
Abstract
Background
The allocation of photosynthate among the parts of plants (e.g., leaves, wood tissues and roots) strongly regulates their growth, and this conditions the terrestrial carbon cycle. Recent studies have shown that atmospheric CO2 and climate change dominate the changes in carbon allocation in plants, but the magnitude and mechanism of its effects remain unclear.
Methods
The Community Atmosphere Biosphere Land Exchange (CABLE) model can accurately simulate the responses of carbon allocation to environmental changes. This study quantifies the contributions of four environmental factors—atmospheric CO2, temperature, precipitation, and radiation—on resource availability and carbon allocation from 1979 to 2014 by using the CABLE model.
Results
The results of the CABLE model showed that rising CO2 significantly reduced carbon allocation to the leaves of plants at a global scale, but the other three environmental factors exhibited contrasting effects that dominated the rise in carbon allocation to the leaves. The increased precipitation and CO2 significantly reduced the light availability and increased carbon allocation to the wooden parts of plants. By contrast, the rising temperature reduced the water availability, resulting in a decrease in carbon allocation to the wooden parts. All four environmental factors consistently exhibited negative effects on carbon allocation to the roots, with rising precipitation causing the largest reduction in carbon allocation to them. Moreover, except for CO2, the effects of the other three environmental factors were heterogeneous owing to their variable interactions in different regions.
Conclusions
The CABLE model can accurately represent the mechanisms of response of resource availability and carbon allocation to environmental changes. Our study highlights the substantial environmental regulation of global carbon allocation. The responses of carbon allocation to global environmental changes need to be extensively studied through ecosystem models based on different hypotheses.
Similar content being viewed by others


Background
Carbon allocation, or the allocation of photosynthate among the parts of plants (e.g., leaves, wood tissues and roots), is one of the most important physiological processes in nature (Xia et al. 2019; Sierra et al. 2022). It determines not only plant growth, but also numerous processes of the ecosystem, including decomposition, carbon and nitrogen sequestration, and the exchange of water between plans and the atmosphere (Aber and Melillo 1991). The carbon allocated to the woody tissues of plants has a longer residence time than that in their leaves or roots, where this significantly influences the global carbon budget (Friedlingstein et al. 1999; McMurtrie and Dewar 2013; Xia et al. 2017; Wei et al. 2022). Carbon allocation between the wooden and non-wooden parts of plants also determines the quality and rate of decomposition of litter (Bird and Torn 2006). Ise et al. (2010) found a large variation in estimates of wooden biomass that stemmed from different assumptions about the coefficients of carbon allocation. The ratio of carbon allocated to the stems of plants in forests according to the vegetation integrative simulator for trace gases (VISIT) model was found to be higher than that of the Biome-BGC model (e.g., 0.517 vs. 0.3 for tropical forests), because of which the wooden biomass simulated by the former model was 178 Pg C higher than that of the latter (Ise et al. 2010). Therefore, reliable estimates of the changes in carbon allocation are crucial for reducing uncertainty in modeling the global carbon cycle.
Carbon allocation varies substantially over space and time in the context of global climate change, but the effects of climate change on carbon allocation are not well understood. For example, a rising CO2 concentration has exhibited divergent effects on carbon allocation in two long-term experiments on CO2 enrichment—an increase in carbon allocation to the wooden parts of plants at Duke but a decrease in it at Oak Ridge—as a result of the different responses of the availability of nitrogen in soil to CO2 enrichment (De Kauwe et al. 2014). Moreover, previous studies have highlighted the inconsistent responses of carbon allocation to the components of plants under warming conditions. Warmer climate has been reported to increase the NPP in boreal forests but reduce the growth of wood, while the limitation in nutrients resulting from rising temperatures leads to greater carbon allocation to the roots (Lapenis et al. 2013). However, there is a dearth of research on how carbon allocation among the components of plants varies with different environmental variables at the global scale, where this is important for understanding the effects of carbon allocation on the terrestrial carbon budget (Xia et al. 2015; Wang et al. 2023).
The modeled response mechanism of carbon allocation to environmental changes remains unclear. Theoretically, the dynamic characteristics of plant carbon allocation result from their response in terms of using carbon to capture the most limiting resources (e.g., light, water, and nutrients) (Sharpe and Rykiel 1991; Kobe et al. 2010). The limitation of critical resources, such as water and nutrients, can enhance carbon allocation to the roots of plants to increase their uptake and alleviate resource deficits (Ikegami et al. 2007; Matzek 2011; Peng et al. 2017; Brunn et al. 2022). Previous studies have suggested that an increase in the ratio of biomass of the roots to that of the shoots may play an important role in the adaptation of plants to drought (Nelson et al. 2004). By contrast, the limitation in the radiation absorbed by them increases carbon allocation to the leaves at the expense of that to the roots to increase absorption of incoming photosynthetically active radiation by them (Wang et al. 2006). Whether the observational data support the assumptions of these models of carbon allocation, and whether the modeled mechanism of the responses of plants to the processes of carbon allocation are correct still require further study.
In this study, we report five modeling experiments based on the Community Atmosphere Biosphere Land Exchange (CABLE) model to illustrate how carbon allocation varies with environmental variables. The objectives of this study are to (1) illustrate the effects of multiple environmental factors on resource availability and carbon allocation, and (2) reveal the response mechanisms of carbon allocation and resource availability to environmental changes according to the CABLE model.
Methods
CABLE model
The CABLE model is a global land-surface model that has been integrated with the CASA (Carnegie–Ames–Stanford approach)-CNP (carbon–nitrogen–phosphorous) model (CASA-CNP). This model can simulate the carbon, nitrogen, phosphorus, water, and energy cycles in terrestrial ecosystems (Wang et al. 2010; Zhang et al. 2011). Four phases of leaf phenology determine the growth and patterns of carbon allocation in leaves in the CABLE model. These are the phases of maximum and steady leaf growth, leaf senescence, and dormancy or no leaves. During the first phase, the ratio of carbon allocation to the leaves (aleaf) is set to 0.8, while those to the wood tissues (awood) and roots (aroot) are set to 0.1 for wooden plant functional types (PFTs), and zero and 0.2, respectively, for non-wooden PFTs. The ratios of carbon allocation (i.e., aleaf_p2, awood_p2, and aroot_p2) in the second phase (phase 2) are calculated according to the resource limitation carbon allocation model. During the third and fourth phases, aleaf is set to zero while awood and aroot are calculated by using the allocation ratios of phase 2 (awood = awood_p2/( awood_p2 + aroot_p2), aroot = aroot_p2/( awood_p2 + aroot_p2)). For evergreen PFTs, the leaf phenology and carbon allocation persist in phase 2 throughout the year.
During the phase of steady leaf growth (i.e., phase 2), the allocation of photosynthate among the components of plants in the CABLE model is regulated by the availability of light, water, and nitrogen (hereinafter called the resource limitation model). This model assumes that the greatest part of the gain in carbon is allocated to the component of the plant that most limits its growth (Fig. 1). If the availability of light is limiting its growth, the plant allocates more carbon to the wood tissues, such as stems. If water and nitrogen are limiting growth, the plant allocates more carbon to the roots (Fig. 1). The resource limitation carbon allocation model uses the scheme developed by Friedlingstein et al. (1999). The allocation coefficients of woody biomes for their leaves (aleaf), woody tissues (awood), and roots (aroot) are as follows:

Fractions of allocation to the leaf (a), wood (b), and root (c) of the plant as functions of light (L), and the minimum values of the availability of water and nitrogen (Min(Water, Nitrogen)) for tree plant functional types (PFTs), with values of l0 = 0.4, s0 = 0.3, and r0 = 0.3. Fractions of allocation for the leaves (d) and roots (e) in grass and crop PFTs, with values of l0 = 0.7 and r0 = 0.3
For non-woody biomes, the allocation coefficients are as follows:
where min (W, N) represents the minimum values of the availability of water (W) and nitrogen (N). The parameters r0 and s0 in the CABLE model describe the proportional allocation to the roots and wood tissues when resources are not limited. For the tree PFTs, both r0 and s0 are set to 0.3, with an allocation of 0.4 to the leaves (l0). For C3 grass and crop PFTs, r0 = 0.3 and l0 = 0.7 (Fig. 1).
Light availability (L) is calculated by:
where LAI is the leaf area index.
Water availability (W) is calculated by:
where fri is the fraction of root in the ith (n = 6) layer of soil, Swi is the volumetric moisture content of soil in the ith layer, and Swilt and Sfield are the volumetric soil moisture contents at the wilting point and the field capacity soil moisture content, respectively.
Nitrogen availability (N) is calculated as the product of the temperature of soil (Ts) and moisture (Ws) (Friedlingstein et al. 1999), where Ts is calculated based on a standard Q10 equation (Potter et al. 1993) and Ws is calculated by using Eq. (7):
where Tsoili (°C) is the temperature of soil in the ith layer, and Q10 = 2.0.
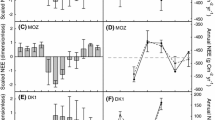
The performance of the resource limitation carbon allocation scheme in CABLE model was evaluated at five natural observation sites and four global change manipulation experiments (precipitation reduction, warming, and elevated atmospheric CO2) in various ecosystems (Table 1 and Fig. 2b, Xia et al. 2017). The experiment at the C4 grass site in Oklahoma showed that the grass allocated more carbon to its roots under warming conditions. A precipitation control experiment at a rainforest site in Caxiuana showed that trees had decreased values of awood, and increased values of aroot and aleaf under drought conditions. The elevated atmospheric CO2 (eCO2) experiment at the evergreen needleleaf tree at the Duke site showed that the tree allocated more carbon to wood under the eCO2 condition. The eCO2 experiment at the deciduous broadleaf tree of the Oak Ridge site showed that the tree allocated more carbon to the roots under the eCO2 condition. The CABLE was able to accurately simulate the responses of carbon allocation to environmental changes at those experimental sites (Xia et al. 2017). We thus used the CABLE model to explore the responses of plant carbon allocation to environmental changes (i.e., the dataset on precipitation, air temperature, and incident shortwave radiation obtained from the Modern-Era Retrospective Analysis for Research and Applications (MERRA), and data on atmospheric CO2 concentration).

Global distribution of plant functional types in the CABLE model (a). The global ecological zones, and locations of the observation sites for carbon allocation (b)
Input data for CABLE model
The CABLE model was driven by the 1-hourly MERRA dataset (Global Modeling and Assimilation Office 2004), which contained data on the air temperature (Tair), precipitation (Prec), specific humidity (Qair), incident longwave radiation (Lwd), incident shortwave radiation (Swd), pressure (Ps) and wind speed (Wind) from 1979 to 2014. The spatial resolution of the data was 0.5° latitude by 0.67° longitude. The annual mean CO2 concentration from 1850 to 2014 was obtained from research by Keeling and Whorf (2005). The fixed PFTs for 2005 were based on data from Lawrence et al. (2012) (Fig. 2a). The PFTs included evergreen needleleaf tree (ENT), evergreen broadleaf tree (EBT), deciduous needleleaf tree (DNT), deciduous broadleaf tree (DBT), shrub (SHB), C3 grass (C3), C4 grass (C4), tundra (TDR), and C3 crop (CROP). The global ecological zone for forest reporting by the FAO was used to analyze the results (Fig. 2b, FAO 2012).
Simulations
To determine the influence of the four environmental variables, i.e., atmospheric CO2 concentration, precipitation, air temperature, and incident shortwave radiation, on the spatiotemporal patterns of carbon allocation, we performed five simulations (Table 2): (1) a control simulation, forced by the original MERRA climatic data and historical CO2 observations from 1979 to 2014; (2) a CO2_fixed simulation, forced by the original MERRA climatic data, with the atmospheric CO2 concentration fixed at the 1979 level (337.0 ppm); and (3) Prec_fixed, (4) Tair_fixed and (5) Swd_fixed simulations, forced by the data on the hourly precipitation, near surface air temperature and incident shortwave radiation of 1979 for all simulation years and historical CO2 observations, respectively. We then quantified the effects of changes in each environmental variable at an inter-annual scale after 1979 on the carbon allocation by calculating the difference in ratios of carbon allocation between the control simulation and simulations in which the inter-annual variations of the given environmental variable after 1979 were not considered (e.g., Prec_fixed simulation).
We conducted a spin-up run and two transient runs for the simulations. For the spin-up run, we ran the model 20 times by recycling the MERRA climatic datasets from 1979 to 2014. The atmospheric CO2 concentration at the 1850 level (280.0 ppm) was used for this. Once the model had reached equilibrium state, it was run in the first transient mode (1850–1978) with varying CO2 concentrations while recycling the MERRA climate datasets from 1979 to 2014. The second transient mode covered the period 1979–2014, and the corresponding CO2 concentrations and climatic variables were used according to the design of the five simulations (Table 2).
Results
Effects of inter-annual changes in environmental variables on carbon allocation
The environmental variables significantly changed from 1979 to 2014 across the globe. Along with a rising atmospheric CO2 concentration, the global mean surface temperature increased by 0.72 °C over 36 years (Fig. 3d). Moreover, the incident shortwave radiation and precipitation exhibited a trend of growth, with rates of increase of 0.12 W m−2 yr−1 and 2.27 mm yr−1, respectively (Fig. 3f, b). However, the climate variables were substantially spatially heterogeneous. Increased temperatures were observed in all plant functional types. The largest increase in the temperature was observed in the tundra, 0.04 °C yr−1 (Table 3), while the lowest increase in temperature increases were observed in evergreen broadleaf tree, at 0.009 °C yr−1 (Table 3). On average, the precipitation showed a trend of decline from 1979 to 2014 only in the evergreen needleleaf tree and the deciduous needleleaf tree, while the largest increase in precipitation occurred in evergreen broadleaf tree (Table 3). Radiation exhibited a large spatial heterogeneity, and significant decreased in the evergreen broadleaf tree, shrub and C4 grass (Table 3).

The trends and inter-annual variations of the global mean precipitation (a, b), air temperature (c, d), incident shortwave radiation (e, f) and atmospheric CO2 concentration (f) from 1979 to 2014
Climate change significant influenced resource availability. The global mean light availability showed a significantly decrease trend from 1979 to 2014 (Fig. 4a and Table 4). The model of carbon allocation in case of resource limitation showed that changes in light availability mainly depended on changes in the LAI. The largest decrease in light availability was caused by the greatest increase in the LAI as a result of the fertilization effect of CO2 (Fig. 4a and d). Compared with the control simulation, the changes in air temperature increased the magnitude of global light availability (Fig. 4a and Table 4). The global precipitation exhibited a trend of an increase (Fig. 3b), and led to an increase in global water availability (i.e., soil moisture) from 1979 to 2014 (Fig. 4b and Table 4). Water availability exhibited the largest decrease in the Tair_fixed simulation (Fig. 4b and Table 4), reflecting the dominant influence of the rising temperature on it. The global mean nitrogen availability showed a significant trend of increase from 1979 to 2014 as a result of the dominant influence of increasing precipitation (Fig. 4c and Table 4).

Inter-annual variations in the global mean availability of light (a), water (b), and nitrogen (c), and leaf area index (LAI, d) in the control simulation (right vertical axis and the red line). Differences in the four variables between the control simulation, and the fixed CO2 (CO2_fixed), fixed precipitation (Prec_fixed), fixed temperature (Tair_fixed), and fixed shortwave radiation (Swd_fixed) simulations (left vertical axis). For example, “Light_Control–Light_CO2_fixed” refers to the difference in light availability between the control and the CO2_fixed simulations, and negative values indicate that changes in CO2 led to a decrease in light availability
The effect of rising CO2 levels on light availability was highly consistent across the globe. More than 83.54% of the global areas of land exhibited a significantly decreased light availability (Fig. 5a) as the LAI increased (Fig. 5d). Increased CO2 concentrations led to an increase in water availability in 63.46% of the global areas of land (Fig. 5b), which in turn controlled the pattern of nitrogen availability except in the northern Amazon (Fig. 5c). Increased CO2 resulted in decreased soil moisture in semi-arid and arid ecological zones (Figs. 2b and 5b). The simulations of the change in precipitation showed that the change in the moisture content of soil, nitrogen availability, and the LAI showed similar patterns to that of the trend of precipitation (Figs. 3a, 5f, g, and h). Thus, light availability exhibited the opposite pattern of change to that of precipitation (Figs. 3a and 5e).

Trends of the differences in the availability of light (a), water (b), and nitrogen (c) as well as the leaf area index (LAI, d) between the control simulation and the fixed CO2 (i.e., CO2_fixed in Table 2) simulation. Trends of differences in the availability of light (e), water (f), and nitrogen (g) as well as the leaf area index (LAI, h) between the control simulation and the fixed precipitation (i.e., Prec_fixed in Table 2) simulation
Compared with those of the atmospheric CO2 and precipitation, the effects of the temperature and shortwave radiation on the availability of resources and the LAI were more complicated (Fig. 6). Increased temperature led to a decrease in the availability of water in soil in 70.97% of the global area of land (Fig. 6b) and increased nitrogen availability at high northern latitudes (60° N to 90° N, Fig. 6c). The decrease in the LAI in the Congo rainforest due to warming was consistent with the decrease in water availability in this region (Figs. 3c, 6b, and d). The effects of shortwave radiation on water availability exhibited the oppose pattern to that of the shortwave radiation (Figs. 3e and 6f). In the Northern Hemisphere, the effect of the shortwave radiation on the LAI was consistent with its own trend (Figs. 3e and 6h), while the increase in the LAI was consistent with that in water availability caused by the change in the shortwave radiation in the Southern Hemisphere (Figs. 3e, 6f, and h).

Trends of the differences in the availability of light (a), water (b), and nitrogen (c) as well as the leaf area index (LAI, d) between the control simulation and the fixed temperature (i.e., Tair_fixed in Table 2) simulation. Trends of differences in the availability of light (e), water (f), and nitrogen (g), and the leaf area index (LAI, h) between the control simulation and the fixed incident shortwave radiation (i.e., Swd_fixed in Table 2) simulation
Effects of climate change on carbon allocation
Global climate change has led to substantial changes in carbon allocation among the three parts of plants, i.e., roots, leaves, and wood tissues. The carbon allocation to the leaves (aleaf) and wood (awood) significantly increased from 1979 to 2014 (Fig. 7a, b, and Table 5), while its allocation to roots (aroot) decreased in this period (Fig. 7c and Table 5). However, the climatic variables had different effects on the changes in carbon allocation. The allocation to leaves was significantly reduced by rising atmospheric CO2 concentration (Fig. 7a and Table 5). By contrast, the changes in precipitation and shortwave radiation enhanced the global mean carbon allocation to the leaves (Fig. 7a and Table 5). The rising atmospheric CO2 and precipitation led to significant increases in awood (Fig. 7b and Table 5), but it significantly decreased with increasing temperature (Fig. 7b and Table 5). The rising precipitation contributed to the largest decrease in aroot (Fig. 7c and Table 5).

Inter-annual variations in the global mean carbon allocation to the leaves (aleaf, a), wood (awood, b), and roots (aroot, c) of plants in the control simulation (right vertical axis and red line). Differences in the ratios of carbon allocation between the control simulation, and the fixed CO2 (CO2_fixed), fixed precipitation (Prec_fixed), fixed temperature (Tair_fixed), and fixed shortwave radiation (Swd_fixed) simulations (left vertical axis). For example, “aleaf_Control–aleaf_CO2_fixed” refers to the difference in values of aleaf between the control and the CO2_fixed simulations, while negative values mean that the changes in CO2 concentration led to decrease in aleaf. CO2 atmospheric CO2 concentration; Prec precipitation; Tair air temperature; Swd incident shortwave radiation
At the regional scale, the rising atmospheric CO2 concentration increased awood to over 80% in woody plants (Fig. 8b). The allocation to leaves decreased with increasing CO2 concentration in the wooden biomes, except at high northern latitudes (60° N to 90° N) and in Australia (Fig. 8a), while aroot exhibited a widespread decrease (Fig. 8c). In response to rising atmospheric CO2 concentration, aroot consistently increased and aleaf consistently decreased in grasslands (Figs. 2, 8a, and c). Increased precipitation enhanced the values of awood (Figs. 8e and 3a), while it led to the largest increase in values of awood in shrubs (Fig. 8e). By contrast, aroot decreased with increasing precipitation (Figs. 8f and 3a). The response of aleaf to precipitation was complicated. For example, it increased with the increasing precipitation in Australia but increased with decreasing precipitation in the Congo Basin (Figs. 8d and 3a). The responses of the ratios of carbon allocation to the temperature were more complicated than those to the CO2 concentration and precipitation because of the divergent influence of the temperature on nitrogen availability (Figs. 8g, h, i, and 6c).

Trends of the differences in carbon allocation to the leaves (aleaf), wood (awood), and roots (aroot) between the control simulation, and fixed CO2 (CO2_fixed), fixed precipitation (Prec_fixed), fixed temperature (Tair_fixed), and fixed shortwave radiation (Swd_fixed) simulations. For example, “aleaf_CO2” refers to the difference in carbon allocation to leaves between the control and the CO2_fixed simulations. CO2 atmospheric CO2 concentration; Prec precipitation; Tair air temperature; Swd incident shortwave radiation
Discussion
The CABLE model was able to accurately simulate the processes of response of resource availability and carbon allocation to environmental changes. Elevated air temperature at the C4 grass site in Oklahoma increased the temperature of soil and reduced the availability of water in it, resulting in more carbon being allocated to the roots to efficiently capture water (McCarthy and Enquist 2007; Xu et al. 2012). A reduction in the rainfall reduced the availability of water in soil while increasing aroot and reducing awood at the Caxiuana site (da Costa et al. 2014). The CABLE model also correctly simulated the decreased availability of water in soil and increased aroot under warming and drought conditions (Xia et al. 2017). At the Duke site, the increase in CO2 concentration enhanced the soil moisture by reducing stomatal conductance and transpiration per unit area of the leaf, and reduced light availability by stimulating leaf growth (McCarthy et al. 2010; Ward et al. 2013; De Kauwe et al. 2014). Both of these effects stimulated the values of awood. The CABLE model was able to accurately represent the effects of eCO2 on the LAI, light and water availability, and carbon allocation (Xia et al. 2017). The increase in aroot at the Oak Ridge site might have been caused by a reduction in the availability of nitrogen in soil (Norby et al. 2010). The CABLE model could not completely reproduce the changes in carbon allocation at this site partly because of the complicated effects of eCO2 on this deciduous forest ecosystem.
The rising atmospheric CO2 concentration enhanced the LAI over most areas of the globe (Fig. 5d). It ultimately influenced carbon allocation by altering the LAI, which is an important property of the ecosystem that determines the surface temperature and soil water balance (Luyssaert et al. 2014; Shen et al. 2015). Rising CO2 concentrations stimulated photosynthesis, which increased the LAI (Zhu et al. 2016). Our simulations showed larger increases in the LAI over low latitudes (Fig. 5d), which is consistent with the results of previous studies (Zhu et al. 2016). The enhanced LAI reduced light availability according to the self-shading rule, but had a weak effect on the availability of water and nitrogen (Table 4). Consequently, changes in these resources enhanced the allocation of carbon to wooden parts of plants and benefited the carbon storage of terrestrial ecosystems (Ise et al. 2010; Poorter et al. 2012).
The effects of eCO2 on water availability were controlled by two processes in the CABLE model: (1) eCO2 decreased the stomatal conductance and transpiration, which led to an increase in the soil moisture. (2) The increased LAI enhanced transpiration and reduced the soil moisture. The results of this study as well as previous ones support the mechanism of response of water availability to the eCO2 as represented by the CABLE model (De Kauwe et al. 2013; Cheng et al. 2014). The decrease in the soil moisture in water-limited PFTs (i.e., shrubs and C3 grass) was caused by the increase in the LAI (Figs. 2, 5b, and d), where this is consistent with the study by Donohue et al. (2009). The C3 grass allocated more carbon to roots under water stress condition (Fig. 8c). With regard to the other PFTs, an increase in CO2 concentration mostly led to an increase in the soil moisture (Fig. 5b). The decrease in carbon allocation to the roots in boreal evergreen needleleaf tree in particular was determined by the increase in the availability of water in soil (Figs. 2, 5b, and 8c).
The effects of precipitation on carbon allocation were substantially spatially heterogeneous owing to inhomogeneous changes in the precipitation and the responses of the ecosystem. Decreased precipitation was found in the most areas of the Northern Hemisphere, and restricted the growth of vegetation and reduced the LAI. However, the decreased precipitation increased soil temperature in the high latitude regions of the Northern Hemisphere (Figs. 3a and 9b) and stimulated vegetation growth (Yuan et al. 2014). Moreover, the increased precipitation in the Amazon rainforest did not enhance the LAI, but resulted in a slight decrease in it instead (Figs. 3a and 5h). One potential cause of this was the reduced radiation with increased precipitation (Myneni et al. 2007). Precipitation dominated the changes in soil moisture as well as nitrogen availability according to the CABLE model (Fig. 5f and g). In general, decreased precipitation reduced the soil moisture and the LAI, and increased the availability of light, while changes in both the soil moisture and light enhanced carbon allocation to the roots but restricted that to the wooden parts and leaves of plants (Waring 1991; Poorter et al. 2012; Perrin and Mitchell 2013).

Differences in the temperature limitation factor [Ts, see Eq. (9)] in the availability of nitrogen between the control simulation, and the simulations in which certain climatic variables were kept constant. For example, “Ts_CO2” refers to the difference in the temperature limitation factor between the control simulation, and the CO2_fixed simulation. CO2 atmospheric CO2 concentration; Prec precipitation; Tair air temperature; Swd incident shortwave radiation
The effects of the temperature and shortwave radiation on carbon allocation are mainly accomplished by influencing the soil moisture and LAI in the CABLE model. Warming increased evaporation and reduced the soil moisture (Figs. 3c, 3e, 6b, and f), where this outcomes is consistent with that reported in previous studies (Xu et al. 2013; Samaniego et al. 2018). The reduced soil moisture led to increased carbon allocation to the roots of plants (Fig. 8i). Warming resulted in an increased LAI in most regions with boreal evergreen needleleaf tree and the tundra (Figs. 2 and 6d), where this is consistent with findings based on remote sensing observations (Piao et al. 2020). This process increased the allocation of carbon to wood tissues in the relevant PFTs (Fig. 8h).
The responses of carbon allocation to climate change were influenced by several uncertainties in the former in the CABLE model. Empirical equations for the availability of nitrogen led to uncertainties in the dynamics of the response of nitrogen in the soil to climate change and its effects on carbon allocation. This empirical equation assumed that spatial variability in the mineralization of nitrogen and the decomposition of the organic matter in soil were identical (Townsend et al. 1995). The availability of nitrogen is calculated as the product of the abiotic factors of the temperature and moisture (Potter et al. 1993). The abiotic factor of the temperature has a standard Q10 formulation, while the availability of nitrogen increases with the temperature. However, the simulations on warming suggested that elevated temperature tended to increase the net nitrogen mineralization in the first year but reduced decrease it in the subsequent years, a process that is attributable to stimulated plant growth and belowground carbon allocation that enhanced the immobilization of microbial nitrogen (Wan et al. 2004). Moreover, the soil moisture strongly regulated the aerobic and anaerobic conditions, which determined the magnitude of nitrification and denitrification as well as the availability of nitrogen (Xu and Prentice 2008). However, the effects of soil moisture on the availability of nitrogen were largely spatially heterogeneous.
Moreover, light condition is one of the most important environmental factors influencing carbon allocation (Iszkuło and Boratyński 2004, 2006). However, the availability of light was not calculated from downwelling radiation, but from leaf area index because the plant at the lower canopy might have had increased access to light through increased allocation of carbon to the stem (Friedlingstein et al. 1999). This method has been used in several models, including Canadian terrestrial ecosystem model (CTEM; Arora and Boer 2005), organizing carbon and hydrology in dynamic Ecosystems (ORCHIDEE; Krinner et al. 2005), and adaptive dynamic global vegetation model (aDGVM; Scheiter and Higgins 2009). Numerous experiments have shown that plants allocate a higher fraction of carbon to stems and leaves under a low incident radiation above the canopy (Poorter et al. 2012). For example, compared with plants grown at higher irradiance, shaded plants generally have a higher specific leaf area (SLA), and allocate more biomass to the leaves than to the roots for the efficient capture of light (Poorter 1999; Franck et al. 2007; Feng and Li 2007). The effect of downwelling radiation on the carbon allocation to the leaves should be considered in the terrestrial ecosystem model (Chen et al. 2020).
Conclusions
In this study, we used the global land-surface model CABLE to investigate the contributions of changes in atmospheric CO2 concentration, precipitation, air temperature, and incident shortwave radiation to global carbon allocation from 1979 to 2014. The model showed that rising atmospheric CO2 concentration had reduced the availability of light and increased that of water at a global scale, where this had enhanced carbon allocation to the wood tissues of plants. By contrast, the global mean precipitation exhibited an increasing trend, leading to an increase in the availability of water and a decrease in that of light. This had led to a reduction in carbon allocation to the roots and an increase in allocation to the wooden parts of plants. Rising atmospheric CO2 concentration had led to a uniform increase in carbon allocation to wooden parts of the plants as well. However, the responses of the ratios of carbon allocation to temperature exhibited a large spatial heterogeneity, and were more complicated than those of the CO2 concentration and precipitation because of the opposite effects of air temperature to the soil temperature and moisture. Although the observations of four manipulation experiments show that the CABLE model can accurately reflect the response mechanism of resource availability and carbon allocation to environmental change, there is still uncertainty in the inference of carbon allocation for different plant functional groups at the global scale. More studies are needed to model and verify the responses of carbon allocation by plants to multiple environmental changes based on models and field experiments in the future.
Availability of data and materials
The data sets used in the current study are available from the corresponding author on reasonable request.
Abbreviations
- CABLE:
-
Community Atmosphere Biosphere Land Exchange
- NPP:
-
Net primary productivity
- CASA-CNP:
-
CASA (Carnegie–Ames–Stanford Approach)-CNP (carbon–nitrogen–phosphorous)
- VISIT:
-
Vegetation Integrative SImulator for Trace gases
- eCO2 :
-
Elevated atmospheric CO2
- aleaf:
-
Allocation coefficients for leaves
- awood:
-
Allocation coefficients for woody tissues
- aroot:
-
Allocation coefficients for roots
- L:
-
Light availability
- W:
-
Water availability
- N:
-
Nitrogen availability
- MERRA:
-
Modern-Era Retrospective analysis for Research and Applications
- Tai:
-
Air temperature
- Prec:
-
Precipitation
- Swd:
-
Incident shortwave radiation
- Lwd:
-
Incident longwave radiation
- Qair:
-
Specific humidity
- Wind:
-
Wind speed
- Ps:
-
Pressure
- PFTs:
-
Plant functional types
- ENT:
-
Evergreen needleleaf tree
- DNT:
-
Deciduous needleleaf tree
- EBT:
-
Evergreen broadleaf tree
- DBT:
-
Deciduous broadleaf tree
- SHB:
-
Shrub
- C3:
-
C3 grass
- C4:
-
C4 grass
- TDR:
-
Tundra
- CROP:
-
C3 crop
- LAI:
-
Leaf area index
- CTEM:
-
Canadian Terrestrial Ecosystem Model
- ORCHIDEE:
-
Organizing Carbon and Hydrology in Dynamic Ecosystems
- aDGVM:
-
Adaptive Dynamic Global Vegetation Model
- SLA:
-
Specific leaf area
References
Aber JD, Melillo JM (1991) Terrestrial ecosystems. Saunders, Philadelphia, Pennsylvania, USA
Arora VK, Boer GJ (2005) A parameterization of leaf phenology for the terrestrial ecosystem component of climate models. Global Change Biol 11(1):39–59
Bird JA, Torn MS (2006) Fine roots vs. needles: a comparison of 13C and 15N dynamics in a ponderosa pine forest soil. Biogeochemistry 79(3):361–382
Brunn M, Hafner BD, Zwetsloot MJ, Weikl F, Pritsch K, Hikino K, Ruehr NK, Sayer EJ, Bauerle TL (2022) Carbon allocation to root exudates is maintained in mature temperate tree species under drought. New Phytol 235(3):965–977
Chen X, Maignan F, Viovy N, Bastos A, Goll D, Wu J, Liu L, Yue C, Peng S, Yuan W, da Conceição AC, O’Sullivan M, Ciais P (2020) Novel representation of leaf phenology improves simulation of Amazonian evergreen forest photosynthesis in a land surface model. J Adv Model Earth Sy 12(1):e2018MS001565
Cheng L, Zhang L, Wang YP, Yu Q, Eamus D (2014) Quantifying the effects of elevated CO2 on water budgets by combining FACE data with an ecohydrological model. Ecohydrology 7(6):1574–1588
da Costa AC, Metcalfe DB, Doughty CE, de Oliveira AA, Neto GF, da Costa MC, Silva Junior JdA, Aragão LE, Almeida S, Galbraith DR (2014) Ecosystem respiration and net primary productivity after 8–10 years of experimental through-fall reduction in an eastern Amazon forest. Plant Ecol Divers 7(1–2):7–24
De Kauwe MG, Medlyn BE, Zaehle S, Walker AP, Dietze MC, Hickler T, Jain AK, Luo Y, Parton WJ, Prentice IC (2013) Forest water use and water use efficiency at elevated CO2: a model-data intercomparison at two contrasting temperate forest FACE sites. Global Change Biol 19(6):1759–1779
De Kauwe MG, Medlyn BE, Zaehle S, Walker AP, Dietze MC, Wang YP, Luo Y, Jain AK, El-Masri B, Hickler T (2014) Where does the carbon go? A model–data intercomparison of vegetation carbon allocation and turnover processes at two temperate forest free-air CO2 enrichment sites. New Phytol 203(3):883–899
Donohue RJ, McVICAR TR, Roderick ML (2009) Climate-related trends in Australian vegetation cover as inferred from satellite observations, 1981–2006. Global Change Biol 15(4):1025–1039
FAO (2012) Global ecological zones for FAO forest reporting: 2010 update. FAO Forestry Paper 179. FAO, Rome, Italy
Feng Y-L, Li X (2007) The combined effects of soil moisture and irradiance on growth, biomass allocation, morphology and photosynthesis in Amomum villosum. Agroforest Syst 71(2):89–98
Franck N, Winkler S, Pastenes C, Infante R (2007) Acclimation to sun and shade of three accessions of the Chilean native berry-crop murta. Agroforest Syst 69(3):215–229
Friedlingstein P, Joel G, Field CB, Fung IY (1999) Toward an allocation scheme for global terrestrial carbon models. Global Change Biol 5(7):755–770
Global Modeling and Assimilation Office (2004) File Specification for Goddard Earth Observing System Data Assimilation System (GEOSDAS) Gridded Output Version 5.3, Report. NASA Goddard Space Flight Cent, Greenbelt, MD
Hui D, Jackson RB (2006) Geographical and interannual variability in biomass partitioning in grassland ecosystems: a synthesis of field data. New Phytol 169(1):85–93
Ikegami M, Whigham DF, Werger MJ (2007) Responses of rhizome length and ramet production to resource availability in the clonal sedge Scirpus olneyi A. Gray. Plant Ecol 189(2):247–259
Ise T, Litton CM, Giardina CP, Ito A (2010) Comparison of modeling approaches for carbon partitioning impact on estimates of global net primary production and equilibrium biomass of woody vegetation from MODIS GPP. J Geophys Res Biogeosci 115:G04025. https://doi.org/10.1029/2010JG001326
Iszkuło G, Boratyński A (2004) Interaction between canopy tree species and European yew Taxus baccata (Taxaceae). Polish J Ecol 52(4):523–531
Iszkuło G, Boratyński A (2006) Analysis of the relationship between photosynthetic photon flux density and natural Taxus baccata seedlings occurrence. Acta Oecol 29(1):78–84
Keeling CD, Whorf TP (2005) Atmospheric CO2 records from sites in the SIO air sampling network. In: Trends: A Compendium of Data on Global Change. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy, Oak Ridge.
Kobe RK, Iyer M, Walters MB (2010) Optimal partitioning theory revisited: nonstructural carbohydrates dominate root mass responses to nitrogen. Ecology 91(1):166–179
Krinner G, Viovy N, de Noblet-Ducoudré N, Ogée J, Polcher J, Friedlingstein P, Ciais P, Sitch S, Prentice IC (2005) A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Global Biogeochem Cycles 19:GB1015. https://doi.org/10.1029/2003GB002199
Lapenis AG, Lawrence GB, Heim A, Zheng C, Shortle W (2013) Climate warming shifts carbon allocation from stemwood to roots in calcium-depleted spruce forests. Global Biogeochem Cycles 27(1):101–107
Lawrence PJ, Feddema JJ, Bonan GB, Meehl GA, O’Neill BC, Oleson KW, Levis S, Lawrence DM, Kluzek E, Lindsay K (2012) Simulating the biogeochemical and biogeophysical impacts of transient land cover change and wood harvest in the community climate system model (CCSM4) from 1850 to 2100. J Clim 25(9):3071–3095
Luyssaert S, Jammet M, Stoy PC, Estel S, Pongratz J, Ceschia E, Churkina G, Don A, Erb K, Ferlicoq M (2014) Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat Clim Change 4(5):389–393
Matzek V (2011) Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment. Biol Invasions 13(12):3005–3014
McCarthy M, Enquist B (2007) Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct Ecol 21(4):713–720
McCarthy HR, Oren R, Johnsen KH, Gallet-Budynek A, Pritchard SG, Cook CW, LaDeau SL, Jackson RB, Finzi AC (2010) Re-assessment of plant carbon dynamics at the Duke free-air CO2 enrichment site: interactions of atmospheric [CO2] with nitrogen and water availability over stand development. New Phytol 185(2):514–528
McMurtrie RE, Dewar RC (2013) New insights into carbon allocation by trees from the hypothesis that annual wood production is maximized. New Phytol 199(4):981–990
Myneni RB, Yang W, Nemani RR, Huete AR, Dickinson RE, Knyazikhin Y, Didan K, Fu R, Juárez RIN, Saatchi SS (2007) Large seasonal swings in leaf area of Amazon rainforests. Proc Natl Acad Sci USA 104(12):4820–4823
Nelson JA, Morgan JA, LeCain DR, Mosier AR, Milchunas DG, Parton BA (2004) Elevated CO2 increases soil moisture and enhances plant water relations in a long-term field study in semi-arid shortgrass steppe of Colorado. Plant Soil 259(1–2):169–179
Norby RJ, Warren JM, Iversen CM, Medlyn BE, McMurtrie RE (2010) CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc Natl Acad Sci USA 107(45):19368–19373
Peng Y, Guo D, Yang Y (2017) Global patterns of root dynamics under nitrogen enrichment. Global Ecol Biogeogr 26(1):102–114
Perrin PM, Mitchell FJ (2013) Effects of shade on growth, biomass allocation and leaf morphology in European yew (Taxus baccata L.). Eur J Forest Res 132(2):211–218
Piao S, Wang X, Park T, Chen C, Lian X, He Y, Bjerke JW, Chen A, Ciais P, Tømmervik H (2020) Characteristics, drivers and feedbacks of global greening. Nat Rev Earth Env 1(1):14–27
Poorter L (1999) Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits. Funct Ecol 13(3):396–410
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2012) Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol 193(1):30–50
Potter CS, Randerson JT, Field CB, Matson PA, Vitousek PM, Mooney HA, Klooster SA (1993) Terrestrial ecosystem production: a process model based on global satellite and surface data. Global Biogeochem Cycles 7(4):811–841
Samaniego L, Thober S, Kumar R, Wanders N, Rakovec O, Pan M, Zink M, Sheffield J, Wood EF, Marx A (2018) Anthropogenic warming exacerbates European soil moisture droughts. Nat Clim Change 8(5):421–426
Scheiter S, Higgins SI (2009) Impacts of climate change on the vegetation of Africa: an adaptive dynamic vegetation modelling approach. Global Change Biol 15(9):2224–2246
Sharpe PJ, Rykiel JrEJ (1991) Modelling integrated response of plants to multiple stresses. In: Mooney HA, Winner WE, Pell EJ (eds) Response of plants to multiple stresses. Academic Press, San Diego, pp 205–224
Shen M, Piao S, Jeong S-J, Zhou L, Zeng Z, Ciais P, Chen D, Huang M, Jin C-S, Li LZ (2015) Evaporative cooling over the Tibetan Plateau induced by vegetation growth. Proc Natl Acad Sci USA 112(30):9299–9304
Sierra CA, Ceballos-Núñez V, Hartmann H, Herrera-Ramírez D, Metzler H (2022) Ideas and perspectives: Allocation of carbon from net primary production in models is inconsistent with observations of the age of respired carbon. Biogeosciences 19(16):3727–3738
Townsend AR, Vitousek PM, Trumbore SE (1995) Soil organic matter dynamics along gradients in temperature and land use on the island of Hawaii. Ecology 76(3):721–733
Wan S, Norby RJ, Pregitzer KS, Ledford J, O’Neill EG (2004) CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots. New Phytol 162(2):437–446
Wang GG, Bauerle WL, Mudder BT (2006) Effects of light acclimation on the photosynthesis, growth, and biomass allocation in American chestnut (Castanea dentata) seedlings. For Ecol Manage 226(1):173–180
Wang Y, Law R, Pak B (2010) A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 7(7):2261–2282
Wang Q, Yan G, Luo X, Liu G, Wang H, Xing Y (2023) Symbiotic mycorrhizal types affect patterns of tree aboveground and belowground C allocation in Northeast China. Ecol Process 12:26
Ward EJ, Oren R, Bell DM, Clark JS, McCarthy HR, Kim H-S, Domec J-C (2013) The effects of elevated CO2 and nitrogen fertilization on stomatal conductance estimated from 11 years of scaled sap flux measurements at Duke FACE. Tree Physiol 33(2):135–151
Waring RH (1991) Responses of evergreen trees to multiple stresses. In: Mooney HA, Winner WE, Pell EJ (eds) Response of plants to multiple stresses. Academic Press, San Diego, pp 371–390
Wei N, Xia J, Zhou J, Jiang L, Cui E, Ping J, Luo Y (2022) Evolution of uncertainty in terrestrial carbon storage in earth system models from CMIP5 to CMIP6. J Clim 35(17):5483–5499
Xia J, Chen Y, Liang S, Liu D, Yuan W (2015) Global simulations of carbon allocation coefficients for deciduous vegetation types. Tellus B Chem Phys Meteorol 67:28016
Xia J, Yuan W, Wang Y-P, Zhang Q (2017) Adaptive carbon allocation by plants enhances the terrestrial carbon sink. Sci Rep 7(1):3341
Xia J, Yuan W, Lienert S, Joos F, Ciais P, Viovy N, Yp W, Wang X, Zhang H, Chen Y (2019) Global patterns in net primary production allocation regulated by environmental conditions and forest stand age: a model-data comparison. J Geophys Res Biogeosci 124(7):2039–2059
Xu R, Prentice I (2008) Terrestrial nitrogen cycle simulation with a dynamic global vegetation model. Global Change Biol 14(8):1745–1764
Xu X, Niu S, Sherry R, Zhou X, Zhou J, Luo Y (2012) Interannual variability in responses of belowground NPP and NPP partitioning to long-term warming and clipping in a tallgrass prairie. Global Change Biol 18:1648–1656
Xu W, Yuan W, Dong W, Xia J, Liu D, Chen Y (2013) A meta-analysis of the response of soil moisture to experimental warming. Environ Res Lett 8:044027
Xu Z, Ren H, Li M-H, Brunner I, Yin J, Liu H, Kong D, Lü X-T, Sun T, Cai J (2016) Experimentally increased water and nitrogen affect root production and vertical allocation of an old-field grassland. Plant Soil 412:369–380
Yuan W, Liu S, Dong W, Liang S, Zhao S, Chen J, Xu W, Li X, Barr A, Black TA (2014) Differentiating moss from higher plants is critical in studying the carbon cycle of the boreal biome. Nat Commun 5:4270. https://doi.org/10.1038/ncomms5270
Zhang Q, Wang Y, Pitman A, Dai Y (2011) Limitations of nitrogen and phosphorous on the terrestrial carbon uptake in the 20th century. Geophys Res Lett 38:L22701. https://doi.org/10.1029/2011GL049244
Zhu Z, Piao S, Myneni RB, Huang M, Zeng Z, Canadell JG, Ciais P, Sitch S, Friedlingstein P, Arneth A (2016) Greening of the Earth and its drivers. Nat Clim Change 6:791–795
Acknowledgements
The MERRA data used here are downloaded at http://disc.sci.gsfc.nasa.gov/daac-bin/FTPSubset.pl. The global ecological zones (GEZ) for FAO forest reporting can be downloaded at https://data.apps.fao.org/map/catalog/static/api/records/2fb209d0-fd34-4e5e-a3d8-a13c241eb61b.
Funding
This study was supported by grants from the National Natural Science Foundation of China (Grant No. 42001094), Scientific Research Project of Tianjin Municipal Education Commission, China (Grant No. 2020KJ002) and Natural Science Foundation of Tianjin, China (Grant No. 18JCQNJC78100).
Author information
Authors and Affiliations
Contributions
WY and JX conceived the research. WY supervised the research. JX and YC conducted the model runs. JX and YC conducted the data analysis. JX and WY drafted the manuscript. All authors contributed to the writing of the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xia, J., Chen, Y., Yuan, W. et al. The effects of multiple environmental factors on global carbon allocation. Ecol Process 12, 60 (2023). https://doi.org/10.1186/s13717-023-00477-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-023-00477-2