CRC by means of a meta-analysis.
The current meta-analysis included 13 case-control studies, which consists of 3465 CRC cases and 3476 controls. Our meta-analysis suggests that the VEGF-A +936C/T gene polymorphism is not associated with the risk of developing CRC. In this investigation, we did not find any evidence of publication bias as shown in Fig. 3. VEGF-A gene polymorphisms have been associated with susceptibility to several cancer types. Previously, two meta-analyses have been performed on −460T/C, −634G/C, +936C/T, −2578C/A, −1154G/A, and + 405C/G [14, 74]. These two meta-analyses indicated that VEGF +936C/T demonstrated no association with colorectal cancer. Our updated meta-analysis results show association among East Asian population which shows concordance to meta-analysis conducted by Gholami et al. [75]. From our study, we have also observed that in the CT vs. CC+TT, OR is 1.5 for East Asians. When compared with the overall pooled odds ratio for the overdominant model (OR = 0.9525; p-value = 0.37577), the East Asian subgroup exhibits an increase in the odds ratio indicating that the SNPs effect varies with ethnicity. Several studies have focused on VEGF-A +936C/T polymorphism for its possible association with colorectal cancer patients. This is the first meta-analysis to be solely conducted on VEGF-A miRNA binding site polymorphism +936C/T in colorectal cancer.
Our study has few possible limitations. (a) Only published studies were included in the meta-analysis, (b) only three databases were searched for relevant articles, (c) Lack of genotype frequency information provided by some published studies, (d) HWE not getting satisfied did not allow the estimation of the best genetic model of inheritance to follow, (e) all case-control studies were obtained from Asian and Caucasian population, (f) and gene-gene and gene-environment interactions were not accounted. Finally, although all cases and controls of each study were explicitly defined with similar inclusion criteria, there may be some potential factors that were not taken into account that may have influenced our results.
Conclusions
To conclude, this meta-analysis suggests that the VEGF-A +936C/T gene polymorphism is not associated with the risk of developing CRC in Caucasian and South Asian population. But the minor allele present in the East Asian population was related to an increased risk of developing CRC. Based on our findings, additional larger population-based studies with diverse ethnic groups are imperative to validate the association of VEGF-A gene polymorphisms and colorectal cancer.
Availability of data and materials
Not applicable.
Abbreviations
- VEGF-A:
-
Vascular endothelial growth factor A
- UTR:
-
Untranslated region
- MiRNA:
-
MicroRNA
- mRNA:
-
Messenger RNA
- CRC:
-
Colorectal cancer
- HIF-1α:
-
Hypoxia-inducible factor 1-alpha
- NOS:
-
Newcastle-Ottawa scale
- HWE:
-
Hardy–Weinberg equilibrium
- OR:
-
Odds ratio
- CI:
-
Confidence interval
References
Haggar FA, Boushey RP. Colorectal cancer epidemiology: incidence, mortality, survival, and risk factors. Clin Colon Rectal Surg. 2009;22(4):191–7. https://doi.org/10.1055/s-0029-1242458.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA-Cancer J Clin. 2018;68(6):394–424. https://doi.org/10.3322/caac.21492.
Rabeneck L, Horton S, Zauber AG, Earle C. Colorectal Cancer. In H: Gelband (Ed) et. al, Cancer: disease control priorities, 3rd ed. Washington (DC): The International Bank for Reconstruction and Development / The World Bank; 2015. p. 101–19.
Wang F, Zhang SD, Xu HM, Zhu JH, Hua RX, Xue WQ, et al. XPG rs2296147 T>C polymorphism predicted clinical outcome in colorectal cancer. Oncotarget. 2016;7(10):11724–32. https://doi.org/10.18632/oncotarget.7352.
Mattis AN, Song G, Hitchner K, Kim RY, Lee AY, Sharma AD, et al. A screen in mice uncovers repression of lipoprotein lipase by microRNA-29a as a mechanism for lipid distribution away from the liver. Hepatology. 2015;61(1):141–52. https://doi.org/10.1002/hep.27379.
Azeem S, Gillani SW, Siddiqui A, Jandrajupalli SB, Poh V, Syed Sulaiman SA. Diet and colorectal cancer risk in Asia-a systematic review. Asian Pac J Cancer Prev. 2015;16(13):5389–96. https://doi.org/10.7314/apjcp.2015.16.13.5389.
Liu L, Zhong R, Wei S, Yin J, Xiang H, Zou L, et al. Interactions between genetic variants in the adiponectin, adiponectin receptor 1 and environmental factors on the risk of colorectal cancer. PLoS One. 2011;6(11):e27301. https://doi.org/10.1371/journal.pone.0027301.
Wei MH, Popescu NC, Lerman MI, Merrill MJ, Zimonjic DB. Localization of the human vascular endothelial growth factor gene, VEGF, at chromosome 6p12. Hum Genet. 1996;97(6):794–7 PMID: 8641698.
Wang L, Ji S, Cheng Z. Association between polymorphisms in vascular endothelial growth factor gene and response to chemotherapies in colorectal cancer: a meta-analysis. PLoS One. 2015;10(5):e0126619. https://doi.org/10.1371/journal.pone.0126619.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97. https://doi.org/10.1016/s0092-8674(04)00045-5.
Chen K, Song F, Calin GA, Wei Q, Hao X, Zhang W. Polymorphisms in microRNA targets: a gold mine for molecular epidemiology. Carcinogenesis. 2008;29:1306–11. https://doi.org/10.1093/carcin/bgn116.
Vincenti V, Cassano C, Rocchi M, Persico G. Assignment of the vascular endothelial growth factor gene to human chromosome 6p21.3. Circulation. 1996;93(8):1493–5. https://doi.org/10.1161/01.cir.93.8.1493.
Watson CJ, Webb NJ, Bottomley MJ, Brenchley PE. Identification of polymorphisms within the vascular endothelial growth factor (VEGF) gene: correlation with variation in VEGF protein production. Cytokine. 2000;12(8):1232–5. https://doi.org/10.1006/cyto.2000.0692.
Zhou LP, Luan H, Dong XH, Jin GJ, Man DL, Shang H. Vascular endothelial growth factor gene polymorphisms and colorectal cancer risk: a meta-analysis. Genet Mol Res. 2011;10(4):3674–88. https://doi.org/10.4238/2011.October.31.13.
Koukourakis MI, Papazoglou D, Giatromanolaki A, Bougioukas G, Maltezos E, Siviridis E. VEGF gene sequence variation defines VEGF gene expression status and angiogenic activity in non-small cell lung cancer. Lung Cancer. 2004;46(3):293–8. https://doi.org/10.1016/j.lungcan.2004.04.037.
Zhao SF, Zhan P, Yang XD, Lu MX, Sun GW, Wang YX, et al. VEGF+936C/T and+ 460C/T gene polymorphisms and oral cancer risk: a meta-analysis. Mol Biol Rep. 2013;40(12):6637–43. https://doi.org/10.1007/s11033-013-2778-x.
Yan Y, Liang H, Li T, Guo S, Li M, Li S, et al. Vascular endothelial growth factor +936C/T polymorphism and breast cancer risk: a meta-analysis of 13 case–control studies. Tumour Biol. 2014;35(3):2687–92. https://doi.org/10.1007/s13277-013-1354-2.
Awata T, Inoue K, Kurihara S, Ohkubo T, Watanabe M, Inukai K, et al. A common polymorphism in the 5’-untranslated region of the VEGF gene is associated with diabetic retinopathy in type 2 diabetes. Diabetes. 2002;51(5):1635–9. https://doi.org/10.2337/diabetes.51.5.1635.
Gu S, Chan WY. Flexible and versatile as a chameleon—sophisticated functions of microRNA-199a. Int J Mol Sci. 2012;13(7):8449–66. https://doi.org/10.3390/ijms13078449.
Zhong J, Huang R, Su Z, Zhang M, Xu M, Gong J, et al. Downregulation of miR-199a-5p promotes prostate adeno-carcinoma progression through loss of its inhibition of HIF-1α. Oncotarget. 2017;8(48):83523–38. https://doi.org/10.18632/oncotarget.18315.
Liu GT, Huang YL, Tzeng HE, Tsai CH, Wang SW, Tang CH. CCL5 promotes vascular endothelial growth factor expression and induces angiogenesis by down-regulating miR-199a in human chondrosarcoma cells. Cancer Lett. 2015;357(2):476–87. https://doi.org/10.1016/j.canlet.2014.11.015.
Ghosh A, Dasgupta D, Ghosh A, Roychoudhury S, Kumar D, Gorain M, et al. MiRNA199a-3p suppresses tumor growth, migration, invasion and angiogenesis in hepatocellular carcinoma by targeting VEGFA, VEGFR1, VEGFR2, HGF and MMP2. Cell Death Dis. 2017;8(3):e2706. https://doi.org/10.1038/cddis.2017.123.
Gadducci A, Sergiampietri C, Lanfredini N, Guiggi I. Micro-RNAs and ovarian cancer: the state of art and perspectives of clinical research. Gynecol Endocrinol. 2014;30(4):266–71. https://doi.org/10.3109/09513590.2013.871525.
Kim BK, Yoo HI, Kim I, Park J, Yoon S. FZD6 expression is negatively regulated by miR-199a-5p in human colorectal cancer. BMB Rep. 2015;48(6):360–6. https://doi.org/10.5483/bmbrep.2015.48.6.031.
Han Y, Kuang Y, Xue X, Guo X, Li P, Wang X, et al. NLK, a novel target of miR-199a-3p, functions as a tumor suppressor in colorectal cancer. Biomed Pharmacother. 2014;68(5):497–505. https://doi.org/10.1016/j.biopha.2014.05.003.
Mudduluru G, Ceppi P, Kumarswamy R, Scagliotti GV, Papotti M, Allgayer H. Regulation of Axl receptor tyrosine kinase expression by miR-34a and miR-199a/b in solid cancer. Oncogene. 2011;30(25):2888–99. https://doi.org/10.1038/onc.2011.13.
Kong Y, Bai PS, Sun H, Nan KJ, Chen NZ, Qi XG. The deoxycholic acid targets miRNA-dependentCAC1 gene expression in multidrug resistance of human colorectal cancer. Int J Biochem Cell Biol. 2012;44(12):2321–32. https://doi.org/10.1016/j.biocel.2012.08.006.
Ye H, Pang L, Wu Q, Zhu Y, Guo C, Deng Y, et al. A critical role of mir-199a in the cell biological behaviors of colorectal cancer. Diagn Pathol. 2015;10(1):65. https://doi.org/10.1186/s13000-015-0260-x.
Hu Y, Liu J, Jiang B, Chen J, Fu Z, Bai F, et al. MiR-199a-5p loss up-regulated DDR1 aggravated colorectal cancer by activating epithelial-to mesenchymal transition related signaling. Dig Dis Sci. 2014;59(9):2163–72. https://doi.org/10.1007/s10620-014-3136-0.
Shweiki D, Itin A, Soffer D, Keshet E. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature. 1992;359(6398):843–5. https://doi.org/10.1038/359843a0.
Harris AL. Hypoxia—a key regulatory factor in tumour growth. Nat Rev Cancer. 2002;2(1):38–47. https://doi.org/10.1038/nrc704.
John B, Enright AJ, Aravin A, Tuschl T, Sander C, Marks DS. Human microRNA targets. PLoS Biol. 2005;2(11):e363. https://doi.org/10.1371/journal.pbio.0020363.
Zhang S, Chen H, Zhao X, Cao J, Tong J, Lu J, et al. REV3L 3′ UTR 460 T> C polymorphism in microRNA target sites contributes to lung cancer susceptibility. Oncogene. 2013;32(2):242–50. https://doi.org/10.1038/onc.2012.32.
Sethupathy P, Collins FS. MicroRNA target site polymorphisms and human disease. Trends Genet. 2008;24(10):489–97. https://doi.org/10.1016/j.tig.2008.07.004.
Xiong F, Wu C, Chang J, Yu D, Xu B, Yuan P, et al. Genetic variation in an miRNA-1827 binding site in MYCL1 alters susceptibility to small-cell lung cancer. Cancer Res. 2011;71(15):5175–81. https://doi.org/10.1158/0008-5472.CAN-10-4407.
Teo MT, Landi D, Taylor CF, Elliott F, Vaslin L, Cox DG, et al. The role of microRNA-binding site polymorphisms in DNA repair genes as risk factors for bladder cancer and breast cancer and their impact on radiotherapy outcomes. Carcinogenesis. 2012;33(3):581–6. https://doi.org/10.1093/carcin/bgr300.
Martorell-Marugan J, Toro-Dominguez D, Alarcon- Riquelme ME, Carmona-Saez P. MetaGenyo: a web tool for meta-analysis of genetic association studies. BMC Bioinformatics. 2017;18:563. https://doi.org/10.1186/s12859-017-1990-4.
Wells G, Shea B, O’connell D, et al. (2000). The Newcastle-Ottawa scale (NOS) for assessing the quality of nonrandomised studies in meta-analyses. http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp.
Yang M, Xiao X, Xing X, Li X, Xia T, Long H. KRAS and VEGF gene 3’-UTR single nucleotide polymorphisms predicted susceptibility in colorectal cancer. PLoS One. 2017;12(3):e0174140. https://doi.org/10.1371/journal.pone.0174140.
Ahmad A, Venizelos N, Hahn-Strömberg V. Prognostic effect of vascular endothelial growth factor +936C/T polymorphism on tumor growth pattern and survival in patients diagnosed with colon carcinoma. J Tumor Res. 2016;2(1):1–6.
Jannuzzi AT, Özhan G, Yanar HT, Alpertunga B. VEGF gene polymorphisms and susceptibility to colorectal cancer. Genet Test Mol Biomarkers. 2015;19(3):133–7. https://doi.org/10.1089/gtmb.2014.0259.
Credidio L, Lima CSP, Leal R, de Ayrizono MLS, Fagundes JJ, Magna LA, et al. C936T polymorphism of the VEGF gene in relation to the risk and the clinical and biological characteristics of sporadic colorectal adenocarcinoma. BMC Res Notes. 2014;7(1):768. https://doi.org/10.1186/1756-0500-7-768.
Lau TP, Roslani AC, Lian LH, Lee PC, Hilmi I, Goh KL, et al. Association between EGF and VEGF functional polymorphisms and sporadic colorectal cancer in the Malaysian population. Genet Mol Res. 2014;13(3):5555–61. https://doi.org/10.4238/2014.July.25.9.
Jang MJ, Jeon YJ, Kim JW, Cho YK, Lee SK, Hwang SG, et al. Association of VEGF and KDR single nucleotide polymorphisms with colorectal cancer susceptibility in Koreans. Mol Carcinog. 2013;52(S1):60–9. https://doi.org/10.1002/mc.21980.
Antonacopoulou AG, Kottorou AE, Dimitrakopoulos FID, Triantafyllia V, Marousi S, Koutras A, et al. VEGF polymorphisms may be associated with susceptibility to colorectal cancer: a case-control study. Cancer Biomark. 2011;10(5):213–7. https://doi.org/10.3233/CBM-2012-0249.
Wu X, Li D, Liu Z, Wan X, Wu Y, Jiang C, et al. Vascular endothelial growth factor 1498C/T, 936C/T polymorphisms associated with increased risk of colorectal adenoma: a Chinese case–control study. Mol Biol Rep. 2011;38(3):1949–55. https://doi.org/10.1007/s11033-010-0316-7.
Wu G, Hasenberg T, Magdeburg R, Bönninghoff R, Sturm JW, Keese M. Association between EGF, TGF-β1, VEGF gene polymorphism and colorectal cancer. World J Surg. 2009;33(1):124–9. https://doi.org/10.1007/s00268-008-9784-5.
Ungerbäck J, Elander N, Dimberg J, Söderkvist P. Analysis of VEGF polymorphisms, tumor expression of VEGF mRNA and colorectal cancer susceptibility in a Swedish population. Mol Med Rep. 2009;2(3):435–9. https://doi.org/10.3892/mmr_00000118.
Bae SJ, Kim JW, Kang H, Hwang SG, Oh D, Kim NK. Gender-specific association between polymorphism of vascular endothelial growth factor (VEGF 936 C> T) gene and colon cancer in Korea. Anticancer Res. 2008;28(2B):1271–6 PMID: 18505065.
Chae YS, Kim JG, Sohn SK, Cho YY, Ahn BM, Moon JH, et al. Association of vascular endothelial growth factor gene polymorphisms with susceptibility and clinicopathologic characteristics of colorectal cancer. J Korean Med Sci. 2008;23(3):421–7. https://doi.org/10.3346/jkms.2008.23.3.421.
Hofmann G, Langsenlehner U, Renner W, Langsenlehner T, Yazdani-Biuki B, Clar H, et al. Common single nucleotide polymorphisms in the vascular endothelial growth factor gene and colorectal cancer risk. J Cancer Res Clin Oncol. 2008;134(5):591–5. https://doi.org/10.1007/s00432-007-0322-x.
Ferrara N, Gerber H, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9:669–76. https://doi.org/10.1038/nm0603-669.
Hein A, Lambrechts D, von Minckwitz G, HaÈberle L, Eidtmann H, Tesch H, et al. Genetic variants in VEGF pathway genes in neoadjuvant breast cancer patients receiving bevacizumab: results from the randomized phase III Gepar Quinto study. Int J Cancer. 2015;137(12):2981–8. https://doi.org/10.1002/ijc.29656.
Della-Morte D, Riondino S, Ferroni P, Palmirotta R, Pastore D, Lauro D, et al. Impact of VEGF gene polymorphisms in elderly cancer patients: clinical outcome and toxicity. Pharmacogenomics. 2015;16(1):61–78. https://doi.org/10.2217/pgs.14.136.
Chen N, Ma CN, Zhao M, Zhang YJ. Role of VEGF gene polymorphisms in the clinical outcome of non-small cell lung cancer. Genet Mol Res. 2015;14(4):16006–11. https://doi.org/10.4238/2015.December.7.13.
Mungunsukh O, Day RM. Transforming growth factor-β1 selectively inhibits hepatocyte growth factor expression via a micro-RNA-199–dependent posttranscriptional mechanism. Mol Biol Cell. 2013;24(13):2088–97. https://doi.org/10.1091/mbc.E13-01-0017.
Ganji PN, Park W, Wen J, Mahaseth H, Landry J, Farris AB, et al. Antiangiogenic effects of ganetespib in colorectal cancer mediated through inhibition of HIF-1α and STAT-3. Angiogenesis. 2013;16(4):903–17. https://doi.org/10.1007/s10456-013-9364-7.
Greener MJ, Sewry CA, Muntoni F, Roberts RG. The 3′-untranslated region of the dystrophin gene–conservation and consequences of loss. Eur J Hum Genet. 2002;10(7):413–20. https://doi.org/10.1038/sj.ejhg.5200822.
Iida K, Kawakami Y, Sone H, Suzuki H, Yatoh S, Isobe K, et al. Vascular endothelial growth factor gene expression in a retinal pigmented cell is up-regulated by glucose deprivation through 3′ UTR. Life Sci. 2002;71(14):1607–14. https://doi.org/10.1016/s0024-3205(02)01842-8.
Ma WJ, Cheng S, Campbell C, Wright A, Furneaux H. Cloning and characterization of HuR, a ubiquitously expressed Elav-like protein. J Biol Chem. 1996;271(14):8144–51. https://doi.org/10.1074/jbc.271.14.8144.
Ramamoorti N, Kumarvelu J, Shanmugasundaram GK, Rani K, Banerjea AC. High frequency of G to a transition mutation in the stromal cell derived factor-1 gene in India, a chemokine that blocks HIV-1 (X4) infection: multiple proteins bind to 3′-untranslated region of SDF-1 RNA. Genes Immun. 2001;2(7):408–10. https://doi.org/10.1038/sj.gene.6363800.
Kim JG, Chae YS, Sohn SK, Cho YY, Moon JH, Park JY, et al. Vascular endothelial growth factor gene polymorphisms associated with prognosis for patients with colorectal cancer. Clin Cancer Res. 2008;14(1):62–6. https://doi.org/10.1158/1078-0432.CCR-07-1537.
Krippl P, Langsenlehner U, Renner W, Yazdani Biuki B, Wolf G, Wascher TC. A common 936 C/T gene polymorphism of vascular endothelial growth factor is associated with decreased breast cancer risk. Int J Cancer. 2003;106(4):468–71. https://doi.org/10.1002/ijc.11238.
Zhang J, Yang J, Chen Y, Mao Q, Li S, Xiong W, et al. Genetic variants of VEGF (rs201963 and + 936C/T) and KDR (rs7667298, rs2305948, and rs1870377) are associated with glioma risk in a Han Chinese population: a case-control study. Mol Neurobiol. 2016;53(4):2610–8. https://doi.org/10.1007/s12035-015-9240-0.
Janardhan B, Vaderhobli S, Bhagat R, Chennagiri Srinivasamurthy P, Venketeshiah Reddihalli P, Gawari R, et al. Investigating impact of vascular endothelial growth factor polymorphisms in epithelial ovarian cancers: a study in the Indian population. PLoS One. 2015;10(7):e0131190. https://doi.org/10.1371/journal.pone.0131190.
Dassoulas K, Gazouli M, Rizos S, Theodoropoulos G, Christoni Z, Nikiteas N, et al. Common polymorphisms in the vascular endothelial growth factor gene and colorectal cancer development, prognosis, and survival. Mol Carcinog. 2009;48(6):563–9. https://doi.org/10.1002/mc.20495.
Jang MJ, Kim JW, Jeon YJ, Chong SY, Oh D, Kim NK, et al. Prognostic significance of vascular endothelial growth factor gene polymorphisms in patients with colorectal cancer. Int J Clin Oncol. 2013;18:1032–41. https://doi.org/10.1007/s10147-012-0493-6.
Jeon YJ, Kim JW, Park HM, Jang HG, Kim JO, Oh J, et al. Interplay between 3’-UTR polymorphisms in the vascular endothelial growth factor (VEGF) gene and metabolic syndrome in determining the risk of colorectal cancer in Koreans. BMC Cancer. 2014;14(1):881. https://doi.org/10.1186/1471-2407-14-881.
Renner W, Kotschan S, Hoffmann C, Barbara OP, Ernst P. A common 936 C/T mutation in the gene for vascular endothelial growth factor is associated with vascular endothelial growth factor plasma levels. J Vasc Res. 2000;37(6):443–8. https://doi.org/10.1159/000054076.
Hu YF, Luscher B, Admon A, Mermod N, Tjian R. Transcription factor AP-4 contains multiple dimerization domains that regulate dimer specificity. Genes Dev. 1990;4(10):1741–52. https://doi.org/10.1101/gad.4.10.1741.
Mermod N, Williams T, Tjian R. Enhancer binding factors AP-4 and AP-1 act in concert to activate SV40 late transcription in vitro. Nature. 1998;332:557–61. https://doi.org/10.1038/332557a0.
Jakubowska A, Gronwald J, Menkiszak J, Go’rski B, Huzarski T, Byrski T, et al. The VEGF_936_C>T 3'UTR polymorphism reduces BRCA1-associated breast cancer risk in Polish women. Cancer Lett. 2008;262(1):71–6. https://doi.org/10.1016/j.canlet.2007.11.029.
Lee SJ, Lee SY, Jeon HS, Park SH, Jang JS, Lee GY, et al. Vascular endothelial growth factor gene polymorphisms and risk of primary lung cancer. Cancer Epidemiol Biomarkers Prev. 2005;14(3):571–5. https://doi.org/10.1158/1055-9965.EPI-04-0472.
Zhao Z, Ba C, Wang W, Wang X, Xue R, Wu X. Vascular endothelial growth factor (VEGF) gene polymorphisms and colorectal cancer: a meta-analysis of epidemiologic studies. Genet Test Mol Biomarkers. 2012;16(12):1390–4. https://doi.org/10.1089/gtmb.2012.0266.
Gholami M, Larijani B, Sharifi F, Hasani-Ranjbar S, Taslimi R, Bastami M, et al. MicroRNA-binding site polymorphisms and risk of colorectal cancer: a systematic review and meta-analysis. Cancer Med. 2019;8(17):7477–99. https://doi.org/10.1002/cam4.2600.
Acknowledgements
We are immensely grateful to the management of Sri Ramachandra Institute of Higher Education (deemed to be University) for supporting our study through the Founder Chancellor Research Fellowship for SSK, ZSS, and NK.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
All authors have read and approved the manuscript. CEJW and TJ conceived the study and designed the experiments. SSK and ZSS conducted and performed the meta-analysis. AR analyzed the data. SSK drafted the manuscript, and NK gave suggestions for the manuscript. CEJW and TJ reviewed the results and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kontham, S.S., Walter, C.E.J., Shankaran, Z.S. et al. A microRNA binding site polymorphism in the 3′ UTR region of VEGF-A gene modifies colorectal cancer risk based on ethnicity: a meta-analysis. J Egypt Natl Canc Inst 34, 18 (2022). https://doi.org/10.1186/s43046-022-00118-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43046-022-00118-3