
Abstract
Background
Entomopathogenic nematodes (EPNs) of the families Heterorhabditidae and Steinernematidae are used as biological control agents of several insect pests in lieu of synthetic pesticides.
Results
Using an insect-baiting technique, naturally occurring EPN isolates were collected from banana and rice fields in the municipality of Hagonoy, the Philippines. A total of 4 isolates were recovered from all the sites namely, HBP1, HBP2, HR3 and HR4. By combining morphometrical and ITS rDNA data, all the isolates were identified as Heterorhabditis indica. To initially assess their infectivity, virulence tests were conducted on the last instar larvae of the super worm, Zophobas morio Fabricius (Coleoptera: Tenebrionidae), the model insect. HR3 isolate had the highest percentage mortality (56%), followed by HBP1 and HBP2 (33%), which were comparable to the available nematode in the market, Heterorhabditis bacteriophora (44%), whereas the lowest was obtained in HR4 isolate (11%).
Conclusions
This study accounted 4 more H. indica isolates in addition to pre-existing isolates in the country, expanding its habitat range and geographic distribution. This also provides baseline information on the potential biocontrol utilization of native EPNs against soil-dwelling insect pests of agricultural crops. Further investigations are required to assess their efficacy against several other insect pests of economic importance for the integrated pest management (IPM) programmes in the country.
Similar content being viewed by others


Background
Entomopathogenic nematodes (EPNs) of the genera Heterorhabditis (Strongyloidea: Heterorhabditidae) and Steinernema (Strongyloidoidea: Steinernematidae) are used as biological control agents (BCA) of several insect pests (Lacey et al. 2015). Together with their respective symbiotic bacteria Photorhabdus and Xenorhabdus (Proteobacteria: Morganellaceae), these associations are considered as pathogenic and lethal (Kaya et al. 2006). EPNs are specific to insects, safe to non-target organisms including humans and other vertebrates and do not pollute the environment (Ehlers and Shapiro-Ilan 2005). The infective juvenile (IJ), also known as Dauer juvenile, is the only free-living stage present and surviving in the soil until it locates and penetrates its hosts. IJs enter the host via natural body openings or by directly penetrating the cuticle until it reaches the haemocoel (Dowds and Peters 2002). Once inside, bacterial cells stored in the IJ gut are released and then proliferate which in turn rapidly kill the insects within 48–72 h (Boemare 2002) or may take longer time for larger insects (Dillman et al. 2012). The nematodes subsequently recover, start feeding on the bacteria-rich cadavers and develop into several generations until the food source is entirely exploited.
EPNs have been documented to occur naturally in different soils, ranging from natural and managed ecosystems of all continents except Antarctica. Several surveys in agricultural areas were done to collect and identify native EPNs for the control of local target pests. To date, around 100 Steinernema and 16 Heterorhabditis have been described worldwide (Bhat et al. 2020a).
Heterorhabditis is a widely distributed genus with several successful stories as BCA of insect pests (Abate et al. 2017). The species H. indica has a global distribution (Bhat et al. 2020a) and with products being formulated and marketed for pest control (Kaya et al. 2006; Guo et al. 2017). In the Philippines, although considered as a megadiverse country, only very few EPNs have been described and reported. The first documented H. indica species was isolated in central Philippines (Cebu, Visayas island) from different vegetable growing areas using morphological characterization (Pascual et al. 2017), whereas in northern Philippines (Batangas, Luzon island), the lone H. indica isolate was described using molecular (ITS rDNA) data (Caoili et al. 2018). Recently, H. indica was also recovered in southern Philippines (Iligan, Mindanao island) by employing the combination of morphology and molecular tools (Leonar and Sumaya 2019). These findings may present useful information in the expanded use of EPNs as BCA in agriculture.
In Philippines, most of local farmers still opt to use synthetic chemicals to control pest infestations. EPNs may offer promising contributions for pest control strategies in these areas. In fact, some EPN species have been reported to be effective on banana pests such as borer (Ping and Xizhan 1991), pseudostem weevil (Lu and Liang 2012), root borer weevil (Mwaitulo et al. 2011) and potential rice pests (Kega et al. 2013).
The present study aimed to (1) isolate naturally occurring or native EPN isolates in banana and rice fields in Hagonoy, Davao del Sur, Philippines; (2) determine the soil parameters which may influence EPN occurrence; (3) identify the isolates using morphometrical data and molecular taxonomic genes; and (4) assess their initial virulence against the last instar larvae of super worm, Zophobas morio Fabricius, 1776 (Coleoptera: Tenebrionidae). This can be baseline information of efficient native isolates for banana and rice and for other major commodities in the country.
Methods
Soil sampling
Soil sampling was carried out in different banana and rice fields of Hagonoy, Davao del Sur, Philippines (6° 40′ North, 125° 19′ East and 21.8 m above sea level). Sampling was conducted from June to July 2019 during the onset of the rainy season. Soil samples were collected at 15 cm depth and placed in plastic containers (Orozco et al. 2014). From each area, a total of 5 subsamples were collected at 100-m distance from each other, following a zigzag pattern and then homogenized to form one composite sample of approximately 2 kg. Soil parameters such as soil temperature, moisture, pH, and texture were also recorded. Soil temperature was determined in situ, using a thermometer; soil moisture was measured by oven drying method for 24 h at 100 °C; and the difference in wet and dry soils were calculated (Buzzi et al. 2010). The soil pH was measured using standard methods in a water suspension (soil: water ratio of 1:2.5, w/v) (Batjes 1995). Finally, identification of soil texture was done by sedimentation method (Taubner et al. 2009) and the percentages of sand, silt and clay in soil samples were determined for the overall soil texture.
Insect-baiting technique and pure culturing
Following the method originally described by Bedding and Akhurst (1975), insect-baiting technique was done with modifications, using the last instar larvae of super worm, Z. morio as baits. Soils were incubated at 25±2 °C and monitored regularly until insect cadaver was evident. All cadavers collected were thoroughly washed with distilled water, disinfected and placed in a modified White trap (White 1927) until emergence of infective juveniles (IJs). IJs were then harvested, cleaned and stored at 10–20 °C. For pure culture and mass production, a series of re-inoculations were carried out using the last instar larvae of Galleria mellonella [L.] (Lepidoptera: Pyralidae) (Hoy et al. 2008).
Morphological identification
A stepwise transfer of IJ, male and female nematodes to anhydrous glycerol was done according to the original description of Seinhorst (1959) with modifications by De Grisse (1969). Three different solutions were used: solution 1 (96% alcohol, 4% formalin and glycerol), solution 2 (4% formalin and glycerol) and solution 3 (pure glycerol). Preliminary observations under the microscope were done to all isolates on some important characters such as the anterior part of IJs, the vulva of females, the spicule, gubernaculum and bursal rays and patterns of males and subsequently identified up to EPN-species groups. Based on the morphological observations, a representative isolate for those isolates with similar morphology was processed for a comprehensive morphometric analysis. Following the description of Nguyen and Hunt (2007), key morphological characters were observed and subsequently measured such as the total body length (L), maximum body width or diameter (MBD), pharynx length (ES), distance from anterior end to excretory pore (EP), distance from anterior end to nerve ring (NR), anal body diameter (ABD), tail length (T) and distance from anterior end to vulva (V) for female nematodes. Additional important characters such as spicule length (SL) gubernaculum length (GL), testis reflexion (TR) and the number and pattern of bursal rays were considered. Based on these, the ratios and percentages were calculated. Using an Olympus BX41 microscope equipped with a Nikon DP27 digital sight camera and a CellSens imaging software, photomicrographs and measurements of the key characters were done. All measurements were calculated in micrometres (μm) except ratio and percentages.
Molecular identification
Extraction of the total genomic DNA was done using a DNA extraction kit following the manufacturer’s protocol (Dongsheng Biotech, Inc.). Briefly, 5 freshly picked IJs from each isolate were placed in 1.5-ml Eppendorf tubes containing few drops of distilled water and were chopped using closed-end micropipette tips. From lysis to elution, a series of different solutions, proteinase K, wash and TE buffers were added accordingly coupled with vortexing and centrifugation processes. The collection tubes containing the extracted DNA were kept at − 20 °C in deep freezer. The Internal Transcribed Spacer 1-5.8S- region of the rDNA (ITS1, 5.8S, ITS2) was amplified using PCR. The following primers were used: TW81 (5-GTTTCC GTA GGT GAA CCT GC-3) as forward and AB28 (5-ATA TGC TTA AGT TCA GCG GGT-3) as reverse primers (Joyce et al. 1994). PCR conditions were programmed as follows: initial denaturation at 94 °C for 5 min, followed by 35 cycles of 94 °C for 1 min, 55 °C for 1 min 30 s, and 72 °C for 2 min, and a final extension at 72 °C for 7 min. Products were sent to Macrogen, Inc. (Seoul, South Korea) for processing purification and unidirectional Sanger sequencing. The obtained sequences were manually curated, trimmed and submitted to the National Center for Biotechnology Information (NCBI) under accession numbers HBP1 (MT458695), HBP2 (MT457558.1), HR3 (MT458704) and HR4 (MT458706). They were nucleotide blasted and a cut of ≥ 97% identity was considered for the same species (Yooyangket et al. 2018; Bhat et al. 2019).
Sequence alignment and phylogenetic analyses
The sequences were edited and compared with those already present in GenBank, using the Basic Local Alignment Search Tool (BLASTN) of the National Center for Biotechnology Information (NCBI) (Altschul et al. 1990). The newly obtained ITS (ITS1, 5.8S, ITS2) rDNA sequences were manually edited using BioEdit 7.2.6 (Hall 1999) and aligned with other relevant segments of same rDNA gene sequences available in GenBank using Clustal Omega alignment. The base substitution model was evaluated using jModeltest2.1.10 (Darriba et al. 2012). Phylogenetic trees were elaborated, using the Bayesian inference method as implemented in the programme MrBayes 3.2.7 (Ronquist et al. 2012). For analysis in jModeltest, the HKY + I + G model was selected. The selected model was initiated with a random starting tree and ran with the Markov chain Monte Carlo (MCMC) for 1 × 106 generations. The Bayesian tree was ultimately visualized using the FigTree programme 1.4.4 (Rambaut 2018). Caenorhabditis elegans (X03680.1) was used as the out group and to root the trees.
Virulence tests
Virulence assays were carried out, following the methods described by Sumaya et al. (2018) and Alonso et al. (2018) with modifications. In the present study, Z. molitor was used as model insect instead of the widely used G. mellonella and Tenebrio molitor (Coleoptera: Tenebrionidae). Three giant last instar larvae of Z. morio were placed into Petri dishes (60 × 15 mm size) with 20 g of sterilized sand at 20% moisture. A total of 300 IJs (100 IJs per insect larva) in 1.3 ml saline solution were inoculated and incubated at 25±2 °C. Control plates had the same set-up but only received 1 ml saline solution. According to Dillman et al. (2012), it may require longer time to kill larger insects. Therefore, mortality of Z. morio larvae were assessed 7 days post-inoculation. Insect cadavers were dissected to confirm the presence of nematodes. This experiment was replicated and repeated 3 times, using different batches of EPNs propagated in vivo. EPN-based products that were currently available in the market (GardeningZone, CA, USA) such as Heterorhabditis bacteriophora and Steinernema feltiae instead of the unavailable H. indica were used for comparison.
Data analyses
Normality tests were first done before carrying out further analysis. Treatment differences amongst 4 H. indica isolates and the commercial strains were analysed using one-way ANOVA. Differences between treatments were compared using post hoc Tukey’s HSD at P ≤ 0.05. All analyses were performed using XLSTAT statistical software 2019.3.1.
Results
Nematode recovery from soil samples
From a total of 10 soil samples collected from banana and rice field soils in Hagonoy, each with 5 subsampling points, 6 soil samples were found EPN-positive with a recovery rate of 60%. All EPN-positive soil samples had the following measurement range: soil temperature (26.2–27 °C); soil moisture (21.2–25.8%); soil pH (6.5–7.17); and soil texture (silty clay- silty loam) (Table 1). The baited nematodes were further pure-cultured, mass produced and maintained in G. mellonella larvae. Only 4 out of 6 EPN-positive areas were processed for morphology and molecular identification. Two nematodes baited from nematode-positive areas such as banana sampling site 4 and rice sampling site 1 were not processed for further experiments due to insufficient number of nematodes which emerged from the cadavers.
Morphology and morphometric analyses
Morphological key characters such as the anterior part of IJs, the vulva of females, the spicule, gubernaculum and bursal rays and patterns of males were observed in all 4 nematode isolates (HBP1, HBP2, HR3 and HR4). All local isolates were observed to have a close resemblance in terms of morphology. These 4 isolates were preliminary identified under species-group Heterorhabditis indica. Consequently, a representative of Hagonoy isolates was selected for comprehensive morphometric analysis of IJs (Table 2), adult males and females (Table 3) and compared with the H. indica type strain from India including the previously described populations in Vietnam and Philippines. The IJ body length of Hagonoy isolate had a mean and range of 482 (403–529) μm, which was found to be slightly smaller than the type strain (528; 479–573 μm) and H.TN48 isolate (563; 348–622 μm) from Vietnam but similar to the F05 isolate (481; 432–560 μm) from the Philippines. The maximum body diameter of 21 (19–23) μm, a ratio of 22 (22–23) and b ratio of 4 (3–4) had similar sizes with the type strain, H.TN48 and F05. Tail length (without sheath) of 48 (42–54) μm was slightly smaller than the F05 isolate.
Concerning male measurements, male isolates of Hagonoy were slightly smaller with the size of 557 (516–629) μm than the type strain 721 (573–788) μm but still within the size range. Additionally, Hagonoy males have a comparable size with H.TN48 isolate from Vietnam (689; 596–818 μm). The anal body diameter was 21 (20–22) μm and found similar to the type strain (26; 22–32 μm) as well as the H.TN48 isolate from Vietnam (24; 21–29 μm). The maximum body diameter was 30 (26–36) μm, which was similar to F05 isolate (35; 26–44 μm) and smaller than the type strains (42; 35–46 μm). The testis reflex (160; 116–204 μm) was bigger than the type strain (91; 35–144 μm) but within the size range. On the other hand, spicule (22; 19–24 μm) and gubernaculum (11; 11–12 μm) lengths were smaller than the type strain, but most importantly, the ratio of spicule and gubernaculum (SP/GL) (0.52; 0.5–0.6) was similar to the type strain (0.5; 0.4–0.6), F05 (0.47; 0.4–0.5) and H.TN48 (0.5; 0.4–0.6) isolates.
Adult females were compared to the type strain only. Protruding vulval lips with transverse slit located in the middle of the body with a percentage (%) 57 (51–75) were observed. The body length was again smaller than the type strain with a size of 1148 (1127–1185) μm, however still within the range (1600; 1200–1800 μm). The anal body diameter and pharynx length have the size and range of 34 (24–40) μm and 123 (105–147) μm, respectively, with comparable sizes in relation to the type strain (26; 22–32 μm) and (131; 120–139 μm). Although some characters had different sizes, however in general IJs, males and females of the representative Hagonoy isolate resemble and were within the size range with the type strain and other H. indica species from Philippines and Vietnam.
Molecular characterization and phylogenetic analyses
In the present study, HBP1, HBP2, HR3 and HR4 isolates yielded approximately 700 bp ITS rDNA sequences and were nucleotide BLASTed with all sequences related to the species available in the NCBI GenBank database. Based on sequence homology, these 4 isolates closely matched with H. indica from India and Heterorhabditis sp. from Thailand, having the following sequence identity similarities: HBP1 (100%), HBP2 (99.85%), HR3 (98.26%) and HR4 (100%). Their respective accession numbers were deposited in the NCBI GenBank. The characterization was further confirmed by phylogenetic analysis, using ITS rDNA sequences of all described type populations of Heterorhabditis species, including present isolates and H. indica populations from India (Fig. 1).

Bayesian inference tree previously and newly sequenced Heterorhabditis indica (bold) and other type described species of Heterorhabditis based on sequences of Internal Transcribed Spacer (ITS1-5.8S-ITS2) rDNA region. Bayesian posterior probabilities (%) are given for each clade. The scale bar shows the number of substitutions per site. Caenorhabditis elegans was used as an outgroup taxon
Virulence against Zophobas morio larvae
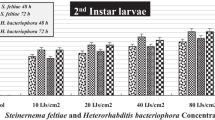
Virulence of the 4 native H. indica isolates was preliminary assessed against the last instar larvae of the superworm, Z. morio, as a model insect under laboratory conditions. A colouration of red to purple indicated Heterorhabditis infection and cadaver dissection was performed to confirm successful infection. Significant differences among treatments was observed (p = 0.01). The highest percentage mortality was obtained in HR3 isolate (56%). This isolate had a mean mortality comparable to the available commercial strain H. bacteriophora (44%). In case of HBP1 and HBP2, similar percentage mortality was observed (33%) that was comparable to the other commercial species, Steinernema feltiae (22%); however, HR4 isolate (11%) showed the lowest mortality (Fig. 2). For the control set-up, which received only water, insect mortality was not observed.

Percentage mortality of the four native Heterorhabditis indica (HBP1, HBP2, HR3, HR4) isolates and the commercial nematodes Heterorhabditis bacteriophora and Steinernema feltiae against Zophobas morio larvae at 25±2 °C 1 week after infective juvenile (IJ) inoculation. Error bars indicate standard deviations with three replications. Different letters above error bars indicate significant differences between strains and lines (Tukey’s HSD test; p ≤ 0.05)
Discussion
In various global surveys on EPN prevalence, varying results have been recorded ranging from low to high recovery or detection. Although in the present study, only few samples were collected (10 soils), mainly focused on banana and rice soils as major crops in Hagonoy, still 6 soil samples harboured EPNs. Such a high recovery can be attributed to several factors, as agreed with Ma et al. (2010) that H. indica may have no particular habitat preference and it could be detected in diverse natural and agricultural habitats. Two isolates from banana sampling site 4 and rice sampling site 1 were recovered, but were not processed for further experiments and these may likely be facultative necromenic nematodes as scavengers (Campos-Herrera et al. 2019).
Concerning different soil parameters, all EPN-positive sites had soil temperature values of 26.2 to 27 °C. These validate the very recent and first report of H. indica in Fiji Islands, where soil temperature of positive sites ranged from 24 to 39 °C (Kour et al. 2020). Meanwhile, the soil moisture content of EPN-positive soils ranged from 21.2 to 25.8%, possibly due to the onset of the rainy season or the regular irrigation by farmers or growers in their banana and rice fields. These results were supported by the studies of Yadav (2012) in India where they found EPNs in soils with moisture ranging from 8 to 25% and by Kour et al. (2020) where moisture levels for H. indica-positive soils ranged from 6 to 24%. However, this contradicted the report of Yooyangket et al. (2018) where EPN-positive samples in Thailand had a moisture content of 1–3% only and no EPNs were detected in moisture levels between 4.0 and 8.0%. EPNs in general require optimal moisture to move around the soil and seek for target insect hosts. With regard to soil pH, a recorded range was between 6.5 and 7.17 with findings similar to those H. indica isolated in Henan, China, with soil pH levels of 6.7–7.8 (Ma et al. 2010) and in Thailand where EPN-positive samples had a soil pH of 6.2–7.0 (Yooyangket et al. 2018). A wider range of soil pH tolerance (from 4.8 to 7) was observed for soils with 7 isolates of H. indica collected in Nepal (Khatri-Chhetri et al. 2010). For the soil texture of obtained samples, it was found to be either silty clay or silty loam, which is considered desirable for rice cultivation to conserve water (Weyers and Gramig 2017) and also suited for banana production (Niranjana et al. 2011). This particular result is in agreement with the results of Kanga et al. (2012) and Abate et al. (2017). Contrary to the findings, most reports have indicated that EPNs, especially Heterorhabditis, were frequently found in or had higher preferences to sandy loam and loamy soils compared to clay soils (Ma et al. 2010). In fact, the H. indica isolated from Luzon island, Philippines was from sandy loam (Caoili et al. 2018) and another isolate from Iligan city in Mindanao island were recovered from sandy soils in the coastal grassland (Leonar and Sumaya 2019). This specific preference and higher recovery of EPNs in sandy soils may be attributed to easier movement and good aeration (Noosidum et al. 2010).
Generally, obtained data on soil parameters suggest insufficient evidence to explicitly pinpoint the specific effects of these soil parameters on EPN prevalence in the areas. What is clear for now is that the response to soil or environmental factors differs with EPN species in these areas. One can likewise argue that preference or response can vary with the same EPN species yet occupies different geographical regimes. Nonetheless, soil parameters such as temperature, moisture, pH and soil texture along with other factors have been recorded to influence the diversity, distribution, survival and infectivity of EPNs (Kanga et al. 2012; Abate et al. 2017; Khashaba et al. 2020). These nematodes for sure have optimal requirements for survival though with some populations co-exist or tolerate extreme conditions, which can theoretically be a form of adaptation to fluctuating environmental conditions.
To further increase resolution and reliability on the obtained EPN species identification, the combination of morphology, morphometric data of a representative strain and molecular tools, using the ITS rDNA region, were employed (Spiridonov 2017). This ITS rDNA is considered a versatile gene marker for nematode identification (Hsieh et al. 2009) and recently considered as an informative marker for species identification, barcoding and phylo-geographical studies of Heterorhabditis species (Dhakal et al. 2020). The result of the morphological and morphometric analysis complemented with our molecular data. The initial diagnosis based on morphology and morphometrics was first belonging to H. indica species-group then narrowed down to H. indica, which was confirmed by sequence results. Although the body length of the IJs were slightly smaller than the type strain from India (Poinar Jr et al. 1992), but is similar to the reported H. indica F05 isolate from Visayas island, Philippines (Pascual et al. 2017). For the males, the length is smaller than the type strain, it was noticed that it was comparable with the IJs from Vietnam (Phan et al. 2003). Other key characters fall within the size range. With the ITS sequences of the local isolates, HBP1 (100%), HBP2 (99.85%), HR3 (98.26%) and HR4 (100%) showed a very high homology to H. indica from India and Heterorhabditis sp. from Thailand.
Many researchers have opted to first use model insects like the lepidopteran Galleria and coleopteran Tenebrio to initially screen and assess the biocontrol potential of native or local isolate collections (Khashaba et al. 2020) before applying on the target insect pest in crop protection. Additionally, these 2 insects are widely used as baits to recover or isolate EPN species from different soils worldwide since they are easy to culture and highly susceptible (Bhat et al. 2020b). Whereas other researchers use these 2 models for large-scale production and most importantly for virulence quality checks prior to application and marketing (Ehlers and Shapiro-Ilan 2005). In this study, we used a last instar larva of a different tenebrionid, Z. morio, to initially assess the virulence of our H. indica local isolates for further uses. Some studies also used Z. morio to evaluate the infectivity or virulence ability of their EPN species (Alonso et al. 2018). Like those abovementioned model insects, Z. morio can be easily cultured and can be effortlessly seen in sand assays. However, the larvae are remarkably large in size and possess thick cuticles, which may be a challenge for EPNs’ rapid penetration and infection. Overall, we have obtained a potential isolate to be evaluated further against target pests under laboratory and field conditions with different nematode concentrations. Further investigations are required to assess its efficacy against several other insect pests of economic importance for the IPM programme in the country.
Conclusions
Initial diagnosis based on morphology and morphometrics was first belonging to H. indica species-group then narrowed down to H. indica, which was confirmed by sequence results. A total of 4 native H. indica isolates were documented from banana (HBP1 and HBP2) and rice (HR3 and HR4) fields, providing additional account of EPN species in the country and extending their habitat ranges and geographical distribution. This study implies that banana and rice field soils in Hagonoy are naturally rich in H. indica with 3 native isolates inducing a high mortality against Z. morio comparable to the commercial strains. Therefore, these isolates could be potentially substantial for the biocontrol of insect pests in the area.
Availability of data and materials
The data and material of this manuscript are available upon reasonable request.
Abbreviations
- IPM:
-
Integrated pest management
- ITS:
-
Internal Transcribed Spacer
- BLAST:
-
Basic Local Alignment Search Tool
- NCBI:
-
National Center for Biotechnology Information
- Z. morio :
-
Zophobas morio
- BCA:
-
Biological Control Agent
- H. indica :
-
Heterorhabditis indica
References
Abate BA, Wingfield MJ, Slippers B, Hurley BP (2017) Commercialisation of entomopathogenic nematodes: should import regulations be revised? Biocontrol Sci Techn 27(2):149–168. https://doi.org/10.1080/09583157.2016.1278200
Alonso V, Nasrolahi S, Dillman AR (2018) Host-specific activation of entomopathogenic nematode infective juveniles. Insects 9(2):59. https://doi.org/10.3390/insects9020059
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215(3):403–410
Batjes NH (1995) A global data set of soil pH properties. Technical Paper 27, International Soil Reference and Information Centre (ISRIC), Wageningen
Bedding RA, Akhurst RJ (1975) A simple technique for the detection of insect parasitic rhabditid nematodes in soil. Nematologica 21:109–110
Bhat AH, Chaubey AK, Askary TH (2020a) Global distribution of entomopathogenic nematodes, Steinernema and Heterorhabditis. Egypt J Biol Pest Co 30:1–5. https://doi.org/10.1186/s41938-020-0212-y
Bhat AH, Chaubey AK, Shokoohi E, Machado RAR (2020b) Molecular and phenotypic characterization of Heterorhabditis indica (Nematoda: Rhabditida) nematodes isolated during a survey of agricultural soils in Western Uttar Pradesh, India. Acta Parasitol 56:1–17. https://doi.org/10.1007/s11686-020-00279-y
Bhat AH, Chaubey AK, Shokoohi E, Mashela PW (2019) Study of Steinernema hermaphroditum (Nematoda, Rhabditida), from the West Uttar Pradesh, India. Acta Parasitol 64(4):720–737. https://doi.org/10.2478/s11686-019-00061-9
Boemare N (2002) Biology, taxonomy and systematics of Photorhabdus and Xenorhabdus. In: Gaugler R (ed) Entomopathogenic nematology. CABI publishing, Wallingford, pp 35–56
Buzzi O, Fityus S, Sloan SW (2010) Use of expanding polyurethane resin to remediate expansive soil foundations. Can Geotech J 47:623–634. https://doi.org/10.1139/T09-132
Campos-Herrera R, Blanco-Pérez R, Bueno-Pallero FÁ, Duarte A, Nolasco G, Sommer RJ, Martín JAR (2019) Vegetation drives assemblages of entomopathogenic nematodes and other soil organisms: Evidence from the Algarve, Portugal. Soil Biol Biochem 128:150–163
Caoili BL, Latina RA, Sandoval RF, Orajay JI (2018) Molecular identification of entomopathogenic nematode isolates from the Philippines and their biological control potential against lepidopteran pests of corn. J Nematol 50(2):99
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 9:772
De Grisse AT (1969) Redescriptionou modifications de quelques techniques utilisé es dans l’étude des nématodesphytoparasitaires. MededelingenRijksfakulteit Land-bouwwetenschappente Gent. 34:351–369
Dhakal M, Nguyen KB, Hunt DJ, Ehlers RU, Spiridonov SE, Subbotin SA (2020) Molecular identification, phylogeny and phylogeography of the entomopathogenic nematodes of the genus Heterorhabditis Poinar, 1976: a multigene approach. Nematology 1(aop):1–7
Dillman AR, Chaston JM, Adams BJ, Ciche TA, Goodrich-Blair H, Stock SP, Sternberg PW (2012) An entomopathogenic nematode by any other name. PLoS Pathog 8(3):e1002527.Dowds BCA, Peters A (2002) Virulence mechanisms. In: Gaugler R (ed) Entomopathogenic nematology. CABI publishing, New York, pp 79–98
Dowds BC, Peters AR (2002) Virulence mechanisms. In: Gaugler R (ed) Entomopathogenic nematology. CABI publishing, Wallingford, pp 79–98
Ehlers RU, Shapiro-Ilan DI (2005) Forum on safety and regulation. In: Grewal PS, Ehlers R-U, Shapiro-Ilan DI (eds). Nematodes as biocontrol agents. CABI publishing, Wallingford, pp. 107–114
Guo W, Yan X, Han R (2017) Adapted formulations for entomopathogenic nematodes, Steinernema and Heterorhabditis spp. Nematology 19(5):587–596
Hall TA (1999) Bioedit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Hoy CW, Grewal PS, Lawrence JL, Jagdale G, Acosta N (2008) Canonical correspondence analysis demonstrates unique soil conditions for entomopathogenic nematode species compared with other free-living nematode species. Biol Control 46(3):371–379
Hsieh FC, Tzeng CY, Tseng JT, Tsai YS, Meng M, Kao SS (2009) Isolation and characterization of the native entomopathogenic nematode, Heterorhabditis brevicaudis, and its symbiotic bacteria from Taiwan. Curr Microbiol 58(6):564–570
Joyce SA, Burnell AM, Powers TO (1994) Characterization of Heterorhabditis isolates by PCR amplification of segments of mtDNA and rDNA genes. J Nematol 26:260–270
Kanga FN, Waeyenberge L, Hauser S, Moens M (2012) Distribution of entomopathogenic nematodes in Southern Cameroon. J Invertebr Pathol 109(1):41–51
Kaya HK, Aguillera MM, Alumai A, Choo HY, De la Torre M, Fodor A, Ganguly S, Hazır S, Lakatos T, Pye A, Wilson M (2006) Status of entomopathogenic nematodes and their symbiotic bacteria from selected countries or regions of the world. Biol Control 38(1):134–155. https://doi.org/10.1016/j.biocontrol.2005.11.004
Kega VM, Kasina M, Olubayo F, Nderitu JH (2013) Management of Maliarpha separatella rag using effective entomopathogenic nematodes and resistant rice cultivars. J Entomol 10:103–109
Khashaba EH, Moghaieb RE, Abd El Azim AM, Ibrahim SA (2020) Isolation, identification of entomopathogenic nematodes, and preliminary study of their virulence against the great wax moth, Galleria mellonella L.(Lepidoptera: Pyralidae). Egypt J Biol Pest Co 30:1–7
Khatri-Chhetri HB, Waeyenberge L, Manandhar HK, Moens M (2010) Natural occurrence and distribution of entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) in Nepal. J Invertebr Pathol 103(1):74–78. https://doi.org/10.1016/j.jip.2009.10.007
Kour S, Khurma U, Brodie G, Hazir S (2020) Natural occurrence and distribution of entomopathogenic nematodes (Steinernematidae, Heterorhabditidae) in Viti Levu, Fiji Islands. J Nematol 52:1–17
Lacey LA, Grzywacz D, Shapiro-Ilan DI, Frutos R, Brownbridge M, Goettel MS (2015) Insect pathogens as biological control agents: back to the future. J Invertebr Pathol 132:1–41
Leonar ALC, Sumaya NHN (2019) Isolation and identification of entomopathogenic nematodes from selected areas in Lanao del Norte, Philippines and their infectivity against Zophobas morio (Coleoptera: Tenebrionidae). In: Entomology 2019, Entomological Society of America
Lu YY, Liang GW (2012) Controlling effect of Steinernema carpocapsae with spraying on banana pseudostem weevil Odoiporus longicollis Oliver in banana plantation. Guangdong Agricultural Sciences 10:33
Ma J, Chen S, Zou Y, Li X, Han R, De Clercq P, Moens M (2010) Natural occurrence of entomopathogenic nematodes in North China. Russ J Nematol 18(2):117–126
Mwaitulo S, Haukeland S, Sæthre MG, Laudisoit A, Maerere AP (2011) First report of entomopathogenic nematodes from Tanzania and their virulence against larvae and adults of the banana weevil Cosmopolites sordidus (Coleoptera: Curculionidae). Int J Trop Insect Sci 31(3):154–161. https://doi.org/10.1017/S1742758411000294
Nguyen KB, Hunt DJ (2007) Entomopathogenic nematodes: systematics, phylogeny and bacterial symbionts. In: Hunt DJ, Perry RN (eds) Nematology monographs and perspectives, Brill NV, The Netherlands
Niranjana KV, Ramamurthy V, Hegde R, Srinivas S, Koyal A, Naidu LG, Sarkar D (2011) Characterization, classification and suitability evaluation of banana growing soils of Pulivendla region, Andhra Pradesh. J Indian Soc Soil Sci 59(1):1–5
Noosidum A, Hodson AK, Lewis EE, Chandrapatya A (2010) Characterization of new entomopathogenic nematodes from Thailand: foraging behavior and virulence to the greater wax moth, Galleria mellonella L.(Lepidoptera: Pyralidae). J Nematol 42(4):281
Orozco RA, Lee MM, Stock SP (2014) Soil sampling and isolation of entomopathogenic nematodes (Steinernematidae, Heterorhabditidae). J Vis Exp 89:e52083. https://doi.org/10.3791/52083
Pascual PR, Pascual ML, Alburo HM (2017) First report of Entomopathogenic nematode Heterorhabditidae (Rhabditida) in organic vegetable farms in Cebu, Philippines. J Agric Manag 20:1–9
Phan KL, Subbotin S, Nguyen NC, Moens M (2003) Heterorhabditis baujardi sp. n. (Rhabditida: Heterorhabditidae) from Vietnam and morphometric data for H. indica populations. Nematology 5:367–382. https://doi.org/10.1163/156854103769224368
Ping XJY, Xizhan W (1991) Control of banana borer with Steinernema carpocapsae. Chinese J Tropical Crops 2:10
Poinar GO Jr, Karunakar GK, David H (1992) Heterorhabditis indicus n. sp. (Rhabditida: Nematoda) from India: separation of Heterorhabditis spp. by infective juveniles. Fundam Appl Nematol 15:467–472
Rambaut A (2018). FigTree, a graphical viewer of phylogenetic trees (Version 1.4.4). Available at http://tree.bio.ed.ac.uk/software/figtree.
Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61(3):539–542
Seinhorst JW (1959) A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 4:67–69
Spiridonov SE (2017) Entomopathogenic nematodes of the families Steineirnematidae and Heterorhabditidae: Morphology and Taxonomy. In: Abd-Elgawad MM, Askary TH, Coupland J (eds) Biocontrol agents: entomopathogenic and slug-parasitic nematodes. CABI publishing, Wallingford, pp 45–63
Sumaya NH, Gohil R, Okolo C, Addis T, Doerfler V, Ehlers RU, Molina C (2018) Applying inbreeding, hybridization and mutagenesis to improve oxidative stress tolerance and longevity of the entomopathogenic nematode Heterorhabditis bacteriophora. J Invertebr Pathol 151:50–58. https://doi.org/10.1016/j.jip.2017.11.001
Taubner H, Roth B, Tippkötter R (2009) Determination of soil texture: comparison of the sedimentation method and the laser-diffraction analysis. J Plant Nutr Soil Sci 72(2):161–171
Weyers SL, Gramig G (2017) Low-input and intensified crop production systems effects on soil health and environment. In: Al-Kaisi MM, Lowery B (eds) Soil health and intensification of agroecosytems. Academic Press, London, pp 277–303
White GF (1927) Scientific apparatus and laboratory methods: a method for obtaining infective nematode larvae from cultures. Science 66:302–303
Yadav AK (2012) Evaluation of the efficacy of three indigenous strains of entomopathogenic nematodes from Meghalaya, India against mustard sawfly, Athalia lugens proxima Klug (Hymenoptera: Tenthredinidae). J Parasit Dis 36(2):175–180
Yooyangket T, Muangpat P, Polseela R, Tandhavanant S, Thanwisai A, Vitta A (2018) Identification of entomopathogenic nematodes and symbiotic bacteria from Nam Nao National Park in Thailand and larvicidal activity of symbiotic bacteria against Aedes aegypti and Aedes albopictus. PloS one 13(4):e0195681
Acknowledgements
We would like to thank the banana and rice farmers of Hagonoy for their active participation and contribution during the field survey and soil sampling. We are grateful to the Nematology Research Group for their technical assistance with some field and laboratory works. Thanks are also due to Dr. Cesar Demayo and Ms. Christine Gonzaga for the assistance they provided during our microscopy works. We acknowledge the Department of Science and Technology (DOST-ASTHRDP) for the scholarship grant to RS. Lastly, our sincerest thanks to the two anonymous reviewers for improving the manuscript.
Funding
This study is partially funded by the Department of Science and Technology (DOST-ASTHRDP) for the field sampling and morphometric analysis.
Author information
Authors and Affiliations
Contributions
All authors contributed significantly to this research study from data analyses including the preparation, writing and review of the manuscript. MLN carried out the experiments. RS assisted in the field sampling and the morphometric part. SA was involved in the conceptualization, molecular analysis and writing. NPDS was involved in conceptualization, design of the study, analysis and writing. AHB was involved in the phylogenetic analysis, writing and editing. NHS conceptualized the study and was in-charge of resources, supervision, analysis and writing. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Navarez, M.L., Sangcopan, R., Aryal, S. et al. Native Philippine Heterorhabditis indica isolates from banana and rice fields and preliminary results of their virulence against the larvae of super worm (Zophobas morio Fabricius Coleoptera: Tenebrionidae). Egypt J Biol Pest Control 31, 46 (2021). https://doi.org/10.1186/s41938-021-00388-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-021-00388-4