
Abstract
The dynamics of the microbial community and functional taxa related to nitrogen (N) removal biogeochemical processes can be important to the development of new cost-effective processes in wastewater treatment. This work consisted of the start-up of an upflow reactor for N-removal by partial nitritation/anammox pathway, working at ambient temperature, during 397 d. After an adaptation to the reactor operational conditions, a stable total N-removal (52% efficiency) was linked to ammonium deletion. High-throughput sequencing of 16S rRNA gene amplicons analysis revealed a relative abundance of about 1% of anammox genus Candidatus Brocadia after 397 d. Nitrosomonas, a nitrifying bacterium also increased the relative abundance, together with the accretion of relative numbers of Denitratisoma and Thiobacillus, recognized as heterotrophic and chemolithoautotrophic denitrifying bacteria, respectively. These findings provide a better understanding of the N-removal by key microbial groups that may be useful to optimize future field application of systems working at ambient temperature.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.1 Introduction
Over the last century, humans have substantially influenced the global nitrogen (N) cycle by increasing both the availability and the mobility of N compounds leading to undesirable noxious effects in the aquatic ecosystems [1]. A particular importance has been given by the European Union (Directive 91/271/EEC) for reducing the discharge of N-containing compounds to the environment.
The removal of N in wastewater treatment plants (WWTPs) is mostly carried out by multi-step microbial processes, and the present technology needs to be upgraded to efficiently remove N from effluents, which is considered crucial for the water environment protection [2]. Conventional biological processes for N-removal involve the autotrophic nitrification from ammonium (NH4+) to nitrate (NO3-), followed by heterotrophic denitrification from NO3- to dinitrogen (N2) gas [3]. This approach is rather costly since it requires aeration for nitrification process, and an external carbon source as an electron donor supply for denitrification, when wastewater has a low organic carbon (C) and N (C:N) ratio [3]. Additionally, these processes carry an associated environmental cost - the production of CO2, CH4 and N2O, important greenhouse gases that contribute to global warming [4].
Anammox process was discovered 26 years ago [5], being a biological process that converts NH4+ to N2 gas with nitrite (NO2-) as an electron acceptor under anoxic conditions [5, 6]. New autotrophic N-removal technologies based on the metabolism of anammox bacteria are considered as a promising cost-saving biological process, which tend to achieve satisfactory N-removal performances from wastewater [4]. The anaerobic and autotrophic nature of this process allows the total reduction of aeration, and simultaneously reduces the energy requirements, eliminating the need of an external organic carbon. Additionally, anammox-based technologies have lower greenhouse gases production [4] and, therefore, reduced environmental impacts. One of the main difficulties that researchers deal when studying the anammox process is the slow growth rate of anammox bacteria, leading to long start-up periods, which remains one of the main obstacles to the widespread anammox application [7]. For proficient N-removal in wastewater, anammox bioreactors generally require additional heating to achieve optimal temperature values (27 to 40 °C) for most anammox species [7], which lowers its energy efficiency. Yet, the presence of anammox bacteria is consistently reported in natural environments with a wide temperature range, including extreme environments, with temperatures as high as 80 °C (e.g., in hot springs), and as low as -5 °C in river or marine sediments [8, 9]. The application of anammox pathway in N-removal bioreactors working at ambient temperature has not been properly explored, and could represent an advantage by improving the energy efficiency in WWTPs.
The operational conditions and the type of reactor can influence the N-removal efficiency [3]. The upflow reactor is one the most used reactors in the last twenty years, since it is a high rate reactor with heterogeneous distribution of substrate [10]. So far, the design and operation of partial nitritation/anammox (PN/A) systems for N-removal were predominantly focused in an engineer perspective, overlooking the ecology and dynamics of microbial communities as a crucial component of these systems. In fact, few studies [11, 12] investigate the interactions among the microbial community composition and structure, as well as the dynamics and biogeochemistry. The microbial community structure in anammox-based reactors is usually characterized by a stable foothold of Planctomycetes phylum, with a noteworthy contribution of Proteobacteria, Chlorobi, Chloroflexi and Bacteroidetes phyla [13]. High-throughput sequencing can provide detailed information about microbial community structures, offering a better understanding of the key microbial groups responsible for the N removal and their interactions. This knowledge could be used to address some limitations in PN/A-based systems, particularly to optimize the configurations and operational conditions to foster key microbial groups, and therefore, reduce the long start-up period. It could also be used to reduce the working temperature in wastewater reactors maintaining similar N-removal proficiency.
This work aims to address the microbial community composition, shifts, and biogeochemistry, as well as to understand how these factors might affect the N-removal performance during the start-up of an upflow reactor at ambient temperature. We hypothesize that the operational conditions used may select/establish the microbial community structure, and the autotrophic functional taxa, particularly nitrifying and anammox bacteria.
2 Materials and methods
2.1 The upflow reactor configuration
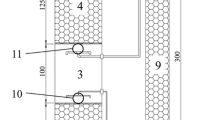
The experimental work was conducted in a poly (methyl methacrylate) acrylic lab-scale upflow reactor with a net working volume of 6.9 L, a diameter of 15 cm, and a height of 50 cm (Fig. 1). A synthetic medium (see below) was fed into the bottom of the reactor by a diaphragm metering pump (SIMDOS 02, KNF, Sursee, Switzerland), which was constantly mixed (100 rpm). All tubes and connectors were made of butyl rubber or polyvinylchloride to prevent air permeability.

Experimental set-up of the laboratorial upflow reactor system
Considering that anammox bacteria has an estimated doubling time of 2-79 d [7], and to retain the sludge seed, and later the anammox biomass in the system, the flowrate of the medium influent was set at 0.1 mL min-1, resulting in a sludge retention time approximately of 48 d during the start-up period. The influent pH was 7.7 ± 0.1, and the reactor functioned at ambient temperature, ranging 14-23 °C. To improve biomass retention and biofilm formation, 13 polyethylene floating bio-balls were placed inside the reactor, occupying approximately 25% of the volume. The reactor was covered with aluminum foil to prevent penetration of light, and to avoid the growth of phototrophic organisms.
2.2 Seed sludge and synthetic medium
The reactor was seeded with 600 mL of wet sludge retrieved from the aeration tank unit from an urban WWTP. The synthetic nutrient medium, used for the anammox bacteria enrichment was modified from van de Graaf et al. [14], and detailed in Table S1. The influent tank and the medium were autoclaved to prevent that any biological residues could alter the medium composition before entering in the bioreactor. To attain hypoxic conditions, the influent medium was always purged with N2 gas for 25 min before the influent bottle was replaced, and at this time the reactor was also purged with N2 gas during the same time.
2.3 Nitrite start-up operational strategy
The NO2- concentration is considered crucial for anammox bacterial growth during the start-up of the reactor, and even modest amounts lead to substrate limitation and slower start-up [15]. To address this limitation, during the first 96 d of this study, we adopted a strategy that included the preparation of the medium with NO2- to promote anammox bacteria. Therefore, during the initial period, the reactor was fed with an influent containing 140 mg N L-1 in the forms of NH4+ and NO2- at a 1:1 ratio. Afterwards, the medium was void of NO2-, expecting that in a PN/A system ammonium oxidizing bacteria (AOB) would produce the NO2- as a result of the first step of nitrification.
2.4 Ammonium, nitrite and nitrate analytical measurement
The NH4+ and NO2- concentrations in effluent samples (retrieved in triplicate with intervals of 10-13 d) were determined colorimetrically using methods described in Grasshoff et al. [16]. The NO3- + NO2- concentrations were determined in the same samples using the spongy cadmium reduction technique [17]. All analyses were performed in triplicate with standard curves generated for each batch. The detection limit was 0.14, 0.03 and 0.42 μg L-1 respectively for NH4+-N, NO2--N, NO3--N, and the precision of determinations was 0.1-8%, depending on the particular nutrient concentration.
2.5 Nitrogen removal efficiencies
The N-removal performance of the reactor was evaluated by the NH4+-N removal efficiency (NH4+RE) and total N-removal efficiency (TNRE) accordingly the following equation:
where Nin and Nout were the inorganic nitrogen (in the form of NH4+-N or total N) concentrations (mg L-1) of the influents and effluents, respectively.
2.6 High-throughput sequencing and bioinformatics analysis
Microbial communities from the initial sludge seed and day 397 samples from the reactor were characterized by next-generation sequencing (NGS). Samples were prepared for Illumina Sequencing by 16S rDNA gene amplification of the prokaryotic community by extracting genomic DNA using PowerDNA Kit for Soil, and purifying using an UltraClean DNA Purification Kit (MO BIO). The genomic DNA were sequenced at Genoinseq facilities (Cantanhede, Portugal).
FASTA files from the merged reads received from to Genoinseq were uploaded and processed for downstream analysis by the NGS analysis pipeline of the SILVA rRNA gene database project (SILVAngs 1.3). Microbial composition of each sample, at different taxonomic levels, was determinate using the pipeline default setting (details can be found in Supplementary Materials), and noticeable shifts (an order of magnitude and/or more than 0.5%) were scrutinized.
2.7 Statistical and data analysis
MS Excel 2016 was used for statistical information, data analysis, and plotting. The NH4+-N, NO2--N, and NO3--N concentrations mean and standard deviations values were calculated. Significant (P < 0.05; P < 0.01; P < 0.001) correlation factors were analysed by correlation matrices. Statistical tests were performed using the commercial software STATISTICA, Version 7, StatSoft (2004).
3 Results
3.1 Nitrogen removal performance during the start-up stage
The variation of N in the form of NH4+, and NO2- concentrations in the influent as well as the NH4+, NO2-, and NO3- concentration profile in the effluent during the 397 d of the start-up period are illustrated in Fig. 2a. The sum of the inorganic N forms in the effluent is also represented in Fig. 2a, whereas the N removal performance, particularly the N in the form of NH4+ removal efficiency (NH4+RE), and TNRE are represented in Fig. 2b. Based in the obtained profiles of N forms, the experimental start-up period of the reactor could be divided into three different stages (Fig. 2): initial (days 0-117), transitional (from day 118-270), and stable (from day 271 onwards).

Reactor performance during the different stages of the start-up, a) Profiles of nitrogen (N) in the form of ammonium (NH4+-N), nitrite (NO2--N) in the influent, and NH4+-N, NO2--N, nitrate (NO3--N), and the sum of N forms (Total N) in the effluent. b) Profile of the removal efficiency of N in the form of ammonia, and total nitrogen
The initial stage was mainly characterized by the “nitrite start-up operational strategy” described in the methodology. This stage was also characterized by high fluctuations in the concentration of N forms. The initial start-up operating phase yielded a production of NO3-, and concomitantly a consumption of NO2- (Fig. 2a). In fact, a negative and significant correlation (r = -0.97; P < 0.001; n = 15) between NO2- and NO3-, and between NH4+ and NO3- (r = -0.77; P < 0.001; n = 15) was observed (Table S2), meaning that NO3- generation appeared to be related with NH4+ and NO2- consumption, pointing to nitrification as the responsible process. Noticeably, after day 47, NO2- was always found at very low concentrations, despite the high input. Also, during the initial stage, at the day 19 the highest NH4+RE during the start-up period (Fig. 2b) was verified reaching 80% with a TNRE of 43%. NH4+RE ranged between 55 and 65% of efficiencies from the day 61 to 117, while TNRE had more fluctuations between 19% at day 61 to 29% at day 96 (Fig. 2b). Noteworthy, during this first stage a positive and significant correlation (r = 0.91; P < 0.001; n = 15) between NH4+ and TN was found in the effluent, and concomitantly a negative correlation in the same magnitude with the NH4+RE suggesting that TN in the effluent was related with transformation of NH4+ in the reactor.
Ending the “nitrite start-up operational strategy”, a clear shift was detected in the profile of N concentrations and removal efficiencies, which paved the way to the transitional stage (from the day 118 to 270) of the start-up period of the reactor (Fig. 2b). Very low nitrite concentrations, frequently below the detection limit, were observed during this second stage. An evident decline of NO3- concentration in the effluent between day 118 and 152 occurred, with a slower decreasing trend towards the end of the experiment (Fig. 2a). A similar trend was observed for the TN concentration in the effluent. Moreover, during this second stage, a positive and significant correlation (r = 0.98; P < 0.001; n = 18) was observed between NO3- and TN in the effluent (Table S2), suggesting that TN removal was related with NO3- transformation pointing to denitrification as the likely responsible process. The NH4+RE showed minor fluctuations ranging between 46 and 62% during the transitional stage. The halt in NO2- addition resulted in an abrupt decline in the TNRE. The values stabilized from day 145 to 190, to increase until the end of the transitional stage, reaching a 36% of efficiency.
On the last stage (from day 271 to 397), the profiles of N forms in the reactor showed a trend to stabilize, being characterized by minor fluctuations (Fig. 2). The NO3- in the effluent gradually decreased at a low rate until day 397, while NO2- remained undetected. This stage was also characterized by a fitful increase of NH4+ and total N in the effluent during the day 271 to 335, and concomitantly a decrease in the NH4+RE and TNRE (Fig. 2). Yet, as from day 335, NH4+-N and total N in the effluent tended to gradually decrease, as removal efficiencies of NH4+RE and TNRE increased, reaching, respectively 54 and 52%. In this last stage, a strong positive correlation (r = 0.8; P < 0.001; n = 23) was observed between NH4+ and TN in the effluent (Table S2). This might indicate that TN removal was related with the fate of NH4+, with anammox as the probable responsible process.
3.2 Diversity and structure of the microbial community
The microbial community structure analysis by the Illumina Miseq platform generated ~147,000 raw reads of the 16S rRNA gene (V4-V5) amplicons with an average length of 404 bp for each of the two samples (initial and day 397) after removing low-quality sequences and trimming the adapters, barcodes, and primers. Of these, ~131,000 reads were merged, and 28,500 rejected at the quality filtering step and chimera removal at the SILVA rRNA gene database project pipeline. A total of 3,534 good-quality reads were classified as ‘No relative’ reads (i.e., without any close relatives) in the data base, and 100,541 reads were taxonomically classified as Bacteria, Archaea, as well as Eukaryota.
The classified sequences were also used for alpha diversity metrics determination (Table 1). All samples had a Good’s coverage (i.e., a measure of coverage of dominant OTUs with more than one sequence) of ~0.99. However, the OTU-level rarefaction curve of samples did not reach a plateau (Fig. S1). The richness in the reactor had a slight shift during the start-up period, revealed by the decreasing trends of 8% of detected OTUs, and of 6% of estimated Chao1. The results of Shannon diversity index also demonstrated that the microbial diversity in the reactor had a minor decreasing trend (3%) over the period of the start-up, although the absolute values were rather high.
A close examination of the relative abundance at the Phylum level showed a relatively stable archaeal community structure, whereas the bacterial community structure shifted distinctly (Fig. 3). It was also noticeable differences at the lower taxonomic level in the microbial community structure of the sludge seed and reactor samples (Fig. 4).

Percentage of the relative abundant of major bacterial phyla, Archaea, and Eukaryotic organisms in the sludge seed and in the upflow reactor

Relative abundance percentage at the lower taxonomic level of the most abundant taxa, and with major noticeable shifts (an order of magnitude and/or more than 0.5%), in the sludge seed (day 0) and after the start-up period (day 397). Colorimetric scale (below) represents increasing percentages of the relative abundance of sequences
3.3 Microbial community composition and major shifts
At higher taxonomic levels, the sludge seed was dominated by the Phylum Proteobacteria (~35%), followed by Bacteroidetes (~18%), Chlorobi (~8%), Chloroflexi (~7%), Planctomycetes (~7%), Acidobacteria (~5%), Actinobacteria (~4%), Verrucomicrobia (~4%), and Armatimonadetes (~3%) (Fig. 3). After the start-up of the reactor, a clear shift in the structure of the microbial community occurred, detected even at the higher taxonomic level. For instance, Chlorobi (~16%) increased 103% in comparison to the relative abundance found in the sludge seed. Increased abundances in Chloroflexi (↑73%), in Cyanobacteria (↑124%), and in Nitrospirae (↑256%) were also observed. On the other hand, other bacterial phyla had their relative abundance decreased, such as Acidobacteria (↓47%), Armatimonadetes (↓76%), and Proteobacteria (↓29.7%), in comparison to the relative abundance found in the sludge seed.
A more detailed bacterial relative abundance at the lower taxonomic level (Phylum, Order, Family, and Genus) is summarized in Fig. 4. For this, assigned OTUs with major shifts (more than 0.5%, and/or an order of magnitude) in the relative abundance in sludge seed and reactor samples were selected. The increase of the relative abundance occurred in the Chlorobi Phylum was noticeable, particularly the genera belonging to the Ignavibacteriales Order. Within the Phylum Chloroflexi, the relative abundance increases of uncultured UTCFX1 bacterium, as well as the SBR1031 was noteworthy. In the Nitrospirae Phylum, an increase of ~1.5% relative abundance of Nitrospira genus could be noted. Although not detected in the sludge seed, genera belonging to the Planctomycetes Phylum and known as anammox bacteria such as Candidatus (Ca.) Jettenia, Ca. Anammoximicrobium, and particularly Ca. Brocadia increased during the start-up period. Despite the overall decrease in abundance verified within Proteobacteria (Fig. 3), some genera showed a relative abundance increase, particularly the Denitratisoma (4.6%), Nitrosomonas (0.7%), and Nitrosospira (1.1%), as well as Thiobacillus (0.8%), all belonging to the Betaproteobacteriales Order.
4 Discussion
This study focused at linking the reactor performance with the likely N cycle biogeochemical processes, and shifts in the microbial community/functional taxa suitable to autotrophic N-removal at ambient temperature. The start-up period of 397 d was divided into three stages based on the fluctuation of the N forms, and the performance of the upflow reactor.
4.1 Initial stage
The N load, particularly the operational strategy of adding NO2- in the mineral influent that fed the reactor, was expected to create niches for autotrophic microorganisms during the start-up period [18]. This study revealed the dynamics of the microbial community towards an increase of N metabolism-related bacteria, including heterotrophs. In fact, two AOB genera, Nitrosomonas and Nitrosospira were enriched in the reactor, in agreement with Chen et al. [19] and Chu et al. [20] studies, which reported the existence of AOB in PN/A systems working, respectively, at room (20–30 °C), and mesophilic (33 °C) temperature. Yet, these authors [19, 20] showed higher AOB abundances comparing to this study. Moreover, Siripong and Rittmann, [21] reported that, at optimal temperature (25-30 °C), the growth rate of Nitrosomonas was higher than that of Nitrosospira, while Nitrosospira was more tolerable to lower temperatures, in agreement with the higher relative abundances of Nitrosospira found in our reactor. Also, the “nitrite start-up operational strategy”, and the experimental conditions during the initial period may also inadvertently selected and enriched the Nitrospira genus. Nitrite oxidizing bacteria (NOB) are usually found in PN/A systems, but at relative low abundances, and tend to decrease with the maturation of the system [19, 22]. The high NH4+RE verified during the initial period and concomitant production of NO3-, pointed to the occurrence of nitrification process. Therefore, even at a lower relative abundance, AOB and NOB were likely involved in the transformation of NH4+ and NO2- into NO3- during the initial period. The extensive NO2- depletion in the effluent might thus be linked to the enrichment of NOB genera, possessing the adequate metabolic machinery (nirK, nirS and nxrAB genes) to metabolize NO2- [23].
During the initial period, the fluctuation in TNRE translated the expected instability of the biological N-removal processes involved. The removal of N from the system seemed to be directly involved with NH4+ removal, since this N form was strongly positively correlated with TN in the effluent. The occurrence of anammox process would simultaneous consume NH4+ and NO2-, however, anammox genera, if present, would likely be outcompeted by the mentioned NOB.
During this stage, TN was negatively correlated with NO3- ruling out the occurrence of other removal process, such as denitrification, which would lead to a decrease in TN. Although the seeding sludge presented a high relative abundance of Rhodanobacter genus, which is able to carry complete denitrification, NGS results showed a decrease of this genus during the start-up period.
4.2 Transition stage
The proposed second stage was characterized by the transitional adaptation of the reactor to zero NO2- input and consequent reduction of TN in the influent. As expected, the TN in the system decreased abruptly during the subsequent 48 d of sludge retention time resulting in negative TNRE. Afterwards, the TN in the effluent stabilized during the following sludge retention time period, and then started to decrease, which might indicate a change of the biogeochemical processes involved in N-removal. One of the most evident results during this stage was the clear NO3- decrease, probably due to the end of nitrite input. This result, associated with minor variation of NH4+, and therefore also in the NH4+RE, suggests that the removal process of NH4+ was not intensely affected by the change in the operational strategy. The very low NO2- concentrations found during this second stage also indicated that NO2- produced by the oxidation of NH4+ by AOB or NO3- reducers would be rapidly metabolized. Formed NO2- was likely metabolized by Nitrospira genus and/or used by anammox bacteria for NH4+ oxidation. Although anammox bacteria were not detected in the seeding sludge, and the microbial composition was not assessed at this transition stage, results showed anammox bacteria enrichment during the start-up period. The use of floating carriers could enable the coexistence between AOB (aerobic) in the biofilm-water interface (outer layer), and anammox bacteria (anaerobic) in the inner layer.
On the other hand, during this stage, the concentration of TN followed the same decreasing trend of NO3-, which was not verified during the initial stage. This feature might indicate denitrification as a plausible process responsible for N-removal from the system during this stage. The presence of denitrifying bacteria in PN/A systems using this synthetic influent was also verified in previous studies [13, 24]. Dissimilatory nitrate reduction to ammonium (DNRA) can also reduce NO3- to NO2-, but is considered a process that conserves N in the system [25], and our results point to the removal of TN.
An enrichment in the Denitratisoma genus, which is classified as heterotrophic denitrifying bacteria, was observed. Indeed, Denitratisoma genus is usually found in high abundance in PN/A reactors working at mesophilic, as well as ambient temperatures [20, 24]. EDTA, an organic carbon compound used in the influent medium (Table S1), as well as the presence of cellular compounds and metabolites released by lysis and decay of organisms might explain the enrichment of other heterotrophic genera in the reactor with NO3- reducing ability (denitrifying and DNRA metabolism), such as Gordonia [26], and Burkholderia-like [27]. Moreover, autotrophic denitrification should also be considered due to the relative abundance increase of known autotrophic bacteria, such as the Thiobacillus genus. This genus is able to grow under both aerobic and anaerobic conditions, oxidizing sulfur compounds as electron donors, and using oxygen, nitrate, and nitrite as electron acceptors [28]. In fact, Dasgupta et al. [29] coupled autotrophic denitrification with PN/A working at room temperature, being Thiobacillus a key player of N removal.
The enrichment of NO3- reducers obtained in this study points to the putative ability for both organotrophic and lithotrophic processes. The presence of these bacteria in PN/A systems could be helpful to the occurrence of a NO2- loop, where NO2- is oxidized to NO3- (by NOB and anammox bacteria), to be reduced back to NO2- by denitrifying bacteria [30]. More studies are needed to screen and confirm the metabolic pathways of these enriched heterotrophic and/or autotrophic denitrifying and DNRA bacteria, which might be important to optimize biological strategies to N-removal from wastewaters in PN/A systems.
4.3 Transition stage
The last proposed stage was characterized by a general stabilization of the N forms profiles, and mainly by the apparent relation between NH4+ and TN removal in the effluent. The highest TNRE was verified during this period, pointing to anammox as the main process responsible for NH4+ removal, and concomitantly TN from the system. This assumption was also supported by the enrichment of genera known as anammox bacteria. Cao et al. [24] also reported 2.3% enrichment in the relative abundance of anammox bacteria, although still higher enrichment comparing to this study. The exerted operational conditions in the reactor enriched three genera known by the anammox ability, Ca. Brocadia, Ca. Jettenia and Ca. Anammoximicrobium. It is important to recognize that the used primers do not target specifically the functional genes of anammox or nitrifying communities, and therefore limits its use in revealing the full bacterial diversity analysis. Yet, these primers can be used to efficiently detect the anammox bacterial community, and for instance it can detect anammox bacteria with relatively small losses in non-halophilic Ca. Brocadia, Ca. Kuenenia, and Ca. Jettenia detection, and with 100% efficiency to detect Ca. Scalindua [31].
The higher relative abundance indicated that the operational conditions, including temperature, were more suitable for Ca. Brocadia than for the other detected genera. Previous studies [22, 24] reported the enrichment and dominance of Ca. Brocadia-like strain in a PN/A system working at low temperature, and therefore, we hypothesize that the enriched Ca. Brocadia temperature range for growth might be lower than the other anammox genera.
In the third stage, NO3- continued to slowly decrease in the effluent with no NO2- accumulation, pointing to partial reduction of NO3- to NO2- likely by heterotrophic bacteria (as previously discussed), and the turnover of NO2- by anammox bacteria for NH4+ oxidation to N2 [5, 6].
Recently a specific clade of Nitrospira genus with the ability to perform the complete ammonia oxidation-comammox [32] was discovered. However, it is not possible to distinguish if the Nitrospira genus assessed by high-throughput sequencing belongs to the comammox clade. Yet, during the third stage, we performed a preliminary assessment of the presence of comammox bacteria in the system by the amplification and sequencing of the amoA gene amplicon. An uncultured Nitrospira sp. clone OTU10 ammonia monooxygenase (amoA) gene with similarity percentage of 92.7% (GenBank: MG387165.1) was detected (data not published). van Kessel et al. [32] suggested that the presence of comammox is compatible with anammox, since the co-occurrence of comammox Nitrospira-like with Ca. Brocadia species was found. This indicates a possible functional link between those two competing organisms. For this reason, more studies are needed to ascertain the real implications of the presence of Nitrospira (comammox) in PN/A systems, and the effect on their performance, towards the enhancement of N-removal efficiency. Also, additional studies are needed to ascertain on the performance and function of Brocadia-like bacteria and AOB, as well as the real implications of the presence of NO3- reducers, such as Thiobacillus genus, as well as Denitratisoma and other heterotrophic genera in a PN/A system.
4.4 Framework of the microbial community dynamics
Proteobacteria, Bacteroidetes, Chlorobi, Chloroflexi, Planctomycetes, Actinobacteria, Acidobacteria, Armatimonadetes, Verrucomicrobia, and Patescibacteria were the most dominant Phyla in the seeding sludge sample. This agrees with other studies [19, 33] that usually describe similar microbial community structures regarding the dominant phyla in sludge from WWTPs.
Proteobacteria, Chlorobi, Chloroflexi, and Bacteroidetes were the dominating Phyla in the reactor after the start-up period, and established a stable foothold in the community, with Planctomycetes Phylum also showing a noteworthy contribution for the microbial assemblage. These microbial groups have been previously reported as the most abundant Phyla in anammox-based reactors [13], working mainly at mesophilic conditions (31–37 °C).
The operational conditions, purging with N2, and particularly the use of a low organic carbon influent, may fostered a negative effect on chemoorganotrophs and on most species classified as aerobic. OTUs, such as the Ca. Competibacter (Proteobacteria Phylum), known to be enriched in phosphorus removal wastewater treatment systems were likely constrained by the different conditions in the reactor [34]. In the same way, the huge decrease in the relative abundance of the uncultured bacterium SJA-28, may be linked to its specific catabolic activity, using acetate, propionate, and/or hydrocarbons as carbon source [34].
The results in this study also disclosed the enrichment of several other OTUs with different metabolic activities. For instance, the Chloroflexi Phylum is frequently found in mesophilic PN/A systems, but can also be found at room and ambient temperatures [13, 35]. The organisms belonging to the Chloroflexi Phylum (e.g., Order Anaerolineales and SBR1031) are considered to be important in the formation, support, and structural organization in the biofilm of the anammox bacterial aggregate [13]. Bacteria belonging to the Chlorobi Phylum (also known as green sulfur bacteria) may compete for cellular compounds and metabolites in PN/A systems [35]. Other closely related taxa, such as Ignavibacterium genus (Ignavibacteriaceae Family), and the uncultured bacterium PHOS-HE36 are recognized to be obligate heterotrophs and facultative anaerobic, and likely have the same metabolic lifestyle using alternative carbon sources [36]. Sedimenticola-like organisms, belonging to the Proteobacteria Phylum, were another group enriched. These bacteria are described as being able to use organic carbon as the sole carbon, and being associated to sulfur oxidation [37]. Additionally, members of the uncultured bacterium OPB56 are also likely heterotrophs that metabolize small organic molecules [36]. To our knowledge, the role of these closely related sulfur-oxidizing microorganisms in PN/A systems has not yet been accessed.
Other microorganisms were enriched during the start-up, for instance the Bryobacter genus (described as strictly aerobic chemo-organotrophic bacteria) that has been reported in a nitrifying biofilm at low (8 °C) temperature [38]. Shu et al. [39] also reported that Bryobacter play a part in the hub of the major microbial assembly in anammox bioreactors. Similar metabolic lifestyles could have favoured Chthonomonas genus belonging to the Armatimonadetes Phylum, the uncultured bacterium belonging to the Microscillaceae Family (Bacteroidetes Phylum), and Luteibacter genus (Xanthomonadales order).
Methanotrophic bacteria belonging to the Methylacidiphilaceae Family within the Phylum Verrucomicrobia were also enriched. These bacteria are considered obligate aerobic capable to growth on methane [40], which might be produced by the anaerobic digestion of decaying biomass [41]. Denitrifying anaerobic methane oxidation coupled with anammox might be relevant in the carbon and nitrogen removal in wastewater treatment plants [42]. Myxobacteria of the genus Sorangium belonging to the Proteobacteria were also increased. To our knowledge, the enrichment of methanotrophic and myxobacteria in PN/A systems was not previously reported, therefore, the underlay metabolism of these organisms during the start-up period remains concealed.
Members affiliated to the candidate phyla Peregrinibacteria, Bacterial Rice Cluster 1, and Wurtsmith Sequence 2 were also enriched. To the best of our knowledge, no study reported this enrichment in PN/A systems, and in fact limited information exists regarding these Candidate phyla. The lack of complete and closed genomes from these groups limits the assessment to the metabolic potential, and therefore limited our considerations about its function in the microbial assembly during the start-up period.
The significance of OTUs not related with the N cycle processes, as well as their role and function within the microbial assemble should be assessed in future studies in order to understand their importance in N-removal processes.
5 Conclusions
This study showed an enrichment of Ca. Brocadia-like bacteria, as well as other flanking key bacterial genera involved in the N cycle, such as AOB, NOB, and heterotrophic and chemolithoautotrophic denitrifying bacteria, together with a positive selection of bacteria from Phyla unrelated with the N cycle processes. Our findings points to Ca. Brocadia-like as a suitable anammox bacteria to implement a PN/A pathway at ambient temperature. Therefore, future scientific efforts should focus on optimizing strategies to enhance the enrichment of Brocadia-like, as well as the flanking bacteria to improve the practical application of the N-removal systems working at ambient temperature.
Availability of data and materials
All data generated during this study are available from the corresponding author (catarina@icbas.up.pt) upon request.
References
Camargo JA, Alonso A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment. Environ Int. 2006;32:831–49.
Carey RO, Migliaccio KW. Contribution of wastewater treatment plant effluents to nutrient dynamics in aquatic systems: a Review. Environ Manage. 2009;44:205–17.
Beylier MR, Balaguer MD, Colprim J, Pellicer-Nacher C, Ni BJ, Smets BF, et al. Biological nitrogen removal from domestic wastewater. In: Moo-Young M, editor. Comprehensive biotechnology. 2nd Amsterdam: Elsevier; 2011. p. 329–40.
Ren Y, Ngo HH, Guo WS, Wang DB, Peng L, Ni BJ, et al. New perspectives on microbial communities and biological nitrogen removal processes in wastewater treatment systems. Bioresour Technol. 2020;297:122491.
Mulder A, van de Graaf AA, Robertson LA, Kuenen JG. Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor. FEMS Microbiol Ecol. 1995;16:177–83.
van de Graaf AA, Mulder A, Debruijn P, Jetten MSM, Robertson LA, Kuenen JG. Anaerobic oxidation of ammonium is a biologically mediated process. Appl Environ Microb. 1995;61:1246–51.
Tomaszewski M, Cema G, Ziembinska-Buczynska A. Influence of temperature and pH on the anammox process: a review and meta-analysis. Chemosphere. 2017;182:203–14.
Jaeschke A, Op den Camp HJ, Harhangi H, Klimiuk A, Hopmans EC, Jetten MSM, et al. 16S rRNA gene and lipid biomarker evidence for anaerobic ammonium-oxidizing bacteria (anammox) in California and Nevada hot springs. FEMS Microbiol Ecol. 2009;67:343–50.
Zhu GB, Xia C, Wang SY, Zhou LL, Liu L, Zhao SY. Occurrence, activity and contribution of anammox in some freshwater extreme environments. Env Microbiol Rep. 2015;7:961–9.
Daud MK, Rizvi H, Akram MF, Ali S, Rizwan M, Nafees M, et al. Review of upflow anaerobic sludge blanket reactor technology: effect of different parameters and developments for domestic wastewater treatment. J Chem-NY. 2018;2018:1596319.
Yang YC, Pan J, Zhou ZC, Wu JP, Liu Y, Lin JG, et al. Complex microbial nitrogen-cycling networks in three distinct anammox-inoculated wastewater treatment systems. Water Res. 2020;168:115142.
Park M, Kim JM, Lee T, Oh YK, Nguyen V, Cho S. Correlation of microbial community with salinity and nitrogen removal in an anammox-based denitrification system. Chemosphere. 2021;263:128340.
Pereira AD, Cabezas A, Etchebehere C, Chernicharo CADL, de Araujo JC. Microbial communities in anammox reactors: a review. Environ Technol Rev. 2017;6:74–93.
van de Graaf AA, de Bruijn P, Robertson LA, Jetten MSM, Kuenen JG. Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor. Microbiol-UK. 1996;142:2187–96.
van der Star WRL, Abma WR, Blommers D, Mulder JW, Tokutomi T, Strous M, et al. Startup of reactors for anoxic ammonium oxidation: experiences from the first full-scale anammox reactor in Rotterdam. Water Res. 2007;41:4149–63.
Grasshoff K, Kremling K, Ehrhardt M. Methods of seawater analysis. 3rd Weinheim: Wiley-VCH; 1999.
Jones MN. Nitrate reduction by shaking with cadmium: alternative to cadmium columns. Water Res. 1984;18:643–6.
Huang Q, Du WL, Miao LL, Liu Y, Liu ZP. Microbial community dynamics in an ANAMMOX reactor for piggery wastewater treatment with startup, raising nitrogen load, and stable performance. AMB Express. 2018;8:156.
Chen R, Yao JQ, Ailijiang N, Liu RS, Fang L, Chen YG. Abundance and diversity of nitrogen-removing microorganisms in the UASB-anammox reactor. PLoS One. 2019;14 e0215615.
Chu ZR, Wang K, Li XK, Zhu MT, Yang L, Zhang J. Microbial characterization of aggregates within a one-stage nitritation-anammox system using high-throughput amplicon sequencing. Chem Eng J. 2015;262:41–8.
Siripong S, Rittmann BE. Diversity study of nitrifying bacteria in full-scale municipal wastewater treatment plants. Water Res. 2007;41:1110–20.
Persson F, Sultana R, Suarez M, Hermansson M, Plaza E, Wilen BM. Structure and composition of biofilm communities in a moving bed biofilm reactor for nitritation-anammox at low temperatures. Bioresour Technol. 2014;154:267–73.
Daims H, Lucker S, Wagner M. A new perspective on microbes formerly known as nitrite-oxidizing bacteria. Trends Microbiol. 2016;24:699–712.
Cao SB, Du R, Li BK, Ren NQ, Peng YZ. High-throughput profiling of microbial community structures in an ANAMMOX-UASB reactor treating high-strength wastewater. Appl Microbiol Biot. 2020;104:6457–67.
Giblin AE, Tobias CR, Song B, Weston N, Banta GT, Rivera-Monroy VH. The importance of dissimilatory nitrate reduction to ammonium (DNRA) in the nitrogen cycle of coastal ecosystems. Oceanography. 2013;26:124–31.
Kim KK, Lee CS, Kroppenstedt RM, Stackebrandt E, Lee ST. Gordonia sihwensis sp. nov., a novel nitrate-reducing bacterium isolated from a wastewater-treatment bioreactor. Int J Syst Evol Micr. 2003;53:1427–33.
Shapleigh JP. Denitrifying prokaryotes. In: Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F, editors. The prokaryotes: prokaryotic physiology and biochemistry. 4th Berlin Heidelberg: Springer; 2013. p. 405–25.
Di Capua F, Ahoranta SH, Papirio S, Lens PNL, Esposito G. Impacts of sulfur source and temperature on sulfur-driven denitrification by pure and mixed cultures of Thiobacillus. Process Biochem. 2016;51:1576–84.
Dasgupta S, Wu S, Goel R. Coupling autotrophic denitrification with partial nitritation-anammox (PNA) for efficient total inorganic nitrogen removal. Bioresour Technol. 2017;243:700–7.
Speth DR, in 't Zandt MH, Guerrero-Cruz S, Dutilh BE, Jetten MSM. Genome-based microbial ecology of anammox granules in a full-scale wastewater treatment system. Nat Commun. 2016;7:11172.
Yang YC, Li M, Li H, Li XY, Lin JG, Denecke M, et al. Specific and effective detection of anammox bacteria using PCR primers targeting the 16S rRNA gene and functional genes. Sci Total Environ. 2020;734:139387.
van Kessel MAHJ, Speth DR, Albertsen M, Nielsen PH, Op den Camp HJ, Kartal B, et al. Complete nitrification by a single microorganism. Nature. 2015;528:555–9.
Zhang L, Shen Z, Fang WK, Gao G. Composition of bacterial communities in municipal wastewater treatment plant. Sci Total Environ. 2019;689:1181–91.
Oyserman BO, Martirano JM, Wipperfurth S, Owen BR, Noguera DR, McMahon KD. Community assembly and ecology of activated sludge under photosynthetic feast-famine conditions. Environ Sci Technol. 2017;51:3165–75.
Lawson CE, Wu S, Bhattacharjee AS, Hamilton JJ, McMahon KD, Goel R, et al. Metabolic network analysis reveals microbial community interactions in anammox granules. Nat Commun. 2017;8:15416.
Hiras J, Wu YW, Eichorst SA, Simmons BA, Singer SW. Refining the phylum Chlorobi by resolving the phylogeny and metabolic potential of the representative of a deeply branching, uncultivated lineage. ISME J. 2016;10:833–45.
Vavourakis CD, Mehrshad M, Balkema C, van Hall R, Andrei AS, Ghai R, et al. Metagenomes and metatranscriptomes shed new light on the microbial-mediated sulfur cycle in a Siberian soda lake. BMC Biol. 2019;17:69.
Young B, Delatolla R, Abujamel T, Kennedy K, Laflamme E, Stintzi A. Rapid start-up of nitrifying MBBRs at low temperatures: nitrification, biofilm response and microbiome analysis. Bioproc Biosyst Eng. 2017;40:731–9.
Shu DT, He YL, Yue H, Yang SC. Effects of Fe (II) on microbial communities, nitrogen transformation pathways and iron cycling in the anammox process: kinetics, quantitative molecular mechanism and metagenomic analysis. RSC Adv. 2016;6:68005–16.
Op den Camp HJ, Islam T, Stott MB, Harhangi HR, Hynes A, Schouten S, et al. Environmental, genomic and taxonomic perspectives on methanotrophic Verrucomicrobia. Env Microbiol Rep. 2009;1:293–306.
Gonzalez-Gil G, Sougrat R, Behzad AR, Lens PNL, Saikaly PE. Microbial community composition and ultrastructure of granules from a full-scale anammox reactor. Microb Ecol. 2015;70:118–31.
van Kessel MAHJ, Stultiens K, Slegers MFW, Cruz SG, Jetten MSM, Kartal B, et al. Current perspectives on the application of N-damo and anammox in wastewater treatment. Curr Opin Biotech. 2018;50:222–7.
Acknowledgements
The authors wish to thank FCT - Foundation for Science and Technology for the support within the scope of UIDB/04423/2020 and UIDP/04423/2020.
Funding
This work was supported by the Project UNNOWN (PTDC/BTA-BTA/31098 /2017), co-financed by COMPETE 2020, Portugal 2020 and the European Union through the ERDF, and by FCT through national funds.
Author information
Authors and Affiliations
Contributions
All authors conceptualized the work. HR analyzed the data; conceptualize and wrote the manuscript. IMWW designed and performed the experiments and nutrient analyses. VSS performed nutrient and molecular biology analyses. AS, ESS, CT, and AAB supervised the work. CT and AAB funded the work. All authors read, revised, and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Supplementary materials.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ribeiro, H., Wijaya, I.M.W., Soares-Santos, V. et al. Microbial community composition, dynamics, and biogeochemistry during the start-up of a partial nitritation-anammox pathway in an upflow reactor. Sustain Environ Res 32, 18 (2022). https://doi.org/10.1186/s42834-022-00130-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42834-022-00130-1