
Abstract
In this paper, we study a diffusion Holling–Tanner predator–prey model with ratio-dependent functional response and Simth growth subject to a homogeneous Neumann boundary condition. Firstly, we use iteration technique and eigenvalue analysis to get the local stability and a Hopf bifurcation at the positive equilibrium. Secondly, by choosing the constant related to delay as bifurcation parameter we obtain periodic solutions near the positive equilibrium. Besides, by using center manifold theory and normal form theory we reflect the stability with Hopf bifurcating periodic solution and bifurcating direction.
Similar content being viewed by others
1 Introduction
Mathematical ecology is a fast and active branch of biomathematics, and the dynamics of biological models are very rich. Thus it is very important for research. The predator–prey model is a very important population dynamics model. It is mainly reflected the factors influencing on the model in the functional response, based on ratio, age, or gender structure, reaction–diffusion, etc. The results of the study are mainly reflected in the stability of the equilibrium point, periodic solution, almost periodic solution, limit cycle, Hopf bifurcation, etc. The functional response is a very important factor to predator–prey model [1–5], It can be divided into several types, such as Hassell–Varley type, Holling types, Beddington–DeAngelies type, Crowley–Martin type, and so on.
In recent years, many authors investigate the diffusive predator–prey model with logistic prey growth rate [6–10] or strong Allee effect in prey [11]. It came out that this assumption is not realistic for a food-limited population under the effect of environmental toxicants. Thus the population dynamics growth restriction is based on the unused available resources [12–16]. This model, also known as the Holling–Tanner model, has been studied for both its mathematical properties and its efficacy for describing real ecological systems such as mite/spider mite, lynx/hare, sparrow/sparrow hawk, and so one by Holling [2], Tanner [3] and Wollkind, Collings, and Logan [4].
In particular, Yue and Wang [16] studied the stability and Hopf bifurcation of a diffusive Holling–Tanner predator–prey model with smith growth subject to Neumann boundary condition:
In realistic systems, delay exists everywhere. We have obtained many interesting conclusions since time delays may have very complex influence on the dynamical behaviors of systems. In the survey papers [12, 17–19], the authors deduced that delay difference equations are much more complicated than ordinary differential equations, because time delay can lead to unstable equilibrium and induce bifurcation. Many results on the Hopf bifurcation of diffusion system can be found in [20–27]. The Hopf bifurcation of the diffusive predator–prey system with delay subject to homogeneous Neumann boundary conditions was studied in [20–23]. The Hopf bifurcation for a delayed predator–prey diffusion system with Dirichlet boundary condition wasonsidered in [24]. The Hopf bifurcation for a predator-prey model with age structure was studied in [27].
Chen et al. [20] have considered a delayed diffusive Leslie–Gower predator–prey system with homogeneous Neumann boundary conditions
Some related work considered the Hopf bifurcation in predator–prey models. According to the results of [20] and the analysis of the stability and bifurcation system (1.1) in [16], we mainly consider the dynamics of the following system:
As far as we know, there are no results on the Hopf bifurcation of system (1.2). In this paper, we investigate the stability of the positive equilibrium, delay-induced Hopf bifurcation, and the properties of Hopf bifurcation such as the direction of the bifurcation and stability of the bifurcating periodic solutions.
2 The stability of equilibrium and the existence of Hopf bifurcation
In this paper, we only investigate the effect of the delay on the stability of the positive equilibrium \(E^{*}(u^{*},v^{*})\) of system (1.2), where
under condition \((H)\): \(\beta <\alpha +\eta \).
For simplicity, in this paper, we only study the case \(\tau_{1}=\tau _{2}=\tau \). Let
The linearization of (1.2) at the equilibrium \(E^{*}\) is
where
with
So, the characteristic equation of (2.1) is
where I is the \(2\times 2\) identity matrix, and \(M_{k}=-k^{2}\operatorname{diag}\{ d_{1},d_{2} \} \), \(k\in N_{0}=\{0,1,2,3,\ldots \}\). This is
When \(\tau =0\), it becomes
where
When \(\tau \neq 0\), assume that iω (\(\omega >0\)) is the root of (2.3). Then we have
Separating the real and imaginary parts of (2.4), we get
which implies that
where
For \(0< k< N_{1}\), we can conclude that (2.6) has only one positive real root
By (2.5) we obtain
for \(k\in \{ 0,1,2,\ldots,N_{1} \} \).
Lemma 2.1
If condition \((H)\) hold, then
for \(j\in N_{0}\).
Lemma 2.2
Let condition \((H)\) hold, \(T_{k}>0\) and \(D_{k}>0\) for \(k\in N_{0}\), and let \(\omega_{k}\) and \(\tau_{k}^{j}\) be defined by (2.7) and (2.8), respectively. Then (2.2) has a pair of purely imaginary roots \(i\omega_{k}\) for each \(k\in \{ 0,1,2, \ldots,N_{1} \} \), and (2.2) has no purely imaginary roots for \(k\geq N_{1}+1\).
Clearly, \(\tau_{k}^{0}=\min_{j\in N_{0}} \{ \tau_{k}^{j} \}\), \(k \in \{0,1,2,\ldots,N_{1}\}\), and from Lemmas 2.1 and 2.2 we know that \(\tau_{0}^{0}=\min \{ \tau_{k}^{j}: 0\leq k\leq N_{1}, j\in N_{0} \} \). Denote \(\tau^{*}=\tau_{0}^{0}\). Let \(\lambda ( \tau)=\alpha (\tau)+i\delta (\tau)\) be the pair of roots of (2.2) near \(\tau =\tau_{k}^{j}\) satisfying \(\alpha ( \tau_{k}^{j} ) =0\) and \(\delta ( \tau_{k}^{j} ) = \omega_{k}\). Then we have the following transversality condition.
Lemma 2.3
For \(k\in \{0,1,2,\ldots,N_{1}\}\) and \(j\in N_{0}\), \(\frac{d \operatorname{Re}(\lambda)}{d \tau }|_{\tau =\tau_{k}^{j}}>0\).
Proof
Differentiating two sides of (2.2), we get
Thus, by (2.5) and (2.7) we have
From Lemmas 2.1–2.3 and from the qualitative theory of partial differential equations we obtain the following results. □
Theorem 2.1
Assume that condition \((H)\) holds and \(\omega_{k}\) and \(\tau_{k}^{j}\) are defined by (2.7) and (2.8), respectively. Denote the minimum of the critical values of delay by \(\tau^{*}=\min_{k,j} \{ \tau_{k}^{j} \} \).
-
(a)
The positive equilibrium \(( u^{*},v^{*} ) \) of system (1.2) is asymptotically stable for \(\tau \in ( 0,\tau^{*} ) \) and unstable for \(( \tau^{*},+\infty ) \);
-
(b)
System (1.2) undergoes Hopf bifurcations near the positive equilibrium \(( u^{*},v^{*} ) \) at \(\tau_{k}^{j}\) for \(k\in \{ 0, 1, 2, \ldots, N_{1} \} \) and \(j\in N_{0}\).
3 Direction of Hopf bifurcation and stability of bifurcating periodic solution
From Theorem 2.1 we know that model (1.2) undergoes Hopf bifurcations near \(E^{*}\) at \(\tau =\tau_{k}^{j}\) and that a family homogeneous and inhomogeneous periodic solutions bifurcate from \(E^{*}\) of (1.2).
In this section, we investigate the stability of these Hopf bifurcations and bifurcating direction by using the normal formal theory of partial differential equation [21–23]. Without loss of generality, denote any of these critical values by \(\tau_{0}\), at which the characteristic equation (1.2) has a pair of simple purely imaginary roots \(i\omega _{0}\).
Setting \(\tilde{u}(\cdot,t)=u(\cdot,\tau t)-u^{*}\), \(\tilde{v}(\cdot,t)=v( \cdot,\tau t)-v^{*}\), \(\tilde{U}(t)= ( \tilde{u}(\cdot,\tau t), \tilde{v}(\cdot,\tau t) ) \), and \(\mathcal{C}= C([-1,0],X)\), \(X= \{ (u,v)\in W^{2,2}(0,\pi)|\frac{\partial u}{\partial x}=\frac{ \partial v}{\partial x}=0 \mbox{ at } x=0,\pi \} \) and then dropping the tildes for simplification of notation, system (1.2) can be written as the equation in the space \(\mathcal{C}\):
where for \(\varphi = ( \varphi_{1},\varphi_{2} ) ^{T}\in \mathcal{C}\), \(L(\mu)(\cdot):\mathcal{C}\rightarrow X \) and \(\tilde{G}:\mathcal{C}\times R\rightarrow X\) are given by
with \(g_{ijkl}^{(n)}=\frac{\partial^{i+j+k+l}\tilde{g}^{(n)} ( u ^{*},v^{*},u^{*},v^{*} ) }{\partial u^{i} \partial v^{j} \partial w^{k}\partial s^{l}}\), \(n=1,2\), and
By direct computation we obtain \(g_{0200}=g_{0002}=g_{1100}=g_{1010}=g _{0011}=0\).
Setting \(\tau =\tau_{0}+\varepsilon \), (3.1) is written as
where
So, \(\varepsilon =0\) is the Hopf bifurcation value for (3.2), and \(\Lambda_{0}= \{ -i\omega_{0}\tau_{0},i\omega_{0}\tau_{0} \} \) is the set of eigenvalues on the imaginary axis of the infinitesimal generator associated with the flow of the following linearized system of (3.2) at the origin:
The eigenvalues of \(\tau_{0}D_{0}\Delta \) on X are \(\mu^{i}_{k}=-d _{i}\tau_{0}k^{2}\), \(i=1,2\), \(k\in N_{0}\), with the corresponding normalized eigenfunctions \(\beta^{i}_{k}\), where
Let \(\mathbf{B}_{k}=\operatorname{span}\{ \langle v(\cdot),\beta_{k}^{i}\rangle \beta_{k}^{i}|v \in \mathcal{C},i=1,2 \} \). Then it is easy to verify that \(L(\tau_{0})(\mathbf{B}_{k})\subset \operatorname{span}\{ \beta_{k}^{1},\beta _{k}^{2} \} \). Assume that \(z_{t}(\theta)\in \mathcal{C}=C ( [-1,0],R ^{2} ) \) and
Then, the linear PDE restricted on \(\mathbf{B}_{k}\) is equivalent to the following FDE on \(\mathcal{C}=C ( [-1,0],R^{2} ) \):
When \(\tau =\tau_{0}\), define \(\eta (\theta)\in BV ( [-1,0],R ) \) such that
and the adjoint bilinear form on \(\mathcal{C}^{*}\times \mathcal{C}\), \(\mathcal{C}^{*}=C ( [0,1],R^{2*} ) \) by
Then, for (3.2) with fixed k, the dual bases \(\Phi_{k}\) and \(\Psi_{k}\) for its the eigenspace P and its dual space \(P^{*}\) are given by
where \(\langle \Phi_{k},\Psi_{k}\rangle =I_{2}\), the \(2\times 2\) identity matrix, and
with \(D= ( 1+2\tau_{0} ( i\omega_{0}+d_{1}k^{2}-b_{11} ) +\frac{ ( i\omega_{0}+d_{1}k^{2}-b_{11} ) e^{2i\omega_{0}\tau_{0}}}{b _{12}b_{21}} ) ^{-1}\).
Following the standard procedure in [21, 23], especially [22], we can obtain the following normal form on the center manifold:
where
with
where
and
where
with
where \(h_{k20}(\theta)\) and \(h_{k11}(\theta)\) are determined by
with
Through the change of variables \(z_{1}=\omega_{1}-i\omega_{2}\), \(z_{2}= \omega_{1}+i\omega_{2}\), and \(\omega_{1}=\rho_{1}\cos \phi\), \(\omega _{2}=\rho_{1}\sin \phi \), the normal form [20] becomes the following polar coordinate system:
where \(\kappa_{k1}=\operatorname{Re}A_{k1}\) and \(\kappa_{k2}=\operatorname{Re}A_{k2}\). Thus, from [28] we see that the sign of \(\kappa_{k1}\kappa_{k2}\) determines the direction of the bifurcation and the sign of \(\kappa_{k2}\) determines the stability of the nontrivial periodic orbits and have following results.
-
(a)
When \(\kappa_{k1}\kappa_{k2}<0\), system (1.2) undergoes Hopf bifurcation at the critical value \(\tau =\tau^{*}\), which is a supercritical bifurcation. Moreover, if \(\kappa_{k2}<0\), then the bifurcating periodic solution is stable; if \(\kappa_{k2}>0\), then the bifurcating periodic solution is unstable.
-
(b)
When \(\kappa_{k1}\kappa_{k2}>0\), system (1.2) undergoes Hopf bifurcation at the critical value \(\tau =\tau^{*}\), which is a subcritical bifurcation. Moreover, if \(\kappa_{k2}<0\), then the bifurcating periodic solution is stable; if \(\kappa_{k2}>0\), then the bifurcating periodic solution is unstable.
4 Numerical simulations
In this section, we give some numerical simulations to support and extend our results by using the mathematical software Matlab.
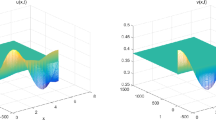
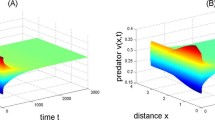
For system (1.2), we choose \(d_{1}=0.2\), \(d_{2}=1\), \(\delta =2\), \(\alpha =0.05\), \(\beta =0.3\), \(h=0.8\), \(c=0.1\), and \((u(x,0),v(x,0))=(0.62+0.005\cos x,0.78+0.005 \cos x)\). Then, a series of calculations show that \((u^{*},v^{*})=(0.6250,0.7813)\) and \(\tau^{*}=6.8222\). Hence \((0.6250,0.7813)\) is locally stable when \(\tau \in [0,\tau^{*})\). When τ crosses through the critical \(\tau^{*}\), \((0.6250,0.7813)\) loses its stability, Hopf bifurcation occurs, and a family of periodic solutions are bifurcation from \((0.6250,0.7813)\). The direction and stability of Hopf bifurcation can be determined by the signs \(\kappa_{k1}\) and \(\kappa_{k2}\); by the procedure of Sect. 3, \(\kappa_{k1}=0.0150\) and \(\kappa_{k2}=1.1544\). So, the Hopf bifurcation occurring at \(\tau^{*}\) is subcritical, and the corresponding Hopf bifurcation periodic orbits are unstable. Taking \(\tau =6.5<6.8222\), the numerical simulation results of system (1.2) are depicted Fig. 1. And taking \(\tau =6.85>6.8222\), the numerical simulation results of system (1.2) are depicted Fig. 2, which is in agreement with the theoretical results.

The positive equilibrium \(( u^{*},v^{*} ) =(0.6250,0.7813)\) of (1.2) is asymptotically stable when \(\tau =6.5< 6.8222\). Here we set parameter values \(d_{1}=0.2\), \(d_{2}=1\), \(\delta =2\), \(\alpha =0.05\), \(\beta =0.3\), \(h=0.8\), \(c=0.1\) and the initial value \((u(x,0),v(x,0))=(0.62+0.005\cos x,0.78+0.005\cos x)\)

There exists unstable spatially homogenous periodic bifurcating from the positive equilibrium \(( u^{*},v^{*} ) =(0.6250,0.7813)\) of (1.2) when \(\tau =6.85>6.8222\). Here we set parameter values \(d_{1}=0.2\), \(d_{2}=1\), \(\delta =2\), \(\alpha =0.05\), \(\beta =0.3\), \(h=0.8\), \(c=0.1\). (A)–(B) The initial value \((u(x,0),v(x,0))=(0.62+0.05\cos x,0.78+0.05\cos x)\); (C)–(D) The initial value \((u(x,0),v(x,0))=(0.62+0.05\sin x,0.78+0.05\sin x)\)
5 Conclusions
This paper mainly consider about the effects of delay τ on dynamics of a diffusive predator-prey model with Simth growth rate under Neumann boundary conditions. Based on the analysis of the characteristic equations, we study the stability of the equilibrium and the existence of delay-induced Hopf bifurcations. Then, by the normal forms on the center manifold, we obtain the results determining the direction and stability of Hopf bifurcation. Finally, choosing the parameter values \(d_{1}=0.2\), \(d_{2}=1\), \(\delta =2\), \(\alpha =0.05\), \(\beta =0.3\), \(h=0.8\), \(c=0.1\), we get the critical value of delay \(\tau^{*}=6.8222\) with a direct computation. From Theorem 2.1 we know that the positive equilibrium \(( u^{*},v^{*} ) =(0.6250,0.7813)\) is asymptotically stable for \(\tau <\tau^{*}\) and unstable for \(\tau >\tau^{*}\). So, system (1.2) undergoes Hopf bifurcation near the positive equilibrium \(( u^{*},v ^{*} ) =(0.6250,0.7813)\) when the delay τ increasingly crosses through the critical value \(\tau^{*}\).
For the predator–prey systems, pattern dynamics is also an interesting topic. Many authors have discussed the pattern dynamics of the diffusion equations [29–33]. In the next topics, we will discuss the pattern dynamics of a diffusive predator–prey model with Simth growth rate.
References
Gause, G.F.: The Struggle for Existence. Hafner, New York (1969)
Holling, C.: The functional response of predator to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 45, 1–60 (1965)
Tanner, J.T.: The stability and the intrinsic growth rates of prey and predator populations. Ecology 56, 855–886 (1975)
Wollkind, D.J., Collings, J.B., Logan, J.A.: Metastability in a temperature dependent model system for trees. Bull. Math. Biol. 50, 379–409 (1988)
May, R.M.: Stability and Complexity in Model Ecosystems. Princeton University Press, Princeton (1973)
Li, X., Jiang, W.: Hopf bifurcation and Turing instability in the reaction–diffusion Holling–Tanner predator–prey model. IMA J. Appl. Math. 78, 287–306 (2013)
Murray, J.D.: Mathematical Biology II. Springer, Heidelberg (2002)
Shi, H., Ruan, S.: Spatial, temporal and spatiotemporal patterns of diffusive predator–prey models with mutual interference. IMA J. Appl. Math. 80, 1534–1568 (2015). https://doi.org/10.1093/imamat/hxv006
Yang, R., Wei, J.: Stability and bifurcation analysis of a diffusive prey–predator system in Holling type III with a prey refuge. Nonlinear Dyn. 79, 631–646 (2015)
Yi, F., Wei, J., Shi, J.: Bifurcation and spatio-temporal patterns in a homogeneous diffusive predator–prey system. J. Differ. Equ. 246, 1944–1977 (2009)
Wang, J., Shi, J., Wei, J.: Dynamics and pattern formation in a diffusive predator–prey system with strong Allee effect in prey. J. Differ. Equ. 251, 1276–1304 (2011)
Fan, M., Wang, K.: Periodicity in a food-limited population model with toxicants and time delays. Acta Math. Appl. Sin. 18, 309–314 (2002)
Gopalsamy, K., Kulenovic, M.R.S., Ladas, G.: Environmental periodicity and time delays in a food-limited population model. J. Math. Anal. Appl. 147, 545–555 (1990)
Smith, F.E.: Population dynamics in Daphnia Magna and a new model for population growth. Ecology 44, 651–663 (1963)
Sivakumar, M., Sambath, M., Balachandran, K.: Stability and Hopf bifurcation analysis of a diffusive predator–prey model with Smith growth. Int. J. Biomath. 8(1), 1550013 (2015)
Yue, Z., Wang, W.: Qualitative analysis of a diffusive ratio-dependent Holling–Tanner predator–prey model with Smith growth. Discrete Dyn. Nat. Soc. 2013, Article ID 267173 (2013). https://doi.org/10.1155/2013/267173
Ruan, S.: On nonlinear dynamics of predator–prey models with discrete delay. Math. Model. Nat. Phenom. 4(2), 140–188 (2009)
Cao, J., Yuan, R.: Bifurcation analysis in a modified Lesile–Gower model with Holling-type II functional response and delay. Nonlinear Dyn. 84(3), 1341–1352 (2016). https://doi.org/10.1007/s11071-015-2572-5
Lian, F., Xu, Y.: Hopf bifurcation analysis of a predator–prey system with Holling-type IV functional response and time delay. Appl. Math. Comput. 215, 1484–1495 (2009)
Chen, S., Shi, J., Wei, J.: Global stability and Hopf bifurcation in a delayed diffusive Leslie–Gower predator–prey system. Int. J. Bifurc. Chaos 22, 1250061 (2012)
Faria, T.: Stability and bifurcation for a delayed predator–prey model and the effect of diffusion. J. Math. Anal. Appl. 254, 433–463 (2001)
Faria, T.: Normal forms and Hopf bifurcation for partial differential equations with delay. Trans. Am. Math. Soc. 352, 2217–2238 (2000)
Song, Y., Peng, Y., Zou, X.: Persistence, stability and Hopf bifurcation in a diffusive ratio-dependent predator–prey model with delay. Int. J. Bifurc. Chaos 24, 1450093 (2014)
Sun, G., Wang, S., Ren, Q.: Effects of time delay and space on herbivore dynamics: linking inducible defenses of plants to herbivore outbreak. Sci. Rep. 5, 11246 (2015)
Li, L., Jin, Z., Li, J.: Periodic solutions in a herbivore–plant system with time delay and spatial diffusion. Appl. Math. Model. 40(7–8), 4765–4777 (2016)
Ma, Z., Huo, H., Xiang, H.: Hopf bifurcation for a delayed predator–prey diffusion system with Dirichlet boundary condition. Appl. Math. Comput. 311, 1–18 (2017)
Tang, H., Liu, Z.: Hopf bifurcation for a predator–prey model with age structure. Appl. Math. Model. 40(2), 726–737 (2016)
Chow, S.N., Hale, J.K.: Methods of Bifurcation Theory. Springer, New York (1982)
Guin, L.N.: Existence of spatial patterns in a predator–prey model with self- and cross-diffusion. Appl. Math. Comput. 226, 320–335 (2014)
Sun, G., Wu, Z., Wang, Z., Jin, Z.: Influence of isolation degree of spatial patterns on persistence of populations. Nonlinear Dyn. 83(1–2), 811–819 (2016)
Sun, G.: Mathematical modeling of population dynamics with Allee effect. Nonlinear Dyn. 85(1), 1–12 (2016)
Sun, G., Wang, C., Wu, Z.: Pattern dynamics of a Gierer–Meinhardt model with spatial effects. Nonlinear Dyn. 88(2), 1385–1396 (2017)
Peng, Y., Ling, H.: Pattern formation in a ratio-dependent predator–prey model with cross-diffusion. Appl. Math. Comput. 331, 307–318 (2018)
Acknowledgements
The authors are grateful to the referee for carefully reading the manuscript and for providing some comments and suggestions which led to improvements in the paper.
Funding
The research of H. Fang was supported by the key project of Provincial Excellent Talents in University of Anhui Province (gxyqZD2016300, gxyqZD2018077) and the Natural Science Foundation of Huangshan University (2017xkjq001). The research of L. Hu was supported by the National Natural Science Foundation of China (11701208). The research of Y. Wu was supported by the Natural Science Foundation of Anhui Province (1708085MA04) and the Key NSF of Anhui Educational Committee (KJ2018A0428).
Author information
Authors and Affiliations
Contributions
All authors contributed equally and read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Fang, H., Hu, L. & Wu, Y. Delay-induced Hopf bifurcation in a diffusive Holling–Tanner predator–prey model with ratio-dependent response and Smith growth. Adv Differ Equ 2018, 285 (2018). https://doi.org/10.1186/s13662-018-1726-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-018-1726-4