
Abstract
Cervical cancer is the first identified human papillomavirus (HPV) associated cancer and the most promising malignancy to be eliminated. However, the ever-changing virus subtypes and acquired multiple drug resistance continue to induce failure of tumor prevention and treatment. The exploration of cervical cancer heterogeneity is the crucial way to achieve effective prevention and precise treatment. Tumor heterogeneity exists in various aspects including the immune clearance of viruses, tumorigenesis, neoplasm recurrence, metastasis and drug resistance. Tumor development and drug resistance are often driven by potential gene amplification and deletion, not only somatic genomic alterations, but also copy number amplifications, histone modification and DNA methylation. Genomic rearrangements may occur by selection effects from chemotherapy or radiotherapy which exhibits genetic intra-tumor heterogeneity in advanced cervical cancers. The combined application of cervical cancer therapeutic vaccine and immune checkpoint inhibitors has become an effective strategy to address the heterogeneity of treatment. In this review, we will integrate classic and recently updated epidemiological data on vaccination rates, screening rates, incidence and mortality of cervical cancer patients worldwide aiming to understand the current situation of disease prevention and control and identify the direction of urgent efforts. Additionally, we will focus on the tumor environment to summarize the conditions of immune clearance and gene integration after different HPV infections and to explore the genomic factors of tumor heterogeneity. Finally, we will make a thorough inquiry into completed and ongoing phase III clinical trials in cervical cancer and summarize molecular mechanisms of drug resistance among chemotherapy, radiotherapy, biotherapy, and immunotherapy.
Similar content being viewed by others


Introduction
Human papillomavirus (HPV) causes an overwhelming majority of cervical cancers (CCs) and an alarmingly increased proportion of oropharyngeal cancers (OPCs). As the earliest discovered HPV-associated cancer, the tumorigenesis and infiltration of cervical cancer are closely relevant to the persistent infection and genome integration of HPV [1, 2].
Although with clear etiology, tumor heterogeneity still exists and gradually becomes a new challenge in the field of HPV-associated cancer research. Three concepts of heterogeneity need to be clarified: inter-patient heterogeneity, inter-tumoral heterogeneity, and intra-tumoral heterogeneity. Differences in tumor phenotypes and genotypes among individuals or distinct tumor sites are defined as inter-patient heterogeneity and inter-tumor heterogeneity respectively. In contrast, intra-tumoral heterogeneity means genomic, transcriptomic, epigenetic, or phenotypic differences within the same tumor lesion which are associated with therapeutic resistance and considerably more challenging [3]. Mendelian law of inheritance suggests that the free combination of genes is an important reason for the emergence of biodiversity [4]. As these probabilistic events encounter Darwinian adaptational selection over time, tumor cells and normal cells will continue to compete in different quadrants of time and space [5]. Genetic intra-tumoral heterogeneity, inter-tumoral heterogeneity, and inter-patient heterogeneity are reflected in a dynamic process of tumorigenesis, invasion, metastasis, or drug resistance [6,7,8]. To overcome the dilemma of precision therapy, it is necessary to break through each of these aspects. The rapid development of sequencing technology provides a platform for revealing tumor heterogeneity. Scanty knowledge has been uncovered on how heterogeneity plays roles in tumor pathogenesis and precision therapy until application of single-cell transcriptome analysis. The advances in single-cell RNA sequencing (scRNA-seq) include distinguishing neoplastic from normal tissue in individual patients and different disease states [9, 10].
In this review, we explore the heterogeneity of cervical cancers from the perspectives of HPV-induced tumorigenesis, internal changes of human genome and molecular mechanisms of drug resistance. The molecular and clinical features of cervical squamous cell carcinoma are discussed in major. In addition, cancer stem cells, cervical adenocarcinoma, and neuroendocrine carcinoma are described respectively in the last chapter. Firstly, we emphasize the significant contribution of alterations of genetic material and HPV gene integration differences in tumorigenesis. Furthermore, we summarize the mechanisms of intra-tumoral and inter-tumoral heterogeneity among inchoate and advanced cancers. Finally, we attempt to explain the huge differences in resistance to therapies among populations through tumor heterogeneity and provide feasible strategies for precise treatment.
Inferring heterogeneity with HPV
Heterogeneity of geographical distribution
In the 1980s, German pathologist Dr Hausen identified HPV as the explicit cause of cervical cancer which opened a new revolution in the etiology treatment of cancer. Moreover, HPV infection is also the cause of multiple cancers in both women and men, including anogenital cancer (anal, vaginal, vulvar, and penial) or head and neck cancer (oropharynx, oral and laryngeal) [11]. Recent studies indicate that over 90% of cervical and anal cancers, over 70% of oropharynx cancers, about 70% of vulvar and vaginal cancers, together with more than 60% of penile cancers are related to HPV [12, 13].
Cervical cancer is the fourth most common cancer in women with 604,127 new cases and 341,831 deaths occurring worldwide in 2020 [14, 15]. The incidence and mortality have shown an obvious geographical imbalance between low-income and middle-income countries (LMICs) with high-income countries in cervical cancer patients. In LMICs, CC is the second most common cancer with an incidence rate of 18.8 per 100 000 women and a mortality rate of 12.4 per 100 000 women. In contrast, as a result of the availability of HPV prophylactic vaccines and standardized screening strategies, the incidence (11.3/100 000 women) and mortality (5.2/100000 women) of cervical cancer have decreased in high-income countries [14]. Vaccination and screening are effective in preventing cervical cancer, but they will impose a huge global economic burden. A systematic review has demonstrated that 106 million women have received at least one dose of HPV vaccine worldwide till 2014, but the HPV vaccination and standardized screening coverage in LMICs are still obviously low [16, 17].The world health organization (WHO) made a call for global action toward CC elimination in 2018, through vaccinating 90% of all girls under the age of fifteen, screening 70% of women at the age of 25, and treating 90% of precancerous lesions. The prediction simulation using the WHO Cervical Cancer Elimination Modelling Consortium (CCEMC) shows that the premature mortality rate of CC in 78 LMICs could be reduced by a third in the next 10 years. The WHO triple-intervention strategy would result in a 96.2% reduction by 2070, and 98.6% reduction by 2120. Famously, vaccination alone could reduce the mortality by 62.7% till 2070 and 89.5% till 2120. It is believed that with concerted global efforts, the incidence of cervical cancer in LMICs will be steadily reduced (Fig. 1) [18]. In 2019, the first domestic bivalent HPV vaccine was released and contributed to the HPV vaccination program in China [19]. This geographical distribution heterogeneity is therefore bound to become uniform gradually with the improvement of the global economic level and the implementation of prevention strategies.

Global map of epidemiological data for tertiary prevention of cervical cancer. A vaccination coverage rates and B cervical cancer screening rates by country in 2014. C incidence and D mortality age-standardized rates per 100,000 by region in 2020. The full-course coverage data among the total female population are illustrated. 78 low-income and lower-middle-income countries involved in the WHO cervical cancer elimination project are highlighted in red. Source: GLOBOCAN 2020
Heterogeneity of HPV infection types
The infection rate besides the infection site of different HPV types is heterogeneous across populations. Fifteen high-risk HPV (HR-HPV) types have been confirmed as carcinogenic viruses, as follows, 16,18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68, 73, and 82. Among them, the cumulative infection rate of HPV16 and 18 accounts for 79% of the squamous-cell carcinomas, and accounts for 95% of the squamous-cell carcinomas together with HPV45, 31, 33, 52, 58, and 35 [20, 21]. A meta-analysis collated data from 115,789 HPV-positive patients has been performed to analyze the distribution of HR-HPV. The percentage of 13 HR-HPV infection distributions under different disease states are demonstrated in Fig. 2, and include HPV16, 18, 31,33, 35, 39, 45, 51, 52, 56, 58, 59 and 68. Among them, HPV16, 18 and 45 infections dominate in invasive cervical cancer( ICC) (ICC: normal ratios 3.1, 1.9 and 1.1, respectively) [22].

Histogram chart of HR-HPV infection distributions by different disease states. Normal: disease-free state; CIN: cervical intraepithelial neoplasia; ICC: invasive cervical cancer
Another systematic meta-analysis collated data from 19 883 HIV-positive patients has been performed and analyzed the distribution of HR-HPV in 2017. Similarly, the ICC: normal ratios of HPV infections are 3.7 (HPV16), 2.5 (HPV18), and 2.6 (HPV45) respectively which is consistent with the conclusion in HIV-uninfected populations [23]. It is illustrated that HPV16, 18 and 45 positivity increase distinctly from normal cytology through squamous intraepithelial lesions to invasive cervical cancer which suggests that we should pay special attention to these types in cervical cancer screening.
On the other hand, in a large sample of healthy people screening, there is data to support a shift in the pre- and post-vaccine prevalence profile. HPV16, 18, 31, 52 and 58 were the most five common infection types in women with normal cytology in the pre-vaccine era [24]. However, infection rates of HPV52, 58, and 56 are increasing in the post-vaccine era [25, 26]. Whether bivalent and quadrivalent vaccines can provide cross-protection is controversial, but there is no doubt that the spectrum of HPV-associated squamous intraepithelial lesions and invasive cervical cancer will continue changing with the introduction of the 9-valent vaccine or even the 11-valent vaccine. Although cervical cancer is being treated earlier and earlier, it is still a constant battle against the ever-changing virus types.
Heterogeneity of anti-viral immunity
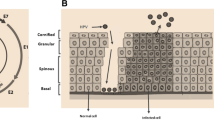
Upon HPV infection, the host cell immediately activates the innate and adaptive immunity to eliminate the virus [27]. Tumor heterogeneity of cervical cancer is reflected in the outcome of the battle between our immune system and virus invasion in the post infection microenvironment (PIM). HPV is undoubtedly one of the most important external factors mediating the heterogeneity of tumor development. HR-HPV type, duration of infection, virulence, human genomic instability and immune clearance will affect the tumorigenesis and development of carcinoma [28]. There are three outcomes of the battle between our immune system and HPV infection. Firstly, the virus is thoroughly cleared by our immune system. Secondly, the overwhelming majority of the virus is cleared, and only minority viruses that lie dormant can escape immunological recognition. Thirdly, the virus escapes immune recognition and integrates into the human genome, resulting in persistent infections and tumorigenesis [29, 30]. Fortunately, persistent high-risk HPV (HR-HPV) infection combined with oncogene genomic integration might lead development of normal cervical cells into intraepithelial neoplasia (CIN) or ICC in decades (Fig. 3).

Clinical outcomes of different anti-viral immune states after HPV infection. A HPVs are completely eliminated by the body’s immunity; B A majority of HPVs is eliminated, a small percentage of latent basal layer stem cells still exist; C HPVs induce immunosuppression, gene integration, CIN and carcinogenesis
PIM has been recognized as a complex and dynamic position with a collection of highly heterogenous cellular or molecular compounds, especially induced by the interaction between HPV-infected keratinocytes and immune cells. Specific cellular immune reactions and break down of immunosuppressive status are essential for effective virus clearance. Insufficient trafficking or maturation of Langerhans cells may lead to antigen-presenting disorder and CD8+ cytotoxic T lymphocyte (CTL) response impairment [31, 32]. Otherwise, the expression of MHC-I on the surface of keratinocytes have been down-regulated after HPV infection and recognition of CTLs will be avoided [33]. Except for CTLs, CD4+ T cells are essential in HPV clearance. An imbalance in T-helper 1(Th1) and Th2-type CD4+ T cells might be associated with immune dysregulation. Furthermore, the malfunction of NK cells is associated with immunosuppression [34].
Heterogeneity of genomic instability and HPV integration
Key characteristics of PIM include immunosuppressive state, oxidative stress response, extracellular matrix (ECM) remodeling, and metabolic reprogramming [35]. Oxidative stress could amplify inflammatory responses and result in accumulation of DNA damage, mutations or genome instability [36]. Expression of matrix metalloproteases is also increased and associated with ECM remodeling and precancerous lesion occurrence [37]. Once the immune microenvironment remodeling that promotes the persistence of HPV infection is established, genomic integration and cytopathic changes occur continuously.
As the overexpression of oncoproteins E6 and E7 in the HPV-infected keratinocytes, E6 disrupts p53 degradation and alteration of cell regulation, on the other hand, E7 induces retinoblastoma (pRb) degradation and promotes cell proliferation [38]. E6 and E7 may also induce genomic instability and lead to carcinogenesis by abrogating cell-cycle checkpoints [39]. Growing evidence suggests that chromosomal instability is also a driving force for the oncogenic transformation of cervical cancer. High chromosomal instability Hela cells exhibit a higher karyotype heterogeneity and are related to KRAS signaling regulation [40].
HPV is a small double-stranded DNA virus whose DNA fragments have the ability to integrate into the human genome. Associations between HPV integration and adjacent host genomic structural variation have been confirmed in HPV-positive cervical cancer cell lines. HPV16 integration has been detected firstly on chromosome 13q22 in SiHa cell lines in 1987 [41]. HPV 16, 18, and 33 viral integration has been detected in cervical squamous cell carcinomas by scientists as early as 1991 [42]. All integration events of the 13 HR-HPV subtypes have already been observed, and an unbalanced distribution of HR-HPV genotypes in cervical cancer has been detected. We have summarized six high-quality studies with HPV integration data of cervical cancer patients through next-generation sequencing (NGS) or whole-genome sequencing (WGS), and the proportion of integration events among different subtypes is analyzed. We can see that the integration of type 16 and 18 accounts for more than 80% of all samples. Other HR-HPVs are HPV45, 31, 33, 52, 58, 59, 39, 56, 68, 35 and 51 in a descending order of integration ratio (Fig. 4) [43,44,45,46,47,48]. We have summarized the high-frequency (more than 4 reported) disrupted genes by HPV integration and listed the hotspots, such as 3q28, 8q24, and 13q22. The top five reported genes are MACROD2, FHIT, POU5F1B, LRP1B and RAD51B (Table 1).

HPV integration normally breaks up the open reading frames of viral E1 and E2 genes which leads to the upregulation of E6 and E7 oncogenes [49]. Genomic instability, HPV integration and gain of telomerase at chromosome 3q26 appear to be strongly associated with genetic events in malignant transformation from CIN to invasive cervical carcinoma. In particular, chromosomal instability may precede genomic integration of oncogenic HPV, while increasing the human telomerase gene copy number occurs after integration as a termination product [50,51,52]. The integration hotspots are non-random and numerous microRNAs are located in the vicinity of integration hotspots and are influenced by the integrated HPV DNA. Highly homologous stretches of HPV16 viral gene E5 and L2 have been detected at the integration hotspots in independent patients which support themselves as quite important events in the integration process [53]. HPV E6E7 alternative transcripts have shown frequent isoforms in HPV16 or HPV18 positive cervical cancer [54]. Multiple frequent integration sites in human genome have been reported and verified through whole genome sequencing, high-throughput RNA, or chromosome conformation capture (Hi-C) sequencing, whereas the patterns of HPV integration in DNA and RNA samples differ significantly. For instance, DLG2, FHIT, HMGA2, KLF12, KLF5, LRP1B, LEPREL1, LINC00392, POU5F1B, and SEMA3D are DNA hotspots [41, 46]. In addition, CASC8, CASC21, ERBB2, RAD51B, RAP2B, TEX41, TP63, TTC6, MACROD2, MIPOL1, and MYC are hotspot genes in RNA samples [55]. DNA breakpoints are prone to an intron, in contrast, RNA breakpoints are prone to the region of EXON [56]. CCDC106 integration on chromosome 19 has been exhibited in altering local chromosome architecture and structure remodeling [57]. Attentionally, the changes in protein expression levels after HPV integration are inconsistent. FHIT and LRP1B are downregulated, while MYC and HMGA2 are elevated. Moreover, the fusion between HPV and human genome may have occurred by microhomology-mediated DNA repair pathways [46].
In terms of prognostic analysis, HPV16 positive status of the pelvic lymph nodes is a significant predictor of recurrent cervical cancer, while HPV16 integrated form is an unfavorable predictor of overall survival [58, 59]. HPV-DNA integration has been detected with association in carcinogenesis and recurrence free survival [60].HPV integration into the common fragile sites may be associated with distant metastasis [61]. Accurate detection of integration sites will continue with the improvement and combination of multidimensional technologies, such as nanopore sequencing and fluorescent in situ hybridization [62, 63]. Detection methods for viral integration sites are changing rapidly, and we believe that the blueprint for HPV integration will become clearer in the next decades.
Inferring heterogeneity with human genomics
Genetic intra-tumor heterogeneity acts as a key challenge in tumor evolution and management which affects patients’ outcomes [6, 64, 65]. The fundamental biological mechanisms underlying intra-tumor heterogeneity include genetic drift, selection, heritable variation, and environmental changes [66, 67]. Somatic mutation of FGFR3 has been identified in a large proportion of cervical cancer by Cappellen et al. as early as 1999 [68]. Nevertheless, at least three driver gene alterations are necessary to convert normal cells to malignant cells [69]. Over the past decades, multiple gene expression profiles and novel through-out sequencing studies have focused on capturing intra-tumor heterogeneity over time and space [70, 71]. Whole genome sequencing data among pan-cancer patients (including cervical cancer) has identified 95.1% subclonal expansions of 1705 tumors which verified the importance of intra-tumor heterogeneity [72]. Several scRNA-seq analyses have also been performed to study intra-tumor heterogeneity at the level of individual cells in cervical cancer. We summarize the intra-tumor heterogeneity of cervical cancer from genomic, transcriptomic and epigenetic alterations under different approaches.
Somatic genomic alterations in cervical cancer
The Catalogue Of Somatic Mutations In Cancer (COSMIC) is the world’s most comprehensive repository of human cancer somatic mutations [73]. Driver hotspots from COSMIC single-base substitution (SBS) mutational signatures are classified into four categories: Deamination, APOBEC, somatic hypermutation, and signature SBS39 [74]. The apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like (APOBEC) which converts cytosine to uracil during RNA editing and retrovirus restriction, has been confirmed in mediating pervasive mutagenesis in human cancers [75]. APOBEC-associated hotspots consist of one to two specific point mutations. In contrast, hotspots associated with somatic hypermutation are characterized by somatic single nucleotide variant (sSNV) clusters in promoter regions, which are clusters of variations in a single nucleotide without any limitations of frequency arisen in somatic cells. APOBEC mutagenesis pattern is associated with 34 common mutational hotspots across multiple cancers and has been identified as the predominant source of mutations in cervical cancers [74, 76]. The high-throughput genotyping platform has been used to interrogate cervical tumors and the consistently high mutation rates of PIK3CA have been confirmed. The APOBEC mutagenesis pattern is associated with nucleotide substitution in the E542K or E545K of PIK3CA, while the non-APOBEC mutagenesis pattern coexists at the same time [44].
The recognized mutated genes are ARID1A, CASP8, EP300, ERBB3, FBXW7, HLA-A, HLA-B, KRAS, MAPK1, NFE2L2, PIK3CA, PTEN, SHKBP1 and TGFBR2 in cervical cancer. It’s worth noting that over 70% of CCs exhibit genomic alterations in PI3K-MAPK and TGFβ signaling pathways [44, 77]. Novel significantly mutated genes have been discovered through deep RNA sequencing approaches and clustering of their mutant allele fraction variants. At least 20% of cervical cancers harbor somatic LKB1 mutations. Approximately 100% of tumors with these mutations harbored single nucleotide substitutions, identifiable monoallelic or biallelic deletions or multiplex ligation probe amplification (MLPA) [78]. Mutational sequencing has identified that 40% of 23 cervical cancer specimens harbored somatic mutations of NOL7, a tumor suppressor gene located on 6p23. Multiple CpG dinucleotides have been detected spanning the first exon or the 5’ untranslated region of NOL7, resulting in its inactivation [79].
There is heterogeneity in gene mutations among different pathological types. PIK3CA mutation rates keep consistent between adenocarcinomas and squamous cell carcinomas. The major mutations in squamous cell carcinomas include EP300, FBXW7, MAPK1, NFE2L2 and EGFR, while KRAS, ELF3, and CBFB in adenocarcinoma [80, 81]. The Cancer Genome Atlas (TCGA) Research Network has identified high frequencies of ARID1A, KRAS, and PTEN mutations in endometrial-like cervical cancers [48]. Mutations in PIK3CA, KRAS, and TP53 have also been detected most commonly in small cell cervical cancer using next generation sequencing [82].
Differential gene expressions in cervical cancer
To discover transcriptomic intra-tumor heterogeneity, previous studies have investigated differential transcript gene expressions between normal and cervical cancer tissues through microarray technologies [83,84,85,86,87,88]. At the RNA level, gene expressions determined by the expression profiling microarray are detected by reverse transcription-polymerase chain reaction (RT-PCR). While at the protein level, the expressions of specific proteins are often described in immunohistochemical (IHC) staining. Multiple-gene transcript signature with differential expressions by cDNA microarray could be used for molecular classification between stage IB and IIB and prediction of response to radiotherapy for advanced cervical cancer [85, 86, 89]. Differential expressions of CDKN2A and PTGES have been identified in invasive cervical cancer versus normal keratinocytes through oligonucleotide microarrays and confirmed through immunohistochemical staining [90]. Apoptotic genes BCL2, BCL2l1, and BIRC2 have been identified as upregulated in late-stage cervical cancer compared to early-stage cases [91]. DPP4, EDN3, FGF14, TAC1 and WNT16 have been indicated simultaneously downregulated and hypermethylated in cervical cancer [92]. Message RNA expression levels of RhoB and STMN1 have been validated associated with overall survival in cervical cancer [93]. A positive correlation has been observed between gene expression of HPV E6/E7 oncogenes and UHMK1 [94].
Expression profiling has been replaced gradually by more accurate sequencing techniques and the search for differential expressed genes (DEGs) in tumors continues. Three DEGs, including RDH12, UBD, and SAA1 have been screened with correlation to tumor size, lymphatic metastasis, and depth of cervical invasion in cervical squamous cell carcinoma through RNA sequencing [95]. Upregulated expression of AKT3 in cervical cancer has been related to resistance to cisplatin [96]. Transcriptome sequencing in HPV16 positive cervical cancer tissues has identified 140 DEGs enriched in cell cycle and DNA repair [97].
Heterogeneity analyzed by single-cell RNA sequencing approaches
Single-cell sequencing is a promising systematic and comprehensive approach to delineating subclone associations and intratumor heterogeneity. Conclusions of single-cell sequencing researches have provided a deeper understanding of specific mechanisms leading to heterogeneity in recent years. The landscape of heterogeneity within 22 cancer cell lines has identified twelve recurrent heterogeneous programs (RHPs) even without the native tumor microenvironment. These RHPs are associated with cell cycle, stress responses, epithelial-mesenchymal transition, and protein metabolism [98]. Focusing on cervical cancer, single-cell RNA sequencing data of 20,938 cells have divided tumor cells into four subpopulations with distinct signature genes and prognoses. Specifically, the cells in the first subpopulation are enriched in immune regulation signaling pathways, such as the ErbB signaling pathway; the cells in the third subpopulation are suggested with high proliferative activity because of their high expression of MKI67, CCNB1 and TOP2A genes. The last two subpopulations are regarded as the original cancer cells and the terminal cancer cells respectively, one with over-expressed stem-related genes SOX2 and ALDHA1 and the other with high expressions of genes enriched in steroid biosynthesis, mismatch repair and peroxisome pathways [99]. Another single-cell RNA sequencing data of 24,371 cells aiming to comprehensively analyze chemotherapy resistant cervical cancer cells have clustered cells into nine subpopulations. Differentially expressed genes enriched in the PI3K/AKT pathway are involved in chemotherapy resistance [100]. The main limitations of microarray and sequencing technologies are detecting variations at the DNA or RNA level rather than the protein level. Validation studies in conjunction with proteomics are essential. Cellular heterogeneity is being characterized in cervical cancer with the advent of single-cell genomics which may provide more accurate information on cancer characteristics, prognostic prediction, and treatment decision selection.
Epigenetic landscape in cervical cancer
Tumor development and drug resistance are sometimes driven by potential gene amplification and deletion, not only somatic genomic alterations but also copy number amplifications, histone modification, and DNA methylation. A large-scale genomic study, including genomic, transcriptomic, and epigenomic landscapes of 118 Ugandan cervical cancer patients has been performed. DNA methylation, histone marks, and gene expression dysregulation differ between A9 and A7 HPV clades. Clade A7 corresponded to a less differentiated phenotype of cervical cancer and lead to a poorer prognosis. Changes in histone modification are associated with HPV integration [101]. Another comprehensive genomic analysis including whole exome sequencing, copy number and methylation analysis of 228 primary cervical cancers has revealed amplifications in immune checkpoint genes PD-L1 and PD-L2, together with lapatinib associated gene BCAR4 [102]. A C-score model according to the chromosomal-arm-level copy number alterations (CNAs) changes of 1q, 2q, 3p, and 7q has been validated to distinguish ICC from normal tissues with 100% sensitivity and specificity [103].
Deregulation of micro-RNA (miRNA), long non-coding RNA (lncRNA) and circular RNA (circRNA) have also been revealed in cervical cancer patients in recent researches. Specifically, miRNAs are small non-coding RNAs which can regulate gene expression through binding to DNA or mRNA [104]. While lncRNAs are long non-coding RNAs which can regulate gene transcription mediated by interacting with chromatin-modifying complexes and miRNAs [105]. CircRNAs are also small non-coding RNAs playing big parts in post-transcription and participate in genetic expression [106]. A type of endogenous RNA, specifically, competing endogenous RNAs (ceRNAs) have been identified to influent the target genes by miRNA and participate in cancer regulation process ultimately [107]. The ceRNA-miRNA-mRNA regulatory axis is gradually explored in cervical cancer research. Both lncRNAs and circRNAs may function as sponges or ceRNAs of miRNAs to regulate mRNA expression [108]. A recent review summarized the reciprocal regulation role of miRNAs, lncRNAs and circRNAs in CC patients. The miRNAs are divided into “oncogenic” miRNAs (miR-10a, miR-19, miR-21, and miR-146a et al.) and “tumor suppressive” miRNAs (miR-29a, miR-214, miR-218, and miR-372 et al.) [109]. Around 14 lncRNAs have shown to be altered and affected important metabolic pathways such as STAT3, wnt/β-catenin, PI3K/AKT, and Notch signaling in cervical cancer [110]. LncRNA XLOC_006390 can serve as a ceRNA and has been verified reversely regulating the expression of miR-331-3p and miR-338-3p, and facilitating tumorigenesis or metastasis in cervical cancer [111]. CircRNA_VPRBP regulates miR-93-5p/FRMD6 axis which lead to inhibited proliferation, migration and invasion of cervical cancer cells [112]. Furthermore, circRNA hsa_circ_0000515 acts as a miR-326 sponge, has been demonstrated to promote cervical cancer progression through upregulated ELK1 expression [113]. These findings might enumerate the regulatory mechanisms of epigenetics in the development of cervical cancer. However, the complexity interaction among diverse non-coding RNAs shows great heterogeneity, which still needs to be further verified.
Inferring heterogeneity with therapeutic diversity
HPV screening and classic three-step diagnostic criteria have been quite normalized and widely used worldwide in the detection of early-stage cervical cancer. According to clinical guidelines, standard surgical treatment is the first-line recommended with satisfactory effect. Results of a completed randomized controlled phase III trial (NCT00002536) have shown no significant differences in stage IB cervical cancer patients with or without neoadjuvant chemotherapy [114]. For locally advanced cervical cancer patients, adjuvant chemotherapy and radiotherapy can improve patients’ outcomes but with increasing controllable toxicity [115]. Nevertheless, a minority of patients have to face distant metastasis, recurrent or persistent cancer and drug resistance. Combination of multiple-agent chemotherapy and biotherapy (Bevacizumab) may be associated with death reduction and prolonged PFS in recurrent/persistent cervical cancer patients (Table 2) [116]. Tumor heterogeneity is undoubtedly an important factor leading to the reverse therapeutic effects among individuals. Diverse treatment is a double-edged sword because it serves as an external factor to promote persistent adaption and selection in the tumor microenvironment. We suggest that cell subclones are derived in response to different therapeutic stimuli and determine patient outcomes (Fig. 5).

Model of clonal progression of cervical cancer. Normal cervical cells may harbor genomic alterations and HPV integration after HPV infection. Some cells regress to normal spontaneously, while others round into clonally invasive carcinoma cells. Overwhelming majority cancer cells are removed or killed during conventional surgery and chemoradiotherapy. A few dormant or new subclones develop into recurrent, persist or metastatic cancer lesions. Systemic therapies (chemotherapy, radiotherapy, biotherapy and immunological therapy) can induce intrinsic or adapted resistant subclones. Resistant subclones contribute to uncontrolled disease and treatment failure
Heterogeneity in terms of chemotherapy resistance
Cervical cancer chemotherapy can be divided into neoadjuvant chemotherapy (NACT) aiming to shrink the mass to facilitate operation, adjuvant chemotherapy or concurrent chemoradiotherapy (CCRT) as maintenance after surgical treatment or standard treatment for locally advanced patients, and palliative chemotherapy for relieving symptoms, pain or prolonging survival in recurrent or metastatic patients [117]. The majority of these patients will receive more than two combination treatments. Most studies on drug resistance have been limited to in vitro experiments, and few studies have been validated in drug-resistant populations. The molecular mechanism of chemotherapeutic resistance has not been fully understood but could be speculated via blocking DNA damage repair, oxidative stress, autophagy, and apoptosis signaling pathways. Both coding and non-coding RNAs participate in chemo-resistance. Non-coding RNAs, including miRNA, lncRNA, and circRNA, are potential therapeutic targets in cancer treatment development. However, its role in the field of drug resistance of cervical cancer remains to be further explored. Genomic rearrangements may occur by selecting effects from chemoradiotherapy which exhibits genetic intra-tumor heterogeneity in advanced cervical cancers. Platinum-paclitaxel combination chemotherapy is recommended as the first-line chemotherapy drugs in multiple solid cancers and we explain their mechanisms of chemotherapy resistance individually [118].
Cisplatin has been used in most studies of platinum resistance. The mechanisms underlying cisplatin resistance in CC are respectively DNA damage repair increase, apoptosis inactivation, epithelial-mesenchymal transition activation, or DNA methylation alteration [119]. For instance, the upregulated expression of COX-2 has been assessed with neoadjuvant cisplatin-based resistance and unfavorable overall survival in locally advanced CC patients [120]. Cisplatin induces chemotherapy resistance of well-differentiated cell line Caski cells by upregulating Src family kinase and interaction with EphA4 through the reactive oxygen species pathway [121]. Inhibiting endogenous EZH2 expression has shown decreased cell metastasis, reversed cisplatin resistance in HeLa cells, and increased antitumor effects in nude mice. Interfering EZH2 expression has been identified correlated with Dicer overexpressed or regulated H3K27 methylation level, which exhibit antitumor activities by interfering the progression of miRNA transcription, and cell cycle and promote cell apoptosis [122]. MALAT1 and PSAT1 could induce resistance in SiHa cells through PI3K/Akt pathway [123, 124].GAS5 could be regulated by P-STAT3 and affect resistance via miR-21/PDCD4 axis [125]. EDC4 could interact with RPA by alleviating DNA damage in cisplatin-resistant HeLa and SiHa cells [126]. IPO4-CEBPD-PRKDC axis is associated with chemoresistance by inhibiting PRKDC-driven DNA damage repair [127]. In addition, an increasing amount of noncoding RNAs have been confirmed and summarized with association to cisplatin resistance [128, 129]. For example, LncRNA HNF1A-AS1 could affect resistance by regulating miR-34b and promoting TUFT1 expression [130]. LncRNA OTUD6B-AS1 could mediate decreased regulation of miR-206 and increased expression of CCND2 [131]. LncRNA NNT-AS1 could improve chemoresistance via the miR-186/HMGB1 axis [132].
Combination chemotherapy with cisplatin and paclitaxel is a standard treatment in recurrent or advanced cervical cancer with an overall response rate of 29%–67% [133, 134]. Meanwhile, confirmed gain of 3q and loss of 11q chromosomes are early events in cancer progression. Subpopulations with differential responses to chemoradiotherapy may be selected into a single intrinsically resistant subpopulation after five weeks of the therapy [135]. Knockdown of Linc00511 could reduce paclitaxel resistance by regulating Bcl-2, MMP-2, MMP-9, MRP1, and P-GP expressions in HeLa cells [136]. Overexpressed miR-214 under paclitaxel treatment could cause an increase in PARP and a decline in PI-3 kinase/Akt levels [137]. Circular RNA CircMYBL2 could enhance paclitaxel resistance by upregulating EGFR mediated by microRNA-665 in vitro and promoting tumor growth in vivo [138].
Heterogeneity in terms of radiotherapy resistance
Radiotherapy for cervical cancer is suitable for locally advanced and recurrent patients or other patients who can’t tolerate surgery. The majority of these patients own a worse prognosis due to advanced FIGO stage. The CCRT is the recommended treatment for advanced cervical cancer compared with radiotherapy alone, because CCRT increases patients’ local control rates and improves prognosis [139]. Integrated bioinformatics analysis on RNA sequencing has identified ten potential biomarkers related to radiotherapy resistance in cervical cancer. The results have indicated overexpression in tumor immune process pathways, including cellular defense response, negative regulation of the immunity, T cell and neutrophil activation, regulation of antigen presentation, and peptidyl-tyrosine autophosphorylation [140]. Other biomarkers, such as HIF-1 could enhance hypoxia-induced radio-resistance via targeting NDRG2 [141]. Overexpressed HOTAIR could promote HIF-1a and lead to radio-resistance in mice [142]. CD147 could induce resistance by regulating the percentage of G2/M phase cells and DNA double-strand breaks repair [143]. RhoC-ROCK2 involved DNA repair pathway is necessary for the radio-resistance phenotype in tumor cells [144]. SEPT9 could affect resistance by interacting with the HMGB1-RB axis and mediating miR-375 [145]. Increased expression of HMGB3 correlated with hTERT could predict poor response to radiotherapy, advanced stage and worse outcome [146]. USP21 is overexpressed in radio-resistant patients and could activate the FOXM1/Hippo signaling pathway [147]. Four specific miRNAs (miR-630, miR-1246, miR-1290, and miR-3138) could promote radio-resistance in vitro [148]. MiR-125 could modulate resistance through the downregulation of CDKN1A [149]. LncRNA UCA1 could promote radio-resistance associated glycolysis in SiHa and HeLa cells via HK2/glycolytic pathway [150]. LncRNA SNHG6 could enhance radio-resistant and promote cell growth via STYX/miR-485-3p axis [151]. Tumor radiotherapy has a certain impact on the TME, for instance, the generation of cancer-associated fibroblasts or macrophages [152, 153].
Heterogeneity in terms of immunotherapy resistance
After failing platinum-based chemotherapy, only about 10% of patients are responsive to additional cytotoxic agents. Immunotherapy of solid tumors is the research hotspot at present aiming to overcome immune suppression in TME and enhance tumor targeted immune attack. The main directions of immunotherapy include immune checkpoint inhibitors, therapeutic antibodies, therapeutic vaccines, cell therapy and small molecule inhibitors. Here we focus on the use of immune checkpoint inhibitors and therapeutic vaccines about the heterogeneity of cervical cancer.
Professors James P Allison and Tasuku Honjo won the 2018 Nobel Prize in Physiology or Medicine for discovering CTLA-4 and PD-1 as immune checkpoints and laying the foundation for tumor immunotherapy. The US Food and Drug Administration has already approved pembrolizumab for advanced cervical cancer patients with positive PD-L1. Clinical trials about the efficacy and safety of Pembrolizumab in advanced cervical cancer have been verified. Objective response rate (ORR) refers to the proportion of patients required for the reduction of the tumor to reach the expected value and to continue to the minimum expected time. ORR is commonly to be seen in evaluating the drug response in cancer patients undergoing clinical trials. The ORR of pembrolizumab in these patients has been increased to 14.6% [154]. Results of the phase III clinical trial of KEYNOTE-826 have expanded the indication for combined immunotherapy for persistent, recurrent or metastatic cervical cancer [155]. Results of the phase I/II clinical trial of CheckMate 358 (nivolumab) have shown an ORR of 26.3% with regardless of PD-L1 expression [156]. Three current trials of combining immunotherapy with chemotherapy for cervical cancer involved angiogenesis inhibitors and ICI combination therapy without conclusions (NCT03912415, NCT03635567, and NCT03556839) [157]. We summarized ongoing phase III clinical trials in cervical cancer and illustrated the effect targets for these therapies (Table 3 and Fig. 6). It can be seen from the current ongoing phase III clinical trials in cervical cancer that PD-1 inhibitors include Pembrolizumab, Camrelizumab, Cemiplimab, Prolgolimab (BCD-100), and QL-1604, while PD-L1 inhibitors include Durvalumab and Atezoliznmab. Newly developed dual targeted drugs AK104 (PD-1 and CTLA-4 inhibitors) and SHR-1701 (PD-L1 and TGFβ inhibitors) have already been used in phase III clinical trials. The sensitivity of immunotherapy mainly depends on the heterogeneity of responses between tumor cells, immune-infiltrating cells, and other stroma cells in the TME. With the further development of scientific research, the refinement of immunotherapy indications marks the arrival of the era of precision therapy.

The potential resistance mechanisms and currently on-going phase III clinical trials’ agents in cervical cancer. The drug resistance in tumor cells showed as up-regulation of immunosuppression, cell proliferation, angiogenesis, cell cycle arrest, DNA repair and down-regulation of apoptosis. T cell anti-tumor immunity may be suppressed through down-regulation of cell cycle progression, IL-2 production, T-cell activation, effector-cell development and up-regulation of apoptosis. Agents of clinical trials have focused on novel immune checkpoint inhibitors including PD-1, CTLA4, PD-L1 and TGFβ
Novel immune checkpoints, for instance, TIGIT (T cell immune receptor with Ig and ITIM domains) have been utilized combined with anti-PD-1 antibody in recurrent or metastatic cervical cancer (NCT04693234). The application of immune checkpoint inhibitors is limited by the heterogeneity of checkpoint expression on tumor cell surface and immune-activated state in TME. Decreased tumor associated lymphocytes and retained HPV E6/E7 gene expressions may promote treatment resistance during chemoradiation therapy in locally advanced cervical cancer patients [158]. Oncogenic E5, E6, and E7 proteins encoded by HR-HPV, especially HPV16 and 18, are implicated in the PD1/PD-L1 pathway leading to increased PD-L1 expression [159,160,161]. B cells are activated by radiation combined with PD-1 blockade and could improve overall survival in HPV-associated squamous cell carcinomas patients [162]. LSD1 inhibitor combined with anti-CD47/PD-L1 monoclonal antibodies could more effectively inhibit tumor growth in a subcutaneous xenograft model because of increasing the expressions of CD47 and PD-L1 [163]. Other driver genes (PI3KCA, PI3KCB, DVL3, WWTR1 and ERBB2) in regulating immune response or immune cell infiltration are with application prospect [164]. Three single-nucleotide polymorphisms (SNPs), specifically PAX8, CLPTM1L, and HLA genes, are replicated in cervical cancer patients and are associated with cervical carcinogenesis through disruption in apoptotic and immune response pathways [165, 166].
Therapeutic vaccines have also shown some success in patients with advanced cervical cancer. An alphavirus-based treatment vaccine combined with sunitinib and irradiation could elicit superior antitumor effects [167]. HPV recombinant vaccine prime-boost could enhance CD8+ T cell mediated tumor cytotoxicity [168]. PD-1 blockade combined with intra-tumoral therapeutic vaccination could elicit HPV16-associated tumor regression in a murine model [169]. The combined application of cervical cancer therapeutic vaccine and immunotherapy has become the general trend at present [170].
Inferring heterogeneity with histological diversity
The histological diversity of cervical cancer is also a manifestation of tumor heterogeneity. There were significant differences in treatment sensitivity and prognosis among different histological types. In the previous paragraphs, we have mainly discussed the characteristics of cervical squamous cell carcinoma, while in this section we will focus on the molecular and clinical characteristics of cervical cancer stem cells, cervical adenocarcinoma cells, and cervical neuroendocrine cell subtypes.
Heterogeneity of cervical cancer stem cell
The clonal evolution model and the cancer stem cell (CSC) model have been used to illustrate intra-tumor heterogeneity. In the first model, stochastic mutations in individual tumor cells form in the tumor microenvironment, the superior sub-clonal cells dominate and proliferate under adaptation and selection [171]. Another model highlights the cellular plasticity and mutational differentiation hierarchy formation generated by CSC clones [172, 173]. We attempt to interpret cervical cancer heterogeneity by describing the cell surface biomarkers, molecular mechanism of stem cell regulation and differences in cytological behavior as follows.
Cervical cancer stem-like cells (CCSC) with an expression pattern of CD44 ( +)CD24(-) surface biomarkers have been isolated from HeLa and SiHa cell lines which present higher capabilities in cell growth, self-renew, chemotherapeutic drug and radiation therapy resistance [174, 175]. Another prolonged Trichostatin A-selected HeLa cell expressing Sox2( +)Oct4( +)Nanog( +) markers display enhanced migration, invasion, and malignancy abilities both in vitro and in vivo, which can also be regulated by STAT3 [176,177,178]. Ubiquitin B has been confirmed as a key gene in the maintenance of Sox2( +)Oct4( +)Nanog( +) CCSC [177]. Hiwi and Gremlin 1 can be regarded as cervical CSC markers because the increased gene expressions facilitate in vitro tumor sphere formation and in vivo tumorigenicity [179, 180]. The extended phenotype of CCSC has been determined with CK-17, p63 + , All + , CD49f + and higher Aldehyde dehydrogenase activity [181]. Besides, the Wnt/beta-catenin pathway is essential to maintain tumorigenicity by microRNA-135a induced CD133( +) CCSC and CCSC related transcription factor levels promoted by LGR5. Wnt3a stimulation may increase tumor sphere size and self-renew [182, 183]. Cancer is a result of uncontrolled cell growth caused by mutations or epigenetic alterations, while cancer stem cell heterogeneity contributes to the whole process of tumorigenesis, recurrence, metastasis and treatment resistance.
Heterogeneity of cervical adenocarcinoma
Cervical adenocarcinomas comprise approximately 25% of cervical cancer in the USA with higher histological heterogeneity compared to squamous cell carcinoma [184]. The World Health Organization (WHO) classification and a more innovative International Endocervical Criteria and Classification (IECC) are commonly recognized classification criteria [185, 186]. The traditional WHO 2014 system divides cervical adenocarcinomas into serous, mucinous, endometrioid, clear cell and some other types based on pathological features. The IECC 2018 system attempts to subdivide adenocarcinomas into HPV-associated (HPVA) and non-HPV-associated (NHPVA) categories [187]. NHPVAs, in particular gastric type is significantly associated with age, horizontal extent, invasive depth or lympho-vascular invasion, advanced stage, worse disease-free survival (DFS) or disease-specific survival (DSS). Among the HPVAs, invasive stratified mucin producing carcinoma subtypes have shown worse DFS and DSS [188]. According to the revised WHO classification 2020, 92.7% of HPVAs can be recognized by the presence of luminal mitoses and apoptosis in addition to mucinous adenocarcinomas [189, 190]. Distinct molecular profiles have been demonstrated between SCC and adenocarcinoma as mentioned above, which suggests that more tailored treatment strategies are necessary [81]. Gastric-type cervical adenocarcinoma has been detected with somatic mutations in TP53, KRAS, CDKN2A, and STK11. Prevalent mutations of PIK3CA and PTEN enriched in the PI3K/Akt/mTOR signaling pathway has also been identified [191]. Potentially driven mutations have been identified in BRAF, ERBB2 and ERBB3. Copy-number aberrations (CNAs) are deletions or expansions of chromosomes/genes in somatic cells. Low levels of CNAs without recurrent amplifications or homozygous deletions are also confirmed [192]. Further similarities and differences genetic heterogeneity between HPVA and HPV-positive squamous cell carcinoma remain to be further studied.
Heterogeneity of other rare histological cervical cancer
Neuroendocrine carcinoma of the cervix (NECC) is a variant of CC with accounts for 1–1.5% [193]. A large meta-analysis with 3538 NECC cases has identified a mean recurrence-free survival of 16 months and overall survival of 40 months [194]. The small cell cervical carcinoma (SCCC) is the most common type of NECC with median overall survival ranging between 10–13 months in advanced SCCC [133]. Adjuvant chemotherapy or chemoradiation is associated with higher five-year survival in 188 SCCC patients [195]. FIGO stage is the unique prognostic factor impacting both overall survival and DFS in a multiple retrospective study with 93 SCCC patients [196]. SCCC is specifically associated with HPV18 infection and its genetic alterations are regulated through PI3K/AKT/mTOR, MAPK, and TP53/BRCA pathways [197]. Driven mutation genes KRAS, PIK3CA, IRS2, SOX2 and homogeneous recombination repair genes are potential therapeutic targets [198].
Conclusions and future perspectives
HPV-associated cervical cancer is a kind of heterogeneous malignant tumor from many perspectives, and its treatment in the advanced stage is extremely difficult. HPV and drug therapy are two extrinsic factors that are closely related to the heterogeneity of cervical cancer. Meanwhile, an in-depth understanding of tumor heterogeneity is a critical issue in developing precision treatment and screening strategies. Our understanding of the molecular and phenotypic heterogeneity in cervical cancer has improved and benefited from the development of deep sequencing and single cell sequencing technology. Nevertheless, it will take time to get breakthrough results on the heterogeneity of tumor microenvironment and treatment responses in advanced cancer. The integration of genomic, transcriptomic and epigenetic information that captures intra-tumoral heterogeneity will reveal the panoramic view of tumor regulatory mechanisms and will promote breakthroughs in precision medicine.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Abbreviations
- CC:
-
Cervical cancer
- CCEMC:
-
Cervical Cancer Elimination Modelling Consortium
- CCRT:
-
Concurrent chemotherapy
- CCSC:
-
Cervical cancer stem-like cell
- ceRNA:
-
Competing endogenous RNA
- CIN:
-
Intraepithelial neoplasia
- circRNA:
-
Circular RNA
- CNA:
-
Copy number alteration
- COSMIC:
-
Catalogue Of Somatic Mutations In Cancer
- CSC:
-
Cancer stem cell
- CTL:
-
Cytotoxic T lymphocyte
- DEG:
-
Differential expressed gene
- DFS:
-
Disease-free survival
- DSS:
-
Disease-specific survival
- ECM:
-
Extracellular matrix
- FIGO:
-
International Federation of Gynecology and Obstetrics
- HPV:
-
Human papillomavirus
- HPVA:
-
HPV-associated
- HR-HPV:
-
High-risk HPV
- ICC:
-
Invasive cervical cancer
- IECC:
-
International Endocervical Criteria and Classification
- IHC:
-
Immunohistochemical
- LMICs:
-
Low-income and middle-income countries
- lncRNA:
-
Long non-coding RNA
- miRNA:
-
Micro-RNA
- MLPA:
-
Multiplex ligation probe amplification
- NACT:
-
Neoadjuvant chemotherapy
- NECC:
-
Neuroendocrine carcinoma of the cervix
- NGS:
-
Next-generation sequencing
- NHPVA:
-
Non-HPV-associated
- OPC:
-
Oropharyngeal cancer
- ORR:
-
Objective response rate
- PD-L1:
-
Programmed death-ligand 1
- PD-L2:
-
Programmed death-ligand 2
- PIM:
-
Post infection microenvironment
- RHP:
-
Recurrent heterogeneous program
- RT-PCR:
-
Reverse transcription-polymerase chain reaction
- SBS:
-
Single-base substitution
- SCC:
-
Squamous cell carcinoma
- SCCC:
-
Small cell cervical carcinoma
- scRNA-seq:
-
Single-cell RNA sequencing
- sSNV:
-
Somatic single nucleotide variant
- TAM:
-
Tumor-related macrophage
- Th1:
-
T-helper 1
- TIGIT:
-
T cell immune receptor with Ig and ITIM domains
- TME:
-
Tumour microenvironment
- WGS:
-
Whole-genome sequencing
- WHO:
-
Word Health Organization
References
Pett M, Coleman N. Integration of high-risk human papillomavirus: a key event in cervical carcinogenesis? J Pathol. 2007;212(4):356–67.
Castellsague X. Natural history and epidemiology of HPV infection and cervical cancer. Gynecol Oncol. 2008;110(3 Suppl 2):S4-7.
Marusyk A, Janiszewska M, Polyak K. Intratumor Heterogeneity: The Rosetta Stone of Therapy Resistance. Cancer Cell. 2020;37(4):471–84.
Cook OF. The Mendelian Inheritance of Mutations. Science. 1908;28(707):86–8.
Williams MJ, Werner B, Barnes CP, Graham TA, Sottoriva A. Identification of neutral tumor evolution across cancer types. Nat Genet. 2016;48(3):238–44.
McGranahan N, Swanton C. Biological and therapeutic impact of intratumor heterogeneity in cancer evolution. Cancer Cell. 2015;27(1):15–26.
Alizadeh AA, Aranda V, Bardelli A, Blanpain C, Bock C, Borowski C, Caldas C, Califano A, Doherty M, Elsner M, et al. Toward understanding and exploiting tumor heterogeneity. Nat Med. 2015;21(8):846–53.
Almendro V, Marusyk A, Polyak K. Cellular heterogeneity and molecular evolution in cancer. Annu Rev Pathol. 2013;8:277–302.
Fan J, Slowikowski K, Zhang F. Single-cell transcriptomics in cancer: computational challenges and opportunities. Exp Mol Med. 2020;52(9):1452–65.
Muller S, Diaz A. Single-Cell mRNA Sequencing in Cancer Research: Integrating the Genomic Fingerprint. Front Genet. 2017;8:73.
Isayeva T, Li Y, Maswahu D, Brandwein-Gensler M. Human papillomavirus in non-oropharyngeal head and neck cancers: a systematic literature review. Head Neck Pathol. 2012;6 Suppl 1(Suppl 1):S104-120.
Szymonowicz KA, Chen J. Biological and clinical aspects of HPV-related cancers. Cancer Biol Med. 2020;17(4):864–78.
Johnson DE, Burtness B, Leemans CR, Lui VWY, Bauman JE, Grandis JR. Head and neck squamous cell carcinoma. Nat Rev Dis Primers. 2020;6(1):92.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–49.
Arbyn M, Weiderpass E, Bruni L, de Sanjose S, Saraiya M, Ferlay J, Bray F. Estimates of incidence and mortality of cervical cancer in 2018: a worldwide analysis. Lancet Glob Health. 2020;8(2):e191–203.
Bruni L, Diaz M, Barrionuevo-Rosas L, Herrero R, Bray F, Bosch FX, de Sanjose S, Castellsague X. Global estimates of human papillomavirus vaccination coverage by region and income level: a pooled analysis. Lancet Glob Health. 2016;4(7):e453-463.
Simms KT, Steinberg J, Caruana M, Smith MA, Lew JB, Soerjomataram I, Castle PE, Bray F, Canfell K. Impact of scaled up human papillomavirus vaccination and cervical screening and the potential for global elimination of cervical cancer in 181 countries, 2020–99: a modelling study. Lancet Oncol. 2019;20(3):394–407.
Canfell K, Kim JJ, Brisson M, Keane A, Simms KT, Caruana M, Burger EA, Martin D, Nguyen DTN, Benard E, et al. Mortality impact of achieving WHO cervical cancer elimination targets: a comparative modelling analysis in 78 low-income and lower-middle-income countries. Lancet. 2020;395(10224):591–603.
Zou Z, Fairley CK, Ong JJ, Hocking J, Canfell K, Ma X, Chow EPF, Xu X, Zhang L, Zhuang G. Domestic HPV vaccine price and economic returns for cervical cancer prevention in China: a cost-effectiveness analysis. Lancet Glob Health. 2020;8(10):e1335–44.
Munoz N, Bosch FX, de Sanjose S, Herrero R, Castellsague X, Shah KV, Snijders PJ, Meijer CJ. International Agency for Research on Cancer Multicenter Cervical Cancer Study G: Epidemiologic classification of human papillomavirus types associated with cervical cancer. N Engl J Med. 2003;348(6):518–27.
Lagheden C, Eklund C, Lamin H, Kleppe SN, Lei J, Elfstrom KM, Sundstrom K, Andrae B, Sparen P, Dillner J. Nationwide comprehensive human papillomavirus (HPV) genotyping of invasive cervical cancer. Br J Cancer. 2018;118(10):1377–81.
Guan P, Howell-Jones R, Li N, Bruni L, de Sanjose S, Franceschi S, Clifford GM. Human papillomavirus types in 115,789 HPV-positive women: a meta-analysis from cervical infection to cancer. Int J Cancer. 2012;131(10):2349–59.
Clifford GM, Tully S, Franceschi S. Carcinogenicity of Human Papillomavirus (HPV) Types in HIV-Positive Women: A Meta-Analysis From HPV Infection to Cervical Cancer. Clin Infect Dis. 2017;64(9):1228–35.
Bruni L, Diaz M, Castellsague X, Ferrer E, Bosch FX, de Sanjose S. Cervical human papillomavirus prevalence in 5 continents: meta-analysis of 1 million women with normal cytological findings. J Infect Dis. 2010;202(12):1789–99.
Soderlund-Strand A, Uhnoo I, Dillner J. Change in population prevalences of human papillomavirus after initiation of vaccination: the high-throughput HPV monitoring study. Cancer Epidemiol Biomarkers Prev. 2014;23(12):2757–64.
Sekine M, Yamaguchi M, Kudo R, J B Hanley S, Hara M, Adachi S, Ueda Y, Miyagi E, Ikeda S, Yagi A, et al. Epidemiologic Profile of Type-Specific Human Papillomavirus Infection after Initiation of HPV Vaccination. Vaccines (Basel). 2020;8(3):425.
Woodman CB, Collins SI, Young LS. The natural history of cervical HPV infection: unresolved issues. Nat Rev Cancer. 2007;7(1):11–22.
Da Silva MLR, De Albuquerque B, Allyrio T, De Almeida VD, Cobucci RNO, Bezerra FL, Andrade VS, Lanza DCF, De Azevedo JCV, De Araujo JMG, et al. The role of HPV-induced epigenetic changes in cervical carcinogenesis (Review). Biomed Rep. 2021;15(1):60.
Roden RBS, Stern PL. Opportunities and challenges for human papillomavirus vaccination in cancer. Nat Rev Cancer. 2018;18(4):240–54.
McBride AA. Human papillomaviruses: diversity, infection and host interactions. Nat Rev Microbiol. 2022;20(2):95–108.
Jiang B, Xue M. Correlation of E6 and E7 levels in high-risk HPV16 type cervical lesions with CCL20 and Langerhans cells. Genet Mol Res. 2015;14(3):10473–81.
Maskey N, Thapa N, Maharjan M, Shrestha G, Maharjan N, Cai H, Liu S. Infiltrating CD4 and CD8 lymphocytes in HPV infected uterine cervical milieu. Cancer Manag Res. 2019;11:7647–55.
Morrison BJ, Steel JC, Morris JC. Reduction of MHC-I expression limits T-lymphocyte-mediated killing of Cancer-initiating cells. BMC Cancer. 2018;18(1):469.
Gray JI, Westerhof LM, MacLeod MKL. The roles of resident, central and effector memory CD4 T-cells in protective immunity following infection or vaccination. Immunology. 2018;154(4):574–81.
Yuan Y, Cai X, Shen F, Ma F. HPV post-infection microenvironment and cervical cancer. Cancer Lett. 2021;497:243–54.
Kawanishi S, Ohnishi S, Ma N, Hiraku Y, Murata M. Crosstalk between DNA Damage and Inflammation in the Multiple Steps of Carcinogenesis. Int J Mol Sci. 2017;18(8):1808.
Zhu D, Ye M, Zhang W. E6/E7 oncoproteins of high risk HPV-16 upregulate MT1-MMP, MMP-2 and MMP-9 and promote the migration of cervical cancer cells. Int J Clin Exp Pathol. 2015;8(5):4981–9.
Longworth MS, Laimins LA. Pathogenesis of human papillomaviruses in differentiating epithelia. Microbiol Mol Biol Rev. 2004;68(2):362–72.
Chen JJ. Genomic Instability Induced By Human Papillomavirus Oncogenes. N Am J Med Sci. 2010;3(2):43–7.
Iemura K, Anzawa H, Funayama R, Iwakami R, Nakayama K, Kinoshita K, Tanaka K. High levels of chromosomal instability facilitate the tumor growth and sphere formation. Cancer Sci. 2022;113(8):2727–37.
el Awady MK, Kaplan JB, O’Brien SJ, Burk RD. Molecular analysis of integrated human papillomavirus 16 sequences in the cervical cancer cell line SiHa. Virology. 1987;159(2):389–98.
Cooper K, Herrington CS, Graham AK, Evans MF, McGee JO. In situ evidence for HPV 16, 18, 33 integration in cervical squamous cell cancer in Britain and South Africa. J Clin Pathol. 1991;44(5):406–9.
Huang J, Qian Z, Gong Y, Wang Y, Guan Y, Han Y, Yi X, Huang W, Ji L, Xu J, et al. Comprehensive genomic variation profiling of cervical intraepithelial neoplasia and cervical cancer identifies potential targets for cervical cancer early warning. J Med Genet. 2019;56(3):186–94.
Ojesina AI, Lichtenstein L, Freeman SS, Pedamallu CS, Imaz-Rosshandler I, Pugh TJ, Cherniack AD, Ambrogio L, Cibulskis K, Bertelsen B, et al. Landscape of genomic alterations in cervical carcinomas. Nature. 2014;506(7488):371–5.
Xu B, Chotewutmontri S, Wolf S, Klos U, Schmitz M, Durst M, Schwarz E. Multiplex Identification of Human Papillomavirus 16 DNA Integration Sites in Cervical Carcinomas. PLoS ONE. 2013;8(6):e66693.
Hu Z, Zhu D, Wang W, Li W, Jia W, Zeng X, Ding W, Yu L, Wang X, Wang L, et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat Genet. 2015;47(2):158–63.
Holmes A, Lameiras S, Jeannot E, Marie Y, Castera L, Sastre-Garau X, Nicolas A. Mechanistic signatures of HPV insertions in cervical carcinomas. NPJ Genom Med. 2016;1:16004.
Cancer Genome Atlas Research N, Albert Einstein College of M, Analytical Biological S, Barretos Cancer H, Baylor College of M, Beckman Research Institute of City of H, Buck Institute for Research on A, Canada’s Michael Smith Genome Sciences C, Harvard Medical S, Helen FGCC, et al. Integrated genomic and molecular characterization of cervical cancer. Nature. 2017;543(7645):378–84.
Zhang R, Shen C, Zhao L, Wang J, McCrae M, Chen X, Lu F. Dysregulation of host cellular genes targeted by human papillomavirus (HPV) integration contributes to HPV-related cervical carcinogenesis. Int J Cancer. 2016;138(5):1163–74.
Hopman AH, Theelen W, Hommelberg PP, Kamps MA, Herrington CS, Morrison LE, Speel EJ, Smedts F, Ramaekers FC. Genomic integration of oncogenic HPV and gain of the human telomerase gene TERC at 3q26 are strongly associated events in the progression of uterine cervical dysplasia to invasive cancer. J Pathol. 2006;210(4):412–9.
Bodelon C, Vinokurova S, Sampson JN, den Boon JA, Walker JL, Horswill MA, Korthauer K, Schiffman M, Sherman ME, Zuna RE, et al. Chromosomal copy number alterations and HPV integration in cervical precancer and invasive cancer. Carcinogenesis. 2016;37(2):188–96.
Akagi K, Li J, Broutian TR, Padilla-Nash H, Xiao W, Jiang B, Rocco JW, Teknos TN, Kumar B, Wangsa D, et al. Genome-wide analysis of HPV integration in human cancers reveals recurrent, focal genomic instability. Genome Res. 2014;24(2):185–99.
Schmitz M, Driesch C, Jansen L, Runnebaum IB, Durst M. Non-random integration of the HPV genome in cervical cancer. PLoS ONE. 2012;7(6):e39632.
Brant AC, Menezes AN, Felix SP, de Almeida LM, Sammeth M, Moreira MAM. Characterization of HPV integration, viral gene expression and E6E7 alternative transcripts by RNA-Seq: A descriptive study in invasive cervical cancer. Genomics. 2019;111(6):1853–61.
Kamal M, Lameiras S, Deloger M, Morel A, Vacher S, Lecerf C, Dupain C, Jeannot E, Girard E, Baulande S, et al. Human papilloma virus (HPV) integration signature in Cervical Cancer: identification of MACROD2 gene as HPV hot spot integration site. Br J Cancer. 2021;124(4):777–85.
Li W, Qi Y, Cui X, Huo Q, Zhu L, Zhang A, Tan M, Hong Q, Yang Y, Zhang H, et al. Characteristic of HPV Integration in the Genome and Transcriptome of Cervical Cancer Tissues. Biomed Res Int. 2018;2018:6242173.
Cao C, Hong P, Huang X, Lin D, Cao G, Wang L, Feng B, Wu P, Shen H, Xu Q, et al. HPV-CCDC106 integration alters local chromosome architecture and hijacks an enhancer by three-dimensional genome structure remodeling in cervical cancer. J Genet Genomics. 2020;47(8):437–50.
Hernadi Z, Szarka K, Sapy T, Krasznai Z, Veress G, Poka R. The prognostic significance of HPV-16 genome status of the lymph nodes, the integration status and p53 genotype in HPV-16 positive cervical cancer: a long term follow up. BJOG. 2003;110(2):205–9.
Ibragimova M, Tsyganov M, Shpileva O, Churuksaeva O, Bychkov V, Kolomiets L, Litviakov N. HPV status and its genomic integration affect survival of patients with cervical cancer. Neoplasma. 2018;65(3):441–8.
Carow K, Golitz M, Wolf M, Hafner N, Jansen L, Hoyer H, Schwarz E, Runnebaum IB, Durst M. Viral-Cellular DNA Junctions as Molecular Markers for Assessing Intra-Tumor Heterogeneity in Cervical Cancer and for the Detection of Circulating Tumor DNA. Int J Mol Sci. 2017;18(10):2032.
Miao Z, Shen J, Zhang FQ, Hu K, Hou XR, Lian X, Sun S, Yan JF, Liu ZK. The relationship between HPV integration and prognosis of cervical cancer. Zhonghua Zhong Liu Za Zhi. 2020;42(12):1014–9.
Yang W, Liu Y, Dong R, Liu J, Lang J, Yang J, Wang W, Li J, Meng B, Tian G. Accurate Detection of HPV Integration Sites in Cervical Cancer Samples Using the Nanopore MinION Sequencer Without Error Correction. Front Genet. 2020;11:660.
Xiong J, Cheng J, Shen H, Ren C, Wang L, Gao C, Zhu T, Li X, Ding W, Zhu D, et al. Detection of HPV and Human Chromosome Sites by Dual-Color Fluorescence In Situ Hybridization Reveals Recurrent HPV Integration Sites and Heterogeneity in Cervical Cancer. Front Oncol. 2021;11:734758.
McGranahan N, Swanton C. Clonal Heterogeneity and Tumor Evolution: Past, Present, and the Future. Cell. 2017;168(4):613–28.
Venkatesan S, Swanton C. Tumor Evolutionary Principles: How Intratumor Heterogeneity Influences Cancer Treatment and Outcome. Am Soc Clin Oncol Educ Book. 2016;35:e141-149.
Lynch M, Ackerman MS, Gout JF, Long H, Sung W, Thomas WK, Foster PL. Genetic drift, selection and the evolution of the mutation rate. Nat Rev Genet. 2016;17(11):704–14.
Turajlic S, Sottoriva A, Graham T, Swanton C. Resolving genetic heterogeneity in cancer. Nat Rev Genet. 2019;20(7):404–16.
Cappellen D, De Oliveira C, Ricol D, de Medina S, Bourdin J, Sastre-Garau X, Chopin D, Thiery JP, Radvanyi F. Frequent activating mutations of FGFR3 in human bladder and cervix carcinomas. Nat Genet. 1999;23(1):18–20.
Reiter JG, Baretti M, Gerold JM, Makohon-Moore AP, Daud A, Iacobuzio-Donahue CA, Azad NS, Kinzler KW, Nowak MA, Vogelstein B. An analysis of genetic heterogeneity in untreated cancers. Nat Rev Cancer. 2019;19(11):639–50.
Ellsworth DL, Blackburn HL, Shriver CD, Rabizadeh S, Soon-Shiong P, Ellsworth RE. Single-cell sequencing and tumorigenesis: improved understanding of tumor evolution and metastasis. Clin Transl Med. 2017;6(1):15.
Levitin HM, Yuan J, Sims PA. Single-Cell Transcriptomic Analysis of Tumor Heterogeneity. Trends Cancer. 2018;4(4):264–8.
Dentro SC, Leshchiner I, Haase K, Tarabichi M, Wintersinger J, Deshwar AG, Yu K, Rubanova Y, Macintyre G, Demeulemeester J, et al. Characterizing genetic intra-tumor heterogeneity across 2,658 human cancer genomes. Cell. 2021;184(8):2239-2254 e2239.
Forbes SA, Bhamra G, Bamford S, Dawson E, Kok C, Clements J, et al. The Catalogue of Somatic Mutations in Cancer (COSMIC). Curr Protoc Hum Genet. 2008; Chapter 10:Unit 10.11. https://doi.org/10.1002/0471142905.hg1011s57.
Wong JKL, Aichmüller C, Schulze M, Hlevnjak M, Elgaafary S, Lichter P, Zapatka M. Association of mutation signature effectuating processes with mutation hotspots in driver genes and non-coding regions. Nat Commun. 2022;13(1):178.
Roberts SA, Lawrence MS, Klimczak LJ, Grimm SA, Fargo D, Stojanov P, Kiezun A, Kryukov GV, Carter SL, Saksena G, et al. An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat Genet. 2013;45(9):970–6.
Cancer Genome Atlas Research Network; Albert Einstein College of Medicine; Analytical Biological Services; Barretos Cancer Hospital; Baylor College of Medicine; Beckman Research Institute of City of Hope; Buck Institute for Research on Aging; Canada's Michael Smith Genome Sciences Centre; Harvard Medical School; Helen F. Graham Cancer Center &Research Institute at Christiana Care Health Services; HudsonAlpha Institute for Biotechnology; ILSbio, LLC; Indiana University School of Medicine; Institute of Human Virology; Institute for Systems Biology; International Genomics Consortium; Leidos Biomedical; Massachusetts General Hospital; McDonnell Genome Institute at Washington University; Medical College of Wisconsin; Medical University of South Carolina; Memorial Sloan Kettering Cancer Center; Montefiore Medical Center; NantOmics; National Cancer Institute; National Hospital, Abuja, Nigeria; National Human Genome Research Institute; National Institute of Environmental Health Sciences; National Institute on Deafness &Other Communication Disorders; Ontario Tumour Bank, London Health Sciences Centre; Ontario Tumour Bank, Ontario Institute for Cancer Research; Ontario Tumour Bank, The Ottawa Hospital; Oregon Health &Science University; Samuel Oschin Comprehensive Cancer Institute, Cedars-Sinai Medical Center; SRA International; St Joseph's Candler Health System; Eli &Edythe L. Broad Institute of Massachusetts Institute of Technology &Harvard University; Research Institute at Nationwide Children's Hospital; Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins University; University of Bergen; University of Texas MD Anderson Cancer Center; University of Abuja Teaching Hospital; University of Alabama at Birmingham; University of California, Irvine; University of California Santa Cruz; University of Kansas Medical Center; University of Lausanne; University of New Mexico Health Sciences Center; University of North Carolina at Chapel Hill; University of Oklahoma Health Sciences Center; University of Pittsburgh; University of São Paulo, Ribeir ão Preto Medical School; University of Southern California; University of Washington; University of Wisconsin School of Medicine &Public Health; Van Andel Research Institute; Washington University in St Louis. Integrated genomic and molecular characterization of cervical cancer. Nature. 2017;543(7645):378–84.
Chung TK, Van Hummelen P, Chan PK, Cheung TH, Yim SF, Yu MY, Ducar MD, Thorner AR, MacConaill LE, Doran G, et al. Genomic aberrations in cervical adenocarcinomas in Hong Kong Chinese women. Int J Cancer. 2015;137(4):776–83.
Wingo SN, Gallardo TD, Akbay EA, Liang MC, Contreras CM, Boren T, Shimamura T, Miller DS, Sharpless NE, Bardeesy N, et al. Somatic LKB1 mutations promote cervical cancer progression. PLoS ONE. 2009;4(4):e5137.
Doci CL, Mankame TP, Langerman A, Ostler KR, Kanteti R, Best T, Onel K, Godley LA, Salgia R, Lingen MW. Characterization of NOL7 gene point mutations, promoter methylation, and protein expression in cervical cancer. Int J Gynecol Pathol. 2012;31(1):15–24.
Analysis identifies mutations linked to cervical cancer. Cancer Discov. 2014;4(3):OF2. https://doi.org/10.1158/2159-8290.CD-NB2014-010.
Wright AA, Howitt BE, Myers AP, Dahlberg SE, Palescandolo E, Van Hummelen P, MacConaill LE, Shoni M, Wagle N, Jones RT, et al. Oncogenic mutations in cervical cancer: genomic differences between adenocarcinomas and squamous cell carcinomas of the cervix. Cancer. 2013;119(21):3776–83.
Frumovitz M, Burzawa JK, Byers LA, Lyons YA, Ramalingam P, Coleman RL, Brown J. Sequencing of mutational hotspots in cancer-related genes in small cell neuroendocrine cervical cancer. Gynecol Oncol. 2016;141(3):588–91.
Yoon JH, Lee JM, Namkoong SE, Bae SM, Kim YW, Han SJ, Cho YL, Nam GH, Kim CK, Seo JS, et al. cDNA Microarray Analysis of Gene Expression Profiles Associated with Cervical Cancer. Cancer Res Treat. 2003;35(5):451–9.
Wong YF, Cheung TH, Tsao GS, Lo KW, Yim SF, Wang VW, Heung MM, Chan SC, Chan LK, Ho TW, et al. Genome-wide gene expression profiling of cervical cancer in Hong Kong women by oligonucleotide microarray. Int J Cancer. 2006;118(10):2461–9.
Harima Y, Togashi A, Horikoshi K, Imamura M, Sougawa M, Sawada S, Tsunoda T, Nakamura Y, Katagiri T. Prediction of outcome of advanced cervical cancer to thermoradiotherapy according to expression profiles of 35 genes selected by cDNA microarray analysis. Int J Radiat Oncol Biol Phys. 2004;60(1):237–48.
Wong YF, Selvanayagam ZE, Wei N, Porter J, Vittal R, Hu R, Lin Y, Liao J, Shih JW, Cheung TH, et al. Expression genomics of cervical cancer: molecular classification and prediction of radiotherapy response by DNA microarray. Clin Cancer Res. 2003;9(15):5486–92.
Song JY, Lee JK, Lee NW, Jung HH, Kim SH, Lee KW. Microarray analysis of normal cervix, carcinoma in situ, and invasive cervical cancer: identification of candidate genes in pathogenesis of invasion in cervical cancer. Int J Gynecol Cancer. 2008;18(5):1051–9.
Bachtiary B, Boutros PC, Pintilie M, Shi W, Bastianutto C, Li JH, Schwock J, Zhang W, Penn LZ, Jurisica I, et al. Gene expression profiling in cervical cancer: an exploration of intratumor heterogeneity. Clin Cancer Res. 2006;12(19):5632–40.
Borkamo ED, Schem BC, Fluge O, Bruland O, Dahl O, Mella O. cDNA microarray analysis of serially sampled cervical cancer specimens from patients treated with thermochemoradiotherapy. Int J Radiat Oncol Biol Phys. 2009;75(5):1562–9.
Santin AD, Zhan F, Bignotti E, Siegel ER, Cane S, Bellone S, Palmieri M, Anfossi S, Thomas M, Burnett A, et al. Gene expression profiles of primary HPV16- and HPV18-infected early stage cervical cancers and normal cervical epithelium: identification of novel candidate molecular markers for cervical cancer diagnosis and therapy. Virology. 2005;331(2):269–91.
Zhu MY, Chen F, Niyazi M, Sui S, Gao DM. Variation in apoptotic gene expression in cervical cancer through oligonucleotide microarray profiling. J Low Genit Tract Dis. 2015;19(1):46–54.
Liu MY, Zhang H, Hu YJ, Chen YW, Zhao XN. Identification of key genes associated with cervical cancer by comprehensive analysis of transcriptome microarray and methylation microarray. Oncol Lett. 2016;12(1):473–8.
Wang S, Chen X. Identification of potential biomarkers in cervical cancer with combined public mRNA and miRNA expression microarray data analysis. Oncol Lett. 2018;16(4):5200–8.
Mousavi SZ, Poortahmasebi V, Mokhtari-Azad T, Shahmahmoodi S, Farahmand M, Farzanehpour M, Jalilvand S. The dysregulation of microarray gene expression in cervical cancer is associated with overexpression of a unique messenger RNA signature. Iran J Microbiol. 2020;12(6):629–35.
Peng G, Dan W, Jun W, Junjun Y, Tong R, Baoli Z, Yang X. Transcriptome profiling of the cancer and adjacent nontumor tissues from cervical squamous cell carcinoma patients by RNA sequencing. Tumour Biol. 2015;36(5):3309–17.
Wang Y, Liu L, Chen Z. Transcriptome profiling of cervical cancer cells acquired resistance to cisplatin by deep sequencing. Artif Cells Nanomed Biotechnol. 2019;47(1):2820–9.
Chen T, Yang S, Xu J, Lu W, Xie X. Transcriptome sequencing profiles of cervical cancer tissues and SiHa cells. Funct Integr Genomics. 2020;20(2):211–21.
Kinker GS, Greenwald AC, Tal R, Orlova Z, Cuoco MS, McFarland JM, Warren A, Rodman C, Roth JA, Bender SA, et al. Pan-cancer single-cell RNA-seq identifies recurring programs of cellular heterogeneity. Nat Genet. 2020;52(11):1208–18.
Li C, Guo L, Li S, Hua K. Single-cell transcriptomics reveals the landscape of intra-tumoral heterogeneity and transcriptional activities of ECs in CC. Mol Ther Nucleic Acids. 2021;24:682–94.
Gu M, He T, Yuan Y, Duan S, Li X, Shen C. Single-Cell RNA Sequencing Reveals Multiple Pathways and the Tumor Microenvironment Could Lead to Chemotherapy Resistance in Cervical Cancer. Front Oncol. 2021;11:753386.
Gagliardi A, Porter VL, Zong Z, Bowlby R, Titmuss E, Namirembe C, Griner NB, Petrello H, Bowen J, Chan SK, et al. Analysis of Ugandan cervical carcinomas identifies human papillomavirus clade-specific epigenome and transcriptome landscapes. Nat Genet. 2020;52(8):800–10.
Burki TK. Novel mutations in cervical cancer. Lancet Oncol. 2017;18(3):e137.
Ren T, Suo J, Liu S, Wang S, Shu S, Xiang Y, Lang JH. Using low-coverage whole genome sequencing technique to analyze the chromosomal copy number alterations in the exfoliative cells of cervical cancer. J Gynecol Oncol. 2018;29(5):e78.
Lai EC. Micro RNAs are complementary to 3’ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat Genet. 2002;30(4):363–4.
Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26–46.
Haddad G, Lorenzen JM. Biogenesis and Function of Circular RNAs in Health and in Disease. Front Pharmacol. 2019;10:428.
Kartha RV, Subramanian S. Competing endogenous RNAs (ceRNAs): new entrants to the intricacies of gene regulation. Front Genet. 2014;5:8.
Basera A, Hull R, Demetriou D, Bates DO, Kaufmann AM, Dlamini Z, Marima R. Competing Endogenous RNA (ceRNA) Networks and Splicing Switches in Cervical Cancer: HPV Oncogenesis, Clinical Significance and Therapeutic Opportunities. Microorganisms. 2022;10(9):1852.
Tornesello ML, Faraonio R, Buonaguro L, Annunziata C, Starita N, Cerasuolo A, Pezzuto F, Tornesello AL, Buonaguro FM. The Role of microRNAs, Long Non-coding RNAs, and Circular RNAs in Cervical Cancer. Front Oncol. 2020;10:150.
Dong J, Su M, Chang W, Zhang K, Wu S, Xu T. Long non-coding RNAs on the stage of cervical cancer (Review). Oncol Rep. 2017;38(4):1923–31.
Luan X, Wang Y. LncRNA XLOC_006390 facilitates cervical cancer tumorigenesis and metastasis as a ceRNA against miR-331-3p and miR-338-3p. J Gynecol Oncol. 2018;29(6): e95.
Shen L, Dang J, Liu S, Xian B, Deng Y, Qu D. CircRNA VPRBP inhibits tumorigenicity of cervical cancer via miR-93-5p/FRMD6 axis. Reprod Sci. 2022;29(8):2251–64.
Tang Q, Chen Z, Zhao L, Xu H. Circular RNA hsa_circ_0000515 acts as a miR-326 sponge to promote cervical cancer progression through up-regulation of ELK1. Aging (Albany NY). 2019;11(22):9982–99.
Eddy GL, Bundy BN, Creasman WT, Spirtos NM, Mannel RS, Hannigan E, O’Connor D. Treatment of (“bulky”) stage IB cervical cancer with or without neoadjuvant vincristine and cisplatin prior to radical hysterectomy and pelvic/para-aortic lymphadenectomy: a phase III trial of the gynecologic oncology group. Gynecol Oncol. 2007;106(2):362–9.
Duenas-Gonzalez A, Zarba JJ, Patel F, Alcedo JC, Beslija S, Casanova L, Pattaranutaporn P, Hameed S, Blair JM, Barraclough H, et al. Phase III, open-label, randomized study comparing concurrent gemcitabine plus cisplatin and radiation followed by adjuvant gemcitabine and cisplatin versus concurrent cisplatin and radiation in patients with stage IIB to IVA carcinoma of the cervix. J Clin Oncol. 2011;29(13):1678–85.
Tewari KS, Sill MW, Monk BJ, Penson RT, Moore DH, Lankes HA, Ramondetta LM, Landrum LM, Randall LM, Oaknin A, et al. Circulating Tumor Cells In Advanced Cervical Cancer: NRG Oncology-Gynecologic Oncology Group Study 240 (NCT 00803062). Mol Cancer Ther. 2020;19(11):2363–70.
Pfaendler KS, Tewari KS. Changing paradigms in the systemic treatment of advanced cervical cancer. Am J Obstet Gynecol. 2016;214(1):22–30.
Koh WJ, Abu-Rustum NR, Bean S, Bradley K, Campos SM, Cho KR, Chon HS, Chu C, Clark R, Cohn D, et al. Cervical Cancer, Version 3.2019, NCCN Clinical Practice Guidelines in Oncology. J Natl Compr Canc Netw. 2019;17(1):64–84.
Zhu H, Luo H, Zhang W, Shen Z, Hu X, Zhu X. Molecular mechanisms of cisplatin resistance in cervical cancer. Drug Des Devel Ther. 2016;10:1885–95.
Ferrandina G, Lauriola L, Distefano MG, Zannoni GF, Gessi M, Legge F, Maggiano N, Mancuso S, Capelli A, Scambia G, et al. Increased cyclooxygenase-2 expression is associated with chemotherapy resistance and poor survival in cervical cancer patients. J Clin Oncol. 2002;20(4):973–81.
Kina S, Kinjo T, Liang F, Nakasone T, Yamamoto H, Arasaki A. Targeting EphA4 abrogates intrinsic resistance to chemotherapy in well-differentiated cervical cancer cell line. Eur J Pharmacol. 2018;840:70–8.
Cai L, Wang Z, Liu D. Interference with endogenous EZH2 reverses the chemotherapy drug resistance in cervical cancer cells partly by up-regulating Dicer expression. Tumour Biol. 2016;37(5):6359–69.
Duan W, Liu X. PSAT1 Upregulation Contributes to Cell Growth and Cisplatin Resistance in Cervical Cancer Cells via Regulating PI3K/AKT Signaling Pathway. Ann Clin Lab Sci. 2020;50(4):512–8.
Wang N, Hou MS, Zhan Y, Shen XB, Xue HY. MALAT1 promotes cisplatin resistance in cervical cancer by activating the PI3K/AKT pathway. Eur Rev Med Pharmacol Sci. 2018;22(22):7653–9.
Fang X, Zhong G, Wang Y, Lin Z, Lin R, Yao T. Low GAS5 expression may predict poor survival and cisplatin resistance in cervical cancer. Cell Death Dis. 2020;11(7):531.
Wu X, Zhong Y, Chen Q, Zhang X, Zhang H. Enhancer of mRNA Decapping protein 4 (EDC4) interacts with replication protein a (RPA) and contributes to Cisplatin resistance in cervical Cancer by alleviating DNA damage. Hereditas. 2020;157(1):41.
Zhou Y, Liu F, Xu Q, Yang B, Li X, Jiang S, Hu L, Zhang X, Zhu L, Li Q, et al. Inhibiting Importin 4-mediated nuclear import of CEBPD enhances chemosensitivity by repression of PRKDC-driven DNA damage repair in cervical cancer. Oncogene. 2020;39(34):5633–48.
Wen X, Liu S, Sheng J, Cui M. Recent advances in the contribution of noncoding RNAs to cisplatin resistance in cervical cancer. PeerJ. 2020;8:e9234.
Masadah R, Rauf S, Pratama MY, Tiribelli C, Pascut D. The Role of microRNAs in the Cisplatin- and Radio-Resistance of Cervical Cancer. Cancers (Basel). 2021;13(5):1168.
Luo X, Wei J, Yang FL, Pang XX, Shi F, Wei YX, Liao BY, Wang JL. Exosomal lncRNA HNF1A-AS1 affects cisplatin resistance in cervical cancer cells through regulating microRNA-34b/TUFT1 axis. Cancer Cell Int. 2019;19:323.
Hou H, Yu R, Zhao H, Yang H, Hu Y, Hu Y, Guo J. LncRNA OTUD6B-AS1 Induces Cisplatin Resistance in Cervical Cancer Cells Through Up-Regulating Cyclin D2 via miR-206. Front Oncol. 2021;11:777220.
Liu Y, Guo R, Qiao Y, Han L, Liu M. LncRNA NNT-AS1 contributes to the cisplatin resistance of cervical cancer through NNT-AS1/miR-186/HMGB1 axis. Cancer Cell Int. 2020;20:190.
Lorusso D, Petrelli F, Coinu A, Raspagliesi F, Barni S. A systematic review comparing cisplatin and carboplatin plus paclitaxel-based chemotherapy for recurrent or metastatic cervical cancer. Gynecol Oncol. 2014;133(1):117–23.
Seol HJ, Ulak R, Ki KD, Lee JM. Cytotoxic and targeted systemic therapy in advanced and recurrent cervical cancer: experience from clinical trials. Tohoku J Exp Med. 2014;232(4):269–76.
Cooke SL, Temple J, Macarthur S, Zahra MA, Tan LT, Crawford RA, Ng CK, Jimenez-Linan M, Sala E, Brenton JD. Intra-tumour genetic heterogeneity and poor chemoradiotherapy response in cervical cancer. Br J Cancer. 2011;104(2):361–8.
Mao BD, Xu P, Xu P, Zhong Y, Ding WW, Meng QZ. LINC00511 knockdown prevents cervical cancer cell proliferation and reduces resistance to paclitaxel. J Biosci. 2019;44(2):44.
Sen P, Ghosal S, Hazra R, Mohanty R, Arega S, Sahu B, Ganguly N. CRISPR-mediated knockdown of miR-214 modulates cell fate in response to anti-cancer drugs in HPV-negative and HPVpositive cervical cancer cells. J Biosci. 2020;45:80.
Dong M, Li P, Xie Y, Wang Z, Wang R. CircMYBL2 regulates the resistance of cervical cancer cells to paclitaxel via miR-665-dependent regulation of EGFR. Drug Dev Res. 2021;82(8):1193–205.
Rose PG, Bundy BN, Watkins EB, Thigpen JT, Deppe G, Maiman MA, Clarke-Pearson DL, Insalaco S. Concurrent cisplatin-based radiotherapy and chemotherapy for locally advanced cervical cancer. N Engl J Med. 1999;340(15):1144–53.
Feng Y, Wang Z, Yang N, Liu S, Yan J, Song J, Yang S, Zhang Y. Identification of Biomarkers for Cervical Cancer Radiotherapy Resistance Based on RNA Sequencing Data. Front Cell Dev Biol. 2021;9:724172.
Liu J, Zhang J, Wang X, Li Y, Chen Y, Li K, Zhang J, Yao L, Guo G. HIF-1 and NDRG2 contribute to hypoxia-induced radioresistance of cervical cancer Hela cells. Exp Cell Res. 2010;316(12):1985–93.
Li N, Meng DD, Gao L, Xu Y, Liu PJ, Tian YW, Yi ZY, Zhang Y, Tie XJ, Xu ZQ. Overexpression of HOTAIR leads to radioresistance of human cervical cancer via promoting HIF-1alpha expression. Radiat Oncol. 2018;13(1):210.
Ju X, Liang S, Zhu J, Ke G, Wen H, Wu X. Extracellular matrix metalloproteinase inducer (CD147/BSG/EMMPRIN)-induced radioresistance in cervical cancer by regulating the percentage of the cells in the G2/m phase of the cell cycle and the repair of DNA Double-strand Breaks (DSBs). Am J Transl Res. 2016;8(6):2498–511.
Pranatharthi A, Thomas P, Udayashankar AH, Bhavani C, Suresh SB, Krishna S, Thatte J, Srikantia N, Ross CR, Srivastava S. RhoC regulates radioresistance via crosstalk of ROCK2 with the DNA repair machinery in cervical cancer. J Exp Clin Cancer Res. 2019;38(1):392.
Jiao X, Zhang S, Jiao J, Zhang T, Qu W, Muloye GM, Kong B, Zhang Q, Cui B. Promoter methylation of SEPT9 as a potential biomarker for early detection of cervical cancer and its overexpression predicts radioresistance. Clin Epigenetics. 2019;11(1):120.
Li Z, Zhang Y, Sui S, Hua Y, Zhao A, Tian X, Wang R, Guo W, Yu W, Zou K, et al. Targeting HMGB3/hTERT axis for radioresistance in cervical cancer. J Exp Clin Cancer Res. 2020;39(1):243.
Li Z, Liu X, Yu H, Wang S, Zhao S, Jiang G. USP21 regulates Hippo signaling to promote radioresistance by deubiquitinating FOXM1 in cervical cancer. Hum Cell. 2022;35(1):333-47.
Zhang B, Chen J, Ren Z, Chen Y, Li J, Miao X, Song Y, Zhao T, Li Y, Shi Y, et al. A specific miRNA signature promotes radioresistance of human cervical cancer cells. Cancer Cell Int. 2013;13(1):118.
Pedroza-Torres A, Campos-Parra AD, Millan-Catalan O, Loissell-Baltazar YA, Zamudio-Meza H, Cantu de Leon D, Montalvo-Esquivel G, Isla-Ortiz D, Herrera LA, Angeles-Zaragoza O, et al. MicroRNA-125 modulates radioresistance through targeting p21 in cervical cancer. Oncol Rep. 2018;39(3):1532–40.
Fan L, Huang C, Li J, Gao T, Lin Z, Yao T. Long noncoding RNA urothelial cancer associated 1 regulates radioresistance via the hexokinase 2/glycolytic pathway in cervical cancer. Int J Mol Med. 2018;42(4):2247–59.
Liu J, Liu X, Li R. LncRNA SNHG6 enhances the radioresistance and promotes the growth of cervical cancer cells by sponging miR-485-3p. Cancer Cell Int. 2020;20:424.
Choo YW, Kang M, Kim HY, Han J, Kang S, Lee JR, Jeong GJ, Kwon SP, Song SY, Go S, et al. M1 Macrophage-Derived Nanovesicles Potentiate the Anticancer Efficacy of Immune Checkpoint Inhibitors. ACS Nano. 2018;12(9):8977–93.
Chu TY, Yang JT, Huang TH, Liu HW. Crosstalk with cancer-associated fibroblasts increases the growth and radiation survival of cervical cancer cells. Radiat Res. 2014;181(5):540–7.
Chung HC, Ros W, Delord JP, Perets R, Italiano A, Shapira-Frommer R, Manzuk L, Piha-Paul SA, Xu L, Zeigenfuss S, et al. Efficacy and Safety of Pembrolizumab in Previously Treated Advanced Cervical Cancer: Results From the Phase II KEYNOTE-158 Study. J Clin Oncol. 2019;37(17):1470–8.
Colombo N, Dubot C, Lorusso D, Caceres MV, Hasegawa K, Shapira-Frommer R, Tewari KS, Salman P, Hoyos Usta E, Yañez E, et al. Pembrolizumab for Persistent, Recurrent, or Metastatic Cervical Cancer. N Engl J Med. 2021;385(20):1856–67.
Naumann RW, Hollebecque A, Meyer T, Devlin MJ, Oaknin A, Kerger J, Lopez-Picazo JM, Machiels JP, Delord JP, Evans TRJ, et al. Safety and Efficacy of Nivolumab Monotherapy in Recurrent or Metastatic Cervical, Vaginal, or Vulvar Carcinoma: Results From the Phase I/II CheckMate 358 Trial. J Clin Oncol. 2019;37(31):2825–34.
Kagabu M, Nagasawa T, Sato C, Fukagawa Y, Kawamura H, Tomabechi H, Takemoto S, Shoji T, Baba T. Immunotherapy for Uterine Cervical Cancer Using Checkpoint Inhibitors: Future Directions. Int J Mol Sci. 2020;21(7):2335.
Cosper PF, McNair C, Gonzalez I, Wong N, Knudsen KE, Chen JJ, Markovina S, Schwarz JK, Grigsby PW, Wang X. Decreased local immune response and retained HPV gene expression during chemoradiotherapy are associated with treatment resistance and death from cervical cancer. Int J Cancer. 2020;146(7):2047–58.
Allouch S, Malki A, Allouch A, Gupta I, Vranic S, Al Moustafa AE. High-Risk HPV Oncoproteins and PD-1/PD-L1 Interplay in Human Cervical Cancer: Recent Evidence and Future Directions. Front Oncol. 2020;10:914.
Zhang L, Zhao Y, Tu Q, Xue X, Zhu X, Zhao KN. The Roles of Programmed Cell Death Ligand-1/ Programmed Cell Death-1 (PD-L1/PD-1) in HPV-induced Cervical Cancer and Potential for their Use in Blockade Therapy. Curr Med Chem. 2021;28(5):893–909.
Rice AE, Latchman YE, Balint JP, Lee JH, Gabitzsch ES, Jones FR. An HPV-E6/E7 immunotherapy plus PD-1 checkpoint inhibition results in tumor regression and reduction in PD-L1 expression. Cancer Gene Ther. 2015;22(9):454–62.
Kim SS, Shen S, Miyauchi S, Sanders PD, Franiak-Pietryga I, Mell L, Gutkind JS, Cohen EEW, Califano JA, Sharabi AB. B Cells Improve Overall Survival in HPV-Associated Squamous Cell Carcinomas and Are Activated by Radiation and PD-1 Blockade. Clin Cancer Res. 2020;26(13):3345–59.
Xu S, Wang X, Yang Y, Li Y, Wu S. LSD1 silencing contributes to enhanced efficacy of anti-CD47/PD-L1 immunotherapy in cervical cancer. Cell Death Dis. 2021;12(4):282.
Wen Y, Zhang S, Yang J, Guo D. Identification of driver genes regulating immune cell infiltration in cervical cancer by multiple omics integration. Biomed Pharmacother. 2019;120:109546.
Bowden SJ, Bodinier B, Kalliala I, Zuber V, Vuckovic D, Doulgeraki T, Whitaker MD, Wielscher M, Cartwright R, Tsilidis KK, et al. Genetic variation in cervical preinvasive and invasive disease: a genome-wide association study. Lancet Oncol. 2021;22(4):548–57.
Franceschi S. Genomic characterisation of cervical cancer and human papillomavirus: new opportunities for precision medicine. Lancet Oncol. 2021;22(4):419–20.
Draghiciu O, Boerma A, Hoogeboom BN, Nijman HW, Daemen T. A rationally designed combined treatment with an alphavirus-based cancer vaccine, sunitinib and low-dose tumor irradiation completely blocks tumor development. Oncoimmunology. 2015;4(10):e1029699.
Sun YY, Peng S, Han L, Qiu J, Song L, Tsai Y, Yang B, Roden RB, Trimble CL, Hung CF, et al. Local HPV Recombinant Vaccinia Boost Following Priming with an HPV DNA Vaccine Enhances Local HPV-Specific CD8+ T-cell-Mediated Tumor Control in the Genital Tract. Clin Cancer Res. 2016;22(3):657–69.
Peng S, Tan M, Li YD, Cheng MA, Farmer E, Ferrall L, Gaillard S, Roden RBS, Hung CF, Wu TC. PD-1 blockade synergizes with intratumoral vaccination of a therapeutic HPV protein vaccine and elicits regression of tumor in a preclinical model. Cancer Immunol Immunother. 2021;70(4):1049–62.
Wendel Naumann R, Leath CA 3rd. Advances in immunotherapy for cervical cancer. Curr Opin Oncol. 2020;32(5):481–7.
Anderson AR, Weaver AM, Cummings PT, Quaranta V. Tumor morphology and phenotypic evolution driven by selective pressure from the microenvironment. Cell. 2006;127(5):905–15.
Sottoriva A, Verhoeff JJ, Borovski T, McWeeney SK, Naumov L, Medema JP, Sloot PM, Vermeulen L. Cancer stem cell tumor model reveals invasive morphology and increased phenotypical heterogeneity. Cancer Res. 2010;70(1):46–56.
Meacham CE, Morrison SJ. Tumour heterogeneity and cancer cell plasticity. Nature. 2013;501(7467):328–37.
Gu W, Yeo E, McMillan N, Yu C. Silencing oncogene expression in cervical cancer stem-like cells inhibits their cell growth and self-renewal ability. Cancer Gene Ther. 2011;18(12):897–905.
Zhang J, Chen X, Bian L, Wang Y, Liu H. CD44+/CD24+-Expressing Cervical Cancer Cells and Radioresistant Cervical Cancer Cells Exhibit Cancer Stem Cell Characteristics. Gynecol Obstet Invest. 2019;84(2):174–82.
Yang Y, Wang Y, Yin C, Li X. Clinical significance of the stem cell gene Oct-4 in cervical cancer. Tumour Biol. 2014;35(6):5339–45.
Tian Y, Ding W, Wang Y, Ji T, Sun S, Mo Q, Chen P, Fang Y, Liu J, Wang B, et al. Ubiquitin B in cervical cancer: critical for the maintenance of cancer stem-like cell characters. PLoS ONE. 2013;8(12):e84457.
Wang H, Cai HB, Chen LL, Zhao WJ, Li P, Wang ZQ, Li Z. STAT3 correlates with stem cell-related transcription factors in cervical cancer. J Huazhong Univ Sci Technolog Med Sci. 2015;35(6):891–7.
Liu W, Gao Q, Chen K, Xue X, Li M, Chen Q, Zhu G, Gao Y. Hiwi facilitates chemoresistance as a cancer stem cell marker in cervical cancer. Oncol Rep. 2014;32(5):1853–60.
Sato M, Kawana K, Fujimoto A, Yoshida M, Nakamura H, Nishida H, Inoue T, Taguchi A, Takahashi J, Adachi K, et al. Clinical significance of Gremlin 1 in cervical cancer and its effects on cancer stem cell maintenance. Oncol Rep. 2016;35(1):391–7.
Ortiz-Sanchez E, Santiago-Lopez L, Cruz-Dominguez VB, Toledo-Guzman ME, Hernandez-Cueto D, Muniz-Hernandez S, Garrido E, Cantu De Leon D, Garcia-Carranca A. Characterization of cervical cancer stem cell-like cells: phenotyping, stemness, and human papilloma virus co-receptor expression. Oncotarget. 2016;7(22):31943–54.
Cao HZ, Liu XF, Yang WT, Chen Q, Zheng PS. LGR5 promotes cancer stem cell traits and chemoresistance in cervical cancer. Cell Death Dis. 2017;8(9):e3039.
Leung CON, Deng W, Ye TM, Ngan HYS, Tsao SW, Cheung ANY, Ziru N, Yuen DCK, Pang RTK, Yeung WSB. MicroRNA-135a-induced formation of CD133+ subpopulation with cancer stem cell properties in cervical cancer. Carcinogenesis. 2020;41(11):1592–604.
Williams NL, Werner TL, Jarboe EA, Gaffney DK. Adenocarcinoma of the cervix: should we treat it differently? Curr Oncol Rep. 2015;17(4):17.
Stolnicu S, Barsan I, Hoang L, Patel P, Terinte C, Pesci A, Aviel-Ronen S, Kiyokawa T, Alvarado-Cabrero I, Pike MC, et al. International Endocervical Adenocarcinoma Criteria and Classification (IECC): A New Pathogenetic Classification for Invasive Adenocarcinomas of the Endocervix. Am J Surg Pathol. 2018;42(2):214–26.
Lu Z, Chen J. Introduction of WHO classification of tumours of female reproductive organs, fourth edition. Zhonghua Bing Li Xue Za Zhi. 2014;43(10):649–50.
Park KJ. Cervical adenocarcinoma: integration of HPV status, pattern of invasion, morphology and molecular markers into classification. Histopathology. 2020;76(1):112–27.
Hodgson A, Olkhov-Mitsel E, Howitt BE, Nucci MR, Parra-Herran C. International Endocervical Adenocarcinoma Criteria and Classification (IECC): correlation with adverse clinicopathological features and patient outcome. J Clin Pathol. 2019;72(5):347–53.
Bulutay P, Haberal N, Ozen O, Erdem O, Zeren EH, Kulac I, et al. Reproducibility of Morphologic Parameters of the International Endocervical Adenocarcinoma Criteria and Classification System and Correlation With Clinicopathologic Parameters: A Multi-Institutional Study. Int J Gynecol Pathol. 2022;41(5):447–58.
Cree IA, White VA, Indave BI, Lokuhetty D. Revising the WHO classification: female genital tract tumours. Histopathology. 2020;76(1):151–6.
Lou H, Villagran G, Boland JF, Im KM, Polo S, Zhou W, Odey U, Juarez-Torres E, Medina-Martinez I, Roman-Basaure E, et al. Genome Analysis of Latin American Cervical Cancer: Frequent Activation of the PIK3CA Pathway. Clin Cancer Res. 2015;21(23):5360–70.
Selenica P, Alemar B, Matrai C, Talia KL, Veras E, Hussein Y, Oliva E, Beets-Tan RGH, Mikami Y, McCluggage WG, et al. Massively parallel sequencing analysis of 68 gastric-type cervical adenocarcinomas reveals mutations in cell cycle-related genes and potentially targetable mutations. Mod Pathol. 2021;34(6):1213–25.
Gadducci A, Carinelli S, Aletti G. Neuroendrocrine tumors of the uterine cervix: A therapeutic challenge for gynecologic oncologists. Gynecol Oncol. 2017;144(3):637–46.
Tempfer CB, Tischoff I, Dogan A, Hilal Z, Schultheis B, Kern P, Rezniczek GA. Neuroendocrine carcinoma of the cervix: a systematic review of the literature. BMC Cancer. 2018;18(1):530.
Cohen JG, Kapp DS, Shin JY, Urban R, Sherman AE, Chen LM, Osann K, Chan JK. Small cell carcinoma of the cervix: treatment and survival outcomes of 188 patients. Am J Obstet Gynecol. 2010;203(4):347 e341-346.
Li J, Ouyang Y, Tao Y, Wang L, Li M, Gao L, Cao X. Small cell carcinoma of the uterine cervix: a multi-institutional experience. Int J Gynecol Cancer. 2020;30(2):174–80.
Xing D, Zheng G, Schoolmeester JK, Li Z, Pallavajjala A, Haley L, Conner MG, Vang R, Hung CF, Wu TC, et al. Next-generation Sequencing Reveals Recurrent Somatic Mutations in Small Cell Neuroendocrine Carcinoma of the Uterine Cervix. Am J Surg Pathol. 2018;42(6):750–60.
Pei X, Xiang L, Chen W, Jiang W, Yin L, Shen X, Zhou X, Yang H. The next generation sequencing of cancer-related genes in small cell neuroendocrine carcinoma of the cervix. Gynecol Oncol. 2021;161(3):779–86.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural Science Foundation of China (81974414, 81772788, 81873430).
Author information
Authors and Affiliations
Contributions
ZQH and QS conceived the manuscript. QS and LLW collected relevant references, drafted the manuscript, and finished the figures and tables. CZ, ZYH and ZQH offered crucial content revision and language polishing. ZQH completed the final manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sun, Q., Wang, L., Zhang, C. et al. Cervical cancer heterogeneity: a constant battle against viruses and drugs. Biomark Res 10, 85 (2022). https://doi.org/10.1186/s40364-022-00428-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-022-00428-7