
Abstract
Background
Entomopathogenic nematodes (EPNs) are promising alternatives since they have many characteristics as a biological control agent against insect pests.
Results
Among indigenous EPN isolated, adapted to local environmental conditions by employing the Galleria baiting strategy, only one sample was positive for the presence of EPNs. The new isolate was identified at the species level using DNA sequencing of the internal transcribed spacer region and the Basic Local Alignment Search Tool search of GenBank showed that the isolate had a high similarity (99%) with that sequence available for Heterorhabditis taysearae. The pathogenicity of the EPN isolate was tested against the cotton leaf worm, Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae), using different concentrations (60, 90, 120 and 150 IJs/larvae). Data showed that 150 IJs/larvae caused 100% mortality rate, followed by 120 IJs/larvae (90%), while 60 IJs/larvae showed the lowest rate (60%) after 72 h.
Conclusions
The present study indicated that the native isolate of EPN could be recommended against S. littoralis as an efficient tool in its control programs.
Similar content being viewed by others

Background
Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae) is a common pest in tropical and subtropical areas around the world (Paniagua Voirol et al. 2018). In Egypt, the pest causes economic losses not only to cotton plants but also to 90 different economic crops from 40 different plant families. The rife use of the pesticides to control pests leads to the development of insect resistance, pollution, and biodiversity loss; it also has significant toxicological consequences for humans, plants, livestock (Malbert-Colas et al. 2020). The modern agriculture strategies aim to reduce misuse of chemical pesticides by using biocontrol agents which are environmentally friendly way to combat pests. Many biocontrol agents such as Entomopathogenic nematodes (EPNs) have been used (El-Gaied et al. 2020). Because of their high adaptability to diverse conditions, large variety of insect hosts, superior power at searching hosts, simple mass production, and ability to resist certain chemical pesticides. The majority of EPNs belong to 2 genera Steinernema and Heterorhabditis, which kill the insects by symbiotic bacteria from the genera Xenorhabdus and Photorhabdus, respectively (Shapiro-Ilan and Dolinsksi 2012). In the infection process, the EPNs infective juveniles (IJs) enter the insect through their normal body openings such as mouth, spiracles, and anus (Liu et al. 2020), and some heterorhabditids have dorsal teeth which help them to enter the insect intersegmental membranes (Burnell et al. 2000). From the nematode gut, the symbiotic bacteria are released causing septicemia (Dziedziech et al. 2020) and insects’ death happens within 48 h (Labaude and Griffin 2018). Relative conservative morphological features of nematodes make their morphological classification of the EPN taxonomy difficult (Peat et al. 2009), so sequence data from nuclear and mitochondrial genetic loci, and whole genomes are the most appropriate way to detect and identify the new isolates of indigenous EPNs that adapted to local environmental and climatic conditions. Nuclear genes have been used extensively, including the external non-transcribed spacer (NTS), the small subunit (SSU) or 18S, the internal transcribed spacer (ITS) region that separates rDNA-coding regions (Khashaba and Abdelazim 2021) composed of ITS-1, 5.8S, and ITS-2; and the large subunit (LSU) or 28S (Lulamba and Serepa-Dlamini 2020).
The aim of this study was to isolate and identify a native EPN species and testing its potency against S. littoralis larvae under laboratory conditions.
Methods
Soil sampling and nematode isolation
Ten soil samples 200–250 g were collected from cabbage field at Ismailia Governorate, Egypt at a depth of 15–20 cm and placed in polyethylene bags. All samples information, location, surrounding vegetation and soil type were recorded then samples transferred to.
to the laboratory. According to Bedding and Akhurst (1975), EPNs were isolated by Galleria-trap method. Last instar of G. mellonella larvae was placed into clean plastic boxes containing soil samples, the boxes turned upside down allow up and down free movement of the wax moth larvae inside the plastic boxes and raise their chance to meet nematode infective juveniles (IJs). The boxes were kept in the dark at 25 °C for 1 week. Every 3 days, dead larvae were collected and rinsed with distilled water then transferred to White traps (Kaya and Stock 1997) to extract the emerging infective juveniles (IJs) in sterile deionized water and stored at 16 °C for molecular characterization.
Nematode isolate identification
Total genomic DNA was extracted from freshly collected 2000 IJs of nematodes using the Bio flux DNA extraction Mini Kit (Bioer Technology Co., Ltd.). 1% agarose gel electrophoresis and spectrophotometer were used to determine the quality and quantity of the DNA. According to Joyce et al. (1994), a PCR reaction was used to amplify the Internal transcribed spacer (ITS) of the rDNA region, using specific primers as follows: forward primer TW81 (5-GTTTCC GTA GGT GAA CCT GC-3) and reverse primer AB28 (5-ATA TGC TTA AGT TCA GCG GGT-3), the PCR conditions were as follows: denaturation at 94 °C for 5 min, followed by 35 cycles of 94 °C for 1 min, 55 °C for 1 min 30 s, and 72 °C for 2 min, then a final extension for 7 min at 72 °C. The products were analyzed by 1.2% agarose gels with TAE buffer. The Wizard® SV Gel and PCR clean-Up system Kit (Promega) were used to purify the PCR products. Products were bi-directionally sequenced with specific PCR primers with a BigDye Terminator Cycle Sequencing Ready Reaction Kit (PE Applied Biosystem, USA) and carried out on ABI PRISM 310 Genetic Analyzer (PE Applied Biosystems, USA) by the Macrogen Inc. service, South Korea. The BLAST search (Basic Local Alignment Search Tool) in NCBI was used to confirm the sequence identification. The sequences obtained were sent and located in the NCBI database with the accession number.
Phylogenetic analysis
The phylogenetic tree was created using the phylogeny.fr platform: http://www.phylogeny.fr/programe (Dereeper et al. 2008). The phylogeny was reconstructed, using the maximum likelihood method (PhyML program), and the reliability for the internal branch was assessed, using the Bootstrapping procedure (100). The tree rendering was performed using with the TreeDyn software.
Pathogenicity of the new EPN isolate against Spodoptera littoralis
Spodoptera littoralis culture
A colony of S. littoralis was reared at 25 ± 1 °C with 70% RH under 16 h: 8 h light: darkness photoperiod. Larvae were fed on fresh castor leaves; in larger jars, the 6th larval instars were allowed to pupate. Emerged moths were supplied by a 10% sugar solution, and they laid their eggs on the provided leaves of Nerium oleander. Egg masses were kept in plastic jars until hatching.
Bioassays
Insecticidal activity of the new isolate was tested using 3rd instar larvae of S. littoralis that placed individually in plastic cups (10 cm diameter and 6 cm deep), filled with 100 gm. sterile wetted sandy soil. Four different concentrations of EPN were prepared (60, 90, 120 and 150 IJs/larvae), and the suspension of nematodes was mixed with the soil then the larvae placed in the cups which covered with plastic lids. For each concentration, 5 replicates were used; the control treatment was carried out using distilled water. The cups were incubated in a dark growth chamber at 26 ± 2 °C. The mortality rate was recorded after 48 h post-treatment.
Statistical analysis
The experimental design used was the randomized complete block design. Bioassay data obtained were subjected to an analysis of variance (Two-way ANOVA) and significant difference values calculated as Tukey’s statistic at P ≤ 0.05 (SAS Institute 2002).
Results
Nematodes’ isolation
Out of 10 samples, collected at the depth of 15–20 cm, only one sample of sandy clay loam soil was positive (10%) and exhibited a total virulence against the G. mellonella larvae based on the symptoms of dead larvae. The nematode was reared on the last instar larvae of G. mellonella, and the emerged nematodes were stored at 16 °C, under laboratory conditions.
Molecular identification and phylogenetic analysis
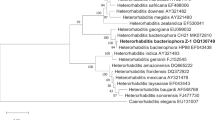
The EPN isolated from the positive samples were further identified to the species level. The PCR amplified product revealed approximately 700 bp of the 5.8S rDNA, ITS1 and ITS2 sequence, and according to the Blast search, the new isolate was belonged to the genus Heterorhabditis and showed 99% similarity with Heterorhabditis taysearae based on the sequence similarity, then submitted to GenBank with accession no. (MW543946.1). The maximum likelihood approach was used to reconstruct the phylogenetic relationships among taysearae species (Fig. 1).

Maximum likelihood ITS phylogenetic trees of Heterorhabditis species using ITS sequence. Based on nucleotide sequences of H. taysearae (alignment length approximately 500 bp using phylo_tree analysis (phylogeny.fr. program)
Potential of the new nematode isolate against Spodoptera littoralis
The new isolate of H. taysearae was tested against 3rd instar S. littoralis larvae using 4 different concentrations (60, 90, 120 and 150 IJs/larvae), and the mortality rates were recorded. Data showed that there was no larval mortality after 24 h of the treatment. Maximum mortality rate was recorded at 150 IJs/larvae (90 and 100%), followed by 120 IJs/larvae (80 and 90%), while the concentration 60 IJs/larvae showed the minimum mortality rate (30 and 60%) after 48 and 72 h post-treatment, respectively (Fig. 2). The data showed that mortality percentages increased by increasing the IJs concentration as well the data presented in Table 1, indicating that the exposure time to different nematode concentration differs significantly (Table 1).

Mortality of Spodoptera littoralis larvae after treatment with Heterorhabditis taysearae at concentrations of 60, 90, 120 and 150 IJs/larva after 48 and 72 h post-treatment
Discussion
Isolating and identifying indigenous EPN populations and species is an important step to attain effective biological control against pests due to their better adaptation to local environment (Nikdel and Niknam 2015). At the present study, a new EPN isolate was recovered from one sample out of 10 samples (10%) from cabbage field at Ismailia governorate, Egypt and the positive sample was classified as sandy clay loam, and these results agree with Khashaba et al. (2020).
DNA sequencing of various regions of the EPN genome has become the most appropriate tool, not only for determining phylogenetic relationships, but also for species categorization as morphological identification is not an accurate method (Stock 2009).
The internal transcribed spacer region of rDNA (ITS-1, 5.8S rDNA, and ITS-2) revealed enough genetic variation to distinguish EPN species and determine their evolutionary relationships (Ngugen et al. 2001).
Based on the molecular identifications and ITS region sequences, the new isolate was grouped by H. taysearae species in a phylogenetic clade. The surveys that achieved by Bhat et al. (2020) in agricultural soils in Egypt supported the present data which showed that heterorhabditid species were more prevailing than steinernematids such as H. indica (Abd El Azim and Khashaba 2021), H. baujardi (Abd El-Rahman 2006), H. aegypti and H. taysearae which were considered as new species from the soils of Egypt (Abd-Elgawad and Ameen 2005). The efficacy of EPNs to control pests may be changeful because it depends on factors such as the nematodes concentrations and exposure duration to the nematodes (Woodring and Kaya 1988). These results were consistent with the present study which suggested that the new EPN isolate concentration of 150 IJs/larvae of the new isolate caused 100% larval mortality, while those of 60 and 90 IJs/larvae showed low mortality rate (60 and 70%), respectively. These data also agree with Sobhy et al. (2020) who reported that the highest mortality (100%) for all larval instars of S. littoralis and Agrotis ipsilon occurred by 200 IJs/dish of the nematode species H. bacteriophora (HP88). Also, Ibrahim et al (2019) who tested the pathogenicity of the EPN, H. zealandica on the last larval instar of greater wax moth, G. mellonella and reported that there was a positive correlation between the concentration of pathogen and the percentage of host mortality.
Conclusions
The present study revealed the isolation of the indigenous EPN species, H. taysearae adapted to the local environment. The pathogenicity of the native EPN isolate indicated that optimization of pathogen concentration resulted to a high mortality rate.
Availability of data and materials
All datasets are presented in the main manuscript.
Abbreviations
- EPN:
-
Entomopathogenic nematodes
- DNA:
-
Deoxyribonucleic acid
- BLAST:
-
Basic Local Alignment Search Tool
- ITS:
-
Internal transcribed spacer region
- NTS:
-
Non-transcribed spacer
- SSU:
-
The small subunit
- LSU:
-
The large subunit
- NCBI:
-
National Center for Biotechnology Information
References
Abd El Azim AM, Khashaba EHK (2021) Genetic variability among three Egyptian isolates of Heterorhabditis indica using a new marker technique (SCoT). Egypt J Biol Pest Control 31:71. https://doi.org/10.1186/s41938-021-00419-0
Abd El-Rahman RM (2006) Isolation and identification of Heterorhabditis baujardi as a first record from Egypt and studying some of its biological traits. Egypt J Appl Sci 8:298–307. https://doi.org/10.1186/s41938-020-0212-y
Abd-Elgawad MMM, Ameen HH (2005) Heterorhabditis egyptiin. sp. (Rhabditida: Heterorhabditidae) from Egypt. Egypt J Agric Res 2:855–870
Bedding RA, Akhurst RJ (1975) A simple technique for the detection of insect parasitic nematodes in soil. Nematologica 21(1):109–110. https://doi.org/10.1163/187529275X00419
Bhat AH, Chaubey AK, Askary TH (2020) Global distribution of entomopathogenic nematodes, Steinernema and Heterorhabditis. Egypt J Biol Pest Control 30:31. https://doi.org/10.1186/s41938-020-0212-y
Burnell AM, Stock SP (2000) Heterorhabditis, Steinernema and their bacterial symbionts-lethal pathogens of insects. Nematology 2:31–42. https://doi.org/10.1163/156854100508872
Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard JF, Guindon S, Lefort V, Lescot M, Claverie JM, Gascuel O (2008) Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res 36:W465–W469. https://doi.org/10.1093/nar/gkn180.Epub
Dziedziech A, Shivankar S, Theopold U (2020) High-resolution infection kinetics of entomopathogenic nematodes entering Drosophila melanogaster. Insects 11(1):60. https://doi.org/10.3390/insects11010060
El-Gaied L, Mahmoud A, Salem R, Elmenofy W, Saleh I, Abulreesh H, Arif I, Osman G (2020) Characterization, cloning, expression and bioassay of vip3 gene isolated from an Egyptian Bacillus thuringiensis against whiteflies. Saudi J Biol Sci 27:1363–1367
Ibrahim SAM, Salem HHA, Taha MA (2019) Dual application of entomopathogenic nematodes and fungi on immune and antioxidant enzymes of the greater wax moth, Galleria mellonella L. Egypt J Biol Pest Control 29:20. https://doi.org/10.1186/s41938-019-0125-9
Institute S (2002) SAS/stat user’s guide. SAS Institute Inc., Cary
Joyce SA, Burnell AM, Powers TO (1994) Characterization of Heterorhabditis isolates by PCR amplification of segments of mtDNA and rDNA genes. J Nematol 26:260–270
Kaya HK, Stock SP (1997) Techniques in insect nematology. In: Lacey LA (ed) Manual of techniques in insect pathology. Academic Press, London, pp 281–324
Khashaba EHK, Abd El Azim AMA (2021) Isolation, identification, and study of the genetic diversity between three entomopathogenic nematodes belonging to Heterorhabditis sp. using ISSR technique. Egypt J Biol Pest Control 31:84. https://doi.org/10.1186/s41938-021-00425-2
Khashaba EHK, Moghaieb REA, Abd El Azim AM, Ibrahim SAM (2020) Isolation, identification of entomopathogenic nematodes, and preliminary study of their virulence against the great wax moth, Galleria mellonella L. (Lepidoptera: Pyralidae). Egypt J Biol Pest Control 30:55. https://doi.org/10.1186/s41938-020-00257-6
Labaude S, Griffin CT (2018) Transmission success of entomopathogenic nematodes used in pest control. Insects 9:72
Liu W, Chen T, Hou R, Chen C, Tu W (2020) The invasion and encapsulation of the entomopathogenic nematode, Steinernema abbasi, in Aedes albopictus (Diptera: Culicidae) Larvae. Insects 11:832
Lulamba TE, Serepa-Dlamini MH (2020) Molecular identification of a Heterorhabditis entomopathogenic nematode isolated from the northernmost region of South Africa. Egypt J Biol Pest Control 30:77
Malbert-Colas A, Drozdz T, Massot M, Bagni T, Chertemps T, Maria A, Maïbèche M, Siaussat D (2020) Effects of low concentrations of deltamethrin are dependent on developmental stages and sexes in the pest moth Spodoptera littoralis. Environ Sci Pollut Res 27:41893–41901
Nguyen K, Maruniak J, Adams B (2001) Diagnostic and phylogenetic utility of the rDNA internal transcribed spacer sequences of steinernema. J Nematol 33:73–83
Nikdel M, Niknam G (2015) Morphological and molecular characterization of a new isolate of entomopathogenic nematode, Steinernema feltiae (Filipjev) (Rhabditida: Steinernematidae) from the Arasbaran forests. Iran J Asia-Pacific Biodivers 8:144–151
Paniagua Voirol LR, Frago E, Kaltenpoth M, Hilker M, Fatouros NE (2018) Bacterial symbionts in Lepidoptera: their diversity, transmission, and impact on the host. Front Microbiol 9:556. https://doi.org/10.3389/fmicb.2018.00556
Peat SM, Hyman BC, Adams BJ (2009) Phylogenetics and population genetics of entomopathogenic and insect-parasitic nematodes. In: Stock P, Vandenberg J, Glazer I, Boemare N (eds) Insect pathogens. Molecular approaches and techniques. CABI, Oxfordshire, pp 166–184. https://doi.org/10.1079/9781845934781.0241
Shapiro-Ilan DI, Han R, Dolinksi C (2012) Entomopathogenic nematode production and application technology. J Nematol 44:206–217
Sobhy HM, Abdel-Bary NA, Harras FA, Faragalla FH, Husseinen HI (2020) Efficacy of entomopathogenic nematodes against Spodoptera littoralis (Boisd.) and Agrotis ipsilon (H.) (Lepidoptera: Noctuidae). Egypt J Biol Pest Control 30:73. https://doi.org/10.1186/s41938-020-00265-6
Stock SP (2009) Molecular approaches and the taxonomy of insect-parasitic and pathogenic nematodes. In: Stock SP, Vandenburg J, Glazer I, Boemare N (eds) Insect pathogens: molecular approaches and techniques. CAB International Press, Wallingford, pp 71–100
Woodring JL, Kaya HK (1988) Steinernematid and heterorhabditid nematodes: a handbook of biology and techniques southern cooperative series bulletin, vol 331. Arkansas Agricultural Experiment Station, Fayetteville, p 30
Acknowledgements
I would like to thank Prof. Dr. Gamal A. El Kady (Plant Protection Department, Faculty of Agriculture, Suez Canal University, Egypt) and prof. Dr. Reda E.A. Moghaieb (Genetic Engineering Research Center, Faculty of Agriculture, Cairo University, Egypt) for helping in the critical reading of the manuscript.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
A.M.A.A. design the experiments and practical work and manuscript writing. The author read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The author declares that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abd El Azim, A.M. Efficacy of the entomopathogenic nematode isolate Heterorhabditis taysearae to control the cotton leafworm, Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Egypt J Biol Pest Control 32, 4 (2022). https://doi.org/10.1186/s41938-022-00503-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00503-z